
Submitted:
31 May 2023
Posted:
01 June 2023
You are already at the latest version
Abstract
APE1 is an essential endodeoxyribonuclease of the base excision repair pathway that maintains genome stability. It was identified as a pivotal factor favoring tumor progression and chemo-resistance through the control of gene expression by a redox-based mechanism. APE1 is overex-pressed and serum-secreted in different cancers, representing a prognostic and predictive factor, and a promising non-invasive biomarker. Strategies directly targeting APE1 funtions led to the identification of inhibitors showing a potential therapeutic value, which some of them are cur-rently in clinical trials. Interestingly, evidence indicates novel roles of APE1 in RNA metabolism still not fully understood, including its activity in processing damaged RNA in chemoresistant phenotypes, regulating oncomiRs maturation, and oxidized-RNA decay. Recent data point out a control role of APE1 in the expression and sorting of oncomiRs, within secreted extracellular vesicles (EVs). This review is focused on giving a portrait of the pros and cons of the last two decades of research aiming at the identification of inhibitors of the redox or the DNA-repair functions of APE1 for the definition of novel targeted therapies for cancer. We will discuss the new perspectives in cancer therapy coming from the unexpected finding of the APE1 role in miRNA processing for personalized therapy.
Keywords:
APE1
; inhibitors
; cancer.
1. A brief introduction to APE1 biology and different functions
The acronym APE1/Ref1 (or more simply APE1) stands for apurinic/apyrimidinic (AP) endondeoxyribonuclease-reduction/oxidation 1, which is a well-known protein with multifunctional roles ranging from the endodeoxyribonuclease activity on DNA and RNA to the hub role in several reduction/oxidation (redox) signaling pathways [1,2,3].
Historically, APE1 has been largely known for its function during the base excision repair (BER) pathway [4], in which non-bulky DNA lesions are repaired. During the BER, APE1 works as AP- endonuclease, being able to cleave abasic sites that are generated spontaneously or by the action of specific glycosylases. The single strand break generated by APE1 cleavage is then brought to complete repair by other downstream BER enzymes (i.e. Polβ, XRCC1, FEN1, Ligase III). Another main cellular role of APE1 is working as a redox hub for several transcription factors (TFs). Indeed, the reduction of some TFs (i.e. NF-κB, p53, Hif-1α, AP-1, Pax-5/8, etc.) by APE1 allows their activation and consequently the initiation of the transcription of specific genes (i.e. IL-8, SIRT-1, VEGF, etc.). An additional role of APE1 in transcriptional regulation is due to its capacity to stabilize G-quadruplex (G4) which are stable conformational structures in the G-rich DNA portion of certain human promoters [5,6,7]. Although, following oxidative stress, the newly formed 8-oxoguanine (8-oxoG) can stall the transcription due to its destabilizing effect, there is evidence that its presence in some promoters induces the formation of BER-stabilized G4 that enhances gene expression [8,9]. In this context, the binding of APE1 to G4 sequences promotes G4 folding. Moreover, the acetylation of APE1 enhances its residence time on DNA and stabilizes G4 structures in cells [10]. In this way, APE1 facilitates transcription factor loading to the promoter modulating the gene expression [11]. Finally, attention will be paved to recently characterized APE1 functions, especially those involved in RNA metabolism [12,13,14,15,16]. Specifically, APE1 has resulted able to cleave abasic RNA [13] and damaged ribonucleotides embedded in DNA [15,16], discovering it as an efficient ribonuclease. Furthermore, only in the last years, concurrent with its novel and unsuspected involvement in RNA-metabolism, it has been described that APE1 can be present in subcellular condensates formed through liquid-liquid phase separation mechanisms (LLPS) [17,18,19] and can be secreted by tumoral cells through extracellular vesicles (EVs) [20].
These intriguing roles of APE1 have been discovered in three decades of constant interest in this protein, delineating them in physiological and pathological contexts and making this protein an attractive therapeutic target for several pathologies, including cancer [21]. Indeed, targeting APE1, after more than 20 years of attempts, still represents an important challenge in cancer therapy. In this review, we will focus on the dysregulation of APE1 in cancer and then we will describe known inhibitors of the principal functions of APE1, paving the way for novel functions involved in chemoresistance and potentially used as new therapeutic targets.
2. APE1 and cancer: a focus on polymorphisms and tissue expression
As previously discussed, being entailed in such focal cellular processes, APE1 dysregulation has a great impact on pathologies like cancer, making it an attractive therapeutical target [2,22]. APE1 dysregulation is involved, at three different levels, in tumour development, as it may concern alterations to its genetic sequence, to its expression, or to its localization [2].
Several studies pointed out the important role of single nucleotide polymorphisms (SNPs), present in APE1 gene, in cancer pathology [23] (Figure 1). The most common and studied APE1 variant is Asp148Glu (D148E), which is present in 48,5% of the population [24]. X-ray crystallography experiments showed that this variant lacks significant structural changes and its considered benign [25]. As a matter of fact, the protein bearing this SNP holds normal AP endonuclease and DNA binding properties, but its 3’-RNA phosphatase and endoribonuclease activities are somehow affected [14,26]. The role of this polymorphism in cancer is controversial, due to several conflicting data in literature [24]. This common variant has been widely studied since more than a hundred publications can be found regarding this topic. Numerous studies and meta-analysis associated D148E variant with an increased risk of different cancers, while others reported the opposite pattern even in the same tumour type [2,24,27]. Another APE1 polymorphic variant associated with cancer and biochemically studied is Arg237Cys (R237C) [28,29]. This substitution is associated with endometrial cancer [28,29] and it has greater consequences on the functional activity of the whole protein [25]. Indeed, the protein X-ray crystal structure revealed that this aminoacidic variation causes significant shifts in adjacent DNA binding residues, leading to a great diminution (3-fold) of APE1 DNA binding ability [25]. Remarkably, this polymorphic variant has a reduced ability to associate with its BER partners, such as polymerase β (Pol β) and XRCC1 [23]. Indeed, the X-ray structure shows that also lysine 244 (K244) changes its position [25], which has been implicated in APE1-Pol β protein-protein interaction [30]. It has also been reported that R237C variant has a reduced AP-endonuclease activity [23], reduced 3’->5’ exonuclease and 3’-damage excision activities [29] and it has a reduced incision capacity close to nucleosomes [31]. One more endometrial tumour-associated variant is Pro112Leu (P112L) [28], which had comparable AP-endonuclease activity to the wild type form [29].
In this review, we performed an analysis on cBioPortal to find APEX1 SNPs or insertion/deletions (IN/DELs) related to different cancer types, considering a curated selection of non-redundant studies (213 studies selected, 69223 samples, 65853 patients) (https://bit.ly/3M9OYa7) [32,33]. The somatic mutation frequency of APEX1 was 0.2% with 108 unique variants. None of the variants detected was a driver mutation for cancer and most of them were sporadic. The most frequent variants were R193C/H, D148E, and H289Y/Q. Interestingly, even if R193C and R193H variants were detected to a comparable extent to D148E in the selected cohort of studies, there is no work in the literature that focuses on this mutation and on its functional impact. According to Mutation Assessor tool [34,35], theoretically, R193H has a low impact on the protein functional activity, while R193C might have a worse impact. There are no functional studies even about H289Y/Q variants, which are predicted to have a neutral impact on protein activity. In Table 1, we report every variant found with the cBioPortal tool for each tumour type.
APE1 overexpression and altered localization are prominent features of several different tumours and most of the time they have been associated with poor prognosis and malignant phenotypes. We further provide a description of how APE1 is altered in cancer and how this impacts tumorigenesis (Table 2).
In bladder cancer (BCa), several studies detected high expression levels of APE1 protein in tumour tissues compared to normal adjacent tissues and they were associated with poor outcomes [37,39,42]. APE1 overexpression was also linked to lympho-vascular invasion features, as high VEGFA levels and infiltration of CD163+ tumour associated macrophages (TAMs) [40]. Nuclei-cytosol distribution of APE1 was variable between high- and low-grade tumours: indeed, low grade cancers displayed increased APE1 levels only in the nucleus, while high grade invasive tumours showed increased positive staining even in the cytoplasm [37]. APE1 is also a promising diagnostic biomarker in BCa, as its levels in serum and urines were increased in contrast to normal healthy controls and they were associated with tumour grade and stage, recurrence, and invasion [36,38,41]. Interestingly, a study observed an increased secretory activity of D148E in bladder cancers, which contributed to increased serum levels of the protein in patients [127].
Concerning hepatocellular carcinoma (HCC), APE1 is upregulated at both transcriptional and translational levels compared to normal liver tissues [79,80,81]. Moreover, the mRNA content increased with tumour progression and it is higher in less differentiated and more aggressive tumours [79]. Patients with greater APE1 protein levels exhibited unfavourable prognosis and lower overall survival [80,81]. Interestingly, both APE1 truncated forms, missing the first 33 residues (N∆33 - 35 kDa) and APE1 full length (37 kDa) were detected in HCC tissue samples and HCC cell lines [82]. APE1 cellular distribution was altered in HCC: while APE1 staining was only nuclear in normal liver tissues, in tumour tissues there was also a significant fraction of the protein in the cytoplasm [83]. Cytoplasmatic APE1 was about three times higher in poorly differentiated tumours and it was associated with reduced overall survival [83]. Noteworthy, the cytoplasmatic staining was associated with APE1 mitochondrial accumulation in grade 1 and grade 2 HCC tumours, while in grade 3 tumours it was not [84]. Even in HCC, APE1 serum levels can be used as a novel diagnostic biomarker [82].
APE1 protein resulted overexpressed also in pancreatic adenocarcinoma (PDAC) tissues and cell lines and it was associated with tumour aggressiveness and poor survival [120,121,122]. The proteolytic form of APE1 N∆33 has been detected even in PDAC tissues, with different abundance versus adjacent non-tumour tissue [53]. Interestingly, the acetylated form of APE1 (acAPE1) was overexpressed in PDAC tumours, while it was almost undetectable in healthy pancreatic tissues [53,85]. acAPE1 holds an increased AP-endonuclease activity, which has been proposed as a cancer mechanism to overcome chemotherapy genotoxic stress and uphold proliferation [85]. APE1 localization in PDAC was mainly nuclear and it was similar between primary tumours and their metastasis [121]. Cytosolic localization was observed only in tumour advanced stages and never in the absence of nuclear localization, while its complete absence was associated with invasion and poor differentiation [99,121].
Concerning prostate cancer (PCa), APE1 protein levels were upregulated compared to normal tissue and benign hypertrophy (BPH) [123,124]. Moreover, higher APE1 levels were observed in tumours bearing TMPRSS2:ERG fusion [124]. APE1 localization was only nuclear in normal prostate tissue and non-cancerous prostate cell lines, while there was an increase in the cytoplasmatic fraction in tumour tissues and tumoral cell lines [123].
APE1 overexpression occurs also in oesophageal carcinomas, like oesophageal adenocarcinoma (EAC) [100,101,102] and oesophageal squamous cell carcinoma (ESCC) [103,104], probably as a mechanism adopted by cancer cells to survive the genotoxic effects of bile reflux [103,104]. APE1 localization was mainly nuclear and it was also associated with worse overall survival in patients receiving platinum chemotherapy [99].
An in silico analysis identified APE1 as a central hub gene for gastric cancer, as its overexpression had a great prognostic value in two analysed datasets (GSE1611533, GSE54129) [62]. Indeed, APE1 was overexpressed at both transcriptional and protein level in gastric cancer [63,64]. APE1 staining was weak in normal non-cancerous gastric tissues, while it was both nuclear and cytoplasmatic in tumour tissues [64]. High levels of APE1 were also correlated with invasion and poor prognosis [64], as its serum levels are a valuable diagnostic biomarker for lymph node metastases prediction [65].
APE1 was also upregulated in salivary gland carcinomas and its levels increased depending on malignant transformation of the tumour [125]. APE1 overexpression was higher in smaller tumours displaying lymph node metastasis and invasive growth [125,126]. APE1 localization was mainly nuclear in every salivary gland tumour subtype, except for adenoid cystic carcinomas in which it showed both nuclear and cytoplasmatic localization [125,126].
Furthermore, the overexpression of APE1 protein and mRNA levels was reported also in non-small-cell lung cancers (NSCLCs) [86,87,88,89]. High APE1 expression was associated with poor prognosis, invasion, and chemoresistance, as its levels increased after treatment with platinum compounds [88]. Moreover, high post treatment APE1 serum levels were correlated with poorer overall survival [90]. Nuclear APE1 staining was associated with favourable patient outcomes [91], while a higher cytoplasmatic localization correlated with poor survival and shorter relapse-free survival (RFS) [92,93,94]. Both full-length and truncated forms were found in lung cancer and interestingly APE1 was prevalently truncated at the N-terminus in NSCLC adjacent non-tumour tissues [53]. AcAPE1 was overexpressed in NSCLC tumours, with a strictly nuclear localization [53,85].
Several studies identified overexpression of APE1 protein in ovarian cancers, which has been associated with advanced tumour stages and decreased overall survival [99,112,113]. Moreover, patients with high levels of APE1 showed more frequent resistance to platinum therapy [99,112,114]. The protein-protein interaction between APE1 and nucleophosmin 1 (NPM1) has been extensively examined in ovarian cancer, as the levels of the two proteins were positively correlated and associated with tumour aggressiveness, malignant phenotype, lymph node metastasis and poor chemosensitivity [112,115]. It has been shown that compounds which impair this interaction can add a synergistic effect to traditional chemotherapeutic molecules [116] APE1 localization seemed to be heterogenous in ovarian cancers, depending on stage and histological subtype [22]. Some studies showed prominent cytoplasmatic staining, which increased from well to poorly differentiated cancers and it was higher in advanced stages tumours [114,115,117,118]. In non-responding to cisplatin cases, the observed APE1 overexpression was mainly at cytoplasmatic level, a feature that was observed also in cisplatin resistant cell-lines [114]. Interestingly, almost 90% of patients with abnormal levels of cytosolic APE1 displayed an abnormal distribution of NPM1 [115]. Additionally, cytosolic APE1 can be considered an independent predictive factor for poor progression free survival and for poor overall survival in ovarian cancers [117]. Other works showed prominent nuclear APE1 staining, which increased during tumorigenesis and was associated with survival time [113,119]. Other studies showed an increase of APE1 in both compartments but with higher nuclear staining, which was associated with cancer aggressiveness, lower debulking after surgery, platinum resistance and lower overall survival [99,114].
For what concerns breast cancer, different studies reported conflicting results about APE1 protein expression. Some works described APE1 overexpression in breast cancers, which was mostly nuclear and correlated with malignant phenotypes and unfavourable prognosis [43,44,45]. Contrary to the pattern of increased acetylation observed in other cancer types [53,85], APE1 acetylation resulted in heterogeneous and deregulated in breast cancer [46]. Even in this case, the functional interaction between APE1 and NPM1 in promoting platinum resistance has been described [47]. In contrast to these findings, another study showed that lower levels of APE1 were associated with tumour aggressiveness and triple negative phenotype [48]. Interestingly, another study showed that, within the Ki-67 low-level expression group, lower levels of APE1 were associated with poor overall survival [44].
APE1 protein levels were upregulated even in cervical tumours and they were associated with Epithelial-to-Mesenchymal transition (EMT), lymph node metastasis and poor radiosensitivity [49,50,51,52]. APE1 localization was widely heterogeneous between cervical tumours, showing mainly nuclear staining [50]. Remarkably, there was a significant difference in subcellular localization of APE1 between radiotherapy non-responding and responding tumour cell lines: indeed, radioresistant cervical tumours cell lines showed higher levels of the cytoplasmatic fraction and lower levels of the nuclear protein, suggesting a role of cytosolic APE1 in radio-resistance promotion [51].
Several studies described an overexpression of APE1 in colorectal cancers (CRC), observing also a gradual increase of its expression during tumour progression [54,55,56,57]. APE1 levels resulted high also in liver metastasis of CRC [58]. APE1 localization in CRC was both nuclear and cytoplasmatic, with most cells displaying mixed or exclusive cytoplasmatic localization [54]. Even in CRC tumour samples and cell lines, nuclear acAPE1 was overexpressed [53,59] and it positively correlated with resistance to 5-Fluorouracil (5-FU) [59]. Moreover, both full-length and truncated forms were detected in colon cancer [53]. Interestingly, the levels of serum APE1 autoantibodies are valuable diagnostic biomarkers for CRC [60].
Regarding glioma, conflicting data are available. Some studies described an overexpression of APE1 in tumoral tissues compared to healthy ones [66,67], with a 13-fold increase of AP-endonuclease activity in 93% of tumours [66]. Glioma radioresistant cell lines displayed higher levels of APE1 compared to responding cell lines [68]. Indeed, an increase in APE1 expression was observed in patients after treatment and recurrence [69]. On the other hand, different studies evidenced low mRNA and protein expression in adult high-grade gliomas, associated with poor overall survival [70,71]. Moreover, APE1 localization in gliomas was predominantly nuclear [70].
For what concerns melanoma, several studies identified APE1 overexpression at both transcriptional and translational levels [95,96,97]. Indeed, APE1 was overexpressed in melanoma cancer cell lines and in clinical samples, showing a prominent nuclear localization in both cases [96,97]. High mRNA levels were associated with vascular invasion, high proliferation rates, poor relapse free survival, and overall survival [95,98]. Patients with higher levels of APE1 also showed a lower response to therapy [98].
APE1 was overexpressed also in another skin tumour, namely cutaneous squamous cell carcinoma (cSCC) [61]. APE1 upregulation in cSCC was associated with increased proliferation and migration by EMT [61]. APE1 was dysregulated in several Head and Neck squamous cell carcinomas (HNSCC). In oral SCC (oSCC), APE1 was overexpressed at protein level and its high expression was significantly correlated with nodal status, shorter overall survival and disease-free survival [72,73]. APE1 localization in oSCC was mainly nuclear, but a translocation to cytoplasm was observed after cisplatin treatment [72,74]. In oSCC, APE1 serum levels are a promising diagnostic biomarker [75]. Indeed, high levels of serum APE1 (sAPE1) are associated with late TNM stages, lymph node metastasis and worse pathological differentiation [75]. Patients with lower levels of sAPE1 went through longer disease-free survival after post-surgery cisplatin therapy and longer overall survival [75]. APE1 overexpression was observed also in laryngeal SCC (LSCC) [76]. APE1 levels resulted upregulated even in sinonasal SCC (sSCC) and SCC with inverted papilloma (SCCwIP), with a vivid nuclear localization associated with metastatization [77]. sSCC tumours showed higher cytoplasmatic staining compared to SCCwIP [77]. Moreover, exclusive cytoplasmatic staining was associated with higher T-stage and histological grade [77]. Lastly, APE1 overexpression characterised also lip SCC (lSCC), with a strong nuclear localization [78].
