
Submitted:
05 June 2023
Posted:
06 June 2023
You are already at the latest version
Abstract
Chicken infectious anemia (CIA) poses a significant threat to the chicken industry in China. De-spite its non-specific symptoms, the disease is often overlooked. This study aimed to conduct a comprehensive analysis of the etiology and pathology of CIA in Guangxi Province, China. Three strains of the chicken infectious anemia virus (CIAV) were isolated from liver samples of diseased 20-week-old chickens. The complete genomes of these strains were sequenced, and experiments on specific pathogen-free (SPF) chicks revealed that the GX21121 strain exhibited high virulence. Histopathological examination of the deceased chicks showed liver cell necrosis, fibrous-serous exudation, inflammatory cell infiltration, hemorrhage in liver tissues, as well as congestion in lung and renal tissues. Phylogenetic analysis of the genome revealed that the three strains had a close genetic relationship to the Heilongjiang wild-type (GenBank KY486144). The genetic evolu-tion of their VP1 genes indicated that all three CIAV isolates belonged to genotype IIIc. In sum-mary, this study demonstrated the genomic diversity of three CIAV strains in adult layer hens. The isolation and characterization of the GX21121 strain as a highly virulent isolate provide val-uable information for further investigations into the etiology, molecular epidemiology, and viral evolution of CIAV.
Keywords:
Chicken infectious anemia virus
; isolation and identification
; pathogenicity
; whole genome
; bioinformatics analysis
1. Introduction
Chicken infectious anemia (CIA) is a viral disease caused by the chicken infectious anemia virus (CIAV) and primarily affects young chicks. It is characterized by aplastic anemia, yellow staining of bone marrow, lymphoid tissue atrophy, and compromised immune function [1,2]. CIAV is a non-enveloped, icosahedral symmetrical virus particle belonging to the Anelloviridae family, with a diameter ranging from 25 nm to 26.5 nm and a genome size of approximately 2.3 kb [3,4,5]. The first isolation of CIAV was reported in Japan in 1970 from a contaminated vaccine [6]. Although chickens are the natural host of CIAV, the virus has also been detected in the feces of humans, mice, dogs, and other birds [7,8,9]. Currently, CIA has been reported in major poultry-raising countries worldwide. The disease primarily affects chicks between 1 and 3 weeks of age [10,11], while adult chickens generally experience subclinical infections when infected with CIAV. Transmission of CIAV occurs through vertical and horizontal routes, such as oral-fecal contamination, and occasionally through contaminated vaccines [12,13]. CIAV can propagate in chicken embryos and lymphoblastoid cell lines but not in chicken embryo fibroblasts, chicken kidney cells, or other primary cells [14]. The MDCC-MSB1 cell line has been widely employed for CIAV isolation in previous studies [15]. However, different CIAV isolates exhibit varying sensitivities and replication rates on MDCC-MSB1 cell lines [16].
The genome of CIAV encodes three viral proteins: the capsid protein (VP1, 51.6 kDa), the viral backbone protein (VP2, 24 kDa), and the apoptosis-inducing protein (VP3, 13.6 kDa) [17,18]. VP1 functions as a structural protein that, in conjunction with VP2, generates a neutralizing epitope. However, research has also shown that VP1 itself contains a neutralization epitope, although variations exist among mutant strains, and the underlying mechanisms are yet to be fully explained [19,20,21]. Acting as the backbone protein, VP2 performs multiple roles in the virus replication process [22,23], while VP3 induces apoptosis in chicken thymic lymphoblasts and primitive hematopoietic cells, leading to severe clinical symptoms such as immunosuppression, hemorrhage, and anemia [24]. The VP1 gene is involved in viral replication and pathogenicity, and extensive studies have been conducted on its sequence variability [25,26,27]. Comparatively, the VP2 and VP3 genes exhibit higher conservation than VP1 [28]. Therefore, the VP1 gene serves as the primary source of variation among CIAV strains. Based on nucleotide sequence variations in the VP1 gene, four distinct genotypes (I, II, III, IV) have been identified worldwide [29,30]. Furthermore, studies have revealed the occurrence of recombination events in the CIAV genome, potentially leading to the emergence of new genotypes [31].
In Guangxi Province, China, a severe outbreak of CIA occurred on a large-scale 20-week-old layer hen farm, resulting in an average of 550 deaths per day over a period of 10 consecutive days. The outbreak was confined to Shed 7, while the remaining nine sheds remained unaffected. The sheds were evenly spaced 60 meters apart, and uniform management practices were implemented across all sheds. Prior to the onset of production, the chickens were grouped and reared separately for a duration of 60 days. All chickens underwent the same routine immunization program for commercial layer farms before grouping. Following grouping, each group received the same immunization program, with no specific immunization against CIAV. However, due to the large-scale operation, variations in vaccine production batches occurred, although efforts were made to ensure that each group received the same batch. Post-mortem examinations and PCR detection of liver tissue samples confirmed that CIAV was the primary causative agent of the infection. The objectives of this study were to isolate and identify CIAV strains, investigate their pathogenicity and genomic characteristics, enhance the understanding of the etiology and molecular biology of CIAV isolates in China, and provide valuable information for the prevention, control, and traceability of CIA.
2. Materials and Methods
2.1. Sample Collection
In December 2021, a total of 50 liver samples were collected from 20-week-old sick layer hens on a farm in Guangxi Province, China.
2.2. Reagents and Materials
Taq DNA polymerase and dNTPs were purchased from TaKaRa Company; DL 2000 DNA Marker was purchased from Shanghai Sangon Bioengineering Technology Service Co., Ltd.; agarose was purchased from OXOID Company; 1×TAE electrophoresis solution, EB nucleic acid dye, PBS solution (pH 7.4, 0.01 M, containing 1000 units/mL penicillin), and virus genome DNA/RNA extraction kit were purchased from Tiangen Biochemical Technology Co., Ltd.; Gel Extraction Kit D2500 was purchased from OMEGA; pMD 19-T vector was purchased from Baobio Engineering (Dalian, China) Co., Ltd. DH5α competent cells were purchased from Thermo Scientific; SPF chicken embryos were purchased from Sichuan Huapai Bioengineering Group Co., Ltd., China; MDCC-MSB1 cells were preserved by our laboratory.
2.3. Primer Design and Synthesis
According to the published CIAV genome sequence (GenBank KY486149.1), Oligo7 software was used to design and synthesize four pairs of primers (Table 1) for detecting CIAV and amplifying the whole genome. Primers were synthesized by Sangon Bioengineering (Shanghai, China) Co., Ltd.
2.4. Sample Handling
Samples were collected and processed for routine Polymerase Chain Reaction (PCR) testing, following the pooling of every 5 samples. The PCR analysis aimed to identify common poultry viruses, including Marek’s disease virus (MD), avian influenza virus (AIV), infectious bursal disease virus (IBDV), infectious bronchitis virus (IBV), adenovirus (ADV), reticuloendotheliosis virus (REV), chicken infectious anemia virus (CIAV), as well as bacterial infections. Upon testing, among the samples examined, only CIAV was detected as positive. To further investigate CIAV, fifteen sick chickens exhibiting typical clinical symptoms associated with CIA were selected. Based on the most prominent PCR amplification bands and the absence of detection for other common poultry diseases, a subset of six samples was chosen for individual CIAV PCR testing. These selected samples were stored at -80°C for subsequent experiments.
The six diseased chicken livers were individually labeled as samples 1 to 6. After being weighed and cut into small pieces, they were thoroughly mixed with PBS solution (pH 7.4, 0.01 M, containing 1000 units/mL penicillin). The mixture was homogenized to obtain a tissue suspension with a concentration of 200 g/L. The tissue suspension was then frozen at -80℃ for 30 minutes and subsequently placed in a constant temperature water bath at 37℃. The suspension was vigorously shaken until complete dissolution, and this freeze-thaw process was repeated three times to ensure proper disruption of the cells. After the freeze-thaw cycles, the suspension was centrifuged at 6000 rpm and 4℃ for 5 minutes. Following centrifugation, 200 μL of the supernatant was collected, and DNA extraction from the viral genome was performed using a DNA/RNA extraction kit. The extracted DNA from each sample was labeled accordingly as 1 to 6 and stored at -80℃ for future use.
