
Submitted:
30 May 2023
Posted:
01 June 2023
You are already at the latest version
Abstract
MicroRNAs (miRNAs) are single-stranded, noncoding RNA molecules that regulate gene expression post-transcriptionally by binding to messenger RNAs. miRNAs are important regulators of gene expression, and their dysregulation is implicated in many human and dog diseases. Most cancers tested, to date, have been shown to express altered miRNA levels, which indicates their potential importance in the oncogenic process. Based on this evidence, numerous miRNAs have been suggested as potential cancer biomarkers for both diagnosis and prognosis. miRNA-based therapies have also been tested in different cancers and have provided measurable clinical benefits to patients. In addition, understanding miRNA biogenesis and regulatory mechanisms in cancer can provide important knowledge about resistance to chemotherapies, leading to more personalized cancer treatment. In this review, we have comprehensively summarized the importance of miRNA in human and dog cancer research. We have discussed the current state of development and potential for miRNA as both a diagnostic marker and a therapeutic target.
Keywords:
miRNAs
; Cancer
; oncomiRs
; tumor suppressor miRNAs
; biomarker
; therapeutics
1. Introduction
miRNAs are small non-coding RNA sequences with an average length of 18-22 bps. To date, 2654 mature miRNAs have been reported in humans [1]. miRNAs play an essential role in biological processes by regulating gene expression at a post-transcription level. miRNAs bind to messenger RNA (mRNA) in the cytoplasm resulting in mRNA degradation or temporary inhibition of translation until needed [2]. Downregulation of a specific miRNA will lead to upregulation of the corresponding proteins’ expression and vice-versa. Conversely, upregulation of miRNA leads to decreased expression of the target protein(s). miRNAs bind at the 3’ UTR, 5’ UTR, and coding regions of mRNA to induce translation repression. miRNAs are also involved in inducing gene transcription by binding within the promoter regions of a gene [3]. miRNA are typically found inside cells; however, a portion of them are shed into the circulation in lipid-coated particles known as exosomes [4]. Circulatory exosomal miRNAs have been identified as possible disease biomarkers, as they are stable in blood and are protected from endogenous RNAse activity [5].
miRNAs play an important role in cancer cell transformation. miRNAs can function as tumor suppressor genes or oncogenes by targeting genes involved in tumor development and progression or cell cycle inhibition, respectively. Since the discovery of microRNAs, they have held great promise for diagnosis, prognosis, and therapy in cancer. Different miRNA profiles can be identified for different tumor types, which could then serve as phenotypic signatures for exploitation in cancer diagnosis, prognosis, and treatment. If miRNA profiles can accurately predict malignancies, this technology could be used as a tool to overcome many diagnostic challenges [6]. In this review we have evaluated the role of miRNA in cancer diagnosis and treatment in humans and dogs.
2. Role of miRNAs in cancer
2.1. Humans:
2.1.1. OncomiRs
OncomiRs are defined as miRNAs that are over-expressed in tumors, repress tumor suppressor mRNAs, and stimulate tumor cell proliferation and metastasis (Figure 1) [7]. There are many different oncomiRNAs with different roles in cancer growth that have been identified so far.
The miR-17-92 cluster (miRs-17, -18a, -19a, -20a, -19b, and -92a) downregulates PTEN (Phosphate and tensin homolog), E2F, the TGF-B signaling pathway, BCL2L11, and thrombospondin-1 (TSP-1) [8]. Functionally it favors tumor growth and is reported as overexpressed in small cell lung cancer, colon cancer, hepatocellular carcinoma, thyroid cancer, colorectal adenoma organoids, and renal cell carcinoma [9,10,11,12,13,14].
miR-21 is associated with Phosphatase and Tensin Homolog (PTEN), Tropomyosin 1 (TPM1), and Programmed Cell Death 4 (PDCD4) downregulation. miR21 overexpression is reported in variety of cancers, such as breast, ovarian, colon, etc. [15,16,17]. Elevated levels of miR-21 were also reported in the serum, plasma, and tumor tissues in breast, lung, ovarian, colon, prostate, pancreatic, and gastric cancer patients [18,19,20,21,22,23,24,25,26,27]. Downregulation of miR-21 reduces cancer proliferation and reverses drug resistance in pancreatic, ovarian, and breast cancers [25,26,27].
miR-181 is an onco-miR that is also up-regulated in a variety of cancer types [28]. miR-181a-5p promotes breast tumor progression through NDRG2-induced activation of the PTEN/AKT signaling pathway and inhibition of SPRY4, PHLPP2, and INPP4B [29,30,31]. miR-181 facilitates prostate cancer cell proliferation by targeting DAX-1 [32]. Similarly, miR-181 upregulation is associated with poor prognosis and survival in oral squamous cell carcinoma and drug resistance in the melanoma [33]. miR-146a is significantly higher in plasma samples from breast cancer patients [34]. miR-221/222 is overexpressed in liver tumorigenesis, breast, colon, and pancreatic tumor [22,35,36,37].
2.1.1. Tumor suppressor miRNAs (TS-miRNAs)
Ts-miRNSa are defined as miRNAs that downregulates cancer progression (Figure 1). The downregulation of tumor suppressor miRNAs plays a crucial role in cancer development and proliferation [38]. TS-miRNAs are more susceptible to mutations due to their location in cancer-associated genomic regions or fragile sites. Downregulation of TS-miRNAs may occur due to dysfunctional proteins involved in their biogenesis or due to any genetic alteration [39]. Inhibition of the expression of important miRNA biogenesis machineries such as Drosha, DCGR8, and Dicer substantially decreases miRNA production and promotes a more transformed cell phenotype [40,41,42,43,44,45].
Loss of TS-miRNA miR-16 is correlated with the progression and expansion of chronic lymphocytic leukemia, gastric, prostate, and pancreatic tumors [46,47,48,49,50]. Let-7 is a tumor suppressor miRNA that targets the Rasa and Myc oncogenes [51]. Ectopic expression of Let-7 induced cell death in lung cancer cells [52]. Let-7 is also reported to target other oncogenes such as HMGA2 and MYCN [53]. Let-7 also acts as a tumor suppressor in breast cancer by inhibiting ERα-mediated cellular malignant growth [54].
miR-29 and miR-34 are tumor suppressor miRNAs whose downregulated expression is associated with the progression and invasion of breast, lung cancer, neuroblastoma and glioblastoma, colon, stomach, osteoblastoma, ovarian, bladder, cervical, cholangiocarcinoma, melanoma, and prostate cancer [55,56,57,58,59]. miR-29 downregulation is also associated with cisplatin resistance in ovarian cancer and elevated cell proliferation in osteosarcoma [60,61,62]. Downregulation of miR-34 is associated with proliferation in pancreatic, lung squamous cell carcinoma, head and neck cancer, colorectal, gastric, and epithelial ovarian cancer [63,64,65,66,67,68]. Elevated expression of miR-362-3p interrupts the cell cycle and inhibits tumor growth resulting in an improved prognosis in colorectal carcinoma patients [69].
Upregulated miR-193b expression results in reduced fatty acid synthase (FASN), which in turn makes triple-negative breast cancer cells more sensitive to the effects of metformin [70]. The expression of miRNA-193b acts as a tumor suppressor in pancreatic cancer and is markedly reduced in tissues with advanced neoplasia. Cell lines transfected with miRNA-193b exhibited significantly decreased proliferation, migration, and invasiveness [71].
The impact of miRNA polymorphisms and their associated impact on cancer risk has been studied [72,73]. SNPs rs3746444 in miR-499 and rs4919510 in miR-608 are significantly associated with an increased risk of lung cancer [74]. A SNP in miRNA-499 increases the risk of prostate cancer [72]. X inactivation-specific transcript (XIST) is a carcinogenic long coding-RNA that is involved in ovarian tumor progression by regulating miR-355/BCL2L2 [75].
2.2. Dogs
Dogs have high similarity to humans in gene sequence and thus gene function. Dogs share the same environmental exposures and risks as humans. Almost 50% of dogs, 10 years old or older, are diagnosed with cancer at some point in their lives [76]. Due to these similarities, dogs are excellent translational models for complex human diseases, such as cancer. As in humans, miRNAs play an important role in canine cancer.
Up-regulation of miRNA-19a, -19b, -17, -5p were reported in T and B cell lymphomas in dogs [77]. Additionally, miRNA-203, -181a, and -218 were reported to be under-expressed in canine lymphoma cell lines and tissues [77]. miRNA-9 enhances mast cell tumor progression [78]. miRNA-145, -203, and -205 are down-regulated in canine melanoma [79,80]. miRNA-123b is significantly over-expressed in B-Cell chronic lymphocytic leukemia (CLL). miRNA-155 is preferentially over-expressed in T lymphocytes and some B-cell CLL, and miRNA-150 is over-expressed in T-Cell CLL in comparison to B-Cell CLL [81]. miRNA-214 promotes apoptosis in hemangiosarcoma, and miRNA dysregulation is also involved in canine splenic hemangiosarcoma [82,83]. miRNA expression profiles differ in canine splenic hemangiosarcoma, nodular hyperplasia, and normal spleens. 22 miRNAs were differentially expressed in canine hemangiosarcoma samples compared to the normal spleen and nodular hyperplasia [82].
In canine mammary tumor (CMT), expression of mi-RNA-141 showed post-transcriptional downregulation of the tumor suppressor gene family INK4A/CDKN2A [84]. miRNA-21 and -29b were reported to be up-regulated in mammary gland tumor tissues, and miR-141 was reported to be over-expressed in canine mammary tumor cell lines, whereas miRNA-31, -34a, and -143/145 were downregulated in canine mammary tissues [84]. Similar miRNAs expression is reported in humans and dogs mammary tumor patients. miR-15a and miR-16 are downregulated in canine ductal carcinomas, while miR-181b, miR-21, miR-29b, and miRlet-7f are upregulated in tubular papillary carcinomas [85]. miR-29b, miR-101, miR-143, and miR-145 expression levels were downregulated and miR-125a expression levels were upregulated in canine mammary tumors compared to normal mammary cells [86].
3. Role of miRNAs in cancer metastasis
3.1. Humans
Metastasis is the spread of cancer cells to a secondary site other than the primary site of tumor formation. Metastasis can either be local, through direct migration, or systemic, via the blood or lymphatics. Most deaths caused by cancer are due to cancer cell metastasis [87]. Like primary cancer, miRNAs has also been noted to play a role in metastasis.
miRNA-10b overexpression was linked with onset of breast cancer and its metastasis to the lung [88,89]. Similarly, overexpression of miRNA-320a in pancreatic cancer strongly contributed to the acquisition of characteristics such as high levels of proliferation, invasion, metastasis, and drug resistance, as well as driving cells towards the epithelial-mesenchymal transition by downregulating PDCD4 [90]. In glioma, miRNA-10b is highly expressed, especially in advanced-grade gliomas [91]. The inhibition of miRNA-10b produces pleiotropic effects (affects growth, invasion, and angiogenesis) in glioma cell lines but not on the growth of normal neurons and astrocytes [91].
Conversely, overexpression of miRNA-320a inhibits the capacity for invasion and migration of breast cancer in vitro, while the silencing of miRNA-320a favors it. This effect is due to the fact that miRNA-320a suppresses the powerful metadherin oncogene. Xenograft experiments (human tumor cell lines grafted in female athymic nude mice) showed that expression of miRNA-320a could inhibit breast cancer metastasis in vivo [92]. Downregulation of miR-335 causes upregulation of TGF-β and its pathway members ROCK1, MAPK1, and LRG1, which leads to significant invasion and migratory potential in neuroblastoma cells [93].
3.2. Dogs
Similar to humans, in dogs, disease progression and metastasis are major indicators of long-term survival in many tumors. Expression analysis of 317 miRNAs from 146 canine mammary tumors reveals significantly different expression profiles between metastatic and non-metastatic groups, thus, signifying the role of miRNAs as a metastasis marker [94]. The downregulation of miRNAs, including miR-29b, miR-101, miR-143, miR-145, and miR-125a, was correlated with progression to metastatic disease compared to non-metastatic canine mammary tumors [94]. miRNA-210 over-expression and miRNA-125a under-expression in canine mammary gland tumors has been shown to drive metastasis [86]. miR-124 regulates canine mammary carcinoma growth and promotes epithelial-mesenchymal transition by targeting CDH2 [95]. The hypoxia-regulated miRNAs profile differs between cell lines of primary and metastatic canine oral melanoma [96]. This profiling showed that metastatic cells are more resistant to hypoxia than primary tumor cells. Downregulation of miRNAs at the 14q32 locus is associated with aggressive osteosarcoma characteristics, including increased metastatic potential and accelerating time to death in both human and canine patients [97].