Furthermore, APE1 levels were upregulated in osteosarcoma and they were associated with poor prognosis and cisplatin resistance [105,106,107,108,109,110,111]. APE1 localization was both nuclear and cytoplasmatic [105,110]. Patients with higher levels of the protein in the cytoplasmatic fraction were less responding to cisplatin treatment and experienced recurrence and metastasis [105].
Therefore, in general, APE1 is significantly overexpressed in different kinds of cancers and subcellular distribution may significantly change depending on the specific tissue and tumoral stage.
3. APE1 as a promising therapeutic target
In the last decades, APE1 has emerged as a promising therapeutic target in cancer, either for its role in DNA repair or for its redox activities. In the next paragraphs, we will dive deeper into these functions of APE1, highlighting the study progression around the discovery of specific inhibitors, principally employed in chemotherapy (Table 3). Last but not least, we will discuss the new roles of APE1 in RNA metabolism and in cell signaling through its secretion, hypothesizing these novel functions as new promising targets in cancer therapy.
3.1. Targeting the APE1 endonuclease activity
The endonuclease function of APE1 is essential for its activity in the BER pathway. This role of APE1 depends mainly on its C-terminal region: the three most important residues involved in this activity are E96, which is implied in the coordination of divalent metals, D210 and H309, that are required for the hydrolysis reaction [128] (Figure 1). Other important residues that mediate different functions can be found in the C-terminal region: indeed, D70 is involved in the 3’-phosphodiesterase activity [129], K98 is required in the Nucleotide Incision Repair (NIR) [130] and F266 is implicated in the 3’ to 5’-exonuclease activity [131].
Previous studies identified different compounds that inhibit endonuclease APE1 activity, in vitro and in human cells, as summarized in different reviews [132,133,134,135]. Various groups, over the years, worked extensively towards the identification of potent small-molecule inhibitors able to target the DNA repair function of APE1, in combination studies, with the rationale that the blockade of APE1 endonuclease activity might have various therapeutic applications, particularly in cancer treatment, by sensitizing cancer cells to DNA-damaging agents and leading to tumor cell death. Although many studies support the inhibition of APE1 as a means of complementing current chemotherapeutic regimens and, accordingly, various chemical inhibitors have been developed, to date, a clinical candidate has yet to be realized. Indeed, despite their high in vitro activity, the toxicity and the selectivity of the majority of the reported endonuclease inhibitors remain to be established. In this section, we attempt to present the major APE1 inhibitors identified thus far and discuss their activity. It is not our scope to revisit all the inhibitors in depth; a comprehensive list of APE1 endonuclease inhibitors is reviewed in [128,135,136]. The published approaches utilized for the development of APE1 endonuclease inhibitors can be mainly categorized into (i) screening of commercially available compounds that were synthesized for targeting other molecules; (ii) computational screening, and (iii) pharmacophore modeling. One of the first studied molecules impairing APE1 repair activity was Methoxyamine (MX), an alkoxyamine derivative that reacts to form an imine with the aldehyde group in the ring-open form of the abasic lesion thereby indirectly blocking APE1 endonuclease activity [137,138]. Since MX was demonstrated to enhance the cytotoxicity effect of alkylating agents such as temozolomide (TMZ) in a wide variety of cancer cell lines both in vitro and in xenografts models [139,140], it advanced to clinical trials; however, to date, clinical studies have not shown success.
A second compound that was first identified as a radiosensitizer of HeLa cells [141] and subsequently reported to be an inhibitor of APE1 by Luo and Kelly in 2004 [142], is Lucanthone or Miracil D. Lucanthone was shown to enhance the cell-killing effect of TMZ and an alkylating agent such as methyl methanesulfonate (MMS) in culture and was further characterized by Naidu and colleagues to bind to the hydrophobic pocket site of APE1 [143]. No other studies were reported successively but its specificity is still debated because part of its inhibitory effect is mediated by its ability to intercalate into DNA and through the inhibition of topoisomerase II and possibly other cellular proteins [21].
Afterward, numerous laboratories relied on high throughput screens (HTS) mainly based on fluorescence assay to identify inhibitors of APE1 endonuclease activity. In general, the identification of the potential hits was followed by different assays aiming to prove specificity and selectivity for APE1 inhibition including AP site incision assay, the ability of the compound to bind DNA per se, and the ability to enhance the cytotoxicity of alkylating agents (i.e. TMZ, MMS). It is worth mentioning that the inhibitors reported so far showed affinities in the µM range, that are not compatible with a suitable pharmaceutical agent and more importantly, none of them have been demonstrated to have utility in pre-clinical animal cancer models. CRT0044876 (7-nitroindole- 2-carboxylic acid) is the first biochemical and biological reported APE1 inhibitor identified through a fluorescein/dabcyl-based AP site incision assay [144]. Madhusudan and colleagues identified the compound CRT0044876 from a screen of a collection of structurally-diverse small molecules. Despite the initial promising results obtained in the potentiation of the cell-killing effects of MMS and TMZ, the reproducibility of this compound has been brought into question [21,145] and because of its poor solubility and permeability, this compound was further neglected until now where its usage has been proposed conjugated with platinum [146]. Considering the weak results obtained with CRT0044876, other screenings were then performed. Using a similar HTS approach, Seiple and colleagues screened the 2000-compound NCI Diversity Set of small molecules and identified aromatic nitroso, carboxylate, sulfonamide, and arylstibonic acid compounds with µM affinities for APE1 protein [147]. Again, also for these compounds, the relatively high inhibitory potency observed in vitro did not find a parallel significant effect in cells. Successively, various one-off studies did not progress to lead optimization. For example, in 2009, prompted by the evidence that APE1 represents an attractive therapeutic target in anticancer drug development, Zawahir and colleagues utilized a pharmacophore-based approach that was used to carry out a virtual screen of a 365,000 small molecule library [148]. The known interactions of APE1 with AP site-containing DNA including components of hydrophobicity, H- bond acceptor, and negatively ionizable features were utilized to design a virtual screen. In the same year, Simeonov and colleagues employing a quantitative HTS screened the commercially available Library of Pharmacologically Active Compounds (LOPAC), identifying 6-hydroxy-DL- DOPA, Reactive blue 2, and myricetin as APE1 possible inhibitors [145]. Although these approaches predicted several molecules, they were not all tested in cell-based assays and thus have not been evaluated for cell permeability.
Successively, Kelley’s group has also used a fluorescence-based high-throughput assay to screen a library of 60,000 small molecule compounds for the ability to inhibit APE1 AP endonuclease activity [149]. The most promising compounds were designated as APE1 Repair Inhibitor AR01, 02, 03, and 06. AR03 is chemically distinct from the previously reported small molecule inhibitors of APE1. This compound was demonstrated to inhibit the cleavage of AP sites in vitro of whole cell extracts and to potentiate the cytotoxicity of TMZ and MMS of glioblastoma SF767 cells. While it is cell-permeable, its planar fused-ring structure may suggest its DNA intercalating ability thus potentially being aspecific.
In 2011, Mohammed and colleagues focused on developing APE1 inhibitors for melanoma and glioma treatment using a structure-based drug design approach [96]. The crystal structure of APE1 was utilized to create four pharmacophore models, including the interactions of the previously identified inhibitor CRT0044876 with active site residues and molecular scaffolds designed to fit the ligand binding site. From the screening of 1679 hits, the authors identified compound 4 (N-(4-fluorophenyl)-2-(4-phenylsulfonyl-2- (p-tolyl)oxazol-5-yl) sulfanyl-acetamide) as the one with the highest AP endonuclease inhibitory activity and the potential to sensitize the activity of MMS and TMZ in both glioma and melanoma cell lines but not in HUVEC cells, suggesting specificity for malignant tissue.
In 2012, a new class of inhibitors of the catalytic endonuclease function was identified by Aiello et al. [150]. Compounds 32-35, which have a 3-benzylcarbamoyl-2-methoxybenzoic acid structure, showed the most active and selective inhibition activity of APE1. These compounds have the potential to be used in combination therapy with 5-fluorodeoxyuridine for colon cancer treatment. In the same year, using a docking-based virtual screening, 15 potential compounds were identified as inhibitors of the APE1 from a library of over 4 million molecules [151]. Two of these compounds, 36 and 37, were found to be potent inhibitors of the protein and could increase the toxicity of MMS. Through molecular dynamics simulations, it was discovered that these compounds may interact with the protein through important binding modes such as hydrogen bonds with specific residues and hydrophobic interactions by virtue of their quinoxaline core. Also in 2012, Simeonov’s group performed a fully-automated HTS using a kinetic fluorescence assay on the NIH Molecular Libraries Small Molecule Repository and other collections, examining each agent at different concentrations [152]. They identified active APE1 inhibitors able to potentiate the genotoxic effect of MMS, leading to an increase in AP sites. The chemical structures of the most effective inhibitors namely MLS001196838, MLS000587064, MLS000737267, MLS000090966, and MLS000863573 would have served as starting points for medicinal chemists to further optimize them.
Another fluorescence-based quantitative HTS of 352 489 small molecules from the NIH Molecular Libraries Small Molecule Repository was performed by Rai and colleagues [153]. APE Inhibitor III (N-(3-(1,3-Benzo[d]thiazol-2-yl)-6-isopropyl-4,5,6,7-tetrahydrothieno[2,3-c]pyridin-2-yl)acetamide) has been demonstrated to potentiate MMS and TMZ activity in HeLa cells. This compound was further used and distributed by the vendors as one of the most promising APE1 inhibitors, however, also this compound has not significantly advanced beyond in vitro studies. Srinivasan et al., using the crystal structure of APE1, computationally constructed molecules that would sterically block its endonuclease site and identified molecules that all contain 2-methyl-4- amino-6,7-dioxoloquinoline structure [154]. The mechanism of action of the compounds was probed by fluorescence and competition studies in T98G glioma cell lines, which indicated for compounds 1 and 4 direct interaction between the inhibitor and the active site of APE1 protein.
In 2015, a pharmacophore model for APE1 small-molecule inhibitors was used to identify new compounds by the means of in silico screening of 10 159 compounds [155]. The virtual docking assay identify four compounds with a 2-methyl-4-amino-6,7-dioxoloquinoline core (AJAY 1-4); AJAY 4 showed the best results in the inhibition of cell growth, however, none of the compounds have advanced in clinical studies.
Another in silico novel approach was pursued by Trilles et al., who guided by X-ray crystal structures of APE1 and computational docking of solvents identify binding hotspots for small organic molecules [156]. Accordingly, they screened a library of macrocycles for inhibiting APE1 endonuclease activity and identified four novel macrocycles that they used as a starting point for designing APE1 ligands. From the initial screening of 66 compounds, four exhibited concentration-dependent inhibition of APE1 endonuclease activity (MC043, MC047, MC042, and MC019). Building on these hits, additional macrocycles were synthesized and macrocyclic lactams 13, 21, 24 have been demonstrated to be more effective in inhibiting APE1 endonuclease function in combination with MMS.
Unfortunately, none of the compounds developed so far have advanced to significant in vivo studies or clinical trials. Interestingly, very recently, two works argued about the specificity of some of the most prominent compounds that are usually sold by vendors as APE1 inhibitors. In the work of Pidugu et al., it has been demonstrated through structural, biophysical, and biochemical approaches that several reported small molecules are weak APE1 inhibitors [157]. In particular, through NMR chemical shift perturbation assay, they showed that CRT0044876 and three similar indole-2-carboxylic acids compounds (5-fluoroindole-2-carboxylic acid [96], 5-nitroindole-2-carboxylic acid and 6-bromoindole-2-carboxylic acid) bind at a pocket of APE1 that is distal from its active site. Furthermore, using Dynamic light scattering (DLS), they also demonstrated that CRT0044876 [144], myricetin [145], and APE Inhibitor III [153] form colloidal aggregates that could sequester APE1 causing non-specific inhibition. For this latter compound, also Xue and Demple recently raised the question about the specificity of this molecule [136]. Since APE1 Knockout lines (CH12F3 [158]) showed equal sensitivity to direct killing by APE Inhibitor III, being even more sensitive to APE Inhibitor III than its wildtype counterpart, the authors claimed possible off-target effects that must be taken into account when using these inhibitors at high dosages.
3.2. Targeting the APE1 redox activity
Unlike the BER function, which is highly conserved from prokaryotes (E. coli exonuclease III) to humans, the redox function is probably unique to mammalians [159]. Whereas the C-terminal of APE1 is mainly involved in the regulation of the endodeoxyribonuclease activity, the N-terminal, unstructured region is strongly implicated in the protein-protein interactions (PPI) and in the activation of several transcription factors (TFs) via a redox mechanism. The redox function of APE1 is exploited by cysteine residues sited at positions 65, 93, and 99 of the unstructured N- terminal region (Figure 1). These residues, are involved in the redox cycle responsible for controlling the reduced state of several TFs [159]. By reducing the TFs , APE1 makes them able to bind DNA. APE1 then returns to its basal state through another reduction, that occurs via a thiol/sulfide exchange with thioredoxin. Among the several TFs regulated by APE1, we include the principal such as NF B [160], AP-1 [160], HIF1 [161], STAT3 [162,163], p53 [164], NRF2 [165], Pax-5 and -8 [166] and others [167]. Given the roles of all these TFs in the cellular biological processes, the effects of APE1 as a redox signaling factor regards principally the promotion of the growth, migration, DDR signaling, and survival in tumor cells as well as inflammation and angiogenesis in the tumor microenvironment. Thus, the inhibition of APE1 redox activity can be a target for slowing growth and progression during tumoral processes. The pharmacological inhibition of APE1 redox activity causes a decrease in the ability of the transcription factors to bind to DNA [167,168,169] and thereby increases the cancer cells’ response to chemotherapeutic agents [170,171].
Differently from the AP-endonuclease inhibitors, testing redox inhibitors resulted in more complications during these years, in part due to the arduous modalities of detection of the redox activity of APE1. In this paragraph, we propose a roundup of literature on a few redox inhibitors that emerged onto the scientific scene.
Dietary agents and several compounds from natural sources, such as soy isoflavones, resveratrol, and curcumin but also the vitamins ascorbate and α-tocopherol [172], were initially tested. Curcumin is a polyphenol with the potential for treatment or prevention of particular human diseases such as oxidative and inflammatory conditions, metabolic syndrome, arthritis, anxiety, hyperlipidemia, and cancers [173]. In 2017, it was demonstrated that Curcumin affects the APE1 redox function inhibiting the transcriptional activity of APE1 on AP-1 and NF- B genes in vitro [174]. For its multiple anti-inflammatories, anti-oxidant and anti-neoplastic properties, Curcumin was enrolled in more than 300 clinical trials. Resveratrol is a naturally occurring polyphenolic compound present in red wine and grapes. It has been demonstrated that it exhibits a neuroprotective role in models of central nervous system diseases, including cerebral ischemia/reperfusion injury [175]. By inhibiting APE1 redox function, resveratrol causes a significantly diminished activity of AP-1 and NF- B proteins in different human cancers models, enhancing the cytotoxicity of chemotherapy [97]. Similarly, utilizing soy isoflavones to block the redox signaling through APE1 and NF-κB dramatically increased prostate cancer cells’ sensitivity to radiation [176].
About ten years ago, the Kelley’ group synthesized a molecule that turned out to be highly promising in the inhibition of APE1 redox activity in several cancer models [22,177]. This molecule [(2E)-2-[(4,5-dimethoxy-2-methyl-3,6-dioxo-1,4-cyclohexadien-1-yl)methylene]-undecanoic Acid, commonly denoted as APX3330 (or E3330) is a quinone derivative. Several studies were then performed by using different pathological models both in vitro and in vivo in which it was demonstrated that APX3330 selectively inhibits NF-κB-mediated gene expression through APE1 binding [178]. In 2009, Zou and colleagues demonstrated that APX3330 blocked the in vitro growth of pancreatic cancer-associated endothelial cells (PCECs) and EPCs, and the differentiation of bone marrow-derived mesenchymal stem cells (BMSCs) into CD31(+) endothelial progeny. Specifically, the effect was attributable to a reduction of H-Ras expression and intracellular nitric oxide (NO) levels, as well as decreased DNA-binding activity of HIF-1 . Inhibition of the APE1 redox function by APX3330 might be a potent therapeutic strategy in solid tumors [179]. Indeed, APX3330 showed anticancer properties in pancreatic cancer, including inhibition of cancer cell growth and migration in several cancer cell lines and xenograft models in mice [180]. APX3330 inhibited the proliferation, migration, and tube formation of retinal vascular endothelial cells in vitro and reduced retinal angiomatous proliferation neovascularization in vivo [181]. As anticipated in the previous paragraphs, elevated expression levels of APE1 have been correlated with more aggressive phenotypes and poor prognosis of NSCLC. Recently, Manguinhas and colleagues demonstrated that APX3330, in combination with cisplatin, reduces H1975 cell viability, migration, and invasion highlighting its use as a boost of cisplatin in NSCLC cells [182]. Moreover, the inhibition of APE1 redox function through APX3330 combined with docetaxel treatment decreased the proliferative rate and cell migration and invasion of breast cancer cells MDA-MB-231 [183]. The APX3330 inhibitory activity was also assessed in pathological angiogenesis, such as retinal neovascularization [184]. Li and colleagues demonstrated that APX3330 treatment suppresses experimental choroidal neovascularization (CNV) in vitro and in vivo, demonstrating that APE1 regulates multiple transcription factors and inflammatory molecules, and is essential for CEC angiogenesis. APE1 is a novel candidate for therapeutically targeting neovascular eye diseases and alleviating the burden associated with anti-VEGF intravitreal injections [185]. Recently, it has been also demonstrated that APX3330 has the potential to be used for the treatment of γ-herpesvirus infection and associated diseases [186].
