
Submitted:
31 May 2023
Posted:
31 May 2023
You are already at the latest version
Abstract
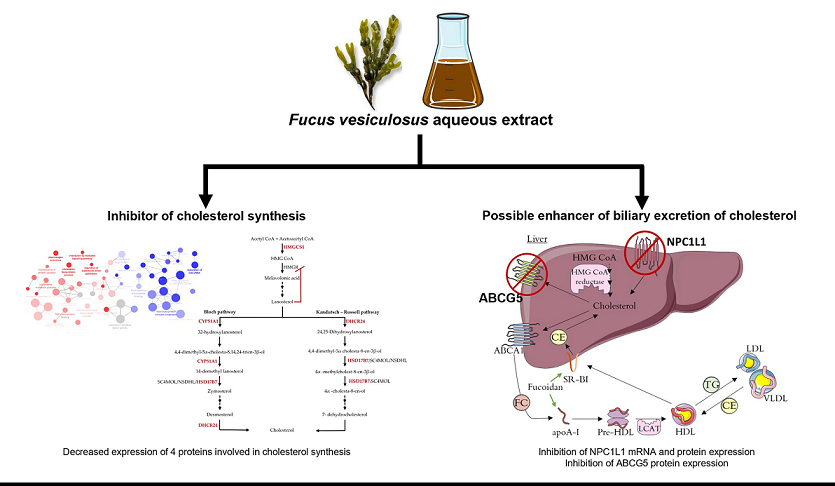
A brown seaweed consumed worldwide, Fucus vesiculosus, has been used to prevent atherosclerosis and hypercholesterolemia, among other uses, however, the mechanisms of action that lead to these effects are not yet fully understood. An F. vesiculosus aqueous extract, prepared as a soup was characterized as rich in sev-eral bioactive compounds, mainly phlorotannin and peptides. This work aims to study the in vitro effect of this extract on the expression of different proteins involved in the synthesis and transport of cholesterol in HepG2 cells, though a proteomic analysis, western blot and qRT-PCR. The results demonstrated that, in liver cells, the extract decreases the expression of 4 proteins involved in cholesterol biosynthesis process and also decrease the expression of two important transporters proteins of cholesterol, NPC1L1 and ABCG5, as well as decrease the NPC1L1 mRNA levels. Our study demonstrates some mechanisms of action of bioactive com-pounds from F. vesiculosus that may explain its previously reported hypocholesterolemic effect.
Keywords:
Fucus vesiculosus
; cholesterol synthesis
; cholesterol excretion
; Ezetimibe
; NPC1L1
; ABCG5
; qRT-PCR
; mRNA
1. Introduction
The principal cause of global mortality is cardiovascular diseases (CVDs), specifically ischemic heart disease (IHD) and stroke [1]. The prevalence of CVDs is linked to unhealthy dietary habits with high consumption of salt, refined carbohydrates and fats, such as cholesterol [2,3]. High levels of cholesterol in blood and tissues are one of the major risks for lethal myocardial infarction and stroke due to the formation of arterial plaques [4]. It is estimated that between 1990 and 2017 the global number of deaths increased by approximately 910,000 due to high levels of non-HDL cholesterol which consequently led to IHD and stroke [5].
Previous studies report that the treatment of high cholesterol levels leads to significant health benefits [1]. The plasma cholesterol levels can be regulated through different mechanisms, such as the de novo cholesterol synthesis, synthesis of bile acids, excretion of cholesterol to bile and intestinal cholesterol absorption [2].
Cholesterol synthesis in liver is a complex, multi-step process involving different enzymes. There are two main possible pathways for cholesterol synthesis the Bloch and the Kandutsch-Russell pathway [6,7]. They share the first steps from acetyl-coA to lanosterol synthesis, after this step, they branch out, although having a few enzymes in common [6,7].
Reverse cholesterol transport (RCT) is an important a pathway that transports cholesterol from non-hepatic tissues to the liver for secretion in bile (White et al., 2010). Although the transport of cholesterol to phospholipids acceptors occurs spontaneously in all cells this is an inefficient process by itself [8]. In RCT three important proteins that play a key role in cholesterol transport are known namely, ATP binding cassette transporter (ABC) A1, ABCG1 and scavenger receptor BI (SR-BI) (Jessup et al., 2006; Song et al., 2012). Cholesterol can also be reabsorbed to the liver by Niemann-Pick C1-Like 1 (NPC1L1) protein, an essential protein that regulates plasma cholesterol levels [2]. In the liver, NPC1L1 has the ability to transport free cholesterol from the canalicular bile back to hepatocytes, thus presenting an opposite mechanism to the ABCG5 / ABCG8 transporter which in turn regulates biliary cholesterol secretion [9]. This protein is also critical to regulating intestinal cholesterol absorption [2]. The NPC1L1 protein is a molecular target of one of the most used drugs in treating hypercholesterolemia, ezetimibe [10]. This drug acts by blocking the internalization of NPC1L1 and consequently decreasing cholesterol uptake [11], leading both to inhibition of intestinal absorption of dietary cholesterol and biliary cholesterol absorption [12].
Another way to regulate blood cholesterol levels is through healthy dietary habits, with the consumption of foods low in saturated fat and nutrients from natural sources, and even complement with the use of natural food supplements. [13].
In recent years, algae have received attention as a food product due to their beneficial effects, namely brown algae, which are rich in several bioactive compounds as phlorotannins [14]. Fucus vesiculosus is a brown algae consumed in different parts of the world and has traditionally been used for several reported beneficial effects, among which: prevention of mineral deficit, weight loss, arthrosis, arthritis, atherosclerosis, viscous blood and hypercholesterolemia, and as an adjuvant for menopause [15,16,17,18]. Particularly regarding the reduction of cholesterol levels, in vivo and in vitro studies with extracts rich in phlorotannins have reported as decreasing the levels of LDL, triglycerides and total cholesterol [19,20,21]. However, the mechanisms of action of brown algae biomolecules that lead to cholesterol-lowering effects remain unclear. Our group have already reported an in vitro study with the aqueous extract of F. vesiculosus, prepared in the form of soup and characterized as rich in phlorotannins and peptides, reported its beneficial effect in vitro in inhibiting the intestinal absorption of cholesterol, as well as in inhibiting its synthesis [22]. Other published study demonstrated that the extract under study in vitro led to the increase of several lipid compounds in HepG2 cells, including fatty acid amides, which are described as inhibitors of the ACAT enzyme and consequently inhibitors of cholesterol absorption and plasma cholesterol levels [23].
The objective of the present work is to evaluate the effect of the aqueous extract of F. vesiculosus, prepared as soup and previously characterized as rich in phlorotannins and peptides [22], on the liver proteins involved in the cholesterol biosynthetic process, through proteomic analysis. It is also intended to study the specific effect of F. vesiculosus extract on important cholesterol transporter proteins NPC1L1 and ABCG5, with molecular assays as qRT-PCR and Western Blot
2. Materials and Methods
2.1. Chemicals
All chemicals were of analytical grade. Water, methanol (MeOH), formic acid and acetonitrile LC-MS grade Optima were purchased from Fisher Scientific (Hampton, USA). Ethanol 96% was bought from Carlo Erba (Peypin, France). Dulbecco's modified Eagle medium (DEMEM), Trypsin, Glutamine, Phosphate-Buffered Saline (PBS), fetal bovine serum (FBS) and Tween 20 were obtained from Lonza® (Verviers, Belgium). Glacial acetic acid and Tris(hydroxymethyl)aminomethane were obtained from Merck Milipore® (Massachusetts, EUA).
