
Submitted:
03 June 2023
Posted:
05 June 2023
You are already at the latest version
Abstract
Anaerobic digestion has emerged as an increasingly popular method for the effective management of organic waste and wastewater with the potential to generate renewable energy. However, efficient digestion depends heavily on the complex and diverse microbial communities mediating this process. This review paper delves into the complex microbiome that exists within anaerobic digestion digesters, playing a critical role in the efficient and sustainable conversion of organic waste into biogas. Its primary focus falls within the scope of how various operational and environmental factors, including temperature, pH, hydraulic retention time, substrate loading rates, and the presence of inhibitory compounds, can substantially impact the microbiome of these systems. Through an extensive analysis of the existing literature, the microbial ecology of anaerobic digestion, including the composition and function of the microbial community, their interactions, and responses to different stressors, are evaluated. By offering critical insights into the distribution patterns of active microbial populations upon alterations in various process parameters, a better understanding of the microbial dynamics within these systems is provided, enhancing their efficiency. Overall, this paper emphasizes the importance of unraveling the microbial evolution within AD reactors to optimize biogas production and sustainable waste management and proposes future perspectives in this direction.
Keywords:
anaerobic digestion
; microbial structure
; microbial interactions
; operating parameters
; digestion efficiency
1. Introduction
Two of the major issues humans face today are the ever-growing energy and climate crisis [1], which compromise the life quality of the present and next generations and the planet's future as a whole. A key sustainable solution to the above might be organic waste's anaerobic digestion (AD), thus generating energy for electricity, heating, and vehicle operation [2]. AD is essentially the process through which organic matter, such as agricultural and industrial waste, livestock residues, municipal solid wastes, and sewage [3] is converted into biogas, containing methane (50-70%) and carbon dioxide (30-50%) [4]. Therefore, AD constitutes a promising practice and an intense focal point in research, as it combines the benefits of producing renewable energy while at the same time, it reduces fossil fuel exploitation, waste disposal, as well as methane emissions stemming from the uncontrolled decomposition of organic waste [3,5].
Biogas generation in AD is the result of a large anaerobic microbial consortium processing organic compounds via four successive steps, namely hydrolysis, acidogenesis, acetogenesis, and methanogenesis [1,3,6,7]. A diverse anaerobic bacterial community conducts the first three steps, while the latter is realized by a subset of archaea, called methanogens [7]. More precisely, during hydrolysis, the initial complex organic substrates, mostly polysaccharides, proteins, and lipids, disintegrate into their monomeric components by hydrolytic bacteria [8,9,10]. These monomers are next fermented by acidogens, producing primarily short-chain fatty acids (SCFAs) (acetate, formate, propionate, butyrate, valerate, and isobutyrate) together with CO2, H2, NH3, and sulfide. Some acidogenesis products, like H2, CO2, acetate, and formate can be immediately transformed into biogas by methanogens. However, many SCFAs need first to be subjected to acetogenesis performed by a bacterial group called syntrophic acetogens, giving rise to acetate, H2, and CO2 [10]. These substrates, together with methyl compounds, are then utilized by methanogens, which can be generally divided into three categories, acetoclastic (consuming acetate), hydrogenotrophic (CO2 reduction using formate or H2), and methylotrophic reduction of methyl groups found in methylated compounds, depending on their metabolic pathway [10,11]. The end products of methanogenesis are methane, water, and CO2 [9], which can then be used in various applications.
Although there are discrete steps during AD, the relationships between bacterial and archaeal communities are complex and intertwined, and a fine balance is required in order to ensure efficient system operation [6,12]. The main bacterial phyla found in AD reactors are Proteobacteria, Firmicutes, Bacteroidetes, and Chloroflexi [9,12], while acetoclastic (Methanosarcinales) [13] and hydrogenotrophic (Methanobacteriales, Methanocellales, Methanococcales) [14] archaeal populations are accountable for most of the methane produced [11]. Figure 1 illustrates the most characteristic bacterial and archaeal groups and their involvement in the various stages of methanogenesis in AD reactors.
It has been shown that environmental factors and process parameters, such as pH, temperature, sludge retention time (SRT), hydraulic retention time (HRT), organic loading rate (OLR), feedstock, etc., can strongly affect the digester microbiota in terms of abundance and diversity [1,5], potentially perturbing the underlying syntrophic relationships amongst them [6]. Interestingly, the interplay between syntrophic acetate-oxidizing bacteria (SAOB), which produce hydrogen, and hydrogenotrophic methanogens, which consume it, is of great importance for maintaining high efficacy and stability of the AD process, as propionate and some other SCFAs may act prohibitively on methanogenesis [10]. Acidogenic bacteria is another group that may hinder methanogenic activity, as they generally grow much faster than methanogens, and under certain operating parameters, they can cause agglomeration of acidic intermediates [15,16], which lower the pH value and inhibit methanogenesis [10]. The reason why methanogens are so susceptible to changes in environmental conditions is that they comprise a low-diversity population, resulting in little functional redundancy [12]. Therefore, it is critical that a highly diverse methanogenic archaeal community be established through optimization of the AD process parameters to promote the growth of such populations [16]. Another rate-limiting step is hydrolysis, especially in cases where intricate substrates, like crop residues, animal manure, and municipal waste are used [10]. These feedstocks are quite intractable and difficult to be hydrolyzed, thus diminishing the performance of AD [10,12]. Readily degradable carbohydrates in other feedstocks, though, may result in acidogenesis surmounting syntrophic acetogenesis and methanogenesis, eventually obstructing methane production [10].
All of the above underline the fact that methane yields of an anaerobic digester are greatly dependent on how external and operating parameters are adjusted since the latter are able to influence the structures and dynamics within the intricate microbial networks of the digester. Hence, there is a strong need to enhance the understanding of which conditions of the AD process affect the various microbial populations and how, in order to be able to modulate those factors in favor of shaping multispecies groups that can tackle environmental fluctuations, while the overall performance of the reactor remains potent. The aim of this review is, therefore, to delineate the impact of numerous AD variables on the functional microbial communities and provide valuable information pertaining to the optimization of biogas generation as a result of a stable and efficient AD process.
2. Feedstock
2.1. Feedstock characteristics
Organic waste used in AD has been traditionally originated from sewage sludge, and animal manure and slurry but has further expanded to bio-waste, such as domestic, agricultural, or municipal waste, food processing, and dedicated energy crops [17,18]. These substrates contain different polysaccharides, proteins, and lipids in different ratios, reflected in the conversion capacity and biogas yields. In general, lipids are very rich in carbon and are characterized by great convertibility (94.8%) to biomethane, followed by proteins (71%) and carbohydrates (50.4%) [19].
A widely used substrate in AD is wastewater sludge (WAS), which, however, may have a low carbon-to-nitrogen (C/N) ratio, resulting in decreased efficacy of the digestion process and, eventually, in poor biogas production. Thus, its co-digestion with other feedstocks can enhance WAS conversion to biomethane [20]. Food waste (FW) can be added to improve digestibility of the less degradable WAS and has been reported to boost AD, having greater organic matter and biodegradability [21]. FW, being a calorie-rich feedstock [22], is associated with elevated biogas yield and thus its utilization in AD has increased over the past years [23]. It is more often, though, used in combination with other sources owing to a number of potentially impeding substances it contains, e.g., total ammonia nitrogen (TAN), Na+, lipids, and volatile fatty acids (VFAs), which upon accumulation can undermine the digester operation. Moreover, it is unpredictable in terms of its characteristics, with great variations in composition [23,24]. The latter, for example, may be due to differential aliment determined by each season of the year [25]. The possible lack of essential trace elements (TE) that contribute significantly to the microbial community metabolism is an additional reason why FW is scarcely used on its own, and in this case, co-digestion with animal manure is recommended to compensate for these crucial elements [8]. Highly proteinaceous substrates, like animal excrements, fish processing residues, stillage liquid phase, and slaughterhouse waste, are readily utilized in commercial biogas plants, as they are associated with increased biogas production. Extensive use, however, may disturb the process stability, causing fluctuations in pH, and elevated amounts of VFAs, such as propionate and acetate [2]. Treatment with manure emanated from poultry needs more attention than the cattle-derived one, because it contains uric acid and undigested proteins in large quantities, leading to the production of free ammonia (NH3) and ammonium ions (NH4+), which are toxic to the microorganisms performing AD. In addition, poultry excrements are rich in lignocellulosic materials, encompassing a vast amount of wood chips and shavings, rendering, along with the high ammonia levels, the anaerobic decomposition of such wastes more resistant [5]. Overall, there are different feedstocks available for digestion in AD plants with distinct characteristics and biomethane generation capacity, but in any case, it is of utmost importance to track any alterations of organic and inorganic compounds of influent wastewater to prevent possible negative impacts on methanogenesis [23].
2.2. Co-digestion of feedstocks
Co-digestion is an effective way to combine the beneficial attributes of specific substrates, achieving a greater equilibrium of nutrients, while diluting the concentration of potentially inhibitory and toxic molecules and providing buffering capacity [26,27,28]. Therefore, mixed substrate systems are associated with enhanced methane yields [27] and more efficient digester operation [28] rendering them more economically profitable [27]. A previous study [26] investigated the putative beneficial effects of adding sugar beet pulp lixiviation (SBPL), a waste produced upon sugar beet processing, in a sewage sludge (SS) digester. The results showed that methane production was higher when combining the two substrates together rather than using them separately, proposing this co-digestion scheme as an efficient waste-processing method.
Additionally, it has been noticed that microbial variety in multi-feedstocks is higher, indicating that a more stable ecosystem is generated, with more functionally specialized microorganisms being apparent [9]. The shift in microbial communities in response to changes in feedstock composition from mono- to co-digestion is largely studied. In thermophilic reactors fed with urban organic waste (UOW), the addition of food and plant materials had a profound effect on microorganism repertoire. Specifically, enrichment with a greater food share led to enhanced activity of members of S1 (Thermotogales), Thermonema, and Lactobacillus, while the reactor treated with a higher proportion of lignocellulosic biomass coming from garden waste and grass clippings, groups like Anaerobaculum, Thermacetogenium, Porphyromonadaceae, Ruminococcaceae, and the lignocellulose-disintegrating Clostridium were considerably favored. Bacteroidetes members were undetected upon sludge-only feeding, but substantially increased in abundance in both reactor types after co-digestion occurred. On the contrary, other bacteria, like Proteobacteria, which were quite abundant during mono-digestion, together with less abundant Dictyoglomi, and Chloroflexi, were no longer observable upon co-digestion. As regards the archaeal population, it remained more stable than bacteria during the change in feedstock, with Methanothermobacter and Methanosarcina being the prevalent taxa during the whole period. Methanosarcina members use both acetoclastic and hydrogenotrophic methanogenic pathways, while Methanothermobacter are implicated in syntrophic acetate oxidation (SAO) through hydrogenotrophic methanogenesis, which specifically led to increased methane production during co-digestion [27].
WAS digestion supplemented with either garden grass or fish waste revealed a shift of methanogenic activity from hydrogenotrophic towards acetoclastic methanogenesis, which denotes that this metabolic pathway is preferred in substrates with high organic matter. The primary genus driving the acetoclastic route was Methanosarcina in both co-digestion schemes, while other groups varied, with Methanoculleus species found mainly after treatment with fish waste and Methanofollis and Methanocorpusculum species being more abundant in grass digesters. Grass digestion led to increased microbial variety, as opposed to fish waste, which released high amounts of NH4+ during its degradation, mediating a significant decrease in microbiome diversity. Clostridiales dominated fish waste digesters, along with Bacteroidales, and, to a lesser degree, the more specific to this substrate Synergistales (genera Defluviitoga) and Petrotogales. Grass digesters contained Bacteroidales, Cloacimonadales, Clostridiales, Betaproteobacteriales, Actinomycetales, and Fibrobacterales, which possess carbohydrate-fermenting capabilities [20].
The addition of lipid-rich Na-Oleate in a cattle manure (CM) digester led to an instant increase in methane production, followed by a more unstable period of dropped yield attributed to total VFA accumulation, ultimately reaching the initial methane levels right after lipid addition [29]. Additional data from other studies also support that balance of the AD process can be recovered, given that long-chain fatty acids (LCFAs) range from 1 to 5 g COD/g VSS [30]. LCFAs are degraded via β-oxidation to acetate and hydrogen/formate by SAOB, which develop syntrophic relationships with hydrogenotrophic archaea, the latter converting acetate, CO2, and hydrogen into methane. The abundance of an already dominant methanogen during CM digestion, Candidatus Methanoculleus thermohydrogenotrophicum, was further reinforced after Na-Oleate supplementation, showing that methanogenesis was mainly hydrogenotrophic [29]. Previous reports on Ca. M. thermohydrogenotrophicum showcase that it is favored only under moderate concentrations of LCFAs [31]. Syntrophic bacteria able to digest fatty acids from Syntrophomonadaceae and Clostridiaceae families assisted the growth of this methanogen, and their abundance was also increased. However, when lipid supplementation ceased, the previously copious Syntrophomonadaceae species were diminished, leading to a subsequent decline of Ca. M. thermohydrogenotrophicum, while a propionate-degrading Bacteroidales species and Methanosarcina thermophila became the main players of AD [29].
The priming effect in co-digestion concerns the accelerated degradation of other organic materials as a result of a labile substrate addition. The underlying mechanism of this interaction, though, remains largely unknown. In a study that swine manure was supplemented with either apple waste (primarily containing labile carbon) or fructose (a pure labile carbon), methane production was greatly enhanced compared to mono-digestion, suggesting that a priming effect could have been driven by labile carbon. This effect likely promoted the breakdown of the readily degradable swine manure by enhancing the activity of microbial groups already implicated in substrate hydrolysis, like Firmicutes, and in acetoclastic methanogenesis. The Firmicutes members mostly affected positively belonged to Clostridia and, specifically, to the acetogenic genera Clostridium and Terrisporobacter, while Methanosarcina and Methanobrevibacter were the predominant methanogens [28].
2.3. Microorganisms present in different types of feedstocks
Intricate feedstock composition imposes the shaping of a more complex microbial population, owning a higher metabolic variety [32]. There are some core phyla commonly found in AD digesters regardless of the substrate. Chloroflexi is able to decompose hardly-degradable organic matter, while Bacteroidetes phylum includes proteolytic microorganisms able to degrade proteins. Firmicutes are established VFA degraders producing acetic acid, and grow in syntrophy with methanogens, while Proteobacteria participate in organic waste decomposition by consuming VFAs [33]. In steady-state digesters the most prevalent archaea belong to the order Methanosarcinales followed by Methanomicrobiales [12]. The latter, together with Methanobacteriales, and Methanococcales are exclusively hydrogenotrophic, while the former use mainly acetate in methane production, with Methanosarcina and Methanothrix (formerly Methanosaeta) being the primary representatives. Methanosarcina, in fact, can also utilize the hydrogenotrophic and methylotrophic biomethanation pathways [34]. Supplementary Figure 1 depicts the dominant microbial taxa implicated in AD for various substrates. It is noted that different groups of microorganisms are observed in each feedstock, highlighting the importance of understanding the microbial ecology of AD systems for optimal performance.
Food waste substrate is generally characterized by a less diverse and more distinct microbial community than wastewater sludge [9,23]. However, it is possible that in AD of food waste a more variable community of functionally significant microbiota can be developed [9]. A study focusing on the microbiota of a FW digester pinpointed that the major bacterial populations were Anaerolineaceae, Rikenellaceae, Proteiniphilum, Clostridiales, and Erysipelotrichaceae from the Chloroflexi, Bacteroidetes, and Firmicutes phyla, while core methanogenic community was composed of the hydrogenotrophic Methanobacteriales and Methanomicrobiales and the acetoclastic Methanosarcinales group [33]. It has also been reported that, in sludge-fed digesters acetogenic bacteria belonged predominantly to Cloacimonadaceae (W27, W5, C. Cloacimonas), Rikenellaceae (DMER64, Blvii28), Mesotoga, Bacteroidetes (vadinHA17), and Smithella. Acetogens in FW digesters, on the other hand, were constituted by Fastidiosipila, Rikenellaceae (RC9, DMER64), Cloacimonadaceae (W27) Proteiniphilum, Fermentimonas, and Syntrophomonas. Concerning the archaeal community, WAS digesters were dominated by Methanolinea (M. tarda), WSA2, Methanohrix (M. concilii), Methanobacterium, Methanoculleus, and Methanobrevibacter, whereas FW digesters contained mainly the groups Methanoculleus (M. bourgensis, M. receptaculi), Methanobacterium (M. petrolearium), Methanomassiliicoccus, and Candidatus Methanoplasma. Despite the differences in microbial composition, which was attributed to the higher TAN and Na+ in the FW digester, the main methanogenesis pathway in both digester types was hydrogenotrophic [23].
Another study showcased the predominance of Ruminococcaceae, Cloacimonadaceae (W5), Syntrophomonadaceae, Tepidimicrobium, Methanothermobacter, and Syntrophaceticus in digesters treated with FW, while Anaerolineaceae, Smithella, Cloacimonadales (W27), Longlinea, and Candidatus Methanofastidiosum were detected at high levels in WAS reactors. Interestingly, mixed waste digesters were mainly composed of Proteiniphilum, Methanosphaera, Fastidiosipila, Syntrophaceticus, and Tepidanaerobacter. From the above, the groups Syntrophaceticus, Tepidanaerobacter, Tepidimicrobium and a representative of Syntrophomonadaceae found in FW and mixed reactors are considered syntrophic acetate-oxidizing bacteria [9]. SAO relationships were quite evident in these digesters, where hydrogenotrophic methanogenesis conducted by Methanobacteriaceae and Methanosarcina, was the main pathway, possibly due to the elevated ammonia levels, which inhibit the growth of acetoclastic methanogens [9]. This is in line with other surveys, where, in response to high ammonia, the major methanogenic populations were the more tolerant hydrogenotrophic ones, with evident enrichment of the genus Methanoculleus [22]. This genus has been, indeed, positively linked to increased ammonia, in contrast with genera, such as Methanocorpusculum, Methanobacterium, and Methanothrix, which are more sensitive to such conditions [34]. On the other hand, it was reported that Methanothrix (M. harundinacea, M. concilii), Methanoculleus (M. bourgensis, M. thermophilus), and Methanobacterium (M. beijingense) were the key-drivers of methanogenesis in FW digestion systems [24]. SAO relations can be formed between several hydrogen-consuming methanogens and acetate-oxidizing bacteria, such as Clostridium ultunense, Syntrophaceticus schinkii, and Tepidanaerobacter acetatoxydans upon ammonia stress [34]. In WAS reactors, Smithella genus was associated with a few methanogens and has been shown to be involved in syntrophic propionate degradation generating acetate, which in turn can be uptaken by acetoclastic methanogens. Another group of interest in these reactors was the candidate class Methanofastidiosa, whose high abundance may imply increased levels of sulfur compounds, since it reduces methylated thiol for methanogenesis [9].