Given its promising and potential anti-angiogenic and antineoplastic activities obtained in vitro, in 2017, APX3330 has been enrolled in the APX_CLN_0011 Phase 1 clinical trial. This trial (ClinicalTrials.gov Identifier: NCT03375086) was a multi-center, open-label, dose-escalation oncology study of APX3330 in patients with advanced solid tumors. The study was completed in 2020 showing the assessment of APX3330 safety, anti-tumor activity, pharmacokinetic and pharmacodynamic profile [187], and the recommendations of Phase 2 study dose. Oral APX3330 demonstrated a favorable safety and tolerability profile and was suitable for Phase 2. In 2020, APX3330 has been enrolled in ZETA-1 Phase 2 clinical trial to evaluate its safety and efficacy to treat diabetic retinopathy (DR) and diabetic macular edema (DME). The trial was completed this year (2023) (ClinicalTrials.gov Identifier: NCT03375086). Oral administration of APX3330 and placebo have demonstrated a favorable ophthalmic and systemic safety and tolerability profile. Additional safety data from the ongoing ZETA-1 trial will be evaluated to further characterize the efficacy and safety of APX3330 for the oral treatment of diabetic eye diseases.
On the wave of the results obtained with APX3330, new analogues of this inhibitor were synthesized including RN8-51, 10-52, and 7-60. Data shown in [188] have demonstrated that the analogue RN8-51 decreased cancer cell growth with little apoptosis demonstrating itself as particularly promising for further anticancer therapeutic development. Kelley and colleagues synthesized novel, second-generation APE1 redox-targeted molecules such as APX2007, APX2009, APX2014, and APX2032 and determined whether they would be protective against neurotoxicity induced by cisplatin or oxaliplatin while not diminishing the platins’ antitumor effect. Specifically, they used ex vivo model of sensory neurons in culture through which they demonstrated that APX2009 [(2E)-2-[(3-methoxy-1,4-dioxo-1,4-dihydronaphthalen-2-yl)methylidene]-N,N-diethylpentanamide] is an effective small molecule that is neuroprotective against cisplatin and oxaliplatin-induced toxicity. APX2009 also demonstrated a strong tumor cell-killing effect in tumor cells and the enhanced tumor cell killing was further substantiated in a more robust three-dimensional pancreatic tumor model [177]. Together, these data suggest that the second-generation compound APX2009 is effective in preventing or reversing platinum-induced CIPN while not affecting the anticancer activity of platins [177]. All three compounds (APX2007, APX2009, and APX2032) demonstrated similar inhibition of NF-κB binding.
Finally, in 2010, Nyland and colleagues described a series of quinones including benzoquinone and naphthoquinone, analogues of the APX3330 with the ability to reduce tumor growth [189].
3.3. Targeting both the APE1 endonuclease and redox activities
A molecule that was shown to inhibit both APE1 endonuclease and redox function is Gossypol [190]. Gossypol is a natural polyphenolic aldehyde that exhibits various effects including antioxidant, anticancer, antiviral, antiparasitic, and antimicrobial activities. It can directly interact with APE1 and enhances the cell-killing effect of MMS and cisplatin. A recent clinical trial (ClinicalTrials.gov Identifier: NCT00540722) aimed to investigate the potential clinical benefit of combining Gossypol with docetaxel and cisplatin in patients with NSCLC who have high expression of APE1 [191]. The trial, designed as a prospective and randomized study, did not show a significant difference between the gossypol and the placebo group, although the gossypol-treated patients had better outcomes of increased progression-free survival (PFS) and overall survival (OS).
AT-101, a derivative of Gossypol and an oral inhibitor of the anti-apoptotic Bcl-2 and Bcl-xL proteins, has been shown to exhibit potent anticancer activity. However, its chemosensitizing effects are not fully understood. AT-101 enhanced the sensitivity of A549 cells to cisplatin in vitro and in vivo by inhibiting APE1-mediated IL-6/STAT3 signaling activation, suggesting its potential use in NSCLC chemotherapy [192]. It was also found to suppress gastric cancer cell migration and renewal and promote chemotherapeutic sensitivity in a gastric cancer model in vivo [193]. The molecular mechanism of its anticancer activity via inhibition of APE1 DNA repair and redox activities is still unclear.
4. Future perspectives from targeting the non-canonical roles of APE1 in miRNA processing
We recently proved that APE1 contributes to the expression of chemoresistance genes via functions in RNA metabolism involving miRs. We found that APE1: i) binds to structured RNAs, including pri-miRs [12,194]; ii) is involved in the processing of miRs implicated in cancer development (e.g. miR-221/222, miR-1246, miR-130b, miR-146a) [195]; iii) is involved in EVs-based sorting of specific miRNA through interaction with hnRNPA2/B1 (in preparation); iv) is a central hub connecting different subnetworks of cancer-associated proteins involved in RNA metabolism and miRs sorting (e.g. NPM1, hnRNPA2/B1, AUF1, FUS, SFPQ) [194,196,197]. We demonstrated that, during genotoxic stress, nuclear APE1 favors the processing and stability of miRs precursors through the association with DROSHA microprocessor complex, impacting, for example, the miR-221/222 axis and in turn, modulating the expression of the tumor suppressor PTEN [12]. Using NSCLC cancer cell lines, we recently defined a signature of 13 miRs (miR-1246, miR-4488, miR-24, miR-183, miR-660, miR-130b, miR-543, miR-200c, miR-376c, miR-218, miR-146a, miR-92b, and miR-33a) that strongly correlate with APE1 expression in human lung cancer and play a central role in cancer cell proliferation and survival [195]. Interestingly, bioinformatic analysis, demonstrated that these miRs possess a strong propensity to contain the RG4 structures in their immature or mature forms (unpublished data). Whether these APE1-regulated miRNAs are responsible for cancer cell response to genotoxic treatment, explaining the role of APE1 in chemoresistance through post-transcriptional mechanisms, is still unknown and should be addressed to understand the central role of APE1 in cancer progression and to define new antitumor strategies. It should be defined whether APE1 recognizes specific oncogenic miR alone or in combination with specific proteins through the detection of regulatory motifs present in the miR structure. On these lines, specific inhibitors of APE1-onco-miR interaction should be explored as potentially novel anticancer strategies for personalized medicine of cancer abs’ on APE1-miR-specific signatures. Work is ongoing in our Lab along these new lines.
5. Secreted APE1 as a novel prognostic non-invasive biomarker of cancer development
We recently showed that APE1 can be secreted (sAPE1) by cancer cells through EVs, including exosomes, during genotoxic stress conditions and it is enzymatically active [8]. However, APE1 presence in the extracellular milieu is still poorly characterized [186,187,188]. sAPE1 expression is actually considered as a novel biomarker for the prognosis of NSCLC as it was proved in a previous study performed on NSCLC patients, in which the levels of sAPE1 resulted significantly more elevated than in healthy controls and were associated with worse progression-free survival [196]. Recently, data obtained by our Lab, were confirmatory of these observations in a cohort of HCC patients [190], in which we found that sAPE1 levels correlated with poor prognosis and were able to discriminate between cancer patients and cirrhotic or healthy donors. The presence of this protein in sera of patients is not solely restricted to cancer diseases but also in inflammatory models, such as coronary artery disease and endotoxemia [191,192]. The biological function of sAPE1 is still completely unknown. An intriguing hypothesis could be that it might act as a paracrine molecule in triggering cell-to-cell communication, important for the local tissue microenvironment inflammatory response.
Also, evidence on the mechanisms responsible for APE1 secretion is lacking, even though the importance of the acetylation, occurring on specific lysine residues sited in the first 33 N-terminal portions of the protein (K27, K31, K32 and K35), has been highlighted in cells treated with the histone deacetylase inhibitor tricostatin A (TSA) [12] and our unpublished data. However, it seems reasonable that APE1 secretion might derive from EVs formation via endosomal sorting complex (ESCRT), due to the protein lacking of a classic secretory signal peptide [193]. This pathway is responsible for the biogenesis and maturation of multivesicular bodies (MVBs), composed of many intraluminal vesicles (ILVs), that are released in the extracellular milieu as exosomes. ILVs formation can occur through several mechanisms and information about the regulation of these processes and the possible differences between the promoted cargo selection is still missing [194].
It is conceivable that these vesicles might be highly shuttled between cells within tumor mass and deliver their content within target cells. This process may fulfill the cancer cells' requirement of a high amount of APE1 to counteract the DNA damage inferred by drugs in a paracrine manner, suggesting that APE1-secretion could represent a novel Damage-associated molecular pattern (DAMP) mechanism, that deserves further in-depth study to develop inhibitors that could specifically target alterations of APE1 secretion in different cancers.
6. Conclusive remarks and future perspectives
Albeit the high potency of many of the compounds aforementioned, additional work is necessary to deliver more specific inhibitors of APE1-altered functions in tumors, which could be useful for clinical trials. While progress has been certainly made in identifying potent APE1 inhibitors, further efforts are needed to specifically achieve selectivity and efficacy. This will require consideration of both the abasic site binding pocket and more distal features of the enzyme that might be important for DNA binding. X-ray crystallography and in vivo experiments would be crucial to expedite rational inhibitor design, validate APE1 as a target, and to explore possible side effects. Moreover, knowing the multiple APE1 cellular function and its relative detailed molecular mechanism could allow us to better target APE1 dysregulated in pathologies (Figure 2).
Targeting a protein-nucleic acid interaction is challenging and this has contributed to the limited success in developing APE1 inhibitors. New approaches are needed for the discovery of novel and selective APE1 inhibitors. In this context, Wilson DM III and colleagues proposed the application of Fragment- and structure-based drug discovery (FBDD/SBDD) methods in the quest for new clinical agents [198]. They applied the ABSOneStepTM platform identifying 25 high-quality crystal structures showing unique and diverse fragment hits bound at the endonuclease site, as well as at a previously unidentified secondary site, overall suggesting multiple novel strategies for inhibiting APE1. Indeed, in addition to direct inhibitors of APE1 nuclease activities, inhibitors against other functions of APE1 may also be clinically valuable. Considering the complex role of APE1, exploring allosteric modes of inhibition, such as disrupting vital interactions between APE1 and other cellular protein partners might be an alternative option [116,199]. For example, we demonstrated that the molecular association with nucleophosmin (NPM1) modulates the endonuclease activity of APE1 [116]. HTS for the disruption of this interaction led to the discovery of three compounds (fiduxosin, spiclomazine and SB 206553); of these, fiduxosin and spiclomazine displayed anti-proliferative activity and sensitized cells to bleomycin. A synergistic effect with platinum drugs was also observed by using these inhibitors in triple negative breast cancer cell model, demonstrating how APE1 could represent a useful therapeutic biomarker also in this type of tumor [47,200]. Similarly, disruption of other APE1 protein interactions or functions can be taken into consideration.
RNA G-quadruplexes (RG4s) are disease-associated non-canonical structures, composed of stacks of guanine tetrads (called G-quartets) kept together by Hoogsteen hydrogen bonds. RG4s are increasingly recognized as fundamental post-transcriptional regulators of gene expression [201]. Interestingly, these elements are widespread in the transcriptome and are particularly enriched in miRs [202]. The folding of these structures can be controlled by their RBP interactors (i.e. hnRNPA2B1, FUS, etc.), cations ( i.e. K+), and small molecule ligands [203], making RG4s highly dynamic. Very recent data underline a regulatory function played by RG4 in miR-maturation through Dicer-inhibition [204] and a potential role in physiological and pathological liquid-liquid phase separation [205,206]. The presence of the rG4 structure in pre-miR exists in equilibrium with the canonical stem-loop structures and this equilibrium regulates the maturation of some miR, such as miR 92b [207]. However, mechanistic information on RG4 function in miR sorting is missing as well as information on the functional role of oxidized guanine (8-oxo) or abasic (AP) sites in the RG4 forming structures, in the stability and biological properties of the miRs in which these structures are present. Understanding whether APE1 function on miR-processing and -decay could be driven by RG4-mediated folding will open mechanistic views as well as translation applications in cancer biology. We are working along these lines.