Reduced expression of miRNA-34a, -134, -544, -382, -1, and -133b are linked with the invasion and migration of tumors and aggressive canine osteosarcoma [97,98,99]. miR-9 promotes a metastatic phenotype of OSA cell lines and is up-regulated in primary OSA tumors [100]. Inhibition of miR-9 reduces migration and invasion. miR-9 regulates gelsolin, which promotes enhanced motility of neoplastic cells [100]. Unlike miR-9, miR-32a reduces or inhibits canine OSA tumor and cell line growth. Metastatic OSA lesions produce 50% less miR-32a than the primary tumors [99]. miRNA-433 is involved in the proliferation, invasion, and migration of dog bladder cancer cells [101].
4. miRNAs as potential cancer biomarkers
4.1. Humans
The development of powerful techniques, such as microRNAome sequencing, microRNA-specific quantitative PCR, and in vivo antisense technologies, is expected to significantly impact clinical oncology in the near future. The characterization of sensitive and specific biomarkers, preferably those that circulate in body fluids, is critical for the timely diagnosis of cancer. Early detection of disease with a minimally invasive screening test could significantly increase the effectiveness of treatment and decrease its cost. The current understanding of the role of miRNAs in the development and progression of cancer has made miRNAs a powerful tool as a cancer biomarker. Several recent studies have revealed that miRNAs are stably detectable in plasma/serum. Due to their release into the circulation and their extraordinary stability, the levels of specific miRNA in plasma and other biological fluids can serve as diagnostic and prognostic biomarkers of diseases (Figure 2) [102].
One challenge in using miRNA as a diagnostic tool is the visualization or detection of the specific miRNA(s) to be tested. A novel DNA nanomachine was developed to selectively visualize miR-21 in cancer cells [103]. The nanomachine was designed with a Zeolite imidazole framework-8 (ZIF-8) metal-organic framework (MOF), assembled with two hairpin probes (Y1 and Y2) labeled with fluorescent dye for signal amplification. After exposure to an acidic extracellular environment, the MOF decomposes and releases the hairpin molecules, which are captured by targeted miR-21. This induces catalytic hairpin assembly (CHA) amplification between the two hairpin molecules and detects the presence of the miRNA even at a low sensitivity of 27pM (64). Thus, targeting microRNA can be an effective approach to developing sensitive diagnostics for cancer cells [104].
When the expression profiles of microRNAs were compared between healthy tissues and tumor samples, unique patterns were identified that could discern between tumor and non-tumor cells [105,106,107,108]. MicroRNA expression profiling appears to be more informative and more powerful in classifying tumor samples by their tissue of origin (something that can be complicated when tumors are diagnosed in advanced stages), tumorigenicity, and degree of differentiation than the profiles of messenger RNA (mRNA) which were traditionally used [106]. For example, a profile of only two hundred microRNAs was shown to be enough to classify poorly differentiated tumors (a frequent clinical problem) with greater precision than using the information of more than sixteen thousand messenger RNAs [106]. Similarly, the effectiveness of miRNAs as biomarkers for tracing cancer's tissue of origin was achieved by measuring 400 samples from 22 tumor tissues and metastases [109].
4.1.1. Diagnosis
Circulating miRNA can also be an effective biomarker for cancer diagnosis. The potential of urinary miRNA biomarkers was studied for the early detection of colorectal cancer. A statistically significant increase in miR-129-1-3p and miR-566 levels in urine samples were found in primary tumor tissues compared to normal tissues [110]. MicroRNAs have been measured in additional tissues and body fluids. Plasma overexpression of miR-21, miR-125b, miR-126, miR-141, let-7, miR-205, and miR-375 were identified in prostate cancer. Similarly, overexpression of miR-20a was found in prostatic tissue, and an increase in miR-21, miR-141, and miR-375 was found in the urine [111,112,113]. The identification of the overexpression of these microRNAs could be useful for the diagnosis of the disease [114,115,116].
Increase in the expression of exosomal miR-23a and miR-1246 allow the detection of disease in its early stages and differentiate between healthy and malignant tissues in colon cancer [117]. In plasma from melanoma patients, miR149-3p, miR150-5p, and miR193a-5p were reported to be higher compared to healthy patients and, therefore, can be used as biomarkers [118]. In saliva from esophageal cancer patients, overexpression of at least four miRNAs was reported (miR-144, miR-10b, miR-451, miR-21) versus healthy patients [119]. This fact has been corroborated in pancreatic cancer where miR-21, miR-23, miR-181a, miR-181b, and miR-196a were overexpressed in patients' saliva compared to controls [120].
4.1.2. Prognosis
The expression profiles of microRNAs can provide important clinical information about the prognosis of patients. MicroRNA profiles correlate with survival in different tumor types, including those in early pathological states. Low levels of expression of the let-7 miRNA family and high levels of miR-155 showed a correlation with a poor prognosis in lung cancer [108]. In a second study on lung cancer, high levels of miR-137, miR372, and miR-182 correlated with poor prognosis, while high levels of miR-221 and let-7a appear to be protective. In addition, the levels of this set of microRNAs were useful in predicting tumor recurrence [121]. A study in colon cancer demonstrated that high levels of miR-21 expression were associated with poor therapeutic response and poor patient prognosis [122].
A study of circulating cell-free microRNA (cf-miRNA) in ovarian cancer patients revealed that high levels of cf-miRNA (miR-92a, -200c, -320b, -320c, -335, -375, -486) are significantly associated with adverse clinical features. This suggests that a panel of cf-miRNA can work as an independent prognostic marker [123].
4.1.3. Tumor staging
It has also been suggested that microRNAs could be relevant in differentiating disease stages, including localized or metastatic cancer. Overexpression of miR-141, miR-200, and miR-375 is linked with the aggressiveness of the prostatic neoplasia [124,125]. In colon cancer, miRNA-122 and some members of the miRNA-200 family are elevated in the plasma of patients with metastatic disease with recurrence [126]. The overexpression of the exosomal miRNAs Let-7g, miR15b, miR-155, and miR-328 can potentially indicate shorter relapse in patients with advanced colon cancer stages [127]. Similarly, overexpression of exosomal miR-328 was linked with the development of liver metastasis in colon cancer patients [127]. In advanced stages, elevated levels of miR-155 indicate resistance to chemotherapy, and elevation of miR-155, miR-200c, and miR-210 indicate local recurrence, metastasis, and poor prognosis [128].
4.1.4. Treatment resistance
The usefulness of microRNAs as indicators of sensitivity or resistance to chemotherapeutic agents has been demonstrated. Overexpression of miR-34a, miR-205, and miR-31 is associated with sensitivity to taxanes. Overexpression of miR-106b has been associated with resistance to radiotherapy, while the overexpression of miR449a confers greater sensitivity to it [129,130,131,132,133,134,135]. MicroRNAs can differentiate between castration-sensitive and castration-resistant prostate cancer. In the latter, miR-125b-2 and miR-708 are decreased and miR-375, miR-141, miR-143, miR-362-5p, miR-214, let-7i and miR-545 are overexpressed [131,135].
The downregulation of miR-452 is linked with acquired resistance to the drug Adriamycin (ADR) or Doxorubicin. Upregulation of miR-452 decreased resistance to ADR in MCF7 cells by abrogating the activity of IGF-1R [136].
It has also been shown that the loss of miRNA-200c correlates with a better prognosis in breast cancer and leads to increased cancer cell sensitivity to Doxorubicin by reducing MDR1 gene expression [137]. Recent studies have suggested that negative regulation of miR-542-3p may contribute to resistance to the breast cancer drug Trastuzumab (Herceptin) since the drug induces the expression of this miRNA in SKBR3 and MCF7/Her2 cells by activating the PI3K-AKT pathway and generating a blockade of the G1/S checkpoint [89]. miRNA-205, at high levels, is predictive of a better response to the TAC NAC regimen (docetaxel, doxorubicin, plus cyclophosphamide). This miRNA improves the chemosensitivity of cancer cells to TAC by suppressing both VEGFA and FGF2. It is downregulated in MCF-7/A02 and CALDOX cells (two drug-resistant derivatives of MCF-7 and Cal51 cells). Thus, the increase in miR-205 can serve as a predictive biomarker and a potential therapeutic target for the treatment [138].
miR-21 is involved in the development of resistance to Gemcitabine through epithelial-to-mesenchymal transition (EMT) and AKT pathway activation. PTEN, a direct target gene of this miRNA, is significantly downregulated in drug-resistant breast cancer cells, and restoration of PTEN expression blocks miR-21-induced EMT and gemcitabine resistance [139].
Aggressive triple-negative breast cancers (TNBC) rapidly develop resistance to chemotherapies. Genotoxic treatments, such as Doxorubicin (Dox), were found to significantly increase the expression of miR-181a in TNBC cells, which increases metastasis. Regulation of this miRNA is orchestrated by the transcription factor STAT3. BAX was also identified as a direct functional target of miR-181a. BAX suppression decreases apoptosis and promotes cell invasion, which is critical in promoting therapeutic resistance and aggressive behavior of TNBC cells after genotoxic treatment. An antagonist of this miRNA may serve as a promising strategy to sensitize TNBC cells to chemotherapy and mitigate metastasis [140].
miR-182, found in triple-negative breast cancer tissues, induces proliferation and invasion of MDA-MB-231 cells through negative regulation of the PFN1 protein. Treatment strategies using miR-182 expression inhibition or PFN1 gene overexpression could also benefit patients with TNBC [141]. High levels of miR-621 in breast cancer predict a better response to chemotherapy since the ectopic overexpression of this miRNA promotes apoptosis and chemosensitivity to drugs by suppressing FBXO11, which causes an Increase in p53 activity, promoting apoptosis in cells exposed to chemotherapeutic agents [142].
The role of miRNA-340 was examined using a lentiviral vector overexpressing miRNA-340 in the triple-negative breast cancer cell line MDA-MB-231. Overexpression of miR-340 decreased the expression of the transcription factor SOX2 and cyclin-dependent P16 and P27 kinase inhibitors. [143]. miR-340 inhibition of these breast cancer associated genes has potential therapeutic use.
An analysis of almost 600 miRNAs in tumors from patients with progressive bladder cancer was performed. The patients were treated with cisplatin and it was found that 15 miRNAs play an important role in the response, 5 in the survival interval, and 3 in both aspects [144].
4.2. Dogs
Circulatory exosomal miRNAs are being studied for their utility as markers for early detection in cancer screening tests using a minimally invasive liquid biopsy approach in dogs [103]. miRNA expression differs between tumor type and stage, which allows for more accurate diagnosis in wide variety of canine tumors [86]. A recent study using Next Generation Sequencing and PCR amplification looking at 13 canine cancer cell lines identified six miRNAs elevated in all cancer types. Of these six miRNAs, cfa-miR-9 was the most overexpressed in all cell lines compared to normal cells [103]. Another study showed that miR-216 and miR-126 showed different circulatory levels depending on tumor type. miR-216 showed elevated levels in the blood of sarcoma patients, while miR-126 showed elevation in epithelial-derived tumor patients [145]. The identification of tumor-specific miRNAs could be an important diagnostic tool in splenic masses where ~75% are malignant, with concurrent poor prognosis (median survival is 110 days) versus the remaining 25% where median survival is over one year [146].
A number of miRNAs have been Identified as specific biomarkers in tumor tissues in several tumor types in dogs. For example, miR-103b, -34a, -106b, and miR-16 can potentially be used as diagnostic biomarkers for transitional cell carcinoma of the bladder in dogs [147,148]. Circulatory miR-214 and -126 are reported as strong predictors of disease-free intervals and overall survival in appendicular OSA patients receiving amputation and chemotherapy [145]. Circulatory miRNA-214 and -126 are also reported as potential biomarkers for diagnosis and prognosis for osteosarcoma and other canine neoplastic diseases [145].
