
Submitted:
02 June 2023
Posted:
06 June 2023
You are already at the latest version
Abstract
Sperm cells must undergo a complex maturation process after ejaculation to be able to fertilize an egg. One component of this maturation is hyperpolarization of the membrane potential to a more negative value. The ion channel responsible for this hyperpolarization, SLO3, was first cloned in 1998, and since then much progress has been made to determine how the channel is regulated and how its function intertwines with various signaling pathways involved in sperm maturation. Although SLO3 was originally thought to be present only in the sperm of mammals, recent evidence suggests that a primordial form of the gene is more widely expressed in some fish species. As with many reproductive genes, SLO3 is rapidly evolving with low conservation between closely related species and different regulatory and pharmacological profiles. Despite these differences, SLO3 appears to have a conserved role in regulating sperm membrane potential and driving large changes in response to stimuli. As with the differences in regulation of the channel, the effect of this hyperpolarization of the membrane potential may vary among mammalian species. Recent discoveries have elucidated the role of SLO3 in these processes in human sperm and provided tools to target the channel to affect human fertility.
Keywords:
membrane hyperpolarization
; SLO3
; contraception
; potassium channels
; sperm
; acrosomal exocytosis
; capacitation
; hyperactivated motility
; SLO channels
; male fertility
1. Introduction
Sperm have a long and complex maturation process that completes after they are expelled from the body in which they are produced. This post-ejaculatory process gives sperm the capacity to fertilize an oocyte and thus is termed capacitation [1,2]. Capacitation occurs in the female genital tract and involves many molecular changes including increases in cyclic AMP, protein tyrosine phosphorylation [3], intracellular pH [4,5,6,7], potassium ion (K+) conductance [8], and intracellular calcium (Ca2+) concentration [7,9,10,11,12]. Additionally, the plasma membrane hyperpolarizes to a more negative potential [8,13,14,15,16,17]. These changes culminate in two major physiological changes. The sperm become hyperactive, which facilitates their release from the oviductal reservoir and helps them penetrate through the cumulus and extracellular matrix surrounding the egg (zona pellucida) [18,19]. Additionally, they undergo acrosomal exocytosis, which helps them penetrate the zona pellucida [20,21,22] and exposes binding sites that allow the sperm membrane to fuse with the membrane of the oocyte [23]. Each step of capacitation is required for normal sperm function, but how each step is regulated and regulates other steps has not been fully determined.
A key component of sperm capacitation in many species from marine invertebrates to mammals is changes in membrane potential [24,25,26]. Generally, membrane potential is determined by the differences in ion concentrations across a membrane and the selective permeability of that membrane to said ions. One of the most prominent ions for controlling membrane potential in sperm is K+. In 1987, K+-dependent transient membrane hyperpolarization was first reported in sea urchin sperm in response to a signal from the egg jelly [26] This hyperpolarization was later shown to also occur in murine and bovine sperm and to be associated with capacitation [13]. Like their mammalian counterparts, human sperm undergo a capacitation-associated hyperpolarization from approximately −40 mV [27] to approximately −58 mV [28].
Several lines of evidence suggest that defects in hyperpolarization can result in infertility. For example, failure to hyperpolarize was correlated with a failure to undergo acrosomal exocytosis in mice [13,25], indicating that sperm membrane hyperpolarization is a key event in sperm capacitation. In humans, electrophysiological studies of patients undergoing in vitro fertilization (IVF) or intra-cytoplasmic sperm injection (ICSI) revealed that ~10% of patients with subfertility have depolarized membrane potentials caused by K+ conductance abnormalities [29]. Sperm isolated from men with idiopathic infertility or asthenozoospermia had a significantly more depolarized membrane potential than those from men with normal fertility [30]. Furthermore, capacitated sperm are more hyperpolarized than non-capacitated sperm [24,31]. In 2020, two groups independently used flow cytometry to quantitate membrane potential in sperm from normozoospermic donors and showed that the ability of sperm to hyperpolarize in capacitating conditions correlated with hyperactivation of motility, acrosomal exocytosis, and success in IVF [32,33].
Given the importance of sperm membrane potential in capacitation and fertilization, many researchers have focused on identifying the responsible K+ channels. In this review, we describe the evidence that the K+ channel SLO3 regulates sperm membrane potential hyperpolarization during capacitation in several species. Additionally, we highlight several challenges that made it difficult to determine whether or not SLO3 is likewise responsible for hyperpolarization in human sperm. Finally, we present new compelling genetic and pharmacologic data confirming that SLO3 is required for human sperm capacitation and fertility.
2. Potassium channels in sperm
Sperm from different species are exposed to drastically different environments, from sea water to unique genital tracts. This means that although hyperpolarization is conserved in many species, the mechanisms that drive it must be tailored to the specific environment. One species in which this mechanism has been explored in depth is the mouse. In 1998 the sperm-specific potassium channel SLO3 was cloned by first conducting a low-stringency NCBI Blast query with a cDNA sequence of mouse Slo1 [34]. A short Slo3 expressed sequence tag was identified and used to screen a mouse testis cDNA library. Two overlapping cDNAs were isolated and joined to make a complete Slo3 coding region. A comparison of the SLO1 and SLO3 protein sequences showed that the two proteins had similar hydrophilicity profiles, a similar core resembling a voltage-gated K+ channel, and long cytoplasmic tail structures. However, the mouse Slo3 sequence was conspicuously missing two Ca2+-binding domains present in the Slo1 cytoplasmic tail, suggesting that the channel is activated by factors other than Ca2+. Instead, the SLO3 channel was found to be activated by intracellular alkalinization.
After the discovery of the SLO3 channel, a potassium current was identified in mouse sperm that shared several key features with the channel [35]. This current, dubbed IKSper, was found to be activated by intracellular pH. The magnitude of the current meant that it was capable of driving large changes in membrane potential [8,35]. These traits matched those of SLO3, and it was confirmed that SLO3 was responsible for IKSper when a SLO3 knockout mouse was generated [35,36,37,38]. Deletion of SLO3 completely abolished the IKSper current. Additionally, sperm from Slo3 knock-out mice lack the hyperpolarization that occurs during capacitation. These sperm also lack the resulting Ca2+ influx through CatSper, the primary Ca2+ channel in sperm that is also necessary for fertility. [36,42]. As a result, Slo3 knockout mice are completely male infertile.
The Slo3 gene appears to have arisen by gene duplication of its close paralogue, Slo1 [43,44,45]. In some non-mammalian animals such as the spotted gar fish (Lepisosteus oculatus) [46] and members of the Salmonidae family, SLO3 has a broad tissue distribution similar to that of SLO1[47]. However, in mammals, its broad tissue distribution has been lost and in all mammalian species where it has been studied, including mice, bovines, and humans, SLO3 is only expressed in sperm. Additionally, as is the case with many reproduction-related genes, SLO3 has evolved rapidly. SLO3 shows only 60% protein sequence identity between mouse, human, and bovine (Figure 1), whereas the SLO1 protein sequence is 90% identical across the same species [48]. Moreover, SLO3 has accumulated species-specific features, which we will discuss later in this review.