A fascinating new field of research relies on the unexpected finding that APE1 can be secreted in the extracellular milieu through EVs. Understanding the intracellular routes responsible for the secretion processes in cancer cells and the role of sAPE1 as a potential paracrine molecule will open new perspectives on precision Medicine.
Author Contributions
All authors have write, read and agreed to the published version of the manuscript.
Funding
This research was funded by AIRC under IG 2017, grant number ID. 19862 (Gianluca Tell).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
The authors thank all the members of the GT lab for fruitful feedbacks.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Antoniali, G.; Malfatti, M.C.; Tell, G. Unveiling the Non-Repair Face of the Base Excision Repair Pathway in RNA Processing: A Missing Link between DNA Repair and Gene Expression? DNA Repair (Amst.) 2017, 56, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Malfatti, M.C.; Antoniali, G.; Codrich, M.; Burra, S.; Mangiapane, G.; Dalla, E.; Tell, G. New Perspectives in Cancer Biology from a Study of Canonical and Non-Canonical Functions of Base Excision Repair Proteins with a Focus on Early Steps. Mutagenesis 2020, 35, 129–149. [Google Scholar] [CrossRef] [PubMed]
- Malfatti, M.C.; Antoniali, G.; Codrich, M.; Tell, G. Coping with RNA Damage with a Focus on APE1, a BER Enzyme at the Crossroad between DNA Damage Repair and RNA Processing/Decay. DNA Repair 2021, 104, 103133. [Google Scholar] [CrossRef]
- Allinson, S.L.; Sleeth, K.M.; Matthewman, G.E.; Dianov, G.L. Orchestration of Base Excision Repair by Controlling the Rates of Enzymatic Activities. DNA Repair 2004, 3, 23–31. [Google Scholar] [CrossRef]
- Hurley, L.H.; Wheelhouse, R.T.; Sun, D.; Kerwin, S.M.; Salazar, M.; Fedoroff, O.Y.; Han, F.X.; Han, H.; Izbicka, E.; Von Hoff, D.D. G-Quadruplexes as Targets for Drug Design. Pharmacol. Ther. 2000, 85, 141–158. [Google Scholar] [CrossRef] [PubMed]
- Fleming, A.M.; Burrows, C.J. G-Quadruplex Folds of the Human Telomere Sequence Alter the Site Reactivity and Reaction Pathway of Guanine Oxidation Compared to Duplex DNA. Chemical Research in Toxicology 2013, 26, 593–607. [Google Scholar] [CrossRef] [PubMed]
- Fleming, A.M.; Zhu, J.; Ding, Y.; Visser, J.A.; Zhu, J.; Burrows, C.J. Human DNA Repair Genes Possess Potential G-Quadruplex Sequences in Their Promoters and 5’-Untranslated Regions. Biochemistry 2018, 57, 991–1002. [Google Scholar] [CrossRef] [PubMed]
- Fleming, A.M.; Burrows, C.J. 8-Oxo-7,8-Dihydroguanine, Friend and Foe: Epigenetic-like Regulator versus Initiator of Mutagenesis. DNA Repair (Amst.) 2017, 56, 75–83. [Google Scholar] [CrossRef]
- Fleming, A.M.; Zhu, J.; Ding, Y.; Burrows, C.J. 8-Oxo-7,8-Dihydroguanine in the Context of a Gene Promoter G-Quadruplex Is an On–Off Switch for Transcription. ACS Chemical Biology 2017, 12, 2417–2426. [Google Scholar] [CrossRef]
- Burra, S.; Marasco, D.; Malfatti, M.C.; Antoniali, G.; Virgilio, A.; Esposito, V.; Demple, B.; Galeone, A.; Tell, G. Human AP-Endonuclease (Ape1) Activity on Telomeric G4 Structures Is Modulated by Acetylatable Lysine Residues in the N-Terminal Sequence. DNA Repair (Amst.) 2019, 73, 129–143. [Google Scholar] [CrossRef]
- Roychoudhury, S.; Pramanik, S.; Harris, H.L.; Tarpley, M.; Sarkar, A.; Spagnol, G.; Sorgen, P.L.; Chowdhury, D.; Band, V.; Klinkebiel, D.; et al. Endogenous Oxidized DNA Bases and APE1 Regulate the Formation of G-Quadruplex Structures in the Genome. Proc. Natl. Acad. Sci. U.S.A. 2020, 117, 11409–11420. [Google Scholar] [CrossRef] [PubMed]
- Antoniali, G.; Serra, F.; Lirussi, L.; Tanaka, M.; D’Ambrosio, C.; Zhang, S.; Radovic, S.; Dalla, E.; Ciani, Y.; Scaloni, A.; et al. Mammalian APE1 Controls MiRNA Processing and Its Interactome Is Linked to Cancer RNA Metabolism. Nat Commun 2017, 8, 797. [Google Scholar] [CrossRef]
- Barnes, T.; Kim, W.-C.; Mantha, A.K.; Kim, S.-E.; Izumi, T.; Mitra, S.; Lee, C.H. Identification of Apurinic/Apyrimidinic Endonuclease 1 (APE1) as the Endoribonuclease That Cleaves c-Myc MRNA. Nucleic Acids Research 2009, 37, 3946–3958. [Google Scholar] [CrossRef] [PubMed]
- Chohan, M.; Mackedenski, S.; Li, W.-M.; Lee, C.H. Human Apurinic/Apyrimidinic Endonuclease 1 (APE1) Has 3′ RNA Phosphatase and 3′ Exoribonuclease Activities. Journal of Molecular Biology 2015, 427, 298–311. [Google Scholar] [CrossRef] [PubMed]
- Malfatti, M.C.; Balachander, S.; Antoniali, G.; Koh, K.D.; Saint-Pierre, C.; Gasparutto, D.; Chon, H.; Crouch, R.J.; Storici, F.; Tell, G. Abasic and Oxidized Ribonucleotides Embedded in DNA Are Processed by Human APE1 and Not by RNase H2. Nucleic Acids Res. 2017, 45, 11193–11212. [Google Scholar] [CrossRef] [PubMed]
- Malfatti, M.C.; Codrich, M.; Dalla, E.; D’Ambrosio, C.; Storici, F.; Scaloni, A.; Tell, G. AUF1 Recognizes 8-Oxo-Guanosine Embedded in DNA and Stimulates APE1 Endoribonuclease Activity. Antioxidants & Redox Signaling 2023, ars.2022.0105. [CrossRef]
- Tosolini, D.; Antoniali, G.; Dalla, E.; Tell, G. Role of Phase Partitioning in Coordinating DNA Damage Response: Focus on the Apurinic Apyrimidinic Endonuclease 1 Interactome. Biomolecular Concepts 2020, 11, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhao, H.; McMahon, A.; Yan, S. APE1 Assembles Biomolecular Condensates to Promote the ATR–Chk1 DNA Damage Response in Nucleolus. Nucleic Acids Research 2022, 50, 10503–10525. [Google Scholar] [CrossRef]
- Dall’Agnese, G.; Dall’Agnese, A.; Banani, S.F.; Codrich, M.; Malfatti, M.C.; Antoniali, G.; Tell, G. Role of Condensates in Modulating DNA Repair Pathways and Its Implication for Chemoresistance. Journal of Biological Chemistry 2023, 104800. [Google Scholar] [CrossRef]
- Mangiapane, G.; Parolini, I.; Conte, K.; Malfatti, M.C.; Corsi, J.; Sanchez, M.; Pietrantoni, A.; D’Agostino, V.G.; Tell, G. Enzymatically Active Apurinic/Apyrimidinic Endodeoxyribonuclease 1 Is Released by Mammalian Cells through Exosomes. J Biol Chem 2021, 296, 100569. [Google Scholar] [CrossRef]
- Fishel, M.L.; Kelley, M.R. The DNA Base Excision Repair Protein Ape1/Ref-1 as a Therapeutic and Chemopreventive Target. Molecular Aspects of Medicine 2007, 28, 375–395. [Google Scholar] [CrossRef]
- Shah, F.; Logsdon, D.; Messmann, R.A.; Fehrenbacher, J.C.; Fishel, M.L.; Kelley, M.R. Exploiting the Ref-1-APE1 Node in Cancer Signaling and Other Diseases: From Bench to Clinic. npj Precision Onc 2017, 1, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Lirussi, L.; Antoniali, G.; D’Ambrosio, C.; Scaloni, A.; Nilsen, H.; Tell, G. APE1 Polymorphic Variants Cause Persistent Genomic Stress and Affect Cancer Cell Proliferation. Oncotarget 2016, 7, 26293–26306. [Google Scholar] [CrossRef] [PubMed]
- Wallace, S.S.; Murphy, D.L.; Sweasy, J.B. Base Excision Repair and Cancer. Cancer Lett 2012, 327, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, A.M.; Stark, W.J.; Flynn, T.S.; Freudenthal, B.D. Molecular and Structural Characterization of Disease-Associated APE1 Polymorphisms. DNA Repair (Amst) 2020, 91–92, 102867. [Google Scholar] [CrossRef]
- Kim, W.C.; Ma, C.; Li, W.-M.; Chohan, M.; Wilson III, D.M.; Lee, C.H. Altered Endoribonuclease Activity of Apurinic/Apyrimidinic Endonuclease 1 Variants Identified in the Human Population. PLoS One 2014, 9, e90837. [Google Scholar] [CrossRef]
- Wilson, D.M.; Kim, D.; Berquist, B.R.; Sigurdson, A.J. Variation in Base Excision Repair Capacity. Mutat Res 2011, 711, 100–112. [Google Scholar] [CrossRef]
- Pieretti, M.; Khattar, N.H.; Smith, S.A. Common Polymorphisms and Somatic Mutations in Human Base Excision Repair Genes in Ovarian and Endometrial Cancers. Mutat Res 2001, 432, 53–59. [Google Scholar] [CrossRef]
- Illuzzi, J.L.; Harris, N.A.; Manvilla, B.A.; Kim, D.; Li, M.; Drohat, A.C.; Iii, D.M.W. Functional Assessment of Population and Tumor-Associated APE1 Protein Variants. PLOS ONE 2013, 8, e65922. [Google Scholar] [CrossRef]
- Abyzov, A.; Uzun, A.; Strauss, P.R.; Ilyin, V.A. An AP Endonuclease 1-DNA Polymerase Beta Complex: Theoretical Prediction of Interacting Surfaces. PLoS Comput Biol 2008, 4, e1000066. [Google Scholar] [CrossRef]
- Hinz, J.M.; Mao, P.; McNeill, D.R.; Wilson, D.M. Reduced Nuclease Activity of Apurinic/Apyrimidinic Endonuclease (APE1) Variants on Nucleosomes: IDENTIFICATION OF ACCESS RESIDUES. J Biol Chem 2015, 290, 21067–21075. [Google Scholar] [CrossRef]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative Analysis of Complex Cancer Genomics and Clinical Profiles Using the CBioPortal. Sci Signal 2013, 6, pl1. [Google Scholar] [CrossRef] [PubMed]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The CBio Cancer Genomics Portal: An Open Platform for Exploring Multidimensional Cancer Genomics Data. Cancer Discov 2012, 2, 401–404. [Google Scholar] [CrossRef]
- Reva, B.; Antipin, Y.; Sander, C. Predicting the Functional Impact of Protein Mutations: Application to Cancer Genomics. Nucleic Acids Research 2011, 39, e118. [Google Scholar] [CrossRef] [PubMed]
- Reva, B.; Antipin, Y.; Sander, C. Determinants of Protein Function Revealed by Combinatorial Entropy Optimization. Genome Biol 2007, 8, R232. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Shin, J.H.; Lee, Y.R.; Joo, H.K.; Song, K.H.; Na, Y.G.; Chang, S.J.; Lim, J.S.; Jeon, B.H. Urinary APE1/Ref-1: A Potential Bladder Cancer Biomarker. Dis Markers 2016, 2016, 7276502. [Google Scholar] [CrossRef]
- Fishel, M.L.; Xia, H.; McGeown, J.; McIlwain, D.W.; Elbanna, M.; Craft, A.A.; Kaimakliotis, H.Z.; Sandusky, G.E.; Zhang, C.; Pili, R.; et al. Antitumor Activity and Mechanistic Characterization of APE1/Ref-1 Inhibitors in Bladder Cancer. Mol Cancer Ther 2019, 18, 1947–1960. [Google Scholar] [CrossRef]
- Shin, J.H.; Choi, S.; Lee, Y.R.; Park, M.S.; Na, Y.G.; Irani, K.; Lee, S.D.; Park, J.B.; Kim, J.M.; Lim, J.S.; et al. APE1/Ref-1 as a Serological Biomarker for the Detection of Bladder Cancer. Cancer Res Treat 2015, 47, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Zeng, J.; Lele, S.; LaGrange, C.A.; Bhakat, K.K. APE1 and SSRP1 Is Overexpressed in Muscle Invasive Bladder Cancer and Associated with Poor Survival. Heliyon 2021, 7, e06756. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-A.; Yang, B.; Tang, T.; Yang, Y.; Zhang, D.; Xiao, H.; Xu, J.; Wang, L.; Lin, L.; Jiang, J. Correlation of APE1 with VEGFA and CD163+ Macrophage Infiltration in Bladder Cancer and Their Prognostic Significance. Oncol Lett 2020, 20, 2881–2887. [Google Scholar] [CrossRef]
- Güllü Amuran, G.; Tinay, I.; Filinte, D.; Ilgin, C.; Peker Eyüboğlu, I.; Akkiprik, M. Urinary Micro-RNA Expressions and Protein Concentrations May Differentiate Bladder Cancer Patients from Healthy Controls. Int Urol Nephrol 2020, 52, 461–468. [Google Scholar] [CrossRef]
- Kumar, M.; Shukla, V.K.; Misra, P.K.; Raman, M.J. Dysregulated Expression and Subcellular Localization of Base Excision Repair (BER) Pathway Enzymes in Gallbladder Cancer. Int J Mol Cell Med 2018, 7, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Liu, C.; Lu, H.; Yin, M.; Shao, C.; Hu, X.; Wu, J.; Wang, Y. The Expression of APE1 in Triple-Negative Breast Cancer and Its Effect on Drug Sensitivity of Olaparib. Tumour Biol 2017, 39, 1010428317713390. [Google Scholar] [CrossRef]
- Woo, J.; Park, H.; Sung, S.H.; Moon, B.-I.; Suh, H.; Lim, W. Prognostic Value of Human Apurinic/Apyrimidinic Endonuclease 1 (APE1) Expression in Breast Cancer. PLoS One 2014, 9, e99528. [Google Scholar] [CrossRef] [PubMed]
- Jian, D.; Li, X.-M.; Dai, N.; Liang, D.-D.; Zhang, G.; Mao, C.-Y.; Wang, D.; Song, G.-B.; Li, M.-X.; Luo, H. Inhibition of APE1 Expression Enhances the Antitumor Activity of Olaparib in Triple-Negative Breast Cancer. Evid Based Complement Alternat Med 2022, 2022, 6048017. [Google Scholar] [CrossRef] [PubMed]
- Poletto, M.; Di Loreto, C.; Marasco, D.; Poletto, E.; Puglisi, F.; Damante, G.; Tell, G. Acetylation on Critical Lysine Residues of Apurinic/Apyrimidinic Endonuclease 1 (APE1) in Triple Negative Breast Cancers. Biochem Biophys Res Commun 2012, 424, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Malfatti, M.C.; Gerratana, L.; Dalla, E.; Isola, M.; Damante, G.; Di Loreto, C.; Puglisi, F.; Tell, G. APE1 and NPM1 Protect Cancer Cells from Platinum Compounds Cytotoxicity and Their Expression Pattern Has a Prognostic Value in TNBC. J Exp Clin Cancer Res 2019, 38, 309. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Fatah, T.M.A.; Perry, C.; Moseley, P.; Johnson, K.; Arora, A.; Chan, S.; Ellis, I.O.; Madhusudan, S. Clinicopathological Significance of Human Apurinic/Apyrimidinic Endonuclease 1 (APE1) Expression in Oestrogen-Receptor-Positive Breast Cancer. Breast Cancer Res Treat 2014, 143, 411–421. [Google Scholar] [CrossRef]
- Herring, C.J.; West, C.M.L.; Wilks, D.P.; Davidson, S.E.; Hunter, R.D.; Berry, P.; Forster, G.; MacKinnon, J.; Rafferty, J.A.; Elder, R.H.; et al. Levels of the DNA Repair Enzyme Human Apurinic/Apyrimidinic Endonuclease (APE1, APEX, Ref-1) Are Associated with the Intrinsic Radiosensitivity of Cervical Cancers. Br J Cancer 1998, 78, 1128–1133. [Google Scholar] [CrossRef] [PubMed]
- Schindl, M.; Oberhuber, G.; Pichlbauer, E.G.; Obermair, A.; Birner, P.; Kelley, M.R. DNA Repair-Redox Enzyme Apurinic Endonuclease in Cervical Cancer: Evaluation of Redox Control of HIF-1alpha and Prognostic Significance. Int J Oncol 2001, 19, 799–802. [Google Scholar] [CrossRef]
- Li, Q.; Wei, X.; Zhou, Z.-W.; Wang, S.-N.; Jin, H.; Chen, K.-J.; Luo, J.; Westover, K.D.; Wang, J.-M.; Wang, D.; et al. GADD45α Sensitizes Cervical Cancer Cells to Radiotherapy via Increasing Cytoplasmic APE1 Level. Cell Death Dis 2018, 9, 1–11. [Google Scholar] [CrossRef]
- Li, Q.; Zhou, Z.-W.; Duan, W.; Qian, C.-Y.; Wang, S.-N.; Deng, M.-S.; Zi, D.; Wang, J.-M.; Mao, C.-Y.; Song, G.; et al. Inhibiting the Redox Function of APE1 Suppresses Cervical Cancer Metastasis via Disengagement of ZEB1 from E-Cadherin in EMT. J Exp Clin Cancer Res 2021, 40, 220. [Google Scholar] [CrossRef]
