Submitted:
01 June 2023
Posted:
02 June 2023
You are already at the latest version
Abstract
Sweet sorghum is an important bioenergy grass and valuable forage with a strong adaptability to saline environments. However, little is known about the mechanisms of sweet sorghum coping with ion toxicity under salt stresses. Here, we first evaluated the salt tolerance of a sweet sorghum cultivar “Lvjuren” and determined its ion accumulation traits under NaCl treatments; then explored key genes involved in Na+, Cl−, K+ and NO3− transport using transcriptome profiling and qRT-PCR method. The results showed that the growth and photosynthesis of sweet sorghum were unaffected by 50 and 100 mM NaCl treatments, indicative of a strong tolerance of this species to salt stresses. Under NaCl treatments, sweet sorghum could efficiently exclude Na+ from shoots and accumulate Cl− in leaf sheaths to avoid their overaccumulation in leaf blades; meanwhile, it possessed a prominent ability to sustain NO3− homeostasis in leaf blades. Transcriptome profiling identified several differentially expressed genes associated with Na+, Cl−, K+ and NO3− transport in roots, leaf sheaths and leaf blades of sweet sorghum after 200 mM NaCl treatment for 6 and 48 h. Moreover, transcriptome data and qRT-PCR results indicated that HKT1;5, CLCc and NPF7.3-1 should be key genes involved in Na+ retention in roots, Cl− accumulation in leaf sheaths and maintenance of NO3− homeostasis in leaf blades, respectively. Many TFs were also identified after NaCl treatment, which should play important regulatory roles in salt tolerance of sweet sorghum. This work lays a preliminary foundation for clarifying the molecular basis underlying the adaptation of sweet sorghum to adverse environments.
Keywords:
soil salinity
; sodium
; chloride
; ion transporters
; transcriptome factors
1. Introduction
Soil salinity is one of the major environmental constraints on plant growth and crop production [1]. There are exceeding 1 billion ha lands affected by salinity worldwide [2]. In China, approximately 30 percent of the total 3.6×107 ha salt-affected lands are potentially arable [3]. Therefore, with population expansion, urban spread and climate change, the use of salinized lands to cultivate crops and forages is an important strategy to ensure food security and promote ecological restoration [4].
Sweet sorghum [Sorghum bicolor (L.) Moench], an annual C4 plant belonging to Poaceae, is a natural variation of grain sorghum [5]. This species is characterized by high fermentable sugars in the juice of the stalks, which makes it attractive as a valuable bioenergy crop [6,7]. Meanwhile, sweet sorghum has been widely used as a forage due to its high biomass and growth rate, as well as prominent palatability and digestibility [8]. Furthermore, different with traditional crop species, sweet sorghum can adapt well to various environmental stresses including salinity, drought and flood, serving as a pioneer plant for recovering saline and marginal lands [5,9]. Combining these eminent traits, there has been increasing practices to evaluate field performance of sweet sorghum in salt-affected lands in China [2,3]. Thus, the understanding of mechanisms employed by sweet sorghum to adapt to adverse environments will lay a theoretical basis for the large-scale cultivation of this species in salinized areas.
Na+ and Cl−, the dominant inorganic ions in saline soil, are toxic to plants by inhibiting enzyme activity, disturbing nutrient balance, hindering photosynthesis, etc. [10,11]. Plants mainly decrease the toxic effects of Na+ and Cl− by excluding or translocating them through ion transporters and channels [12,13]. It has been found that most species in Poaceae cope with Na+-toxicity by maintaining a low Na+ content in leaves (termed Na+ exclusion trait) [4], which is mainly achieved by restricting the long-distance transport of Na+ from roots into shoots, and several key proteins involved in this process such as HKT1;5 and HAK4 has been identified [14,15]. Physiological studies on wheat (Triticum aestivum) and rice (Oryza sativa) find that Cl− content in leaves or shoots was much higher than that in roots under NaCl treatments [16,17], indicating that these species cannot efficiently restrict the transport of Cl− into shoots under salt stress. Although the Cl− tolerance in the model plant Arabidopsis and some Cl−-sensitive plants such as Glycine max and Citus spp. has been investigated, and key proteins mediating Cl− transport such as CLCs, NPFs and CCC1 in these plant species have been functional characterized in recent years [18], the mechanisms underlying how Poaceae plants adapt to Cl−-toxicity still remain elusive.
Leaf sheath at the base of leaf tissue is a special structure evolved by Poaceae plants, which plays an essential role in supporting leaf blades, reserving nutrients and resisting chilling stress [19]. Furthermore, it has been proven that durum wheat (Triticum turgidum L. subsp. durum Desf.) can accumulate large amounts of Na+ in leaf sheaths to decrease Na+ content in leaf blades under salt stresses [20], suggesting that leaf sheath might act as a useful “ion reservoir” in shoots to avoid ion overaccumulation in leaf blades. Therefore, the further study on the role of leaf sheath in decreasing ion toxicity would provide new insights for the understanding of salt tolerance mechanisms of Poaceae plants.
Given the strong resistance of sweet sorghum to abiotic stresses, it is considered an important resource for exploring mechanisms and gene resources that can be used in the improvement of crop responses to environmental stresses [2]. Researchers have investigated ion accumulation characteristics in roots and shoots of sweet sorghum under saline conditions [21]. However, Na+ and Cl− accumulation traits in leaf sheaths of this species have not been reported. In addition, although transcriptome factors such as WRKY50 have been proven to play a key role in regulating Na+ transport in sweet sorghum under NaCl stress [22], the study on the function of Na+ and Cl− transporters or channels in salt tolerance of sweet sorghum still lags behind.
In this study, we first evaluated the physiological response of a sweet sorghum cultivar to NaCl treatments by measuring growth and photosynthesis indexes, and then determined Na+, K+, Cl− and NO3− contents in roots, leaf sheaths and leaf blades under NaCl treatments. Subsequently, we investigated the expression changes of genes involved in ion transport or encoding transcriptome factors after NaCl treatment by transcriptome sequencing. Finally, we analyzed expression patterns of HKT1;5, CLCc and NPF7.3 in sweet sorghum after NaCl treatment using qRT-PCR method.
2. Results
2.1. The Effect of NaCl Treatments on Growth and Photosynthesis of Sweet Sorghum
After 50 and 100 mM NaCl treatments, leaf blades of seedlings were healthy, while leaf blades of 200 mM NaCl treated seedlings were visually wilting (Figure 1A). Compared with the control, 50 mM NaCl treatment had no effect on plant height (PH), fresh weight (FW) and dry weight (DW) of roots and shoots, as well as shoot water content (WC) (Figure 1B,C). 100 mM NaCl treatment significantly decreased PH, while it had no effect on tissue biomass and shoot WC when compared with the control (Figure 1B,C). Differently, in comparison with the control, 200 mM NaCl treatment sharply declined abovementioned parameters (excepted for root DW, Figure 1B,C).
The net photosynthetic rate (Pn) and stomatal conductance (Gs) under 50 and 100 mM NaCl treatments were maintained at the same level as those under the control condition; in contrast, Pn and Gs under 200 mM NaCl treatment were significantly decreased (Figure 2A,B). In comparison with the control, all the NaCl treatments had no effect on chlorophyll b content, but 100 and 200 mM NaCl treatments significantly decreased chlorophyll a content (Figure 2C,D). These results suggested that sweet sorghum cultivar “Lvjuren” could well tolerate to 50 and 100 mM NaCl treatments, while its growth and photosynthesis were inhibited by 200 mM NaCl treatment.
2.2. The Ion Contents in Different Tissues of Sweet Sorghum under NaCl Treatments
Compared with the control, Na+ content in roots, leaf sheaths and leaf blades was gradually increased after 50-200 mM NaCl treatments (Figure 3A). It was obvious that Na+ content in roots was much higher than in shoots under NaCl treatments (Figure 3A). Meanwhile, in shoots, leaf sheath Na+ content under all treatments was significantly higher than leaf blade Na+ content (Figure 3A). In comparison with the control, all NaCl treatments significantly decreased K+ content in roots and leaf sheaths; differently, only 200 mM NaCl treatment obviously decreased K+ content in leaf blades (Figure 3B).
After treatment with 50-200 mM NaCl, Cl− content in all tissues was dramatically increased when compared with that under the control condition (Figure 3C). In contrast to tissue Na+ distribution pattern, Cl− content in roots was clearly lower than in shoots under all treatments (Figure 3C). Meanwhile, Cl− content in leaf sheaths was approximately 2 times higher than in leaf blades (Figure 3C). Compared with the control, 100 and 200 mM NaCl treatments significantly decreased NO3− content in roots and leaf sheaths, while all the salt treatments had no effect on NO3− content in leaf blades (Figure 3D).
2.3. RNA-seq Analysis of Sweet Sorghum under NaCl Stress
After irrigation with Hoagland solution (C), or treatment with 200 mM NaCl (S) for 6 and 48h, we collected root (R), leaf sheath (LS) and leaf blade (LB) samples for transcriptome sequencing. In total, 36 mRNA sequencing libraries (C6R1-3, S6R1-3, C6LS1-3, S6LS1-3, C6LB1-3, S6LB1-3, C24R1-3, S24R1-3, C24LS1-3, S24LS1-3, C24LB1-3, S24LB1-3) were finally generated. As shown in Table S2, in each library, at least 200 million clean reads were obtained by RNA-seq, with clean bases > 6.01 Gb and GC content > 51.5%. The Q30 value of these libraries was more than 87.87% (Table S2). By mapping the clean reads in each library to the sorghum reference genome sequence (NCBI accession number: GCF_000003195.3), it was found that the percentage of mapped reads in these libraries was from 82.37% to 92.53% (Table S3). There were totally 5952 new genes (termed Sorghum_bicolor_newGene_1, 2, 3…..) that cannot mapped to the reference genome sequence, among which 2574 members were functionally annotated by alignment against protein databases (Table S4).
2.4. Identification of DEGs Related to Ion Transport in Roots, Leaf Sheaths and Leaf Blades after NaCl Treatment for 6 and 24 h
After 200 mM NaCl treatment for 6 h, in total, 3843 (2355 upregulated, 1488 downregulated), 1895 (688 upregulated, 1207 downregulated) and 2355 (1409 upregulated, 946 downregulated) DEGs were identified in roots, leaf sheaths and leaf blades, respectively; after NaCl treatment for 48 h, in total, 1933 (1219 upregulated, 714 downregulated), 1270 (713 upregulated, 557 downregulated) and 4103 (1960 upregulated, 2143 downregulated) DEGs were identified in roots, leaf sheaths and leaf blades, respectively (Figure S1).
Subsequently, we analyzed the effects of NaCl treatment on transcript levels of DEGs related to Na+, K+, Cl− and NO3− transport in different tissues. As shown in Figure 4A and Table S5, after NaCl treatment for 6 h, 33 upregulated DEGs, including NHX, CHX, CCX, NCX, CNGC, HKT, KEA, HAK/KT/KUP, AKT and KOR that are probably involved in Na+ and/or K+ transport, CLC, SLAH, ALMT, NPF that are probably involved in Cl− and/or NO3− transport, as well as H+-ATPase and Ca2+-ATPase that provide H+ or Ca2+ pump for ion transport, 15 downregulated DEGs, including KEA, HAK/KT/KUP, AKT, KOR, CLC, SLAH and NPF, were identified in roots of sweet sorghum. After NaCl treatment for 48 h, the number of DEGs related to ion transport was declined, but 1 CNGC, CLC, ALMT and H+-ATPase, 2 CHX, CCX and SLAH, 5 HAK/KT/KUP and 11 NPF were still upregulated (Figure 4B and Table S6). In leaf sheaths, after NaCl treatment for 6 and 48 h, only 11 and 12 upregulated DEGs related to Na+, K+, Cl− and NO3− transport were identified, respectively, and several members that were detective in roots such as NHX, CHX, CNGC, KEA, KOR, SLAH and ALMT were not identified (Figure 5, Tables S7 and S8). In leaf blades, after NaCl treatment for 6 and 48 h, 17 and 19 upregulated DEGs, as well as 8 and 26 downregulated DEGs, respectively, were identified (Figure 6, Tables S9 and S10). It was found that some DEGs such as AKT, KEA and SLAH were only detective in leaf blades under NaCl treatment for 48 h (Figure 6, Tables S9 and S10), suggesting that these genes might be mainly responsible for long-term salt stress.