Relevant research revealed the dominance of Clostridiales and Bacteroidales in digesters with mixed bio-waste composed of agricultural waste, manure, and municipal solid waste (MSW), while agricultural residue reactors were especially enriched with the genus MBA03 of the Firmicutes phylum. MSW digesters supplemented with green waste had Clostridiales, Cloacamonales and Bacteroidales in high levels, and WAS digesters contained additionally Sphingobacteriales and Syntrophobacterales in prevalence. Cloacimonetes phylum candidates are regarded to be implicated in propionate syntrophic oxidation, rendering them important mediators in the AD of sludge waste. In terms of archaea, WAS reactors were prevailed by Candidatus Methanofastidiosum (WCHA1-57), which is considered to participate in methylated thiol reduction. Archaeal communities of mixed and agricultural waste digesters were quite different from WAS, with the first being enriched with Methanoculleus and Methanothrix, while in the second, Methanosarcina took the lead, probably due to the higher ammonia and total solids content in this substrate. Of note, MBA03 and Methanosarcina concomitant presence in agricultural waste reactors indicates a putative syntrophic association via direct interspecies electron transfer (DIET) taking place during the AD of agricultural residues [17].
2.4. Effect of the inoculum
The influence of the starting inoculum on AD long-term operation, efficiency, and microbial composition is of great concern [35]. It is generally documented that inoculum is an important factor determining the system’s constancy and ability to begin the digestion process [36,37]. Quintero et al. concluded that the appropriate inoculum could lead to higher methane yields, enhanced digestion rate, micronutrient enrichment, reduced lag phase, and AD robustness [38]. Especially during the start-up period, which is the most limiting step, inoculum plays a critical role, particularly in batch digestion [37]. Sewage sludge, solid organic wastes, fresh cow dung or digested manure are some extensively used inocula. The synthesis, as well as the proportion of the inoculum used in relation to a specific substrate, must be considered in order to ensure a beneficial biomethanation rate, and limit potential hindrances associated with the accumulation of toxic compounds [39]. Fine-tuned substrate to inoculum (S:I) ratio provides the basis for a balanced microbial community and facilitates the initial and most crucial step of hydrolysis [37]. In general, a low S:I ratio is more preferable in AD of high total solids (TS) content, aiming to augment the number of methanogens so as to achieve a rapid and effective beginning of the digestion. Nonetheless, it is proposed that every substrate has a unique S:I ratio according to the concentration of VFAs and the capacity to be maintained at certain levels [40].
Although it is suggested that anaerobic incubation of the inoculum prior to loadings can contribute to increased methane production and shorter lag period, owing to the buffering potential of the incubated inoculum, its source seems to influence the AD performance only in the case of non-incubated inoculum [37]. Other findings support that long-term digester performance, hydrolytic, acetogenic, and methanogenic rates, and microbial structure were unaffected by the origin of the inoculum since they all eventually converged in the different digesters after some period. Concerning the microbiota in particular, there was a shift of distinct compositions towards a core community shaped by the process operational parameters, suggesting that these deterministic factors rather than the starting microbial composition of the inoculum are the primary drivers [35]. Similar results were obtained from an additional study, where digesters with separate inoculum origin, developed comparable microbiome and digestion efficiency after 4 HRTs under the same treatment. However, the starting inoculum proved critical for the time required to achieve stable process operation, and the microbial structure development was dependent on the levels of VFAs and ammonia in the inoculum [36]. Importantly, it became evident that the usage of inoculum with similar characteristics to the feeding substrate aided the establishment of the final microbial community, confirming previous hypotheses that it would be beneficial to opt for an inoculum source resembling the preferred key influential factors, including feedstock, pH, and reactor temperature, in order to mitigate the adjustment time for the microbiome [9,26]. The amount of inoculum is also an important parameter to consider, as large amounts are likely to promote high levels of microbial activity, which in turn prevent the system from becoming overloaded with nutrients or other substances that could inhibit or slow down microbial growth [26].
3. Total solids
AD systems are divided into 3 main categories, based on the amount of total solids (TS) present in the substrate: wet AD, with ≤10% TS, semi-dry or high-solids AD (HSAD), with 10-20% TS and dry AD, having a TS content greater than 20% [33,41]. HSAD and dry technology have gained popularity and appear as promising approaches, as they encompass several benefits, like smaller reactor volume requirement, lower energy needed for heating [33,42], increased resistance to certain inhibitors [42], reduced wastewater production, and greater volumetric methane productivity [41]. Moreover, wet digestion of organic waste with increased levels of solids, a significant water quantity would be required for substrate dilution [43]. Nonetheless, the inadequate amount of water in these AD systems results in an inefficient exchange of matter and uneven nourishment for the microorganisms in the digester. In fact, co-digestion is usually implemented in order to supply the necessary nutrients for a balanced bacterial growth [41]. Therefore, the amount of TS in solid waste has a significant effect on the efficiency of anaerobic digestion, particularly with regard to the production of biogas and methane [33,43], occasionally leading to poorer performance [42].
There is a number of studies investigating how microbial communities react in response to variations in the TS content. More precisely, Abbassi-Guendouz et al. observed a decline in the overall production of methane during the mesophilic digestion of cardboard upon TS rise from 10% to 25%, which was attributed to the restricted transfer of material that occurred at higher TS levels [44]. Likewise, in an HSAD system with TS of 15.7%, methanogenic activity was notably lower compared to that observed in the corresponding and low-solids digester (5.7% TS), due to hindrance in the mass transfer caused by the reduced water content in the first system [45]. Conversely, this effect seems to have been overcome in a pilot-scale sludge HSAD (15% TS) equipped with an upgraded stirring mechanism that facilitated mixing, where the functioning of the HSAD resembled that of a low-solids AD system in terms of organic solid removal rates and biogas production [46]. Moreover, in a sewage sludge AD system, it was found that upon increment of TS content from 10% to 20%, reduction of volatile solids (VS) and methane yield only slightly dropped, probably due to high ammonia, while the HSAD could operate on 4-6 times higher OLR, indicating considerably higher volumetric methane production rate [47]. Interestingly, Arelli et al. observed a substantial boost of 70-85% in biogas production, when the TS increased from 25% to 30% in an AD system of FW and CM co-digestion [48]. Generally, drawing a clear conclusion concerning the impact of TS content on the performance of AD systems is challenging. After all, the efficiency of AD as regards methane production is also affected by the organic constituents of the different substrates, such as their carbohydrate, protein, and lipid contents [43].
TS content in feedstocks affects not only the AD performance, but also the microbial structure within the system [33]. As denoted in various studies, changes in fermentation conditions can lead to alterations in the predominant microbial community and metabolic pathways. This is because microorganisms possess differing levels of adaptability to such changes [42]. More precisely, in a study of lab-scale FW AD with increasing TS content from 5% to 20%, a shift in species composition of bacterial and archaeal communities was documented. Three major bacterial phyla were present in the FW digester: Chloroflexi, Bacteroidetes, and Firmicutes. At high TS content, a decreasing trend of Choroflexi, which was represented by genera of the Anaerolineaceae family, was observed. Conversely, Bacteroidetes were augmented as the TS content increased, with Rikenellaceae spp. and Proteiniphilum being the prevalent genera, indicating their importance in protein hydrolysis. Firmicutes abundance, mainly represented by Clostridiales and Erysipelotrichaceae, remained at comparable levels throughout the experiment, suggesting their role as syntrophic acetogenic bacteria. Furthermore, as the feeding TS contents increased, the proportionate quantities of other phyla such as Proteobacteria, Spirochaetes (Spirochaeta), and Tenericutes (Acholeplasma) also increased, playing important roles in FW degradation. Concerning methanogens, Methanosarcina was found to be the dominant methanogen regardless of the TS content, indicating that acetoclastic methanogenesis was the principal pathway of methane production. In the low-solids anaerobic digester, hydrogenotrophic Methanoculleus was the second more copious methanogen, though its abundance declined upon TS content increment, in contrast with Methanosarcina, which slightly increased, due to higher acetate. Therefore, the hydrogenotrophic pathway had a smaller role in generating methane in high-solid AD. The increase of TS below 20% was found to have a positive effect on biogas production and methane content due to the greater amount of readily degradable substrate for microorganisms [33].
Han et al. compared the microbiome of wet (TS ≤ 10%) and semi-dry (TS 10%-20%) digesters treating various types of FW and found that two distinct microbial communities were formed. Porphyromonadaceae, Sphingobacteriaceae, and Syntrophomonadaceae families were predominant under wet conditions, while the semi-dry digesters were characterized by Clostridiaceae, Patulibacteraceae, Pseudonocardiaceae, Lachnospiraceae, Rikenellaceae. Methanobacteriaceae were highly abundant in the wet digesters and correlated positively with Syntrophomonadaceae, whereas they were barely detectable in digesters with increased TS content. On the contrary, Methanomicrobiaceae were evident under wet conditions and found to be the most prevalent in the semi-dry digesters. These results indicate that methanogenesis was mainly conducted via the hydrogenotrophic pathway. Soluble chemical oxygen demand (SCOD) to chemical oxygen demand (COD) ratio, VS and COD removal efficiency, and methane production were significantly higher in the wet digesters than in the semi-dry digesters, while in the latter, TAN was relatively elevated. Moreover, VFA concentration was notably reduced in high TS digesters and was attributed to the lower hydrolysis rate due to limited mass transfer in these reactors, thus confining acidogenesis [49].
In another study, the effects of FW with agricultural waste co-digestion in an HSAD system with 20% total solids were examined. These two substrates combined in a ratio of 5:1, respectively, yielded the highest biogas production. Bacteroidetes, Synergistetes Firmicutes, Patescibacteria, and Cloacimonadota were the dominant phyla, represented by the hydrolytic genera Proteiniphilum, Fastidiosipila, Gallicola, Aminobacterium, Syntrophaceticus, and W5053. Of these, Aminobacterium, Syntrophaceticus, and Gallicola were strongly correlated with VFA levels and CH4 production, indicating their role in the biodegradation of the co-digested substrate. Methanogenic communities consisted primarily of Methanobacterium and Methanosphaera, followed by Methanoculleus, and Methanosarcina, implying that hydrogenotrophic methanogenesis was predominant, with methylotrophic pathway also present, since Methanosphaera can utilize CO2 to produce methane. Overall, this HSAD co-digestion system exhibited great buffering capacity and even at high concentrations of NH4+-N, the system remained unaffected [41].
Wang et al. investigated the AD performance of digesters that co-digested pig manure (PM) and FW at varying TS concentrations (5%-20%). According to the findings, the specific methane yield did not vary substantially as the TS content raised from 5% to 15%. However, at 20% TS content, a reduction was obvious. Wet digesters were highly enriched in Chloroflexi (Anaerolineaceae), and to a lesser extent in Firmicutes, and Proteobacteria, while an opposite trend was observed in the dry digesters, with Ruminococcaceae, Clostridium sensu stricto, Syntrophomonas, and Terrisporobacter being the most prevalent. Methanogenesis was syntrophically supported by the last three bacterial groups which promoted the degradation of organic acids, albeit in dry systems the methanogenic process was hampered due to the presence of high VFA and ammonia levels. Dry digesters were characterized by a shift from acetoclastic to hydrogenotrophic/methylotrophic methanogenesis. Wet AD greatly involved Methanothrix, but the proliferation of this genus was inhibited as a result of VFA and ammonia accumulation in the dry digesters, leading to an enrichment of other more versatile methanogens, such as Methanosarcina, Methanosphaerula, Methanoculleus, and Methanobrevibacter, which utilize H2 and/or CO2 for methane production [43].
A study of digesters that were treating sewage sludge with increasing TS content also revealed a change in microbial communities. The primary bacterial phyla detected in all reactors were Firmicutes, Bacteroidetes, WS6, Proteobacteria and Chloroflexi. In digesters with TS 10% & 15% named HSAD1, Firmicutes were the most abundant phylum developing syntrophy with acetogens, while in TS 17% & 19% (HSAD2), they were markedly decreased. Bacteroidetes, on the other hand, became prevalent in HSAD2, putatively because of the greater amount of protein that was added to the denser feedstock. Of note, as the TS content increased from 10% to 19%, there was an apparent increase in the abundance of the archaeal community, indicating a rise in archaeal diversity. This helped to sustain consistent methanogenesis in the digester with denser concentration of solids and ensured a steady-state methane yield. The main methanogenic pathway was the acetoclastic in all HSAD systems, with Methanosarcina being the most ubiquitous genus, overpowering Methanothrix, likely because the abundance of intermediary substances, such as VFAs or free ammonia created an unfavorable environment for Methanothrix. However, at higher solids levels, Methanosarcina population was significantly diminished, concurrently with enrichment of more hydrogenotrophic or methylotrophic methanogens. Hence, at such extreme concentrations of ammonia, the resilience of Methanosarcina was still not as high as certain hydrogenotrophs. These included Methanobacterium of the class Methanobacteria, and Methanospirillum, Methanoculleus, and Methanimicrococcus of Methanomicrobia. In addition, the genera Methanomassiliicoccus and Ca. Methanomethylophilus of Thermoplasmata, which produce methane by reducing methyl compounds, were also favored [42]. Table 1 summarizes the microbial populations that are positively correlated with each TS content AD category.
4. Ammonia concentration
The production of biogas can be greatly affected by the presence of substances that are harmful to microorganisms. Ammonia levels that are less than 200 mg L-1 can benefit AD since microbial metabolism depends highly on nitrogen [29]. In addition, nitrogen also provides sufficient buffering capacity to the system, helping prevent acidification during the initial stage [36]. The level of residual ammonia is not solely influenced by the ammonia delivered from protein degradation via the deamination of amino acids. Since it is consumed by microbes as a nitrogen source, its eventual concentration in AD digesters is designated by the intricate equilibrium between its anabolic consumption and catabolic production. This balance is strongly reliant on the coordinated actions of microorganisms within the digester [50]. It has been reported that 3 g NH4+-N L-1 is the highest ammonia level that does not cause any adverse effects in sludge mesophilic reactors. In contrast, levels greater than 5 g L-1 seem to hinder AD activity and lead to VFA accumulation [51]. The threshold for NH3 in mesophilic reactors is between 0.14-0.28 g NH3 L-1 [2]. In an aqueous solution, total ammonia nitrogen (TAN) can be found in two forms: free ammonia nitrogen (FAN-NH3) and ammonium nitrogen (NH4+), which are in equilibrium [29].
Two parameters that gravely influence the proportion of FAN-NH3 and NH4+ are pH and temperature, with their increment causing a rise in NH3, which is considered the primary inhibitor among the two forms. Consequently, microbial communities are subjected to high toxicity levels [5]. Elevated FAN levels can obstruct the AD process both directly and indirectly. This involves passive diffusion of FAN into the cells of bacteria and methanogens, which can directly hinder the essential metabolic enzymes, and disrupt the pH and proton motive force (PMF) within the microbes' cytoplasm [23,24]. In addition, when the C:N ratio is very low, because of high protein content, NH4+ levels in the environment will likely be augmented, impeding the growth of anaerobic microorganisms [19]. In fact, it has been found that C:N ratio under 50 can cause complete cessation of methane production [32]. Moreover, solid-state AD is more prone to generating excessive amounts of ammonia nitrogen compared to liquid AD. This is attributed to the greater organic loading and reduced water content in the solid-state process [52].
The stages of degradation that rely on acetoclastic or hydrogenotrophic methanogens are the ones that are most affected by ammonia levels. As a result, the pathways higher up in the degradation process are also impacted [2,5]. The community that tends to show the most notable sensitivity are the acetoclastic methanogens [2,50]. Adaptation to high ammonia exposure involves a shift to SAO coupled with hydrogenotrophic methanogenesis as the principal method for converting acetate [2,5,50]. SAO is a two-step process involving the oxidation of acetate to produce H2 and CO2 by SAOB, and the subsequent hydrogenotrophic methanogenesis conducted by the corresponding archaeal population. Though under standard conditions, the first reaction is not thermodynamically favorable, it can occur if hydrogen pressure is reduced by hydrogenotrophic methanogenesis through interspecies hydrogen transfer (IHT) [53]. Hydrogenotrophic pathway is the primary route in many environments, as hydrogenotrophs have a lower threshold for hydrogen compared to acetogens for acetate. Additionally, the energy yield from the conversion of carbon dioxide and hydrogen to methane is greater than that from the conversion of acetate [32].
A major concern raised by elevated ammonia levels is that they appear to spawn the accumulation of VFAs, especially propionate and acetate [2,51], which is associated with instability of the AD processes [51]. All the more evidence supports that consistent CH4 production is contingent on acetogenesis, rather than methanogenesis, as NH4+ exerts irreversible limiting effects on the syntrophic bacteria that oxidize acetate and propionate, causing VFA accumulation [54]. Propionate appears to be a key factor that governs the shift in microbial communities, leading to the production of methane in environments with high NH4+ [51]. Hence, the pathway for propionate degradation is an important topic of investigation. This would include syntrophic propionate-oxidizing bacteria (SPOB), which could help alleviate this issue by oxidizing propionate under conditions that constrain the activity of acetoclastic methanogens [2]. Notably this population seems to be negatively affected by high ammonia [51], except for some ammonia-resilient SPOB [55]. Therefore, the numbers of such groups could be a deterministic factor for an effective methane production [51].