Glycine, Bovine Serum Albumin (BSA), Igepal® CA-630, Iodoacetamide, Urea, Ammonium bicarbonate and Glucose were purchased from Sigma-Aldrich (Barcelona, Spain). Pierce™ DTT (Dithiothreitol), Pierce™ Trypsin Protease MS Grade, mini Protein Gel NuPAGE™ 4 to 12% Bis-Tris, Bolt® MOPS Transfer Buffer (20X), Bolt® MOPS SDS Running Buffer (20X), PageRuler™ Prestained Protein Ladder and 4X Bolt™ LDS Sample Buffer were obtained from Thermo Fisher Scientific (Waltham, USA), Coomassie Brilliant Blue R-250 was purchased from BIORAD® (Hercules, USA). NZYBlue Protein Marker, 5× SDS-PAGE Sample Loading Buffer, NZY Total RNA Isolation kit, NZY First-Strand cDNA Synthesis Kit and NZYSpeedy qPCR Green Master Mix ROX plus were purchased from Nzytech® (Lumiar, Portugal). AmershamTM ProtranTM Premium 0.45 µm Nitrocellulose Blotting Membrane, AmershamTM ECLTM Prime Western Blotting Detection Reagents, AmershamTM ECLTM Prime Blocking Agent and Amersham ECL™ anti-rabbit IgG, horseradish peroxidase-linked whole antibody (from donkey) were obtained from VWR International (Radnor, USA). ABCG5 and NPC1L1 antibodies were purchased from Novus Biologicals. NPC1L1 and GAPDH primers were purchased from Alfagene (Lisbon, Portugal).
2.2. Algae Extract Preparation and Identification of Compounds
Whole dried brown seaweed F. vesiculosus Linnaeus from the North Atlantic Ocean was purchased fromCeleiro diet., Lisbon, Portugal (imported by Américo Duarte Paixão Lda, Lot number 03ALG2731901). The aqueous extraction and the purification by Solid Phase Extraction (SPE) of F. vesiculosus were performed as described in [22]. The identification of the extract compounds was performed though Liquid Chromatography by High Resolution Mass Spectrometry (LC-HRMS/MS) using an Elute OLE UHPLC system interfaced with a quadrupole time-of-flight (QqToF) Impact II mass spectrometer equipped with an electrospray source (ESI) (Bruker DaltoniK GmbH, Bremen, German), the results were previously presented in (André et al., 2020).
2.3. Cell Culturing
Caco-2 cells (ECACC 86010202), a human colorectal adenocarcinoma epithelial cell line and HepG2 (ECACC 85011430), a human hepatocellular liver carcinoma cell line, were cultured in DMEM supplemented with 10% and 20% FBS, respectively, and 2 mM L-glutamine at 37 °C in an atmosphere with 5% CO2. The culture cells were kept in sub-confluence with trypsinization every 72h.
2.4. Membrane Protein Extraction and SDS-PAGE Electrophoresis
HepG2 cells seeded in T75 flasks were under contact with the F. vesiculosus extract at 0.25 mg/mL (IC30) [23]] and culture medium without FBS (control) during 24h. After incubation time, the cells were washed twice with PBS, scraped with water and transferred to a pre-weight eppendorf tube. The cells were then lyophilized in a Heto PowerDry 3000 apparatus (Thermo Fisher Scientific, Waltham, USA). Approximately 3 mg of cells of control and cells exposed to extract were used to obtain the fractions of membrane proteins using the Mem-PER Plus Membrane Protein Extraction Kit (Thermo Scientific™) following the manufacturer’s indications. The different samples of both protein fractions were separated under reducing conditions in NuPAGE 4 to 12% gradient gels (Invitrogen™, Carlsbad, USA) using a Mini Gel Tank (Invitrogen™, Carlsbad, USA) according to the manufacturer’s instructions. The gels were stained with 40% of Coomassie R-250 blue, 50% of methanol and 10% of glacial acetic acid during 1h and distaining with a solution of 7.5 % glacial acetic acid, 10 % ethanol and 82.5 % distilled water overnight. Gels were photographed using ImageQuant LAS 50 (GE Healthcare Life Sciences®, Illinois, USA) and the images were analysed using ImageJ software.
2.5. In-Gel Protein Digestion, Nano-LC−ESI−MS/MS and DataAnalysis
In-gel protein digestion was performed as described in [24]. The nLC-MS/MS analysis of the resulting peptide digests was performed as described in [25], using an Ultimate 3000 nLC apparatus coupled to a UHR-QqTOF IMPACT HD apparatus (Bruker Daltonics, Bremen, German) with a CaptiveSpray ion source (Bruker Daltonics, Bremen, German). Raw LC−MS/MS data were processed in MaxQuant (V.1.6.10.43) for automated protein identification. MS raw files were analyzed by MaxQuant software, version 1.6.10.43 [26], and peptide lists were searched against the human Uniprot FASTA database. A contaminant database generated by the Andromeda search engine [27] was configured with cysteine carbamidomethylation as a fixed modification and N-terminal acetylation and methionine oxidation as variable modifications. We set the false discovery rate (FDR) to 0.01 for protein and peptide levels with a minimum length of seven amino acids for peptides, and the FDR was determined by searching a reverse database. Enzyme specificity was set as C terminal to arginine and lysine as expected using trypsin. A maximum of two missed cleavages were allowed. Data processing was performed using Perseus (version 1.6.2.3) with default settings [28].
All proteins and peptides matching the reversed database were filtered out. Subcellular localization and gene ontology analysis were performed using STRING online resources at https://string-db.org/, and ClueGo plug-in in Cytoscape (V3.9.0), respectively [29].
2.6. Western Blot Analysis
HepG2 cells were seeded in T25 culture flasks and after confluence were exposed to DMEM medium without FBS containing 0.25 mg/mL of F. vesiculosus aqueous extract purified by SPE (F. vesiculosus extract). After 24h, the cells were scraped and collected with water and lyophilized. The cells were dissolved in lysis buffer (Igepal 4%, DTT 1%, Urea 6 M), at a concentration of 0.025 mg of cell/ mL, followed by sonication, for 5 min, and centrifugation for 10 min at 10 000 rpm. The cells precipitated were used for western blot following the protocol described in [30]. The assays were performed in triplicate and the results are presented in terms of mean and standard deviation.
2.7. Real Time Quantitative PCR
HepG2 cells were seeded in T75 culture flasks and after confluence were exposed to DEMEM medium without FBS, to 0.25 mg/mL of F. Vesiculosus aqueous extract and to 100 µM of ezetimibe. 24 h post exposition, RNA was harvested from cells using NZY First-Strand cDNA Synthesis Kit. Each experiment of RNA extraction was carried out with duplicate samples. cDNA was synthesized from 1 µg of RNA using NZY First-Strand cDNA Synthesis Kit following the manufacturer’s protocol. The qRT-PCR was performed in triplicates using NZYSpeedy qPCR Green Master Mix (2x), ROX plus. The primers sequence used to amplify GAPDH and NPC1L1 genes were described in [31]. Reactions were performed in the Applied Biosystems 7500 Real-Time PCR System (Thermo Fisher Scientific, Waltham, USA), and the real-time PCR program consisted of 40 cycles (95 °C for 15 s and 62 °C for 30 s) after initial 10 min incubation at 95 °C. The expression of NPC1L1 was determined relative to GAPDH and data are presented as mean values with standard deviations.
2.8. Statistical Analysis
The data were expressed as mean ± standard deviation using the Microsoft® Excel 2016 software. Statistical analysis was performed using one-way analysis of variance (ANOVA) using software developed by Microsoft® with p-value ≤ 0.05 considered as significant.