2.4.1. PCR Detection
The primer CIAV ④ in Table 1 was used to perform PCR amplification of the extracted DNA in 2.4. A total 20 μL PCR reaction system included extracted DNA 1 μL, Taq enzyme 10 μL, ddH2O 7 μL, upstream and downstream primers 1 μL each; reaction condition was as follows: pre-denaturation at 95 ℃ for 5 min; denaturation at 94 ℃ for 30 s, annealing at 50.5 ℃ for 30 s, extending at 72 ℃ for 30 s, 30 cycles; extending at 72 ℃ for 7 min; storing at 16 ℃ to end the reaction. 5 μL of the PCR product was electrophoresed in a 1.5 % agarose gel at 120 V and 200 mA for 28 min.
2.4.2. Virus Isolation and Identification
The samples initially tested positive for CIAV using PCR in section 2.4.1. To eliminate potential viral contaminants, an equal amount of chloroform was added to the supernatants of the samples under aseptic conditions. The mixture was incubated for 15 minutes. Subsequently, to remove potential bacterial contamination, the supernatants were exposed to a 70℃ water bath for 5 minutes. The treated virus suspensions were then inoculated into the yolk sacs of three 6-day-old SPF chicken embryos, with each chicken embryo receiving a volume of 0.15 mL. 13 dpi, the SPF chicken embryos were dissected aseptically to collect the allantoic fluids, liver, spleen, thymus, and bursa of Fabricius.
The collected tissues were thoroughly homogenized to create a tissue suspension with a concentration of 200 g/L. After homogenization, the mixture was centrifuged at 3000 rpm and 4℃ for 20 minutes. The resulting supernatant was carefully collected, and to ensure purity, it was filtered through a 0.45 μm filter. This process yielded the isolated virus stock, which was ready for further analysis and experimentation.
The isolated virus stock was subjected to three successive passages using the aforementioned method. After each passage, the harvested virus stock was analyzed using PCR. If all the PCR results for the harvested virus stock were positive, it indicated the successful isolation of a CIAV strain.
2.4.3. MDCC-MSB1 Cells Inoculation of CIAV Isolate
The isolated viral suspension was inoculated onto 5×103~5×105 MDCC-MSB1 cells and cultured for 4 days at 37 ℃ under 5% CO2 condition to obtain the first passage of cell culture virus. DNA was extracted from the harvested cells and culture media for PCR detection. The CIAV-positive culture was continuously passaged up to the 5th passage for observing cytopathic effects (CPE). If CPE was observed or PCR results were positive, the virus strain was deemed to be adapted to the MDCC-MSB1 cell line.
2.5. Whole Genome Amplification of CIAV Isolates
Using the four pairs of primers listed in Table 1, we performed PCR amplification on the chicken embryo isolates to obtain four partially overlapping sequences with estimated lengths of 1010 bp, 751 bp, 1055 bp, and 540 bp. The amplification reactions were carried out in a 25 μL reaction system as described in Table 2. The reaction mixture consisted of 2 μL of total supernatant DNA, 12.5 μL of Taq enzyme, 8.5 μL of ddH2O, and 1 μL each of the upstream and downstream primers. The amplified products were visualized by electrophoresis on a 1.5% agarose gel, and the PCR amplification products were then extracted using the OMEGA Gel Extraction Kit D2500. Subsequently, the purified products were ligated into the pMD19-T vector for sequencing. The obtained sequencing results were analyzed using DNA STAR software SeqMan Pro, which allowed for the assembly of the sequences to obtain the full-length genome of the CIAV strain isolated from the chicken farm.
2.6. Experimental Infection of 1-Day-Old SPF Chicks with CIAV
Eighteen 1-day-old SPF chicks were intramuscularly inoculated with 0.3 mL (104 EID50) of the chicken embryo passage virus solution per chick in the leg, while an additional 6 chicks were inoculated with equal doses of saline as a control. The two groups of chicks were housed separately in appropriate conditions to ensure their comfort. Throughout the duration of the experiment, any deceased test chicks were promptly dissected and subjected to testing. Organs exhibiting lesions were carefully selected, fixed in 10% formalin, and subsequently embedded in paraffin for the preparation of pathological histological sections. These sections were then stained using hematoxylin and eosin (HE), and images were captured using a 3DHISTECH (Hungary) Pannoramic 250 digital section scanner to observe any histopathological changes.
2.7. Genome-Wide Bioinformatics Analysis
2.7.1. Genome-Wide Genetic Evolutionary Analysis
In this study, a total of 75 reference strains of CIAV isolated from different periods and regions were selected based on their full sequences available in GenBank (see Appendix A). The whole genome sequences obtained through amplification in our study were aligned with these reference strains using MEGA-X software, employing the Clustal W algorithm. The genetic evolutionary tree was constructed using the Neighbor-Joining (NJ) method, with 1000 bootstrap replicates. To determine the genotype of the CIAV strain from the chicken farm under investigation, the typing method proposed by Ducatez, M.F. et al. [29] was employed. To explore the possibility of recombination events among the selected sequences, we utilized the RDP 4.0 software [32]. Multiple methods, including RDP, Geneconv, Bootscan, Maxchi, Chimaera, Siscan, and 3Seq, were employed with a P value adjusted to 0.05. The recombination events identified were further validated using SimPlot v.3.5.1 [33].
3. Results
3.1. Pathological Changes
3.2. PCR Test Results of the Samples
All the selected six liver samples, which exhibited characteristic pathological changes, were tested positive for CIAV. The electrophoresis analysis revealed a band size of approximately 500 bp, which corresponded to the expected target band (Figure 2a).
3.3. Virus Isolation Results
After three consecutive passages in SPF chicken embryos, PCR testing of the chicken embryo yolk sac culture virus nucleic acid confirmed three positive results for CIAV (Figure 2b), indicating the successful isolation of three CIAV isolates. These isolates were named GX21121, GX21122, and GX21123. Among these isolates, strain GX21121 exhibited persistent liver lesions and specific PCR bands even after three additional passages. Therefore, strain GX21121 was selected for further investigation of its pathogenicity.
3.4. MDCC-MSB1 Culture Results
After incubating the MDCC-MSB1 cells with CIAV strain GX21121 for 5 days, the PCR test consistently yielded negative results. This trend continued even after four consecutive passages. The experiment was repeated three times, and each time the PCR test yielded negative results. Based on these findings, it can be concluded that the GX21121 strain isolated from the SPF chicken embryo yolk sac is not adapted to the MDCC-MSB1 cell line.
3.5. Whole Genome Amplification Results
The results of PCR amplification of the four fragments are shown below (Figure 2c). which were consistent with expectations. Three CIAV complete gene sequences of 2298 kb referring to CIAV isolates GX21121, GX21122, and GX21123 (GenBank accession numbers: OQ267594, OQ267595, OQ267596). In this study were obtained after sequencing and splicing. Based on the gene annotation information, a mockup conforming to the whole gene of the three CIAV strains was drawn (Figure 2d).