- Bhakat, K.K.; Sengupta, S.; Adeniyi, V.F.; Roychoudhury, S.; Nath, S.; Bellot, L.J.; Feng, D.; Mantha, A.K.; Sinha, M.; Qiu, S.; et al. Regulation of Limited N-Terminal Proteolysis of APE1 in Tumor via Acetylation and Its Role in Cell Proliferation. Oncotarget 2016, 7, 22590–22604. [Google Scholar] [CrossRef] [PubMed]
- Kakolyris, S.; Kaklamanis, L.; Engels, K.; Turley, H.; Hickson, I.D.; Gatter, K.C.; Harris, A.L. Human Apurinic Endonuclease 1 Expression in a Colorectal Adenoma-Carcinoma Sequence. Cancer Res 1997, 57, 1794–1797. [Google Scholar] [PubMed]
- Lou, D.; Zhu, L.; Ding, H.; Dai, H.-Y.; Zou, G.-M. Aberrant Expression of Redox Protein Ape1 in Colon Cancer Stem Cells. Oncol Lett 2014, 7, 1078–1082. [Google Scholar] [CrossRef]
- Codrich, M.; Comelli, M.; Malfatti, M.C.; Mio, C.; Ayyildiz, D.; Zhang, C.; Kelley, M.R.; Terrosu, G.; Pucillo, C.E.M.; Tell, G. Inhibition of APE1-Endonuclease Activity Affects Cell Metabolism in Colon Cancer Cells via a P53-Dependent Pathway. DNA Repair (Amst) 2019, 82, 102675. [Google Scholar] [CrossRef]
- Kühl Svoboda Baldin, R.; Austrália Paredes Marcondes Ribas, C.; de Noronha, L.; Veloso da Silva-Camargo, C.C.; Santos Sotomaior, V.; Martins Sebastião, A.P.; Vasconcelos de Castilho, A.P.; Rodrigues Montemor Netto, M. Expression of Parkin, APC, APE1, and Bcl-XL in Colorectal Polyps. J Histochem Cytochem 2021, 69, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Noike, T.; Miwa, S.; Soeda, J.; Kobayashi, A.; Miyagawa, S. Increased Expression of Thioredoxin-1, Vascular Endothelial Growth Factor, and Redox Factor-1 Is Associated with Poor Prognosis in Patients with Liver Metastasis from Colorectal Cancer. Hum Pathol 2008, 39, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Zeng, J.; Roychoudhury, S.; Biswas, P.; Mohapatra, B.; Ray, S.; Dowlatshahi, K.; Wang, J.; Band, V.; Talmon, G.; et al. Targeting Histone Chaperone FACT Complex Overcomes 5-Fluorouracil Resistance in Colon Cancer. Mol Cancer Ther 2020, 19, 258–269. [Google Scholar] [CrossRef]
- Huajun, W.; Ying, F.; Hongxing, Z.; Weifeng, S.; Pingyang, S.; Mingde, H.; Guoguang, L. Clinical Value of Combined Detection of Serum APE1-Aabs and CEACAM-1 in the Diagnosis of Colorectal Cancer. Eur Rev Med Pharmacol Sci 2018, 22, 1286–1289. [Google Scholar] [CrossRef]
- Deng, X.; Zhen, P.; Niu, X.; Dai, Y.; Wang, Y.; Zhou, M. APE1 Promotes Proliferation and Migration of Cutaneous Squamous Cell Carcinoma. J Dermatol Sci 2020, 100, 67–74. [Google Scholar] [CrossRef]
- Ajucarmelprecilla, A.; Pandi, J.; Dhandapani, R.; Ramanathan, S.; Chinnappan, J.; Paramasivam, R.; Thangavelu, S.; Mohammed Ghilan, A.-K.; Aljohani, S.A.S.; Oyouni, A.A.A.; et al. In Silico Identification of Hub Genes as Observing Biomarkers for Gastric Cancer Metastasis. Evid Based Complement Alternat Med 2022, 2022, 6316158. [Google Scholar] [CrossRef] [PubMed]
- Manoel-Caetano, F.S.; Rossi, A.F.T.; Calvet de Morais, G.; Severino, F.E.; Silva, A.E. Upregulation of the APE1 and H2AX Genes and MiRNAs Involved in DNA Damage Response and Repair in Gastric Cancer. Genes Dis 2019, 6, 176–184. [Google Scholar] [CrossRef]
- Qing, Y.; Li, Q.; Ren, T.; Xia, W.; Peng, Y.; Liu, G.-L.; Luo, H.; Yang, Y.-X.; Dai, X.-Y.; Zhou, S.-F.; et al. Upregulation of PD-L1 and APE1 Is Associated with Tumorigenesis and Poor Prognosis of Gastric Cancer. Drug Des Devel Ther 2015, 9, 901–909. [Google Scholar] [CrossRef]
- Wei, X.; Li, Y.-B.; Li, Y.; Lin, B.-C.; Shen, X.-M.; Cui, R.-L.; Gu, Y.-J.; Gao, M.; Li, Y.-G.; Zhang, S. Prediction of Lymph Node Metastases in Gastric Cancer by Serum APE1 Expression. J Cancer 2017, 8, 1492–1497. [Google Scholar] [CrossRef]
- Bobola, M.S.; Blank, A.; Berger, M.S.; Stevens, B.A.; Silber, J.R. Apurinic/Apyrimidinic Endonuclease Activity Is Elevated in Human Adult Gliomas. Clin Cancer Res 2001, 7, 3510–3518. [Google Scholar] [PubMed]
- Scott, T.L.; Wicker, C.A.; Suganya, R.; Dhar, B.; Pittman, T.; Horbinski, C.; Izumi, T. Polyubiquitination of Apurinic/Apyrimidinic Endonuclease 1 by Parkin. Mol Carcinog 2017, 56, 325–336. [Google Scholar] [CrossRef]
- Naidu, M.D.; Mason, J.M.; Pica, R.V.; Fung, H.; Peña, L.A. Radiation Resistance in Glioma Cells Determined by DNA Damage Repair Activity of Ape1/Ref-1. J Radiat Res 2010, 51, 393–404. [Google Scholar] [CrossRef]
- Hudson, A.L.; Parker, N.R.; Khong, P.; Parkinson, J.F.; Dwight, T.; Ikin, R.J.; Zhu, Y.; Chen, J.; Wheeler, H.R.; Howell, V.M. Glioblastoma Recurrence Correlates With Increased APE1 and Polarization Toward an Immuno-Suppressive Microenvironment. Front Oncol 2018, 8, 314. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xiao, H.; Luo, Q.; Li, M.; Wei, S.; Zhu, X.; Xiao, H.; Chen, L. Low APE1/Ref-1 Expression Significantly Correlates with MGMT Promoter Methylation in Patients with High-Grade Gliomas. Int J Clin Exp Pathol 2016, 9, 9562–9568. [Google Scholar]
- Perry, C.; Agarwal, D.; Abdel-Fatah, T.M.A.; Lourdusamy, A.; Grundy, R.; Auer, D.T.; Walker, D.; Lakhani, R.; Scott, I.S.; Chan, S.; et al. Dissecting DNA Repair in Adult High Grade Gliomas for Patient Stratification in the Post-Genomic Era. Oncotarget 2014, 5, 5764–5781. [Google Scholar] [CrossRef]
- Hsia, K.-T.; Liu, C.-J.; Mar, K.; Lin, L.-H.; Lin, C.-S.; Cheng, M.-F.; Lee, H.-S.; Chiu, S.-Y. Impact of Apurinic/Apyrimidinic Endonuclease 1/Redox Factor-1 on Treatment Response and Survival in Oral Squamous Cell Carcinoma. Head Neck 2016, 38, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Wicker, C.A.; Takiar, V.; Suganya, R.; Arnold, S.M.; Brill, Y.M.; Chen, L.; Horbinski, C.M.; Napier, D.; Valentino, J.; Kudrimoti, M.R.; et al. Evaluation of Antioxidant Network Proteins as Novel Prognostic Biomarkers for Head and Neck Cancer Patients. Oral Oncol 2020, 111, 104949. [Google Scholar] [CrossRef] [PubMed]
- Santana, T.; Sá, M.C.; de Moura Santos, E.; Galvão, H.C.; Coletta, R.D.; Freitas, R. de A. DNA Base Excision Repair Proteins APE-1 and XRCC-1 Are Overexpressed in Oral Tongue Squamous Cell Carcinoma. J Oral Pathol Med 2017, 46, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Li, Y.; Kong, J.; Li, C. Apurinic/Apyrimidinic Endonuclease 1/Redox Factor-1 Could Serve as a Potential Serological Biomarker for the Diagnosis and Prognosis of Oral Squamous Cell Carcinoma. J Oral Maxillofac Surg 2019, 77, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lun, L.; Jiang, X.; Wang, Y.; Li, X.; Du, G.; Wang, J. APE1 Facilitates PD-L1-Mediated Progression of Laryngeal and Hypopharyngeal Squamous Cell Carcinoma. Int Immunopharmacol 2021, 97, 107675. [Google Scholar] [CrossRef]
- Lee, J.W.; Jin, J.; Rha, K.-S.; Kim, Y.M. Expression Pattern of Apurinic/Apyrimidinic Endonuclease in Sinonasal Squamous Cell Carcinoma. Otolaryngol Head Neck Surg 2012, 147, 788–795. [Google Scholar] [CrossRef]
- Souza, L.R.; Fonseca-Silva, T.; Pereira, C.S.; Santos, E.P.; Lima, L.C.; Carvalho, H.A.; Gomez, R.S.; Guimarães, A.L.S.; De Paula, A.M.B. Immunohistochemical Analysis of P53, APE1, HMSH2 and ERCC1 Proteins in Actinic Cheilitis and Lip Squamous Cell Carcinoma. Histopathology 2011, 58, 352–360. [Google Scholar] [CrossRef]
- Di Maso, V.; Mediavilla, M.G.; Vascotto, C.; Lupo, F.; Baccarani, U.; Avellini, C.; Tell, G.; Tiribelli, C.; Crocè, L.S. Transcriptional Up-Regulation of APE1/Ref-1 in Hepatic Tumor: Role in Hepatocytes Resistance to Oxidative Stress and Apoptosis. PLoS One 2015, 10, e0143289. [Google Scholar] [CrossRef]
- Sun, Z.; Zhu, Y.; Aminbuhe; Fan, Q.; Peng, J.; Zhang, N. Differential Expression of APE1 in Hepatocellular Carcinoma and the Effects on Proliferation and Apoptosis of Cancer Cells. BST 2018, 12, 456–462. [CrossRef]
- Lu, X.; Zhao, H.; Yuan, H.; Chu, Y.; Zhu, X. High Nuclear Expression of APE1 Correlates with Unfavorable Prognosis and Promotes Tumor Growth in Hepatocellular Carcinoma. J Mol Histol 2021, 52, 219–231. [Google Scholar] [CrossRef]
- Pascut, D.; Sukowati, C.H.C.; Antoniali, G.; Mangiapane, G.; Burra, S.; Mascaretti, L.G.; Buonocore, M.R.; Crocè, L.S.; Tiribelli, C.; Tell, G. Serum AP-Endonuclease 1 (SAPE1) as Novel Biomarker for Hepatocellular Carcinoma. Oncotarget 2019, 10, 383–394. [Google Scholar] [CrossRef]
- Di Maso, V.; Avellini, C.; Crocè, L.S.; Rosso, N.; Quadrifoglio, F.; Cesaratto, L.; Codarin, E.; Bedogni, G.; Beltrami, C.A.; Tell, G.; et al. Subcellular Localization of APE1/Ref-1 in Human Hepatocellular Carcinoma: Possible Prognostic Significance. Mol Med 2007, 13, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Bazzani, V.; Barchiesi, A.; Radecka, D.; Pravisani, R.; Guadagno, A.; Di Loreto, C.; Baccarani, U.; Vascotto, C. Mitochondrial Apurinic/Apyrimidinic Endonuclease 1 Enhances MtDNA Repair Contributing to Cell Proliferation and Mitochondrial Integrity in Early Stages of Hepatocellular Carcinoma. BMC Cancer 2020, 20, 969. [Google Scholar] [CrossRef]
- Sengupta, S.; Mantha, A.K.; Song, H.; Roychoudhury, S.; Nath, S.; Ray, S.; Bhakat, K.K. Elevated Level of Acetylation of APE1 in Tumor Cells Modulates DNA Damage Repair. Oncotarget 2016, 7, 75197–75209. [Google Scholar] [CrossRef] [PubMed]
- Long, K.; Gu, L.; Li, L.; Zhang, Z.; Li, E.; Zhang, Y.; He, L.; Pan, F.; Guo, Z.; Hu, Z. Small-Molecule Inhibition of APE1 Induces Apoptosis, Pyroptosis, and Necroptosis in Non-Small Cell Lung Cancer. Cell Death Dis 2021, 12, 1–15. [Google Scholar] [CrossRef]
- Yoo, D.G.; Song, Y.J.; Cho, E.J.; Lee, S.K.; Park, J.B.; Yu, J.H.; Lim, S.P.; Kim, J.M.; Jeon, B.H. Alteration of APE1/Ref-1 Expression in Non-Small Cell Lung Cancer: The Implications of Impaired Extracellular Superoxide Dismutase and Catalase Antioxidant Systems. Lung Cancer 2008, 60, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Li, Q.; Li, Y.; Duan, W.; Huang, C.; Zheng, X.; Sun, L.; Luo, J.; Wang, D.; Zhang, S.; et al. Prediction of Survival Prognosis of Non-Small Cell Lung Cancer by APE1 through Regulation of Epithelial-Mesenchymal Transition. Oncotarget 2016, 7, 28523–28539. [Google Scholar] [CrossRef]
- Gu, X.; Cun, Y.; Li, M.; Qing, Y.; Jin, F.; Zhong, Z.; Dai, N.; Qian, C.; Sui, J.; Wang, D. Human Apurinic/Apyrimidinic Endonuclease SiRNA Inhibits the Angiogenesis Induced by X-Ray Irradiation in Lung Cancer Cells. Int J Med Sci 2013, 10, 870–882. [Google Scholar] [CrossRef]
- Zhang, S.; He, L.; Dai, N.; Guan, W.; Shan, J.; Yang, X.; Zhong, Z.; Qing, Y.; Jin, F.; Chen, C.; et al. Serum APE1 as a Predictive Marker for Platinum-Based Chemotherapy of Non-Small Cell Lung Cancer Patients. Oncotarget 2016, 7, 77482–77494. [Google Scholar] [CrossRef]
- Kakolyris, S.; Giatromanolaki, A.; Koukourakis, M.; Kaklamanis, L.; Kanavaros, P.; Hickson, I.D.; Barzilay, G.; Georgoulias, V.; Gatter, K.C.; Harris, A.L. Nuclear Localization of Human AP Endonuclease 1 (HAP1/Ref-1) Associates with Prognosis in Early Operable Non-Small Cell Lung Cancer (NSCLC). J Pathol 1999, 189, 351–357. [Google Scholar] [CrossRef]
- Puglisi, F.; Aprile, G.; Minisini, A.M.; Barbone, F.; Cataldi, P.; Tell, G.; Kelley, M.R.; Damante, G.; Beltrami, C.A.; Di Loreto, C. Prognostic Significance of Ape1/Ref-1 Subcellular Localization in Non-Small Cell Lung Carcinomas. Anticancer Res 2001, 21, 4041–4049. [Google Scholar] [PubMed]
- Wu, H.-H.; Chu, Y.-C.; Wang, L.; Tsai, L.-H.; Lee, M.-C.; Chen, C.-Y.; Shieh, S.-H.; Cheng, Y.-W.; Lee, H. Cytoplasmic Ape1 Expression Elevated by P53 Aberration May Predict Survival and Relapse in Resected Non-Small Cell Lung Cancer. Ann Surg Oncol 2013, 20 Suppl 3, S336–347. [Google Scholar] [CrossRef]
- Wu, H.-H.; Cheng, Y.-W.; Chang, J.T.; Wu, T.-C.; Liu, W.-S.; Chen, C.-Y.; Lee, H. Subcellular Localization of Apurinic Endonuclease 1 Promotes Lung Tumor Aggressiveness via NF-KappaB Activation. Oncogene 2010, 29, 4330–4340. [Google Scholar] [CrossRef]
- Abbotts, R.; Jewell, R.; Nsengimana, J.; Maloney, D.J.; Simeonov, A.; Seedhouse, C.; Elliott, F.; Laye, J.; Walker, C.; Jadhav, A.; et al. Targeting Human Apurinic/Apyrimidinic Endonuclease 1 (APE1) in Phosphatase and Tensin Homolog (PTEN) Deficient Melanoma Cells for Personalized Therapy. Oncotarget 2014, 5, 3273–3286. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, M.Z.; Vyjayanti, V.N.; Laughton, C.A.; Dekker, L.V.; Fischer, P.M.; Wilson, D.M.; Abbotts, R.; Shah, S.; Patel, P.M.; Hickson, I.D.; et al. Development and Evaluation of Human AP Endonuclease Inhibitors in Melanoma and Glioma Cell Lines. Br J Cancer 2011, 104, 653–663. [Google Scholar] [CrossRef]
- Yang, S.; Irani, K.; Heffron, S.E.; Jurnak, F.; Meyskens, F.L. Alterations in the Expression of the Apurinic/Apyrimidinic Endonuclease-1/Redox Factor-1 (APE/Ref-1) in Human Melanoma and Identification of the Therapeutic Potential of Resveratrol as an APE/Ref-1 Inhibitor. Mol Cancer Ther 2005, 4, 1923–1935. [Google Scholar] [CrossRef] [PubMed]
- Guida, M.; Tommasi, S.; Strippoli, S.; Natalicchio, M.I.; De Summa, S.; Pinto, R.; Cramarossa, A.; Albano, A.; Pisconti, S.; Aieta, M.; et al. The Search for a Melanoma-Tailored Chemotherapy in the New Era of Personalized Therapy: A Phase II Study of Chemo-Modulating Temozolomide Followed by Fotemustine and a Cooperative Study of GOIM (Gruppo Oncologico Italia Meridionale). BMC Cancer 2018, 18, 552. [Google Scholar] [CrossRef]
- Al-Attar, A.; Gossage, L.; Fareed, K.R.; Shehata, M.; Mohammed, M.; Zaitoun, A.M.; Soomro, I.; Lobo, D.N.; Abbotts, R.; Chan, S.; et al. Human Apurinic/Apyrimidinic Endonuclease (APE1) Is a Prognostic Factor in Ovarian, Gastro-Oesophageal and Pancreatico-Biliary Cancers. Br J Cancer 2010, 102, 704–709. [Google Scholar] [CrossRef]
- Hong, J.; Chen, Z.; Peng, D.; Zaika, A.; Revetta, F.; Washington, M.K.; Belkhiri, A.; El-Rifai, W. APE1-Mediated DNA Damage Repair Provides Survival Advantage for Esophageal Adenocarcinoma Cells in Response to Acidic Bile Salts. Oncotarget 2016, 7, 16688–16702. [Google Scholar] [CrossRef]
- Bhat, A.A.; Lu, H.; Soutto, M.; Capobianco, A.; Rai, P.; Zaika, A.; El-Rifai, W. Exposure of Barrett’s and Esophageal Adenocarcinoma Cells to Bile Acids Activates EGFR-STAT3 Signaling Axis via Induction of APE1. Oncogene 2018, 37, 6011–6024. [Google Scholar] [CrossRef]