3. Results and Discussion
3.1. Effect of F. Vesiculosus on Hepatic Proteins
Different studies on the hypocholesterolemic effect of different brown seaweeds have been carried out, but the molecular mechanism of the bioactive compounds in brown seaweeds that lead to a decrease in total cholesterol levels is still not fully understood. The effect of F. vesiculosus extract on hepatic proteins was evaluated through SDS-PAGE electrophoresis of membrane proteins from HepG2 cells exposed to a non-cytotoxic concentration of F. vesiculosus extract [23] (Figure 1).
The extract caused several changes in the relative intensity of different proteins (Figure 1A-B), namely, 65 % of the bands analysed using the ImageJ program show significant changes (ANOVA test with p-value ≥ 0.05) in their intensity in the presence of the extract compared to the control (Figure 1B).
Proteomic Analysis
A proteomic analysis was performed to identify some proteins from HepG2 cells present in the previous electrophoretic gel presented. For the proteomic assay, gel bands from distinct zones of the gel with different molecular weights (highlighted in red in Figure 1B) and different intensities were removed from both control and cell under the effect of the extract. A total of 809 protein groups were identified, from which 671 were detected in the control experiments and 695 in the extract. 68.9% (557) of the protein groups were detected in both control and test cells, whereas 114 proteins were exclusively present in the control cells, and 138 were exclusively present in cells exposed to the extract (Figure 2).
Proteins expressed only in the control and those expressed only in cells exposed to the extract were analysed in different databases depending on the different objectives intended with the analysis of the results. The analysis of gene ontology (GO) terms biological process (BP) and molecular function (MP) were analysed using ClueGO Cytoscape App. The networks in Figure 3A show the GO terms more significantly enriched for control cells (red) and cells exposed to the extract (blue), followed by the statistics graphs representing the percentage of gene by term for the control cells (Figure 3B) and for the cells exposed to extract (Figure 3C). Analysing Figure 3, it is possible to observe that the proteins from both groups participate in several biological processes with different molecular functions; however, in this study, we intended to highlight the proteins from the cholesterol biosynthetic process due to the previous reported hypocholesterolemic effect of F. vesiculosus aqueous extract under study [22,23].
The KEGG and Reactome pathways in which the proteins under study participate were also analysed using ClueGO Cytoscape App (Figure 4).
The protein Lanosterol 14-alpha demethylase (CYP51A1), enzyme 3-hydroxysterol 24-reductase (DHCR24), 3-Hydroxy-3-Methylglutaryl-CoA Synthase 1 (HMGCS1), and Hydroxysteroid 17-Beta Dehydrogenase 7 (HSD17B7) involved in the cholesterol biosynthesis pathway (Figure 5) were detected in the control cells, whereas they were not detected in the cells after 24h contact with the F. vesiculosus extract.
DHCR24 catalyses the final step of the Bloch pathway of cholesterol synthesis and also catalyses the first step of Kandutsch – Russell pathway [7], as shown in Figure 5. Previous studies with HepG2 cells demonstrated that the inhibition of DHCR24 leads to decreased cholesterol production [37]. HSD17B7 is another enzyme common to both pathways of cholesterol synthesis in different steps (Figure 5), and a previous study demonstrated that the lack of this enzyme inhibits the de novo cholesterol biosynthesis [38]. CYP51A1 is known as a critical cholesterologenic enzyme essential for regulating cholesterol biosynthesis [39]. In previous studies, inhibition of CYP51A1 has shown to lead to significant reductions in serum levels of total cholesterol and serum low-density proteins [39]. Furthermore, inhibition of CYP51A1 leads to accumulation of lanosterol which, in turn, induces the degradation and ubiquitination of HMG CoA reductase enzyme (HMGR), the rate-limiting step in cholesterol synthesis [39,40]. In a previous study, F. vesiculosus extract also shows a significant capacity to in vitro inhibit the HMGR with an half inhibitory concentration (IC50) of 4.16 µg/mL [22]. The other protein from the cholesterol biosynthesis pathway, HMGCS1, catalyses the condensation of acetyl-CoA with acetoacetyl-CoA to form HMG-CoA (Figure 5) (https://www.uniprot.org/uniprot/Q01581), and its inhibition consequently leads to inhibition of cholesterol synthesis [41].
As mentioned, these four proteins were only detected in control cells, which means that the presence of the extract is leading to a decrease in the expression of these proteins, leading in turn to the inhibition of cholesterol synthesis. The description of these molecular mechanisms of action of the extract on proteins involved in the synthesis of cholesterol is novel, which is in line with the hypocholesterolemic effect already reported [17].
3.2. Effect of Fucus Vesiculosus on Cholesterol Transporters
F. Vesiculosus Aqueous Extract Decrease Hepatic Expression of NPC1L1 and ABCG5
A western blotting and qRT-PCR techniques were performed to study the specific effect of F. vesiculosus extract on the hepatic expression of two important cholesterol transporter proteins, NPC1L1 and ABCG5.
This extract was previous characterized through LC-HRMS/MS analysis and the results demonstrated that phlorotannin derivatives and small peptides represented 94% of the intensities detected. As the extract under study is rich in phenolic compounds [22], namely phlorotannins, the western blot technique was performed using the same weight of cells per sample [30]. The standard protocol for this technique is based on standardizing samples based on protein amount however, previous studies [30,32] have reported that different phenolic compounds affect cellular protein content, making it impossible to use the total protein content and internal control proteins to do the standardizing of samples.
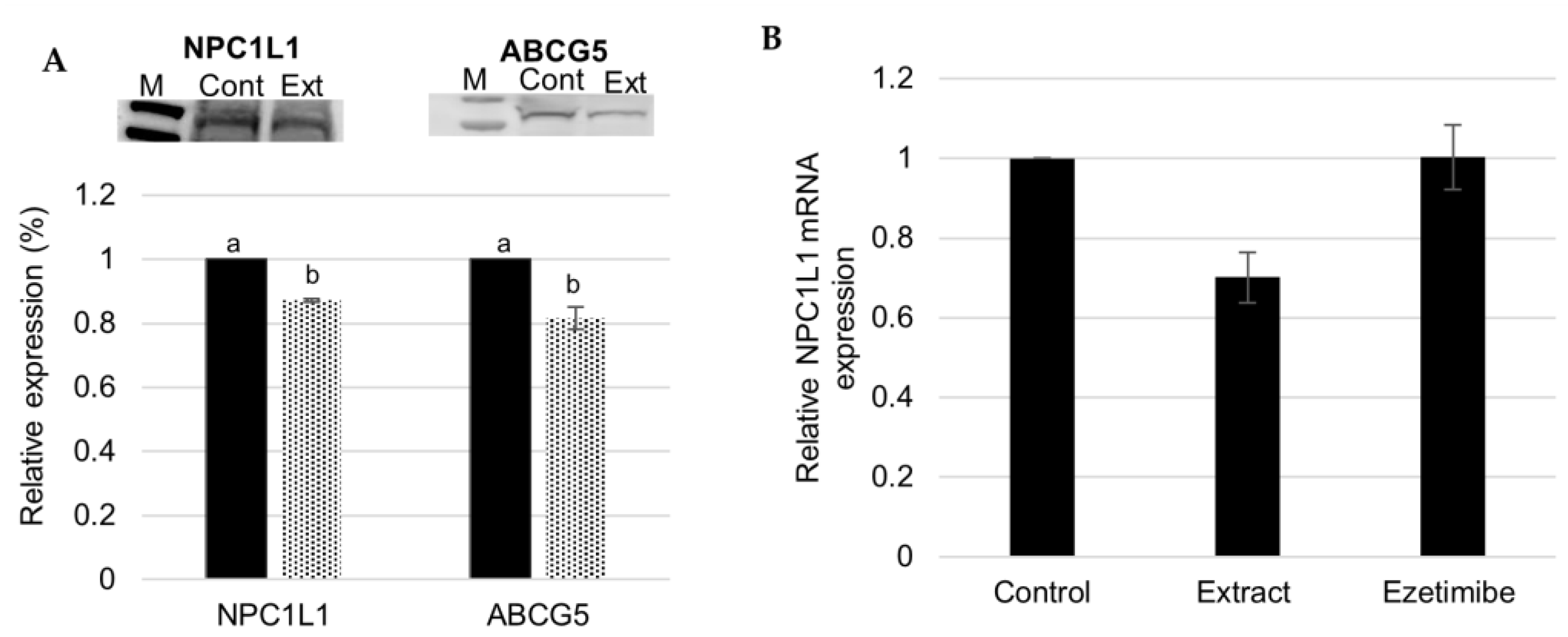
Figure 6A shows that, in HepG2 cells, the extract led to a decrease of NPC1L1 expression by 12.76 ± 0.47 %. The qRT-PCR demonstrated that the extract also led to NPC1L1 mRNA inhibition, being the relative expression of NPC1L1 in the presence of the extract of 0.70 ± 0.06 % in relation to control, which means an inhibition of approximately 30 % (Figure 6).