The heat map showed that, in roots, there were 24 DEGs related to ion transport after NaCl treatment for both 6 and 48 h, the majority of these DEGs was upregulated (Figure 7A). In leaf sheaths, only 4 upregulated DEGs (CLCc, NPF3.1, NPF7.3-1 and P-Ca2+-ATPase 10) were identified after NaCl treatment for both 6 and 48 h (Figure 7B). In leaf blades, although 9 DEGs were identified after NaCl treatment for both 6 and 48 h, only 3 members (HAK12, CLCf and P-Ca2+-ATPase 7) were upregulated (Figure 7C).
2.5. Identification of DEGs Encoding Transcription Factor in Roots, Leaf Sheaths and Leaf Blades after NaCl Treatment for 6 h
As the expression of transcription factors genes (TFs) changes rapidly in response to abiotic stresses [23], we analyzed differentially expressed TFs in tissues of sweet sorghum after salt treatment for 6 h. As shown in Figure 8A, 216 upregulated TFs and 72 downregulated TFs were identified in roots, and these DEGs were categorized into WRKY, MYB, NAC, bHLH, AP2/ERF, bZIP, MADS-box, HSF, ZF, GRAS families. In leaf sheaths and leaf blades, the number of differentially expressed TFs was less than in roots (Figure 8B,C). Besides, it was noticed that the number of upregulated TFs in all tissues were much more than downregulated TFs (Figure 8).
The venn diagram showed that, among the above differentially expressed TFs, 194, 31 and 71 members were specifically identified in roots, leaf sheaths and leaf blades, respectively (Figure 9A). As 25 differentially expressed TFs were detected in all three tissues (Figure 9A), we then analyzed the expression changes of these TFs, and found that only 2 members (MYB59 and bHLH101) in roots, leaf sheaths and leaf blades were downregulated, while other 23 members in all tissues were upregulated (Figure 9B). Moreover, the log2 ratio of NAC25, bZIP60 and HSF C-2a in roots, leaf sheaths and leaf blades were > 3 (meaning that the transcripts of these genes increased more than 23 folds after NaCl treatment, Figure 9B), which indicated that these TFs might play key roles in the salt tolerance of sweet sorghum.
2.6. Validation of RNA-Seq Results
To verify RNA-seq data, the relative expression levels of 20 randomly selected genes were analyzed by qRT-PCR method. Then, the correlation between RNA-seq results and qRT-PCR results was determined. As shown in Figure S2, the R2 in roots, leaf sheaths and leaf blades after salt treatment for 6 and 48 h, respectively, was more than 0.89, indicating that the RNA-seq data were reliable.
2.7. Expression Pattern of HKT1;5, CLCc and NPF7.3 in Sweet Sorghum under NaCl Treatments
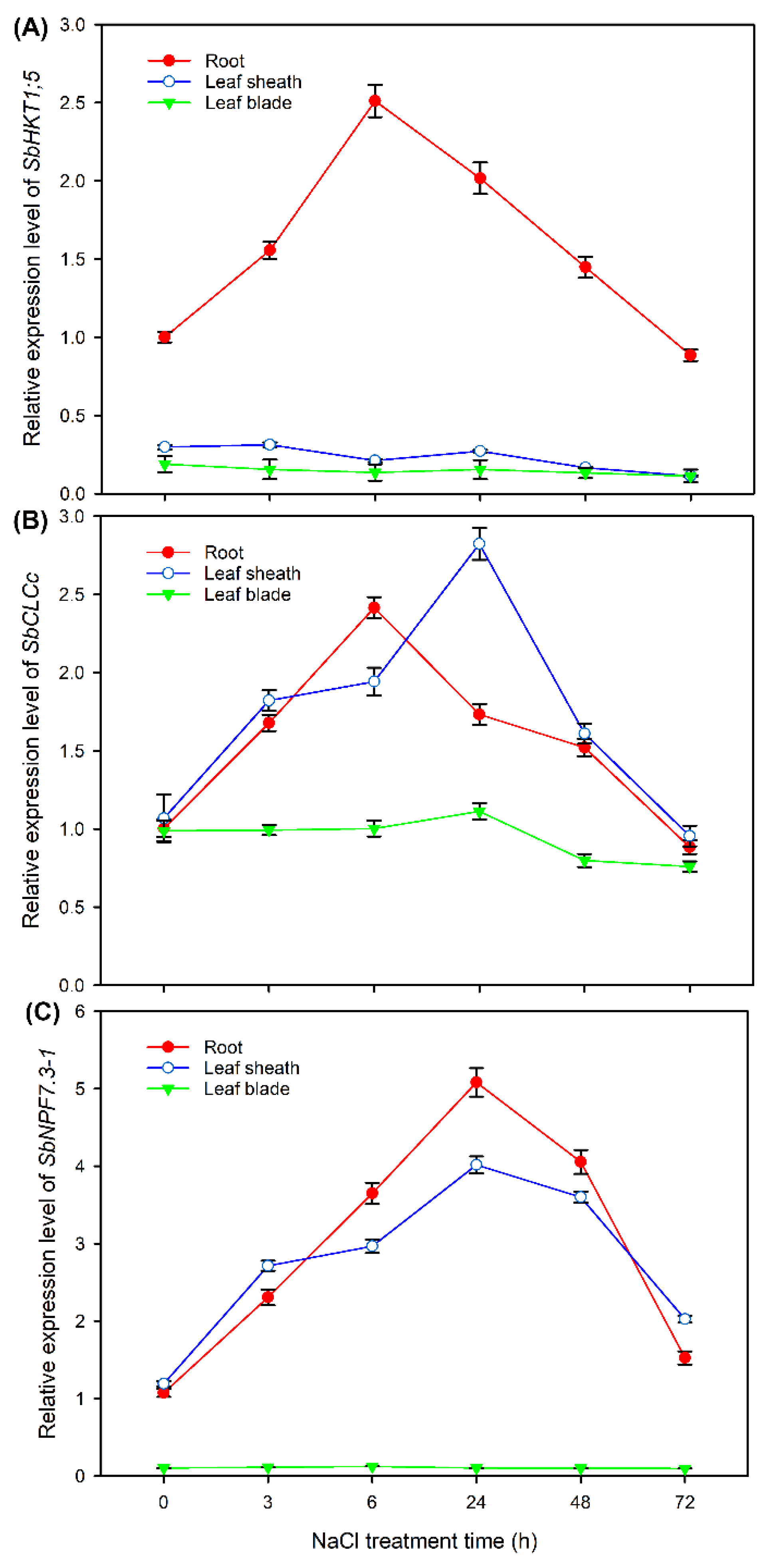
Sweet sorghum showed strong abilities to restrict Na+ accumulation in shoots, reserving large amounts of Cl− in leaf sheaths and maintaining high NO3− content in leaf blades (Figure 2). HKT1;5 has been reported to play a key role in restricting the long-distance transport of Na+ from roots into shoots of rice [14]. CLCc has be thought to facilitate Cl− accumulation in plant tissues by mediating vacuolar Cl− compartmentalization [24], and NPF7.3 (also named as NRT1.5) help to the transport of NO3− into shoots by mediating the efflux of NO3− from root stele to xylem sap [25]. As transcriptome data showed that the expression of HKT1;5 in roots of sweet sorghum was upregulated after NaCl treatment for 6 h (Table S5), and the expression of CLCc and NPF7.3-1 in roots and leaf sheaths of sweet sorghum was also upregulated after salt treatment for both 6 and 48 h (Figure 7), we analyzed in detail the expression pattern of these three genes in response to 200 mM NaCl treatment using qRT-PCR method.
As shown in Figure 10A, under normal conditions (NaCl treatment for 0 h), SbHKT1;5 was mainly expressed in roots; moreover, its relative expression level in roots was increased after NaCl treatment for 3 and 6 h, and then gradually decreased with the prolonged treatment time. Although SbCLCc showed no tissue specific expression under normal conditions, its expression was induced only in roots and leaf sheaths by NaCl treatment for 3-48 h (Figure 10B). SbNPF7.3-2 dominantly expressed in roots and leaf sheaths; it was noticed that the expression of this gene in roots and leaf sheaths was sharply increased (more than 3 folds) after NaCl treatment for 6-48 h (Figure 10C).
3. Discussion
3.1. Sweet Sorghum Could Efficiently Exclude Na+ from Shoots and Accumulate Cl− in Leaf Sheaths under NaCl Stress
The ability to maintain a low Na+ content in shoots or leaves, which is termed Na+ exclusion trait, is vital for the salt tolerance of plant species in Poaceae [4,26]. In this study, Na+ content in shoots, especially in leaf blades, was much lower than that in roots of sweet sorghum under 50-200 mM NaCl treatments (Figure 3A). Differently, A study on grain sorghum cultivars has shown that Na+ content in leave blades is close to or even higher than in roots under 200 mM NaCl treatment [27]. The Na+ exclusion trait is mainly achieved by the retrieval of Na+ from root xylem sap to restrict Na+ transport from roots into shoots [14,15]. Therefore, in comparison with grain sorghum, sweet sorghum possesses a stronger ability to restrict the long-distance transport of Na+ under salt stress. It has been reported that, when Na+ is translocated into shoots of durum wheat, leaf sheath could accumulate the majority of Na+ to decrease Na+ content in leaf blades [20]. Our results also showed that Na+ content in leaf sheaths was clearly higher than that in leaf blades of sweet sorghum under NaCl treatments (Figure 3A), suggesting that the retention of Na+ in leaf sheaths should be also important for sweet sorghum coping with Na+ toxicity to leaf blades.
Studies have found that Cl− content in shoots of wheat, rice and grain sorghum is higher than that in roots under NaCl stress [16,17,27], suggesting that Poaceae plants should not evolve Cl− exclusion trait from shoots. However, the mechanisms of plants in this family coping with Cl−-toxicity are still elusive so far. Wei et al. [21] found that sweet sorghum could also accumulate much more Cl− in shoots than in roots under salt stress. Similarly, in this study, Cl− content in leaf sheaths and leaf blades of sweet sorghum was clearly higher than that in roots under NaCl treatments (Figure 3C). These results suggest that sweet sorghum could transport the majority of Cl− into shoots under saline conditions. As the equilibrium potential of cell membranes is negative [28], the accumulation of Cl− in shoots might help to balance the positive charge of Na+ for the maintenance of membrane stability. It was observed that Cl− content in leaf sheaths was nearly 2-fold higher than that in leaf blades under NaCl treatments (Figure 3C), indicating that leaf sheath of sweet sorghum also serves an indispensable “Cl− reservoir” to avoid Cl− overaccumulation in leaf blades under salt stresses. Therefore, the large accumulation of Cl− in leaf sheaths should be a key process of sweet sorghum alleviating Cl−-toxicity.