4.1. Effect of elevated ammonia on microbial communities
In mesophilic sludge digesters with increased ammonia load, Candidatus Brevefilum of the phylum Chloroflexi was recognized as a possible mediator of syntrophic propionate oxidation, assisting the moderation of propionate accumulation, and permitting the continuation of methane production despite the extreme inhibition levels due to high ammonia. This genus was present, together with Ca. Cloacimonas, and Syntrophorhabdus, which are also putative SPOB, in the highest ammonia concentration level of 5 g NH4+-N L-1. Nonetheless, Ca. Brevefilum was the only group that increased under such conditions, denoting a pronounced tolerance. Other groups that dominated the increased NH4+ digesters were the hydrogen-producing Paraclostridium, Enterococcus, and Romboutsia, along with Proteiniphilum and Turicibacter. The archaeal community was mainly composed of the acetoclastic Methanothrix in all reactors, though its numbers gravely decreased compared to those in digesters with mild ammonia concentrations. This aligns with the fact that this genus is generally sensitive to NH4+ [51]. More precisely, Methanothrix are obligatory acetate-consumers that exhibit a greater affinity for acetate, but have slower growth rates, when compared to Methanosarcina. Hence, Methanothrix dominate the digesters when acetate concentrations are low [42,50]. However, this genus possesses thin filaments, which increase the exposure surface, rendering it more vulnerable to toxic compounds, such as high ammonia [42]. Methanosarcina, on the other hand, have greater development rates and can assemble into irregular cell clumps. Thus, they are more resilient to variations in pH and elevated levels of VFAs and ammonia [33,42]. Adding to that, Methanosarcina can harness both the acetoclastic and hydrogenotrophic methanogenesis, making them more resistant to certain inhibitors of the acetoclastic route unlike Methanothrix [33]. Therefore, in this study Methanosarcina, being more adaptive, was profoundly enriched, together with the hydrogenotrophs Methanoculleus and Methanobacterium, owing to their ability to withstand high VFA concentrations. These prevailed methanogens probably facilitated the effective utilization of acetate, hydrogen, and CO2 generated by hydrogen-producing bacteria and SPOB, providing CH4 even in the presence of high NH4+ levels [51].
The effects of increased ammonia (0.14-0.28 g NH3 L-1) were assessed in reactors co-digesting household waste and albumin operating at 37 °C. The results showcased that SAO was the main route for creating methane from acetate, with Clostridium ultunense, Syntrophaceticus schinkii, and Tepidanaerobacter acetatoxydans being the most prominent syntrophic bacteria [2]. The latter two were also found enriched in a wastewater plant treated with high grease waste proportion, growing in syntrophy with Methanosarcina to utilize acetate [9]. Moreover, the genus Syntrophaceticus has been reported as capable of enduring disturbances resulting from the accumulation of VFAs and ammonia inhibition and can preserve the efficiency of AD even when exposed to exceptionally high levels of ammonia [41]. In contradiction to prior research findings, SAO was dominant in the presence of various acetate concentrations ranging from > 0.1 to 70 mM, suggesting that a low acetate concentration is sufficient for a SAO-mediated operation. This implies that high acetate levels may not necessarily lead to the development of SAO in digesters operating under high-ammonia mesophilic regime. Methanogenic archaea were mainly constituted by the hydrogenotrophic Methanoculleus bourgensis of the class Methanomicrobiales, while Methanosarcina acetivorans, M. mazei (Methanosarcinaceae) and Methanobrevibacter smithii (Methanobacteriales) were also present. However, acetoclastic Methanotrichaceae members were not detected. The effective operation under mesophilic conditions with high ammonia levels in these digesters where M. bourgensis prevailed, highlighted the crucial role of this methanogenic group in achieving a potent performance [2].
Chicken waste, a highly enriched in ammonium substrate, was fed to mesophilic reactors with increasing OLR in order to study the effects on microbial communities and digester performance. Over the course of the experiment, TAN and FAN concentration reached 4.56 g L-1 and 416 mg L-1, respectively. High ammonia levels caused a diminution of the initially predominant Bacteroidales members Marinilabiaceae and Porphyromonadaceae, whereas Clostridium, as well as Gallicola had an opposite trend. Thus, it was proposed that SAO was adopted as an alternate acetate-consuming pathway, since Clostridium has been a well-documented genus participating in syntrophic acetate oxidation. With regard to archaea, at the beginning the most abundant group was Methanobacterium, however when ammonia and VFA levels increased, Methanosarcina became prevalent, possibly cooperating with Clostridium in SAO as hydrogen-utilizing methanogen [5]. These two methanogenic groups are known to be resistant under ammonia-stressed conditions [56]. Of note, in this system Methanobacteriaceae outcompeted Methanomicrobiaceae, in contrast to other studies where at high levels of ammonia, Methanomicrobiales group was crucial in acetate utilization through SAO, while Methanobacteriales development was impeded [50,57].
Lee et al. compared the microbial communities between mesophilic digesters fed with sewage sludge and digesters treated with food waste and observed that TAN was higher in FW reactors, ranging from 1.7 to 6.5 g TAN L-1, compared to that of sludge digesters (0.1–2.2 g TAN L-1) and led to overall lower AD performance in the former. Methanoculleus was the primary methanogenic genus in FW digesters, with Methanoculleus receptaculi outcompeting Methanoculleus bourgensis, while on the other hand, both of these two species were augmented in the sludge digesters. This implies that M. receptaculi and M. bourgensis may have similar competitiveness when exposed to high ammonia concentrations within the range of 0.1-2.2 g TAN L-1 (2-145 mg FAN L-1), but when ammonia concentrations are higher within the range of 1.7-6.5 g TAN L-1 (70-1.184 mg FAN L-1), M. receptaculi may outperform M. bourgensis. Moreover, Methanobacterium petrolearium seems to be quite sensitive to ammonia since its abundance decreased at even 2.2 g TAN L-1 (140 mg FAN L-1). Cloacimonadales W27 was highly associated with M. receptaculi and M. bourgensis in the digesters, suggesting that this group could act as a syntrophic partner to these hydrogenotrophic methanogens under high ammonia conditions [23].
In another research where high ammonia in FW and sludge co-digestion mesophilic systems was studied, NH4+ levels were approximately 1.6 times higher in the digester fed nearly almost with FW, compared to the digester with a smaller share of FW. This higher ammonia concentration was accompanied by decreased methane yields of this digester. Clostridia was the most prevalent class, with Caldicoprobacter algeriensis, Clostridium senegalense, Ercella succinigene, and Gelria glutamica being the most prominent species. Microbacter and Proteiniphilum belonging to Porphyromonadaceae family of Bacteroidia and Defluviitoga tunisiensis of Thermotogae were also relatively abundant. Of these groups, G. glutamica and D. tunisiensis seemed to be slightly positively correlated with high NH4+ levels. Specifically for G. glutamica it has been observed to coexist with hydrogenotrophic methanogens, which in this study were mainly represented by M. bourgensis, followed by Methanomassiliicoccus luminyensis, indicating that methane production was achieved through the hydrogenotrophic pathway in this digester driven by the increased ammonia [25].
Kalamaras et al. examined the acclimation of microbial structures in response to stepwise increase in ammonia using consecutive batch cultures and manure-based substrate. The ultimate concentration of ammonia reached the extreme value of 9 g TAN L-1 but CH4 production was already markedly decreased by 6 g TAN L-1 compared to levels up to 5 g TAN L-1. The highest TAN reactor was also characterized by a poorer microbial variety. The activity of cellulose-degrading bacteria generally diminished as the concentration of ammonia increased, with the exception of Cellulosilyticum ruminicola (Cellulosilyticum) and to Alkaliflexus imshenetskii (Ruminofilibacter), which were not inhibited by the rising NH4+ levels. Possible syntrophic associations were attributed to S. zehnderi, S. bryanti and S. sapovorans, which all belong to the genus Syntrophomonas and are known to break down butyrate and certain LCFAs in syntrophy with hydrogenotrophs. Clostridium chauvoei, which can generate acetate and butyrate by utilizing pyruvate, was also identified as a potent SAO bacterium. Along with the increment in ammonia, these microorganisms showed an increasing activity as well. Other bacteria with consistent activity in all batch cultures were Hydrogenispora ethanolica and Acetomicrobium hydrogeniformans, both of which are thought to develop syntrophic relations with hydrogenotrophs. The archaeal community was mainly dominated by Methanosarcina flavescens and M. bourgensis, which demonstrated enhanced activity regardless of the ammonia concentration. This indicated that the above-mentioned species appear to be tolerant to increased TAN and that hydrogenotrophic methanogenesis was adopted under such regime. Other hydrogenotrophs belonged to the genera Methanobrevibacter and Methanocorpusculum, with the former slightly increasing in activity, while the latter decreasing considerably upon ammonia stress. In addition, the initially highly abundant methylotroph Methanomassiliicoccus luminyensis showed a marked decrease in consequence of elevated ammonia, however other methylotrophic groups, albeit being less active at low ammonia, displayed an opposite trend upon TAN augmentation. Overall, gradual TAN exposure resulted in effective microbial adaptation for concentrations up to 5 g TAN L−1 [29].
A practice commonly described to tackle high ammonia concentrations and optimize system performance is introducing TE. Adding a TE mixture composed of iron, cobalt, selenium, nickel tungsten, and hydrochloric acid to mesophilic digesters led to alterations in the abundances of the acetogenic bacterial consortium and a robust enrichment of M. bourgensis. Furthermore, it substantially increased biomethanation and digester performance, while alleviating VFA accumulation [2]. Moreover, it has been reported that zeolite addition in a mesophilic digester treating high-ammonia substrate resulted in increased Methanosarcina numbers and enhanced removal efficiency of TAN and VFA uptake, thus promoting AD stability. In this system, however, a substantial CH4 augmentation was not evident [5]. Another study found that TE Fe, Ni, Zn, and Mo played a crucial role in stabilizing the anaerobic co-digestion of food waste and sewage sludge at high organic loads. These elements reduced the accumulation of inhibitory substances, such as propionic acid and ammonia, resulting in higher biogas yield and biodegradability. At higher OLR, Cu and Co were also added to handle the more acute ammonia buildup. This supplementation could lower ammonia levels and maintain stability for OLR up to 14 g L-1 [58]. Overall, these reports highlight the importance of TE in enhancing performance and process stability under high ammonia pressure. In Table 2, the microbial groups that are resilient under increased ammonia conditions are outlined.
5. Temperature
5.1. Different temperature regimes
AD can be conducted at various temperature ranges, spanning from mesophilic (30-40 °C) to thermophilic (50-65 °C) conditions [19,59]. The operation of digesters at hyper-mesophilic temperatures, namely from 40 to 44 °C has also been observed in the few past years [60]. It has been established that raising the temperature in anaerobic digestion systems promotes a higher metabolic activity of the microorganisms involved in the biochemical processes [8], leading to an acceleration in the substrate degradation rate and a more robust methane production [59,61]. This is attributed to the increased activity of hydrolytic enzymes, which lower the hydraulic retention time (HRT) [62] by enhancing hydrolysis rates [8,61], allowing the effective digestion even of more complex and challenging substrates, like lignocellulosic material [52,62]. Moreover, the growth rates of thermophilic methanogens are two to three times higher than those of their mesophilic counterparts, contributing to the greater metabolic rates observed in these systems [24]. Hence, thermophilic reactors can be reduced in volume, which implies fewer construction costs, while efficiently producing biogas [26]. Of note, the influence that temperature has on AD is primarily related to the degradation rate, rather than the overall biodegradability of a substance [63].
Nevertheless, despite their efficacy, thermophilic digesters are prone to lower process stability compared to mesophilic digesters [24], which can be at least partially justified by the fact that they are characterized by a more limited microbial population variety [8]. Thus, thermophilic microorganisms, including methanogens [26], are particularly susceptible to alterations in operational and environmental conditions [62]. Attempts for adaptation of these communities to increased temperatures may entail the peril of reactor acidification [62] and rises in temperature of more than 1-2 °C could cause a substantial decline in methane production [26]. Notably, a disruption in the balance between the fermentative and methanogenic populations can be incited by the unremitting VFA accumulation under thermophilic conditions [64], as a result of higher ammonia content [51]. Therefore, managing and refining the thermophilic process is a challenging task, which is why the majority of biogas facilities still employ mesophilic AD to generate biogas. These systems benefit from greater stability and favorable growth conditions, thus being less vulnerable to toxic compounds, such as ammonia, while also effectively processing a wider range of biomass and waste materials [26]. Furthermore, most methanogens are mesophilic [26] and under such conditions several routes (acetoclastic, hydrogenotrophic, and methylotrophic) contribute to methane production, whereas thermophilic temperatures dictate a shift to mainly hydrogenotrophic pathway [56]. Noteworthy is also the fact that mesophilic AD involves fewer heating expenses [52]. Overall, the preferred operating temperature may vary depending on the special characteristics of each biogas plant, including substrate feeding and residence time.
5.2. Changes in microbial composition due to temperature variation
Several research teams have observed the differences in microbial population structure between mesophilic and thermophilic digesters, which are summarized in Supplementary Figure 2. For instance, de Jonge et al. compared 18 full-scale reactors that processed different rations of food waste with temperatures ranging from 37 °C to 58 °C. The major factor that contributed to the microbiome shaping was temperature. More precisely, in mesophilic reactors, Firmicutes, Bacteroidetes, Cloacimonetes, and Chloroflexi prevailed in general. Abundant genera within digesters with a high share of FW or mixed wastes were Fastidiosipila, Syntrophaceticus, Tissierella, Methanosphaera, Methanobacterium, and Methanosarcina, while those digesters treated primarily with wastewater sludge were represented by Candidatus Cloacamonas, Methanoregulaceae and Candidatus Methanofastidiosum. It was inferred that hydrogenotrophic methanogenesis was the main pathway for CH4 production in FW-rich digesters, owing to the extensive presence of hydrogenotrophs and their possible association with SAO bacteria, like Syntrophaceticus and Tepidanaerobacter [9]. In WAS digesters, the high abundance of the candidate class Methanofastidiosa, could indicate increased levels of sulfur compounds within the reactor. The phyla Thermotogae, Firmicutes, Bacteroidetes and Synergistetes dominated the thermophilic digesters, with the MBA03 family and the genera Defluviitoga, Anaerobaculum, and Methanothermobacter being prominent. The dominance of the latter suggests that hydrogenotrophic methanogenesis also governs the thermophilic digesters [27]. Another study that observed twenty full-scale anaerobic digesters treating FW under mesophilic (35-39 °C) and thermophilic (54-57 °C) conditions, also found the genus Defluviitoga in great numbers in the thermophilic digesters, while Methanoculleus was the most ubiquitous methanogen in both temperatures. Some other mesophilic digesters were dominated by either Methanothrix, Methanospirillum, or Methanobacterium, and a general trend toward hydrogenotrophic methanogenesis was observed in both types of digesters [22].
Another study evaluated the microbial communities involved in AD of FW full-scale biogas plants operating at different temperatures (35-36 °C or 50-57.5 °C) and the factors influencing these communities. All reactors had comparable performance in terms of biogas yield. Firmicutes and Bacteriodetes were the dominant phyla, with Fastidiosipila, Petrimonas, Syntrophomonas, VadinBC27 wastewater-sludge group, Acholeplasma, and Sphaerochaeta being shared in all digesters. High temperatures seemed to favor the growth of Tepidanaerobacter, Proteiniphilum, and Caldicobrobacter, whereas Christensenellaceae R-7 group and Rikenellaceae RC9 gut group were more abundant in mesophilic digesters. Caldicobrobacter, along with Ca. Caldatribacterium and Ruminiclostridium were poorly associated with COD removal efficacy, while closely linked with acetate and Na+ at elevated temperatures. It was, therefore, proposed that they could act as indicators of process instability at thermophilic regimes. Notably, the proportion of Proteiniphilum was greater in thermophilic rather than mesophilic digesters, despite all identified Proteiniphilum species being mesophilic and growing in the temperature range of 15-45 °C. This observation suggested the possible existence of uncultured thermophilic species within this genus. Concerning the archaeal populations, members of the genera Methanothrix (M. harundinacea, M. concilii), Methanoculleus (M. bourgensis) and Methanobacterium (M. beijingense) were enriched in mesophilic reactors. Methanoculleus (M. bourgensis, M. thermophilus) and Methanobacterium (M. beijingense) genera were also highly present in thermophilic conditions. Ammonia concentration in these digesters was twice as much as that in lower temperatures, which could have led to impediment of acetoclastic methanogens and justifies the total prevalence of hydrogenotrophs [24]. Interestingly, in a slightly different setting of digesters operating under high ammonia, a rise of temperature from 37 °C to 42 °C caused a subtle increase in ammonia levels, along with an augmentation of Methanomicrobiales and Methanosarcinaceae, albeit with a decline of M. bourgensis abundance [2].
Heyer et al. investigated the performance and the microbial communities of mesophilic (33-35 °C) and thermophilic (40-54 °C) biogas plants that digested a wide variety of substrates. Thermophile digesters were enriched with orders, such as Thermotogales, Deinococcales, and Spirochaetales, while the presence of Clostridiales and Thermoanaerobacteriales was linked to lower temperatures. The archaeal community in thermophilic reactors was composed mainly of Thermococcales, Methanococcales, and Methanosarcinales, whereas Methanobacteriales and Methanomicrobiales were most abundant in the mesophilic digesters. Positive associations between the latter and members of Clostridiales and Thermoanaerobacteriales were indicative of SAO relationships. Metaproteomics analysis revealed that more proteins involved in the pathways for methane production and cellular transport were detected in mesophilic digesters. This was consistent with the fact that at moderate temperatures, methane can be generated through various routes such as acetoclastic, hydrogenotrophic, and methylotrophic, while at high temperatures the hydrogenotrophic pathway is the primary means of generating methane. Overall, high temperature was correlated with enhanced biogas productivity [56].