Previous studies have reported that overexpression of NPC1L1 in the liver causes an inhibitory effect on biliary cholesterol secretion, once re-absorbs cholesterol from bile leading to high levels of liver cholesterol and thus the risk of developing atherosclerosis and other CVD [2,33]. Consequently, the overexpression of hepatic NPC1L1 may aggravate diet-induced atherosclerosis [2]. The F. vesiculosus extract effect on the inhibition of NPC1L1 mRNA and protein expression could, therefore, be beneficial by increasing biliary excretion of cholesterol and, consequently, contribute to decrease the risk of atherosclerosis.
One of the drugs most used for the therapy of hypercholesterolemia is ezetimibe, which acts to block NPC1L1 internalization and consequently decrease cholesterol uptake [11]. Previous studies also reported that ezetimibe treatment does not affects protein or mRNA expression of NPC1L1 [34]. Considering that, the extract was seen to inhibit the expression of both NPC1L1 mRNA and protein, the study here reported could be a starting point for the search for new therapeutic strategy showing a different mechanism of action than ezetimibe.
F. vesiculosus aqueous extract also decreased the hepatic expression of ABCG5 by 18.40 ± 3.53 % (Figure 6A) relatively to the control cells. In the liver, the ABCG5 and the ABCG8 transporter form a complex (ABCG5/ABCG8) that mediates the cholesterol secretion to bile [33]. Based on this knowledge, one would expect that the extract would induce hepatic expression of ABCG5 to promote hepatic excretion of cholesterol in the bile, but this was not observed. Our hypothesis is that, as the extract leads to the inhibition of cholesterol synthesis, as demonstrated in the previous results presented in section 3.2 and as observed in a previous in vitro study [22], there is less need to eliminate cholesterol from liver into the bile, leading, consequently, to a lower expression of the ABCG5 protein. The observed inhibition of NPC1L1 expression also leads to lower cholesterol levels in the liver which also leads to a lesser need to eliminate cholesterol from this organ into the bile. The effect of statins, another group of drugs often prescribed to lower cholesterol levels in the expression of the ABCG5/ABCG8 transporter is still an area under study. Statins primarily acts by inhibiting HMGR leading to cholesterol synthesis inhibition. Studies in vivo demonstrated that some statins increase the expression of ABCG5 leading to increased biliary excretion of cholesterol [35,36]. By contrast, other studies reported that atorvastatin and pravastatin decrease the expression of ABCG5 at the intestinal and hepatic level [34,35].
Further in vivo studies and with primary cells are needed to clarify the effect of F. vesiculosus extract on biliary cholesterol excretion and on blood and liver cholesterol levels and to better understand its inhibitory effect towards ABCG5 protein expression.
4. Conclusions
This study describes some mechanisms of action of Fucus vesiculosus, rich in phlorotannins and peptides on different liver proteins, proving its previously described hypocholesterolemic effect. The proteomic study demonstrated the inhibitory effect of this extract on the expression of four different proteins involved in cholesterol biosynthetic process, resulting in the inhibition of its synthesis. With Western blot and qRT-PCR techniques was observed that this extract has the capacity to decrease the protein expression as well as the mRNA expression of an important cholesterol transporter protein, namely NPC1L1 which results in decrease of biliary excretion of cholesterol as consequently reduce the atherosclerosis risk. Due to inhibitory effect of this extract on cholesterol synthesis and on the expression of the cholesterol transporter NPC1L1, a decrease in the expression of the ABCG5 transporter was also observed in the cells exposed to the extract, since there is less cholesterol to be removed from the liver.
Author Contributions
Conceptualization: R.A. and M.L.S.; writing—original draft preparation: R.A.; writing—review and editing: M.L.S., M.B., C.A., R.P., H.M.S; supervision: M.L.S., M.B., H.M.S.; supervision of Real-time quantitative PCR assay: C.A., M.B.; supervision of proteomic assay: H.M.S., project administration and funding acquisition: M.L.S. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by UIDB/04046/2020 and UIDP/04046/2020 Centre grants from FCT, Portugal (to BioISI); research project PTDC/BIA-BQM/28355/2017; IPL/2019/AMar2Fun. Centro de Química Estrutural is a Research Unit (RU) funded by FCT, projects UIDB/00100/2020 and UIDP/00100/2020. Institute of Molecular Sciences is an Associate Laboratory funded by FCT, project LA/P/0056/2020. R.A. is recipient of a fellowship from BioSys PhD programme PD65-2012 (Ref PD/BD/142861/2018) from FCT (Portugal). PROTEOMASS Scientific Society is acknowledged by the funding provided to the Laboratory for Biological Mass Spectrometry Isabel Moura (#PM001/2019 and #PM003/2016). H.M.S acknowledges the FC/MECTES for his research contract FCT Investigator Program 2015 (IF/00007/2015) and the funding provided by the Associate Laboratory for Green Chemistry LAQV which is financed by national funds from FCT/MCTES (UIDB/50006/2020), (UIDP/50006/2020).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Roth GA, Mensah GA, Johnson CO, Addolorato G, Ammirati E, Baddour LM, et al. Global Burden of Cardiovascular Diseases and Risk Factors, 1990-2019: Update From the GBD 2019 Study. J Am Coll Cardiol 2020;76:2982–3021. [CrossRef]
- Yamamoto H, Yamanashi Y, Takada T, Mu S, Tanaka Y, Komine T, et al. Hepatic expression of Niemann-Pick C1-like 1, a cholesterol reabsorber from bile, exacerbates western diet-induced atherosclerosis in LDL receptor mutant mice S. Mol Pharmacol 2019;96:47–55. [CrossRef]
- Garcia-Gutierrez E, Sayavedra LBT-RM in FS. Diet, Microbiota and the Gut-Brain Axis. Ref. Modul. Food Sci., Elsevier; 2021. [CrossRef]
- Wilkins E., Wilson I., Wickramasinghe K., Bhatnagar P., Leal J., Luengo-Fernandez R., et al. European Cardiovascular Disease Statistics 2017 edition. Eur Hear Network, Brussels 2017:192.