- Sriramajayam, K.; Peng, D.; Lu, H.; Zhou, S.; Bhat, N.; McDonald, O.G.; Que, J.; Zaika, A.; El-Rifai, W. Activation of NRF2 by APE1/REF1 Is Redox-Dependent in Barrett’s Related Esophageal Adenocarcinoma Cells. Redox Biol 2021, 43, 101970. [Google Scholar] [CrossRef]
- Han, G.; Tian, Y.; Duan, B.; Sheng, H.; Gao, H.; Huang, J. Association of Nuclear Annexin A1 with Prognosis of Patients with Esophageal Squamous Cell Carcinoma. Int J Clin Exp Pathol 2014, 7, 751–759. [Google Scholar]
- Song, J.; Futagami, S.; Nagoya, H.; Kawagoe, T.; Yamawaki, H.; Kodaka, Y.; Tatsuguchi, A.; Gudis, K.; Wakabayashi, T.; Yonezawa, M.; et al. Apurinic/Apyrimidinic Endonuclease-1 (APE-1) Is Overexpressed via the Activation of NF-ΚB-P65 in MCP-1-Positive Esophageal Squamous Cell Carcinoma Tissue. J Clin Biochem Nutr 2013, 52, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, Z.; Li, Q.; Zhang, L.; Cheng, Y.; Zhong, Z. Mitochondrial APE1 Promotes Cisplatin Resistance by Downregulating ROS in Osteosarcoma. Oncol Rep 2020, 44, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, Z.; Zhang, L.; Zhong, Z. Cytoplasmic APE1 Promotes Resistance Response in Osteosarcoma Patients with Cisplatin Treatment. Cell Biochemistry and Function 2020, 38, 195–203. [Google Scholar] [CrossRef]
- Wang, D.; Luo, M.; Kelley, M.R. Human Apurinic Endonuclease 1 (APE1) Expression and Prognostic Significance in Osteosarcoma: Enhanced Sensitivity of Osteosarcoma to DNA Damaging Agents Using Silencing RNA APE1 Expression Inhibition. Mol Cancer Ther 2004, 3, 679–686. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, Y.; Yuan, Z.; Wang, C.; Shi, Y. Predicting Chemosensitivity in Osteosarcoma Prior to Chemotherapy: An Investigational Study of Biomarkers with Immunohistochemistry. Oncology Letters 2012, 3, 1011–1016. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.; Qing, Y.; Dai, N.; Li, M.; Qian, C.; Yang, Y.; Cheng, Y.; Li, Z.; Zhang, S.; Zhong, Z.; et al. Apurinic/Apyrimidinic Endonuclease 1 Induced Upregulation of Fibroblast Growth Factor 2 and Its Receptor 3 Induces Angiogenesis in Human Osteosarcoma Cells. Cancer Sci 2014, 105, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Shan, J.; Dai, N.; Zhong, Z.; Qing, Y.; Yang, Y.; Zhang, S.; Li, C.; Sui, J.; Ren, T.; et al. Apurinic/Apyrimidinic Endonuclease 1 Regulates Angiogenesis in a Transforming Growth Factor β-Dependent Manner in Human Osteosarcoma. Cancer Sci 2015, 106, 1394–1401. [Google Scholar] [CrossRef]
- Dai, N.; Qing, Y.; Cun, Y.; Zhong, Z.; Li, C.; Zhang, S.; Shan, J.; Yang, X.; Dai, X.; Cheng, Y.; et al. MiR-513a-5p Regulates Radiosensitivity of Osteosarcoma by Targeting Human Apurinic/Apyrimidinic Endonuclease. Oncotarget 2016, 9, 25414–25426. [Google Scholar] [CrossRef]
- Londero, A.P.; Orsaria, M.; Tell, G.; Marzinotto, S.; Capodicasa, V.; Poletto, M.; Vascotto, C.; Sacco, C.; Mariuzzi, L. Expression and Prognostic Significance of APE1/Ref-1 and NPM1 Proteins in High-Grade Ovarian Serous Cancer. Am J Clin Pathol 2014, 141, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Lu, R.; Xie, S.; Zheng, H.; Wang, H.; Wang, Y.; Sun, J.; Gao, X.; Guo, L. APE1 Overexpression Promotes the Progression of Ovarian Cancer and Serves as a Potential Therapeutic Target. Cancer Biomark 2016, 17, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, J.; Xiang, D.; Wang, D.; Xin, X. Alterations in the Expression of the Apurinic/Apyrimidinic Endonuclease-1/Redox Factor-1 (APE1/Ref-1) in Human Ovarian Cancer and Indentification of the Therapeutic Potential of APE1/Ref-1 Inhibitor. Int J Oncol 2009, 35, 1069–1079. [Google Scholar] [PubMed]
- Fan, X.; Wen, L.; Li, Y.; Lou, L.; Liu, W.; Zhang, J. The Expression Profile and Prognostic Value of APE/Ref-1 and NPM1 in High-Grade Serous Ovarian Adenocarcinoma. APMIS 2017, 125, 857–862. [Google Scholar] [CrossRef]
- Poletto, M.; Malfatti, M.C.; Dorjsuren, D.; Scognamiglio, P.L.; Marasco, D.; Vascotto, C.; Jadhav, A.; Maloney, D.J.; Wilson, D.M.; Simeonov, A.; et al. Inhibitors of the Apurinic/Apyrimidinic Endonuclease 1 (APE1)/Nucleophosmin (NPM1) Interaction That Display Anti-Tumor Properties. Mol Carcinog 2016, 55, 688–704. [Google Scholar] [CrossRef]
- Sheng, Q.; Zhang, Y.; Wang, R.; Zhang, J.; Chen, B.; Wang, J.; Zhang, W.; Xin, X. Prognostic Significance of APE1 Cytoplasmic Localization in Human Epithelial Ovarian Cancer. Med Oncol 2012, 29, 1265–1271. [Google Scholar] [CrossRef]
- Moore, D.H.; Michael, H.; Tritt, R.; Parsons, S.H.; Kelley, M.R. Alterations in the Expression of the DNA Repair/Redox Enzyme APE/Ref-1 in Epithelial Ovarian Cancers. Clin Cancer Res 2000, 6, 602–609. [Google Scholar]
- Tanner, B.; Grimme, S.; Schiffer, I.; Heimerdinger, C.; Schmidt, M.; Dutkowski, P.; Neubert, S.; Oesch, F.; Franzen, A.; Kölbl, H.; et al. Nuclear Expression of Apurinic/Apyrimidinic Endonuclease Increases with Progression of Ovarian Carcinomas. Gynecologic Oncology 2004, 92, 568–577. [Google Scholar] [CrossRef]
- Pramanik, S.; Chen, Y.; Song, H.; Khutsishvili, I.; Marky, L.A.; Ray, S.; Natarajan, A.; Singh, P.K.; Bhakat, K.K. The Human AP-Endonuclease 1 (APE1) Is a DNA G-Quadruplex Structure Binding Protein and Regulates KRAS Expression in Pancreatic Ductal Adenocarcinoma Cells. Nucleic Acids Res 2022, 50, 3394–3412. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhou, S.; Sandusky, G.E.; Kelley, M.R.; Fishel, M.L. Reduced Expression of DNA Repair and Redox Signaling Protein APE1/Ref-1 Impairs Human Pancreatic Cancer Cell Survival, Proliferation, and Cell Cycle Progression. Cancer Invest 2010, 28, 885–895. [Google Scholar] [CrossRef]
- Fishel, M.L.; Jiang, Y.; Rajeshkumar, N.V.; Scandura, G.; Sinn, A.L.; He, Y.; Shen, C.; Jones, D.R.; Pollok, K.E.; Ivan, M.; et al. Impact of APE1/Ref-1 Redox Inhibition on Pancreatic Tumor Growth. Mol Cancer Ther 2011, 10, 1698–1708. [Google Scholar] [CrossRef] [PubMed]
- Kelley, M.R.; Cheng, L.; Foster, R.; Tritt, R.; Jiang, J.; Broshears, J.; Koch, M. Elevated and Altered Expression of the Multifunctional DNA Base Excision Repair and Redox Enzyme Ape1/Ref-1 in Prostate Cancer. Clin Cancer Res 2001, 7, 824–830. [Google Scholar]
- Juhnke, M.; Heumann, A.; Chirico, V.; Höflmayer, D.; Menz, A.; Hinsch, A.; Hube-Magg, C.; Kluth, M.; Lang, D.S.; Möller-Koop, C.; et al. Apurinic/Apyrimidinic Endonuclease 1 (APE1/Ref-1) Overexpression Is an Independent Prognostic Marker in Prostate Cancer without TMPRSS2:ERG Fusion. Mol Carcinog 2017, 56, 2135–2145. [Google Scholar] [CrossRef]
- Silva, L.P.; Santana, T.; Sedassari, B.T.; de Sousa, S.M.; Sobral, A.P.V.; Freitas, R. de A.; Barboza, C.A.G.; de Souza, L.B. Apurinic/Apyrimidinic Endonuclease 1 (APE1) Is Overexpressed in Malignant Transformation of Salivary Gland Pleomorphic Adenoma. Eur Arch Otorhinolaryngol 2017, 274, 3203–3209. [Google Scholar] [CrossRef]
- Felix, F.A.; da Silva, L.P.; Lopes, M.L.D. de S.; Sobral, A.P.V.; Freitas, R. de A.; de Souza, L.B.; Barboza, C.A.G. DNA Base Excision Repair and Nucleotide Excision Repair Proteins in Malignant Salivary Gland Tumors. Arch Oral Biol 2021, 121, 104987. [Google Scholar] [CrossRef]
- Lee, Y.R.; Lim, J.S.; Shin, J.H.; Choi, S.; Joo, H.K.; Jeon, B.H. Altered Secretory Activity of APE1/Ref-1 D148E Variants Identified in Human Patients With Bladder Cancer. Int Neurourol J 2016, 20, S30–37. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wilson, D.M. Human Apurinic/Apyrimidinic Endonuclease 1. Antioxid. Redox Signal. 2014, 20, 678–707. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Acosta, V.M.; Ruiz-Perez, L.M.; Yang, W.; Gonzalez-Pacanowska, D.; Vidal, A.E. Identification of a Residue Critical for the Excision of 3’-Blocking Ends in Apurinic/Apyrimidinic Endonucleases of the Xth Family. Nucleic Acids Research 2009, 37, 1829–1842. [Google Scholar] [CrossRef] [PubMed]
- Timofeyeva, N.A.; Koval, V.V.; Ishchenko, A.A.; Saparbaev, M.K.; Fedorova, O.S. Lys98 Substitution in Human AP Endonuclease 1 Affects the Kinetic Mechanism of Enzyme Action in Base Excision and Nucleotide Incision Repair Pathways. PLoS ONE 2011, 6, e24063. [Google Scholar] [CrossRef]
- Whitaker, A.M.; Flynn, T.S.; Freudenthal, B.D. Molecular Snapshots of APE1 Proofreading Mismatches and Removing DNA Damage. Nature Communications 2018, 9. [Google Scholar] [CrossRef]
- Wilson, D.M.; Simeonov, A. Small Molecule Inhibitors of DNA Repair Nuclease Activities of APE1. Cellular and Molecular Life Sciences 2010, 67, 3621–3631. [Google Scholar] [CrossRef] [PubMed]
- Al-Safi, R.I.; Odde, S.; Shabaik, Y.; Neamati, N. Small-Molecule Inhibitors of APE1 DNA Repair Function: An Overview. Curr Mol Pharmacol. 2012, 5, 14–35. [Google Scholar] [CrossRef]
- Laev, S.S.; Salakhutdinov, N.F.; Lavrik, O.I. Inhibitors of Nuclease and Redox Activity of Apurinic/Apyrimidinic Endonuclease 1/Redox Effector Factor 1 (APE1/Ref-1). Bioorg Med Chem 2017, 25, 2531–2544. [Google Scholar] [CrossRef]
- Caston, R.A.; Gampala, S.; Armstrong, L.; Messmann, R.A.; Fishel, M.L.; Kelley, M.R. The Multifunctional APE1 DNA Repair-Redox Signaling Protein as a Drug Target in Human Disease. Drug Discov Today 2021, 26, 218–228. [Google Scholar] [CrossRef]
- Xue, Z.; Demple, B. Knockout and Inhibition of Ape1: Roles of Ape1 in Base Excision DNA Repair and Modulation of Gene Expression. Antioxidants (Basel) 2022, 11, 1817. [Google Scholar] [CrossRef] [PubMed]
- Liuzzi, M.; Weinfeld, M.; Paterson, M.C. Selective Inhibition by Methoxyamine of the Apurinic/Apyrimidinic Endonuclease Activity Associated with Pyrimidine Dimer-DNA Glycosylases from Micrococcus Luteus and Bacteriophage T4. Biochemistry 1987, 26, 3315–3321. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Gerson, S.L. Therapeutic Impact of Methoxyamine: Blocking Repair of Abasic Sites in the Base Excision Repair Pathway. Curr Opin Investig Drugs 2004, 5, 623–627. [Google Scholar] [PubMed]
- Liu, L.; Nakatsuru, Y.; Gerson, S.L. Base Excision Repair as a Therapeutic Target in Colon Cancer. Clin Cancer Res 2002, 8, 2985–2991. [Google Scholar] [PubMed]
- Fishel, M.L.; He, Y.; Smith, M.L.; Kelley, M.R. Manipulation of Base Excision Repair to Sensitize Ovarian Cancer Cells to Alkylating Agent Temozolomide. Clin Cancer Res 2007, 13, 260–267. [Google Scholar] [CrossRef]
- Bases, R. Enhancement of X-Ray Damage in HeLa Cells by Exposure to Lucanthone (Miracil D) Following Radiation. Cancer Res 1970, 30, 2007–2011. [Google Scholar]
- Luo, M.; Kelley, M.R. Inhibition of the Human Apurinic/Apyrimidinic Endonuclease (Ape1) Repair Activity and Sensitization of Breast Cancer Cells to DNA Alkylating Agents with Lucanthone. Anticancer Research 2004, 24, 2127–2134. [Google Scholar]
- Naidu, M.D.; Agarwal, R.; Pena, L.A.; Cunha, L.; Mezei, M.; Shen, M.; Wilson, D.M.; Liu, Y.; Sanchez, Z.; Chaudhary, P.; et al. Lucanthone and Its Derivative Hycanthone Inhibit Apurinic Endonuclease-1 (APE1) by Direct Protein Binding. PLoS ONE 2011, 6, e23679. [Google Scholar] [CrossRef] [PubMed]
- Madhusudan, S.; Smart, F.; Shrimpton, P.; Parsons, J.L.; Gardiner, L.; Houlbrook, S.; Talbot, D.C.; Hammonds, T.; Freemont, P.A.; Sternberg, M.J.E.; et al. Isolation of a Small Molecule Inhibitor of DNA Base Excision Repair. Nucleic Acids Research 2005, 33, 4711–4724. [Google Scholar] [CrossRef] [PubMed]
- Simeonov, A.; Kulkarni, A.; Dorjsuren, D.; Jadhav, A.; Shen, M.; McNeill, D.R.; Austin, C.P.; Wilson, D.M. Identification and Characterization of Inhibitors of Human Apurinic/Apyrimidinic Endonuclease APE1. PLoS ONE 2009, 4, e5740. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Fu, D.; Xu, Y.; Wang, X.; Deng, X.; Zhou, S.; Du, F.; Cui, X.; Deng, Y.; Tang, Z. Pt(IV) Prodrug as a Potential Antitumor Agent with APE1 Inhibitory Activity. J Med Chem 2022, 65, 15344–15357. [Google Scholar] [CrossRef] [PubMed]
- Seiple, L.A.; Cardellina, J.H.; Akee, R.; Stivers, J.T. Potent Inhibition of Human Apurinic/Apyrimidinic Endonuclease 1 by Arylstibonic Acids. Mol Pharmacol 2008, 73, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Zawahir, Z.; Dayam, R.; Deng, J.; Pereira, C.; Neamati, N. Pharmacophore Guided Discovery of Small-Molecule Human Apurinic/Apyrimidinic Endonuclease 1 Inhibitors. J Med Chem 2009, 52, 20–32. [Google Scholar] [CrossRef]
- Bapat, A.; Glass, L.S.; Luo, M.; Fishel, M.L.; Long, E.C.; Georgiadis, M.M.; Kelley, M.R. Novel Small-Molecule Inhibitor of Apurinic/Apyrimidinic Endonuclease 1 Blocks Proliferation and Reduces Viability of Glioblastoma Cells. Journal of Pharmacology and Experimental Therapeutics 2010, 334, 988–998. [Google Scholar] [CrossRef]
- Aiello, F.; Shabaik, Y.; Esqueda, A.; Sanchez, T.W.; Grande, F.; Garofalo, A.; Neamati, N. Design and Synthesis of 3-Carbamoylbenzoic Acid Derivatives as Inhibitors of Human Apurinic/Apyrimidinic Endonuclease 1 (APE1). ChemMedChem 2012, 7, 1825–1839. [Google Scholar] [CrossRef]
- Ruiz, F.M.; Francis, S.M.; Tintoré, M.; Ferreira, R.; Gil-Redondo, R.; Morreale, A.; Ortiz, Á.R.; Eritja, R.; Fàbrega, C. Receptor-Based Virtual Screening and Biological Characterization of Human Apurinic/Apyrimidinic Endonuclease (Ape1) Inhibitors. ChemMedChem 2012, 7, 2168–2178. [Google Scholar] [CrossRef]
- Dorjsuren, D.; Kim, D.; Vyjayanti, V.N.; Maloney, D.J.; Jadhav, A.; Wilson, D.M.; Simeonov, A. Diverse Small Molecule Inhibitors of Human Apurinic/Apyrimidinic Endonuclease APE1 Identified from a Screen of a Large Public Collection. PLoS ONE 2012, 7, e47974. [Google Scholar] [CrossRef] [PubMed]
- Rai, G.; Vyjayanti, V.N.; Dorjsuren, D.; Simeonov, A.; Jadhav, A.; Wilson, D.M.; Maloney, D.J. Synthesis, Biological Evaluation, and Structure–Activity Relationships of a Novel Class of Apurinic/Apyrimidinic Endonuclease 1 Inhibitors. Journal of Medicinal Chemistry 2012, 55, 3101–3112. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, A.; Wang, L.; Cline, C.J.; Xie, Z.; Sobol, R.W.; Xie, X.-Q.; Gold, B. Identification and Characterization of Human Apurinic/Apyrimidinic Endonuclease-1 Inhibitors; American Chemical Society, 2012; Vol. 51; ISBN 00062960.