3. Structure and gating of the SLO3 pore-forming subunits
The pore-forming components of SLO channels are formed by homo-tetramers of α-subunits. Generally, SLO family α-subunits resemble those of voltage-gated K+ channels in having transmembrane domains symmetrically arranged around a water-filled, K+ selective pore. However, SLO2.1 and SLO2.2 channels have six transmembrane domains (S1-S6) and thus have intracellular N- and C-termini as is common with members of the voltage-gated K+ channel family (Figure 2) [49,50]. In contrast, SLO1 and SLO3 have seven transmembrane domains (S0-S6) and thus have extracellular N-termini.
The cytosolic domains of SLO family channels contain two regulators of K+ conductance (RCK) domains, RCK1 and RCK2. These domains sense several intracellular signals and confer each subfamily with distinctive properties [51,52,53,54,55,56]. For example, in SLO1, both RCK1 and RCK2 contain Ca2+ sensors [56,57,58]. The “calcium bowl” in RCK2 is composed of a highly conserved string of aspartate residues, which are negatively charged [59]. SLO2.1 and SLO2.2 are also modulated by Na+, Cl-, and activation of G-protein-coupled receptors [49,50,60,61,62,63]. The cytosolic domain may also be a point of interaction between monomers of different SLO family channels.
The gating of SLO3 is similar to that of SLO1, as the opening of both channels is allosterically regulated by movement of a voltage sensor. This movement is driven by transmembrane potential and conformational change of the cytosolic gating ring induced by intracellular ligand binding. However, there are two important differences between the sensitivities of SLO1 and SLO3 channels to ligands. First, SLO1 is activated by acidification, whereas SLO3 is activated by alkalization [34,64]. Second, SLO1 has several Ca2+ binding sites and is activated by a broad range of Ca2+ concentrations [56,59,64]. Because of this, SLO1 can function over a broad range of voltages. In contrast, mouse SLO3 is insensitive to calcium and human SLO3 is several orders of magnitude less sensitive to Ca2+ than SLO1 [65] and functions in a narrow voltage range near the sperm resting potential.
SLO1 and SLO3 are both sensitive to pH. SLO1 has two histidine residues in the gating ring, which may act as proton sensors and open the channel in response to low intracellular pH [64]. The mechanism of pH modulation of SLO3 is unknown. The half-activation point of SLO3 by pH is estimated to be around 7.7 [66], which is close to the pKa of histidine. With the recently solved structures of the human SLO3 gating ring [67] and complete SLO1 channel [68,69,70], the key residues governing SLO3 regulation by protons should be revealed soon.
4. Structure and function of the SLO3 β and γ subunits
SLO1 and SLO3 α subunits are associated with several accessory subunits that regulate their expression and biophysical properties [71,72,73]. The α subunit of SLO1 channels is usually associated with auxiliary β- and γ-subunits [74,75,76,77,78], including β1-4 and γ1-2. These auxiliary subunits influence channel pharmacological and gating properties.
An important regulator of SLO3 is γ2, also known as leucine-rich-repeat-containing protein 52 (LRRC52) [79]. This subunit is abundantly expressed in the testis and predominantly interacts with SLO3. It is composed of a single transmembrane segment with an N-terminal extracellular peptide and a short cytoplasmic C-terminal tail [80]. Expression of γ2 depends on SLO3 expression as Slo3-/- mice do not express measurable γ2. The primary effect of γ2 is to shift the activation of SLO3 to more negative potentials, as seen in mouse, rat, and human channels [67,80,81]. Importantly, mouse SLO3 currents in the presence of γ2 activate at membrane voltage ranges similar to those of IKSper [80]. The importance of γ2 for SLO3 activity is evident in γ2-/- knockout mice, as IKSper currents in sperm from γ2-/- mice activate slower and their activation curve is shifted to more positive potentials than the currents from wild-type sperm. Additionally, alkaline pH is less able to hyperpolarize γ2-/- sperm than wild-type sperm. As a consequence of all this, the γ2-/- male mice are severely subfertile [82], indicating that the γ2 subunit is essential for male fertility.
Mouse and human SLO3 somewhat vary in their responses to γ2. For example, functional expression of human SLO3 requires γ2, and the activation rate and pH-sensitivity of human SLO3 channels expressed in Xenopus oocytes are increased by γ2. Although mouse SLO3 can be heterologously expressed without γ2, it has increased expression when this subunit is present [65,67,80,83]. Additionally, mouse SLO3 currents display a minor increase in pH sensitivity and activates at more negative potentials [67] when co-expressed with γ2. Nonetheless, γ2 is an important regulator of mouse SLO3. This was shown by expressing mouse SLO3 in Xenopus oocytes. Currents obtained from expressing mouse SLO3 alone exhibited a different pH- and voltage dependence than IKSper. However, when co-expressed with γ2, SLO3 produced currents that resembles the native IKSper currents [80].
Unlike γ2, γ1 (LRRC26), γ3 (LRRC55), and γ4 (LRRC38) are minimally expressed in the testis [78,80]. Co-expression of γ1 or γ4 with mouse SLO3 in Xenopus oocytes yielded a slight shift towards activation at more negative potentials [80]. Co-expression of γ3 had no effect on mouse or rat SLO3 currents [80]. Thus, γ1, γ3, and γ4 do not appear to play a substantial role in SLO3 regulation.
Because SLO1 is regulated by β subunits, the effects of these subunits on SLO3 has been examined. β1-3 are minimally expressed in the mouse testis and do not appear to functionally regulate SLO3 [76,84,85]. It has even been shown that replacing the mouse SLO1 tail with that of mouse SLO3 ablates the effect of β1 on the channel [86]. Moreover, if co-expressed in Sf9 cells, β1, β3a, and β3b all immunoprecipitate with mouse SLO3 but do not affect the channel gating [85]. β4 is expressed in mouse testis, and co-expression of β4 can increase surface expression, macro-conductance, and activation kinetics of mouse SLO3 channels in Xenopus oocytes [85]. This indicates that only β4 selectively modulates SLO3 expression and function. In humans, β3 and β4 mRNAs are both expressed in the testis [75] but little is known regarding the effects of β subunits on human SLO3.
5. Challenges in determining whether SLO3 is responsible for human sperm hyperpolarization
Both mouse and human sperm undergo hyperpolarization that is required for capacitation. Moreover, mouse and human sperm both have similar K+ currents, IKSper and hKSper, respectively. Although SLO3 was conclusively demonstrated to be responsible for mouse sperm hyperpolarization in 2010 [36], four key challenges have impeded our understanding of whether SLO3 is responsible for hyperpolarization in human sperm. These include differences in pH and Ca2+ regulation, voltage sensitivity, functional relationships with CatSper, and pharmacology.