- Taddei C, Zhou B, Bixby H, Carrillo-Larco RM, Danaei G, Jackson RT, et al. Repositioning of the global epicentre of non-optimal cholesterol. Nature 2020;582:73–7. [CrossRef]
- Singh P, Saxena R, Srinivas G, Pande G, Chattopadhyay A. Cholesterol biosynthesis and homeostasis in regulation of the cell cycle. PLoS One 2013;8:e58833. [CrossRef]
- Mitsche MA, McDonald JG, Hobbs HH, Cohen JC. Flux analysis of cholesterol biosynthesis in vivo reveals multiple tissue and cell-type specific pathways. Elife 2015;4:e07999. [CrossRef]
- Jessup W, Gelissen IC, Gaus K, Kritharides L. Roles of ATP binding cassette transporters A1 and G1, scavenger receptor BI and membrane lipid domains in cholesterol export from macrophages. Curr Opin Lipidol 2006;17:247–57. [CrossRef]
- Tang W, Jia L, Ma Y, Xie P, Haywood J, Dawson PA, et al. Ezetimibe restores biliary cholesterol excretion in mice expressing Niemann-Pick C1-Like 1 only in liver. Biochim Biophys Acta 2011;1811:549–55. [CrossRef]
- Huff MW, Pollex RL, Hegele RA. NPC1L1: evolution from pharmacological target to physiological sterol transporter. Arterioscler Thromb Vasc Biol 2006;26:2433–8. [CrossRef]
- Ge L, Wang J, Qi W, Miao H-H, Cao J, Qu Y-X, et al. The cholesterol absorption inhibitor ezetimibe acts by blocking the sterol-induced internalization of NPC1L1. Cell Metab 2008;7:508–19. [CrossRef]
- Landis MN, Adams DR. Drugs for the Skinternist. In: Wolverton SE, editor. Compr. Dermatologic Drug Ther. Fourth Edi, Elsevier; 2021, p. 430-444.e3. [CrossRef]
- Stone NJ, Robinson JG, Lichtenstein AH, Bairey Merz CN, Blum CB, Eckel RH, et al. 2013 ACC/AHA guideline on the treatment of blood cholesterol to reduce atherosclerotic cardiovascular risk in adults: A report of the american college of cardiology/american heart association task force on practice guidelines. Circulation 2014;129. [CrossRef]
- André R, Pacheco R, Bourbon M, Serralheiro ML. Brown Algae Potential as a Functional Food against Hypercholesterolemia: Review. Foods 2021;10:234. [CrossRef]
- Ososki AL, Lohr P, Reiff M, Balick MJ, Kronenberg F, Fugh-Berman A, et al. Ethnobotanical literature survey of medicinal plants in the Dominican Republic used for women’s health conditions. J Ethnopharmacol 2002;79:285–98. [CrossRef]
- Geukens K, Wijnhoven L. Assessment report on Fucus vesiculosus L ., thallus Herbal preparations in solid dosage form for oral use. vol. 44. 2014.
- Skibola CF. The effect of Fucus vesiculosus, an edible brown seaweed, upon menstrual cycle length and hormonal status in three pre-menopausal women: a case report. BMC Complement Altern Med 2004;4:10. [CrossRef]
- Romm A, Hardy ML, Mills S. Endocrine Disorders and Adrenal Support. Bot. Med. Women’s Heal. Second edi, St. Louis (Missouri: Elsevier; 2017, p. 186–210.
- Yoon NY, Kim HR, Chung HY, Choi JS. Anti-hyperlipidemic effect of an edible brown algae, Ecklonia stolonifera, and its constituents on poloxamer 407-induced hyperlipidemic and cholesterol-fed rats. Arch Pharm Res 2008;31:1564–71. [CrossRef]
- Shin H-C, Kim SH, Park Y, Lee BH, Hwang HJ. Effects of 12-week oral supplementation of Ecklonia cava polyphenols on anthropometric and blood lipid parameters in overweight Korean individuals: a double-blind randomized clinical trial. Phytother Res 2012;26:363–8. [CrossRef]
- Yeo A-R, Lee J, Tae IH, Park S-R, Cho YH, Lee BH, et al. Anti-hyperlipidemic Effect of Polyphenol Extract (Seapolynol(TM)) and Dieckol Isolated from Ecklonia cava in in vivo and in vitro Models. Prev Nutr Food Sci 2012;17:1–7. [CrossRef]
- André R, Guedes L, Melo R, Ascensão L, Pacheco R, Vaz PD, et al. Effect of food preparations on in vitro bioactivities and chemical components of fucus vesiculosus. Foods 2020;9:1–20. [CrossRef]
- André R, Guedes R, López J, Serralheiro ML. Untargeted metabolomic study of HepG2 cells under the effect of Fucus vesiculosus aqueous extract. Rapid Commun Mass Spectrom 2021;35. [CrossRef]
- Oliveira E, Araújo JE, Gómez-Meire S, Lodeiro C, Perez-Melon C, Iglesias-Lamas E, et al. Proteomics analysis of the peritoneal dialysate effluent reveals the presence of calciumregulation proteins and acute inflammatory response. Clin Proteomics 2014;11:1–8. [CrossRef]
- Jorge S, Capelo JL, Laframboise W, Dhir R, Lodeiro C, Santos HM. Development of a Robust Ultrasonic-Based Sample Treatment to Unravel the Proteome of OCT-Embedded Solid Tumor Biopsies. J Proteome Res 2019;18:2979–86. [CrossRef]
- Cox J, Mann M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat Biotechnol 2008;26:1367–72. [CrossRef]
- Cox J, Neuhauser N, Michalski A, Scheltema RA, Olsen J V, Mann M. Andromeda: A Peptide Search Engine Integrated into the MaxQuant Environment. J Proteome Res 2011;10:1794–805. [CrossRef]
- Tyanova S, Cox J. Perseus: A Bioinformatics Platform for Integrative Analysis of Proteomics Data in Cancer Research. In: von Stechow L, editor. Methods Mol. Biol. 2018, New York, NY: Springer New York; 2018, p. 133–48. [CrossRef]
- Bindea G, Mlecnik B, Hackl H, Charoentong P, Tosolini M, Kirilovsky A, et al. ClueGO: a Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009;25:1091–3. [CrossRef]
- Ressaissi A, Attia N, Pacheco R, Falé PL, Serralheiro MLM. Cholesterol transporter proteins in HepG2 cells can be modulated by phenolic compounds present in Opuntia ficus-indica aqueous solutions. J Funct Foods 2020;64:103674. [CrossRef]
- Zou J, Feng D. Lycopene reduces cholesterol absorption through the downregulation of Niemann-Pick C1-like 1 in Caco-2 cells. Mol Nutr Food Res 2015;59:2225–30. [CrossRef]
- Falé PL, Amaral F, Amorim Madeira PJ, Sousa Silva M, Florêncio MH, Frazão FN, et al. Acetylcholinesterase inhibition, antioxidant activity and toxicity of Peumus boldus water extracts on HeLa and Caco-2 cell lines. Food Chem Toxicol an Int J Publ Br Ind Biol Res Assoc 2012;50:2656–62. [CrossRef]
- Xie P, Jia L, Ma Y, Ou J, Miao H, Wang N, et al. Ezetimibe inhibits hepatic Niemann-Pick C1-Like 1 to facilitate macrophage reverse cholesterol transport in mice. Arterioscler Thromb Vasc Biol 2013;33:920–5. [CrossRef]
- Kawase A, Hata S, Takagi M, Iwaki M. Pravastatin modulate niemann-pick C1-like 1 and ATP-binding cassette G5 and G8 to influence intestinal cholesterol absorption. J Pharm Pharm Sci 2015;18:765–72. [CrossRef]
- Tremblay AJ, Lamarche B, Lemelin V, Hoos L, Benjannet S, Seidah NG, et al. Atorvastatin increases intestinal expression of NPC1L1 in hyperlipidemic men. J Lipid Res 2011;52:558–65. [CrossRef]
- Tang W, Ma Y, Yu L. Plasma cholesterol is hyperresponsive to statin in ABCG5/ABCG8 transgenic mice. Hepatology 2006;44:1259–66. [CrossRef]
- El-Darzi N, Astafev A, Mast N, Saadane A, Lam M, Pikuleva IA. N,N-Dimethyl-3β-hydroxycholenamide Reduces Retinal Cholesterol via Partial Inhibition of Retinal Cholesterol Biosynthesis Rather Than its Liver X Receptor Transcriptional Activity . Front Pharmacol 2018;9:827.