Cutaneous mast cell tumor is a common neoplasms in dogs. Dysregulation of 63 miRNAs (18 up- and 45- downregulated) was reported in mast cell tumor tissues compared to the adjacent healthy tissues. miR-21, miR-379, and miR-885 are promising biomarkers to identify patients with mast cell tumors and lymph node metastasis [149]. Canine mammary carcinoma patients have differentially expressed miRNAs. Circulatory miR-19b and miR-18a can potentially be used as biomarkers for diagnosis and prognosis in canine mammary cancer patients [150]. Similarly, miR-21, miR-155, miR-9, miR-34a, miR-143/145, and miR-31 have altered (up and down regulated) expression in canine mammary tumor cells and may serve as potential targets for breast cancer therapeutics [84].
The development of accurate blood tests based on miRNA expression could provide veterinarians with more information for more effective treatment. Minimally invasive liquid biopsy in canine multicentric lymphoma is proposed to evaluate the potential outcome of chemotherapy. Circulatory miR-205, miR-222, miR-20a, and miR-93 can be used as molecular signatures to predict non-responsive cases [151]. Similarly, circulatory miRNAs can also be used as a biomarker to depict cardiotoxicity in dogs due to the prolonged use of the chemotherapeutic drug doxorubicin. miR-502 downregulation is a potential biomarker to predict cardiotoxicity before other echocardiographic parameters [152]. Exosomal miR-143 and miR-221 are significantly increased in metastatic melanoma patients and can be used as biomarkers to identify metastatic melanoma [153]. miR-15b and miR-342-3p isolated from plasma can be used as non-invasive biomarkers to differentiate glioma from other intracranial diseases in dogs [154]. miR-214 and miR-126 are over-secreted in malignant endothelial proliferated diseases [83]. Therefore, these miRNAs have the potential as a diagnostic biomarker for malignant endothelial proliferated diseases in dogs.
Uveal melanoma is a primary intraocular tumor in dogs with no effective means of predicting metastasis. Fourteen miRNAs have been identified that exhibit significant differences between metastatic and non-metastasizing tumors. miRNA expression profile of these 14 miRNAs may be used to predict metastasis and for therapeutic purposes [155].
Research on canine tumors has implications in human medicine; however, canine miRNAs are much less well-studied than human miRNAs. There is a significant crossover in miRNAs identified in the two species [156]. In mammary tumors, a study showed nine of ten overexpressed miRNAs in canine tumors are the same as in human mammary tumors [85]. Circulating miR-214 and miR-126 showed similar patterns in human and canine patients based on tumor type and disease progression [145]. Down-regulation of miR-1, miR-34, and upregulation of miR-9 and miR-106b cluster have been correlated with increased proliferation and invasion of osteosarcoma in humans and dogs [156]. The crossover between canine and human miRNA expression patterns in tumors poses a unique research opportunity. The similarities allow for easy interspecies application and the advancement of diagnostic technology and treatments in both species simultaneously.
5. Role of miRNAs in cancer treatment
5.1. Humans
As stated above, each cancer possesses a specific combination of miRNAs, either overexpressed oncomiRNAs targeting tumor suppressor genes or downregulated tumor suppressor miRNAs targeting oncogenes [157]. This profile of expressed miRNAs may be used to establish a "fingerprint" that could potentially identify specific tumor types and even subtypes with a given tumor. Since miRNAs are involved in cancer cell gene regulation, these may provide excellent opportunities to design personalized therapeutics for cancer patients. miRNA-based anti-cancer therapies have recently generated interest either as monotherapies or in combination with other cancer therapies. Targeting oncomiRNAs induces expression of tumor suppressor genes, which in turn enhance tumor cell killing and promote tumor regression [158]. However, physiological and cellular barriers hamper the in vivo efficacy of anti-miRNA technologies.
As previously mentioned, one of the first miRNAs detected in the human genome, miR-21, is overexpressed in glioblastoma [159] and could be used as a therapeutic target in this type of cancer. In glioblastoma cells, the additive interaction of antisense oligonucleotide inhibitors to both miRNA-21 and miRNA-10b may constitute an effective therapeutic strategy to control glioblastoma growth by inhibiting oncogene expression and inducing tumor suppressor gene expression. miRNA-21 inhibitors also interrupt the activity of the EGFR pathway, thereby increasing the expression of PDCD4 and TPM1 and reducing the activities of matrix metalloproteinases (MMPs) [159]. Inhibition of NADPH oxidase (NOX) dramatically lowered the invasive potential of lung cancer in vitro by decreasing miRNA-21-expression, [160].
When miRNA inhibitors are co-administered with an anti-cancer agent, they can induce synergistic effects (e.g., in glioblastoma) [161]. A concern in miRNA modulation strategies is the proper identification, in silico, of miRNA inhibitors or analogs that can effectively inhibit or mimic the function of specific miRNAs to achieve miRNA loss or gain of function, respectively. Another challenge to miRNA-directed therapies' efficiency is the long-term release of these miRNA inhibitors or mimics at their specific target sites. A new form of miRNA inhibitor delivery has been developed to answer these concerns, specifically targeting miRNA-155 in an acidic tumor microenvironment in murine lymphoma. To achieve this, peptide nucleic acid anti-miRs were attached to a peptide with a low pH-induced transmembrane structure (pHLIP). This construct could target the tumor microenvironment and transport anti-miRs under acidic conditions across the plasma membrane. This approach evades the hepatic barrier (removal of foreign proteins from circulation by the hepatic reticulo-endothelial system) and facilitates miRNA targeting through a non-specific endocytic pathway [162]. An alternative miRNA inhibitor delivery strategy using R3V6 peptide was evaluated as a transporter of antisense oligodeoxynucleotides [163]. Serum stability assays showed that R3V6 protected miRNA inhibitors from nucleases more efficiently than polyethyleneimine (PEI; 25kDa, PEI25k). In an in vitro transfection assay, R3V6 transported antisense oligodeoxynucleotide anti-miRNA-21 into cells more efficiently than PEI (25kDa, PEI25k) and lipofectamine [163].
microRNA can also serve as a candidate for developing oncolytic virotherapy. A new miRNA-modified Coxsackievirus B3 (CVB3) was developed by inserting miR-145/143, miR-1, and miR-216 target sequences into the 5' untranslated region (5' UTR) of the CVB3 genome. miR-145/143 is downregulated in tumors, miR-1is muscle-specific, and miR-216 is pancreas selective [164]. The virus is downregulated in any cell expressing any of the 3 miRNAs but is replication-competent in cells, such as a tumor, that do not express any of the three. This novel miRNA-modified oncolytic virus inhibited triple-negative breast cancer growth in immunocompromised mouse models [164].
Chemotherapy and miRNA therapy combinations have shown synergistically increased antineoplastic activities. The combination of a miRNA-21 inhibitor and taxol is an effective therapeutic strategy to control the growth of glioblastoma multiforme (GBM) by inhibiting the expression and phosphorylation of STAT3 in vitro [165].
5.2. Dogs
Canine hemangiosarcoma has an extremely poor prognosis. Upregulation of miR-214 induces apoptosis in hemangiosarcoma cell lines. Intraperitoneal administration of synthetic miR-214 (miR-214/5AE) exhibits anti-tumor effects in a murine model of canine hemangiosarcoma. It induces apoptosis and prohibits cell proliferation [166]. Similarly, intratumoral administration of synthetic miRNA-205 (miR-205BP/S3) can be used to treat canine malignant melanoma. Administration of miR-205BP/S3 in eleven dogs lead to five complete remissions, three dogs with stable diseases, and three cases of progressive disease [167].
Novel miRNA vectors are being explored to induce oncolysis and disease remission in solid tumors leading to a new wave of cancer treatments [168]. Canine osteosarcoma patients with metastatic disease are have a poor prognosis. miR-34a suppresses the oncogene Eag-1, and the downregulation of miR-34a has been correlated with the progression of canine osteosarcoma. In vitro and in vivo models showed that administration of miR-34a inhibited osteosarcoma progression and decreased Eag-1 production [101]. A bioengineered miRNA prodrug (tRNA/miR-34a) was successfully processed into mature miR-34a in canine osteosarcoma cells. The administration of tRNA/miR-34a in murine models with canine OSA xenografts caused delayed tumor growth, increased necrosis and apoptosis, and reduced cellular proliferation [168]. The tRNA/miR-34a treatment showed 32% less tumor growth and more prolonged survival versus the control groups [168]. The emergence of new research on the effects of miRNA on tumor progression allows for novel treatment development.
6. Future Perspective and Summary
miRNAs are clearly involved in cancer progression and metastasis in both humans and dogs. As stated above, a wide range of miRNAs can be utilized as cancer biomarkers and for cancer therapeutics. A cancer diagnosis is often delayed to the point that cancer has progressed significantly and metastasis has occurred. miRNAs show promise as a biomarker for earlier cancer detection and, therefore, more effective treatment. miRNAs can be secreted into the bloodstream, are stable, as a circulating free miRNAs or are protected from endogenous RNAse enzyme activity by encapsulation in exosomes or proteins. These biomarkers can serve to screen patients for potential undiagnosed tumors. A positive result would trigger a search for the tumor and potentially therapy to treat the primary tumor and reduce the chance of metastases. miRNAs may potentially serve as markers for staging tumors, including the presence or absence of metastases, based on the stage of disease at the time of sample collection. All this is achievable using minimally invasive, “liquid biopsy” approaches to monitor tumor progression, response to therapy, and relapse. However, to realize the full potential of miRNAs in these areas, more research is required regarding the use of miRNAs as therapeutic biomarkers.
Author Contributions
Conceptualization, P.A.; Resources, A.C. and D.J.P.; Writing—Original Draft Preparation, A.C. and D.J.P; Writing—Review and Editing, B.F.S and P.A.; Visualization, P.A.; Supervision, P.A.; Project Administration, P.A.; All authors have read and agreed to the published version of the manuscript.
Acknowledgments
The authors would like to acknowledge the Scott-Ritchey Research Center at the College of Veterinary Medicine, Auburn University, Alabama, for financial support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- www.miRBase.org. Available online: https://www.mirbase.org/cgi-bin/browse.pl.