2.1. Challenge 1: Differences in pH and Ca2+ regulation
Although IKSPer and hKSper have several similarities, these currents have important differences, including that hKSper is less sensitive to pH than is IKSper. Additionally, unlike the IKSper current, the hKSper current is sensitive to intracellular Ca2+ [65,87]. Given these reports, Mannowetz et al. suggested that hKSper was conducted by SLO1 rather than by SLO3 [67]. Later, Brenker et al. [65] showed that human SLO3 expressed in Xenopus oocytes and Chinese hamster ovary cells is sensitive to both Ca2+ and pH. The Ca2+ concentrations required to activate SLO3 in both sperm and heterologously expressed in cells were similar (60-1000 µM) and higher than those needed to activate SLO1 [56,65]. Due to these findings these investigators concluded that SLO3 is likely responsible for the hKSper current.
SLO3 from mice [67,88,89], rats [81], bovines [88], and humans [65] are activated by intracellular alkalinization, though the origin of this pH sensitivity is unresolved. Mouse SLO3 channels are more pH-sensitive than bovine channels and contain two histidine residues in the RCK1 domain, whereas bovine channels have only one histidine [88]. However, substituting one of the histidines in the mouse channel did not significantly alter its pH sensitivity, suggesting that this region is not central to pH regulation. We recently identified a natural structural variant (C382R) of human SLO3 with enhanced pH sensitivity [48]. This variant inserts a positive charge in the -B helix of the N-lobe in RCK1, a critical region likely to interact with the pore-gating region of the channel and may enhance the effectiveness of transduction between the gating ring and the cytoplasmic pore-gating domain. This variant was the third most common single nucleotide polymorphism (SNP) in SLO3 in the NIH SNP database, possibly indicating positive selection pressure. However, the effect of this SNP on fertility is unknown [48].
2.2. Challenge 2: Differences in voltage sensitivity
The voltage range of activation of the alpha subunit of SLO3 expressed by itself in heterologous systems varies widely between species. Bovine SLO3 activates at more negative potentials than mouse SLO3. The half-maximal activation of conductance of mouse is at +70-77 mV [34,88] and near +0.5 mV in bovines [88]. Our lab generated a series of chimeric constructs between bovine and mouse α-subunits to map the functional domain responsible for this difference [88]. Our data revealed that the amino acid residues determining this property were distributed over a loop of the RCK1 domain. This region contains the greatest sequence and length polymorphisms among RCK1 domains of SLO3 channels from many species. This region also has unusually low sequence conservation between bovine and mouse SLO3 channels. Specifically, we found that introducing this region from the bovine channel into the mouse conferred a similar voltage sensitivity as in the bovine channel, but the reverse was not true. This implies that the RCK1 domain of mouse SLO3 is important for voltage sensitivity, but the bovine channel contains other unknown regulatory domains. Expression of the human SLO3 alpha subunit by itself does not result in measurable whole-cell currents. Human SLO3 whole-cell currents can only be obtained in the presence of the human γ2 subunit [65,83,90]. The human SLO3 channel, even in the presence of the γ2 subunit activates at more positive potentials than mouse or bovine SLO3 in Xenopus oocytes [65].
2.3. Challenge 3: Differences in functional relationship with CatSper
In mouse sperm capacitation, the role of SLO3 is extensively characterized (Figure 3). During capacitation, two major ion channels become active, the SLO3 K+ channel and the CatSper Ca2+ channel. Both channels are essential for fertilization, as knocking out either of them confers male infertility [34,35,36,38,42,91,92,93,94,95]. It is accepted in the field that the increase in intracellular Ca2+ necessary to promote hyperactivated motility in the sperm is conducted by CatSper channels [7,35,38,42]. Furthermore, this activation of CatSper seems to be subsequent to and dependent on SLO3 activation, as CatSper activation in Slo3-/- mice is impaired and can be rescued by alkaline depolarization [11]. A working hypothesis suggests that when sperm are exposed to bicarbonate and an alkaline external pH in the female genital tract, it initially results in an increase in sperm intracellular pH, possibly through the activation of the sperm-specific Na+/H+ exchanger (sNHE) (unpublished data). This rise in intracellular pH activates SLO3 channels, leading to cell hyperpolarization due to potassium efflux. This hyperpolarization can potentially enhance the driving force and influx of calcium ions through CatSper channels, which exhibit a weak voltage-dependence [35]. Additionally, it may further activate sNHE, which contains a putative voltage sensor that could be activated by membrane hyperpolarization [11]. Activation of the sNHE would increase intracellular pH, further stimulating CatSper, SLO3 and establishing a positive feedback loop. Notably, recent results from Chávez et al. in 2020 found more evidence that membrane hyperpolarization induced an increase in intracellular pH of individual sperm [96]. Ultimately, the activation of CatSper channels triggers calcium influx, leading to the necessary elevation in intracellular calcium concentration for sperm hyperactivation.
In human sperm the temporal sequence of activation of these channels is debated (Figure 3). Because human SLO3 is activated by intracellular Ca2+ it was suggested that SLO3 is activated downstream of CatSper in human sperm [97,98]. Contrary to this model, it has been shown that human sperm undergo transient Ca2+ oscillations that inactivate CatSper and are inhibited by hyperpolarization [99,100]. According to this SLO3 activity precedes CatSper activity in human sperm as it does in mouse sperm. More experiments need to be done to determine the exact sequence of events in human sperm.
The precise details of the SLO3 and CatSper interaction may differ in other ways between mouse and human sperm. For instance, even though sNHE is expressed in human sperm, the proton channel Hv1 has been suggested to have a significant role in the alkalinization of human sperm [101,102]. Hv1 is activated by a combination of the pH gradient and membrane depolarization, and it may also be activated by the removal of zinc after ejaculation and during sperm passage through the female genital tract [101,103]. Activation of this channel could be crucial in raising intracellular sperm pH and subsequently facilitating the activation of CatSper channels. However, in mouse sperm, the Hv1 proton channel does not appear to play a significant role, as the Hv1 knockout mice maintain their fertility [104]. Progesterone at low concentrations activates human CatSper currents however it does not have an effect on the mouse currents [65].
2.4. Challenge 4: Differences in pharmacology
Both mouse and human SLO3 exhibit similar responses to certain inhibitors. For example, low concentrations of Ba2+ have been found to inhibit mouse SLO3 currents [24,105,106]. This inhibition also extends to hyperpolarization of CHO cells transfected with human SLO3 and human sperm during capacitation [83]. Additionally, high concentrations of TEA have comparable effects on both human and mouse SLO3 currents [83,105,106]. However, it's important to note that low concentrations of TEA do not hinder human sperm hyperpolarization during capacitation [83] and have no impact on hKSper currents [65] or human SLO3 in transfected HEK-293 cells [107]. This indicates that, similar to mouse SLO3, human SLO3 is only inhibited by high concentrations of TEA. Another widely used potassium channel blocker, 4-AP, does not exhibit inhibitory effects on either human or mouse SLO3 when applied externally [108,109]. In contrast, two other inhibitors, quinidine and clofilium, demonstrate strong inhibition of both mouse [105,106,108] and human SLO3 [65,109]. Despite these similarities, mouse and human SLO3 differ in their responses to several other inhibitors.