- Jokela H, Rantakari P, Lamminen T, Strauss L, Ola R, Mutka A-L, et al. Hydroxysteroid (17beta) dehydrogenase 7 activity is essential for fetal de novo cholesterol synthesis and for neuroectodermal survival and cardiovascular differentiation in early mouse embryos. Endocrinology 2010;151:1884–92. [CrossRef]
- Wang Y, Rogers PM, Su C, Varga G, Stayrook KR, Burris TP. Regulation of cholesterologenesis by the oxysterol receptor, LXRalpha. J Biol Chem 2008;283:26332–9. [CrossRef]
- Song B-L, Javitt NB, DeBose-Boyd RA. Insig-mediated degradation of HMG CoA reductase stimulated by lanosterol, an intermediate in the synthesis of cholesterol. Cell Metab 2005;1:179–89. [CrossRef]
- Ercal B, Crawford PA. Ketone Body Metabolism in the Neonate. In: Polin RA, Abman SH, Rowitch DH, Benitz WE, Fox WWBT-F and NP (Fifth E, editors. Fetal Neonatal Physiol. 5th ed., Elsevier; 2017, p. 370–9. [CrossRef]
Figure 1.
Effect of F. vesiculosus aqueous extract on soluble membrane proteins of HepG2. A - Gel obtained from SDS-PAGE of the soluble membrane protein fraction of HepG2 cells under the effect of: (Cont), cell culture media and (Ext), 0.25 mg/mL of F. vesiculosus extract; (M), marker proteins. B - Heat map generated from SDS-PAGE gel analyse with Image J software reflecting the protein band intensity of soluble membrane protein fraction of HepG2 cells exposed to (Cont) cell culture media and (Ext) F. vesiculosus extract. Cells samples are arranged in columns, protein band intensity, and estimated molecular weight in rows. Dark blue shades correspond to a high-intensity protein band; light blue shades correspond to a low-intensity protein band.
Figure 1.
Effect of F. vesiculosus aqueous extract on soluble membrane proteins of HepG2. A - Gel obtained from SDS-PAGE of the soluble membrane protein fraction of HepG2 cells under the effect of: (Cont), cell culture media and (Ext), 0.25 mg/mL of F. vesiculosus extract; (M), marker proteins. B - Heat map generated from SDS-PAGE gel analyse with Image J software reflecting the protein band intensity of soluble membrane protein fraction of HepG2 cells exposed to (Cont) cell culture media and (Ext) F. vesiculosus extract. Cells samples are arranged in columns, protein band intensity, and estimated molecular weight in rows. Dark blue shades correspond to a high-intensity protein band; light blue shades correspond to a low-intensity protein band.
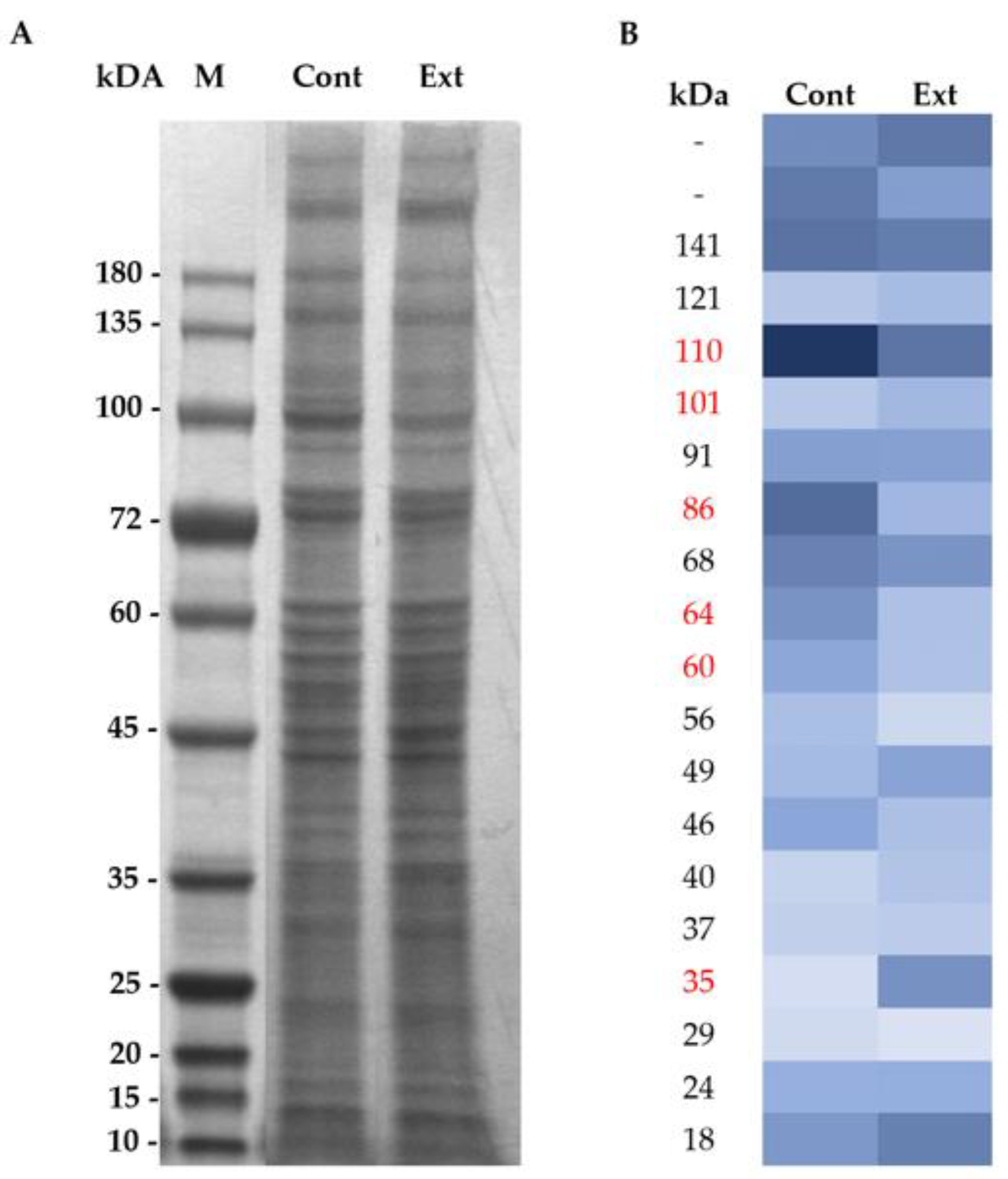
Figure 2.
Venn diagram showing the differentially expressed proteins from comparative proteomic analysis of HepG2 cells control and cells exposed to 0.25 mg/mL of F. vesiculosus (http://bioinformatics.psb.ugent.be/webtools/Venn/).
Figure 2.
Venn diagram showing the differentially expressed proteins from comparative proteomic analysis of HepG2 cells control and cells exposed to 0.25 mg/mL of F. vesiculosus (http://bioinformatics.psb.ugent.be/webtools/Venn/).