- Feng, Z.; Kochanek, S.; Close, D.; Wang, L.; Srinivasan, A.; Almehizia, A.A.; Iyer, P.; Xie, X.-Q.; Johnston, P.A.; Gold, B. Design and Activity of AP Endonuclease-1 Inhibitors. J Chem Biol 2015, 8, 79–93. [Google Scholar] [CrossRef]
- Trilles, R.; Beglov, D.; Chen, Q.; He, H.; Wireman, R.; Reed, A.; Chennamadhavuni, S.; Panek, J.S.; Brown, L.E.; Vajda, S.; et al. Discovery of Macrocyclic Inhibitors of Apurinic/Apyrimidinic Endonuclease 1. J Med Chem 2019, 62, 1971–1988. [Google Scholar] [CrossRef] [PubMed]
- Pidugu, L.S.; Servius, H.W.; Sevdalis, S.E.; Cook, M.E.; Varney, K.M.; Pozharski, E.; Drohat, A.C. Characterizing Inhibitors of Human AP Endonuclease 1. PLoS One 2023, 18, e0280526. [Google Scholar] [CrossRef]
- Masani, S.; Han, L.; Yu, K. Apurinic/Apyrimidinic Endonuclease 1 Is the Essential Nuclease during Immunoglobulin Class Switch Recombination. Molecular and Cellular Biology 2013, 33, 1468–1473. [Google Scholar] [CrossRef]
- Georgiadis, M.M.; Luo, M.; Gaur, R.K.; Delaplane, S.; Li, X.; Kelley, M.R. Evolution of the Redox Function in Mammalian Apurinic/Apyrimidinic Endonuclease. Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis 2008, 643, 54–63. [Google Scholar] [CrossRef]
- Ando, K.; Hirao, S.; Kabe, Y.; Ogura, Y.; Sato, I.; Yamaguchi, Y.; Wada, T.; Handa, H. A New APE1/Ref-1-Dependent Pathway Leading to Reduction of NF- B and AP-1, and Activation of Their DNA-Binding Activity. Nucleic Acids Research 2008, 36, 4327–4336. [Google Scholar] [CrossRef]
- Logsdon, D.P.; Grimard, M.; Luo, M.; Shahda, S.; Jiang, Y.; Tong, Y.; Yu, Z.; Zyromski, N.; Schipani, E.; Carta, F.; et al. Regulation of HIF1α under Hypoxia by APE1/Ref-1 Impacts CA9 Expression: Dual Targeting in Patient-Derived 3D Pancreatic Cancer Models. Mol Cancer Ther 2016, 15, 2722–2732. [Google Scholar] [CrossRef]
- Ray, S.; Lee, C.; Hou, T.; Bhakat, K.K.; Brasier, A.R. Regulation of Signal Transducer and Activator of Transcription 3 Enhanceosome Formation by Apurinic/Apyrimidinic Endonuclease 1 in Hepatic Acute Phase Response. Molecular Endocrinology 2010, 24, 391–401. [Google Scholar] [CrossRef]
- Cardoso, A.A.; Jiang, Y.; Luo, M.; Reed, A.M.; Shahda, S.; He, Y.; Maitra, A.; Kelley, M.R.; Fishel, M.L. APE1/Ref-1 Regulates STAT3 Transcriptional Activity and APE1/Ref-1–STAT3 Dual-Targeting Effectively Inhibits Pancreatic Cancer Cell Survival. PLoS ONE 2012, 7, e47462. [Google Scholar] [CrossRef]
- Jayaraman, L.; Murthy, K.G.; Zhu, C.; Curran, T.; Xanthoudakis, S.; Prives, C. Identification of Redox/Repair Protein Ref-1 as a Potent Activator of P53. Genes Dev. 1997, 11, 558–570. [Google Scholar] [CrossRef] [PubMed]
- Fishel, M.L.; Wu, X.; Devlin, C.M.; Logsdon, D.P.; Jiang, Y.; Luo, M.; He, Y.; Yu, Z.; Tong, Y.; Lipking, K.P.; et al. Apurinic/Apyrimidinic Endonuclease/Redox Factor-1 (APE1/Ref-1) Redox Function Negatively Regulates NRF2. Journal of Biological Chemistry 2015, 290, 3057–3068. [Google Scholar] [CrossRef]
- Tell, G.; Scaloni, A.; Pellizzari, L.; Formisano, S.; Pucillo, C.; Damante, G. Redox Potential Controls the Structure and DNA Binding Activity of the Paired Domain. Journal of Biological Chemistry 1998, 273, 25062–25072. [Google Scholar] [CrossRef] [PubMed]
- Evans, A.R.; Limp-Foster, M.; Kelley, M.R. Going APE over Ref-1. Mutation Research/DNA Repair 2000, 461, 83–108. [Google Scholar] [CrossRef] [PubMed]
- Xanthoudakis, S.; Curran, T. Identification and Characterization of Ref-1, a Nuclear Protein That Facilitates AP-1 DNA-Binding Activity. The EMBO Journal 1992, 11, 653–665. [Google Scholar] [CrossRef]
- Xanthoudakis, S.; Miao, G.; Wang, F.; Pan, Y.C.; Curran, T. Redox Activation of Fos-Jun DNA Binding Activity Is Mediated by a DNA Repair Enzyme. The EMBO Journal 1992, 11, 3323–3335. [Google Scholar] [CrossRef]
- Arlt, A.; Vorndamm, J.; Breitenbroich, M.; Fölsch, U.R.; Kalthoff, H.; Schmidt, W.E.; Schäfer, H. Inhibition of NF-ΚB Sensitizes Human Pancreatic Carcinoma Cells to Apoptosis Induced by Etoposide (VP16) or Doxorubicin. Oncogene 2001, 20, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Arlt, A.; Gehrz, A.; Müerköster, S.; Vorndamm, J.; Kruse, M.-L.; Fölsch, U.R.; Schäfer, H. Role of NF-ΚB and Akt/PI3K in the Resistance of Pancreatic Carcinoma Cell Lines against Gemcitabine-Induced Cell Death. Oncogene 2003, 22, 3243–3251. [Google Scholar] [CrossRef]
- Raffoul, J.J.; Heydari, A.R.; Hillman, G.G. DNA Repair and Cancer Therapy: Targeting APE1/Ref-1 Using Dietary Agents. Journal of Oncology 2012, 2012, 1–11. [Google Scholar] [CrossRef]
- Gupta, S.C.; Patchva, S.; Aggarwal, B.B. Therapeutic Roles of Curcumin: Lessons Learned from Clinical Trials. AAPS J 2013, 15, 195–218. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhong, C.; Wang, Q.; Chen, W.; Yuan, Y. Curcumin Is an APE1 Redox Inhibitor and Exhibits an Antiviral Activity against KSHV Replication and Pathogenesis. Antiviral Research 2019, 167, 98–103. [Google Scholar] [CrossRef]
- Escobar, I.; Xu, J.; Jackson, C.W.; Stegelmann, S.D.; Fagerli, E.A.; Dave, K.R.; Perez-Pinzon, M.A. Resveratrol Preconditioning Protects Against Ischemia-Induced Synaptic Dysfunction and Cofilin Hyperactivation in the Mouse Hippocampal Slice. Neurotherapeutics 2023. [Google Scholar] [CrossRef]
- Raffoul, J.J.; Banerjee, S.; Singh-Gupta, V.; Knoll, Z.E.; Fite, A.; Zhang, H.; Abrams, J.; Sarkar, F.H.; Hillman, G.G. Down-Regulation of Apurinic/Apyrimidinic Endonuclease 1/Redox Factor-1 Expression by Soy Isoflavones Enhances Prostate Cancer Radiotherapy In Vitro and In Vivo. Cancer Research 2007, 67, 2141–2149. [Google Scholar] [CrossRef]
- Kelley, M.R.; Wikel, J.H.; Guo, C.; Pollok, K.E.; Bailey, B.J.; Wireman, R.; Fishel, M.L.; Vasko, M.R. Identification and Characterization of New Chemical Entities Targeting Apurinic/Apyrimidinic Endonuclease 1 for the Prevention of Chemotherapy-Induced Peripheral Neuropathy. J Pharmacol Exp Ther 2016, 359, 300–309. [Google Scholar] [CrossRef]
- Shimizu, N.; Sugimoto, K.; Tang, J.; Nishi, T.; Sato, I.; Hiramoto, M.; Aizawa, S.; Hatakeyama, M.; Ohba, R.; Hatori, H.; et al. High-Performance Affinity Beads for Identifying Drug Receptors. Nat Biotechnol 2000, 18, 877–881. [Google Scholar] [CrossRef]
- Zou, G.-M.; Karikari, C.; Kabe, Y.; Handa, H.; Anders, R.A.; Maitra, A. The Ape-1/Ref-1 Redox Antagonist E3330 Inhibits the Growth of Tumor Endothelium and Endothelial Progenitor Cells: Therapeutic Implications in Tumor Angiogenesis. J. Cell. Physiol. 2009, 219, 209–218. [Google Scholar] [CrossRef]
- Zou, G.-M.; Maitra, A. Small-Molecule Inhibitor of the AP Endonuclease 1/REF-1 E3330 Inhibits Pancreatic Cancer Cell Growth and Migration. Molecular Cancer Therapeutics 2008, 7, 2012–2021. [Google Scholar] [CrossRef] [PubMed]
- Jiang, A.; Gao, H.; Kelley, M.R.; Qiao, X. Inhibition of APE1/Ref-1 Redox Activity with APX3330 Blocks Retinal Angiogenesis in Vitro and in Vivo. Vision Research 2011, 51, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Manguinhas, R.; Fernandes, A.S.; Costa, J.G.; Saraiva, N.; Camões, S.P.; Gil, N.; Rosell, R.; Castro, M.; Miranda, J.P.; Oliveira, N.G. Impact of the APE1 Redox Function Inhibitor E3330 in Non-Small Cell Lung Cancer Cells Exposed to Cisplatin: Increased Cytotoxicity and Impairment of Cell Migration and Invasion. Antioxidants 2020, 9, 550. [Google Scholar] [CrossRef] [PubMed]
- Guerreiro, P.S.; Corvacho, E.; Costa, J.G.; Saraiva, N.; Fernandes, A.S.; Castro, M.; Miranda, J.P.; Oliveira, N.G. The APE1 Redox Inhibitor E3330 Reduces Collective Cell Migration of Human Breast Cancer Cells and Decreases Chemoinvasion and Colony Formation When Combined with Docetaxel. Chem Biol Drug Des 2017, 90, 561–571. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, X.; Zhou, T.; Kelley, M.R.; Edwards, P.A.; Gao, H.; Qiao, X. Suppression of Choroidal Neovascularization Through Inhibition of APE1/Ref-1 Redox Activity. Invest. Ophthalmol. Vis. Sci. 2014, 55, 4461. [Google Scholar] [CrossRef]
- Hartman, G.D.; Lambert-Cheatham, N.A.; Kelley, M.R.; Corson, T.W. Inhibition of APE1/Ref-1 for Neovascular Eye Diseases: From Biology to Therapy. IJMS 2021, 22, 10279. [Google Scholar] [CrossRef]
- Hu, J.; Wang, Y.; Yuan, Y. Inhibitors of APE1 Redox Function Effectively Inhibit γ-Herpesvirus Replication in Vitro and in Vivo. Antiviral Research 2021, 185, 104985. [Google Scholar] [CrossRef] [PubMed]
- Shahda, S.; Lakhani, N.J.; O’Neil, B.; Rasco, D.W.; Wan, J.; Mosley, A.L.; Liu, H.; Kelley, M.R.; Messmann, R.A. A Phase I Study of the APE1 Protein Inhibitor APX3330 in Patients with Advanced Solid Tumors. JCO 2019, 37, 3097–3097. [Google Scholar] [CrossRef]
- Kelley, M.R.; Luo, M.; Reed, A.; Su, D.; Delaplane, S.; Borch, R.F.; Nyland, R.L.; Gross, M.L.; Georgiadis, M.M. Functional Analysis of Novel Analogues of E3330 That Block the Redox Signaling Activity of the Multifunctional AP Endonuclease/Redox Signaling Enzyme APE1/Ref-1. Antioxidants & Redox Signaling 2011, 14, 1387–1401. [Google Scholar] [CrossRef]
- Nyland, R.L.; Luo, M.; Kelley, M.R.; Borch, R.F. Design and Synthesis of Novel Quinone Inhibitors Targeted to the Redox Function of Apurinic/Apyrimidinic Endonuclease 1/Redox Enhancing Factor-1 (Ape1/Ref-1). Journal of Medicinal Chemistry 2010, 53, 1200–1210. [Google Scholar] [CrossRef]
- Qian, C.; Li, M.; Sui, J.; Ren, T.; Li, Z.; Zhang, L.; Zhou, L.; Cheng, Y.; Wang, D. Identification of a Novel Potential Antitumor Activity of Gossypol as an APE1/Ref-1 Inhibitor. Drug Des Devel Ther 2014, 8, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, X.; Zhang, L.; Li, M.; Dai, N.; Luo, H.; Shan, J.; Yang, X.; Xu, M.; Feng, Y.; et al. A Randomized, Double-Blind, Placebo-Controlled Study of B-Cell Lymphoma 2 Homology 3 Mimetic Gossypol Combined with Docetaxel and Cisplatin for Advanced Non-Small Cell Lung Cancer with High Expression of Apurinic/Apyrimidinic Endonuclease 1. Invest New Drugs 2020, 38, 1862–1871. [Google Scholar] [CrossRef]
- Ren, T.; Shan, J.; Qing, Y.; Qian, C.; Li, Q.; Lu, G.; Li, M.; Li, C.; Peng, Y.; Luo, H.; et al. Sequential Treatment with AT-101 Enhances Cisplatin Chemosensitivity in Human Non-Small Cell Lung Cancer Cells through Inhibition of Apurinic/Apyrimidinic Endonuclease 1-Activated IL-6/STAT3 Signaling Pathway. Drug Des Devel Ther 2014, 8, 2517–2529. [Google Scholar] [CrossRef]
- Wei, X.; Duan, W.; Li, Y.; Zhang, S.; Xin, X.; Sun, L.; Gao, M.; Li, Q.; Wang, D. AT101 Exerts a Synergetic Efficacy in Gastric Cancer Patients with 5-FU Based Treatment through Promoting Apoptosis and Autophagy. Oncotarget 2016, 7, 34430–34441. [Google Scholar] [CrossRef]
- Poletto, M.; Vascotto, C.; Scognamiglio, P.L.; Lirussi, L.; Marasco, D.; Tell, G. Role of the Unstructured N-Terminal Domain of the HAPE1 (Human Apurinic/Apyrimidinic Endonuclease 1) in the Modulation of Its Interaction with Nucleic Acids and NPM1 (Nucleophosmin). Biochem. J. 2013, 452, 545–557. [Google Scholar] [CrossRef]
- Antoniali, G.; Dalla, E.; Mangiapane, G.; Zhao, X.; Jing, X.; Cheng, Y.; De Sanctis, V.; Ayyildiz, D.; Piazza, S.; Li, M.; et al. APE1 Controls DICER1 Expression in NSCLC through MiR-33a and MiR-130b. Cell. Mol. Life Sci. 2022, 79, 446. [Google Scholar] [CrossRef] [PubMed]
- Vascotto, C.; Cesaratto, L.; Zeef, L.A.H.; Deganuto, M.; D’Ambrosio, C.; Scaloni, A.; Romanello, M.; Damante, G.; Taglialatela, G.; Delneri, D.; et al. Genome-Wide Analysis and Proteomic Studies Reveal APE1/Ref-1 Multifunctional Role in Mammalian Cells. PROTEOMICS 2009, 9, 1058–1074. [Google Scholar] [CrossRef]
- Ayyildiz, D.; Antoniali, G.; D’Ambrosio, C.; Mangiapane, G.; Dalla, E.; Scaloni, A.; Tell, G.; Piazza, S. Architecture of The Human Ape1 Interactome Defines Novel Cancers Signatures. Sci Rep 2020, 10, 28. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.M.; Deacon, A.M.; Duncton, M.A.J.; Pellicena, P.; Georgiadis, M.M.; Yeh, A.P.; Arvai, A.S.; Moiani, D.; Tainer, J.A.; Das, D. Fragment- and Structure-Based Drug Discovery for Developing Therapeutic Agents Targeting the DNA Damage Response. Prog Biophys Mol Biol 2021, 163, 130–142. [Google Scholar] [CrossRef]
- Codrich, M.; Degrassi, M.; Malfatti, M.C.; Antoniali, G.; Gorassini, A.; Ayyildiz, D.; De Marco, R.; Verardo, G.; Tell, G. APE1 Interacts with the Nuclear Exosome Complex Protein MTR4 and Is Involved in Cisplatin- and 5-Fluorouracil-Induced RNA Damage Response. The FEBS Journal 2022, febs.16671. [CrossRef]
- Garutti, M.; Pelizzari, G.; Bartoletti, M.; Malfatti, M.C.; Gerratana, L.; Tell, G.; Puglisi, F. Platinum Salts in Patients with Breast Cancer: A Focus on Predictive Factors. IJMS 2019, 20, 3390. [Google Scholar] [CrossRef]
- Dumas, L.; Herviou, P.; Dassi, E.; Cammas, A.; Millevoi, S. G-Quadruplexes in RNA Biology: Recent Advances and Future Directions. Trends in Biochemical Sciences 2021, 46, 270–283. [Google Scholar] [CrossRef]
- Kwok, C.K.; Sahakyan, A.B.; Balasubramanian, S. Structural Analysis Using SHALiPE to Reveal RNA G-Quadruplex Formation in Human Precursor MicroRNA. Angew. Chem. Int. Ed. 2016, 55, 8958–8961. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, J.; Santos, T.; Miranda, A.; Alexandre, D.; Teixeira, B.; Simões, P.; Lopes-Nunes, J.; Cruz, C. Ligands as Stabilizers of G-Quadruplexes in Non-Coding RNAs. Molecules 2021, 26, 6164. [Google Scholar] [CrossRef]
- Koralewska, N.; Szczepanska, A.; Ciechanowska, K.; Wojnicka, M.; Pokornowska, M.; Milewski, M.C.; Gudanis, D.; Baranowski, D.; Nithin, C.; Bujnicki, J.M.; et al. RNA and DNA G-Quadruplexes Bind to Human Dicer and Inhibit Its Activity. Cell. Mol. Life Sci. 2021, 78, 3709–3724. [Google Scholar] [CrossRef] [PubMed]
- Asamitsu, S.; Shioda, N. Potential Roles of G-Quadruplex Structures in RNA Granules for Physiological and Pathological Phase Separation. The Journal of Biochemistry 2021, 169, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, M.; Duncan, S.; Yang, X.; Abdelhamid, M.A.S.; Huang, L.; Zhang, H.; Benfey, P.N.; Waller, Z.A.E.; Ding, Y. G-Quadruplex Structures Trigger RNA Phase Separation. Nucleic Acids Research 2019, 47, 11746–11754. [Google Scholar] [CrossRef] [PubMed]
- Mirihana Arachchilage, G.; Dassanayake, A.C.; Basu, S. A Potassium Ion-Dependent RNA Structural Switch Regulates Human Pre-MiRNA 92b Maturation. Chemistry & Biology 2015, 22, 262–272. [Google Scholar] [CrossRef]
Figure 1.
Human APE1 structure and key residues. Schematic representation of APE1 primary sequence, in which the most important aminoacids are highlighted. In the first 33 residues, required for protein-protein interaction, residues K6 and K7 (green, ο) are involved in the shuttling between the nucleus and cytoplasm. Residues K27, K31, K32 and K35 can be acetylated and are essential for proteosomal cleavage. The redox regulatory region in included between aminoacid 35 to 125 in which the main residues involved are C65, C93, and C99 (red). The endonuclease domain spans between 65 to 318 (residues indicated in blue, •). Specifically, E96 is involved in divalent metal coordination, while D210 and H309 have functions in the hydrolytic reaction. Other important residues in the endonuclease domain are D70, which is implicated in the 3’-phosphodiesterase activity, K98, important for the nucleotide incision repair (NIR) and F266, which is involved in the 3’ to 5’-exonuclease activity. Lastly, P112L, D148E, R193C/H, R237C, and H298Y/Q (yellow, ο) are some of the polymorphisms of APE1 that will be discussed in this Review. Below, the tridimensional structure of APE1 is reported (PDB ID: 7SVB). Created with BioRender.com.
Figure 1.
Human APE1 structure and key residues. Schematic representation of APE1 primary sequence, in which the most important aminoacids are highlighted. In the first 33 residues, required for protein-protein interaction, residues K6 and K7 (green, ο) are involved in the shuttling between the nucleus and cytoplasm. Residues K27, K31, K32 and K35 can be acetylated and are essential for proteosomal cleavage. The redox regulatory region in included between aminoacid 35 to 125 in which the main residues involved are C65, C93, and C99 (red). The endonuclease domain spans between 65 to 318 (residues indicated in blue, •). Specifically, E96 is involved in divalent metal coordination, while D210 and H309 have functions in the hydrolytic reaction. Other important residues in the endonuclease domain are D70, which is implicated in the 3’-phosphodiesterase activity, K98, important for the nucleotide incision repair (NIR) and F266, which is involved in the 3’ to 5’-exonuclease activity. Lastly, P112L, D148E, R193C/H, R237C, and H298Y/Q (yellow, ο) are some of the polymorphisms of APE1 that will be discussed in this Review. Below, the tridimensional structure of APE1 is reported (PDB ID: 7SVB). Created with BioRender.com.