The first study that examined hKSper currents in human sperm revealed that the current was inhibited by the canonical SLO1 inhibitors charybdotoxin, iberiotoxin, and paxilline [87]. Although later studies confirmed some of these findings, others have presented conflicting results. For example, flow cytometry measurements of membrane potential showed that charybdotoxin and iberiotoxin each strongly inhibited human sperm hyperpolarization during capacitation [24,83]. However, neither drug blocked more than 50% of currents in heterologously expressed human SLO3 currents in CHO cells [83]. Later recordings showed no effect of Iberiotoxin on hKSper in sperm [65] or on human SLO3 currents heterologously expressed in HEK cells [107]. These discrepancies may be caused by different recording conditions used [65,87]. We recently measured the effect of iberiotoxin on human SLO3 expressed in HEK-293 cells and found that it inhibited SLO3 at about 20-fold higher concentrations than SLO1 and had a much wider inhibition curve, making its inhibition of SLO3 currents highly susceptible to different recording conditions [90]. Paxilline on the other hand maintained a strong selectivity for SLO1 over human SLO3, as was observed in mice, making it a more useful tool for comparing the inhibition of SLO1 or SLO3 channels across species [90,108].
Both clofilium and quinidine consistently inhibit hKSper currents under physiological conditions [65,109], leading to the prevention of hyperactivation and hyperpolarization of human sperm [65,83,109]. However, determining the extent of SLO3 channel inhibition by quinidine and clofilium in sperm, as well as their impact on sperm physiological properties, is challenging due to the presence of CatSper in these cells [110]. CatSper channels can conduct large currents carried by monovalent cations, which may contribute to the total outward K+ current measured in sperm and potentially be mistaken for SLO3 currents. Consequently, quinidine and clofilium might also inhibit CatSper channels [38]. Supporting this notion, Mansell et al. reported that clofilium and quinidine inhibit CatSper channels in human sperm [109]. The off-target effects of these inhibitors on CatSper, as well as their lack of selectivity against other potassium channels, could contribute to their effects on hyperpolarization and hyperactivation. To avoid this confusion, the application of divalent ions, such as 2 mM or higher Ca2+, in the external solution can significantly inhibit CatSper from carrying such currents [92,111].
Additionally, studying these channels in heterologous systems can help characterize the inhibitory effects of these drugs on specific channels. For instance, it has been demonstrated that clofilium inhibits human SLO3 currents expressed in CHO cells [24].
Making the situation more complex, inhibitors of CatSper channels can also exert inhibitory effects on SLO3 channels. For example, Mibefradil, a CatSper inhibitor, shows slight and reversible inhibition of mouse SLO3 [106].
Therefore, a dual approach should be employed to characterize the inhibitory effects of these drugs. This includes investigating the inhibitory effects of the drugs on heterologously expressed channels as well as channels recorded in native cells. Fortunately, such a dual approach is feasible for SLO3 channels, which are well-expressed in both Xenopus oocytes and mammalian cell lines. However, achieving expression of CatSper channels in a heterologous system has not yet been accomplished, posing a challenge for their characterization. A summary of data on the effects of inhibitors on potassium currents in native cells and in heterologous systems are presented in Table 1 and Table 2 respectively.
6. Newly discovered variants and inhibitors confirm that SLO3 is responsible for human sperm hyperpolarization
Two recent papers provide genetic evidence supporting the role of SLO3 in human fertility. Lv et al. reported that a missense mutation and a splice variant of human SLO3 channels are associated with male infertility [94]. However, it should be noted that the male patient in the study presented with asthenozoospermia, a condition characterized by reduced or absent motile sperm [112]. This disorder is not known to be associated with SLO3-deficient mice, as these mice exhibit normal sperm count and motility [36]. Therefore, the presence of this condition suggests that other sperm functional defects unrelated to SLO3 function may have contributed to the infertility observed [94].
In a more compelling case implicating human SLO3 in infertility, a man carrying a missense mutation of the Slo3 gene (c.1237A>T: Ile413Phe) exhibited sperm that failed to hyperpolarize, undergo acrosome reaction, and achieve successful fertilization in in vitro fertilization (IVF) procedures [95]. However, in intracytoplasmic sperm injection (ICSI), where sperm capacitation is not required, fertilization was successful, as expected for a mutation in the Slo3 gene. To further confirm the role of SLO3 in this phenotype, the authors generated a mouse line in which the endogenous Slo3 gene carried the same missense mutation found in the affected men. These mice also exhibited infertility. These findings provide clear evidence that SLO3 is necessary for fertility in both humans and mice and suggest its conserved role in acrosomal exocytosis [95].
We recently described a new inhibitor, VU0546110, which is more than 40-fold selective for human SLO3 over SLO1 [90]. This inhibitor completely inhibited hKSper, confirming that SLO1 channels do not meaningfully contribute to the current. This inhibition also had physiological effects, significantly inhibiting hyperpolarization, hyperactivation, and the acrosome reaction in human sperm. These downstream effects provide further evidence that human SLO3 is necessary for sperm hyperpolarization and fertility.
7. Conclusions and future directions
The two paralogues Slo1 and Slo3 apparently have a common ancestor that underwent a gene duplication event approximately 450 million years. Since then, the older gene, Slo1, has remained highly conserved in species as diverse as the nematode C. elegans worm and homo sapiens. Conversely, Slo3 has been subject to wide evolutionary divergence, functioning in many tissue types in the spotted gar [47] but only functioning in spermatozoa in other species [34]. As a sperm-specific gene in mammals, Slo3 continues to rapidly evolve to acquire new functional properties, such as Ca2+-dependent activation in human sperm [65]. In mammals, SLO3 K+ channels hyperpolarize sperm to ensure that Ca2+ and possibly also pH reach optimal internal values to drive hyperactivation and regulated acrosomal exocytosis. These actions are essential for sperm to reach and penetrate an oocyte. Because SLO3 plays such an important role in male fertility, it is becoming the focus of studies of possible causes of male infertility and is being targeted to develop new contraceptives.