- Fabian, M.R.; Sonenberg, N.; Filipowicz, W. Regulation of mRNA translation and stability by microRNAs. Annual review of biochemistry 2010, 79, 351–379. [Google Scholar] [CrossRef] [PubMed]
- Broughton, J.P.; Lovci, M.T.; Huang, J.L.; Yeo, G.W.; Pasquinelli, A.E. Pairing beyond the Seed Supports MicroRNA Targeting Specificity. Mol Cell 2016, 64, 320–333. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenbach, H. The clinical relevance of circulating, exosomal miRNAs as biomarkers for cancer. Expert review of molecular diagnostics 2015, 15, 1159–1169. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nature cell biology 2007, 9, 654–659. [Google Scholar] [CrossRef]
- Ahmad, N.; Haider, S.; Jagannathan, S.; Anaissie, E.; Driscoll, J.J. MicroRNA theragnostics for the clinical management of multiple myeloma. Leukemia 2014, 28, 732–738. [Google Scholar] [CrossRef]
- Svoronos, A.A.; Engelman, D.M.; Slack, F.J. OncomiR or Tumor Suppressor? The Duplicity of MicroRNAs in Cancer. Cancer Res 2016, 76, 3666–3670. [Google Scholar] [CrossRef]
- Mogilyansky, E.; Rigoutsos, I. The miR-17/92 cluster: a comprehensive update on its genomics, genetics, functions and increasingly important and numerous roles in health and disease. Cell Death Differ 2013, 20, 1603–1614. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Han, C.; Wu, T. MiR-17-92 cluster promotes hepatocarcinogenesis. Carcinogenesis 2015, 36, 1213–1222. [Google Scholar] [CrossRef]
- Tsuchida, A.; Ohno, S.; Wu, W.; Borjigin, N.; Fujita, K.; Aoki, T.; Ueda, S.; Takanashi, M.; Kuroda, M. miR-92 is a key oncogenic component of the miR-17-92 cluster in colon cancer. Cancer Sci 2011, 102, 2264–2271. [Google Scholar] [CrossRef]
- Takakura, S.; Mitsutake, N.; Nakashima, M.; Namba, H.; Saenko, V.A.; Rogounovitch, T.I.; Nakazawa, Y.; Hayashi, T.; Ohtsuru, A.; Yamashita, S. Oncogenic role of miR-17-92 cluster in anaplastic thyroid cancer cells. Cancer Sci 2008, 99, 1147–1154. [Google Scholar] [CrossRef]
- Hayashita, Y.; Osada, H.; Tatematsu, Y.; Yamada, H.; Yanagisawa, K.; Tomida, S.; Yatabe, Y.; Kawahara, K.; Sekido, Y.; Takahashi, T. A polycistronic microRNA cluster, miR-17-92, is overexpressed in human lung cancers and enhances cell proliferation. Cancer Res 2005, 65, 9628–9632. [Google Scholar] [CrossRef] [PubMed]
- Martens-de Kemp, S.R.; Komor, M.A.; Hegi, R.; Bolijn, A.S.; Tijssen, M.; de Groen, F.L.M.; Depla, A.; van Leerdam, M.; Meijer, G.A.; Fijneman, R.J.A.; et al. Overexpression of the miR-17-92 cluster in colorectal adenoma organoids causes a carcinoma-like gene expression signature. Neoplasia 2022, 32, 100820. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Zhao, X.; Zhang, Z.; Xiong, M.; Wang, Y.; Sun, Y.; He, B.; Zhu, J. The diagnostic and prognostic value of the miR-17-92 cluster in hepatocellular carcinoma: A meta-analysis. Front Genet 2022, 13, 927079. [Google Scholar] [CrossRef] [PubMed]
- Menon, A.; Abd-Aziz, N.; Khalid, K.; Poh, C.L.; Naidu, R. miRNA: A Promising Therapeutic Target in Cancer. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Ozgun, A.; Karagoz, B.; Bilgi, O.; Tuncel, T.; Baloglu, H.; Kandemir, E.G. MicroRNA-21 as an indicator of aggressive phenotype in breast cancer. Onkologie 2013, 36, 115–118. [Google Scholar] [CrossRef]
- Echevarria-Vargas, I.M.; Valiyeva, F.; Vivas-Mejia, P.E. Upregulation of miR-21 in cisplatin resistant ovarian cancer via JNK-1/c-Jun pathway. PLoS One 2014, 9, e97094. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, C.; Shan, S.; Liu, X.; Jiang, Z.; Ren, T. TLR4/ROS/miRNA-21 pathway underlies lipopolysaccharide instructed primary tumor outgrowth in lung cancer patients. Oncotarget 2016, 7, 42172–42182. [Google Scholar] [CrossRef]
- Si, H.; Sun, X.; Chen, Y.; Cao, Y.; Chen, S.; Wang, H.; Hu, C. Circulating microRNA-92a and microRNA-21 as novel minimally invasive biomarkers for primary breast cancer. J Cancer Res Clin Oncol 2013, 139, 223–229. [Google Scholar] [CrossRef]
- Baez-Vega, P.M.; Echevarria Vargas, I.M.; Valiyeva, F.; Encarnacion-Rosado, J.; Roman, A.; Flores, J.; Marcos-Martinez, M.J.; Vivas-Mejia, P.E. Targeting miR-21-3p inhibits proliferation and invasion of ovarian cancer cells. Oncotarget 2016, 7, 36321–36337. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, Z.; Gao, C.; Chen, P.; Chen, J.; Liu, W.; Xiao, S.; Lu, H. miR-21 plays a pivotal role in gastric cancer pathogenesis and progression. Lab Invest 2008, 88, 1358–1366. [Google Scholar] [CrossRef]
- Yaman Agaoglu, F.; Kovancilar, M.; Dizdar, Y.; Darendeliler, E.; Holdenrieder, S.; Dalay, N.; Gezer, U. Investigation of miR-21, miR-141, and miR-221 in blood circulation of patients with prostate cancer. Tumour Biol 2011, 32, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Toiyama, Y.; Takahashi, M.; Hur, K.; Nagasaka, T.; Tanaka, K.; Inoue, Y.; Kusunoki, M.; Boland, C.R.; Goel, A. Serum miR-21 as a diagnostic and prognostic biomarker in colorectal cancer. J Natl Cancer Inst 2013, 105, 849–859. [Google Scholar] [CrossRef]
- Simonian, M.; Mosallayi, M.; Mirzaei, H. Circulating miR-21 as novel biomarker in gastric cancer: Diagnostic and prognostic biomarker. J Cancer Res Ther 2018, 14, 475. [Google Scholar] [CrossRef] [PubMed]
- Sicard, F.; Gayral, M.; Lulka, H.; Buscail, L.; Cordelier, P. Targeting miR-21 for the therapy of pancreatic cancer. Mol Ther 2013, 21, 986–994. [Google Scholar] [CrossRef]
- Si, M.L.; Zhu, S.; Wu, H.; Lu, Z.; Wu, F.; Mo, Y.Y. miR-21-mediated tumor growth. Oncogene 2007, 26, 2799–2803. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.K.; Blansit, K.; Kiet, T.; Sherman, A.; Wong, G.; Earle, C.; Bourguignon, L.Y. The inhibition of miR-21 promotes apoptosis and chemosensitivity in ovarian cancer. Gynecol Oncol 2014, 132, 739–744. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Pang, X.; Luan, F. Diagnosis value of miR-181, miR-652, and CA72-4 for gastric cancer. J Clin Lab Anal 2022, 36, e24411. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Shen, R.; Yan, Y.; Deng, L. miR-186 promotes tumor growth in cutaneous squamous cell carcinoma by inhibiting apoptotic protease activating factor-1. Experimental and therapeutic medicine 2018, 16, 4010–4018. [Google Scholar] [CrossRef]
- Zhai, Z.; Mu, T.; Zhao, L.; Li, Y.; Zhu, D.; Pan, Y. MiR-181a-5p facilitates proliferation, invasion, and glycolysis of breast cancer through NDRG2-mediated activation of PTEN/AKT pathway. Bioengineered 2022, 13, 83–95. [Google Scholar] [CrossRef]
- Strotbek, M.; Schmid, S.; Sanchez-Gonzalez, I.; Boerries, M.; Busch, H.; Olayioye, M.A. miR-181 elevates Akt signaling by co-targeting PHLPP2 and INPP4B phosphatases in luminal breast cancer. Int J Cancer 2017, 140, 2310–2320. [Google Scholar] [CrossRef]
- Tong, S.J.; Liu, J.; Wang, X.; Qu, L.X. microRNA-181 promotes prostate cancer cell proliferation by regulating DAX-1 expression. Exp Ther Med 2014, 8, 1296–1300. [Google Scholar] [CrossRef] [PubMed]
- Barbato, A.; Iuliano, A.; Volpe, M.; D'Alterio, R.; Brillante, S.; Massa, F.; De Cegli, R.; Carrella, S.; Salati, M.; Russo, A.; et al. Integrated Genomics Identifies miR-181/TFAM Pathway as a Critical Driver of Drug Resistance in Melanoma. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Abrahamsson, A.; Dabrosin, C. Tissue specific expression of extracellular microRNA in human breast cancers and normal human breast tissue in vivo. Oncotarget 2015, 6, 22959–22969. [Google Scholar] [CrossRef] [PubMed]
- Tao, K.; Yang, J.; Guo, Z.; Hu, Y.; Sheng, H.; Gao, H.; Yu, H. Prognostic value of miR-221-3p, miR-342-3p and miR-491-5p expression in colon cancer. Am J Transl Res 2014, 6, 391–401. [Google Scholar] [PubMed]
- Pineau, P.; Volinia, S.; McJunkin, K.; Marchio, A.; Battiston, C.; Terris, B.; Mazzaferro, V.; Lowe, S.W.; Croce, C.M.; Dejean, A. miR-221 overexpression contributes to liver tumorigenesis. Proc Natl Acad Sci U S A 2010, 107, 264–269. [Google Scholar] [CrossRef]
- Chen, W.X.; Hu, Q.; Qiu, M.T.; Zhong, S.L.; Xu, J.J.; Tang, J.H.; Zhao, J.H. miR-221/222: promising biomarkers for breast cancer. Tumour Biol 2013, 34, 1361–1370. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Sevignani, C.; Dumitru, C.D.; Hyslop, T.; Noch, E.; Yendamuri, S.; Shimizu, M.; Rattan, S.; Bullrich, F.; Negrini, M.; et al. Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc Natl Acad Sci U S A 2004, 101, 2999–3004. [Google Scholar] [CrossRef]
- Macfarlane, L.A.; Murphy, P.R. MicroRNA: Biogenesis, Function and Role in Cancer. Curr Genomics 2010, 11, 537–561. [Google Scholar] [CrossRef]
- Yang, W.; Lee, D.Y.; Ben-David, Y. The roles of microRNAs in tumorigenesis and angiogenesis. Int J Physiol Pathophysiol Pharmacol 2011, 3, 140–155. [Google Scholar]