K+ and NO3− are essential macronutrients for plant growths and both act as inorganic osmotica, therefore, the maintenance of K+ and NO3− homeostasis is vital for plant adaptations to saline environments [1,29]. However, as Na+ and Cl− would compete for biding sites of K+ and NO3− transporters or channels, the uptake and accumulation of K+ and NO3− in most glycophytes are severely inhibited under salt stress because [30,31]. In this study, K+ content in leaf blades was maintained relatively stable under 50 and 100 NaCl treatments, while significantly declined under 200 mM NaCl (Figure 3B), suggesting that sweet sorghum could maintain K+ homeostasis in leaf blades under low and moderate salt stresses. Interestingly, NO3− content in leaf blades under 50-200 mM NaCl treatments was the same as that under the control condition (Figure 3D), indicative of a prominent ability for maintaining NO3− homeostasis in leaf blades of sweet sorghum under severe salt stresses.
3.2. The Genes Related to Ion Transport Play Key Roles in the Salt Tolerance of Sweet Sorghum
Plants relieves Na+ and Cl− toxicity by excluding them from roots, restricting the transport of them into shoots, or sequestering them in vacuoles of photosynthetic organs, and these processes are dominated by ion transporters and channels [12,18]. In this study, we identified many DEGs related to Na+ and Cl− transport in different tissues of sweet sorghum after 200 mM NaCl treatment by transcriptome sequencing. It was obvious that the number of DEGs in roots was more than that in leaf sheaths and leaf blades (Figure 4, Figure 5 and Figure 6), suggesting that the root primarily controls ion transport under salt stresses. HKT1;5 and HAK4 are thought to be involved in restricting long-distance transport of Na+ from roots into shoots in rice and maize by mediating the retrieval of Na+ from root xylem sap [14,15]. In our transcriptome data, no expression change of HAK4 in sweet sorghum was found, however, the expression of HKT1;5 in roots was upregulated after salt treatment for 6 h (Table S5). Moreover, qRT-PCR results showed that the relative expression level of HKT1;5 in roots was substantially increased under NaCl treatment for 3-24 h (Figure 10A). Taken together, HKT1;5 should play a key role in the Na+ exclusion trait of sweet sorghum.
Our physiological results showed that the leaf sheath of sweet sorghum can accumulate large amount of Na+ and Cl− to restrict their transport into leaf blades (Figure 3). It has been reported that HKT1;4 functions in the retention of Na+ in leaf sheaths of durum wheat under saline conditions [32]. Interestingly, our transcriptome data showed that the transcript of HKT1;4 was only detective in leaf sheaths of sweet sorghum, and moreover, its expression was upregulated after NaCl treatment for 6 h (Table S7), suggesting that HKT1;4 should also play a key role in Na+ accumulation in leaf sheaths of sweet sorghum. The vacuolar sequestration of Cl− mediated by chloride channel CLC dominates the accumulation of Cl− in plant tissues [24,33]. In our transcriptome data, the expression of CLCc in leaf sheaths of sweet sorghum was upregulated after NaCl treatment for both 6 and 48 h (Figure 7A). Meanwhile, the relative expression levels of CLCc in roots and leaf sheaths showed an increase trend under NaCl treatment for 3-48 h (Figure 10B), indicating that CLCc should play a vital role in the accumulation of Cl− in roots and leaf sheaths of sweet sorghum under saline conditions.
The sequestration of Na+ and Cl− in vacuoles of photosynthetic organs is essential for the salt tolerance of plants [34,35]. The tonoplast-located NHX (e.g. NHX1 and NHX2) are key proteins mediating the transport of Na+ into vacuoles [36,37]. However, in this study, the expression of NHX1 and NHX2 in leaf blades of sweet sorghum was downregulated after NaCl treatment for 6 and 24 h (Table S9 and S10), suggesting that there might be other molecular components involved in vacuolar sequestration of Na+ in leaf blades of sweet sorghum. In Arabidopsis and Pugionium cornutum, CLCg is proven to mediate vacuolar Cl− sequestration in shoots [33,35]. In our transcriptome data, two transcripts of CLCg (named SbCLCg-1 and SbCLCg-2) were identified in sweet sorghum, and the expression of SbCLCg-1 was upregulated, while the expression of SbCLCg-2 was downregulated in leaf blades after NaCl treatment for 48 h (Table S10). Therefore, SbCLCg-1 should be an indispensable transporter mediating the sequestration of Cl− in cell vacuole of leaf blades in sweet sorghum under salt stresses.
Sweet sorghum possesses a prominent ability to maintain NO3− homeostasis in leaf blades under salt stresses (Figure 3D). In the model plant Arabidopsis, NPF7.3 is thought to mediate NO3− loading into root xylem and therefore, involved in the long-distance transport NO3− from roots into shoots [38]. However, the expression of NRT1.5 in roots of Arabidopsis is suppressed by salt stress [25]. Differently, in our transcriptome data, 2 transcripts of NPF7.3 (named NPF7.3-1 and NPF7.3-2) were identified, and the expressions of both genes in roots were upregulated after NaCl treatment for both 6 and 48 h (Figure 7A), suggesting that sweet sorghum could enhance the translocation of NO3− into shoots by upregulating the expression of NPF7.3-1 and NPF7.3-2 in roots. Moreover, the expression of NPF7.3-1 in leaf sheaths was also upregulated after salt treatment for both 6 and 48 h (Figure 7B), and qRT-PCR results verified that its expression level in leaf sheaths sharply increased under NaCl treatment for 3-48 h (Figure 10C). Given NPF7.3 mediates NO3− efflux at the cellular level [38], we speculate that, when NO3− is transported into shoots of sweet sorghum, NPF7.3-2 functions in the efflux of NO3− from leaf sheath cells, and thus helping to the transport of NO3− into leaf blades.
In addition, our results also found that the expression of several H+-ATPase and Ca2+-ATPase was upregulated in sweet sorghum after NaCl treatments (Figure 4, Figure 5 and Figure 6), suggesting that these ATPase should provide H+ and Ca2+ pumps for the transmembrane transport of ions such as Na+, K+, Cl− and NO3− and therefore, are also involved in the maintenance of ion homeostasis of sweet sorghum under salt stresses.
3.3. Identification of Key Transcriptome Factors Involved in the Salt Tolerance of Sweet Sorghum
TFs are key regulatory genes involved in plant adaptations to environmental stresses [39]. Recent studies have reported the importance of TFs in the salt tolerance of sweet sorghum, for example, SbWRKY50 could drive the expression of SOS1 and HKT1 to regulate ion homeostasis, and SbbHLH85 enhances Na+ absorption by roots [22,40]. In this study, to identify key TFs regulating salt tolerance of sweet sorghum, we analyzed the differentially expressed TFs after NaCl treatment for 6 h by transcriptome sequencing. Our results identified hundreds of TFs in roots, leaf sheaths and leaf blades of sweet sorghum, and the majority were upregulated after salt treatment (Figure 8). Furthermore, 25 differentially expressed TFs were detective in all tissues, among which the expression of NAC25, bZIP60 and HSF C-2a were upregulated more than 8-fold in roots, leaf sheaths and leaf blades (Figure 9). Therefore, these three TFs might play essential regulatory roles in the adaptation of sweet sorghum to salt stresses. Moreover, in the present study, we identified 31 differentially expressed TFs exclusively in leaf sheaths of sweet sorghum after NaCl treatment (Figure 9A). The further study on these TFs is likely to elucidate the function of leaf sheath in the salt tolerance of sweet sorghum.
3.4. Sweet Sorghum Possesses a Strong Photosynthetic Ability under Salt Stresses
Photosynthesis is a vital process of primary metabolism, provides a large extent of energy and carbohydrates for plant growth and development [41]. However, photosynthesis of most plant species is generally inhibited under saline conditions as a consequence of lessened CO2 availability due to stomatal closure, disturbed chloroplast light energy capture, hindered photosynthetic electron flow and carbon assimilation capacity [42]. Differently, it has been reported that the photosynthesis rate, stomatal pore size and PSII photochemical efficiency of a salt-tolerant sweet sorghum cultivar are all maintained at high levels under NaCl treatments [43]. In the present study, it was found that sweet sorghum cultivar “Lvjuren” showed a high salt tolerance as its growth was unaffected by 50 and 100 mM NaCl treatments (Figure 1). Furthermore, the net photosynthesis rate and stomatal conductance of “Lvjuren” under 50 and 100 mM NaCl treatments were maintained stable, and chlorophyll b content was unaffected when external NaCl concentration was up to 200 mM (Figure 2). All these results suggested that sweet sorghum possesses a strong photosynthetic ability under saline environments.
The cultivation of sweet sorghum in large-scale salinized areas is thought to be a promising approach to ensure food security and promote ecological restoration [2,3]. For this purpose, the strong photosynthetic ability of sweet sorghum under saline environments could (i) provide large amounts of resources (leaves and stalks) for producing of silage and hay; (ii) develop roots and shoots for sand fixation and soil reservation and, (iii) accumulate sugars for energy production. Researchers have analyzed the expression changes of genes involved in photosynthetic processes and sugar biosynthesis under NaCl treatments using transcriptome sequencing [43]. Therefore, elucidating the mechanisms of sweet sorghum maintaining photosynthesis would provide an important theoretical basis for the cultivation of this species in marginal lands.
4. Materials and methods
4.1. Plant Material and Growth Conditions
Seeds of “Lvjuren”, a sweet sorghum cultivar in China, were obtained from Beijing Best Grass Industry Co., Ltd. (Beijing, China). The seeds were sterilized in 75% ethanol and then sown in plastic pots (pot size: 10 cm in height, 10 cm in bottom diameter and 12 cm in top diameter, 4-5 seeds per pot) filled with coarse silica sand with a particle diameter of about 0.5 cm, and irrigated with modified Hoagland solution consisting of 5 mM KNO3, 1 mM KH2PO4, 1 mM MgSO4, 1 mM Ca(NO3)2, 60 μM Fe-citrate, 50 μM H3BO3, 10 μM MnCl2, 1.6 μM ZnSO4, 0.6 μM CuSO4, and 0.05 μM Na2MoO4. The pH of the culture solution was adjusted to 5.7. The growth conditions were as follows: a constant temperature of 28°C, 16 h light period with the light flux density of approximately 500 mmol m–2 s–1, and relative humidity of approximately 60%-80%.
After 3 weeks, the seedlings were thinned out to one uniform plant in each pot. Then, seedlings were exposed to Hoagland solution supplemented with 0 (control), 50, 100 and 200 mM NaCl. 100 and 200 mM NaCl treatments were increased by 50 mM each day until final concentrations were achieved to avoid salt shock. All treatment solutions were changed every 2 d to maintain a constant NaCl concentration. After 10 d, seedlings were harvested for the measurement of physiological parameters. Six replicate seedlings were used for all measurements (n = 6).
4.2. Determination of Plant Height, Tissue Biomass and Shoot Water Content
The plant height (PH) of individual seedlings was measured first; then the root and shoot were separated, and fresh weight (FW) was measured. All samples were finally oven-dried at 80°C for 3 d to measure dry weight (DW). The shoot water content was calculated as (FW-DW)/DW [29].
4.3. Measurements of Photosynthesis-related Parameters
The leaf net photosynthesis rate (Pn) and stomatal conductance (Gs) were measured with LI-6400 photosynthetic measuring apparatus (LI-COR Biosciences, Lincoln, NE, USA) according to Ma et al. [44]. The chlorophyll in leaf samples was extracted with 80% acetone and 95% ethanol (1:1, v/v) in the dark for 24 h, after centrifugation, the absorbances at 645 nm and 663 nm of supernatant were measured using a UV spectrophotometer (UV-2102C, Unico Instrument Co., Ltd, Shanghai, China). The chlorophyll a and b contents were calculated according to Inskeep and Bloom [45].