Author Contributions
Conceptualization, M.D.L and C.S.M.; writing—original draft preparation, M.D.L., J.J.F., P.L, S.B., A.B., K.A., M.P., and C.S.M.; writing—review and editing, M.D.L. and C.S.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors would like to thank Deborah J. Frank and Lawrence Salkoff for editing this manuscript and Anthony Bartley for preparation of Figure 2.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chang, M.C. Fertilizing Capacity of Spermatozoa Deposited into the Fallopian Tubes. Nature 1951, 168, 697–698. [Google Scholar] [CrossRef]
- AUSTIN, C.R. Observations on the Penetration of the Sperm in the Mammalian Egg. Aust J Sci Res B 1951, 4, 581–596. [Google Scholar] [CrossRef]
- Visconti, P.E.; Moore, G.D.; Bailey, J.L.; Leclerc, P.; Connors, S.A.; Pan, D.; Olds-Clarke, P.; Kopf, G.S. Capacitation of Mouse Spermatozoa. II. Protein Tyrosine Phosphorylation and Capacitation Are Regulated by a CAMP-Dependent Pathway. Development 1995, 121, 1139–1150. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Oberdorf, J.A.; Florman, H.M. PH Regulation in Mouse Sperm: Identification of Na+-, Cl−-, and[Formula]Dependent and Arylaminobenzoate-Dependent Regulatory Mechanisms and Characterization of Their Roles in Sperm Capacitation. Dev Biol 1996, 173, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Vredenburgh-Wilberg, W.L.; Parrish, J.J. Intracellular PH of Bovine Sperm Increases during Capacitation. Mol Reprod Dev 1995, 40, 490–502. [Google Scholar] [CrossRef] [PubMed]
- Breitbart, H.; Breitbart, H. Signaling Pathways in Sperm Capacitation and Acrosome Reaction. 2003.
- Ferreira, J.J.; Lybaert, P.; Puga-Molina, L.C.; Santi, C.M. Conserved Mechanism of Bicarbonate-Induced Sensitization of CatSper Channels in Human and Mouse Sperm. Front Cell Dev Biol 2021, 9, 2614. [Google Scholar] [CrossRef] [PubMed]
- Chávez, J.C.; de la Vega-Beltrán, J.L.; Escoffier, J.; Visconti, P.E.; Treviño, C.L.; Darszon, A.; Salkoff, L.; Santi, C.M. Ion Permeabilities in Mouse Sperm Reveal an External Trigger for SLO3-Dependent Hyperpolarization. PLoS One 2013, 8, e60578. [Google Scholar] [CrossRef] [PubMed]
- DasGupta, S.; Mills, C.L.; Fraser, L.R. Ca2+-Related Changes in the Capacitation State of Human Spermatozoa Assessed by a Chlortetracycline Fluorescence Assay. J Reprod Fertil 1993, 99, 135–143. [Google Scholar] [CrossRef]
- BALDI, E.; CASANO, R.; FALSETTI, C.; KRAUSZ, C.; MAGGI, M.; FORTI, G. Intracellular Calcium Accumulation and Responsiveness to Progesterone in Capacitating Human Spermatozoa. J Androl 1991, 12, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Chávez, J.C.; Ferreira, J.J.; Butler, A.; De La Vega Beltrán, J.L.; Treviño, C.L.; Darszon, A.; Salkoff, L.; Santi, C.M. SLO3 K+ Channels Control Calcium Entry through CATSPER Channels in Sperm. Journal of Biological Chemistry 2014, 289, 32266–32275. [Google Scholar] [CrossRef]
- Ferreira, J.J.; Cassina, A.; Irigoyen, P.; Ford, M.; Pietroroia, S.; Peramsetty, N.; Radi, R.; Santi, C.M.; Sapiro, R. Increased Mitochondrial Activity upon CatSper Channel Activation Is Required for Mouse Sperm Capacitation. Redox Biol 2021, 48, 102176. [Google Scholar] [CrossRef]
- Zeng, Y.; Clark, E.N.; Florman, H.M. Sperm Membrane Potential: Hyperpolarization during Capacitation Regulates Zona Pellucida-Dependent Acrosomal Secretion. Dev Biol 1995, 171, 554–563. [Google Scholar] [CrossRef]
- Santi, C.M.; Orta, G.; Salkoff, L.; Visconti, P.E.; Darszon, A.; Treviño, C.L. K+ and Cl- Channels and Transporters in Sperm Function. Curr Top Dev Biol 2013, 102, 385–421. [Google Scholar] [CrossRef]
- Arnoult, C.; Kazam, I.G.; Visconti, P.E.; Kopf, G.S.; Villaz, M.; Florman, H.M. Control of the Low Voltage-Activated Calcium Channel of Mouse Sperm by Egg ZP3 and by Membrane Hyperpolarization during Capacitation. Proc Natl Acad Sci U S A 1999, 96, 6757–6762. [Google Scholar] [CrossRef]
- Muñoz-Garay, C.; de la Vega-Beltrán, J.L.; Delgado, R.; Labarca, P.; Felix, R.; Darszon, A. Inwardly Rectifying K+ Channels in Spermatogenic Cells: Functional Expression and Implication in Sperm Capacitation. Dev Biol 2001, 234, 261–274. [Google Scholar] [CrossRef] [PubMed]
- Gunderson, S.J.; Puga Molina, L.C.; Spies, N.; Balestrini, P.A.; Buffone, M.G.; Jungheim, E.S.; Riley, J.; Santi, C.M. Machine-Learning Algorithm Incorporating Capacitated Sperm Intracellular PH Predicts Conventional in Vitro Fertilization Success in Normospermic Patients. Fertil Steril 2021, 115, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Demott, R.P.; Suarez, S.S. Hyperactivated Sperm Progress in the Mouse Oviduct. Biol Reprod 1992, 46, 779–785. [Google Scholar] [CrossRef] [PubMed]
- SUAREZ, S.S.; DAI, X. -B; DEMOTT, R.P.; REDFERN, K.; MIRANDO, M.A. Movement Characteristics of Boar Sperm Obtained from the Oviduct or Hyperactivated In Vitro. J Androl 1992, 13, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Yanagimachi, R. Fertility of Mammalian Spermatozoa: Its Development and Relativity. Zygote 1994, 2, 371–372. [Google Scholar] [CrossRef] [PubMed]
- Buffone, M.G.; Rodriguez-Miranda, E.; Storey, B.T.; Gerton, G.L. Acrosomal Exocytosis of Mouse Sperm Progresses in a Consistent Direction in Response to Zona Pellucida. J Cell Physiol 2009, 220, 611–620. [Google Scholar] [CrossRef] [PubMed]