The effects of temperature were assessed in sludge digestion of wastewater plants operating at either 34 °C, 38 °C or 42 °C. Raising the temperature to 42 °C caused an imbalance in the process, as implied by the decrease in methane production, together with a rise of VFA and ammonia levels. On the other hand, when the temperature decreased to 34 °C, the amount of specific methane produced was comparable to that of 38 °C, though with a minor biogas production rate and sometimes foaming. Increasing the temperature also impacted the methanogenic population, whilst the drop to 34 °C had no significant effect. More precisely, under hyper-mesophilic conditions, Methanomicrobiales, the dominant order at 34 °C and 38 °C, was outcompeted by Methanosarcinaceae and Methanobacteriales. The similar numbers of both acetoclastic and hydrogenotrophic methanogens was attributed to the ammonia increase up to mid-optimal levels for the growth of these two groups and neither of them was favored more. As regards the lower temperature, it was speculated that Microthrix parvicella, whose preferred temperature is below 35 °C, or other filamentous bacteria were likely related to the foaming situation observed. All in all, digesters operating at 38 °C had the most efficient process performance [60].
Shaw et al. compared the microbiome structure in mesophilic (37 °C) and thermophilic (55 °C) digesters using swine manure as substrate. Results showed that the amount of methane produced under moderate- and high-temperature conditions was similar at all time periods measured. The microbial composition at mesophilic conditions exhibited greater variety with a larger number of enriched families being involved in biogas production. Conversely, most abundant microbial groups at high temperatures were more implicated specifically in methanogenesis [59]. Accordingly, it has been reported that at temperatures lower than 65 °C, microorganisms affiliated to methanogens were the predominant population, whereas at higher degrees, acidogenic bacteria were more prevalent [65]. Spirochaetaceae and Enterococcaceae families that participate in hydrolysis and acidogenesis, respectively, promoting the breakdown of organic compounds, were more increased at 37 °C. Moreover, Synergistaceae was also abundant in these reactors, whereas Lachnospiraceae, Enterobacteriaceae, and Planococcaceae were mostly found in thermophilic digesters. Families, like Ruminococcaceae, Clostridiaceae, Corynebacteriaceae, and Prevotellaceae were observed at both temperatures. The higher level of digestion at thermophilic temperatures can result in the accumulation of certain VFAs, thus potentially lowering the pH. However, the stable pH levels in both mesophilic and thermophilic digesters observed in this study suggest that higher temperatures may have led to an earlier shift towards the later stages (acetogenesis or methanogenesis) of AD, rather than hydrolysis or acidogenesis. Methanogenic populations differed between the two temperatures. The families Methanotrichaceae, Methanocorpusculaceae, Methanoregulaceae, and Methanomassiliicoccaceae were found in high richness at 37 °C. On the other hand, high temperatures favored the proliferation of Methanobacteriaceae and Methanomicrobiaceae. Methanocorpusculaceae and Methanomicrobiaceae, which dominated at each temperature, were the most influential methanogens, indicating that in both cases, methanogenesis was driven by the hydrogenotrophic pathway [59].
5.3. Effect of temperature combined with OLR changes
A study of microbial responses to increasing OLR from 1.0 to 2.5 g VS L−1 day−1 in mesophilic (35 °C) and thermophilic (55 °C) reactors treating FW as a substrate revealed that the mesophilic temperatures showed better and consistent performance in terms of methane production compared to the thermophilic ones. This was because mesophilic reactors had a higher diversity and even distribution of bacterial species, in contrast to thermophilic, in which acetate and propionate concentrations accumulated up to much higher levels upon OLR increase, showing instability. Firmicutes and Synergistetes were detected in both temperatures, but the dominant phyla for each digester were Thermotogae, Bacteroidetes, Spirochabacter, Proteobacteria and Tenericutes at high temperatures and Bacteroidetes, Chloroflexi, Actinobacteria, and Spirochaetes at mesophilic ones. Genera, like Defluviitoga and Fervidobacterium, Gelria, Lachnospiraceae uncultured, Ruminococcaceae Incertae Sedis, Sporanaerobacter, Tepidanaerobacter, Petrobacter, and Anaerobaculum were enriched in thermophilic digesters, whereas at the lower temperature Kosmotoga, Synergistaceae uncultured, Petrimonas, Paludibacter, Proteiniphilum, Sporanaerobacter, and Prevotella dominated the reactors. Along with OLR elevation, most of these genera had fluctuations in their relative abundance, indicating the influence of this parameter. There was no significant difference in the methanogen variety among mesophilic and thermophilic digesters, but there was a clear difference concerning genus composition. Methanosarcina was an omnipresent methanogen in both digesters, while Methanothrix was mainly found in mesophilic, and Methanothermobacter and Methanoculleus were more abundant in thermophilic conditions. At high temperatures, hydrogenotrophic methanogenesis played a more important role in methane production compared to moderate ranges, and Defluviitoga was a dominant bacterial genus that contributed to the syntrophic hydrogen production in thermophilic digesters [64].
In a similar study, AD at 35 °C and 55 °C of corn stalks or wheat straw while increasing the OLR, showed that raising the temperature at stable OLR of 5 g VS L−1 day−1 led to a notable boost in the production of methane, especially in the case of wheat straw digestion. The amount of biogas produced increased as more substrate was added, and the process could handle higher substrate loads up to 45 g VS L−1 day−1. However, additional substrate did not result in more biogas production, owing to possible inhibition or limitations in mass transfer. In this high-temperature setting, the most copious phyla were Firmicutes, Proteobacteria, Bacteroidetes, Spirochaetes, Synergistetes, Chloroflexi, Coprothermobacterota, and Actinobacteria. Among these phyla, the most abundant classes were Clostridia, which can tolerate great amounts of VFAs, followed by Bacteroidia, which are largely involved in substrate degradation and are crucial for preventing acidosis. Bacterial genera detected in the 55 °C digester included Proteiniphilum, Proteiniborus, Pseudomonas, Advenella, Treponema, Gracilibacter, Parabacteroides, Variimorphobacter, Comamonas, Anaerobacterium, Ruminiclostridium, Acetomicrobium, and Thermoclostridium. The methanogenic community consisted of Methanocorpusculum, Methanobacterium, Methanoculleus, Methanoregula, Methanosarcina, Methanomassiliicoccus, which, with the exception of the latter two, are hydrogenotrophic. Methanocorpusculum aggregans and Methanobacterium formicicum are typically found in moderate temperatures and their presence in a bioreactor that operates at high temperatures had not been documented before [61].
5.4. Microbial adaptation upon temperature alteration
Studies that evaluate the adaptation of AD digesters of upon operating temperature changes have also been conducted. Tukacs-Hajos et al. investigated the anaerobic fermentation of sugar beet-pressed pulp in pilot-scale digesters, focusing on the thermophilic adaptation of a mesophilic microbial community in digesters operating at 38 °C. The process involved a gradual increase in temperature until 55 °C, where at steady-state, greater biogas yield was observed, despite the higher VFA levels. Metagenomics analysis revealed that the hydrolytic and acidogenic bacteria could faster than the methanogenic archaea, with Proteobacteria decreasing in numbers, while Thermotogae and Synergistetes significantly increasing. The abundance of different methanogen types also changed in response to the temperature increase, indicating a shift from acetoclastic to hydrogenotrophic methanogenesis. This change in acetate consumption might have affected the acidogenic consortia, possibly explaining the apparent decrease in Firmicutes at steady-state. Mesophilic digesters were dominated by genera, like Methanothrix, Methanosphaera, Methanoregula, Methanospirillum, while in thermophilic conditions the main groups benefitted were Methanothermobacter and Methanoculleus. Methanosarcina, though, remained the most abundant genus during the whole experimental period. Overall, the rise of Thermotogae was pivotal in stabilizing the system after a significant increase in acid levels due to temperature increment. This also underlines that the temperature gradient applied during the transition from mesophilic to thermophilic operation is significant for effective acclimation [62].
In another study, AD of swine manure was initially operated at 37 °C and once the reactor reached steady state, the temperature was lowered to 23 °C for eight days. The sudden temperature decrease caused a drop in biogas and methane production, and a decrease in the removal efficiency of COD and TS. However, when the temperature was raised back to 37 °C, there was an instant recovery of pH value and methane production returned to the initial levels. Temperature disturbance differently affected the various microorganisms in terms of both composition and function. Groups, such as Porphyromonadaceae, Ruminococcaceae, Bacteroidetes and Bacteroidales were favored, while Synergistaceae and the acetogenesis-related Syntrophaceae dropped in numbers. Moreover, the acidogenic bacteria Veillonellaceae and Flavobacteriaceae were outcompeted by Enterococcaceae following the temperature changes. Interestingly, the abundance of Fibrobacteraceae and Gracilibacteraceae was raised during the disturbance period, however after temperature restoration, the former decreased while the latter decreased. The study also revealed a shift within hydrogenotrophic methanogens from Methanocorpusculaceae and Methanoregulaceae to Methanomicrobiaceae, Methanobacteriaceae and Methanospirillaceae. Furthermore, the acetoclastic Methanotrichaceae and Methanosarcinaceae were replaced by the methylotrophic Methanomassiliicoccaceae. These findings indicated that the disturbance in temperature was sufficient to shift the microbiome to a novel steady-state [66].
6. Volatile Fatty Acids (VFAs)
6.1. Factors causing VFA accumulation
Acetic acid is known to be the primary carbon source for biomethanation through AD. Most of the other VFAs, such as propionic acid, are also transformed into acetic acid before microbes incorporate them into the methane metabolism [67]. Under standard conditions, the transformation of certain VFAs, like propionate and butyrate, towards acetate, H2, and CO2 requires energy absorption, rendering it challenging to happen without external aid during AD. This step is, therefore, considered rate-limiting for the process [68]. In order to evaluate the stability of AD, the concentration of VFAs is a critical metric [5]. Partial alkalinity, pH and VFAs are credible signs of process imbalance in low buffering capacity reactors, but in greatly buffered systems, pH alterations can be minor, thus VFAs are regarded as the sole dependable index for monitoring [69].
Being a vital component in the methanogenic process by serving as substrates, VFAs are positively linked to methane generation, when their concentration remains within a particular range. However, it is important to note that excessive aggregation of VFAs can have an inhibitory effect on AD, ultimately leading to system failure [67]. There is a number of factors accountable for VFA accumulation, mainly linked to turbulences in the conditions of AD [26]. For instance, VFAs can overly increase, when high organic loading [62] or certain attributes of the influent feedstock, such as enhanced carbohydrate content [20], may result in acute generation of VFAs, which cannot be entirely utilized by methanogens [24]. In addition, if the concentration of obstructive compounds, such as Na+, TAN, and lipids in the reactor surpasses the levels that are impeding to methanogenic activity, they can lead to a buildup of unconsumed VFAs [24].
Another critical parameter is temperature, as VFA accumulation can be noticed at higher temperatures [26,62], probably due to the sensitivity of acetoclastic methanogens to temperature [26]. Moreover, elevated TS content in the digester may reduce the hydrolysis rate, thus which subsequently restricting the acidogenic process [49]. Regardless of the underlying cause, accumulated VFAs may lead to reactor acidification and pH drop [5,26]. Under such conditions, the growth of methanogenic communities is constrained [26], since VFAs interfere with their metabolism by entering the cytoplasm and dissociating into VFA anions and protons [19]. Ultimately, the AD operation might completely cease [5].
6.2. Effect of VFAs on microbial communities
Comprehending the behavior of microbial communities responsible for VFA production or valorization is essential to process optimization. Studies on this topic commonly induce VFA buildup by increasing the OLR. For instance, Mathai et al. used mixed waste (FW, municipal, brewery, etc.) digesters with different OLRs (1-6 g COD L-1 day-1) to assess the alterations of the microbial community makeup upon VFA elevation. Digesters that operated at an OLR of 5 g COD L-1 day-1 or higher experienced significant process disturbance, as indicated by an accumulation of VFAs, a decline in pH levels, and an 80% diminution in the rate of methane production. Microorganisms responsible for propionate and butyrate declined in numbers as the OLR increased. These groups included Syntrophobacter, Pelotomaculum and Smithella for propionate and Syntrophomonas for butyrate. From their kinetics characteristics, it was inferred that Syntrophobacter have greater affinity for propionate, whereas Pelotomaculum have higher specific growth rates. Upon higher OLR the acetoclastic Methanothrix showed a gradual decrease, together with a concomitant rise in Methanoculleus and the SAOB Tepidanaerobacter acetatoxydans, indicating a change in acetate consumption from being broken down via acetoclastic methanogenesis to being oxidized through SAO. Owing to their slower growth rates compared to acetoclastic methanogens, SAOB did not process acetate quick enough to alleviate its excessive amounts. Of note, notwithstanding the accumulation of acetate, Methanosarcina were not favored, probably due to the abrupt onset of adverse growth conditions [70].
Similar findings were observed in another study, in which acidification was induced via OLR increase in reactors fed with sugar beet pulp. The pH was then mitigated with external addition of NaOH and NaHCO3 to adjust its value. During the late acidosis phase, CH4 production decreased, while upon recovery of the pH, total VFA levels dropped, and efficient methane production was restored. Key bacterial phyla detected initially were Bacteroidetes, Firmicutes, and candidate phylum WWE1, represented by Porphyromonadaceae, Lachnospiraceae, and Cloacamonaceae, respectively. Dominant archaeal families comprised Methanomicrobiaceae, Methanobacteriaceae and Methanotrichaceae. When increased OLR was employed, replicate reactors began to show diverged microbial communities, with the majority of microorganisms displaying functional adaptation redundancy to the environmental alterations. Bacteroidetes were characterized by much less diversity than Firmicutes, albeit being more resistant to the elevated OLR and low pH. One reactor was also enriched in Thermotogae phylum. Surprisingly, in contrast to other reports, archaeal richness and diversity increased upon acidosis. The formerly predominant Methanothrix was outcompeted by Methanoculleus, signifying that hydrogenotrophic methanogenesis became the prevailing pathway. However, none of the common SAO bacteria was detected. After pH restoration, Methanosarcina increased in numbers replacing Methanoculleus and the candidate genus vadinCA11, which is potentially halophilic, was also detected. Concerning bacteria, Porphyromonadaceae and Bacteroidaceae (Bacteroidetes), as well as Sedimentibacter, Clostridium bolteae and unclassified Epulopiscium (Firmicutes) were favored. The presence of the propionate-producing C. bolteae was consistent with the elevated propionate levels observed at that period. All in all, there was an apparent differential evolvement of microbial communities in the replicate reactors upon exposure to unfavorable conditions [65].
Additionally, in AD of lignocellulosic biomass at high OLR, led to the buildup of VFAs, along with a decrease in both pH and methane output. Microbial composition analysis revealed the prevalence of Proteiniphilum, Marinilabiliaceae, Synergistaceae Bacteroidaceae, Ruminococcaceae, and Lachnospiraceae, while syntrophic bacteria, such as Syntrophaceae and Syntrophomonadaceae were only detected in small numbers. Methanosarcina were the main archaeal group, followed by Methanobacterium, thus the acetoclastic methanogenic pathway was considered to be predominant in this study, during VFA accumulation. When microaeration was implemented to tackle high VFA concentration, Proteiniphilum appeared to be pivotal in the transformation of VFAs into CO2 [71].
Moreover, another study focused on methanogenic population changes upon elevated VFA levels in mesophilic and thermophilic anaerobic digester plants. Results showed that despite the significant increase in VFAs, pH values and methane generation remained quite stable, and methanogens were mostly unaffected. This was explained by the addition of cow manure in the substrate composition, which is rich in bicarbonate and ammonia and has likely contributed to system buffering. Similar to the aforementioned study, Methanosarcina abundance was augmented upon VFA increment in the mesophilic reactor, which was consistent with previous work underlining that the growth of this genus is promoted by acetate accumulation. Conversely, the numbers of Methanobacterium and Methanoculleus were not influenced by the high VFA presence. Methanothermobacter was the predominant genus under thermophilic conditions, however, VFA accumulation did not alter its abundance. Other abundant genera comprised Methanosarcina, which exhibited an increase, as well as Methanobacterium and Methanoculleus, which were rather unaffected by the VFA uprise [69]. A relatively different response of the methanogenic community was documented in a thermophilic reactor treating synthetic wastewater, where Methanoculleus was initially the most highly enriched methanogen but decreased in abundance during VFA buildup. Contrarily, Methanothermobacter and Methanosarcina increased in abundance during this period. Upon system stabilization, Methanosarcina selectively increased in abundance, indicating its contribution to the degradation of acetate, and remained stable afterwards. Of note, Methanothermobacter decreased to the point of being the least abundant methanogen, while Methanoculleus became predominant until the end of the experiment [72].
In a study where the influence of certain VFAs on microbiota and methane yield under thermophilic conditions was assessed, it was found that while the presence of increased formic acid resulted in evident methane production, elevated levels of acetic, butyric, and propionic acid, significantly suppressed methanogenesis and triggered a rise in hydrogen concentration. The hydrogenotrophic Methanoculleus thermophilus and Methanothermobacter wolfeii were the two dominant species and were both positively linked to biogas production. Exposure to high levels of propionic and butyric acid led to substantial diminution of these two species. Μ. thermophilus in particular, though considered a major player in fatty acid utilization under low VFA conditions, was also sensitive to formic acid accumulation. M. wolfeii, on the other hand, was promoted by increased formic loads and became the predominant species accounting for the enhanced methane generation under such conditions. An opposite trend was observed with high acetic acid concentrations, where M. wolfeii was inhibited, whilst Μ. thermophilus was not, indicating that this methanogen might have been favored by syntrophic acetate-utilizing bacteria [73].