4.4. Measurement of Ion Contents in Tissues
The root, leaf sheath and leaf blade samples of seedings were first put in 120°C oven for 20 min, then thoroughly dried at 80°C. After that, samples were incubated in 100 mM acetic acid at 90°C for 2 h, then Na+ and K+ contents in tissues were determined using a flame spectrophotometer (Model 410 Flame; Sherwood Scientific, Ltd., Cambridge, UK) according to Wang et al. [46]. Samples were also incubated with deionized water at 100°C for 2 h, then tissue Cl− content was determined using a chloride analyzer (Model 926, Sherwood Scientific Ltd., Cambridge, UK) [33], and tissue NO3− content was determined by the colorimetric method as described by Drechsler et al. [47].
4.5. Transcriptome Sequencing
For transcriptome sequencing, the seedlings of sweet sorghum cultivar “Lvjuren” were cultured as described in Section 2.1. After 3 weeks, uniform seedlings were divided into the control (C) group and salt treatment (S) group. In C group, seedlings were irrigated with Hoagland solution for 6 and 48 h, then root (R), leaf sheath (LS) and leaf blade (LB) samples were collected and labeled as C6R, C6LS, C6LB, C48R, C48LS and C48LB, respectively. In S group, seedlings were treatment with 200 mM NaCl for 6 and 48 h, then root, leaf sheath and leaf blade samples were collected and labeled as S6R, S6LS, S6LB, S48R, S48LS and S48LB, respectively. Each sample had three biological replicates (n = 3).
Total RNA was extracted from above samples, and then converted into mRNA sequencing libraries using Illumina HiSeq platform (Biomarker Technologies Co., Ltd., Beijing, China). Therefore, a total of 36 independent libraries were sequenced. The raw reads were filtered by removal of the adaptor sequences and low-quality sequence reads to obtain high-quality clean reads. Subsequently, the clean reads were mapped to the Sorghum bicolor reference genome sequence (NCBI accession number: GCF_000003195.3, https://www.ncbi.nlm.nih.gov/genome/?term=sorghum). The reads with a perfect match or one mismatch were further analyzed and annotated based on the reference genome. Finally, the gene function was annotated by aligning against protein databases including Nr; Nt; Pfam; KOG/COG; Swiss-Prot; KO and GO.
4.6. Differentially Expressed Genes Analysis
Gene expression levels in each library were determined by the fragments per kilobase of transcript per million fragments mapped (FPKM) method as described by Mortazavi et al. [48]. Differential expression analysis of each treatment group against the corresponding control group (e.g. C6R-1, C6R-2, C6R-3 vs S6R-1, S6R-2, S6R-3) was performed using the DESeq2 software [49]. The results were corrected for multiple tests, and the false discovery rate (FDR) was used to determine the threshold p-value in multiple tests (Ma et al., 2016). In this study, a gene with FDR<0.001 and absolute value of log2(FPKMtreated/FPKMcontrol)>1 was termed as differentially expressed gene (DEG). Finally, we analyzed DEGs related to Na+, K+, Cl− and NO3− transport after salt treatment for 6 and 48 h, and screened DEGs encoding transcriptional factors after salt treatment for 6 h.
4.7. Validation of RNA-sequencing Results
To confirm the reliability of RNA sequencing (RNA-seq) results, we randomly selected 20 genes in transcriptome data, and determined the relative expression levels of these genes using qRT-PCR method with a StepOnePlus Real-Time PCR Thermocycler (ABI PRISM 7500, USA) [33]. Sweet sorghum internal reference gene SbActin1 was used as a control. The gene specific primers for qRT-PCR were listed in Table S1. Finally, the correlation analysis between RNA-seq and qRT-PCR results was performed.
4.8. Analysis of Expression Pattern of HKT1;5, CLCc and NPF7.3 in Sweet Sorghum under NaCl Treatment
From our transcriptome data, we selected three key genes HKT1;5, CLCc and NPF7.3-2 to investigate their expression pattern under NaCl treatment. 3-week-old sweet sorghum seedlings were treated with 200 mM NaCl for 0, 3, 6, 24, 48 and 72 h, respectively. Then, the root, leaf sheath and leaf blade samples were harvested, the total RNA in these samples was extracted and the first-strand cDNA was synthesized using the PrimeScript™ RT Master Mix (Perfect Real Time) kit (TaKaRa, Biotech Co., Ltd., Dalian, China). The relative expression levels of these genes in above samples were determined by qRT-PCR method. The primers for HKT1;5, CLCc and NPF7.3-2 were listed in Table S1.
4.9. Data Analysis
Six replicate seedlings were used for all physiological parameter measurements (n = 6), three replicate seedlings were used for transcriptome sequencing and qRT-PCR analysis (n = 3). The results for the physiological parameters and qRT-PCR are all presented as the mean with standard deviation (SD). The data were subjected to one-way analysis of variance (ANOVA) using SPSS 19.0 statistical software (SPSS Inc., Chicago, IL, USA) followed by Tukey’s HSD to detect significant differences between means at a significance level of P < 0.05.
5. Conclusions
In conclusion, our results demonstrated that sweet sorghum is a typical Na+ exclusion plant species that can maintain a low Na+ content in shoots under salt stress. Although sweet sorghum cannot restrict Cl− translocation into shoots, it decreases Cl− toxicity to leaf blades by the large accumulating of Cl− in leaf sheaths. Furthermore, sweet sorghum shows a prominent ability for maintaining NO3− homeostasis in leaf blades under NaCl treatments. Transcriptome sequencing identified many key genes involved in Na+, K+, Cl− and NO3− transport in roots, leaf sheaths and leaf blades of sweet sorghum after NaCl treatment. Furthermore, the increased expression of HKT1;5, CLCc and NPF7.3-1 after salt treatment is conducive to retention of Na+ in roots, accumulation of Cl− in leaf sheath and maintaining a high NO3− content in leaf blades, respectively. In addition, many TFs play essential regulatory roles in the salt tolerance of sweet sorghum. These results provide theoretical basis for the large-scale cultivation of sweet sorghum in salinized areas.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, Huan Guo and Yan-Nong Cui; Data curation, Huan Guo, Chun-Ya Nie and Zhen Li; Methodology, Huan Guo, Chun-Ya Nie, Zhen Li, Jie Kang and Xiao-Long Wang; Software, Chun-Ya Nie and Zhen Li; Supervision, Yan-Nong Cui; Writing – original draft, Huan Guo; Writing – review & editing, Yan-Nong Cui.
Funding
This research was financially supported by the Natural Science Basic Research Program of Shaanxi Province (2023-JC-QN-0255), National Natural Science Foundation of China (32101253), China Postdoctoral Science Foundation (2022M712610) and Scientific Startup Foundation for Doctors of Northwest A and F University (2452021106).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare that they have no conflict of interest to this work.
Abbreviations
NHX: sodium/hydrogen exchanger; CHX: Cation/H+ antiporter; CCX: Cation/calcium exchanger; NCX: Sodium/calcium exchanger; CNGC: cyclic nucleotide-gated ion channel; HKT: high affinity K+ transporter; KEA: K+ efflux antiporter; KT/KUP/HAK: potassium transporter; AKT: inward rectifying K+ channel; KOR: outward rectifying K+ channel; CLC: chloride channel; SLAH: slow-type anion channel associated homolog; ALMT: aluminium activated malate transporter; NPF: nitrate transporter1/peptide transporter; CCC: cation chloride cotransporter; WRKY: WRKY DNA-binding domain transcription factor; MYB: MYB-like DNA-binding transcription factor; NAC: NAC domain-containing transcription factor; bHLH: helix-loop-helix DNA-binding domain transcription factor; AP2: AP2 domain transcription factor; ERF: Ethylene-responsive transcription factor; bZIP: basic leucine-zipper transcription factor; MADS-box: MADS-box transcription factor; HSF: Heat stress transcription factor; ZF: Zinc finger; GRAS: GRAS domain transcription factor; Nr: NCBI non-redundant protein sequences; Nt: NCBI non-redundant nucleotide sequences; Pfam: Protein family; KOG/COG: Clusters of Orthologous Groups of proteins; Swiss-Prot: A manually annotated and reviewed protein sequence database; KO: KEGG Ortholog database; GO: Gene Ontology.
References
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yu, F.; Xie, P.; Sun, S, Qiao, X.; Tang, S.; Chen, C.; Yang, S.; Mei, C.; Yang, D.; Wu, Y.; Xia, R.; Li, X.; Lu, J.; Liu, Y.; Xie, X.; Ma, D.; Xu, X.; Liang, Z.; Feng, Z.; Huang, X.; Yu, H.; Liu, G.; Wang, Y.; Li, J.; Zhang, Q.; Chen, C.; Ouyang, Y.; Xie, Q. A Gγ protein regulates alkaline sensitivity in crops. Science 2023, 379, eade8416. [CrossRef]
- Li, J.; Lei, S.; Gong, H.; Liu, Z.; Zhang, Y.; Ouyang, Z. Field performance of sweet sorghum in salt-affected soils in China: A quantitative synthesis. Environ. Res. 2023, 222, 115362. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; James, R.A.; Xu, B.; Athman, A.; Conn, S.J.; Jordans, C.; Byrt, C.S.; Hare, R.A.; Tyerman, S.D.; Tester, M.; Plett, D.; Gilliham, M. Wheat grain yield on saline soils is improved by an ancestral Na+ transporter gene. Nat. Biotechnol. 2012, 30, 360–364. [Google Scholar] [CrossRef]
- Zheng, L.Y.; Guo, X.S.; He, B.; Sun, L.J.; Peng, Y.; Dong, S.S.; Liu, T.F.; Jiang, S.; Ramachandran, S.; Liu, C.M.; Jing, H.C. Genome-wide patterns of genetic variation in sweet and grain sorghum (Sorghum bicolor). Genome Biol. 2011, 12, R114. [Google Scholar] [CrossRef] [PubMed]
- Regassa, T.H.; Wortmann, C.S. Sweet sorghum as a bioenergy crop: Literature review. Biomass Bioenerg. 2014, 64, 348–355. [Google Scholar] [CrossRef]
- López-Sandin, I.; Zavala-García, F.; Levin, L.; Ruiz, H.A.; Hernández-Luna, C.E.; Gutiérrez-Soto, G. Evaluation of bioethanol production from sweet sorghum variety roger under different tillage and fertilizer treatments. BioEnerg. Res. 2021, 14, 1058–1069. [Google Scholar] [CrossRef]