- Breitbart, H.; Rubinstein, S.; Lax, Y. Regulatory Mechanisms in Acrosomal Exocytosis. Rev Reprod 1997, 2, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Inoue, N.; Ikawa, M.; Okabe, M. The Mechanism of Sperm–Egg Interaction and the Involvement of IZUMO1 in Fusion. Asian J Androl 2011, 13, 81. [Google Scholar] [CrossRef] [PubMed]
- López-González, I.; Torres-Rodríguez, P.; Sánchez-Carranza, O.; Solís-López, A.; Santi, C.M.; Darszon, A.; Treviño, C.L. Membrane Hyperpolarization during Human Sperm Capacitation. Mol Hum Reprod 2014, 20, 619–629. [Google Scholar] [CrossRef]
- De La Vega-Beltran, J.L.; Sánchez-Cárdenas, C.; Krapf, D.; Hernandez-González, E.O.; Wertheimer, E.; Treviño, C.L.; Visconti, P.E.; Darszon, A. Mouse Sperm Membrane Potential Hyperpolarization Is Necessary and Sufficient to Prepare Sperm for the Acrosome Reaction. J Biol Chem 2012, 287, 44384–44393. [Google Scholar] [CrossRef] [PubMed]
- González-Martínez, M.; Darszon, A. A Fast Transient Hyperpolarization Occurs during the Sea Urchin Sperm Acrosome Reaction Induced by Egg Jelly. FEBS Lett 1987, 218, 247–250. [Google Scholar] [CrossRef]
- Linares-Hernández, L.; Guzmán-Grenfell, A.M.; Hicks-Gomez, J.J.; González-Martínez, M.T. Voltage-Dependent Calcium Influx in Human Sperm Assessed by Simultaneous Optical Detection of Intracellular Calcium and Membrane Potential. Biochim Biophys Acta Biomembr 1998, 1372, 1–12. [Google Scholar] [CrossRef]
- Patrat, C.; Serres, C.; Jouannet, P. Progesterone Induces Hyperpolarization after a Transient Depolarization Phase in Human Spermatozoa. Biol Reprod 2002, 66, 1775–1780. [Google Scholar] [CrossRef]
- Brown, S.G.; Publicover, S.J.; Mansell, S.A.; Lishko, P. V.; Williams, H.L.; Ramalingam, M.; Wilson, S.M.; Barratt, C.L.R.; Sutton, K.A.; Da Silva, S.M. Depolarization of Sperm Membrane Potential Is a Common Feature of Men with Subfertility and Is Associated with Low Fertilization Rate at IVF. Human Reproduction 2016, 31, 1147–1157. [Google Scholar] [CrossRef]
- Calzada, L.; Tellez, J. Defective Function of Membrane Potential (Ψ) on Sperm of Infertile Men. Arch Androl 1997, 38, 151–155. [Google Scholar] [CrossRef]
- Brukman, N.G.; Nuñez, S.Y.; Puga Molina, L. del C.; Buffone, M.G.; Darszon, A.; Cuasnicu, P.S.; da Ros, V.G. Tyrosine Phosphorylation Signaling Regulates Ca 2+ Entry by Affecting Intracellular PH during Human Sperm Capacitation. J Cell Physiol 2019, 234, 5276–5288. [Google Scholar] [CrossRef]
- Molina, L.C.P.; Gunderson, S.; Riley, J.; Lybaert, P.; Borrego-Alvarez, A.; Jungheim, E.S.; Santi, C.M. Membrane Potential Determined by Flow Cytometry Predicts Fertilizing Ability of Human Sperm. Front Cell Dev Biol 2020, 7, 387. [Google Scholar] [CrossRef]
- Baro Graf, C.; Ritagliati, C.; Torres-Monserrat, V.; Stival, C.; Carizza, C.; Buffone, M.G.; Krapf, D. Membrane Potential Assessment by Fluorimetry as a Predictor Tool of Human Sperm Fertilizing Capacity. Front Cell Dev Biol 2020, 7, 383. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, M.; Wei, A.; Yuan, A.; Gaut, J.; Saito, M.; Salkoff, L. Slo3, a Novel PH-Sensitive K+ Channel from Mammalian Spermatocytes. J Biol Chem 1998, 273, 3509–3516. [Google Scholar] [CrossRef] [PubMed]
- Navarro, B.; Kirichok, Y.; Clapham, D.E. KSper, a PH-Sensitive K+ Current That Controls Sperm Membrane Potential. Proc Natl Acad Sci U S A 2007, 104, 7688–7692. [Google Scholar] [CrossRef]
- Santi, C.M.; Martínez-López, P.; de la Vega-Beltrán, J.L.; Butler, A.; Alisio, A.; Darszon, A.; Salkoff, L. The SLO3 Sperm-Specific Potassium Channel Plays a Vital Role in Male Fertility. FEBS Lett 2010, 584, 1041–1046. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.H.; Yang, C.; Kim, S.T.; Lingle, C.J.; Xia, X.M. Deletion of the Slo3 Gene Abolishes Alkalizationactivated K+ Current in Mouse Spermatozoa. Proc Natl Acad Sci U S A 2011, 108, 5879–5884. [Google Scholar] [CrossRef]
- Zeng, X.H.; Navarro, B.; Xia, X.M.; Clapham, D.E.; Lingle, C.J. Simultaneous Knockout of Slo3 and CatSper1 Abolishes All Alkalization- and Voltage-Activated Current in Mouse Spermatozoa. Journal of General Physiology 2013, 142, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2—a Multiple Sequence Alignment Editor and Analysis Workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed]
- Troshin, P. V.; Procter, J.B.; Sherstnev, A.; Barton, D.L.; Madeira, F.; Barton, G.J. JABAWS 2.2 Distributed Web Services for Bioinformatics: Protein Disorder, Conservation and RNA Secondary Structure. Bioinformatics 2018, 34, 1939–1940. [Google Scholar] [CrossRef]
- Troshin, P. V.; Procter, J.B.; Barton, G.J. Java Bioinformatics Analysis Web Services for Multiple Sequence Alignment-JABAWS:MSA. Bioinformatics 2011, 27, 2001–2002. [Google Scholar] [CrossRef]
- Zeng, X.-H.; Yang, C.; Kim, S.T.; Lingle, C.J.; Xia, X.-M. Deletion of the Slo3 Gene Abolishes Alkalization-Activated K+ Current in Mouse Spermatozoa. Proc Natl Acad Sci U S A 2011, 108, 5879–5884. [Google Scholar] [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-Based Map of the Human Proteome. Science (1979) 2015, 347. [Google Scholar] [CrossRef] [PubMed]
- The Human Protein Atlas. Available online: https://www.proteinatlas.org/ (accessed on 18 July 2022).