Other works that correlated the abundance of different microbial groups with VFAs have showed that, for instance Syntrophomonas were associated with low propionate levels and high acetate can be positively correlated with Methanosarcina, Coprothermobacter, Syntrophomonas, Clostridium, and Solibacillus from the phylum Firmicutes. Syntrophomonas was also positively influenced by butyrate, along with Anaerobaculum [27]. Moreover, Proteiniphilum growth is inhibited at elevated acetate, propionate, and butyrate levels, and it is thus considered quite sensitive to acidosis and high VFAs. Caldicoprobacter, Candidatus Caldatribacterium, and Ruminiclostridium are three genera possibly indicative of imbalance in thermophilic AD due to acidification, since they produce acetate via sugar fermentation [24]. Pelotomaculum thermopropionicum, Proteiniphilum acetatigenes, and Ercella succinigenes have also been associated with increased acetate concentration. Conversely, Acholeplasma brassicae and Desulfotomaculum thermocisternum seem to have no positive correlation with acetate levels [25]. Interestingly, apart from Methanosarcina, which are resilient at increased VFAs [27], Methanotrichaceae have also shown an increasing trend with VFA elevation and their relationship with bacteria that degrade LCFAs has been reported [50]. The candidate archaeal class WSA2 is also worth mentioning, since it is the first known methanogen exclusively involved in methanogenesis through methylated thiol reduction, utilizing carbon sources such as acetate, malonate, and particularly propionate [17]. Given that reactors may experience propionate accumulation under less ideal conditions [17], and that propionate is a marker of acidogenesis and methanogenesis imbalance, causing inhibition of the latter [50], WSA2 might contribute to overall AD process stability [17]. Important propionate oxidizing bacteria seem to be found within the candidate phylum Cloacimonetes, probably participating in syntrophic associations with methanogens [17]. All in all, it can be seen that there is a clear influence of VFAs on the abundance of different microbial groups, with some being positively linked to certain VFAs while others being inhibited.
7. pH
7.1. The role of pH and its variation
A suitable pH level must be maintained to ensure efficient functioning of AD and optimum decomposition of VFAs. A VFA:alkalinity ratio from 0.0 to 0.1 indicates a stable system, whereas values among 0.1 and 0.4 suggest auspicious operational conditions with low to no risk of acidification. The acidity of VFAs is typically counterbalanced by the alkalizing effect of methanogens, which produce bicarbonate, ammonia, and carbon dioxide, ultimately maintaining a stable pH in the system [26]. Microbial community structure and activity are significantly affected by pH, as it plays a crucial role in regulating them. Acetoclastic methanogens, in specific, have a strict optimum pH range favoring their growth, and their performance can be severely suppressed at pH values lower than 6.2 [8]. Moreover, pH is a crucial parameter that impacts the ability of trace elements to dissolve in AD systems [74]. Acidification is a frequent operational failure observed in numerous AD reactors and can occur when VFAs accumulate, often due to substrate overloading or the usage of readily degradable feeding sources. Temperature decrease can also contribute to this phenomenon. High VFA levels denote an imbalance disequilibrium between VFA generation by fermentative and acidogenic bacteria, and their utilization by acetogenic bacteria and methanogenic archaea [65]. Indeed, a decrease in pH seems to promote the production of VFAs via enhancing the acidogenic activity, while concurrently impeding methanogenic activity [26], thus resulting in reduced methane production [65].
High VFA levels can directly impede methanogenesis, and this effect is more pronounced at lower pH levels. This is due to the ratio of undissociated, and dissociated forms being altered by pH. The undissociated exerts more toxic effects by permeating through the cell membrane and lowering the intracellular pH, which ultimately causes cell activity impairment. Given that acidogenesis typically occurs quickly and may contribute to the VFA buildup, further decreasing the pH value, it may potentially lead to diminution or complete obstruction of methanogenesis. Hydrolytic bacteria are also fast-growing microorganisms and are more tolerable to changes of the pH unlike methanogens [75]. Acidosis can be moderated by using substrates containing high protein content, which, due to NH4+ enrichment, exert buffering properties and prevent pH drop [19]. For anaerobic microbes to effectively break down proteinic compounds, the pH needs to be above 6, since the ability of FW-derived proteins to dissolve may be diminished at lower pH levels [22]. Mesophilic conditions seem to promote organic nitrogen degradation, therefore resulting in a slightly basic pH compared to higher temperatures [8]. Overall, acidification in AD plants is involved in complications, such as decreased methane generation and the production of a digestion residue so acidic that is not suitable for subsequent fertilization purposes [65].
7.2. Influence of pH on microbial communities
As stated above, balance within the microbial community is evidently affected by pH changes. Table 3 contains the positively affected microbial communities under different pH ranges. Studies have been mainly focused on hydrolytic and acidogenic populations, as their structure seems to be quite dependent on pH value. Some reports mention an increased relative abundance of Proteobacteria, Aminicenantes, Actinobacteria, and Nitrospirae at acidic pH of 6.0 and a lower Firmicutes presence compared to higher pH values [52]. Another study observed lower AD performance upon pH level increase to 8.6, where percentages of Petrimonas and Proteiniphilum were elevated, suggesting that these two genera may be tolerant to alkaline conditions. Proteiniphilum, though, seems to be negatively affected by acidic pH values [24]. Other reported alkalophiles are Tissierella and Tepidimicrobium, which thrived at pH 9.0 under thermophilic conditions [76].
The bacterial composition among different pH values was assessed by Zhou et al. during the digestion of cabbage waste. Measurements of VFA levels showed greater amounts at pH 7.0 in the beginning compared to these of pH 5.0, 6.0, and the uncontrolled pH reactors, but they declined towards the end of the fermentation process at pH 7.0, primarily because acetate levels were diminished. The production of VFAs was likely inhibited during the initial fermentation stage due to the low pH levels, as evidenced by the relatively low VFA levels at pH 5.0 and uncontrolled pH. Thus, such low pH values were responsible for hindering the hydrolysis of cellulosic content and the operation of cellulolytic bacteria. Microbial analysis revealed the presence of common degrading phyla, namely Firmicutes, Bacteroidetes, Proteobacteria, Actinobacteria, and Chloroflexi. The latter, with the class Anaerolineae was detected in significant numbers only in the inoculum, possibly due to competitive inhibition. Bacteroidetes abundance was favored in pH 6.0 but declined under more acidic conditions, unlike Firmicutes, which followed opposite trends. Bacteroidia were found to be associated with acetate and propionate production and Clostidia, which immensely proliferated as the fermentation progressed, mediated the production of acetate and butyrate. The abundance of Lactobacillus from class Bacilli was reduced in all pH samples along with fermentation time, denoting that lactic acid was broken down and consumed [77]. This genus was also found abundant in acidic reactors (pH <6) of FW digestion [22]. In pH 6.0 the main fermentative genera were Bacteroides, Clostridium IV, Dialister, and Ruminococcus, while lower pH favored Clostridium IV, Clostridium sensu stricto, unclassified_Ruminococcaceae [77].
Initial pH adjustment also plays a role in shaping the microbial communities as detected in a study of cow manure AD where pH was set at 5.5, 6.5, 7.5, or 8.5. Firmicutes, Bacteroidetes, Actinobacteria, and Spirochaetes were the general prevalent phyla. The pH level of 8.5 resulted in the greatest yield of VFAs, followed by the pH 7.5 digester. pH values in all reactors dropped within 4 days of operation to a level between 6.0-5.0 and continued to be stable thereafter, suggesting that the acidogenic phase was predominant. Firmicutes abundance was increased in all pH values, while the Actinobacteria population showed a substantial rise at the beginning, followed by a considerable decrease until the end of the experiment. The most abundant groups detected at the early digestion period were Bifidobacterium (all pH values), Treponema (pH 8.5), Prevotellaceae YAB2003 group, Prevotella, and Lachnospiraceae (pH 6.5, 7.5, 8.5). These bacteria were predominantly hydrolytic and decreased in numbers with time. The later fermentation phase was characterized by the Firmicutes representatives Romboutsia, Turicibacter, Peptostreptococcaceae, Clostridium sensu stricto 1, Paeniclostridium, Ruminococcaceae, Fonticella and Caproiciproducens. Romboutsia and Paeniclostridium had the highest numbers in all digesters, which remained consistent along the procedure. Clostridium sensu stricto 1 had an increasing trend at pH 5.5, 7.5, and 8.5 with the highest abundance at the latter pH value. Fonticella was undetected initially, though its abundance was later enhanced at pH 6.5 and 7.5, unlike in pH 5.5 and 8.5 reactors, where it decreased significantly. The population of Ruminococcaceae initially increased and was higher at pH 6.5, but then diminished, while Caproiciproducens was more enriched at pH 5.5, but only after a period of time. Finally, Turicibacter showed an increasing trend at pH 5.5, whereas in the rest digesters remained stable. The microbial community's metabolic activity varied significantly due to the impact of the initial pH on different acidogenic pathways. [78].
Investigation of the microbial consortia at different pH conditions (6.0, 7.0, and 8.0) during mesophilic AD of swine manure disclosed changes in both bacterial and archaeal populations, accompanied by a shift in the methanogenesis pathway. The hydrolytic Clostridiales and Bacteroidales dominated all digesters. The family Porphyromonadaceae (Bacteroidales) was more abundant at pH 7.0, followed by pH 8.0, while it was undetectable at pH 6.0. Interestingly, the bacterium Solibacillus silvestris, also present in the inoculum, was highly observed in pH 8.0 reactors. This microorganism is capable of producing an extracellular bioemulsifier glycoprotein which reinforces the tolerance at a variety of different pH. Moreover, an unclassified SAO group, which utilizes formate and H2/CO2 was identified at pH 8.0. Sphaerochaeta (Spirochaetes) seemed to be favored at neutral pH and is known to produce acetate, ethanol, and formate to be used in methanogenesis. Setting pH to 7.0 benefitted the growth of Methanocorpusulum, which can utilize H2/CO2 or formate to produce CH4, whereas at pH 6.0 and 8.0 Methanosarcina dominated. VFA degradation and methane production were more pronounced in the pH 7.0 conditions. In general, both alkaline and acidic environments led to reduction in bacterial diversity and a minor alteration in archaeal diversity. The structure of microbes participating in the degradation phase might affect the methane production pathways [79]. In conclusion, pH plays a critical role in determining the diversity and abundance of microorganisms in anaerobic digestion. It is, therefore, essential to maintain the optimal pH range for the growth and operation of specific microbial populations that are responsible for the efficient degradation of organic matter and the production of products, which can then be used in methanogenesis.
8. Hydraulic Retention Time (HRT) & Organic Loading Rate (OLR)
8.1. HRT and OLR in anaerobic digestion systems
HRT is a controllable factor in anaerobic digestion that determines the average time that a substance spends in the system. It can be easily adjusted by changing the flow rate without the need for chemicals. In continuously stirred tank reactor (CSTR), HRT plays a critical role in regulating the degradation time, microbial growth, and the level of organic matter degradation, as well as the intermediate substance buildup [80]. It is, thus, essential to pinpoint an appropriate HRT to achieve a stable and effective AD system. Methanogens have a growth rate of approximately 3.6 days, so short HRTs are not typically preferred for methane production [81]. Depending on the substrate feeding the optimum HRT may vary, for instance lignocellulosic biomass require a longer HRT due to their resistance to microbial breakdown. In terms of reducing the costs and improving process performance shorter HRTs are more preferable [82]. However, lowering the HRT below a certain threshold can result in the loss of methanogens, VFA and hydrogen accumulation and a decline in pH [80]. Therefore, to ensure optimal conditions for each bacterial group and preserve their maximum potential, two-stage AD is considered a promising approach, as it combines shorter HRT, efficient substrate degradation, and greater energy yields. This system divides the different phases of biogas production into a hydrolysis-acidogenesis stage followed by acetogenesis-methanogenesis stage, where in the former VFA and H2 are produced, and the latter includes VFA conversion to methane and CO2 [81]. Staged digestion systems usually use HRTs of 2-4 days for the first reactor, while it has been noted that running the methanogenic stage with HRTs below 8-10 days may result in instability of the methanogenic process [8].
Apart from HRT, OLR has also a significant impact on the efficiency of the process and digester stability [83], affecting the growth and metabolic activity of microbial communities [83,84]. AD of slowly digestible substrates (e.g., cellulosic materials) may lead to poor methane production, therefore these digesters are typically operated at increased OLRs with feedstocks containing high TS content as a way to address this issue [71]. Nevertheless, if the OLR is raised beyond a specific level, it may result in significant VFA accumulation [85] via rapid conversion of organic matter through fermentation [22], thus inhibiting methanogenesis [86]. This results in the disruption of the balance between syntrophic acetogenic bacteria and methanogens as it affects the hydrogen partial pressure in the system [71]. In addition, excessively high or unstable OLR can lead to a reduction in the diversity of microbial communities present in the system [25], underscoring the importance of maintaining stable and favorable conditions for microbial growth and activity. In order to maintain high methane yields upon increasing OLR within acceptable values, co-digestion is often utilized [85].
Distinguishing between the effects of HRT or OLR is challenging, as alterations in HRT are often linked to changes also in OLR in continuous anaerobic digesters [87]. When the HRT is decreased for a certain substrate constitution, the OLR increases, which implies that microorganisms are exposed to a higher substrate amount per unit of time [80]. Altering the HRT and OLR can cause a cascade effect on the different stages of the digestion process, leading to changes in the functional activity of enzymes, and trophic interactions among microorganisms [87]. Moreover, changes in these two parameters highly determine the quantity and retention time of inoculum added to the digester, thus affecting the adaptation time and the kinetics of microorganisms [88]. High HRT and a low OLR seem to favor hydrogenotrophic methanogenesis and one-carbon metabolism of bacteria, therefore promoting SAO. This is consistent with the slow growth rates of bacteria performing SAO. Conversely, there has been a positive association of high OLRs and low HRTs with the occurrence of acetoclastic methanogenesis as well as bacterial glycolysis, while SAO is not favored [56]. Methanosarcina has been reported to withstand high OLRs, while some Methanothrix species can grow at short HRTs, regardless of the elevated organic acid levels [34]. AD systems generally produce more methane per unit volume by combining higher OLR and shorter HRT [80].
8.2. HRT & OLR impact on microbial communities
8.2.1. HRT
Peces et al. attempted to dissociate the effect of HRT from that of OLR in mesophilic digesters treating various substrates, by decreasing the HRT (15 to 2 days) while maintaining the OLR constant. According to the results, reducing the HRT led to a decline in methane output and a rise in VFAs, accompanied by a decrease in COD removal and lower microbial diversity. The hydrolysis rate and hydrogenotrophic methanogenesis were not affected by HRT diminution, but the activity of acetogens as well as acetoclastic methanogenesis, were found to be influenced. Regarding the hydrolytic groups, at shorter HRTs the proportion of the Sphingobacterium genus and the candidate family SB-5 increased, while there was a decrease in abundance of Marinilabiliaceae, DMER64 (Rinkenellaceae), Lentimicrobium, and SR-FBR-L83. The Mesotoga genus (Thermotogota) was dominant at longer HRTs but was replaced by the candidate AUTHM297 at lower HRTs. Anaerolineaceae, had a high abundance at HRTs of 4 days and 2 days, but it was not detected in brewery wastewater samples. Firmicutes and Spirochaetes were not significantly influenced by the change in HRT. With HRT decrease, the abundance of certain acetogenic bacteria, such as Syntrophorhabdus, Syntrophobacter, Smithella, and Candidatus Cloacimonas, declined, except for Lactivibrio. Methanothrix was the most ubiquitous methanogen across all HRTs and digesters and did not fluctuate in numbers due to changes in HRT. The hydrogenotrophic Methanoculleus and Methanoregula were more dominant at longer HRTs (15 and 8 days), whereas Methanobacterium tended to grow at lower HRTs. This study showcased that HRT per se did not cause methanogenic community washout, putatively due to biofilm formation, which was observed at 2-day HRT, thus sustaining stable methane production even under such conditions [80].
Another study focusing on the effects of reducing the HRT found that in thermophilic co-digestion systems (sludge with food and plant waste) with HRT decreasing from 30 to 10 days sugar-fermenting microorganisms, such as Thermonema, S1 (Thermotogales), Caldicoprobacter, the SAO Thermacetogenium, and Lactobacillus increased as HRT dropped, until 10-day HRT, where they decreased in abundance except for Thermonema and S1, indicating process inhibition, which led to reduced methane production. On the other hand, the numbers of Acinetobacter, Solibacillus, Dictyoglomus, Proteiniclasticum, Exiguobacterium, Fervidobacterium, Bacillus, Allochromatium and SMB53 decreased. Throughout HRT decrease, the prevailing genera were cellulolytic (S1, Thermonema), acid-degrading (Anaerobaculum) and proteolytic (Coprothermobacter). At 10-day HRT, S1 and Thermonema were the only prominent bacteria, probably owing to their higher growth rates, which increased their tolerance at shorter HRT. The predominant pathway for methane formation was hydrogenotrophic methanogenesis coupled with SAO, with the Methanothermobacter being the dominant methanogen except for the 10-day HRT where Methanosarcina became prevalent [27]. In a similar thermophilic AD system of FW and sludge co-digestion, the interspecies syntrophic relationships that developed at short HRTs, significantly promoted hydrogenotrophic and mixotrophic methanogenesis, mediated by Methanoculleus thermophilus and Methanosarcina thermophila, respectively [87].
In a semi-pilot scale mesophilic reactor treating FW, HRT of 124 days was accompanied by the highest methane yield, as well as COD and VS removal efficiency. A drop of HRT from 124 to 62 days resulted in VFA and ammonia accumulation and methane production begun to decline until 31-day HRT. The dominant phylum throughout the whole experiment was Bacteroidetes, with Bacteroides, Dysgonomonas, Proteiniphilum, DMER64, and uncultured Sphingobacteriia being the main genera. In fact, the abundance of the first three was enhanced upon HRT drop. Firmicutes followed in abundance and their major representatives were Streptococcus and Enterococcus from the Bacilli class, as well as Sedimentibacter, Tepidimicrobium, and Syntrophomonas belonging to Clostridia. While Bacilli decreased as the HRT was diminished, Clostridia had an increasing trend. Other phyla present in the system were Proteobacteria (Proteus), Patescibacteria (Parcubacteria), Cloacimonetes (Candidatus Cloacimonas and LNR A2-18), Spirochaetes (Sediminispirochaeta and Treponema), Synergistetes (Synergistia), and Actinobacteria (Actinomyces). Proteobacteria were enriched with HRT decrease, while Patescibacteria were almost undetectable at 31-day HRT. All the other phyla had fluctuating abundances during the whole period. As regards methanogens, Methanobrevibacter, Methanothrix, and Methanobacterium were initially the most abundant and hydrogenotrophic methanogenesis was the most dominant pathway. Upon HRT reduction, Methanosarcina became dominant at 62-day HRT, followed by Methanobacterium and Methanocorpusculum, while at 41-day HRT the latter prevailed along with the methylotrophic Methanimicrococcus. These methanogens were partially washed-out when the HRT decreased to 31 days, which together with acidogenic bacteria increment, led to process failure [89].