3.6. CIAV GX21121 Pathogenicity to One-Day-Old SPF Chicks
After inoculating the experimental SPF chicks with CIAV GX21121, a total of 18 chicks were monitored. Two chicks died at 7 dpi, five chicks died at 14 dpi, and the remaining 11 chicks died successively between 15 and 29 dpi. No deaths were observed in the negative control (NC) group. When compared to the NC, the deceased chicks infected with CIAV at 7 and 14 dpi exhibited significant clinical signs of depression and rough feathers. Varying degrees of scattered subcutaneous hemorrhagic spots were observed. Additionally, the infected chicks showed pale muscles, yellow staining of the liver with white necrotic foci at the edges, swollen and hemorrhaged kidneys at 7 dpi, pale kidneys at 14 dpi, swollen spleen with white necrotic foci on the surface at 7 dpi, and necrosis at the edge of the spleen at 14 dpi. The 7 dpi chicks displayed thymus enlargement and hemorrhage, while the 14-day-old dead chicks showed thymus atrophy. Selected significant changes between the experimental group (EG) and the negative control (NC) are presented below (Figure 3a–h). Microscopic examination of histopathological sections from the 7 dpi deceased chicks revealed liver cell necrosis, fibrous serous exudation, inflammatory cell infiltration, and hemorrhage in some liver tissues. Lung tissues exhibited fibrinoid exudation and inflammation. Renal sections showed renal cell infiltration, fibrous tissue hyperplasia and congestion, renal tubular degeneration and necrosis, inflammatory cell infiltration, and fibrous tissue hyperplasia, among other pathological changes (Figure 3s–u). CIAV nucleic acids were detected positive in all experimental infected chicks.
s-u. Microscopic examination of the histopathological changes in the liver tissue of the 7 dpi chicks (s) revealed the absence of clear lobular division and disordered arrangement of hepatic cords. Extensive hepatocellular necrosis was observed, characterized by cytoplasmic shrinkage or dissolution, nuclear pyknosis, and disappearance. Patchy hepatocyte necrosis resulted in the formation of necrotic foci. Inflammatory cells, primarily neutrophils, infiltrated the area, accompanied by fibro-serous exudation. The necrotic foci were also associated with bleeding, visible red blood cell aggregation, and the accumulation of light-staining fluid around their edges. No significant inflammatory cell infiltration was observed in the liver sinusoids. The intrahepatic portal area, interlobular arteries, interlobular veins, and interlobular bile duct structures remained intact, with no apparent fibrous tissue proliferation or inflammatory cell infiltration. In the lung tissue (t), the surface was covered with pleura, without visible lobular division. The connective tissue of the pleura extended deep into the lung parenchyma, dividing it into several pulmonary lobes. The bronchial structure at all levels of lung tissue appeared normal, with relatively neat arrangement of the bronchial ciliated epithelium. Fibrin-like exudate was present inside the bronchi, and the bronchial lumen contained a light-staining substance. The structure of the third-level bronchi, pulmonary lobules, and pulmonary capillaries was clear. Inflammatory cells, mainly segmented or rod-shaped neutrophils, infiltrated into the loose connective tissue around the blood vessels. Some fibrous tissue proliferation was observed in the lung tissue, with elongated nucleus-shaped fibroblasts. A significant aggregation of red blood cells was observed, while no other notable pathological changes were observed. In the kidney tissue (u), the renal capsule remained intact, and the boundary between the cortex and medulla was relatively clear. The structure of the cortical glomeruli was complete and clear, with no obvious degeneration or necrosis. The renal tubules were arranged relatively regularly. Some renal tubular degeneration and necrosis were observed, with degenerated and necrotic renal tubular epithelial cells exhibiting cytoplasmic dissolution, nuclear shrinkage, and disintegration, resulting in the disappearance of renal tubule structures in severely affected areas, forming necrotic foci accompanied by fibrous tissue proliferation. Inflammatory cells, including neutrophils and lymphocytes, infiltrated the area, along with elongated nucleus-shaped fibroblasts. No significant inflammatory infiltration or fibrosis was observed in the interstitium.
3.7. Genome-Wide Bioinformatics Analysis
The phylogenetic analysis of the complete genomes of the isolated strains and reference strains revealed close relationships among the three isolated strains. GX21122 and GX21123 formed a distinct minor branch, indicating a closer evolutionary relationship between them compared to GX21121 (Figure 4A). The reference strains that exhibited the closest genetic relationship were obtained from the northeastern region of China (GenBank KY486144, KY486155, KY486149). Nucleotide homology analysis showed a high level of similarity among the isolated strains, with the highest homology of 99.74% observed with the reference strains (GenBank KY486144), which is consistent with the phylogenetic tree based on complete genomes. The VP1 phylogenetic tree demonstrated a tight clustering of the VP1 sequences of the three isolated strains, along with the Heilongjiang strain (GenBank KY486144) and the Jilin strain (GenBank KY486155), all belonging to genotype IIIc (Figure 4B). Analysis of the VP1 amino acid sequences revealed the presence of glutamine at position 394 in all three isolated strains, which is considered a characteristic feature of virulent CIAV strains [34].
Recombination analysis using RDP 4.0 software indicated that GX21122 and GX21123 were potential recombinant strains involved in the same recombination event. The major parent was the Guangxi strain (GenBank MN103406) of genotype IV, while the minor parent was not detected. The predicted recombination region was identified between nucleotide positions 1790-2210. This recombination event was supported by seven out of nine methods used in RDP 4.0 (Table 3 and Figure 4C,E). The SimPlot analysis confirmed these results, showing a potential fragment recombination event of GX21122 and GX21123 with the region of 1679-2132 nt (Figure 4D,F). No potential recombination signal was detected for GX21121.
4. Discussion
This study investigates an outbreak of disease that occurred in a 20-week-old laying hen farm in Guangxi Province, China. The affected hens exhibited clinical symptoms and pathological changes consistent with anemia, including depression, pale combs, and muscles. we successfully isolated and identified three strains of CIAV, namely GX21121, GX21122, and GX21123, using SPF chicken embryo yolk sac inoculation. The full genome sequences of these strains were obtained through PCR amplification and splicing, and their corresponding GenBank accession numbers are OQ267594, OQ267595, and OQ267596. Through pathogen isolation, identification, and comprehensive genome analysis, we confirmed the presence of CIAV on this particular farm.
Epidemiological surveys have shown that the prevalence of CIAV infection in chicken flocks in China ranges from 40% to 70%. It is particularly common in chickens aged 1 to 3 weeks, as they are more susceptible to the virus due to the associated immunosuppression [10,11]. However, it is noteworthy that this outbreak occurred in 20-week-old laying hens, which are generally considered less susceptible to CIAV infection. CIAV infection can induce immunosuppression in chickens, making them more susceptible to other pathogens. Co-infections involving CIAV and other pathogens have been documented in clinical cases [35,36,37], suggesting a possible association with this outbreak. However, laboratory testing conducted on the affected chickens in this study only detected the presence of CIAV, ruling out common viral and bacterial diseases. Considering the clinical symptoms observed, it is highly likely that CIAV was the primary causative agent of this outbreak. It is important to note that the limited scope of pathogen testing should be taken into consideration, but these findings emphasize the potential impact of a CIAV outbreak on poultry farms. The results suggest that CIAV-induced immunosuppression can manifest in various clinical signs and lesions within the flock, which may vary depending on the age of the chickens.
No CPE were observed in MDCC-MSB1 cells infected with the GX21121 strain, even after repeating the experiment five times. Additionally, consistent negative results were obtained from PCR testing, indicating that the isolate did not exhibit adaptability to MDCC-MSB1 cells. This observation is consistent with previous reports that have highlighted variations in CIAV infectivity among different subtypes of MSB1 cells [15]. Different CIAV strains display differential sensitivity depending on the MSB1 cell subtype. For example, the CIA-1 strain [38] may not replicate in MSB1-L cells but shows limited replication in MSB1-S cells, while both cell subtypes are susceptible to the CUX-1 strain [39]. The underlying reasons for these differences are not yet fully understood. Our analysis primarily focused on the preliminary assessment of GX21121’s adaptability to MDCC-MSB1 cells. However, subsequent animal pathogenicity experiments confirmed the significant virulence of GX21121, indicating that the invasiveness of CIAV in cells is not strongly correlated with its pathogenicity. It is important to note that definitive conclusions regarding cell cultivation conditions could not be drawn from this study. Nevertheless, these findings provide preliminary insights into investigating the differential infectivity of CIAV in MSB1 cells.