Figure 3.
(A) - Networking of ClueGO analysis of significant enrichment GO biological process and Molecular function (Term P-value corrected with Bonferroni step down, p< 0.05), where are represented the term enrichment of proteins from HepG2 control cells (red),proteins from HepG2 cells exposed to F. vesiculosus (blue), and terms from both clusters (grey). (B) - Bar chart representing the percentage of gene by term of GO Biological process and GO molecular function from HepG2 control cells and HepG2 exposed to F. vesiculosus (C).
Figure 3.
(A) - Networking of ClueGO analysis of significant enrichment GO biological process and Molecular function (Term P-value corrected with Bonferroni step down, p< 0.05), where are represented the term enrichment of proteins from HepG2 control cells (red),proteins from HepG2 cells exposed to F. vesiculosus (blue), and terms from both clusters (grey). (B) - Bar chart representing the percentage of gene by term of GO Biological process and GO molecular function from HepG2 control cells and HepG2 exposed to F. vesiculosus (C).
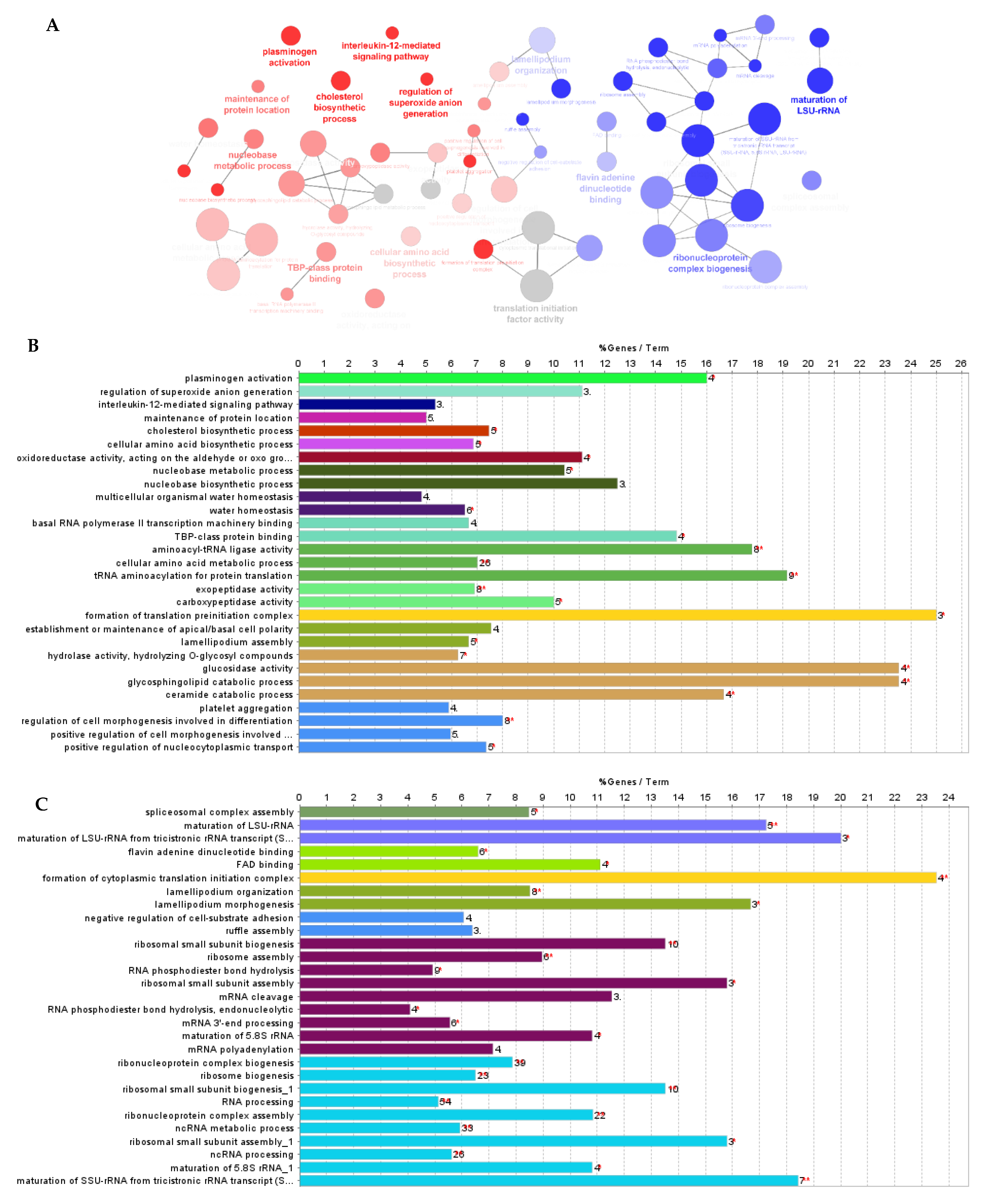
Figure 4.
– (A) - Networking of ClueGO significant enrichment pathways analysis from KEEG and Reactome (Term P-value corrected with Bonferroni step down, p< 0.05), where are represented the terms enrichment of proteins from HepG2 control cells (red), HepG2 cells exposed to F. vesiculosus (blue) and terms from both clusters (grey). (B) - Bar chart representing the percentage of gene by term of each enriched pathway from HepG2 control cells (B) and HepG2 exposed to F. vesiculosus (C).
Figure 4.
– (A) - Networking of ClueGO significant enrichment pathways analysis from KEEG and Reactome (Term P-value corrected with Bonferroni step down, p< 0.05), where are represented the terms enrichment of proteins from HepG2 control cells (red), HepG2 cells exposed to F. vesiculosus (blue) and terms from both clusters (grey). (B) - Bar chart representing the percentage of gene by term of each enriched pathway from HepG2 control cells (B) and HepG2 exposed to F. vesiculosus (C).