Gaby et al. investigated the impact of HRT diminution (20 to 13 days) on mesophilic digesters and observed no significant reduction in methane generation. Clostridium dominated in the beginning, but it was outcompeted by Syntrophomonas and the uncharacterized genus W22 upon HRT reduction. Other groups, like Tissierellaceae, Ruminococcaceae, and Bacteroidaceae decreased in abundance, whereas Porphyromonadaceae slightly increased. Methanosarcina and Methanoculleus were the predominant archaea at 20-day HRT, indicating the conduction of mainly hydrogenotrophic methanogenesis, however when HRT decreased to 13 days, these groups were reduced in numbers, while Methanobacterium and Methanothrix were evidently increased. Methanospirillum was only detected at longer HRT, whereas Methanothermobacter was mostly present at shorter HRT. Overall, these digesters were characterized by high richness of hydrogenotrophic and acetoclastic methanogens, putatively maintaining comparable methane yields despite the lower HRT [8].
8.2.2. OLR
Changes in OLR can also critically influence the microbial composition of an AD digester, thus affecting the whole process effectiveness. For example, Ziganshina et al. found that increasing the OLR from 1.0 to 3.5 g VS L-1 day-1 in mesophilic digesters fed with poultry waste while keeping the HRT constant led to a dramatic decrease in Bacteroidetes, unlike Firmicutes, which were greatly enhanced. The less abundant Tenericutes, represented by Acholeplasmataceae, also increased, while Synergistetes, with the main family being Synergistaceae, was diminished. The most enriched groups within Firmicutes were unidentified Erysipelotrichaceae, Clostridium, and Gallicola, while Bacteroidetes comprised mainly by Marinilabiaceae and Porphyromonadaceae. Archaeal diversity was lower, with Methanobacterium and Methanosarcina dominating during the whole period. While the former was more abundant at OLR 1.0 g VS L-1 day-1, it was outcompeted by Methanosarcina upon OLR increase. It was suggested that elevated OLR promoted acetate utilization via SAO, with Clostridium forming syntrophic relationships with Methanosarcina, as well as with the hydrogenotrophs Methanobacterium and Methanoculleus. Under such conditions, ammonia and VFA accumulation was observed and pH value increased [5].
In a mesophilic co-digestion reactor of vinegar residue and swine manure, OLR of 5.0 g VS L-1 day-1 was the maximum that the system could tolerate and when OLR was increased to 6 g VS L-1 day-1 the accumulation of VFAs led to a cessation of feeding for a period of 7 days. Upon recovery, the digester operated at 5.5 g VS L-1 day-1 and methane production was similar to that before. The acid-producing Bacteroidetes with Ruminofilibacter and VadinBC27 wastewater sludge group and Firmicutes with Sedimentibacter, Ruminiclostridium, Christensenellaceae R-7 group, Syntrophomonas, Acetivibrio were initially the dominant phyla. Candidatus Cloacimonetes (Ca. Cloacamonas) and Spirochaetes (Treponema 2) Fibrobacterota, and Proteobacteria were also highly present. Uprise of OLR at 6 g VS L-1 day-1 led to Bacteroidetes decrease, while Firmicutes increased. Moreover, Candidatus Cloacimonetes and Proteobacteria also decreased in numbers. Spirochaetes, though, seemed to benefit from high polysaccharide content and increased upon the inhibition and recovery period. In the genus level, Ca. Cloacamonas and VadinBC27 wastewater sludge group initially increased but after OLR reached 6 g VS L-1 day-1 they diminished. Ruminofilibacter and Treponema 2 had increasing trends upon OLR increase and were also present during the recovery stage. Sedimentibacter also showed a positive relation with OLR, since its abundance was enhanced at high OLR, indicating a significant involvement in SAO during that period. At stable-state, the acetoclastic Methanothrix was the main methanogen, followed by the less abundant Methanosarcina, Methanobacterium, and Methanobrevibacter. At an OLR of 6.0 g VS L-1 day-1, Methanosarcina and Methanobrevibacter increased, while Methanothrix was greatly reduced. During the recovery stage, the proportions of Methanothrix and Methanobrevibacter declined, whereas Methanosarcina became the predominant population, consuming the residual acetate but also acting as hydrogenotroph [85].
In another study, during mesophilic co-digestion of restaurant grease waste and municipal wastewater sludge, overloading was caused by increasing the grease waste content. Upon stable operation, Methanothrix and Methanomicrobium were the major methanogens, while when OLR increased from 3.84 to 8.00 g COD L-1 day-1, Methanosarcina was favored, along with Methanomicrobium, but with a reduction in Methanothrix. This indicates a shift towards hydrogenotrophic methanogenesis. Concurrently, the digester was characterized by low pH, high VFA levels and decreased biogas production. Regarding the bacterial community, Ca. Cloacamonas of the phylum Spirochaetes was the most ubiquitous group and remained stable in numbers in spite of loading changes. However, Escherichia (Proteobacteria) was found increased upon substrate overload, while Anaerolinea (Chloroflexi) was reduced. The main driver for microbial composition changes was VFA accumulation [90].
While most studies focus on the effects of stepwise increase of OLR, in actual AD process, load shocks and periodic fluctuations in OLR resulting from variations in substrate quality and operational factors are more common. Such conditions were investigated by Wu et al., who used mesophilic and thermophilic AD reactors fed with sludge. During the disturbance period VFA and ammonia levels were similar to those at stable-state, possibly due to the high buffering capacity of the systems. However, parameters like biogas yield, gas production rate and volatile TS removal efficiency were more variable. In the mesophilic reactor the dominant phyla were Proteobacteria, Chloroflexi, Nitrospirae, and Spirochaetes, while in the thermophilic were Firmicutes, Acidobacteria, Cloacimonetes, and Chloroflexi. Interestingly, When the OLR fluctuation began, only the microbial community thermophilic digester experienced changes, with the hydrolytic norank_o_Aminicenantales (Acidobacteria) and Acetomicrobium (Synergistetes) increasing in abundance. Conversely, the VFA oxidizers W5 (Cloacimonetes) and unclassified_f_Lentimicrobiaceae (Bacteroidetes) were diminished. Concerning the archaeal communities, mesophilic reactor was governed by Methanothrix and the hydrogenotrophs Ca. Methanofastidiosum and Methanospirillum, while in thermophilic reactor Methanosarcina dominated, followed by Methanothrix. OLR disturbance had no effect on the methanogenic community in thermophilic conditions except for Methanomassiliicoccus, which was augmented. The overall stable performance of this digester was attributed to functional redundancy of the microbial populations [91].
Braz et al. studied the impacts of an abrupt organic overload of glycerol on the microbial community and process disturbance in sludge digesters. Short after the OLR shock, VFA accumulation was observed accompanied by a mitigation of methane production. Bacteroidetes, with the family Rikenellaceae, and Firmicutes prevailed at stable conditions and during the shock period followed by Cloacimonetes, candidate division SR1, and Proteobacteria. During the glycerol shock, Erysipelotrichaceae (Firmicutes) and Acholeplasmataceae (Tenericutes) increased in numbers, whereas Chloroflexi, and especially Anaerolineaceae, were reduced. After the OLR overload, Firmicutes continued to increase, mainly owing to a marked enrichment of Anaerosinus (Veillonellaceae family) and Clostridium sensu stricto 1, while Bacteroidetes dropped significantly. Methanogenic community consisted primarily of Methanothrix and Methanoculleus genera together with the candidate genus WCHA1-57 (Ca. Methanofastidiosum). After the OLR shock, there was an instant increase in Methanoculleus and the candidate phylum Woesearchaeota, whose activity is currently unknown. Upon acidification, Methanoculleus prevailed, while Methanothrix was diminished, indicating a shift to hydrogenotrophic methanogenesis. This study showcased the ability of microbial populations in AD to rapidly respond to organic overloading, with certain alterations occurring before changes in performance parameters were apparent [92].
8.2.3. Comparison of HRT versus OLR changes
The microbial community response to alterations in either HRT or OLR has been assessed in several studies. It has been found that changes in these two parameters shaped distinct microbial communities as a result. The most variable phyla were Actinobacteria, Firmicutes and Bacteroidetes, although a core community seemed to be present in all the reactors. The dominant populations initially consisted of Proteobacteria and Actinobacteria, however the latter were greatly diminished at increased OLR, while Firmicutes were favored. Microorganisms regarded as biomarkers for OLR increase included Actinomycetaceae, Ruminococcaceae, and Methanotrichaceae, whereas HRT was linked to increments in the family GZKB119, Pseudomonadaceae, Peptococcaceae and Methanomicrobiaceae. The enhanced degradation of organic matter upon higher OLR led to increased levels of acetic acid, which could explain the higher abundance of the acetoclastic Methanotrichaceae and is consistent with the elevated methane production observed. On the other hand, a shift towards SAO accompanied by hydrogenotrophic methanogenesis has been previously reported at lower HRTs, accounting for the increase in Methanomicrobiaceae. Interestingly, the HRT had a minor impact on the microorganisms, whereas OLR alteration led to a more varied community [83].
Similar conclusions were also drawn by Christou et al., who conducted a bioaugmentation experiment, supplying manure digesters functioning at elevated ammonia with ammonia-adapted microbial inoculum. Successful bioaugmentation was only achieved in the digesters with OLR 3.02 and 4.0 g VS L-1 day-1, along with an increase in methane production, while the one operating at 2.09 g VS L-1 day-1 eventually deteriorated. The fermentative Clostridiaceae, Turicibacter and Romboutsia, as well as the Bacteroidetes genus PeH15, increased upon bioaugmentation. However, Firmicutes members were negatively correlated with OLR increase, while PeH15 had a rising tendency. The syntrophic VFA oxidizers Syntrophomonas (butyrate) and Pelotomaculum (propionate) followed contrary trends, in that Syntrophomonas was favored by bioaugmentation, although it decreased at high OLR, while Pelotomaculum had the opposite fate. Archaeal community at low OLR shifted from the methylotrophic Methanomethylophilaceae to Methanobacteriaceae. In contrast, upon higher OLR, a substantial increase in Methanobacteriaceae and Methanosarcina was observed after bioaugmentation, but methylotrophic methanogenesis was also present, suggesting that all three pathways co-occurred. HRT reduction after bioaugmentation led to a considerable enhancement in Methanobacteriaceae at the higher OLR (4.0 g VS L-1 day-1), while at OLR 3.02 g VS L-1 day-1 Methanosarcina were the most abundant methanogen. However, lowering HRT had little impact on reactor performance, which could be due to successful bioaugmentation in the digesters [84].
9. Functional redundancy
When disruptions occur in an AD system, microbiota depend on three core mechanisms, namely resistance, resilience, and functional redundancy, to sustain their functionality. The greater the biodiversity of microorganisms, the higher the ecosystem solidity. This is attributed to functional redundancy, which refers to different species having overlapping activities, thus helping to offset the adverse impacts that may arise from the elimination of vital yet sensitive to disturbances microorganisms [17,24]. On the other hand, it has been suggested that microbial groups with lower diversity but with complementary pathways, which is referred to as niche complementarity, can also lead to stable performance. This can be explained by the lower direct competition for nutrients observed in these systems, promoting structural stability. Furthermore, a lower number of species may be sufficient for preserving the stability of an ecosystem if one species relies on the function of another for its survival. In other words, a productive ecosystem may require less species diversity, and a greater variety may not entail a better suited community to the AD process [17].
The above are in line with the so-called "insurance hypothesis" proposed by Yachi & Loreau, which states that the necessary species richness for functional redundancy in an ecosystem is determined by the interactions and responses of distinct species to environmental changes. If there is greater diversity in the way species respond, then the number of species required for redundancy could be reduced [93]. It has been proposed that efficient biogas production relies on a combination of increased richness and low evenness of the microbiota, suggesting that both dominant species and less abundant ones are needed to compose a highly performing community that offers robustness to the system [56]. The concept of core microbial populations has driven many researchers into identifying key bacterial and archaeal populations widely present in AD systems that are likely carrying out the most critical ecological functions within their specific environment. It is therefore crucial to outline the core groups in terms of structure and function as to better comprehend the AD process and valorize this knowledge for future management of the digesters to the benefit of methane production [17].
10. Current status
AD, as a type of biomass transformation approach, holds great promise for renewable energy generation owing to its environmentally sustainable characteristics and its potential to convert a wide range of substrates into various energy sources, thus reducing the global carbon footprint [94]. The versatility of AD technology lies not only in its ability to utilize feedstock sources, but also in its application on both small and large scales, which encourage the production of biogas worldwide. It is important, though, that in order for biomethane to be considered a renewable energy source and be adopted in the market, it needs to meet certain criteria outlined in the renewable fuel standard [18]. Overall, AD has a great potential to contribute to sustainable waste management and reduction of greenhouse gas emissions.
It is common knowledge that AD involves various types of microbial metabolic processes. Therefore, it is essential to examine the interrelation among the microbial network dynamics, physicochemical operational parameters and process effectiveness [25]. To improve the management of AD systems ensuring optimal energy recovery, it is crucial to gain a more profound comprehension of the microbial communities within digesters and how they relate to the operational parameters regulating the AD environment. This understanding is essential to enable practical applications that can successfully enhance AD performance [9,27]. Of note, since the efficiency of AD relies on the concerted microbial interplay among hydrolytic and acidogenic bacteria, syntrophic bacteria, and methanogens [19], it is important to take into consideration the different growth attributes and responsiveness that these microorganisms have [24]. For example, methanogenic populations typically exhibit greater susceptibility to fluctuations in AD operating conditions compared to bacteria responsible for hydrolysis and acidogenesis [1].
The functions carried out by the diverse microbial communities involved in AD processes are complex and multifaceted [27] and are influenced by a number of interdependent factors, making it challenging to understand the interactions between process parameters and microbes [22]. Insufficient understanding of these interactions could be considered a barrier to refining AD performance and achieving sustainable biogas production [22]. Moreover, even if two reactors function under comparable parameters, distinct microbial consortia may evolve due to stochastic processes [95]. In order to design impactful techniques to control the behavior of microorganisms in AD, further research is required regarding the underlying principles of microbial plasticity in response to environmental conditions using advanced methodologies, such as metagenomics and computational analyses [1]. This will aid the identification of microbial populations that can either serve as markers for monitoring or be added in the context of bioaugmentation to facilitate the reestablishment of regular AD operation in malfunctioning digesters [10].
11. Future perspectives
Since anaerobic digestion is a widely utilized technology for the conversion of organic waste into renewable energy, there is a significant need for further exploration of the interaction between operational factors and microbial dynamics to obtain a more comprehensive understanding of the AD process. This would ultimately lead to more efficient and sustainable biogas production. The integration and further development of advanced molecular techniques, such as metagenomics, metatranscriptomics, and metaproteomics, is critical for a more accurate characterization of microbial communities in AD systems. Additionally, there is a need to explore the long-term effects of operational factors on the stability and resilience of microbial communities in AD digesters. Moreover, research can be expanded to evaluate the potential use of probiotics or other microbial additives to enhance process efficiency and stability in the face of various environmental stresses. Ultimately, by improving our understanding of the complex interplay between operational factors and microbial communities, we can work towards optimizing AD processes for more sustainable and efficient waste management.
12. Conclusion
In conclusion, the study of the microbial dynamics in anaerobic digesters is a complex and ever-evolving field, influenced by a range of operational factors. Understanding these factors and their effects on the microbiome is essential for optimizing biogas production and improving the efficiency of anaerobic digestion. Further research is needed to fully understand the interactions between these factors and microbial dynamics, and to develop strategies for optimizing anaerobic digestion processes.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Figure 1: Microbial groups found highly abundant in anaerobic digesters with different types of feedstocks. FW, food waste; WAS, wastewater sludge; CW, crop waste; CO-D, co-digestion; fish, fishery waste; SM, swine manure; MSW, municipal solid waste; CM, cow manure. Supplementary Figure 2: Microbial taxa commonly found in anaerobic digestion reactors at different temperature ranges.
Author Contributions
Conceptualization, T.S. and A.G.C.; investigation, E.K.; data curation, E.K. and A.G.C.; writing—original draft preparation, E.K.; writing—review and editing, T.S., V.T. and A.G.C.; visualization, A.G.C.; supervision, T.S. and V.T.; project administration, T.S. and V.T.; funding acquisition, T.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was carried out as part of the project «μ2GAS» (Project code: ΚΜΡ6-0143562) under the framework of the Action «Investment Plans of Innovation» of the Operational Program «Central Macedonia 2014-2020», that is co-funded by the European Regional Development Fund and Greece.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare that there are no competing interests concerning the publication of this manuscript.