All 1-day-old SPF chicks infected with the GX21121 strain experienced mortality within 29 days, with acute deaths observed at 7 dpi. These infected chicks displayed clinical symptoms such as depression and showed macroscopic and microscopic pathological changes, including scattered subcutaneous hemorrhagic lesions, liver necrosis foci, and severe anemia. It is important to note that GX21121 was isolated from a flock that had not been vaccinated against CIAV. Considering that only one shed of the poultry farm experienced morbidity and mortality, with no previous outbreaks of other infectious diseases, the likelihood of the outbreak being caused by feed or husbandry management factors is considered low. Additionally, the large scale of the poultry farm and separate groups of birds subjected to the same immunization procedures but potentially differing vaccine batches suggest the possibility of the virus being introduced through vaccines [2,4]. Further analysis of the complete genome sequence revealed a close genetic relationship between the isolated strains and three strains from the northeastern region of China, all belonging to genotype IIIc. Recombination analysis identified reliable recombination signals between GX21122 and GX21123 within nucleotide positions 1670-2130. The parental strains involved in the recombination event were genotype IV field strains prevalent in Guangxi. This recombination event occurred within amino acid positions 282aa-433aa of the VP1 protein. Interestingly, previous reports have highlighted significant differences between genotypes IIIc and IV in the VP1 amino acid sequence, particularly within positions 294aa-448aa [40]. Therefore, it is highly likely that recombination occurred between the parental strains in this region, resulting in the emergence of a new strain with an altered genotype. Multiple recombination events were detected during the analysis, highlighting the importance of recombination as a crucial mechanism for generating new CIAV strains and genotypes. Despite the overall conservation of CIAV sequences, it is important to acknowledge evolutionary trends, as this disease outbreak may indicate the onset of evolutionary changes in the infectivity characteristics of the virus.
Indeed, vaccination is an effective measure to control CIAV and prevent outbreaks of the disease. The outbreak described in this study can be attributed to the lack of vaccination against CIAV on the affected poultry farm. The study aimed to analyze the genetic diversity and evolution of the three CIAV strains isolated from a large-scale layer hen farm in Guangxi Province, China. The genomic characteristics of the isolated strains were examined, and the pathogenicity of the GX21121 isolate was clarified. By analyzing the genetic diversity and evolution of the isolated strains, the study provides important insights and references for the prevention, control, and traceability of CIA. Understanding the genomic characteristics of the strains and clarifying the pathogenicity of GX21121 can aid in developing effective control strategies, including vaccination programs, to prevent future outbreaks and mitigate the impact of the disease on poultry farms. The study’s findings serve as valuable materials and references for stakeholders involved in the management and control of CIA in poultry populations.
5. Conclusion
In this study, an outbreak of disease in a 20-week-old laying hen farm in Guangxi Province, China was investigated. The study revealed that the clinical symptoms and pathological changes associated with CIAV infection vary depending on the age of the chickens. Three CIAV strains were isolated from the affected farm and subjected to comprehensive analysis. The whole genome analysis of the isolated strains provided insights into their genetic characteristics and evolutionary relationships. The study also examined the possibility of recombination events among the strains. Additionally, an animal pathogenicity experiment confirmed the high virulence of the GX21121 isolate, as evidenced by the development of typical CIA lesions and the 100% mortality rate observed in 1-day-old SPF chicks. The findings of this study contribute valuable information to the understanding of CIAV pathogenicity, genomic characteristics, and evolution. This information can be utilized for further research on the etiology, molecular epidemiology, and viral evolution of CIAV. The study serves as a foundation for future studies aimed at improving prevention, control, and management strategies for CIAV outbreaks in poultry farms.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, Z.Y. and Z.H.; methodology, Z.Y.; software, Z.Y.; validation, Z.Y. and Z.H.; formal analysis, Z.Y.; investigation, Z.Y.; resources, Z.Y.; data curation, Z.Y.; writing—original draft preparation, Z.Y.; writing—review and editing, Z.H.; supervision, Z.H..All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the “Innovative Graduate Research Project” provided by Southwest Minzu University (ZD2022633).
Institutional Review Board Statement
The animal experiments were conducted in accordance with the protocol of the National Act on the Use of Experimental Animals(China) and were approved by the Sichuan Committee on Laboratory Animals (approval number SYXK(川)2019-216).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
All data generated in the present study was available in the published manuscript and its Supplementary Materials.
Acknowledgments
We acknowledge the extraordinary support given by the administrative personnel of our Department, especially Dr. Lucia Ferroni and Dr. Monica della Pietra. We also thank Prof. Paolo Fais of the University of Bologna for the useful early discussions about PFAS.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
| No. |
Accession NO. | Country | Isolate | Year | Nucleotides (bp) |
Host |
| 1 | ON596886 | China | GX2112 | 2021 | 2298 | Chicken |
| 2 | MH186142 | China | CIAV-Shanxi7 | 2018 | 2291 | Chicken |
| 3 | HM134240 | China | \ | 2010 | 2298 | Chicken |
| 4 | KU050680 | China | GD-101 | 2015 | 2298 | Chicken |
| 5 | KU645520 | China | HN1405 | 2014 | 2298 | Chicken |
| 6 | KU845735 | China | CIAV-F10 | 2015 | 2298 | Chicken |
| 7 | KX447637 | China | LY-2 | 2016 | 2298 | Chicken |
| 8 | KX811526 | China | SD15 | 2015 | 2298 | Chicken |
| 9 | KY486144 | China | HLJ15170 | 2015 | 2298 | Chicken |
| 10 | KY486151 | China | NX15140 | 2015 | 2298 | Chicken |
| 11 | KY486155 | China | LN15170 | 2015 | 2298 | Chicken |
| 12 | MK089240 | China | 17CC0509 | 2017 | 2298 | Chicken |
| 13 | MK089243 | China | 17SY0902 | 2017 | 2298 | Chicken |
| 14 | MK887171 | China | N1 | 2016 | 2298 | Chicken |
| 15 | MN103406 | China | GX1904B | 2019 | 2298 | Chicken |
| 16 | MN649259 | China | GX1907B | 2019 | 2298 | Chicken |
| 17 | MT091001 | China | TBC19 | 2019 | 2298 | Chicken |