- Macqueen, D.J.; Johnston, I.A. A Well-Constrained Estimate for the Timing of the Salmonid Whole Genome Duplication Reveals Major Decoupling from Species Diversification. Proceedings of the Royal Society B: Biological Sciences 2014, 281. [Google Scholar] [CrossRef] [PubMed]
- Braasch, I.; Gehrke, A.R.; Smith, J.J.; Kawasaki, K.; Manousaki, T.; Pasquier, J.; Amores, A.; Desvignes, T.; Batzel, P.; Catchen, J.; et al. The Spotted Gar Genome Illuminates Vertebrate Evolution and Facilitates Human-Teleost Comparisons. Nature Genetics 2016 48:4 2016, 48, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Vicens, A.; Andrade-López, K.; Cortez, D.; Gutiérrez, R.M.; Treviño, C.L. Premammalian Origin of the Sperm-Specific Slo3 Channel. FEBS Open Bio 2017, 7, 382–390. [Google Scholar] [CrossRef]
- Geng, Y.; Ferreira, J.J.; Dzikunu, V.; Butler, A.; Lybaert, P.; Yuan, P.; Magleby, K.L.; Salkoff, L.; Santi, C.M. A Genetic Variant of the Sperm-Specific SLO3 K+ Channel Has Altered PH and Ca2+ Sensitivities. J Biol Chem 2017, 292, 8978–8987. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, A.; Joiner, W.J.; Wu, M.; Yang, Y.; Sigworth, F.J.; Kaczmarek, L.K. Slick (Slo2.1), a Rapidly-Gating Sodium-Activated Potassium Channel Inhibited by ATP. J Neurosci 2003, 23, 11681–11691. [Google Scholar] [CrossRef]
- Yuan, A.; Santi, C.M.; Wei, A.; Wang, Z.W.; Pollak, K.; Nonet, M.; Kaczmarek, L.; Crowder, C.M.; Salkoff, L. The Sodium-Activated Potassium Channel Is Encoded by a Member of the Slo Gene Family. Neuron 2003, 37, 765–773. [Google Scholar] [CrossRef]
- Jiang, Y.; Pico, A.; Cadene, M.; Chait, B.T.; MacKinnon, R. Structure of the RCK Domain from the E. Coli K+ Channel and Demonstration of Its Presence in the Human BK Channel. Neuron 2001, 29, 593–601. [Google Scholar] [CrossRef]
- Peng Yuan; Leonetti, M. D.; Pico, A.R.; Hsiung, Y.; MacKinnon, R. Structure of the Human BK Channel Ca2+-Activation Apparatus at 3.0 Å Resolution. Science (1979) 2010, 329, 182–186. [Google Scholar] [CrossRef]
- Marty, A. Ca-Dependent K Channels with Large Unitary Conductance in Chromaffin Cell Membranes. Nature 1981 291:5815 1981, 291, 497–500. [Google Scholar] [CrossRef]
- Pallotta, B.S.; Magleby, K.L.; Barrett, J.N. Single Channel Recordings of Ca2+-Activated K+ Currents in Rat Muscle Cell Culture. Nature 1981 293:5832 1981, 293, 471–474. [Google Scholar] [CrossRef]
- Latorre, R.; Vergara, C.; Hidalgo, C. Reconstitution in Planar Lipid Bilayers of a Ca2+-Dependent K+ Channel from Transverse Tubule Membranes Isolated from Rabbit Skeletal Muscle. Proc Natl Acad Sci U S A 1982, 79, 805–809. [Google Scholar] [CrossRef]
- Schreiber, M.; Yuan, A.; Salkoff, L. Transplantable Sites Confer Calcium Sensitivity to BK Channels. Nature Neuroscience 1999 2:5 1999, 2, 416–421. [Google Scholar] [CrossRef]
- Xia, X.M.; Zeng, X.; Lingle, C.J. Multiple Regulatory Sites in Large-Conductance Calcium-Activated Potassium Channels. Nature 2002, 418, 880–884. [Google Scholar] [CrossRef]
- Geng, Y.; Deng, Z.; Zhang, G.; Budelli, G.; Butler, A.; Yuan, P.; Cui, J.; Salkoff, L.; Magleby, K.L. Coupling of Ca2+ and Voltage Activation in BK Channels through the AB Helix/Voltage Sensor Interface. Proc Natl Acad Sci U S A 2020, 117, 14512–14521. [Google Scholar] [CrossRef]
- Schreiber, M.; Salkoff, L. A Novel Calcium-Sensing Domain in the BK Channel. Biophys J 1997, 73, 1355–1363. [Google Scholar] [CrossRef]
- Santi, C.M.; Ferreira, G.; Yang, B.; Gazula, V.-R.; Butler, A.; Wei, A.; Kaczmarek, L.K.; Salkoff, L. Opposite Regulation of Slick and Slack K Channels by Neuromodulators. 2006. [CrossRef]
- Li, P.; Halabi, C.M.; Stewart, R.; Butler, A.; Brown, B.; Xia, X.; Santi, C.; England, S.; Ferreira, J.; Mecham, R.P.; et al. Sodium-Activated Potassium Channels Moderate Excitability in Vascular Smooth Muscle. J Physiol 2019, 597, 5093–5108. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.J.; Butler, A.; Stewart, R.; Gonzalez-Cota, A.L.; Lybaert, P.; Amazu, C.; Reinl, E.L.; Wakle-Prabagaran, M.; Salkoff, L.; England, S.K.; et al. Oxytocin Can Regulate Myometrial Smooth Muscle Excitability by Inhibiting the Na+-Activated K+ Channel, Slo2.1. J Physiol 2019, 597, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, A.; Gan, L.; Kaczmarek, L.K. Localization of the Slack Potassium Channel in the Rat Central Nervous System. Journal of Comparative Neurology 2002, 454, 241–254. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.; Xu, R.; Heinemann, S.H.; Hoshi, T. Reciprocal Regulation of the Ca2+ and H+ Sensitivity in the SLO1 BK Channel Conferred by the RCK1 Domain. Nat Struct Mol Biol 2008, 15, 403–410. [Google Scholar] [CrossRef]
- Brenker, C.; Zhou, Y.; Muller, A.; Echeverry, F.A.; Trotschel, C.; Poetsch, A.; Xia, X.M.; Bonigk, W.; Lingle, C.J.; Kaupp, U.B.; et al. The Ca2+-Activated K+ Current of Human Sperm Is Mediated by Slo3. Elife 2014, 2014, 1–19. [Google Scholar] [CrossRef]
- Zhang, X.; Zeng, X.; Lingle, C.J. Slo3 K+ Channels: Voltage and PH Dependence of Macroscopic Currents. Journal of General Physiology 2006, 128, 317–336. [Google Scholar] [CrossRef]
- Leonetti, M.D.; Yuan, P.; Hsiung, Y.; MacKinnon, R. Functional and Structural Analysis of the Human SLO3 PH- and Voltage-Gated K+ Channel. Proc Natl Acad Sci U S A 2012, 109, 19274–19279. [Google Scholar] [CrossRef]
- Hite, R.K.; Tao, X.; MacKinnon, R. Structural Basis for Gating the High-Conductance Ca2+ -Activated K+ Channel. Nature 2017, 541, 52–57. [Google Scholar] [CrossRef]