- Qu, H.; Zheng, L.; Song, H.; Jiao, W.; Li, D.; Fang, E.; Wang, X.; Mei, H.; Pu, J.; Huang, K.; et al. microRNA-558 facilitates the expression of hypoxia-inducible factor 2 alpha through binding to 5'-untranslated region in neuroblastoma. Oncotarget 2016, 7, 40657–40673. [Google Scholar] [CrossRef]
- Baradaran, B.; Shahbazi, R.; Khordadmehr, M. Dysregulation of key microRNAs in pancreatic cancer development. Biomed Pharmacother 2019, 109, 1008–1015. [Google Scholar] [CrossRef]
- Wen, J.; Fu, J.; Zhang, W.; Guo, M. Genetic and epigenetic changes in lung carcinoma and their clinical implications. Mod Pathol 2011, 24, 932–943. [Google Scholar] [CrossRef]
- Zhao, L.; Duan, Y.T.; Lu, P.; Zhang, Z.J.; Zheng, X.K.; Wang, J.L.; Feng, W.S. Epigenetic Targets and their Inhibitors in Cancer Therapy. Curr Top Med Chem 2018, 18, 2395–2419. [Google Scholar] [CrossRef]
- Link, S.; Grund, S.E.; Diederichs, S. Alternative splicing affects the subcellular localization of Drosha. Nucleic Acids Research 2016, 44, 5330–5343. [Google Scholar] [CrossRef]
- Xia, L.; Zhang, D.; Du, R.; Pan, Y.; Zhao, L.; Sun, S.; Hong, L.; Liu, J.; Fan, D. miR-15b and miR-16 modulate multidrug resistance by targeting BCL2 in human gastric cancer cells. Int J Cancer 2008, 123, 372–379. [Google Scholar] [CrossRef]
- Shen, J.; Wan, R.; Hu, G.; Yang, L.; Xiong, J.; Wang, F.; Shen, J.; He, S.; Guo, X.; Ni, J.; et al. miR-15b and miR-16 induce the apoptosis of rat activated pancreatic stellate cells by targeting Bcl-2 in vitro. Pancreatology 2012, 12, 91–99. [Google Scholar] [CrossRef]
- Musumeci, M.; Coppola, V.; Addario, A.; Patrizii, M.; Maugeri-Sacca, M.; Memeo, L.; Colarossi, C.; Francescangeli, F.; Biffoni, M.; Collura, D.; et al. Control of tumor and microenvironment cross-talk by miR-15a and miR-16 in prostate cancer. Oncogene 2011, 30, 4231–4242. [Google Scholar] [CrossRef]
- Cimmino, A.; Calin, G.A.; Fabbri, M.; Iorio, M.V.; Ferracin, M.; Shimizu, M.; Wojcik, S.E.; Aqeilan, R.I.; Zupo, S.; Dono, M.; et al. miR-15 and miR-16 induce apoptosis by targeting BCL2. Proc Natl Acad Sci U S A 2005, 102, 13944–13949. [Google Scholar] [CrossRef]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent deletions and down-regulation of micro- RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc Natl Acad Sci U S A 2002, 99, 15524–15529. [Google Scholar] [CrossRef]
- Johnson, S.M.; Grosshans, H.; Shingara, J.; Byrom, M.; Jarvis, R.; Cheng, A.; Labourier, E.; Reinert, K.L.; Brown, D.; Slack, F.J. RAS is regulated by the let-7 microRNA family. Cell 2005, 120, 635–647. [Google Scholar] [CrossRef]
- Kumar, M.S.; Erkeland, S.J.; Pester, R.E.; Chen, C.Y.; Ebert, M.S.; Sharp, P.A.; Jacks, T. Suppression of non-small cell lung tumor development by the let-7 microRNA family. Proc Natl Acad Sci U S A 2008, 105, 3903–3908. [Google Scholar] [CrossRef]
- Buechner, J.; Tomte, E.; Haug, B.H.; Henriksen, J.R.; Lokke, C.; Flaegstad, T.; Einvik, C. Tumour-suppressor microRNAs let-7 and mir-101 target the proto-oncogene MYCN and inhibit cell proliferation in MYCN-amplified neuroblastoma. Br J Cancer 2011, 105, 296–303. [Google Scholar] [CrossRef]
- Sun, X.; Qin, S.; Fan, C.; Xu, C.; Du, N.; Ren, H. Let-7: a regulator of the ERalpha signaling pathway in human breast tumors and breast cancer stem cells. Oncol Rep 2013, 29, 2079–2087. [Google Scholar] [CrossRef]
- Slusarz, A.; Pulakat, L. The two faces of miR-29. J Cardiovasc Med (Hagerstown) 2015, 16, 480–490. [Google Scholar] [CrossRef]
- Zhang, W.; Wu, Q.; Liu, Y.; Wang, X.; Ma, C.; Zhu, W. LncRNA HOTAIR Promotes Chemoresistance by Facilitating Epithelial to Mesenchymal Transition through miR-29b/PTEN/PI3K Signaling in Cervical Cancer. Cells Tissues Organs 2022, 211, 16–29. [Google Scholar] [CrossRef]
- Vera, O.; Bok, I.; Jasani, N.; Nakamura, K.; Xu, X.; Mecozzi, N.; Angarita, A.; Wang, K.; Tsai, K.Y.; Karreth, F.A. A MAPK/miR-29 Axis Suppresses Melanoma by Targeting MAFG and MYBL2. Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef]
- Hozaka, Y.; Seki, N.; Tanaka, T.; Asai, S.; Moriya, S.; Idichi, T.; Wada, M.; Tanoue, K.; Kawasaki, Y.; Mataki, Y.; et al. Molecular Pathogenesis and Regulation of the miR-29-3p-Family: Involvement of ITGA6 and ITGB1 in Intra-Hepatic Cholangiocarcinoma. Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef]
- Grassilli, S.; Bertagnolo, V.; Brugnoli, F. Mir-29b in Breast Cancer: A Promising Target for Therapeutic Approaches. Diagnostics (Basel) 2022, 12. [Google Scholar] [CrossRef]
- Zhang, W.; Qian, J.X.; Yi, H.L.; Yang, Z.D.; Wang, C.F.; Chen, J.Y.; Wei, X.Z.; Fu, Q.; Ma, H. The microRNA-29 plays a central role in osteosarcoma pathogenesis and progression. Mol Biol (Mosk) 2012, 46, 622–627. [Google Scholar] [CrossRef]
- Yu, P.N.; Yan, M.D.; Lai, H.C.; Huang, R.L.; Chou, Y.C.; Lin, W.C.; Yeh, L.T.; Lin, Y.W. Downregulation of miR-29 contributes to cisplatin resistance of ovarian cancer cells. Int J Cancer 2014, 134, 542–551. [Google Scholar] [CrossRef]
- Li, M.H.; Wu, Z.Y.; Wang, Y.; Chen, F.Z.; Liu, Y. Expression of miR-29 and STAT3 in osteosarcoma and its effect on proliferation regulation of osteosarcoma cells. Eur Rev Med Pharmacol Sci 2019, 23, 7275–7282. [Google Scholar] [CrossRef]
- Ji, Q.; Hao, X.; Zhang, M.; Tang, W.; Yang, M.; Li, L.; Xiang, D.; Desano, J.T.; Bommer, G.T.; Fan, D.; et al. MicroRNA miR-34 inhibits human pancreatic cancer tumor-initiating cells. PLoS One 2009, 4, e6816. [Google Scholar] [CrossRef]
- Corney, D.C.; Hwang, C.I.; Matoso, A.; Vogt, M.; Flesken-Nikitin, A.; Godwin, A.K.; Kamat, A.A.; Sood, A.K.; Ellenson, L.H.; Hermeking, H.; et al. Frequent downregulation of miR-34 family in human ovarian cancers. Clin Cancer Res 2010, 16, 1119–1128. [Google Scholar] [CrossRef]
- Wu, X.; Cheng, Y.L.; Matthen, M.; Yoon, A.; Schwartz, G.K.; Bala, S.; Taylor, A.M.; Momen-Heravi, F. Down-regulation of the tumor suppressor miR-34a contributes to head and neck cancer by up-regulating the MET oncogene and modulating tumor immune evasion. J Exp Clin Cancer Res 2021, 40, 70. [Google Scholar] [CrossRef]
- Sun, D.; Wu, Y.; Zhang, S.; Han, Y.; Shen, J.; Zheng, W.; Wei, L.; Liu, Y.; Ren, L.; Gu, Z.; et al. Distinct roles of miR-34 family members on suppression of lung squamous cell carcinoma. Biomed Pharmacother 2021, 142, 111967. [Google Scholar] [CrossRef]
- Shi, L.; Fan, B.; Chen, D.; Guo, C.; Xiang, H.; Nie, Y.; Zhong, D.; Shi, X. Human cytomegalovirus protein UL136 activates the IL-6/STAT3 signal through MiR-138 and MiR-34c in gastric cancer cells. Int J Clin Oncol 2020, 25, 1936–1944. [Google Scholar] [CrossRef]
- Dehghan, R.; Najafi, R.; Azizi Jalilian, F.; Saidijam, M.; Radaei, Z.; Zamani, A.; Ezati, R.; Asna-Ashari, F.; Amini, R. A promising effect of zerumbone with improved anti-tumor-promoting inflammation activity of miR-34a in colorectal cancer cell lines. Mol Biol Rep 2021, 48, 203–218. [Google Scholar] [CrossRef]
- Christensen, L.L.; Tobiasen, H.; Holm, A.; Schepeler, T.; Ostenfeld, M.S.; Thorsen, K.; Rasmussen, M.H.; Birkenkamp-Demtroeder, K.; Sieber, O.M.; Gibbs, P.; et al. MiRNA-362-3p induces cell cycle arrest through targeting of E2F1, USF2 and PTPN1 and is associated with recurrence of colorectal cancer. Int J Cancer 2013, 133, 67–78. [Google Scholar] [CrossRef]
- Wahdan-Alaswad, R.S.; Cochrane, D.R.; Spoelstra, N.S.; Howe, E.N.; Edgerton, S.M.; Anderson, S.M.; Thor, A.D.; Richer, J.K. Metformin-Induced Killing of Triple-Negative Breast Cancer Cells Is Mediated by Reduction in Fatty Acid Synthase via miRNA-193b. Hormones and Cancer 2014, 5, 374–389. [Google Scholar] [CrossRef]
- Li, J.; Kong, F.; Wu, K.; Song, K.; He, J.; Sun, W. miR-193b directly targets STMN1 and uPA genes and suppresses tumor growth and metastasis in pancreatic cancer. Mol Med Rep 2014, 10, 2613–2620. [Google Scholar] [CrossRef]
- Hashemi, M.; Moradi, N.; Ziaee, S.A.; Narouie, B.; Soltani, M.H.; Rezaei, M.; Shahkar, G.; Taheri, M. Association between single nucleotide polymorphism in miR-499, miR-196a2, miR-146a and miR-149 and prostate cancer risk in a sample of Iranian population. J Adv Res 2016, 7, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.G.; Zhou, X.M.; Cui, Z.G.; Hou, G. Effects of common polymorphisms in miR-146a and miR-196a2 on lung cancer susceptibility: a meta-analysis. J Thorac Dis 2016, 8, 1297–1305. [Google Scholar] [CrossRef]
- Li, D.; Zhu, G.; Di, H.; Li, H.; Liu, X.; Zhao, M.; Zhang, Z.; Yang, Y. Associations between genetic variants located in mature microRNAs and risk of lung cancer. Oncotarget 2016, 7, 41715–41724. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Wang, N.; Duan, G. Long non-coding RNA XIST regulates ovarian cancer progression via modulating miR-335/BCL2L2 axis. World J Surg Oncol 2021, 19, 165. [Google Scholar] [CrossRef] [PubMed]
- Eckstein, S. Dogs and Cancer: Get the Facts. Availabe online: https://pets.webmd.