| 18 | MW423616 | China | TZC1910 | 2019 | 2298 | Chicken |
| 19 | MW579762 | China | GX2020-D6 | 2020 | 2298 | Chicken |
| 20 | MW660821 | China | SDSPF2020 | 2020 | 2298 | SPF |
| 21 | MZ540762 | China | YN04 | 2020 | 2298 | Chicken |
| 22 | MH186141 | China | CIAV-Guangdong11 | 2018 | 2291 | Chicken |
| 23 | KF224927 | China | GD-C-12 | 2013 | 2298 | Chicken |
| 24 | AF475908 | China | \ | 2002 | 2298 | Chicken |
| 25 | MH186138 | China | CIAV-Hebei12 | 2018 | 2293 | Chicken |
| 26 | MH186137 | China | CIAV-Heilongjiang16 | 2018 | 2295 | Unstated |
| 27 | MZ668717 | China | HN2021-1415 | 2021 | 2298 | Chicken |
| 28 | MK770259 | China | XH16 | 2016 | 2298 | Chicken |
| 29 | KM496310 | China | SC-PS | 2013 | 2298 | Chicken |
| 30 | EF176599 | China | C14 | 2007 | 2298 | Unstated |
| 31 | KM496303 | China | SC-HY | 2014 | 2298 | Chicken |
| 32 | KM496305 | China | SC-MS | 2014 | 2298 | Chicken |
| 33 | KM496307 | China | SC-MZ42A | 2013 | 2298 | Chicken |
| 34 | KM496309 | China | SC-NC2 | 2013 | 2298 | Chicken |
| 35 | KJ728816 | China | 4 | 2012 | 2298 | Chicken |
| 36 | MT795928 | China | 1512TW | 2016 | 2298 | Chicken |
| 37 | MT799753 | China | 1635TW | 2017 | 2298 | Chicken |
| 38 | MT799771 | China | 1874TW, | 2018 | 2298 | Chicken |
| 39 | AY846844 | China | TJBD40 | 2004 | 2298 | Unstated |
| 40 | AF311892 | Alabama | 98D02152 | 2003 | 2298 | Unstated |
| 41 | KJ872514 | Argentina | CAV-18 | 2007 | 2298 | Chicken |
| 42 | AF395114 | Bangladesh | BD-3 | 2004 | 2298 | Unstated |
| 44 | MG846491 | Brazil | RS/BR/15/1R | 2015 | 2298 | Chicken |
| 45 | MT671980 | Brazil | CAV | 2018 | 2298 | Chicken |
| 46 | MG827098 | Egypt | CAV-CA1-2015 | 2015 | 2298 | Chicken |
| 47 | MG827099 | Egypt | CAV-GZ2-2016 | 2016 | 2298 | Chicken |
| 48 | MG827100 | Egypt | CAV-SK4-2017 | 2017 | 2298 | Chicken |
| 49 | MH001570 | Egypt | CAV-EG-28 | 2017 | 2298 | Chicken |
| 50 | MT268633 | Egypt | EG-ElBeheira-2019 | 2019 | 2298 | Chicken |
| 51 | KY053900 | India | CAV/NAM/TANUVAS/09 | 2009 | 2263 | Chicken |
| 52 | MZ868644 | India | NIAB-DPR | 2020 | 2298 | Chicken |
| 53 | MT239353 | Iran | UT-Zahraee | 2018 | 2298 | Chicken |
| 54 | MT813071 | Italy | CIAV/IT/CK/855/17 | 2017 | 2189 | Chicken |
| 55 | MT813078 | Italy | CIAV/IT/CK/1188/19 | 2019 | 2186 | Chicken |
| 56 | AY040632 | Malaysia | 3-1P60 | 2001 | 2298 | Chicken |
| 57 | MZ666102 | Tunisia | CIAV_TN_7-15 | 2019 | 2181 | Chicken |
| 58 | MT259325 | Turkey | GDA6K | 2015 | 2296 | Chicken |
| 59 | MW219799 | Turkey | CIAV/2011/Akcdg/TUR | 2011 | 2298 | Chicken |
| 60 | MW219800 | Turkey | CIAV/2014/Elzg-66/TUR | 2014 | 2298 | Chicken |
| 61 | MH536106 | Vietnam | G17.3.1 | 2017 | 2298 | Chicken |
| 62 | D10068 | \ | 26P4 (vaccine) | 2007 | 2298 | Cells |
| 63 | AF313470 | USA | Del Ros (vaccine) | 2000 | 2294 | Unstated |
| 64 | M81223 | \ | CUX-1 (vaccine) | 1993 | 2298 | Unstated |
| 65 | EF683159 | Australia | 3711 (vaccine) | 2007 | 2279 | Unstated |
| 66 | MH001564 | Egypt | CAV-EG-26 | 2017 | 2298 | Chicken |
| 67 | JF507715 | South Korea | CIAV89-69 | 1991 | 2298 | Chicken |
| 68 | AB119448 | Japan | G6 | 2009 | 2298 | Chicken |
| 69 | DQ016140 | Slovenia | 69 | 2006 | 1350 | Unstated |
| 70 | JQ308213 | Chile | CL37 | 2008 | 1831 | Unstated |
| 71 | KY888892 | China | 1102PT01 | 2011 | 1823 | Chicken |
| 72 | MK423873 | Vietnam | Vietnam/PT1/17 | 2019 | 1823 | Chicken |
| 73 | KP780288 | Vietnam | CAV/Vietnam/HN1/13 | 2018 | 1823 | Chicken |
| 74 | DQ991394 | USA | 01-4201 | 2007 | 2298 | Unstated |
| 75 | KY888907 | China | 1310CH05 | 2013 | 1823 | Chicken |
References
- Shao, H.; Li, J.; Yuan, H.; Ji, L.; Zhang, J.; Jin, W.; Qian, K.; Ye, J.; Qin, A. Isolation and Molecular Characteristics of a CIAV Isolate From Pigeons, China. Front. Vet. Sci. 2021, 8, 669154. [CrossRef]
- Li, Y.; Wang, J.; Chen, L.; Wang, Q.; Zhou, M.; Zhao, H.; Chi, Z.; Wang, Y.; Chang, S.; Zhao, P. Genomic Characterization of CIAV Detected in Contaminated Attenuated NDV Vaccine: Epidemiological Evidence of Source and Vertical Transmission From SPF Chicken Embryos in China. Front. Vet. Sci. 2022, 9, 930887. [CrossRef]
- Rosario, K.; Breitbart, M.; Harrach, B.; Segalés, J.; Delwart, E.; Biagini, P.; Varsani, A. Revisiting the Taxonomy of the Family Circoviridae: Establishment of the Genus Cyclovirus and Removal of the Genus Gyrovirus. Arch. Virol. 2017, 162, 1447–1463. [CrossRef]
- Miller, M.M.; Ealey, K.A.; Oswald, W.B.; Schat, K.A. Detection of Chicken Anemia Virus DNA in Embryonal Tissues and Eggshell Membranes. Avian Dis. 2003, 47, 662–671. [CrossRef]
- Noteborn, M.H.; de Boer, G.F.; van Roozelaar, D.J.; Karreman, C.; Kranenburg, O.; Vos, J.G.; Jeurissen, S.H.; Hoeben, R.C.; Zantema, A.; Koch, G. Characterization of Cloned Chicken Anemia Virus DNA That Contains All Elements for the Infectious Replication Cycle. J. Virol. 1991, 65, 3131–3139. [CrossRef]
- Yuasa, N.; Yoshida, T.T. Isolation and Some Characteristics of an Agent Inducing Anemia in Chicks. Avian Dis. 1979, 23, 366. [CrossRef]
- Chu, D.K.W.; Poon, L.L.M.; Chiu, S.S.S.; Chan, K.H.; Ng, E.M.; Bauer, I.; Cheung, T.K.; Ng, I.H.Y.; Guan, Y.; Wang, D.; et al. Characterization of a Novel Gyrovirus in Human Stool and Chicken Meat. J. Clin. Virol. Off. Publ. Pan Am. Soc. Clin. Virol. 2012, 55, 209–213. [CrossRef]
- Fatoba, A.J.; Adeleke, M.A. Chicken Anemia Virus: A Deadly Pathogen of Poultry. Acta Virol. 2019, 63, 19–25. [CrossRef]
- Fang, L.; Li, Y.; Wang, Y.; Fu, J.; Cui, S.; Li, X.; Chang, S.; Zhao, P. Genetic Analysis of Two Chicken Infectious Anemia Virus Variants-Related Gyrovirus in Stray Mice and Dogs: The First Report in China, 2015. BioMed Res. Int. 2017, 2017, 6707868. [CrossRef]
- Kim, H.-R.; Kwon, Y.-K.; Bae, Y.-C.; Oem, J.-K.; Lee, O.-S. Molecular Characterization of Chicken Infectious Anemia Viruses Detected from Breeder and Broiler Chickens in South Korea. Poult. Sci. 2010, 89, 2426–2431. [CrossRef]
- Miller, M.M.; Schat, K.A. Chicken Infectious Anemia Virus: An Example of the Ultimate Host–Parasite Relationship. Avian Dis. 2004, 48, 734–745. [CrossRef]
- Todd, D. Avian Circovirus Diseases: Lessons for the Study of PMWS. Vet. Microbiol. 2004, 98, 169–174. [CrossRef]
- Li, Y.; Hu, Y.; Cui, S.; Fu, J.; Wang, Y.; Cui, Z.; Fang, L.; Chang, S.; Zhao, P. Molecular Characterization of Chicken Infectious Anemia Virus from Contaminated Live-Virus Vaccines. Poult. Sci. 2017, 96, 1045–1051. [CrossRef]
- Zhang, X.; Liu, Y.; Wu, B.; Sun, B.; Chen, F.; Ji, J.; Ma, J.; Xie, Q. Phylogenetic and Molecular Characterization of Chicken Anemia Virus in Southern China from 2011 to 2012. Sci. Rep. 2013, 3, 3519. [CrossRef]
- Yuasa, N. Propagation and Infectivity Titration of the Gifu-1 Strain of Chicken Anemia Agent in a Cell Line (MDCC-MSB1) Derived from Marek’s Disease Lymphoma. Natl. Inst. Anim. Health Q. (Tokyo) 1983, 23, 13–20.
- Akiyama, Y.; Kato, S. Two Cell Lines from Lymphomas of Marek’s Disease. Biken J. 1974, 17, 105–116.