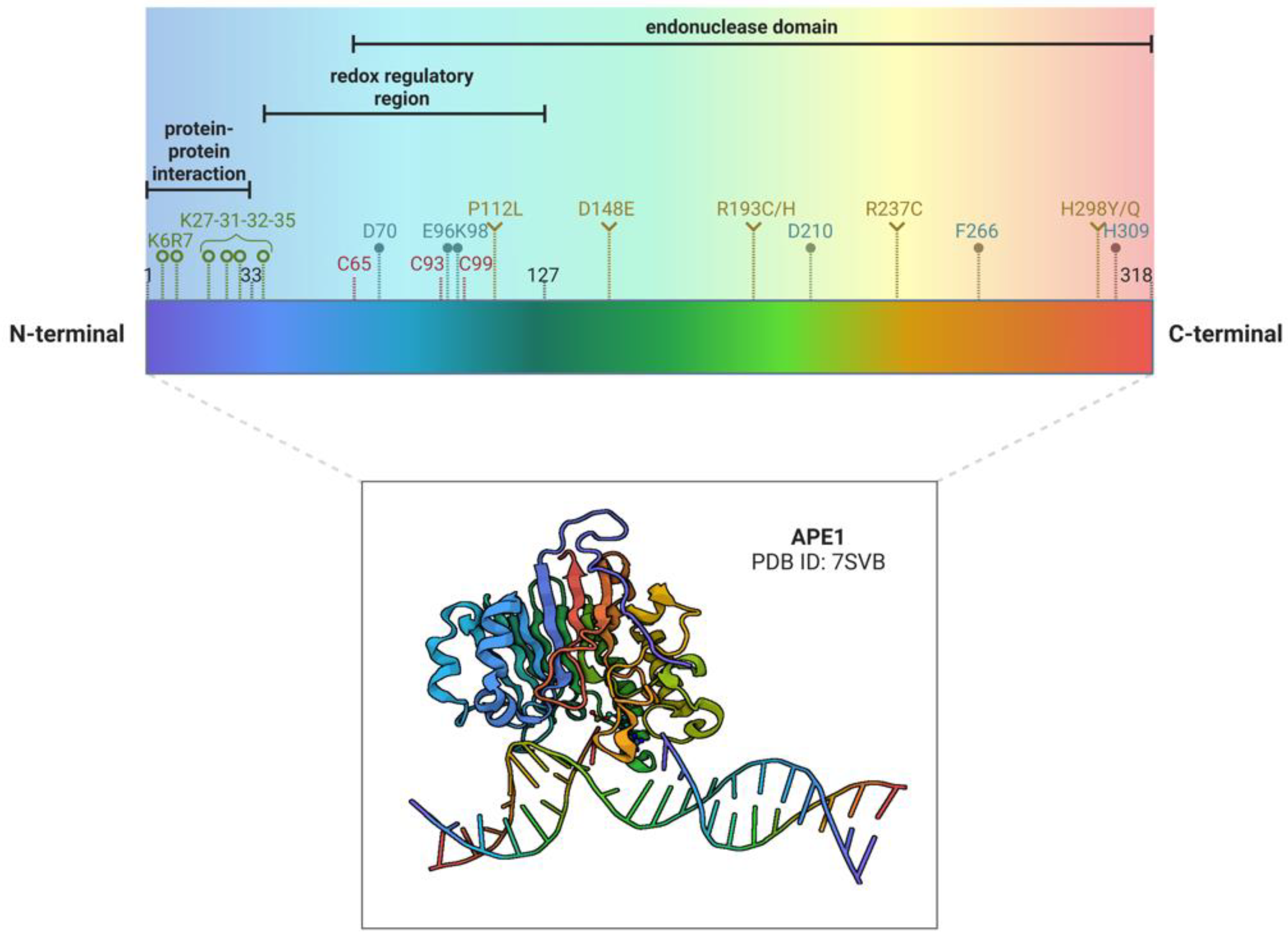
Figure 2.
Illustration of the main functions of APE1 and relative inhibitors. The DNA repair mediated by the endonuclease activity of APE1 is inhibited by Methoxyamine, APE Inhibitor III, and Gossypol (blue blunt arrows). The gene expression promoted by the redox activity of APE1 on transcriptional factors (TFs) is inhibited by Gossypol, Curcumin, Resveratrol, and APX3330 (green blunt arrows). Especially through its N-terminal region, APE1 is involved in several PPI, including Nucleophosmin 1 (NPM1). This interaction is inhibited by Fiduxosin, SB 206553, and Spiclomazine (fuchsia blunt arrows). Recent findings have shown that APE1 is secreted by EVs in the microambient and is involved in RNA metabolism, including the regulation of miRNA processing. If these novel features of APE1 can be inhibited is still unknown (brown blunt arrows) and an interesting starting point for future explorations. For each inhibitor, pillows are drawn when the inhibitor has been enrolled in one or more clinical trials. Created with BioRender.com.
Figure 2.
Illustration of the main functions of APE1 and relative inhibitors. The DNA repair mediated by the endonuclease activity of APE1 is inhibited by Methoxyamine, APE Inhibitor III, and Gossypol (blue blunt arrows). The gene expression promoted by the redox activity of APE1 on transcriptional factors (TFs) is inhibited by Gossypol, Curcumin, Resveratrol, and APX3330 (green blunt arrows). Especially through its N-terminal region, APE1 is involved in several PPI, including Nucleophosmin 1 (NPM1). This interaction is inhibited by Fiduxosin, SB 206553, and Spiclomazine (fuchsia blunt arrows). Recent findings have shown that APE1 is secreted by EVs in the microambient and is involved in RNA metabolism, including the regulation of miRNA processing. If these novel features of APE1 can be inhibited is still unknown (brown blunt arrows) and an interesting starting point for future explorations. For each inhibitor, pillows are drawn when the inhibitor has been enrolled in one or more clinical trials. Created with BioRender.com.
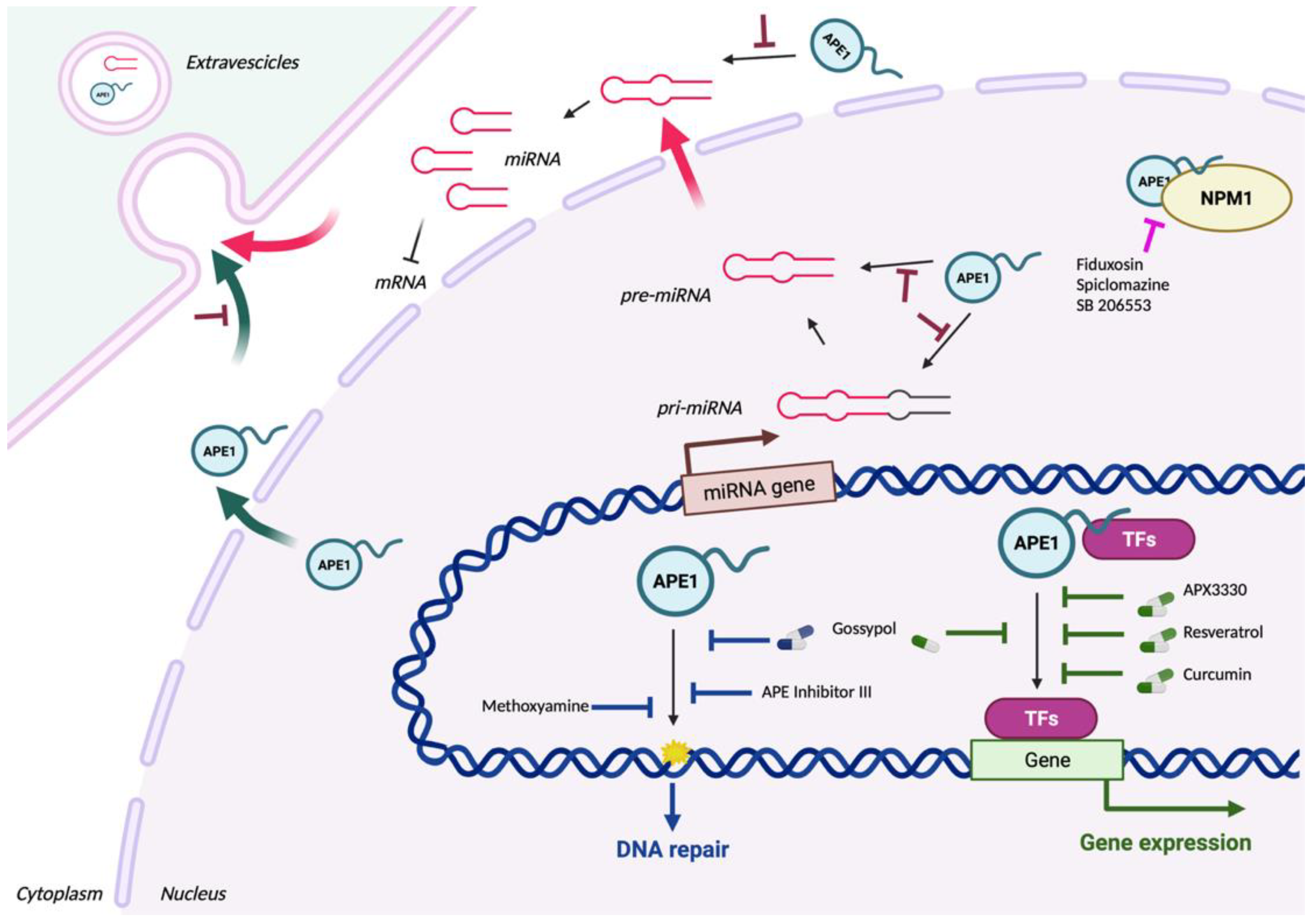

Table 1.
APE1 polymorphisms in different cancer types were detected with the cBioPortal tool.
| Cancer | Polymorphisms |
|---|---|
| Adrenocortical carcinoma | D283H; X20_splice |
| Ampullary cancer | M271T |
| Bladder cancer | D15H; S146L; Q109E; E217* |
| Bone cancer | V131M |
| Breast cancer | L244Tfs*8; Q51*; H289Y; G5A; K7Rfs*75; R185W; A60G |
| Cervical cancer | R281C; K7R |
| Colorectal cancer | A273T; R193C; R221H; G130D; R247Q; A230D; M271I; R221H; R181*; E242D; R274*; P293S; P49Qfs*33; L220I; T233M; N226Efs*26; V131M; L291Vfs*6; R221C; K63E; D210N; R281H; X82_splice; E101D; K77N; P331H; G306S; G132D |
| Endometrial cancer | R193H; S164L; R221H; A170V; P223L; V84I; V278D; R281H; K228T |
| Gastric cancer | R281C; P122A; K27N |
| Glioma | H289Q; Q245R; G80E; D142N |
| Head and Neck cancer | P48H |
| Leukaemia and Lymphoma | R181Q; L17P; K7R; R187H |
| Liver cancer | G5E; W280S; L220P; W280R; D50Rfs*28; G8R; H289Y, N226Efs*26; Y264_G279del |
| Lung cancer | G41C; V206Cfs*11; V142Sfs*8; E16Q; P331T; E149Q; X147_splice; R177*; D90H; Q51*;X237_splice; R193C; R28S; M271del; V206Cfs*11; S115F; G8R; I146V; D148E |
| Melanoma | G241W; E16K; G127V; R136S; P122T; D148E; A263V; K7Rfs*75; L108F; V69L |
| Oesophageal cancer | E46D; D251N; M270Nfs*14; F165V; N102I; L291Vfs*6; K3R; G145D |
| Ovarian cancer | Q95*, V168I; R193H; L291Vfs*6 |
| Pancreatic cancer | R193C; M271del |
| Prostate cancer | P139Q; A30T; R187H |
| Renal cancer | E149* |
| Sarcoma | R187L; K35Rfs*11; K35Q |
| Skin cancer | P89S |
Table 2.
APE1 dysregulation in tumours: from its expression and distribution to its value as a diagnostic biomarker.
Table 2.
APE1 dysregulation in tumours: from its expression and distribution to its value as a diagnostic biomarker.
| Cancer | Expression | Diagnostic value | Localization | Refs |
|---|---|---|---|---|
| Bladder cancer (BCa) |
Protein overexpression, associated with poor survival and invasion. | Serum and urine levels as a diagnostic biomarker. |
|
[36,37,38,39,40,41,42] |
| Breast cancer |
|
n.d. | Nuclear localization. | [43,44,45,46,47,48] |
| Cervical cancer |
High protein expression associated with lymph node metastasis, EMT and decreasing radiosensitivity. | n.d. |
|
[49,50,51,52] |
| Colorectal cancer (CRC) |
|
Serum APE1-autoantibodies levels as a diagnostic biomarker. |
|
[53,54,55,56,57,58,59,60] |
| Cutaneous Squamous Cell carcinoma (cSCC) |
Protein overexpression in tumour tissues, which promotes proliferation of cancer cells and migration by EMT. | n.d. | n.d. | [61] |
| Gastric carcinoma |
mRNA and protein upregulation, correlated with lymph node metastasis, depth of invasion and poor prognosis. | Serum levels as a diagnostic biomarker for metastasis prediction. | Nuclear and cytoplasmatic localization. | [62,63,64,65] |
| Glioma |
|
n.d. | Nuclear localization. | [66,67,68,69,70,71] |
| Head and Neck Squamous Cell carcinoma (HNSCC) |
|
In oral SCC (oSCC): Serum levels as a diagnostic biomarker, with high levels correlated with late TNM stages, lymph node metastasis and worse pathologic differentiation. |
higher cytoplasmatic staining in sSCC, associated with T-stage and histological grade.
|
[72,73,74,75,76,77,78] |
| Liver cancer |
|
Serum levels as a diagnostic biomarker. |
|
[79,80,81,82,83,84] |
| Lung cancer |
|
High post-treatment serum levels are associated with lower overall survival. |
|
[53,85,86,87,88,89,90,91,92,93,94] |
| Melanoma | mRNA and protein overexpression associated with vascular invasion, high mitotic rate, lower response to therapy and poor prognosis. | n.d. | Nuclear localization. | [95,96,97,98] |
| Oesophageal carcinoma (EAC) | Protein overexpression in tumour tissues, associated with worse overall survival. | n.d. | Mainly nuclear localization. | [99,100,101,102,103,104] |
| Osteosarcoma |
|
n.d. |
|
[105,106,107,108,109,110,111] |
| Ovarian cancer |
Protein overexpression in tumour tissues, associated with advanced stages, platinum resistance and poor chemosensitivity, decreased overall survival and lymph node metastasis. | n.d. |
|
[99,112,113,114,115,116,117,118,119] |
| Pancreatic adenocarcinoma (PDAC) |
|
n.d. | Strong nuclear staining in tumour tissues, with cytosol staining only in advanced stages. | [53,85,99,120,121,122] |
| Prostate carcinoma (PCa) |
Protein overexpression in cancer samples. | n.d. | Nuclear and cytoplasmatic localization. | [123,124] |
| Salivary gland carcinoma |
|
n.d. | Mainly nuclear staining, with nuclear and cytosolic staining in some malignant forms. | [125,126] |
Table 3.
List of the principal APE1 inhibitors. The principal APE1 inhibitors are grouped for their APE1 function inhibited including the endonuclease activity, the redox activity, and the protein-protein interaction. For each inhibitor, the IUPAC name, the PubChem CID, the molecular formula and weight, and the structure are reported. In the end, the main references for each inhibitor are indicated. For more detailed information, refer to the text.
Table 3.
List of the principal APE1 inhibitors. The principal APE1 inhibitors are grouped for their APE1 function inhibited including the endonuclease activity, the redox activity, and the protein-protein interaction. For each inhibitor, the IUPAC name, the PubChem CID, the molecular formula and weight, and the structure are reported. In the end, the main references for each inhibitor are indicated. For more detailed information, refer to the text.
| APE1 function inhibited | Name | IUPAC name | PubChem CID | Molecular Formula | Molecular Weight (g/mol) | Structure | Refs |
|---|---|---|---|---|---|---|---|
| Endonuclease | Methoxyamine | O-methylhydroxylamine | 4113 | CH5NO | 47.057 | 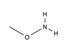 |
[137,138,139,140] |
| Lucanthone | 1-[2-(diethylamino)ethylamino]-4-methylthioxanthen-9-one | 10180 | C20H24N2OS | 340.5 |  |
[141,142,143] | |
| CRT0044867 | 7-Nitroindole-2-carboxylic acid | 81409 | C9H6N2O4 | 206.15 | 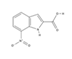 |
[21,144,145,146] | |
| Myricetin | 3,5,7-trihydroxy-2-(3,4,5-trihydroxyphenyl)chromen-4-one | 5281672 | C15H10O8 | 318.23 |  |
[145] | |
| AR03 | 2,4,9-Trimethylbenzo[b][1,8]naphthyridin-5-amine | 698490 | C15H15N3 | 237.30 |  |
[149] | |
| APE Inhibitor III | N-[3-(1,3-benzothiazol-2-yl)-6-isopropyl-4,5,6,7-tetrahydrothieno [2,3-c]pyridin-2-yl]acetamide | 3581333 | C19H21N3OS2 | 371.5 |  |
[153,154] | |
| Redox | Curcumin | (1E,6E)-1,7-bis(4-hydroxy-3-methoxyphenyl)hepta-1,6-diene-3,5-dione | 969516 | C21H20O6 | 368.4 | 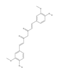 |
[173,174] |
| Resveratrol | 5-[(E)-2-(4-hydroxyphenyl)ethenyl]benzene-1,3-diol | 445154 | C14H12O3 | 228.24 | 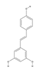 |
[97,175] | |
| APX3330 | (2E)-2-[(4,5-dimethoxy-2-methyl-3,6-dioxocyclohexa-1,4-dien-1-yl)methylidene]undecanoic acid | 6439397 | C21H30O6 | 378.5 | 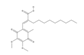 |
[22,177,178,179,180,181,182,183,184,185,186,187] | |
| APX2009 | (2E)-N,N-diethyl-2-[(3-methoxy-1,4-dioxonaphthalen-2-yl)methylidene]pentanamide | 71618575 | C21H25NO4 | 355.4 | 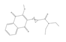 |
[177] | |
| Endonuclease Redox | Gossypol | 7-(8-formyl-1,6,7-trihydroxy-3-methyl-5-propan-2-ylnaphthalen-2-yl)-2,3,8-trihydroxy-6-methyl-4-propan-2-ylnaphthalene-1-carbaldehyde | 3503 | C30H30O8 | 518.6 | 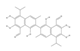 |
[190,191] |
| protein-protein interaction | Fiduxosin | 5-[4-[(3aR,9bR)-9-methoxy-3,3a,4,9b-tetrahydro-1H-chromeno[3,4-c]pyrrol-2-yl]butyl]-12-phenyl-8-thia-3,5,10,13-tetrazatricyclo[7.4.0.02,7]trideca-1(13),2(7),9,11-tetraene-4,6-dione | 172307 | C30H29N5O4S | 555.6 |  |
[47,116,200] |
| Spiclomazine | 8-[3-(2-chlorophenothiazin-10-yl)propyl]-1-thia-4,8-diazaspiro[4.5]decan-3-one | 65714 | C22H24ClN3OS2 | 446.0 |  |
[47,116,200] | |
| SB 206553 | 1-methyl-N-pyridin-3-yl-6,7-dihydropyrrolo[2,3-f]indole-5-carboxamide | 5163 | C17H16N4O | 292.33 |  |
[47,116,200] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