- Yang, Z.; Li, J.L.; Liu, L.N.; Xie, Q.; Sui, N. Photosynthetic regulation under salt stress and salt-tolerance mechanism of sweet sorghum. Front. Plant Sci. 2020, 10, 1722. [Google Scholar] [CrossRef]
- Cui, J.; Ren, G.; Qiao, H.; Xiang, X.; Huang, H.; Chang, J. Comparative transcriptome analysis of seedling stage of two sorghum cultivars under salt stress. J. Plant Growth Regul. 2018, 37, 986–998. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Plant salt tolerance: adaptations in halophytes. Ann. Bot. 2015, 115, 327–331. [Google Scholar] [CrossRef]
- Geilfus, C.M. Review on the significance of chlorine for crop yield and quality. Plant Sci. 2018, 270, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Bazihizina, N.; Colmer, T.D.; Cuin, T.A.; Mancuso, S.; Shabala, S. Friend or foe? Chloride patterning in halophytes. Trends Plant Sci. 2019, 24, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, N.I.; Yamaji, N.; Yamamoto, H.; Okubo, K.; Ueno, H.; Costa, A.; Tanoi, K.; Matsumura, H.; Fujii-Kashino, M.; Horiuchi, T.; Nayef, M.A.; Shabala, S.; An, G.; Ma, J.F.; Horie, T. OsHKT1;5 mediates Na+ exclusion in the vasculature to protect leaf blades and reproductive tissues from salt toxicity in rice. Plant J. 2017, 91, 657–670. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Liang, X.; Wang, L.; Cao, Y.; Song, W.; Shi, J.; Lai, J.; Jiang, C. A HAK family Na+ transporter confers natural variation of salt tolerance in maize. Nat. Plants 2019, 5, 1297–1308. [Google Scholar] [CrossRef] [PubMed]
- Kingsbury, R. W.; Epstein, E. Salt sensitivity in wheat, a case for specific ion toxicity. Plant Physiol. 1986, 80, 651–654. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.Q.; Gao, X.H.; Sun, W.; An, J.; Zhao, Y.X.; Zhang, H. Cloning and functional characterization of a cation-chloride cotransporter gene OsCCC1. Plant Mol. Biol. 2011, 75, 567–578. [Google Scholar] [CrossRef]
- Li, B.; Tester, M.; Gilliham, M. Chloride on the move. Trends Plant Sci. 2017, 22, 236–248. [Google Scholar] [CrossRef]
- Louarn, G.; Andrieu, B.; Giauffret, C. A size-mediated effect can compensate for transient chilling stress affecting maize (Zea mays) leaf extension. New Phytol. 2010, 187, 106–118. [Google Scholar] [CrossRef]
- James, R.A.; Davenport, R.J.; Munns, R. Physiological characterization of two genes for Na+ exclusion in durum wheat, Nax1 and Nax2. Plant Physiol. 2016, 142, 1537–1547. [Google Scholar] [CrossRef]
- Wei, X.; Liu, L.; Lu, C.; Yuan, F.; Han, G.; Wang, B. SbCASP4 improves salt exclusion by enhancing the root apoplastic barrier. Planta 2021, 254, 81. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Li, J.; Sui, Y.; Han, G.; Zhang, Y.; Guo, S.; Sui, N. The sweet sorghum SbWRKY50 is negatively involved in salt response by regulating ion homeostasis. Plant Mol. Biol. 2020, 102, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.N.; Wang, F.Z.; Yang, C.H.; Yuan, J.Z.; Guo, H.; Zhang, J.L.; Wang, S.M.; Ma, Q. Transcriptomic profiling identifies candidate genes involved in the salt tolerance of the xerophyte Pugionium cornutum. Genes 2019, 10, 1039. [Google Scholar] [CrossRef] [PubMed]
- Jossier, M.; Kroniewicz, L.; Dalmas, F.; Thiec, D.L.; Ephritikhine, G.; Barbier-Brygoo, H.; Vavasseur, A.; Filleur, S.; Leonhardt, N. The Arabidopsis vacuolar anion transporter, AtCLCc, is involved in the regulation of stomatal movements and contributes to salt tolerance. Plant J. 2010, 64, 563–576. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Z.; Lv, X.F.; Li, J.Y.; Yi, H.Y.; Gong, J.M. Arabidopsis NRT1.5 is another essential component in the regulation of nitrate reallocation and stress tolerance. Plant Physiol. 2012, 159, 1582–1590. [Google Scholar] [CrossRef]
- Horie, T.; Hauser, F.; Schroeder, J.I. HKT transporter-mediated salinity resistance mechanisms in Arabidopsis and monocot crop plants. Trends Plant Sci. 2009, 14, 660–668. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.S.; Zou, Q.; Zhao, K.F. Effect of NaCl stress on ionic contents in different organs of sorghum plants. Acta Agronomica Sinica 2000, 26, 845–850. [Google Scholar]
- Teakle, N.L.; Tyerman, S.D. Mechanisms of Cl– transport contributing to salt tolerance. Plant Cell Environ. 2010, 33, 566–589. [Google Scholar] [CrossRef]
- Cui, Y.N.; Li, X.T.; Yuan, J.Z.; Wang, F.Z.; Guo, H.; Xia, Z.R.; Wang, S.M.; Ma, Q. Chloride is beneficial for growth of the xerophyte Pugionium cornutum by enhancing osmotic adjustment capacity under salt and drought stresses. J. Exp. Bot. 2020, 71, 4215–4231. [Google Scholar] [CrossRef]
- Tyerman, S.D.; Skerrett, I.M. Root ion channels and salinity. Sci. Hortic-Amsterdam 1999, 78, 175–235. [Google Scholar] [CrossRef]
- Reich, M.; Aghajanzadeh, T.; Helm, J.; Parmar, S.; Hawkesford, M.J.; De Kok, L.J. Chloride and sulfate salinity differently affect biomass, mineral nutrient composition and expression of sulfate transport and assimilation genes in Brassica rapa. Plant Soil 2017, 411, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.B.; Spielmeyer, W.; Lagudah, E.S.; James, R.A.; Platten, J.D.; Dennis, E.S.; Munns, R. A sodium transporter (HKT7) is a candidate for Nax1, a gene for salt tolerance in durum wheat. Plant Physiol. 2006, 142, 1718–1727. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.N.; Lin, Z.R.; Cai, M.M.; Liu, R.W.; Wang, S.M.; Ma, Q. PcCLCg is involved in the accumulation of Cl− in shoots for osmotic adjustment and salinity resistance in the Cl−-tolerant xerophyte Pugionium cornutum. Plant Soil. 2023. [Google Scholar] [CrossRef]
- Apse, M.P.; Blumwald, E. Na+ transport in plants. FEBS Lett. 2007, 581, 2247–2254. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.T.; Agorio, A.; Jossier, M.; Depre, S.; Thomine, S.; Filleur, S. Characterization of the chloride channel-like, AtCLCg, involved in chloride tolerance in Arabidopsis thaliana. Plant Cell Physiol. 2016, 57, 764–775. [Google Scholar] [CrossRef]
- Teng, X.X.; Cao, W.L.; Lan, H.X.; Tang, H.J.; Bao, Y.M.; Zhang, H.S. OsNHX1, an Na+/H+ antiporter gene, can enhance salt tolerance in rice plant through more effective accumulation of toxic Na+ in leaf mesophyll and bundle sheath cells. Acta Physiol. Plant. 2007, 39, 113–125. [Google Scholar] [CrossRef]
- Bao, A.K.; Du, B.Q.; Touil, L.; Kang, P.; Wang, Q.L.; Wang, S.M. Co-expression of tonoplast cation/H+ antiporter and H+-pyrophosphatase from xerophyte Zygophyllum xanthoxylum improves alfalfa plant growth under salinity, drought and field conditions. Plant Biotechnol. J. 2016, 14, 964–975. [Google Scholar] [CrossRef]
- Lin, S.H.; Kuo, H.F.; Canivenc, G.; Lin, C.S.; Lepetit, M.; Hsu, P.K.; Tillard, P.; Lin, H.L.; Wang, Y.Y.; Tsai, C.B.; Gojon, A.; Tsay, Y.F. Mutation of the Arabidopsis NRT1.5 nitrate transporter causes defective root-to-shoot nitrate transport. Plant Cell 2008, 20, 2514–2528. [Google Scholar] [CrossRef]
- Wang, H.; Wang, H.; Shao, H.; Tang, X. Recent advances in utilizing transcription factors to improve plant abiotic stress tolerance by transgenic technology. Front. Plant Sci. 2016, 7, 67. [Google Scholar] [CrossRef]
- Song, Y.; Li, S.; Zheng, H.; Han, G.; Sun, X.; Yang, W.; Wang, H.; Zhuang, K.; Kong, F.; Meng, Q.; Sui, N. SbbHLH85, a bHLH member, modulates resilience to salt stress by regulating root hair growth in sweet sorghum. Theor. Appl. Genet. 2022, 135, 201–206. [Google Scholar] [CrossRef]
- Nouri, M.Z.; Moumeni, A.; Komatsu, S. Abiotic stresses: Insight into gene regulation and protein expression in photosynthetic pathways of plants. Int. J. Mol. Sci. 2015, 16, 20392–20416. [Google Scholar] [CrossRef] [PubMed]
- Chaves, M.M.; Costa, J.M.; Zarrouka, O.; Pinheiro, C.; Lopes, C.M.; Pereira, J.S. Controlling stomatal aperture in semi-arid regions: The dilemma of saving water or being cool? Plant Sci. 2016, 251, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Sui, N.; Yang, Z.; Liu, M.; Wang, B. Identification and transcriptomic profiling of genes involved in increasing sugar content during salt stress in sweet sorghum leaves. BMC Genomics 2015, 16, 534. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Yue, L.J.; Zhang, J.L.; Wu, G.Q.; Bao, A.K.; Wang, S.M. Sodium chloride improves photosynthesis and water status in the succulent xerophyte Zygophyllum xanthoxylum. Tree Physiol. 2012, 32, 4–13. [Google Scholar] [CrossRef]
- Inskeep, W.P.; Bloom, P.R. Extinction coefficients of chlorophyll-a and chlorophyll-b in N,N-dimethylformamide and 80-percent acetone. Plant Physiol. 1985, 77, 483–485. [Google Scholar] [CrossRef]
- Wang, S.M.; Zhang, J.L.; Flowers, T.J. Low-affinity Na+ uptake in the halophyte Suaeda maritima. Plant Physiol. 2007, 145, 559–571. [Google Scholar] [CrossRef]
- Drechsler, N.; Zheng, Y.; Bohner, A.; Nobmann, B.; von Wiren, N.; Kunze, R.; Rausch, C. Nitrate-dependent control of shoot K homeostasis by the nitrate transporter1/peptide transporter family member NPF7.3/NRT1.5 and the stelar K+ outward rectifier SKOR in Arabidopsis. Plant Physiol. 2015, 169, 2832–2847. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef]
- De Vega, J.J.; Teshome, A.; Klaas, M.; Grant, J.; Finnan, J.; Barth, S. Physiological and transcriptional response to drought stress among bioenergy grass Miscanthus species. Biotechnol. Biofuels 2021, 14, 60. [Google Scholar] [CrossRef]
Figure 1.
Effects of 50-200 mM NaCl treatments on growth of sweet sorghum cultivar “Lvjuren”. (A) growth photograph, (B) plant height, (C) fresh weight, (D) dry weight, (E) shoot water content. Data are means (±SD), n=6. Different letters indicate significant differences as determined using Tukey’s HSD test (P<0.05).
Figure 1.
Effects of 50-200 mM NaCl treatments on growth of sweet sorghum cultivar “Lvjuren”. (A) growth photograph, (B) plant height, (C) fresh weight, (D) dry weight, (E) shoot water content. Data are means (±SD), n=6. Different letters indicate significant differences as determined using Tukey’s HSD test (P<0.05).