- Renshaw, R.W.; Soiné, C.; Weinkle, T.; O’Connell, P.H.; Ohashi, K.; Watson, S.; Lucio, B.; Harrington, S.; Schat, K.A. A Hypervariable Region in VP1 of Chicken Infectious Anemia Virus Mediates Rate of Spread and Cell Tropism in Tissue Culture. J. Virol. 1996, 70, 8872–8878. [CrossRef]
- Lacorte, C.; Lohuis, H.; Goldbach, R.; Prins, M. Assessing the Expression of Chicken Anemia Virus Proteins in Plants. Virus Res. 2007, 129, 80–86. [CrossRef]
- Koch, G.; van Roozelaar, D.J.; Verschueren, C.A.; van der Eb, A.J.; Noteborn, M.H. Immunogenic and Protective Properties of Chicken Anaemia Virus Proteins Expressed by Baculovirus. Vaccine 1995, 13, 763–770. [CrossRef]
- Douglas, A.J.; Phenix, K.; Mawhinney, K.A.; Todd, D.; Mackie, D.P.; Curran, W.L. Identification of a 24 KDa Protein Expressed by Chicken Anaemia Virus. J. Gen. Virol. 1995, 76 ( Pt 7), 1557–1562. [CrossRef]
- Trinh, D.Q.; Ogawa, H.; Bui, V.N.; Baatartsogt, T.; Kizito, M.K.; Yamaguchi, S.; Imai, K. Characterization of MAbs to Chicken Anemia Virus and Epitope Mapping on Its Viral Protein, VP1. J. Gen. Virol. 2015, 96, 1086–1097. [CrossRef]
- Noteborn, M.H.; Kranenburg, O.; Zantema, A.; Koch, G.; de Boer, G.F.; van der Eb, A.J. Transcription of the Chicken Anemia Virus (CAV) Genome and Synthesis of Its 52-KDa Protein. Gene 1992, 118, 267–271. [CrossRef]
- Peters, M.A.; Jackson, D.C.; Crabb, B.S.; Browning, G.F. Chicken Anemia Virus VP2 Is a Novel Dual Specificity Protein Phosphatase. J. Biol. Chem. 2002, 277, 39566–39573. [CrossRef]
- Wang, J.; Li, Y.; Zhang, Y.; Chen, L.; Fang, L.; Chang, S.; Wang, Y.; Zhao, P. Construction of Chicken Infectious Anemia Virus Infectious Clone and Study on Its Pathogenicity. Front. Microbiol. 2022, 13, 1016784. [CrossRef]
- Simultaneous Expression of Recombinant Baculovirus-Encoded Chicken Anaemia Virus (CAV) Proteins VP1 and VP2 Is Required for Formation of the CAV-Specific Neutralizing Epitope - PubMed Available online: https://pubmed.ncbi.nlm.nih.gov/9880024/ (accessed on 18 February 2023).
- Chowdhury, S.M.Z.H.; Omar, A.R.; Aini, I.; Hair-Bejo, M.; Jamaluddin, A.A.; Md-Zain, B.M.; Kono, Y. Pathogenicity, Sequence and Phylogenetic Analysis of Malaysian Chicken Anaemia Virus Obtained after Low and High Passages in MSB-1 Cells. Arch. Virol. 2003, 148, 2437–2448. [CrossRef]
- Hailemariam, Z.; Omar, A.R.; Hair-Bejo, M.; Giap, T.C. Detection and Characterization of Chicken Anemia Virus from Commercial Broiler Breeder Chickens. Virol. J. 2008, 5, 128. [CrossRef]
- Farkas, T.; Tanaka, A.; Kai, K.; Kanoe, M. Cloning and Sequencing of the Genome of Chicken Anaemia Virus (CAV) TK-5803 Strain and Comparison with Other CAV Strains. J. Vet. Med. Sci. 1996, 58, 681–684. [CrossRef]
- Ducatez, M.F.; Owoade, A.A.; Abiola, J.O.; Muller, C.P. Molecular Epidemiology of Chicken Anemia Virus in Nigeria. Arch. Virol. 2006, 151, 97–111. [CrossRef]
- Ou, S.-C.; Lin, H.-L.; Liu, P.-C.; Huang, H.-J.; Lee, M.-S.; Lien, Y.-Y.; Tsai, Y.-L. Epidemiology and Molecular Characterization of Chicken Anaemia Virus from Commercial and Native Chickens in Taiwan. Transbound. Emerg. Dis. 2018, 65, 1493–1501. [CrossRef]
- Van Dong, H.; Tran, G.T.H.; Van Nguyen, G.; Dao, T.D.; Bui, V.N.; Huynh, L.T.M.; Takeda, Y.; Ogawa, H.; Imai, K. Chicken Anemia Virus in Northern Vietnam: Molecular Characterization Reveals Multiple Genotypes and Evidence of Recombination. Virus Genes 2019, 55, 643–653. [CrossRef]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and Analysis of Recombination Patterns in Virus Genomes. Virus Evol. 2015, 1, vev003. [CrossRef]
- Lole, K.S.; Bollinger, R.C.; Paranjape, R.S.; Gadkari, D.; Kulkarni, S.S.; Novak, N.G.; Ingersoll, R.; Sheppard, H.W.; Ray, S.C. Full-Length Human Immunodeficiency Virus Type 1 Genomes from Subtype C-Infected Seroconverters in India, with Evidence of Intersubtype Recombination. J. Virol. 1999, 73, 152–160. [CrossRef]
- Li, Y.; Fang, L.; Cui, S.; Fu, J.; Li, X.; Zhang, H.; Cui, Z.; Chang, S.; Shi, W.; Zhao, P. Genomic Characterization of Recent Chicken Anemia Virus Isolates in China. Front. Microbiol. 2017, 8, 401. [CrossRef]
- Miles, A.M.; Reddy, S.M.; Morgan, R.W. Coinfection of Specific-Pathogen-Free Chickens with Marek’s Disease Virus (MDV) and Chicken Infectious Anemia Virus: Effect of MDV Pathotype. Avian Dis. 2001, 45, 9–18.
- Gimeno, I.M.; Schat, K.A. Virus-Induced Immunosuppression in Chickens. Avian Dis. 2018, 62, 272–285. [CrossRef]
- Li, X.; Zhang, K.; Pei, Y.; Xue, J.; Ruan, S.; Zhang, G. Development and Application of an MRT-QPCR Assay for Detecting Coinfection of Six Vertically Transmitted or Immunosuppressive Avian Viruses. Front. Microbiol. 2020, 11, 1581. [CrossRef]
- Lucio, B.; Schat, K.A.; Shivaprasad, H.L. Identification of the Chicken Anemia Agent, Reproduction of the Disease, and Serological Survey in the United States. Avian Dis. 1990, 34, 146–153.
- Renshaw, R.W.; Soiné, C.; Weinkle, T.; O’Connell, P.H.; Ohashi, K.; Watson, S.; Lucio, B.; Harrington, S.; Schat, K.A. A Hypervariable Region in VP1 of Chicken Infectious Anemia Virus Mediates Rate of Spread and Cell Tropism in Tissue Culture. J. Virol. 1996, 70, 8872–8878. [CrossRef]
- Rao, P.L.; Priyanka, E.; Kannaki, T.R.; Subbiah, M. Whole Genome Analysis and Molecular Characterization of Chicken Infectious Anemia Virus from an Outbreak in a Layer Flock Reveals Circulation of Genogroup IIIb in South India. Virus Res. 2022, 308, 198649. [CrossRef]
Figure 1.
Typical lesions of sick and dead chickens. (a) Cockscomb pale. (b) pale chest muscles. (c) liver diffused hemorrhage and hepatic necrosis.
Figure 1.
Typical lesions of sick and dead chickens. (a) Cockscomb pale. (b) pale chest muscles. (c) liver diffused hemorrhage and hepatic necrosis.