References
- Xu, R.Z.; Fang, S.Y.; Zhang, L.; Huang, W.X.; Shao, Q.Q.; Fang, F.; Feng, Q.; Cao, J.S.; Luo, J.Y. Distribution patterns of functional microbial community in anaerobic digesters under different operational circumstances: A review. Bioresource Technology 2021, 341. [Google Scholar] [CrossRef] [PubMed]
- Westerholm, M.; Muller, B.; Isaksson, S.; Schnurer, A. Trace element and temperature effects on microbial communities and links to biogas digester performance at high ammonia levels. Biotechnology for Biofuels 2015, 8. [Google Scholar] [CrossRef] [PubMed]
- Fontana, A.; Patrone, V.; Puglisi, E.; Morelli, L.; Bassi, D.; Garuti, M.; Rossi, L.; Cappa, F. Effects of geographic area, feedstock, temperature, and operating time on microbial communities of six full-scale biogas plants. Bioresource Technology 2016, 218, 980–990. [Google Scholar] [CrossRef]
- Panigrahi, S.; Dubey, B.K. A critical review on operating parameters and strategies to improve the biogas yield from anaerobic digestion of organic fraction of municipal solid waste. Renewable Energy 2019, 143, 779–797. [Google Scholar] [CrossRef]
- Ziganshina, E.E.; Belostotskiy, D.E.; Ilinskaya, O.N.; Boulygina, E.A.; Grigoryeva, T.V.; Ziganshin, A.M. Effect of the Organic Loading Rate Increase and the Presence of Zeolite on Microbial Community Composition and Process Stability During Anaerobic Digestion of Chicken Wastes. Microbial Ecology 2015, 70, 948–960. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.K.; Lee, S.H.; Kim, Y.; Park, H.D. Current understanding and perspectives in anaerobic digestion based on genome-resolved metagenomic approaches. Bioresource Technology 2022, 344. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.P.; Wu, X.Y.; Zhou, B.Q.; Wang, Y.; Sun, Y.; Wang, Y.F.; Chen, Z.B.; Zhang, J.F. Effect of one step temperature increment from mesophilic to thermophilic anaerobic digestion on the linked pattern between bacterial and methanogenic communities. Bioresource Technology 2019, 292. [Google Scholar] [CrossRef]
- Gaby, J.C.; Zamanzadeh, M.; Horn, S.J. The effect of temperature and retention time on methane production and microbial community composition in staged anaerobic digesters fed with food waste. Biotechnology for Biofuels 2017, 10. [Google Scholar] [CrossRef]
- de Jonge, N.; Davidsson, A.; Jansen, J.L.; Nielsen, J.L. Characterisation of microbial communities for improved management of anaerobic digestion of food waste. Waste Management 2020, 117, 124–135. [Google Scholar] [CrossRef]
- Lim, J.W.; Park, T.; Tong, Y.W.; Yu, Z. Chapter One - The microbiome driving anaerobic digestion and microbial analysis. In Advances in Bioenergy; Li, Y., Khanal, S.K., Eds.; Elsevier, 2020; Volume 5, pp. 1–61. [Google Scholar]
- Langer, S.G.; Gabris, C.; Einfalt, D.; Wemheuer, B.; Kazda, M.; Bengelsdorf, F.R. Different response of bacteria, archaea and fungi to process parameters in nine full-scale anaerobic digesters. Microbial Biotechnology 2019, 12, 1210–1225. [Google Scholar] [CrossRef]
- Carballa, M.; Regueiro, L.; Lema, J.M. Microbial management of anaerobic digestion: exploiting the microbiome-functionality nexus. Current Opinion in Biotechnology 2015, 33, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Fenchel, T.; King, G.M.; Blackburn, T.H. Chapter 1 - Bacterial Metabolism. In Bacterial Biogeochemistry (Third Edition); Fenchel, T., King, G.M., Blackburn, T.H., Eds.; Academic Press: Boston, 2012; pp. 1–34. [Google Scholar]
- Sarmiento, B.F.; Leigh, J.A.; Whitrnan, W.B. GENETIC SYSTEMS FOR HYDROGENOTROPHIC METHANOGENS. Methods in Enzymology: Methods in Methane Metabolism, Pt A 2011, 494, 43–73. [Google Scholar] [CrossRef]
- Han, G.; Shin, S.G.; Cho, K.; Lee, J.; Kim, W.; Hwang, S. Temporal variation in bacterial and methanogenic communities of three full-scale anaerobic digesters treating swine wastewater. Environmental Science and Pollution Research 2019, 26, 1217–1226. [Google Scholar] [CrossRef] [PubMed]
- Zielinska, M.; Cydzik-Kwiatkowska, A.; Zielinski, M.; Debowski, M. Impact of temperature, microwave radiation and organic loading rate on methanogenic community and biogas production during fermentation of dairy wastewater. Bioresource Technology 2013, 129, 308–314. [Google Scholar] [CrossRef]
- Calusinska, M.; Goux, X.; Fossepre, M.; Muller, E.E.L.; Wilmes, P.; Delfosse, P. A year of monitoring 20 mesophilic full-scale bioreactors reveals the existence of stable but different core microbiomes in bio-waste and wastewater anaerobic digestion systems. Biotechnology for Biofuels 2018, 11. [Google Scholar] [CrossRef]
- Achinas, S.; Achinas, V.; Euverink, G.J.W. A Technological Overview of Biogas Production from Biowaste. Engineering 2017, 3, 299–307. [Google Scholar] [CrossRef]
- Saha, S.; Basak, B.; Hwang, J.H.; Salama, E.; Chatterjee, P.K.; Jeon, B.H. Microbial Symbiosis: A network towards Biomethanation. Trends in Microbiology 2020, 28, 968–984. [Google Scholar] [CrossRef]
- Puig-Castellvi, F.; Cardona, L.; Bouveresse, D.J.R.; Cordella, C.B.Y.; Mazeas, L.; Rutledge, D.N.; Chapleur, O. Assessment of the microbial interplay during anaerobic co-digestion of wastewater sludge using common components analysis. Plos One 2020, 15. [Google Scholar] [CrossRef]
- Yun, Y.M.; Cho, S.K.; Kim, H.W.; Jung, K.W.; Shin, H.S.; Kim, D.H. Elucidating a synergistic effect of food waste addition on the enhanced anaerobic digestion of waste activated sludge. Korean Journal of Chemical Engineering 2015, 32, 1542–1546. [Google Scholar] [CrossRef]
- Shin, S.G.; Han, G.; Lee, J.; Shin, J.; Hwang, S. A snapshot of microbial community structures in 20 different field-scale anaerobic bioreactors treating food waste. Journal of Environmental Management 2019, 248. [Google Scholar] [CrossRef]
- Lee, J.; Kim, E.; Han, G.; Tongco, J.V.; Shin, S.G.; Hwang, S. Microbial communities underpinning mesophilic anaerobic digesters treating food wastewater or sewage sludge: A full-scale study. Bioresource Technology 2018, 259, 388–397. [Google Scholar] [CrossRef]
- Lee, J.; Shin, S.G.; Han, G.; Koo, T.; Hwang, S. Bacteria and archaea communities in full-scale thermophilic and mesophilic anaerobic digesters treating food wastewater: Key process parameters and microbial indicators of process instability. Bioresource Technology 2017, 245, 689–697. [Google Scholar] [CrossRef]
- Sposob, M.; Moon, H.S.; Lee, D.; Kim, T.H.; Yun, Y.M. Comprehensive analysis of the microbial communities and operational parameters of two full-scale anaerobic digestion plants treating food waste in South Korea: Seasonal variation and effect of ammonia. Journal of Hazardous Materials 2020, 398. [Google Scholar] [CrossRef]
- Montanes, R.; Solera, R.; Perez, M. Anaerobic co-digestion of sewage sludge and sugar beet pulp lixiviation in batch reactors: Effect of temperature. Bioresource Technology 2015, 180, 177–184. [Google Scholar] [CrossRef]
- Fitamo, T.; Treu, L.; Boldrin, A.; Sartori, C.; Angelidaki, I.; Scheutz, C. Microbial population dynamics in urban organic waste anaerobic co-digestion with mixed sludge during a change in feedstock composition and different hydraulic retention times. Water Research 2017, 118, 261–271. [Google Scholar] [CrossRef]
- Lin, Q.; De Vrieze, J.; Fang, X.Y.; Li, L.J.; Li, X.Z. Labile carbon feedstocks trigger a priming effect in anaerobic digestion: An insight into microbial mechanisms. Bioresource Technology 2022, 344. [Google Scholar] [CrossRef]
- Kalamaras, S.D.; Vasileiadis, S.; Karas, P.; Angelidaki, I.; Kotsopoulos, T.A. Microbial adaptation to high ammonia concentrations during anaerobic digestion of manure-based feedstock: biomethanation and 16S rRNA gene sequencing. Journal of Chemical Technology and Biotechnology 2020, 95, 1970–1979. [Google Scholar] [CrossRef]
- Pereira, M.A.; Sousa, D.Z.; Mota, M.; Alves, M.M. Mineralization of LCFA associated with anaerobic sludge: Kinetics, enhancement of methanogenic activity, and effect of VFA. Biotechnology and Bioengineering 2004, 88, 502–511. [Google Scholar] [CrossRef]
- Kougias, P.G.; Campanaro, S.; Treu, L.; Zhu, X.Y.; Angelidaki, I. A novel archaeal species belonging to Methanoculleus genus identified via de-novo assembly and metagenomic binning process in biogas reactors. Anaerobe 2017, 46, 23–32. [Google Scholar] [CrossRef]
- Niu, Q.; Kobayashi, T.; Takemura, Y.; Kubota, K.; Li, Y.Y. Evaluation of functional microbial community's difference in full-scale and lab-scale anaerobic digesters feeding with different organic solid waste: Effects of substrate and operation factors. Bioresource Technology 2015, 193, 110–118. [Google Scholar] [CrossRef]
- Yi, J.; Dong, B.; Jin, J.W.; Dai, X.H. Effect of Increasing Total Solids Contents on Anaerobic Digestion of Food Waste under Mesophilic Conditions: Performance and Microbial Characteristics Analysis. Plos One 2014, 9. [Google Scholar] [CrossRef]
- Ziganshin, A.M.; Ziganshina, E.E.; Kleinsteuber, S.; Nikolausz, M. Comparative Analysis of Methanogenic Communities in Different Laboratory-Scale Anaerobic Digesters. Archaea-an International Microbiological Journal 2016, 2016. [Google Scholar] [CrossRef]
- Peces, M.; Astals, S.; Jensen, P.D.; Clarke, W.P. Deterministic mechanisms define the long-term anaerobic digestion microbiome and its functionality regardless of the initial microbial community. Water Research 2018, 141, 366–376. [Google Scholar] [CrossRef]
- Duan, N.; Kougias, P.G.; Campanaro, S.; Treu, L.; Angelidaki, I. Evolution of the microbial community structure in biogas reactors inoculated with seeds from different origin. Science of The Total Environment 2021, 773, 144981. [Google Scholar] [CrossRef]
- Demichelis, F.; Tommasi, T.; Deorsola, F.A.; Marchisio, D.; Fino, D. Effect of inoculum origin and substrate-inoculum ratio to enhance the anaerobic digestion of organic fraction municipal solid waste (OFMSW). Journal of Cleaner Production 2022, 351. [Google Scholar] [CrossRef]
- Quintero, M.; Castro, L.; Ortiz, C.; Guzman, C.; Escalante, H. Enhancement of starting up anaerobic digestion of lignocellulosic substrate: fique's bagasse as an example. Bioresource Technology 2012, 108, 8–13. [Google Scholar] [CrossRef]
- Bella, K.; Rao, P.V. Anaerobic co-digestion of cheese whey and septage: Effect of substrate and inoculum on biogas production. Journal of Environmental Management 2022, 308. [Google Scholar] [CrossRef]
- Li, Y.Y.; Wang, Y.Q.; Yu, Z.H.; Lu, J.X.; Li, D.Y.; Wang, G.Y.; Li, Y.; Wu, Y.; Li, S.Y.; Xu, F.Q.; et al. Effect of inoculum and substrate/inoculum ratio on the performance and methanogenic archaeal community structure in solid state anaerobic co-digestion of tomato residues with dairy manure and corn stover. Waste Management 2018, 81, 117–127. [Google Scholar] [CrossRef]
- Bao, R.; Wei, Y.F.; Guan, R.L.; Li, X.J.; Lu, X.B.; Rong, S.Y.; Zuo, X.Y.; Yuan, H.R. High-solids anaerobic co-digestion performances and microbial community dynamics in co-digestion of different mixing ratios with food waste and highland barley straw. Energy 2023, 262. [Google Scholar] [CrossRef]
- Liu, C.; Li, H.; Zhang, Y.Y.; Si, D.D.; Chen, Q.W. Evolution of microbial community along with increasing solid concentration during high-solids anaerobic digestion of sewage sludge. Bioresource Technology 2016, 216, 87–94. [Google Scholar] [CrossRef]
- Wang, Z.Z.; Jiang, Y.; Wang, S.; Zhang, Y.Z.; Hu, Y.S.; Hu, Z.H.; Wu, G.X.; Zhan, X.M. Impact of total solids content on anaerobic co-digestion of pig manure and food waste: Insights into shifting of the methanogenic pathway. Waste Management 2020, 114, 96–106. [Google Scholar] [CrossRef]
- Abbassi-Guendouz, A.; Brockmann, D.; Trably, E.; Dumas, C.; Delgenes, J.P.; Steyer, J.P.; Escudie, R. Total solids content drives high solid anaerobic digestion via mass transfer limitation. Bioresource Technology 2012, 111, 55–61. [Google Scholar] [CrossRef]
- Liu, C.; Li, H.; Zhang, Y.Y.; Chen, Q.W. Characterization of methanogenic activity during high-solids anaerobic digestion of sewage sludge. Biochemical Engineering Journal 2016, 109, 96–100. [Google Scholar] [CrossRef]
- Liao, X.C.; Li, H. Biogas production from low-organic-content sludge using a high-solids anaerobic digester with improved agitation. Applied Energy 2015, 148, 252–259. [Google Scholar] [CrossRef]
- Duan, N.N.; Dong, B.; Wu, B.; Dai, X.H. High-solid anaerobic digestion of sewage sludge under mesophilic conditions: Feasibility study. Bioresource Technology 2012, 104, 150–156. [Google Scholar] [CrossRef]
- Arelli, V.; Begum, S.; Anupoju, G.R.; Kuruti, K.; Shailaja, S. Dry anaerobic co-digestion of food waste and cattle manure: Impact of total solids, substrate ratio and thermal pre treatment on methane yield and quality of biomanure. Bioresource Technology 2018, 253, 273–280. [Google Scholar] [CrossRef]
- Han, G.; Shin, S.G.; Lee, J.; Shin, J.; Hwang, S. A comparative study on the process efficiencies and microbial community structures of six full-scale wet and semi-dry anaerobic digesters treating food wastes. Bioresource Technology 2017, 245, 869–875. [Google Scholar] [CrossRef]
- Choi, G.; Kim, H.; Lee, C. Long-term monitoring of a thermal hydrolysis-anaerobic co-digestion plant treating high-strength organic wastes: Process performance and microbial community dynamics. Bioresource Technology 2021, 319. [Google Scholar] [CrossRef]
- Duc, L.V.; Miyagawa, Y.; Inoue, D.; Ike, M. Identification of key steps and associated microbial populations for efficient anaerobic digestion under high ammonium or salinity conditions. Bioresource Technology 2022, 360. [Google Scholar] [CrossRef]
- Su, L.H.; Sun, X.; Liu, C.W.; Ji, R.T.; Zhen, G.Y.; Chen, M.; Zhang, L.J. Thermophilic Solid-State Anaerobic Digestion of Corn Straw, Cattle Manure, and Vegetable Waste: Effect of Temperature, Total Solid Content, and C/N Ratio. Archaea-an International Microbiological Journal 2020, 2020. [Google Scholar] [CrossRef]
- Fotidis, I.A.; Karakashev, D.; Kotsopoulos, T.A.; Martzopoulos, G.G.; Angelidaki, I. Effect of ammonium and acetate on methanogenic pathway and methanogenic community composition. Fems Microbiology Ecology 2013, 83, 38–48. [Google Scholar] [CrossRef]
- Zhang, H.; Yuan, W.D.; Dong, Q.; Wu, D.; Yang, P.J.; Peng, Y.; Li, L.; Peng, X.Y. Integrated multi-omics analyses reveal the key microbial phylotypes affecting anaerobic digestion performance under ammonia stress. Water Research 2022, 213. [Google Scholar] [CrossRef]