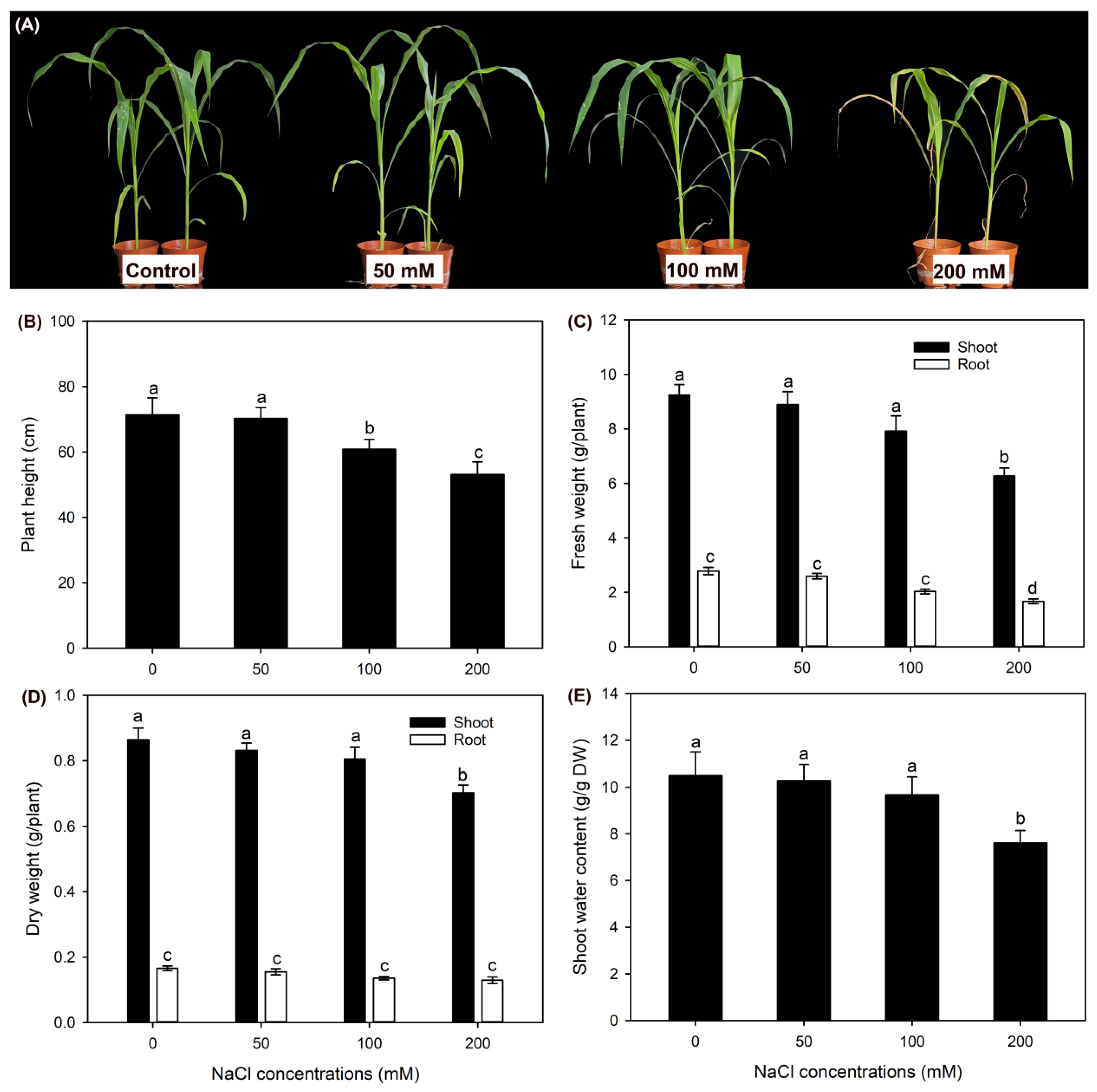
Figure 2.
Effects of 50-200 mM NaCl treatments on photosynthesis of sweet sorghum cultivar “Lvjuren”. (A) net photosynthetic rate, (B) stomatal conductance, (C) chlorophyll a content, (D) chlorophyll b content.
Figure 2.
Effects of 50-200 mM NaCl treatments on photosynthesis of sweet sorghum cultivar “Lvjuren”. (A) net photosynthetic rate, (B) stomatal conductance, (C) chlorophyll a content, (D) chlorophyll b content.
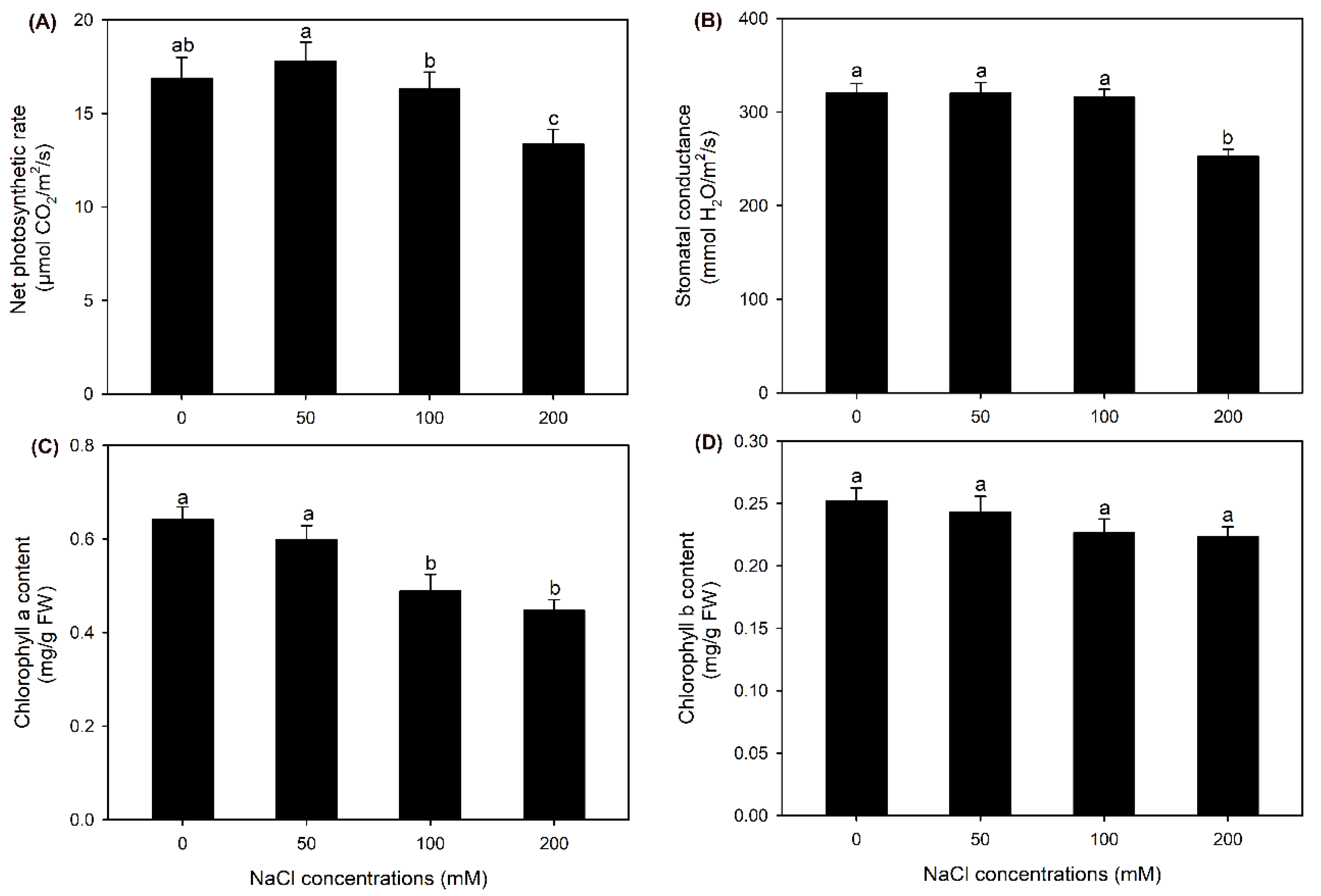
Figure 3.
Effects of 50-200 mM NaCl treatments on tissue Na+ (A), K+ (B), Cl− (C) and NO3− (D) contents of sweet sorghum cultivar “Lvjuren”.
Figure 3.
Effects of 50-200 mM NaCl treatments on tissue Na+ (A), K+ (B), Cl− (C) and NO3− (D) contents of sweet sorghum cultivar “Lvjuren”.
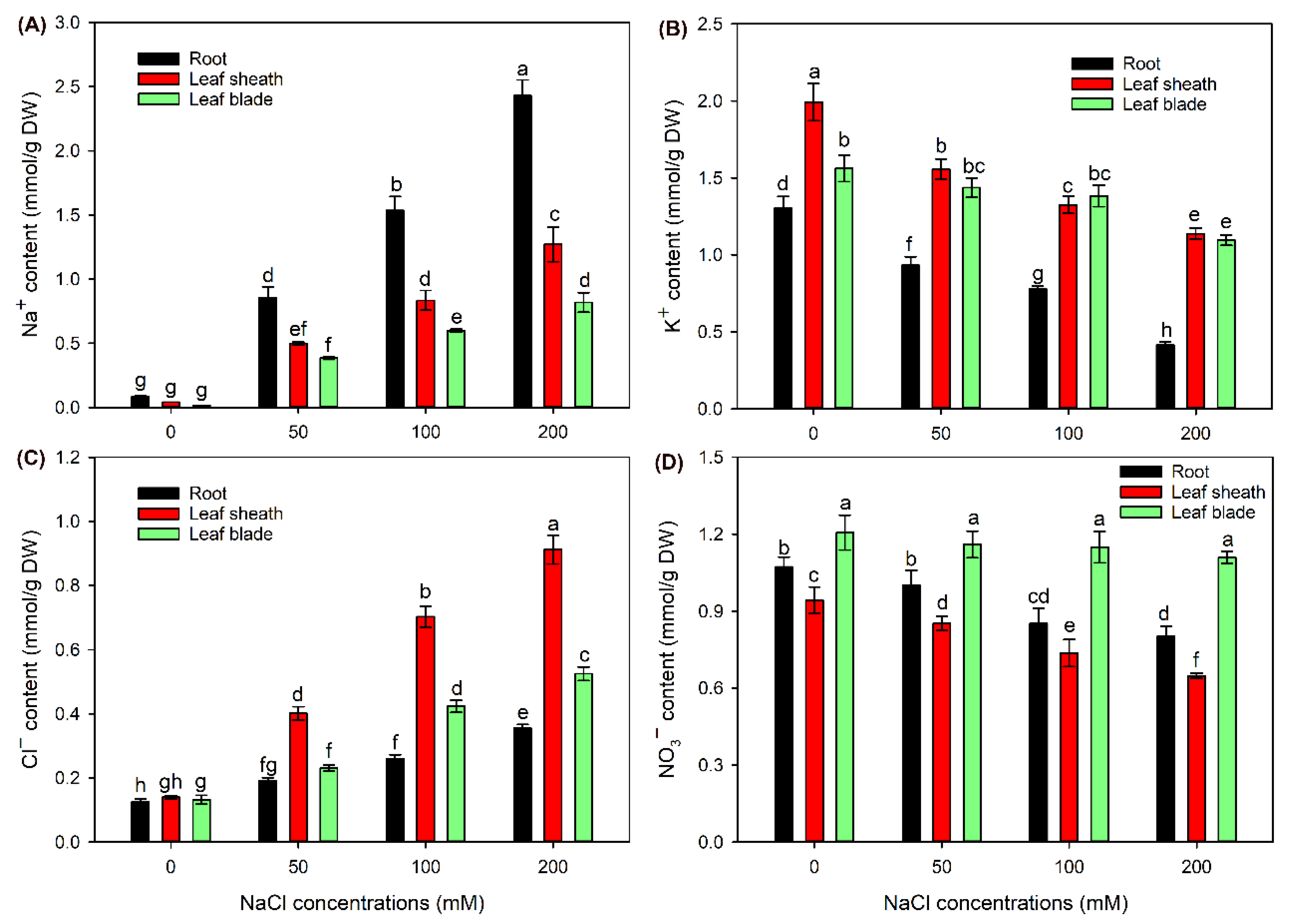
Figure 4.
The number of DEGs related to Na+, K+, Cl− and NO3− transport in roots of sweet sorghum cultivar “Lvjuren” after 200 mM NaCl treatment for 6 (A) and 24 (B) h, respectively. The red upward arrow and green downward arrow show the total number of upregulated DEGs and downregulated DEGs, respectively.
Figure 4.
The number of DEGs related to Na+, K+, Cl− and NO3− transport in roots of sweet sorghum cultivar “Lvjuren” after 200 mM NaCl treatment for 6 (A) and 24 (B) h, respectively. The red upward arrow and green downward arrow show the total number of upregulated DEGs and downregulated DEGs, respectively.