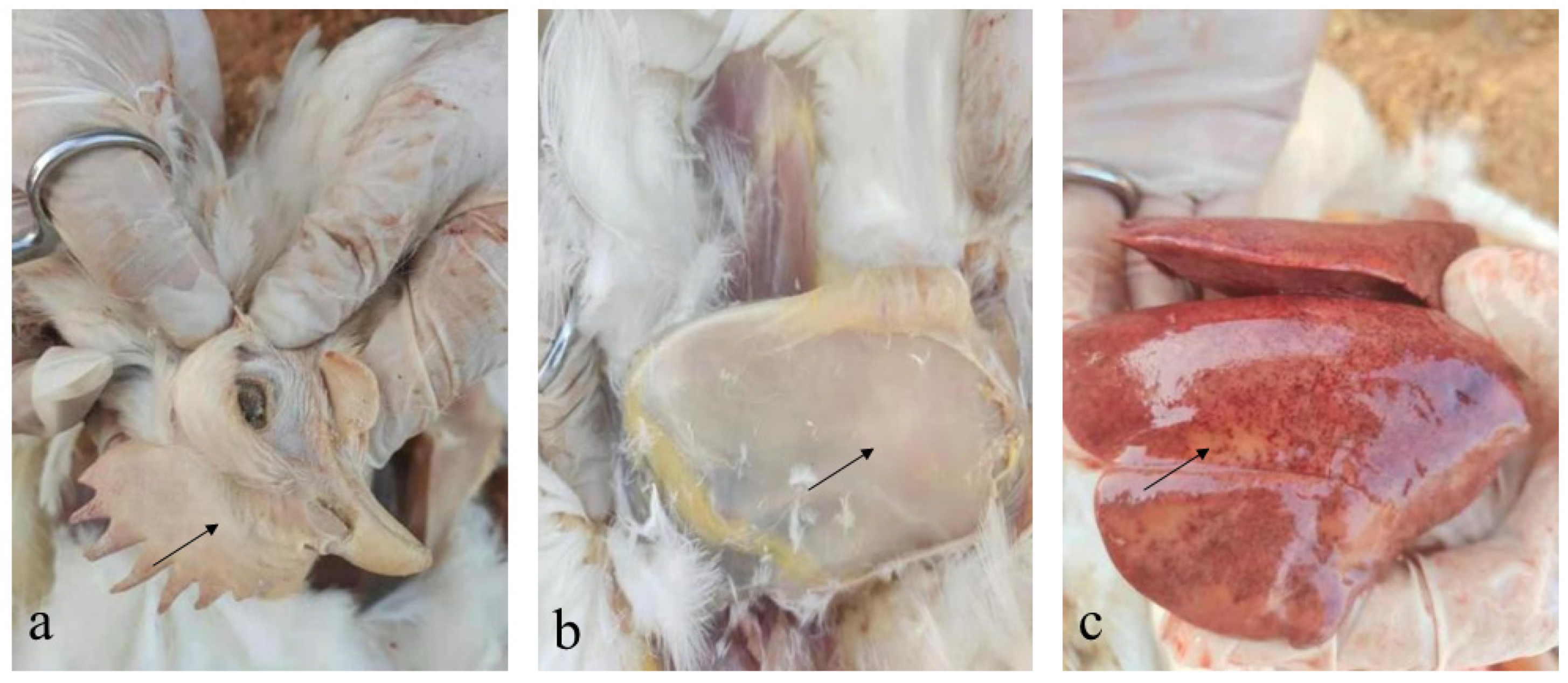
Figure 2.
(a) PCR detection results of the 6 liver samples suspected to be CIAV positive. Note: M: DNA Marker (DL 2000); 1~6: clinical samples; -: negative control. (b) PCR detection results of three CIAV isolates. Note: M: DNA Marker (DL 2000); 1~3: GX21121, GX21122, and GX21123 viruses isolated from 3 liver samples; -: negative control; +: positive control. (c) Four fragments of GX21121 gene amplification electrophoresis results. Note: M: DNA Marker (DL 2000); CIAV①~④: Four amplified fragments with 4 primers. CIAV GX21121 DNA simulation map by SnapGene software. Note: The yellow arrow part is the coding region, encoding 3 proteins: VP1, VP2, VP3.
Figure 2.
(a) PCR detection results of the 6 liver samples suspected to be CIAV positive. Note: M: DNA Marker (DL 2000); 1~6: clinical samples; -: negative control. (b) PCR detection results of three CIAV isolates. Note: M: DNA Marker (DL 2000); 1~3: GX21121, GX21122, and GX21123 viruses isolated from 3 liver samples; -: negative control; +: positive control. (c) Four fragments of GX21121 gene amplification electrophoresis results. Note: M: DNA Marker (DL 2000); CIAV①~④: Four amplified fragments with 4 primers. CIAV GX21121 DNA simulation map by SnapGene software. Note: The yellow arrow part is the coding region, encoding 3 proteins: VP1, VP2, VP3.
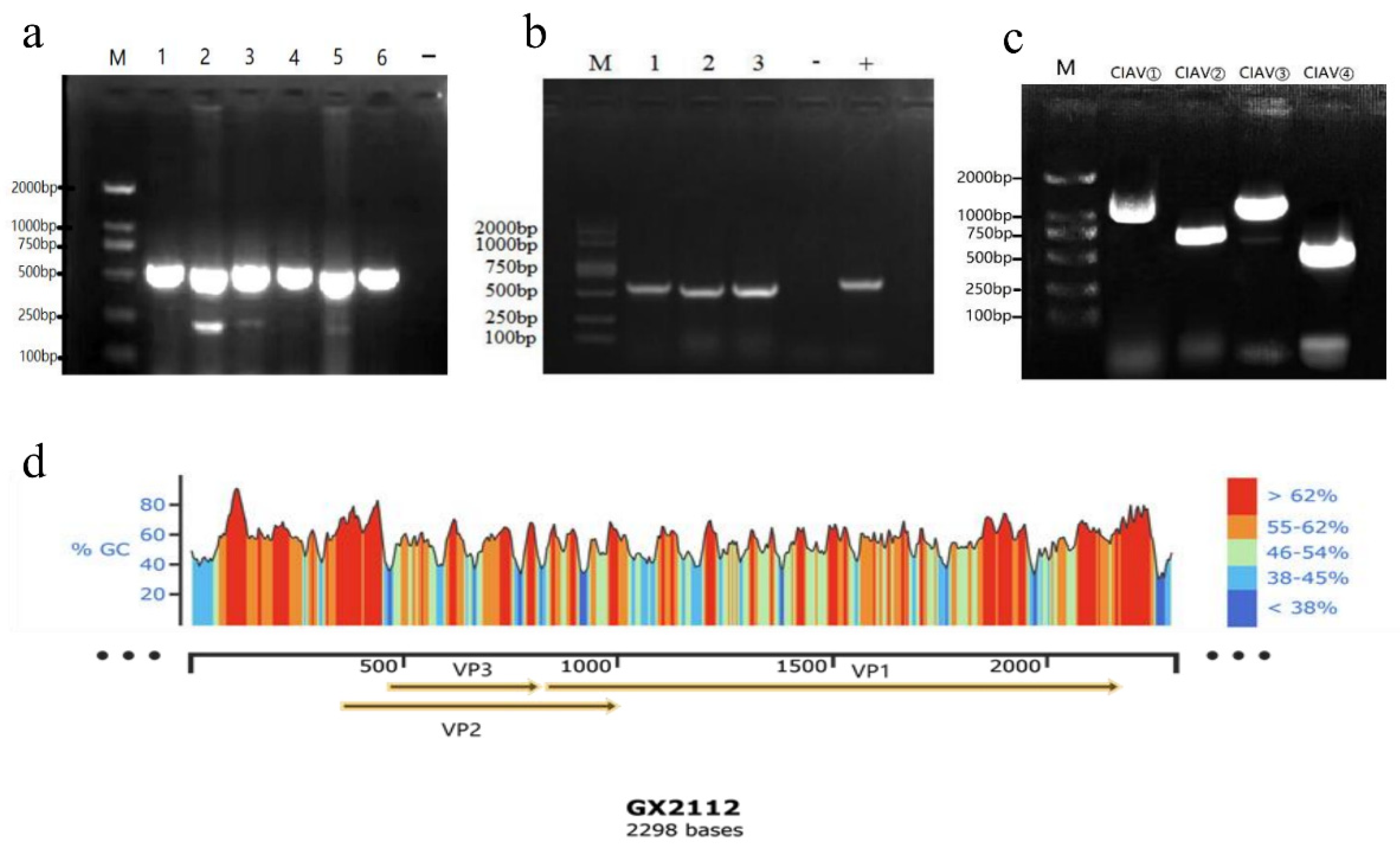
Figure 3.
a-h. Pathological examination of experimental SPF chicks revealed significant differences between the infected chicks and the NC, The infected chicks exhibited notable signs of depression and rough feathers compared to the NC (a, b); The EG displayed scattered subcutaneous hemorrhagic spots and pale muscles, which were not observed in the NC (c,d); The infected chicks also showed a yellowish appearance in the liver with surface foci of necrosis, while the NC did not exhibit such lesions (e,f). The kidneys of the infected chicks appeared pale, without any apparent lesions observed in the NC (g,h). Note: dpi refers to days post-inoculation, EG refers to the experimental group, and NC refers to the negative control.