- Westerholm, M.; Calusinska, M.; Dolfing, J. Syntrophic propionate-oxidizing bacteria in methanogenic systems. Fems Microbiology Reviews 2022, 46. [Google Scholar] [CrossRef]
- Heyer, R.; Benndorf, D.; Kohrs, F.; De Vrieze, J.; Boon, N.; Hoffmann, M.; Rapp, E.; Schluter, A.; Sczyrba, A.; Reichl, U. Proteotyping of biogas plant microbiomes separates biogas plants according to process temperature and reactor type. Biotechnology for Biofuels 2016, 9. [Google Scholar] [CrossRef]
- Ziganshin, A.M.; Liebetrau, J.; Proter, J.; Kleinsteuber, S. Microbial community structure and dynamics during anaerobic digestion of various agricultural waste materials. Applied Microbiology and Biotechnology 2013, 97, 5161–5174. [Google Scholar] [CrossRef]
- Bardi, M.J.; Aminirad, H. Synergistic effects of co-trace elements on anaerobic co-digestion of food waste and sewage sludge at high organic load. Environmental Science and Pollution Research 2020, 27, 18129–18144. [Google Scholar] [CrossRef]
- Shaw, G.T.W.; Liu, A.C.; Weng, C.Y.; Chou, C.Y.; Wang, D.Y. Inferring microbial interactions in thermophilic and mesophilic anaerobic digestion of hog waste. Plos One 2017, 12. [Google Scholar] [CrossRef]
- Moestedt, J.; Ronnberg, J.; Nordell, E. The effect of different mesophilic temperatures during anaerobic digestion of sludge on the overall performance of a WWTP in Sweden. Water Science and Technology 2017, 76, 3213–3219. [Google Scholar] [CrossRef]
- Kabaivanova, L.; Petrova, P.; Hubenov, V.; Simeonov, I. Biogas Production Potential of Thermophilic Anaerobic Biodegradation of Organic Waste by a Microbial Consortium Identified with Metagenomics. Life-Basel 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Tukacs-Hajos, A.; Pap, B.; Maroti, G.; Szendefy, J.; Szabo, P.; Retfalvi, T. Monitoring of thermophilic adaptation of mesophilic anaerobe fermentation of sugar beet pressed pulp. Bioresource Technology 2014, 166, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Angelidaki, I.; Sanders, W. Assessment of the anaerobic biodegradability of macropollutants. Re/Views in Environmental Science & Bio/Technology 2004, 3, 117–129. [Google Scholar] [CrossRef]
- Guo, X.H.; Wang, C.; Sun, F.Q.; Zhu, W.J.; Wu, W.X. A comparison of microbial characteristics between the thermophilic and mesophilic anaerobic digesters exposed to elevated food waste loadings. Bioresource Technology 2014, 152, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Goux, X.; Calusinska, M.; Lemaigre, S.; Marynowska, M.; Klocke, M.; Udelhoven, T.; Benizri, E.; Delfosse, P. Microbial community dynamics in replicate anaerobic digesters exposed sequentially to increasing organic loading rate, acidosis, and process recovery. Biotechnology for Biofuels 2015, 8. [Google Scholar] [CrossRef]
- Shaw, G.T.W.; Weng, C.Y.; Chen, C.Y.; Weng, F.C.H.; Wang, D.Y. A systematic approach re-analyzing the effects of temperature disturbance on the microbial community of mesophilic anaerobic digestion. Scientific Reports 2019, 9. [Google Scholar] [CrossRef]
- Liu, J.; Zuo, X.Y.; Peng, K.; He, R.; Yang, L.Y.; Liu, R.F. Biogas and Volatile Fatty Acid Production During Anaerobic Digestion of Straw, Cellulose, and Hemicellulose with Analysis of Microbial Communities and Functions. Applied Biochemistry and Biotechnology 2022, 194, 762–782. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, C.X.; Yuan, Z.W.; Wang, R.M.; Angelidaki, I.; Zhu, G.F. Syntrophy mechanism, microbial population, and process optimization for volatile fatty acids metabolism in anaerobic digestion. Chemical Engineering Journal 2023, 452. [Google Scholar] [CrossRef]
- Franke-Whittle, I.H.; Walter, A.; Ebner, C.; Insam, H. Investigation into the effect of high concentrations of volatile fatty acids in anaerobic digestion on methanogenic communities. Waste Management 2014, 34, 2080–2089. [Google Scholar] [CrossRef]
- Mathai, P.P.; Nicholes, M.S.; Venkiteshwaran, K.; Brown, C.M.; Morris, R.L.; Zitomer, D.H.; Maki, J.S. Dynamic shifts within volatile fatty acid-degrading microbial communities indicate process imbalance in anaerobic digesters. Applied Microbiology and Biotechnology 2020, 104, 4563–4575. [Google Scholar] [CrossRef]
- Nguyen, D.; Wu, Z.Y.; Shrestha, S.; Lee, P.H.; Raskin, L.; Khanal, S.K. Intermittent micro-aeration: New strategy to control volatile fatty acid accumulation in high organic loading anaerobic digestion. Water Research 2019, 166. [Google Scholar] [CrossRef]
- Hori, T.; Haruta, S.; Ueno, Y.; Ishii, M.; Igarashi, Y. Dynamic transition of a methanogenic population in response to the concentration of volatile fatty acids in a thermophilic anaerobic digester. Applied and Environmental Microbiology 2006, 72, 1623–1630. [Google Scholar] [CrossRef]
- Wagner, A.O.; Malin, C.; Lins, P.; Illmer, P. Effects of various fatty acid amendments on a microbial digester community in batch culture. Waste Management 2011, 31, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.F.; Hu, K.; Zheng, Z.H.; Zhang, Y.; Guo, S.Y.; Zhao, X.L.; Cui, Z.J.; Wang, X.F. Effects of adding EDTA and Fe2+ on the performance of reactor and microbial community structure in two simulated phases of anaerobic digestion. Bioresource Technology 2019, 275, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Venkiteshwaran, K.; Bocher, B.; Maki, J.; Zitomer, D. Relating Anaerobic Digestion Microbial Community and Process Function : Supplementary Issue: Water Microbiology. Microbiology Insights 2015, 8s2, MBI–S33593. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.G.; Yang, X.; Yao, X.F. Effects of pH on the biodegradation characteristics of thermophilic micro-aerobic digestion for sludge stabilization. Rsc Advances 2019, 9, 8379–8388. [Google Scholar] [CrossRef]
- Zhou, X.N.; Lu, Y.; Huang, L.; Zhang, Q.; Wang, X.Y.; Zhu, J.Y. Effect of pH on volatile fatty acid production and the microbial community during anaerobic digestion of Chinese cabbage waste. Bioresource Technology 2021, 336. [Google Scholar] [CrossRef] [PubMed]
- Castro-Ramos, J.J.; Solis-Oba, A.; Solis-Oba, M.; Calderon-Vazquez, C.L.; Higuera-Rubio, J.M.; Castro-Rivera, R. Effect of the initial pH on the anaerobic digestion process of dairy cattle manure. Amb Express 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zhang, R.; Liu, F.W.; Yong, X.Y.; Wu, X.Y.; Zheng, T.; Jiang, M.; Jia, H.H. Biogas production and microbial community shift through neutral pH control during the anaerobic digestion of pig manure. Bioresource Technology 2016, 217, 44–49. [Google Scholar] [CrossRef]
- Peces, M.; Astals, S.; Jensen, P.D.; Clarke, W.P. Transition of microbial communities and degradation pathways in anaerobic digestion at decreasing retention time. New Biotechnology 2021, 60, 52–61. [Google Scholar] [CrossRef]
- Tena, M.; Perez, M.; Solera, R. Effect of hydraulic retention time on the methanogenic step of a two-stage anaerobic digestion system from sewage sludge and wine vinasse: Microbial and kinetic evaluation. Fuel 2021, 296. [Google Scholar] [CrossRef]
- Shi, X.S.; Dong, J.J.; Yu, J.H.; Yin, H.; Hu, S.M.; Huang, S.X.; Yuan, X.Z. Effect of Hydraulic Retention Time on Anaerobic Digestion of Wheat Straw in the Semicontinuous Continuous Stirred-Tank Reactors. Biomed Research International 2017, 2017. [Google Scholar] [CrossRef]
- Xu, R.; Yang, Z.H.; Zheng, Y.; Liu, J.B.; Xiong, W.P.; Zhang, Y.R.; Lu, Y.; Xue, W.J.; Fan, C.Z. Organic loading rate and hydraulic retention time shape distinct ecological networks of anaerobic digestion related microbiome. Bioresource Technology 2018, 262, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Christou, M.L.; Vasileiadis, S.; Karpouzas, D.G.; Angelidaki, I.; Kotsopoulos, T.A. Effects of organic loading rate and hydraulic retention time on bioaugmentation performance to tackle ammonia inhibition in anaerobic digestion. Bioresource Technology 2021, 334. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Meng, X.S.; Zhou, G.N.; Zhou, Z.Z.; Zheng, T.; Bai, Y.E.; Yuan, H.R.; Huhe, T. Effects of organic loading rates on the anaerobic co-digestion of fresh vinegar residue and pig manure: Focus on the performance and microbial communities. Biochemical Engineering Journal 2022, 183. [Google Scholar] [CrossRef]
- Yang, S.M.; Luo, F.; Yan, J.; Zhang, T.L.; Xian, Z.Y.; Huang, W.Y.; Zhang, H.G.; Cao, Y.J.; Huang, L. Biogas production of food waste with in-situ sulfide control under high organic loading in two-stage anaerobic digestion process: Strategy and response of microbial community. Bioresource Technology 2023, 373. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Jiao, P.; Zhang, M.; Wu, P.; Zhang, Y.; Wang, Y.; Xu, K.; Yu, J.; Ma, L. Impacts of organic loading rate and hydraulic retention time on organics degradation, interspecies interactions and functional traits in thermophilic anaerobic co-digestion of food waste and sewage sludge. Bioresource Technology 2023, 370, 128578. [Google Scholar] [CrossRef] [PubMed]
- Parajuli, A.; Khadka, A.; Sapkota, L.; Ghimire, A. Effect of Hydraulic Retention Time and Organic-Loading Rate on Two-Staged, Semi-Continuous Mesophilic Anaerobic Digestion of Food Waste during Start-Up. Fermentation-Basel 2022, 8. [Google Scholar] [CrossRef]
- Pramanik, S.K.; Suja, F.B.; Pramanik, B.K. Effects of hydraulic retention time on the process performance and microbial community structure of an anaerobic single-stage semi-pilot scale reactor for the treatment of food waste. International Biodeterioration & Biodegradation 2020, 152. [Google Scholar] [CrossRef]
- Razaviarani, V.; Buchanan, I.D. Reactor performance and microbial community dynamics during anaerobic co-digestion of municipal wastewater sludge with restaurant grease waste at steady state and overloading stages. Bioresource Technology 2014, 172, 232–240. [Google Scholar] [CrossRef]
- Wu, Z.L.; Zhang, Q.; Xia, Z.Y.; Gou, M.; Sun, Z.Y.; Tang, Y.Q. The responses of mesophilic and thermophilic anaerobic digestion of municipal sludge to periodic fluctuation disturbance of organic loading rate. Environmental Research 2023, 218. [Google Scholar] [CrossRef]
- Braz, G.H.R.; Fernandez-Gonzalez, N.; Lema, J.M.; Carballa, M. The time response of anaerobic digestion microbiome during an organic loading rate shock. Applied Microbiology and Biotechnology 2018, 102, 10285–10297. [Google Scholar] [CrossRef]
- Yachi, S.; Loreau, M. Biodiversity and ecosystem productivity in a fluctuating environment: The insurance hypothesis. Proceedings of the National Academy of Sciences of the United States of America 1999, 96, 1463–1468. [Google Scholar] [CrossRef] [PubMed]
- Roopnarain, A.; Rama, H.; Ndaba, B.; Bello-Akinosho, M.; Bamuza-Pemu, E.; Adeleke, R. Unravelling the anaerobic digestion 'black box': Biotechnological approaches for process optimization. Renewable & Sustainable Energy Reviews 2021, 152. [Google Scholar] [CrossRef]
- Lv, Z.; Leite, A.F.; Harms, H.; Richnow, H.H.; Liebetrau, J.; Nikolausz, M. Influences of the substrate feeding regime on methanogenic activity in biogas reactors approached by molecular and stable isotope methods. Anaerobe 2014, 29, 91–99. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Key bacterial and archaeal groups engaging in different methanogenesis stages within anaerobic digestion reactors. VFA, Volatile Fatty Acid; SAOB, Syntrophic Acetate-oxidizing Bacteria.
Figure 1.
Key bacterial and archaeal groups engaging in different methanogenesis stages within anaerobic digestion reactors. VFA, Volatile Fatty Acid; SAOB, Syntrophic Acetate-oxidizing Bacteria.
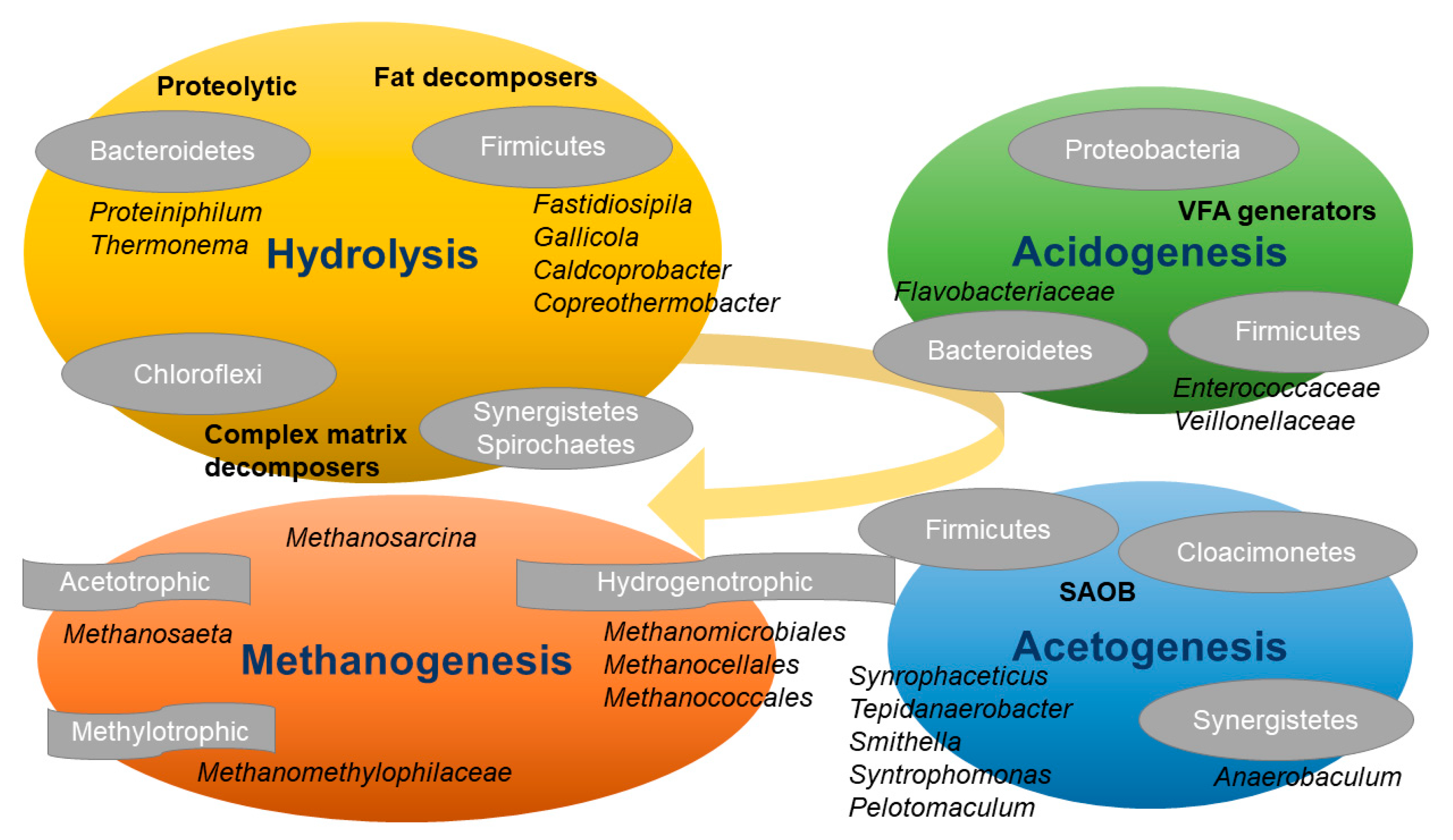

Table 1.
Favored microbial communities at digesters with different TS content.
| TS content | Biogas Yield | Favored | |
|---|---|---|---|
| Wet AD | ≤10% | Increased [45] |
Porphyromonadaceae Sphingobacteriaceae Syntrophomonadaceae [49] |
| HSAD | 10-20% | Declined [44] Increased [33,42] |
Clostridiaceae Patulibacteraceae Pseudonocardiaceae Lachnospiraceae Rikenellaceae Methanobacteriaceae [49] |
| Dry AD | >20% | Increased [41,48] Declined [43] |
Proteiniphilum Fastidiosipila, Gallicola Aminobacterium Syntrophaceticus W5053 [41] |
Table 2.
Microbial communities favored at different ammonia-rich digesters. SPOB, Syntrophic Propionate Oxidizing Bacteria; SAOB Syntrophic Acetate Oxidizing Bacteria.
Table 2.
Microbial communities favored at different ammonia-rich digesters. SPOB, Syntrophic Propionate Oxidizing Bacteria; SAOB Syntrophic Acetate Oxidizing Bacteria.
| Digester | Microbial communities favored | Role | Reference |
|---|---|---|---|
| Mesophilic sludge digesters |
Ca. Brevefilum (Chloroflexi) Ca. Cloacimonas Syntrophorhabdus |
SPOB | [51] |
|
Paraclostridium Enterococcus Romboutsia Proteiniphilum Turicibacter |
hydrogen-producing | [51] | |
| Methanosarcina | acetoclastic and hydrogenotrophic methanogenesis | [9,33,51] | |
| Mesophilic reactors co-digesting household waste and albumin |
Clostridium ultunense Syntrophaceticus schinkii Tepidanaerobacter acetatoxydans |
SAOB | [2,41] |
| Mesophilic reactors with chicken waste |
Marinilabiaceae Porphyromonadaceae |
SAOB | [5] |
| Methanosarcina | cooperating with Clostridium for hydrogenotrophic methanogenesis | [5,56] | |
| Mesophilic digesters fed with sewage sludge and FW | Methanoculleus receptaculi | [23] | |
| Cloacimonadales W27 | syntrophic partner to hydrogenotrophic methanogens | [23] | |
|
Caldicoprobacter algeriensis Clostridium senegalense Ercella succinigene Gelria glutamica |
[25] | ||
|
Gelria glutamica Defluviitoga tunisiensis |
positively correlated with high NH4+ | [25] | |
| Batch cultures and manure-based substrate |
Cellulosilyticum ruminicola (Cellulosilyticum) Alkaliflexus imshenetskii (Ruminofilibacter) |
syntrophic associations | [29] |
| Hydrogenispora ethanolica Acetomicrobium hydrogeniformans | [29] | ||
| Methanosarcina flavescens | [29] |
Table 3.
Microbial communities favored at differed pH ranges digesters.
| pH | Microbial communities favoured | Reference |
|---|---|---|
| Acidic | Proteobacteria, Aminicenantes, Actinobacteria, and Nitrospirae | [52] |
| Bacteroidetes | [77] | |
|
Clostridium IV, Clostridium sensu stricto, unclassified_Ruminococcaceae |
[77] | |
| Caproiciproducens | [78] | |
| Turicibacter | [78] | |
| Methanosarcina | [79] | |
| Neutral | Sphaerochaeta (Spirochaetes) | [79] |
| Basic | Petrimonas and Proteiniphilum | [24] |
| Tissierella and Tepidimicrobium | [76] | |
| Porphyromonadaceae | [79] | |
| Solibacillus silvestris | [79] | |
| Methanosarcina | [79] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