- Uhl, E.; Krimer, P.; Schliekelman, P.; Tompkins, S.M.; Suter, S. Identification of altered MicroRNA expression in canine lymphoid cell lines and cases of B- and T-Cell lymphomas. Genes, chromosomes & cancer 2011, 50, 950–967. [Google Scholar] [CrossRef]
- Fenger, J.M.; Bear, M.D.; Volinia, S.; Lin, T.Y.; Harrington, B.K.; London, C.A.; Kisseberth, W.C. Overexpression of miR-9 in mast cells is associated with invasive behavior and spontaneous metastasis. BMC cancer 2014, 14, 84. [Google Scholar] [CrossRef]
- Noguchi, S.; Mori, T.; Hoshino, Y.; Yamada, N.; Maruo, K.; Akao, Y. MicroRNAs as tumour suppressors in canine and human melanoma cells and as a prognostic factor in canine melanomas. Veterinary and comparative oncology 2013, 11, 113–123. [Google Scholar] [CrossRef]
- Noguchi, S.; Mori, T.; Hoshino, Y.; Yamada, N.; Nakagawa, T.; Sasaki, N.; Akao, Y.; Maruo, K. Comparative study of anti-oncogenic microRNA-145 in canine and human malignant melanoma. The Journal of veterinary medical science 2012, 74, 1–8. [Google Scholar] [CrossRef]
- Gioia, G.; Mortarino, M.; Gelain, M.E.; Albonico, F.; Ciusani, E.; Forno, I.; Marconato, L.; Martini, V.; Comazzi, S. Immunophenotype-related microRNA expression in canine chronic lymphocytic leukemia. Veterinary immunology and immunopathology 2011, 142, 228–235. [Google Scholar] [CrossRef]
- Grimes, J.A.; Prasad, N.; Levy, S.; Cattley, R.; Lindley, S.; Boothe, H.W.; Henderson, R.A.; Smith, B.F. A comparison of microRNA expression profiles from splenic hemangiosarcoma, splenic nodular hyperplasia, and normal spleens of dogs. BMC veterinary research 2016, 12. [Google Scholar] [CrossRef]
- Heishima, K.; Mori, T.; Sakai, H.; Sugito, N.; Murakami, M.; Yamada, N.; Akao, Y.; Maruo, K. MicroRNA-214 Promotes Apoptosis in Canine Hemangiosarcoma by Targeting the COP1-p53 Axis. PloS one 2015, 10, e0137361. [Google Scholar] [CrossRef]
- Lutful Kabir, F.M.; DeInnocentes, P.; Bird, R.C. Altered microRNA Expression Profiles and Regulation of INK4A/CDKN2A Tumor Suppressor Genes in Canine Breast Cancer Models. Journal of cellular biochemistry 2015, 116, 2956–2969. [Google Scholar] [CrossRef] [PubMed]
- Boggs, R.M.; Wright, Z.M.; Stickney, M.J.; Porter, W.W.; Murphy, K.E. MicroRNA expression in canine mammary cancer. Mamm Genome 2008, 19, 561–569. [Google Scholar] [CrossRef] [PubMed]
- von Deetzen, M.C.; Schmeck, B.T.; Gruber, A.D.; Klopfleisch, R. Malignancy Associated MicroRNA Expression Changes in Canine Mammary Cancer of Different Malignancies. ISRN veterinary science 2014, 2014, 148597. [Google Scholar] [CrossRef]
- Negrini, M.; Calin, G.A. Breast cancer metastasis: a microRNA story. Breast Cancer Res 2008, 10, 203. [Google Scholar] [CrossRef]
- Ma, L.; Reinhardt, F.; Pan, E.; Soutschek, J.; Bhat, B.; Marcusson, E.G.; Teruya-Feldstein, J.; Bell, G.W.; Weinberg, R.A. Therapeutic silencing of miR-10b inhibits metastasis in a mouse mammary tumor model. Nature biotechnology 2010, 28, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Yang, L.; Zhang, J. MiRNA5423p downregulation promotes trastuzumab resistance in breast cancer cells via AKT activation. Oncol Rep 2015, 33, 1215–1220. [Google Scholar] [CrossRef]
- Wang, W.; Zhao, L.; Wei, X.; Wang, L.; Liu, S.; Yang, Y.; Wang, F.; Sun, G.; Zhang, J.; Ma, Y.; et al. MicroRNA-320a promotes 5-FU resistance in human pancreatic cancer cells. Sci Rep 2016, 6, 27641. [Google Scholar] [CrossRef]
- Teplyuk, N.M.; Uhlmann, E.J.; Gabriely, G.; Volfovsky, N.; Wang, Y.; Teng, J.; Karmali, P.; Marcusson, E.; Peter, M.; Mohan, A.; et al. Therapeutic potential of targeting microRNA-10b in established intracranial glioblastoma: first steps toward the clinic. EMBO Mol Med 2016, 8, 268–287. [Google Scholar] [CrossRef]
- Yu, J.; Wang, J.G.; Zhang, L.; Yang, H.P.; Wang, L.; Ding, D.; Chen, Q.; Yang, W.L.; Ren, K.H.; Zhou, D.M.; et al. MicroRNA-320a inhibits breast cancer metastasis by targeting metadherin. Oncotarget 2016, 7, 38612–38625. [Google Scholar] [CrossRef]
- Lynch, J.; Fay, J.; Meehan, M.; Bryan, K.; Watters, K.M.; Murphy, D.M.; Stallings, R.L. MiRNA-335 suppresses neuroblastoma cell invasiveness by direct targeting of multiple genes from the non-canonical TGF-beta signalling pathway. Carcinogenesis 2012, 33, 976–985. [Google Scholar] [CrossRef] [PubMed]
- Bulkowska, M.; Rybicka, A.; Senses, K.M.; Ulewicz, K.; Witt, K.; Szymanska, J.; Taciak, B.; Klopfleisch, R.; Hellmen, E.; Dolka, I.; et al. MicroRNA expression patterns in canine mammary cancer show significant differences between metastatic and non-metastatic tumours. BMC cancer 2017, 17, 728. [Google Scholar] [CrossRef]
- Ren, X.; Fan, Y.; Shi, D.; Xu, E.; Liu, Y. MicroRNA-124 inhibits canine mammary carcinoma cell proliferation, migration and invasion by targeting CDH2. Res Vet Sci 2022, 146, 5–14. [Google Scholar] [CrossRef]
- Hino, Y.; Rahman, M.M.; Lai, Y.C.; Husna, A.A.; Chen, H.W.; Hasan, M.N.; Nakagawa, T.; Miura, N. Hypoxic miRNAs expression are different between primary and metastatic melanoma cells. Gene 2021, 782, 145552. [Google Scholar] [CrossRef] [PubMed]
- Sarver, A.L.; Thayanithy, V.; Scott, M.C.; Cleton-Jansen, A.M.; Hogendoorn, P.C.; Modiano, J.F.; Subramanian, S. MicroRNAs at the human 14q32 locus have prognostic significance in osteosarcoma. Orphanet journal of rare diseases 2013, 8, 7. [Google Scholar] [CrossRef]
- Leonardo, L.; Laura, P.; Serena, B.M. miR-1 and miR-133b expression in canine osteosarcoma. Res Vet Sci 2018, 117, 133–137. [Google Scholar] [CrossRef]
- Lopez, C.M.; Yu, P.Y.; Zhang, X.; Yilmaz, A.S.; London, C.A.; Fenger, J.M. MiR-34a regulates the invasive capacity of canine osteosarcoma cell lines. PloS one 2018, 13, e0190086. [Google Scholar] [CrossRef] [PubMed]
- Fenger, J.M.; Roberts, R.D.; Iwenofu, O.H.; Bear, M.D.; Zhang, X.; Couto, J.I.; Modiano, J.F.; Kisseberth, W.C.; London, C.A. MiR-9 is overexpressed in spontaneous canine osteosarcoma and promotes a metastatic phenotype including invasion and migration in osteoblasts and osteosarcoma cell lines. BMC Cancer 2016, 16, 784. [Google Scholar] [CrossRef]
- Xu, X.; Zhu, Y.; Liang, Z.; Li, S.; Xu, X.; Wang, X.; Wu, J.; Hu, Z.; Meng, S.; Liu, B.; et al. c-Met and CREB1 are involved in miR-433-mediated inhibition of the epithelial-mesenchymal transition in bladder cancer by regulating Akt/GSK-3beta/Snail signaling. Cell death & disease 2016, 7, e2088. [Google Scholar] [CrossRef]
- Takasaki, S. Roles of microRNAs in cancers and development. Methods Mol Biol 2015, 1218, 375–413. [Google Scholar] [CrossRef]
- Agarwal, P.; Crepps, M.P.; Stahr, N.A.; Kretzschmar, W.P.; Harris, H.C.; Prasad, N.; Levy, S.E.; Smith, B.F. Identification of canine circulating miRNAs as tumor biospecific markers using Next-Generation Sequencing and Q-RT-PCR. Biochem Biophys Rep 2021, 28, 101106. [Google Scholar] [CrossRef] [PubMed]
- Yao, S.; Zhao, X.; Wang, L.; Chen, F.; Gong, H.; Chen, C.; Cai, C. pH-activated DNA nanomachine for miRNA-21 imaging to accurately identify cancer cell. Mikrochim Acta 2022, 189, 266. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.G.; Calin, G.A.; Meloon, B.; Gamliel, N.; Sevignani, C.; Ferracin, M.; Dumitru, C.D.; Shimizu, M.; Zupo, S.; Dono, M.; et al. An oligonucleotide microchip for genome-wide microRNA profiling in human and mouse tissues. P Natl Acad Sci USA 2004, 101, 9740–9744. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA expression profiles classify human cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Volinia, S.; Calin, G.A.; Liu, C.G.; Ambs, S.; Cimmino, A.; Petrocca, F.; Visone, R.; Iorio, M.; Roldo, C.; Ferracin, M.; et al. A microRNA expression signature of human solid tumors defines cancer gene targets. Proc Natl Acad Sci U S A 2006, 103, 2257–2261. [Google Scholar] [CrossRef]
- Yanaihara, N.; Caplen, N.; Bowman, E.; Seike, M.; Kumamoto, K.; Yi, M.; Stephens, R.M.; Okamoto, A.; Yokota, J.; Tanaka, T.; et al. Unique microRNA molecular profiles in lung cancer diagnosis and prognosis. Cancer Cell 2006, 9, 189–198. [Google Scholar] [CrossRef]
- Rosenfeld, N.; Aharonov, R.; Meiri, E.; Rosenwald, S.; Spector, Y.; Zepeniuk, M.; Benjamin, H.; Shabes, N.; Tabak, S.; Levy, A.; et al. MicroRNAs accurately identify cancer tissue origin. Nat Biotechnol 2008, 26, 462–469. [Google Scholar] [CrossRef]