Figure 3.
a-h. Pathological examination of experimental SPF chicks revealed significant differences between the infected chicks and the NC, The infected chicks exhibited notable signs of depression and rough feathers compared to the NC (a, b); The EG displayed scattered subcutaneous hemorrhagic spots and pale muscles, which were not observed in the NC (c,d); The infected chicks also showed a yellowish appearance in the liver with surface foci of necrosis, while the NC did not exhibit such lesions (e,f). The kidneys of the infected chicks appeared pale, without any apparent lesions observed in the NC (g,h). Note: dpi refers to days post-inoculation, EG refers to the experimental group, and NC refers to the negative control.

Figure 4.
(A) Genome-wide phylogenetic tree; (B) Phylogenetic tree of VP1; (C, E)The result of the Bootstrap algorithm in RDP recombination analysis for GX21122 and GX21123, the pink area represents the “Tract of sequence with a recombinant origin” indicating the predicted region where recombination may have occurred. The intertwined area of multiple sequences represents the predicted potential recombination site or fragment; (D, F) The result of the Simplot algorithm in SimPlot recombination analysis for GX21122 and GX21123.
Figure 4.
(A) Genome-wide phylogenetic tree; (B) Phylogenetic tree of VP1; (C, E)The result of the Bootstrap algorithm in RDP recombination analysis for GX21122 and GX21123, the pink area represents the “Tract of sequence with a recombinant origin” indicating the predicted region where recombination may have occurred. The intertwined area of multiple sequences represents the predicted potential recombination site or fragment; (D, F) The result of the Simplot algorithm in SimPlot recombination analysis for GX21122 and GX21123.
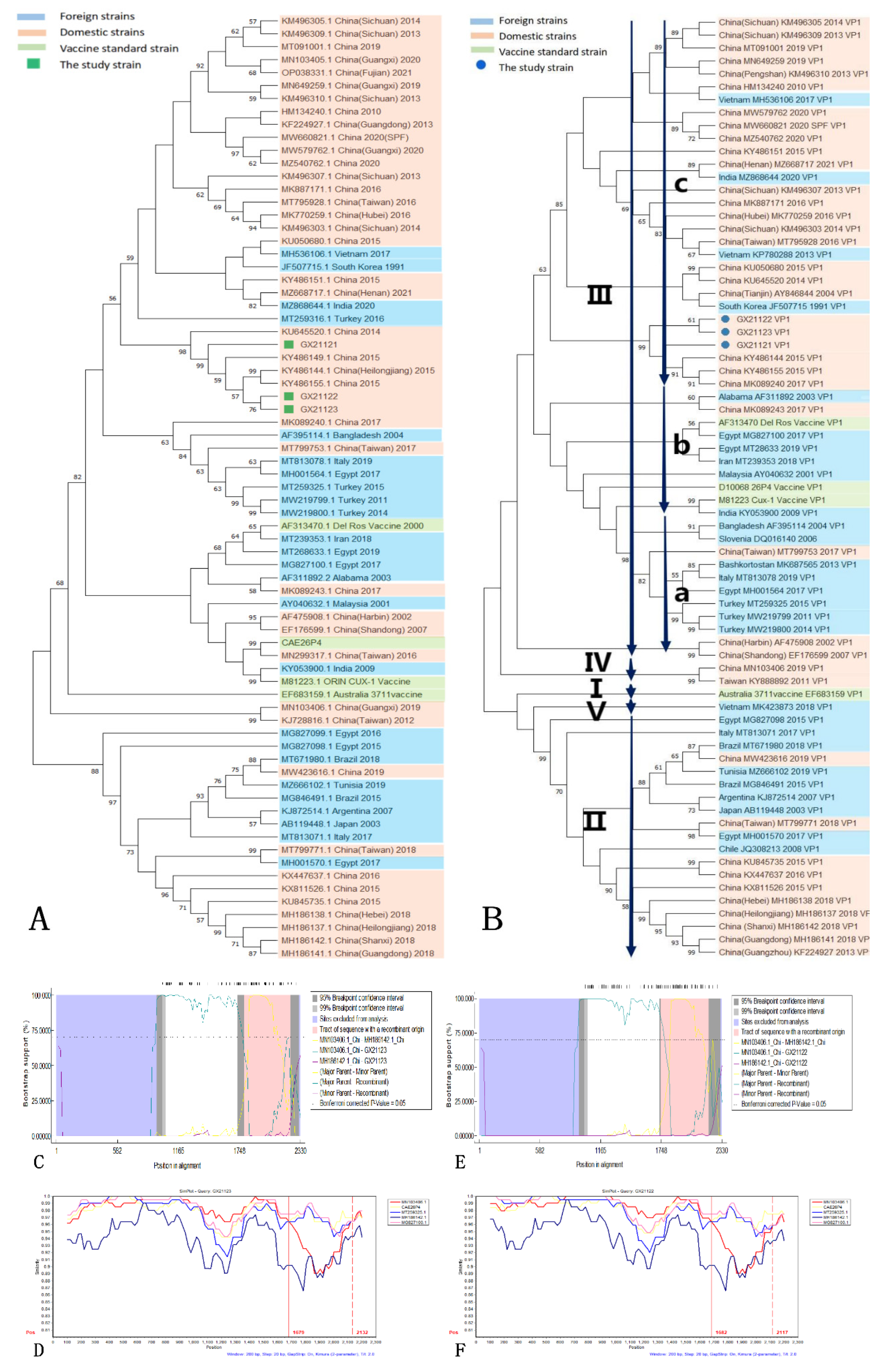

Table 1.
Primers used for CIAV DNA detection and whole genome amplification.
| Primers | (Primer sequence 5’ to 3’) | Length |
|---|---|---|
| CIAV ①-F | GACCGATCAACCCAAGCCTCC | 1010 bp |
| CIAV ①-R | ATCTTCCCGGTCGCATAAGCA | |
| CIAV ②-F | CTTGCCGGTTCTTTAATCACCCT | 751 bp |
| CIAV ②-R | CTCTTACCCAGCTGCCACACC | |
| CIAV ③-F | CTACATGGCAGCACCCGCATC | 1055 bp |
| CIAV ③-R | TCCGGCACATTCTTA(G)AAACCAG | |
| CIAV ④-F | AATGAACGCTCTCCAAGAAG | 540 bp |
| CIAV ④-R | AGCGGATAGTCATAGTAGAT |
Table 2.
Whole genome amplification reaction conditions.
| primers | pre-denatured | denatured×35 | Annealing×35 | extension×35 | post elongation |
|---|---|---|---|---|---|
| CIAV ① | 95 ℃, 5 min | 94 ℃, 30 s | 61 ℃, 30 s | 72 ℃, 70 s | 72 ℃, 7 min |
| CIAV ② | \ | \ | 62 ℃, 30 s | 72 ℃, 45 s | 72 ℃, 7 min |
| CIAV ③ | \ | \ | 58.7 ℃, 30 s | 72 ℃, 70 s | 72 ℃, 7 min |
| CIAV ④ | \ | \ | 50.5 ℃, 30 s | 72 ℃, 30 s | 72 ℃, 8 min |
Table 3.
Recombination statistics of GX21122 and GX21123 using RDP 4.0.
| Method | Recombination p value |
|---|---|
| RDP | 1.403×10-02 |
| GENECONV | 2.624×10-02 |
| BootScan | 3.598×10-02 |
| MaxChi | 4.106×10-03 |
| Chimaera | 1.071×10-02 |
| SiScan | 2.675×10-04 |
| 3Seq | 6.864×10-03 |
*The recombination events showing p value<0.05 were regarded reliable.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