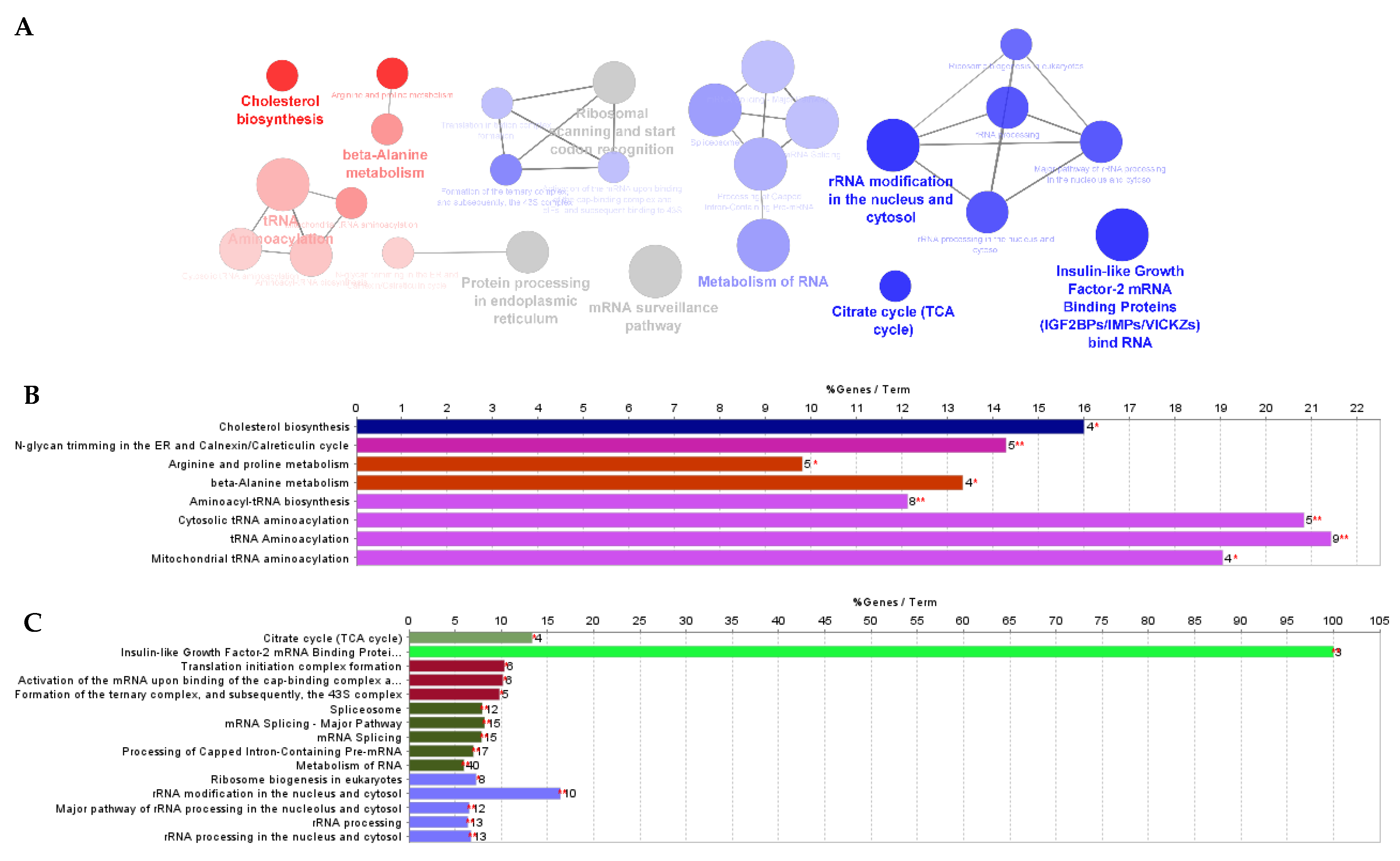
Figure 5.
Cholesterol synthesis pathway. The pathway proteins that proteomic analysis revealed to be present just in HepG2 control cells and not in HepG2 cells exposed to the extract are highlighted in red.
Figure 5.
Cholesterol synthesis pathway. The pathway proteins that proteomic analysis revealed to be present just in HepG2 control cells and not in HepG2 cells exposed to the extract are highlighted in red.
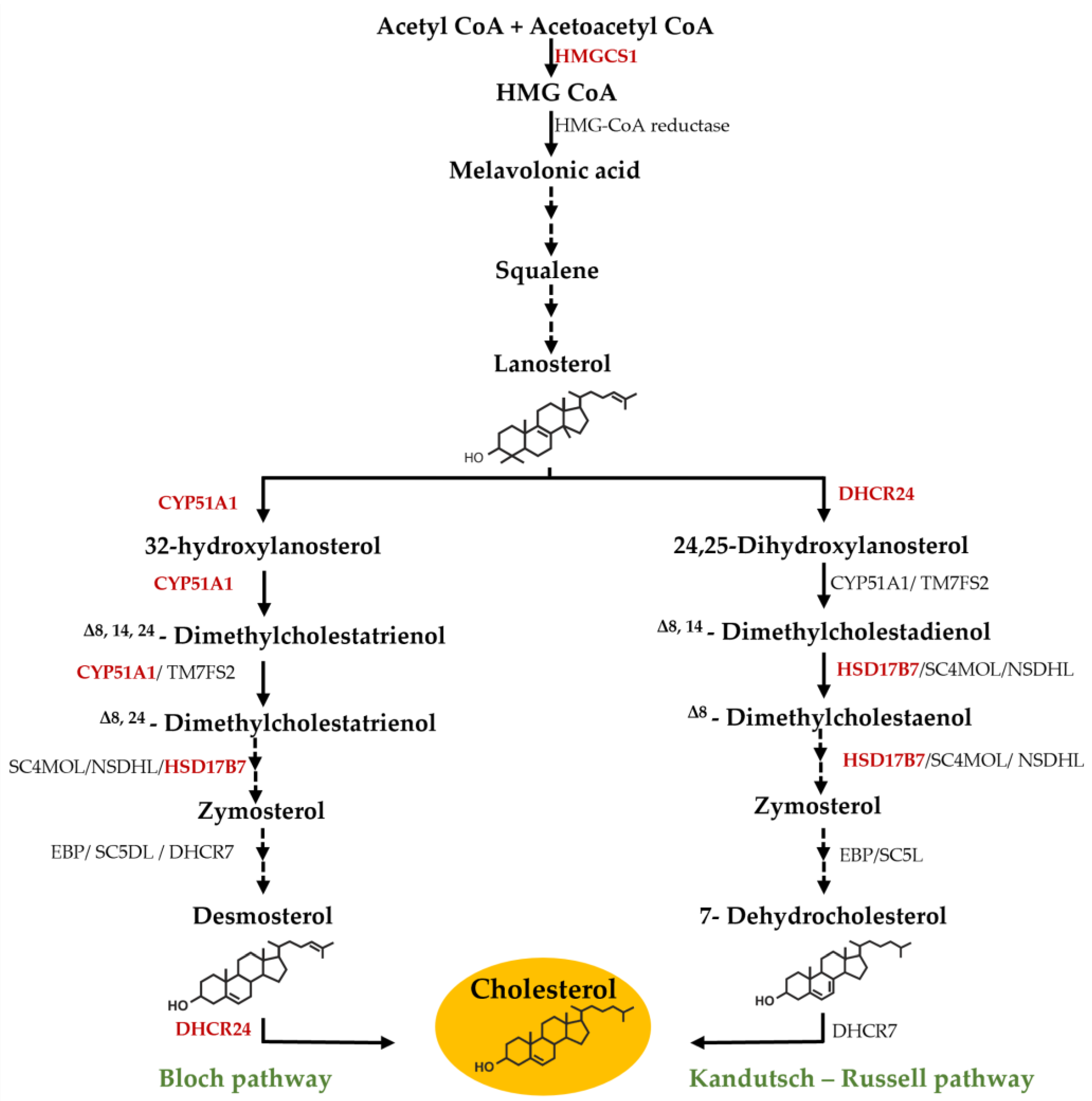
Figure 6.
(A) Western blot results representing the effect of F. vesiculosus aqueous extract (0.25 mg/mL) on the protein expression of NPC1L1 and ABCG5 in HepG2 cells using the same weight of cells (0.025 mg of cell/ mL of lysis buffer), (B) relative NPC1L1 mRNA expression in HepG2 under the effect of F. vesiculosus aqueous extract (0.25 mg/mL) and ezetimibe (100 µM) determined by qRT-PCR (normalised to GAPDH). Different superscript letters (a–b) correspond to values of relative expression for each gene that can be considered statistically different (p ≤ 0.05).
Figure 6.
(A) Western blot results representing the effect of F. vesiculosus aqueous extract (0.25 mg/mL) on the protein expression of NPC1L1 and ABCG5 in HepG2 cells using the same weight of cells (0.025 mg of cell/ mL of lysis buffer), (B) relative NPC1L1 mRNA expression in HepG2 under the effect of F. vesiculosus aqueous extract (0.25 mg/mL) and ezetimibe (100 µM) determined by qRT-PCR (normalised to GAPDH). Different superscript letters (a–b) correspond to values of relative expression for each gene that can be considered statistically different (p ≤ 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