- Iwasaki, H.; Shimura, T.; Kitagawa, M.; Yamada, T.; Nishigaki, R.; Fukusada, S.; Okuda, Y.; Katano, T.; Horike, S.I.; Kataoka, H. A Novel Urinary miRNA Biomarker for Early Detection of Colorectal Cancer. Cancers (Basel) 2022, 14. [Google Scholar] [CrossRef]
- Bidarra, D.; Constancio, V.; Barros-Silva, D.; Ramalho-Carvalho, J.; Moreira-Barbosa, C.; Antunes, L.; Mauricio, J.; Oliveira, J.; Henrique, R.; Jeronimo, C. Circulating MicroRNAs as Biomarkers for Prostate Cancer Detection and Metastasis Development Prediction. Front Oncol 2019, 9, 900. [Google Scholar] [CrossRef]
- Downs-Holmes, C.; Silverman, P. Breast cancer: overview & updates. Nurse Pract 2011, 36, 20–26. [Google Scholar] [CrossRef]
- Vanacore, D.; Boccellino, M.; Rossetti, S.; Cavaliere, C.; D'Aniello, C.; Di Franco, R.; Romano, F.J.; Montanari, M.; La Mantia, E.; Piscitelli, R.; et al. Micrornas in prostate cancer: an overview. Oncotarget 2017, 8, 50240–50251. [Google Scholar] [CrossRef]
- Guo, X.; Han, T.; Hu, P.; Guo, X.; Zhu, C.; Wang, Y.; Chang, S. Five microRNAs in serum as potential biomarkers for prostate cancer risk assessment and therapeutic intervention. Int Urol Nephrol 2018, 50, 2193–2200. [Google Scholar] [CrossRef]
- Jackson, B.L.; Grabowska, A.; Ratan, H.L. MicroRNA in prostate cancer: functional importance and potential as circulating biomarkers. BMC Cancer 2014, 14, 930. [Google Scholar] [CrossRef]
- Lyu, J.; Zhao, L.; Wang, F.; Ji, J.; Cao, Z.; Xu, H.; Shi, X.; Zhu, Y.; Zhang, C.; Guo, F.; et al. Discovery and Validation of Serum MicroRNAs as Early Diagnostic Biomarkers for Prostate Cancer in Chinese Population. Biomed Res Int 2019, 2019, 9306803. [Google Scholar] [CrossRef]
- Ogata-Kawata, H.; Izumiya, M.; Kurioka, D.; Honma, Y.; Yamada, Y.; Furuta, K.; Gunji, T.; Ohta, H.; Okamoto, H.; Sonoda, H.; et al. Circulating exosomal microRNAs as biomarkers of colon cancer. PLoS One 2014, 9, e92921. [Google Scholar] [CrossRef]
- Fogli, S.; Polini, B.; Carpi, S.; Pardini, B.; Naccarati, A.; Dubbini, N.; Lanza, M.; Breschi, M.C.; Romanini, A.; Nieri, P. Identification of plasma microRNAs as new potential biomarkers with high diagnostic power in human cutaneous melanoma. Tumour Biol 2017, 39, 1010428317701646. [Google Scholar] [CrossRef]
- Xie, Z.; Chen, G.; Zhang, X.; Li, D.; Huang, J.; Yang, C.; Zhang, P.; Qin, Y.; Duan, Y.; Gong, B.; et al. Salivary microRNAs as promising biomarkers for detection of esophageal cancer. PLoS One 2013, 8, e57502. [Google Scholar] [CrossRef]
- Humeau, M.; Vignolle-Vidoni, A.; Sicard, F.; Martins, F.; Bournet, B.; Buscail, L.; Torrisani, J.; Cordelier, P. Salivary MicroRNA in Pancreatic Cancer Patients. PLoS One 2015, 10, e0130996. [Google Scholar] [CrossRef]
- Yu, S.L.; Chen, H.Y.; Chang, G.C.; Chen, C.Y.; Chen, H.W.; Singh, S.; Cheng, C.L.; Yu, C.J.; Lee, Y.C.; Chen, H.S.; et al. MicroRNA signature predicts survival and relapse in lung cancer. Cancer Cell 2008, 13, 48–57. [Google Scholar] [CrossRef]
- Schetter, A.J.; Leung, S.Y.; Sohn, J.J.; Zanetti, K.A.; Bowman, E.D.; Yanaihara, N.; Yuen, S.T.; Chan, T.L.; Kwong, D.L.; Au, G.K.; et al. MicroRNA expression profiles associated with prognosis and therapeutic outcome in colon adenocarcinoma. JAMA 2008, 299, 425–436. [Google Scholar] [CrossRef]
- Gahlawat, A.W.; Witte, T.; Haarhuis, L.; Schott, S. A novel circulating miRNA panel for non-invasive ovarian cancer diagnosis and prognosis. Br J Cancer 2022, 127, 1550–1556. [Google Scholar] [CrossRef]
- Farran, B.; Dyson, G.; Craig, D.; Dombkowski, A.; Beebe-Dimmer, J.L.; Powell, I.J.; Podgorski, I.; Heilbrun, L.; Bolton, S.; Bock, C.H. A study of circulating microRNAs identifies a new potential biomarker panel to distinguish aggressive prostate cancer. Carcinogenesis 2018, 39, 556–561. [Google Scholar] [CrossRef]
- Shukla, K.K.; Misra, S.; Pareek, P.; Mishra, V.; Singhal, B.; Sharma, P. Recent scenario of microRNA as diagnostic and prognostic biomarkers of prostate cancer. Urol Oncol 2017, 35, 92–101. [Google Scholar] [CrossRef]
- Maierthaler, M.; Benner, A.; Hoffmeister, M.; Surowy, H.; Jansen, L.; Knebel, P.; Chang-Claude, J.; Brenner, H.; Burwinkel, B. Plasma miR-122 and miR-200 family are prognostic markers in colorectal cancer. Int J Cancer 2017, 140, 176–187. [Google Scholar] [CrossRef]
- Monzo, M.; Santasusagna, S.; Moreno, I.; Martinez, F.; Hernandez, R.; Munoz, C.; Castellano, J.J.; Moreno, J.; Navarro, A. Exosomal microRNAs isolated from plasma of mesenteric veins linked to liver metastases in resected patients with colon cancer. Oncotarget 2017, 8, 30859–30869. [Google Scholar] [CrossRef]
- Chen, J.; Wang, W.; Zhang, Y.; Chen, Y.; Hu, T. Predicting distant metastasis and chemoresistance using plasma miRNAs. Med Oncol 2014, 31, 799. [Google Scholar] [CrossRef]
- Bhatnagar, N.; Li, X.; Padi, S.K.; Zhang, Q.; Tang, M.S.; Guo, B. Downregulation of miR-205 and miR-31 confers resistance to chemotherapy-induced apoptosis in prostate cancer cells. Cell Death Dis 2010, 1, e105. [Google Scholar] [CrossRef]
- Li, B.; Shi, X.B.; Nori, D.; Chao, C.K.; Chen, A.M.; Valicenti, R.; White Rde, V. Down-regulation of microRNA 106b is involved in p21-mediated cell cycle arrest in response to radiation in prostate cancer cells. Prostate 2011, 71, 567–574. [Google Scholar] [CrossRef]
- Mao, A.; Zhao, Q.; Zhou, X.; Sun, C.; Si, J.; Zhou, R.; Gan, L.; Zhang, H. MicroRNA-449a enhances radiosensitivity by downregulation of c-Myc in prostate cancer cells. Sci Rep 2016, 6, 27346. [Google Scholar] [CrossRef]
- Nagesh, P.K.B.; Chowdhury, P.; Hatami, E.; Boya, V.K.N.; Kashyap, V.K.; Khan, S.; Hafeez, B.B.; Chauhan, S.C.; Jaggi, M.; Yallapu, M.M. miRNA-205 Nanoformulation Sensitizes Prostate Cancer Cells to Chemotherapy. Cancers (Basel) 2018, 10. [Google Scholar] [CrossRef]
- Ni, J.; Bucci, J.; Chang, L.; Malouf, D.; Graham, P.; Li, Y. Targeting MicroRNAs in Prostate Cancer Radiotherapy. Theranostics 2017, 7, 3243–3259. [Google Scholar] [CrossRef]
- Thieu, W.; Tilki, D.; de Vere White, R.; Evans, C.P. The role of microRNA in castration-resistant prostate cancer. Urol Oncol 2014, 32, 517–523. [Google Scholar] [CrossRef]
- Wang, Y.; Lieberman, R.; Pan, J.; Zhang, Q.; Du, M.; Zhang, P.; Nevalainen, M.; Kohli, M.; Shenoy, N.K.; Meng, H.; et al. miR-375 induces docetaxel resistance in prostate cancer by targeting SEC23A and YAP1. Mol Cancer 2016, 15, 70. [Google Scholar] [CrossRef]
- Hu, Q.; Gong, J.P.; Li, J.; Zhong, S.L.; Chen, W.X.; Zhang, J.Y.; Ma, T.F.; Ji, H.; Lv, M.M.; Zhao, J.H.; et al. Down-regulation of miRNA-452 is associated with adriamycin-resistance in breast cancer cells. Asian Pac J Cancer Prev 2014, 15, 5137–5142. [Google Scholar] [CrossRef]
- Safaei, S.; Amini, M.; Najjary, S.; Mokhtarzadeh, A.; Bolandi, N.; Saeedi, H.; Alizadeh, N.; Javadrashid, D.; Baradaran, B. miR-200c increases the sensitivity of breast cancer cells to Doxorubicin through downregulating MDR1 gene. Exp Mol Pathol 2022, 125, 104753. [Google Scholar] [CrossRef]
- Hu, Y.; Qiu, Y.; Yagüe, E.; Ji, W.; Liu, J.; Zhang, J. miRNA-205 targets VEGFA and FGF2 and regulates resistance to chemotherapeutics in breast cancer. Cell Death & Disease 2016, 7, e2291–e2291. [Google Scholar] [CrossRef]
- Wu, Z.H.; Tao, Z.H.; Zhang, J.; Li, T.; Ni, C.; Xie, J.; Zhang, J.F.; Hu, X.C. MiRNA-21 induces epithelial to mesenchymal transition and gemcitabine resistance via the PTEN/AKT pathway in breast cancer. Tumour Biol 2016, 37, 7245–7254. [Google Scholar] [CrossRef]
- Niu, J.; Xue, A.; Chi, Y.; Xue, J.; Wang, W.; Zhao, Z.; Fan, M.; Yang, C.H.; Shao, Z.M.; Pfeffer, L.M.; et al. Induction of miRNA-181a by genotoxic treatments promotes chemotherapeutic resistance and metastasis in breast cancer. Oncogene 2016, 35, 1302–1313. [Google Scholar] [CrossRef]
- Liu, H.; Wang, Y.; Li, X.; Zhang, Y.J.; Li, J.; Zheng, Y.Q.; Liu, M.; Song, X.; Li, X.R. Expression and regulatory function of miRNA-182 in triple-negative breast cancer cells through its targeting of profilin 1. Tumour Biol 2013, 34, 1713–1722. [Google Scholar] [CrossRef]
- Xue, J.; Chi, Y.; Chen, Y.; Huang, S.; Ye, X.; Niu, J.; Wang, W.; Pfeffer, L.M.; Shao, Z.M.; Wu, Z.H.; et al. MiRNA-621 sensitizes breast cancer to chemotherapy by suppressing FBXO11 and enhancing p53 activity. Oncogene 2016, 35, 448–458. [Google Scholar] [CrossRef]
- Mohammadi Yeganeh, S.; Vasei, M.; Tavakoli, R.; Kia, V.; Paryan, M. The effect of miR-340 over-expression on cell-cycle-related genes in triple-negative breast cancer cells. Eur J Cancer Care (Engl) 2017, 26. [Google Scholar] [CrossRef]