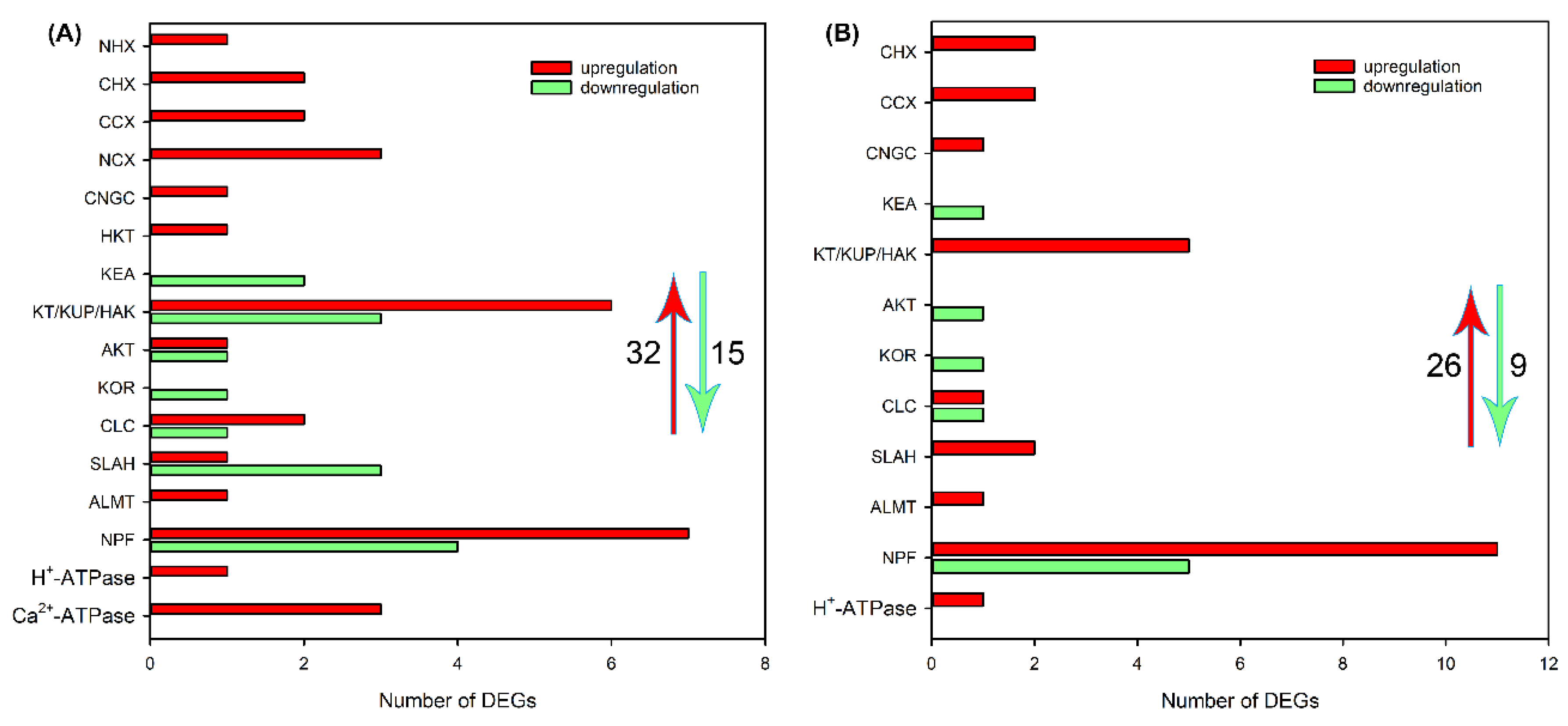
Figure 5.
The number of DEGs related to Na+, K+, Cl− and NO3− transport in leaf sheaths of sweet sorghum cultivar “Lvjuren” after 200 mM NaCl treatment for 6 (A) and 24 (B) h, respectively.
Figure 5.
The number of DEGs related to Na+, K+, Cl− and NO3− transport in leaf sheaths of sweet sorghum cultivar “Lvjuren” after 200 mM NaCl treatment for 6 (A) and 24 (B) h, respectively.
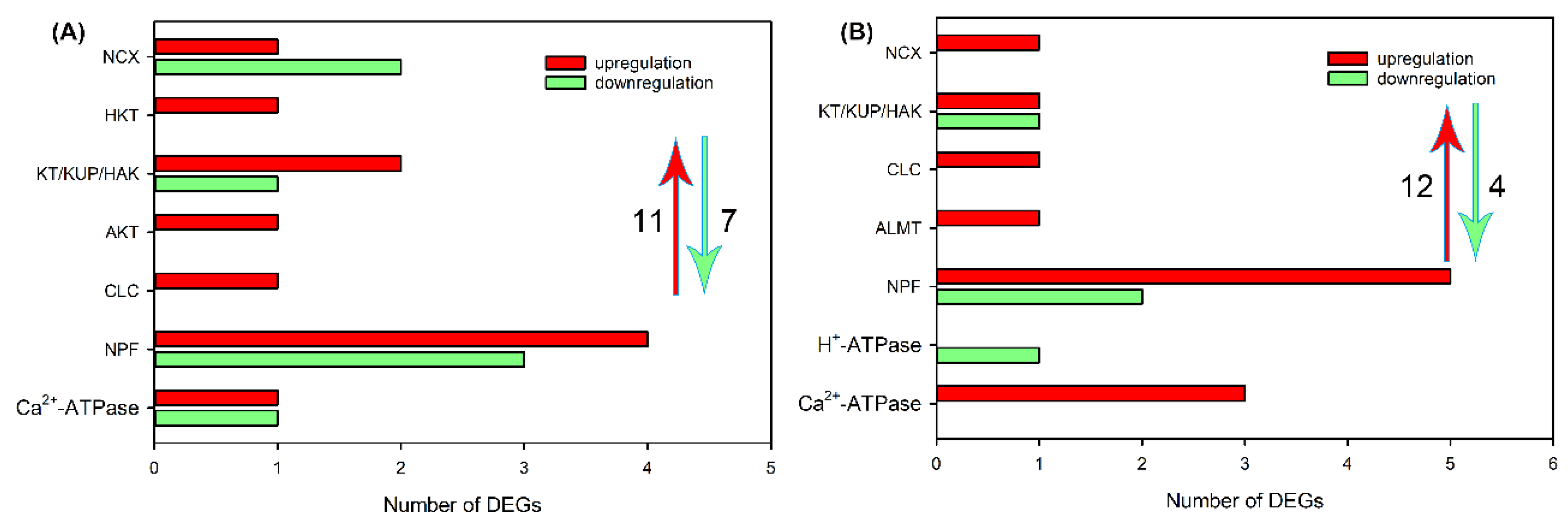
Figure 6.
The number of DEGs related to Na+, K+, Cl− and NO3− transport in leaf blades sweet sorghum cultivar “Lvjuren” after 200 mM NaCl treatment for 6 (A) and 24 (B) h, respectively.
Figure 6.
The number of DEGs related to Na+, K+, Cl− and NO3− transport in leaf blades sweet sorghum cultivar “Lvjuren” after 200 mM NaCl treatment for 6 (A) and 24 (B) h, respectively.
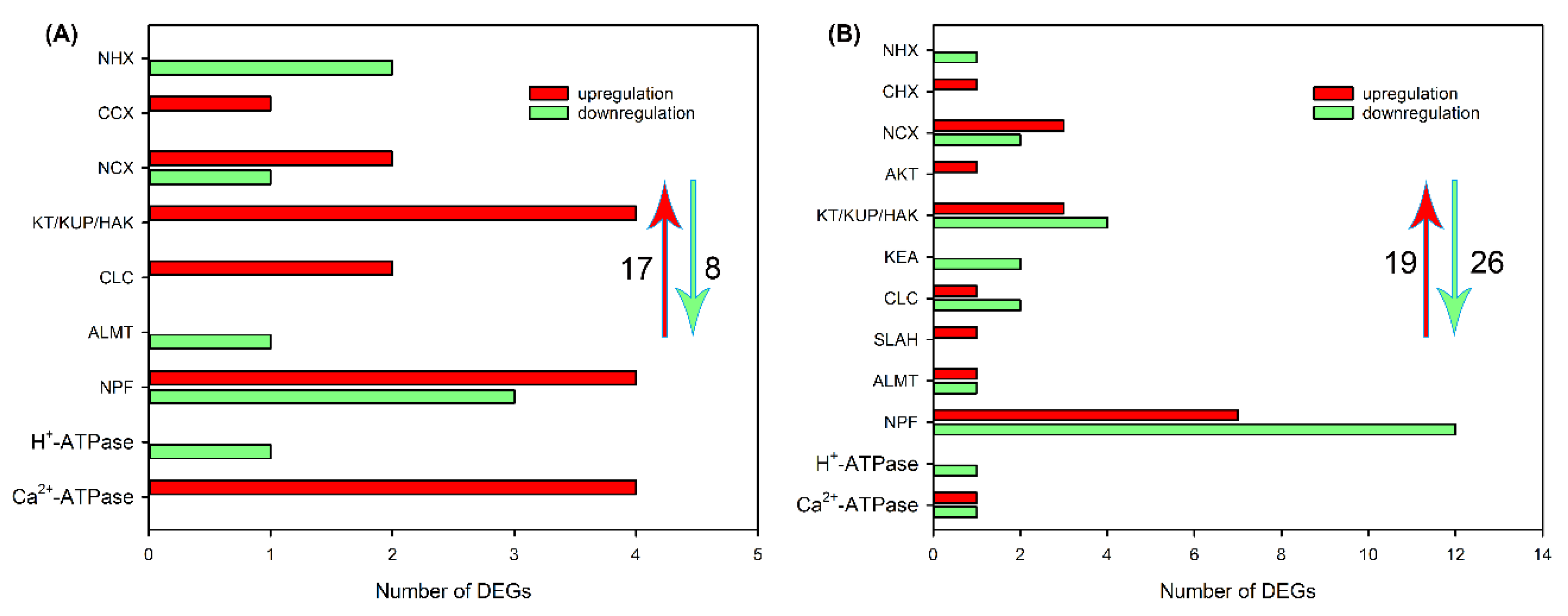
Figure 7.
Heat maps showing the expression changes of DEGs related to ion transport that were detective in roots (A), leaf sheaths (B) and leaf blades (C) after 200 mM NaCl treatment for both 6 and 48 h.
Figure 7.
Heat maps showing the expression changes of DEGs related to ion transport that were detective in roots (A), leaf sheaths (B) and leaf blades (C) after 200 mM NaCl treatment for both 6 and 48 h.
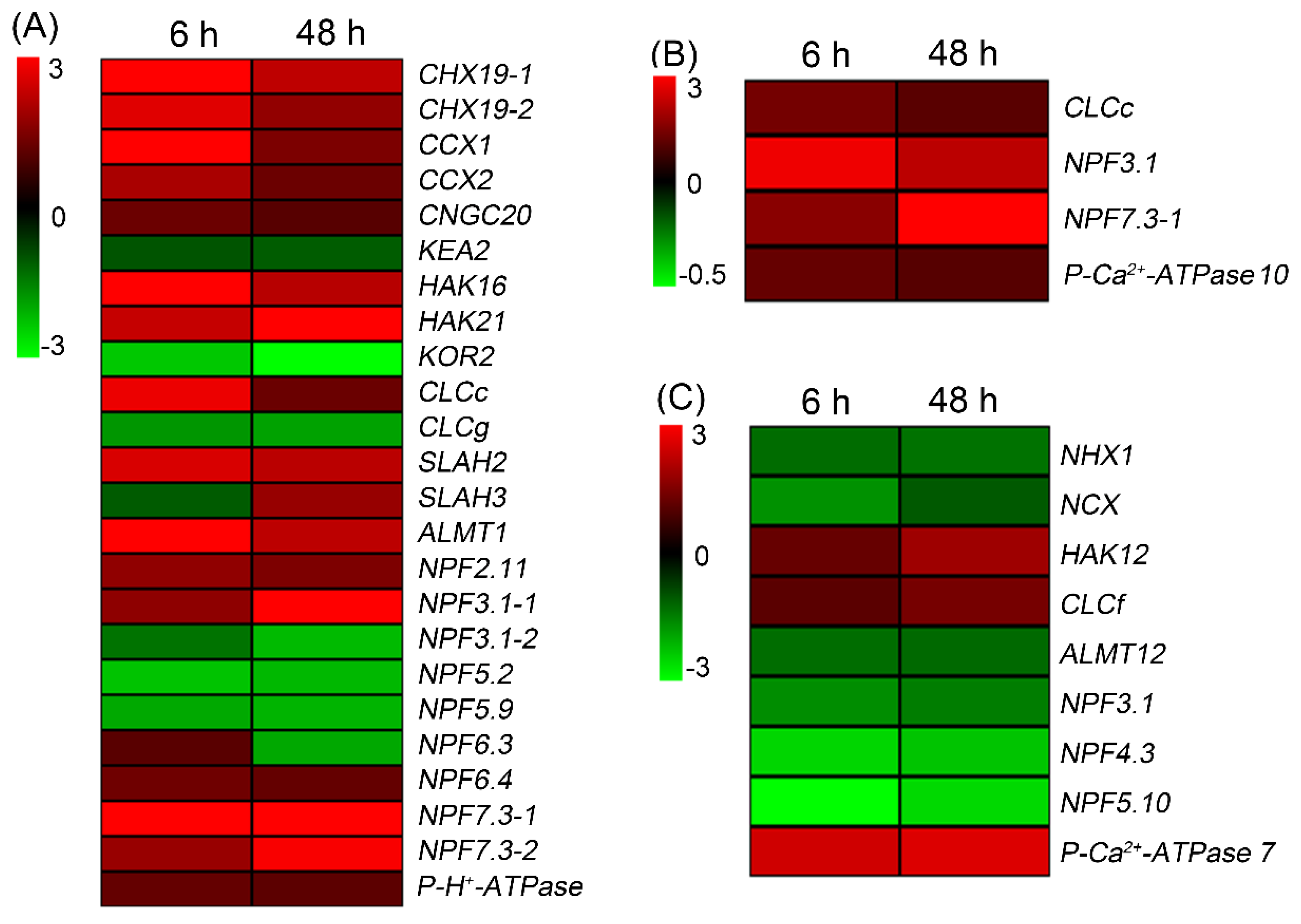
Figure 8.
The number of DEGs encoding transcriptome factor in roots (A), leaf sheaths (B) and leaf blades (C), respectively, after 200 mM NaCl treatment for 6 h.
Figure 8.
The number of DEGs encoding transcriptome factor in roots (A), leaf sheaths (B) and leaf blades (C), respectively, after 200 mM NaCl treatment for 6 h.
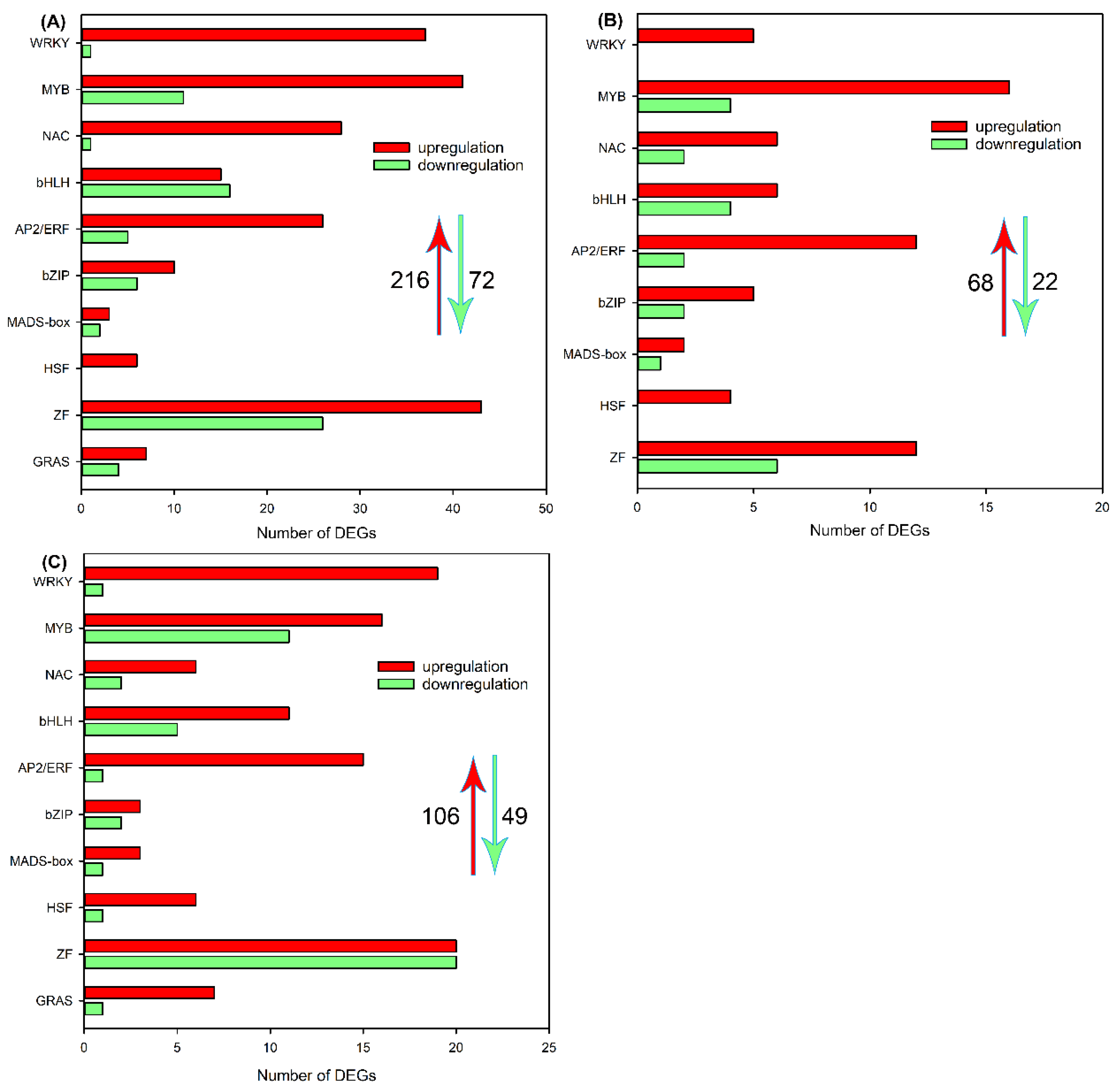
Figure 9.
Venn diagrams showing the number of exclusive and common DEGs encoding transcriptome factors in roots, leaf sheaths and leaf blades (A); heat maps showing the expression changes of DEGs encoding transcriptome factors in all tissues after 200 mM NaCl treatment for 6 h (B).
Figure 9.
Venn diagrams showing the number of exclusive and common DEGs encoding transcriptome factors in roots, leaf sheaths and leaf blades (A); heat maps showing the expression changes of DEGs encoding transcriptome factors in all tissues after 200 mM NaCl treatment for 6 h (B).
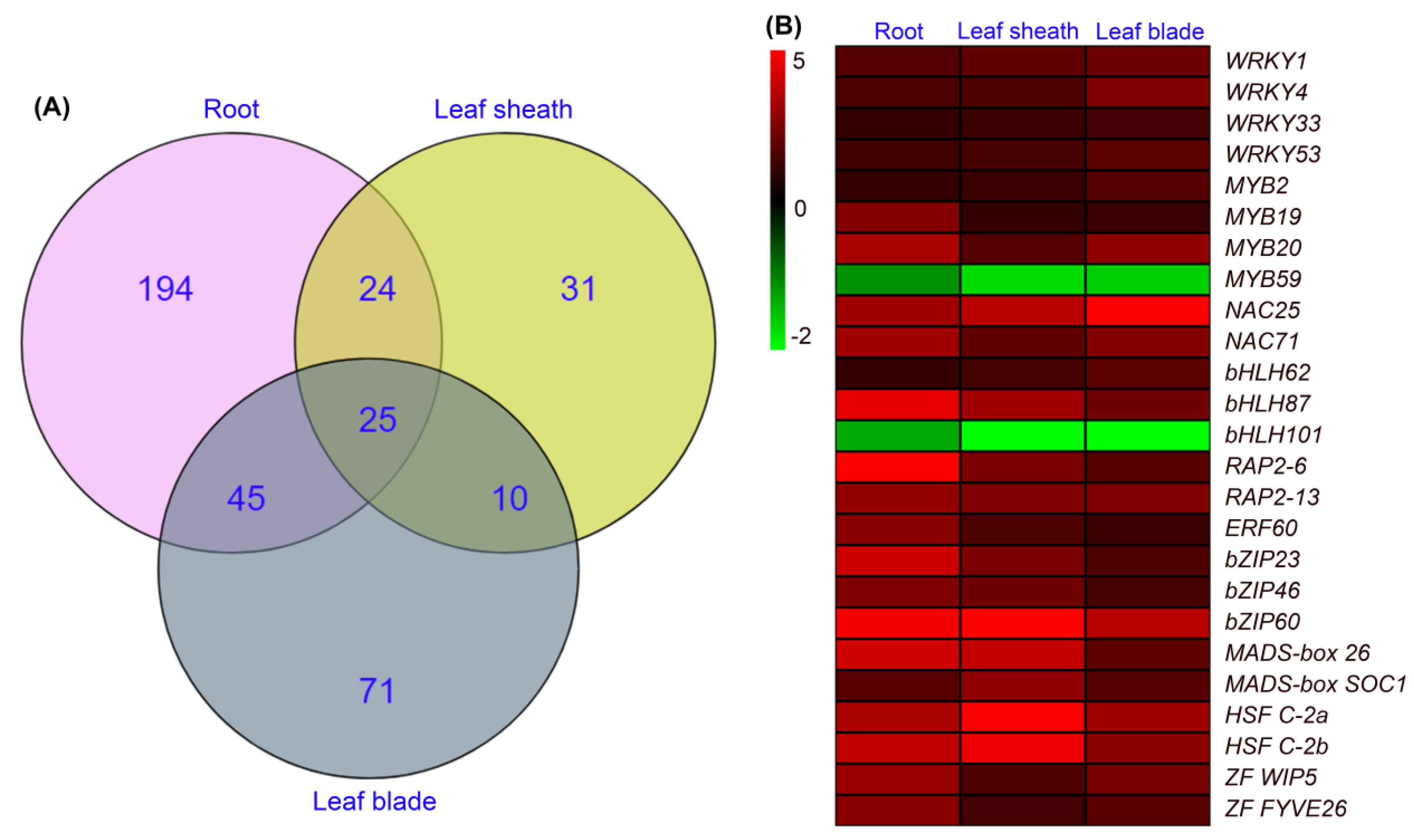
Figure 10.
The relative expression levels of SbHKT1;5 (A), SbCLCc (B) and SbNPF7.3-1 (C) in roots, leaf sheaths and leaf blades of sweet sorghum cultivar “Lvjuren” after 200 mM NaCl treatment for 0, 3, 6, 24, 48 and 72 h., respectively. Data are means (±SD), n=3.
Figure 10.
The relative expression levels of SbHKT1;5 (A), SbCLCc (B) and SbNPF7.3-1 (C) in roots, leaf sheaths and leaf blades of sweet sorghum cultivar “Lvjuren” after 200 mM NaCl treatment for 0, 3, 6, 24, 48 and 72 h., respectively. Data are means (±SD), n=3.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