- Nordentoft, I.; Birkenkamp-Demtroder, K.; Agerbaek, M.; Theodorescu, D.; Ostenfeld, M.S.; Hartmann, A.; Borre, M.; Orntoft, T.F.; Dyrskjot, L. miRNAs associated with chemo-sensitivity in cell lines and in advanced bladder cancer. BMC Med Genomics 2012, 5, 40. [Google Scholar] [CrossRef]
- Heishima, K.; Ichikawa, Y.; Yoshida, K.; Iwasaki, R.; Sakai, H.; Nakagawa, T.; Tanaka, Y.; Hoshino, Y.; Okamura, Y.; Murakami, M.; et al. Circulating microRNA-214 and -126 as potential biomarkers for canine neoplastic disease. Sci Rep 2017, 7, 2301. [Google Scholar] [CrossRef]
- Cleveland, M.J.; Casale, S. Incidence of malignancy and outcomes for dogs undergoing splenectomy for incidentally detected nonruptured splenic nodules or masses: 105 cases (2009-2013). J Am Vet Med Assoc 2016, 248, 1267–1273. [Google Scholar] [CrossRef]
- Kent, M.S.; Zwingenberger, A.; Westropp, J.L.; Barrett, L.E.; Durbin-Johnson, B.P.; Ghosh, P.; Vinall, R.L. MicroRNA profiling of dogs with transitional cell carcinoma of the bladder using blood and urine samples. BMC veterinary research 2017, 13, 339. [Google Scholar] [CrossRef]
- Vinall, R.L.; Kent, M.S.; deVere White, R.W. Expression of microRNAs in urinary bladder samples obtained from dogs with grossly normal bladders, inflammatory bladder disease, or transitional cell carcinoma. American journal of veterinary research 2012, 73, 1626–1633. [Google Scholar] [CrossRef]
- Zamarian, V.; Ferrari, R.; Stefanello, D.; Ceciliani, F.; Grieco, V.; Minozzi, G.; Chiti, L.E.; Arigoni, M.; Calogero, R.; Lecchi, C. miRNA profiles of canine cutaneous mast cell tumours with early nodal metastasis and evaluation as potential biomarkers. Sci Rep 2020, 10, 18918. [Google Scholar] [CrossRef]
- Fish, E.J.; Martinez-Romero, E.G.; DeInnocentes, P.; Koehler, J.W.; Prasad, N.; Smith, A.N.; Bird, R.C. Circulating microRNA as biomarkers of canine mammary carcinoma in dogs. J Vet Intern Med 2020, 34, 1282–1290. [Google Scholar] [CrossRef]
- Garnica, T.K.; Lesbon, J.C.C.; Avila, A.; Rochetti, A.L.; Matiz, O.R.S.; Ribeiro, R.C.S.; Zoppa, A.; Nishiya, A.T.; Costa, M.T.; de Nardi, A.B.; et al. Liquid biopsy based on small extracellular vesicles predicts chemotherapy response of canine multicentric lymphomas. Sci Rep 2020, 10, 20371. [Google Scholar] [CrossRef]
- Beaumier, A.; Robinson, S.R.; Robinson, N.; Lopez, K.E.; Meola, D.M.; Barber, L.G.; Bulmer, B.J.; Calvalido, J.; Rush, J.E.; Yeri, A.; et al. Extracellular vesicular microRNAs as potential biomarker for early detection of doxorubicin-induced cardiotoxicity. J Vet Intern Med 2020, 34, 1260–1271. [Google Scholar] [CrossRef]
- Husna, A.A.; Rahman, M.M.; Lai, Y.C.; Chen, H.W.; Hasan, M.N.; Nakagawa, T.; Miura, N. Identification of melanoma-specific exosomal miRNAs as the potential biomarker for canine oral melanoma. Pigment Cell Melanoma Res 2021, 34, 1062–1073. [Google Scholar] [CrossRef]
- Narita, M.; Nishida, H.; Asahina, R.; Nakata, K.; Yano, H.; Dickinson, P.J.; Tanaka, T.; Akiyoshi, H.; Maeda, S.; Kamishina, H. Expression of microRNAs in plasma and in extracellular vesicles derived from plasma for dogs with glioma and dogs with other brain diseases. Am J Vet Res 2020, 81, 355–360. [Google Scholar] [CrossRef]
- Starkey, M.P.; Compston-Garnett, L.; Malho, P.; Dunn, K.; Dubielzig, R. Metastasis-associated microRNA expression in canine uveal melanoma. Vet Comp Oncol 2018, 16, 81–89. [Google Scholar] [CrossRef]
- Leonardi, L.; Scotlandi, K.; Pettinari, I.; Benassi, M.S.; Porcellato, I.; Pazzaglia, L. MiRNAs in Canine and Human Osteosarcoma: A Highlight Review on Comparative Biomolecular Aspects. Cells 2021, 10. [Google Scholar] [CrossRef]
- Sethi, S.; Ali, S.; Sethi, S.; Sarkar, F.H. MicroRNAs in personalized cancer therapy. Clin Genet 2014, 86, 68–73. [Google Scholar] [CrossRef]
- Costa, P.M.; Pedroso de Lima, M.C. MicroRNAs as Molecular Targets for Cancer Therapy: On the Modulation of MicroRNA Expression. Pharmaceuticals (Basel) 2013, 6, 1195–1220. [Google Scholar] [CrossRef]
- Liu, Y.; Zheng, M.; Jiao, M.; Yan, C.; Xu, S.; Du, Q.; Morsch, M.; Yin, J.; Shi, B. Polymeric nanoparticle mediated inhibition of miR-21 with enhanced miR-124 expression for combinatorial glioblastoma therapy. Biomaterials 2021, 276, 121036. [Google Scholar] [CrossRef]
- Yan, S.; Liu, G.; Pei, C.; Chen, W.; Li, P.; Wang, Q.; Jin, X.; Zhu, J.; Wang, M.; Liu, X. Inhibition of NADPH oxidase protects against metastasis of human lung cancer by decreasing microRNA-21. Anticancer Drugs 2015, 26, 388–398. [Google Scholar] [CrossRef]
- Dong, C.G.; Wu, W.K.; Feng, S.Y.; Wang, X.J.; Shao, J.F.; Qiao, J. Co-inhibition of microRNA-10b and microRNA-21 exerts synergistic inhibition on the proliferation and invasion of human glioma cells. Int J Oncol 2012, 41, 1005–1012. [Google Scholar] [CrossRef]
- Cheng, C.J.; Bahal, R.; Babar, I.A.; Pincus, Z.; Barrera, F.; Liu, C.; Svoronos, A.; Braddock, D.T.; Glazer, P.M.; Engelman, D.M.; et al. MicroRNA silencing for cancer therapy targeted to the tumour microenvironment. Nature 2015, 518, 107–110. [Google Scholar] [CrossRef]
- Song, H.; Oh, B.; Choi, M.; Oh, J.; Lee, M. Delivery of anti-microRNA-21 antisense-oligodeoxynucleotide using amphiphilic peptides for glioblastoma gene therapy. J Drug Target 2015, 23, 360–370. [Google Scholar] [CrossRef]
- Bahreyni, A.; Liu, H.; Mohamud, Y.; Xue, Y.C.; Zhang, J.; Luo, H. A new miRNA-Modified coxsackievirus B3 inhibits triple negative breast cancer growth with improved safety profile in immunocompetent mice. Cancer Lett 2022, 548, 215849. [Google Scholar] [CrossRef]
- Ren, Y.; Zhou, X.; Mei, M.; Yuan, X.B.; Han, L.; Wang, G.X.; Jia, Z.F.; Xu, P.; Pu, P.Y.; Kang, C.S. MicroRNA-21 inhibitor sensitizes human glioblastoma cells U251 (PTEN-mutant) and LN229 (PTEN-wild type) to taxol. BMC Cancer 2010, 10, 27. [Google Scholar] [CrossRef]
- Yoshikawa, R.; Maeda, A.; Ueno, Y.; Sakai, H.; Kimura, S.; Sawadaishi, T.; Kohgo, S.; Yamada, K.; Mori, T. Intraperitoneal administration of synthetic microRNA-214 elicits tumor suppression in an intraperitoneal dissemination mouse model of canine hemangiosarcoma. Vet Res Commun 2022, 46, 447–457. [Google Scholar] [CrossRef]
- Yoshikawa, R.; Mori, T.; Noguchi, S.; Akao, Y.; Maruo, K.; Kitade, Y. Synthetic microRNA-205 exhibited tumour suppression in spontaneous canine malignant melanoma by intratumoral injection. Vet Comp Oncol 2019, 17, 407–412. [Google Scholar] [CrossRef]
- Alegre, F.; Ormonde, A.R.; Snider, K.M.; Woolard, K.; Yu, A.M.; Wittenburg, L.A. A genetically engineered microRNA-34a prodrug demonstrates anti-tumor activity in a canine model of osteosarcoma. PLoS One 2018, 13, e0209941. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
miRNAs can be classified as oncomiRs and tumor suppressors. A. OncomiRs suppress tumor suppressor gene translation and promote tumor cell growth through constitutive overexpression. B. Tumor suppressor miRNAs inhibit tumorigenesis and subsequent cancer development by suppressing the translation of mRNAs that encode for oncogenes. C. Hallmarks of carcinogenesis. This figure was created using Biorender.
Figure 1.
miRNAs can be classified as oncomiRs and tumor suppressors. A. OncomiRs suppress tumor suppressor gene translation and promote tumor cell growth through constitutive overexpression. B. Tumor suppressor miRNAs inhibit tumorigenesis and subsequent cancer development by suppressing the translation of mRNAs that encode for oncogenes. C. Hallmarks of carcinogenesis. This figure was created using Biorender.
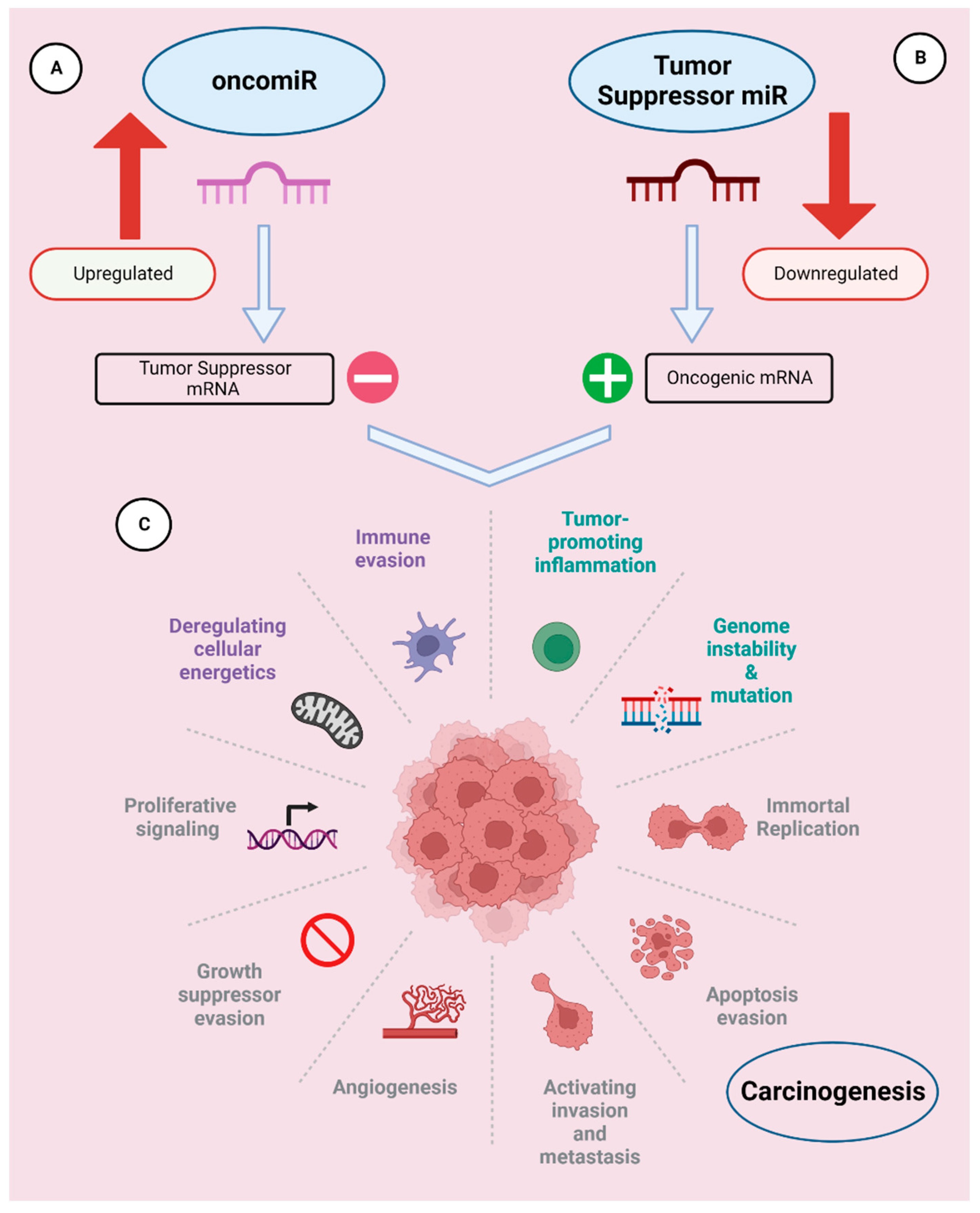
Figure 2.
MiRNAs as cancer biomarkers: potential clinical applications. miRNAs are isolated from tumor tissue, saliva, blood, plasma, serum, etc., and then profiled for miRNA expression. miRNAs profiling helps in cancer diagnosis, cancer staging, and therapeutics. This figure was created using Biorender.
Figure 2.
MiRNAs as cancer biomarkers: potential clinical applications. miRNAs are isolated from tumor tissue, saliva, blood, plasma, serum, etc., and then profiled for miRNA expression. miRNAs profiling helps in cancer diagnosis, cancer staging, and therapeutics. This figure was created using Biorender.
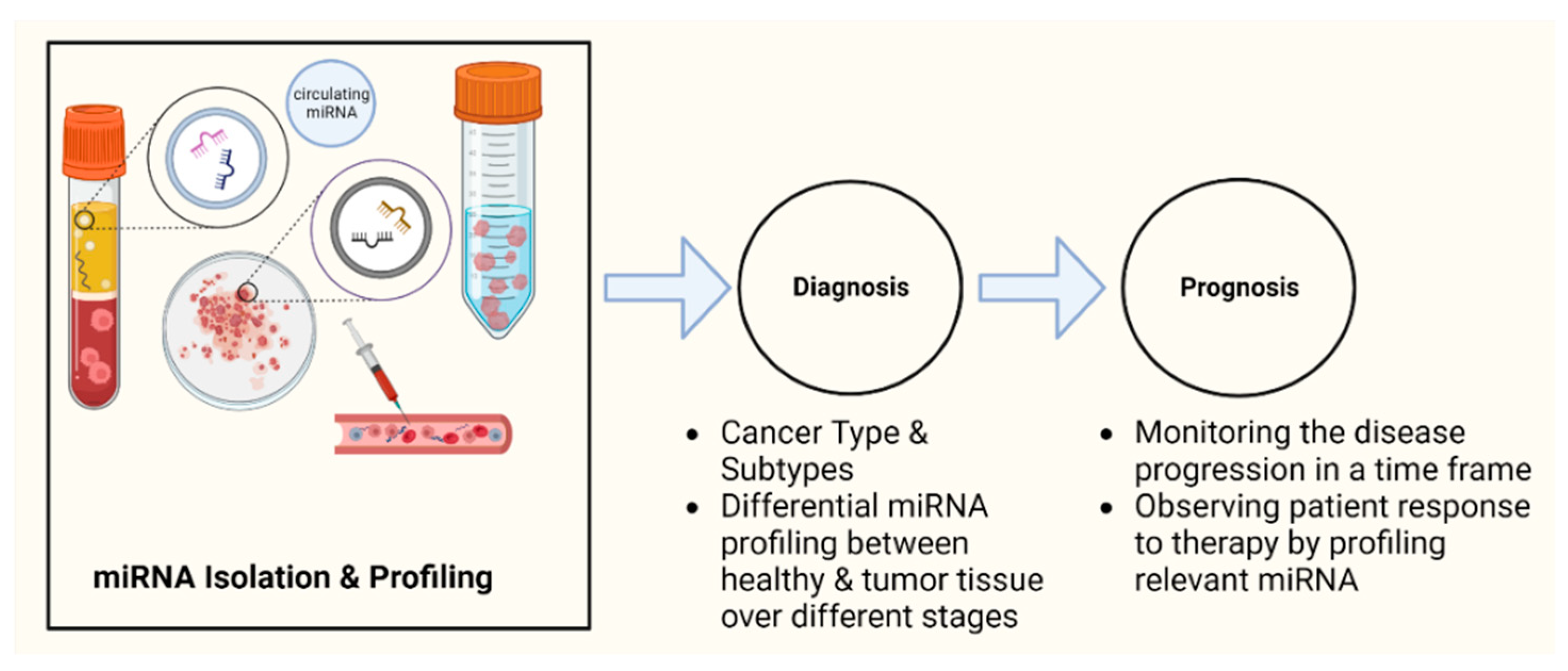
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