- Tao, X.; Hite, R.K.; MacKinnon, R. Cryo-EM Structure of the Open High-Conductance Ca2+ -Activated K+ Channel. Nature 2017, 541, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Tao, X.; Mackinnon, R. Molecular Structures of the Human Slo1 K+ Channel in Complex with B4. Elife 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Niu, X.; Wu, R.S.; Chudasama, N.; Yao, Y.; Jin, X.; Weinberg, R.; Zakharov, S.I.; Motoike, H.; Marx, S.O.; et al. Location of Modulatory β Subunits in BK Potassium Channels. Journal of General Physiology 2010, 135, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Li, Q.; Yan, J. The Leucine-Rich Repeat Domains of BK Channel Auxiliary γ Subunits Regulate Their Expression, Trafficking, and Channel-Modulation Functions. Journal of Biological Chemistry 2022, 298, 101664. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Perez, V.; Xia, X.-M.; Lingle, C.J. Functional Regulation of BK Potassium Channels by Γ1 Auxiliary Subunits. Proc Natl Acad Sci U S A 2014, 111, 4868–4873. [Google Scholar] [CrossRef] [PubMed]
- Behrens, R.; Nolting, A.; Reimann, F.; Schwarz, M.; Waldscḧtz, R.; Pongs, O. HKCNMB3 and HKCNMB4, Cloning and Characterization of Two Members of the Large-Conductance Calcium-Activated Potassium Channel β Subunit Family. FEBS Lett 2000, 474, 99–106. [Google Scholar] [CrossRef]
- Brenner, R.; Jegla, T.J.; Wickenden, A.; Liu, Y.; Aldrich, R.W. Cloning and Functional Characterization of Novel Large Conductance Calcium-Activated Potassium Channel β Subunits, HKCNMB3 and HKCNMB4. Journal of Biological Chemistry 2000, 275, 6453–6461. [Google Scholar] [CrossRef]
- Uebele, V.N.; Lagrutta, A.; Wade, T.; Figueroa, D.J.; Liu, Y.; McKenna, E.; Austin, C.P.; Bennett, P.B.; Swanson, R. Cloning and Functional Expression of Two Families of β-Subunits of the Large Conductance Calcium-Activated K+ Channel. Journal of Biological Chemistry 2000, 275, 23211–23218. [Google Scholar] [CrossRef]
- Yan, J.; Aldrich, R.W. LRRC26 Auxiliary Protein Allows BK Channel Activation at Resting Voltage without Calcium. Nature 2010 466:7305 2010, 466, 513–516. [Google Scholar] [CrossRef]
- Yan, J.; Aldrich, R.W. BK Potassium Channel Modulation by Leucine-Rich Repeat-Containing Proteins. Proc Natl Acad Sci U S A 2012, 109, 7917–7922. [Google Scholar] [CrossRef] [PubMed]
- Dolan, J.; Walshe, K.; Alsbury, S.; Hokamp, K.; O’Keeffe, S.; Okafuji, T.; Miller, S.F.C.; Tear, G.; Mitchell, K.J. The Extracellular Leucine-Rich Repeat Superfamily; a Comparative Survey and Analysis of Evolutionary Relationships and Expression Patterns. BMC Genomics 2007, 8, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Zeng, X.-H.; Zhou, Y.; Xia, X.-M.; Lingle, C.J. LRRC52 (Leucine-Rich-Repeat-Containing Protein 52), a Testis-Specific Auxiliary Subunit of the Alkalization-Activated Slo3 Channel. Proc Natl Acad Sci U S A 2011, 108, 19419–19424. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.M.; Zhong, Z.G.; Du, X.R.; Zhang, F.F.; Guo, Q.; Liu, Y.; Tang, Q.Y.; Zhang, Z. Cloning and Characterization of the Rat Slo3 (KCa5.1) Channel: From Biophysics to Pharmacology; 2020; Vol. 177; ISBN 0000000280.
- Zeng, X.-H.; Yang, C.; Xia, X.-M.; Liu, M.; Lingle, C.J. SLO3 Auxiliary Subunit LRRC52 Controls Gating of Sperm KSPER Currents and Is Critical for Normal Fertility. Proc Natl Acad Sci U S A 2015, 112, 2599–2604. [Google Scholar] [CrossRef]
- Sánchez-Carranza, O.; Torres-Rodríguez, P.; Darszon, A.; Treviño, C.L.; López-González, I. Pharmacology of HSlo3 Channels and Their Contribution in the Capacitation-Associated Hyperpolarization of Human Sperm. Biochem Biophys Res Commun 2015, 466, 554–559. [Google Scholar] [CrossRef]
- Jiang, Z.; Wallner, M.; Meera, P.; Toro, L. Human and Rodent MaxiK Channel β-Subunit Genes: Cloning and Characterization. Genomics 1999, 55, 57–67. [Google Scholar] [CrossRef]
- Yang, C.-T.; Zeng, X.-H.; Xia, X.-M.; Lingle, C.J. Interactions between β Subunits of the KCNMB Family and Slo3: Β4 Selectively Modulates Slo3 Expression and Function. PLoS One 2009, 4, e6135. [Google Scholar] [CrossRef]
- Qian, X.; Nimigean, C.M.; Niu, X.; Moss, B.L.; Magleby, K.L. Slo1 Tail Domains, but Not the Ca2+ Bowl, Are Required for the Β1 Subunit to Increase the Apparent Ca2+ Sensitivity of BK Channels. Journal of General Physiology 2002, 120, 829–843. [Google Scholar] [CrossRef] [PubMed]
- Mannowetz, N.; Naidoo, N.M.; Choo, S.A.S.; Smith, J.F.; Lishko, P. V. Slo1 Is the Principal Potassium Channel of Human Spermatozoa. Elife 2013, 2013, e01009. [Google Scholar] [CrossRef] [PubMed]
- Santi, C.M.; Butler, A.; Kuhn, J.; Wei, A.; Salkoff, L. Bovine and Mouse SLO3 K + Channels. Journal of Biological Chemistry 2009, 284, 21589–21598. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Young, S.; Krenz, H.; Tüttelmann, F.; Röpke, A.; Krallmann, C.; Kliesch, S.; Zeng, X.H.; Brenker, C.; Strünker, T. The Ca2+ Channel CatSper Is Not Activated by CAMP/PKA Signaling but Directly Affected by Chemicals Used to Probe the Action of CAMP and PKA. J Biol Chem 2020, 295, 13181. [Google Scholar] [CrossRef] [PubMed]
- Lyon, M.; Li, P.; Ferreira, J.J.; Lazarenko, R.M.; Kharade, S. V.; Kramer, M.; McClenahan, S.J.; Days, E.; Bauer, J.A.; Spitznagel, B.D.; et al. A Selective Inhibitor of the Sperm-Specific Potassium Channel SLO3 Impairs Human Sperm Function. Proceedings of the National Academy of Sciences 2023, 120, e2212338120. [Google Scholar] [CrossRef]
- Ren, D.; Navarro, B.; Perez, G.; Jackson, A.C.; Hsu, S.; Shi, Q.; Tilly, J.L.; Clapham, D.E. A Sperm Ion Channel Required for Sperm Motility and Male Fertility. Nature 2001, 413, 603–609. [Google Scholar] [CrossRef]
- Kirichok, Y.; Navarro, B.; Clapham, D.E. Whole-Cell Patch-Clamp Measurements of Spermatozoa Reveal an Alkaline-Activated Ca2+ Channel. Nature 2006, 439, 737–740. [Google Scholar] [CrossRef]
- Carlson, A.E.; Westenbroek, R.E.; Quill, T.; Ren, D.; Clapham, D.E.; Hille, B.; Garbers, D.L.; Babcock, D.F. CatSper1 Required for Evoked Ca2+ Entry and Control of Flagellar Function in Sperm. Proc Natl Acad Sci U S A 2003, 100, 14864–14868. [Google Scholar] [CrossRef]
- Lv, M.; Liu, C.; Ma, C.; Yu, H.; Shao, Z.; Gao, Y.; Liu, Y.; Wu, H.; Tang, D.; Tan, Q.; et al. Homozygous Mutation in SLO3 Leads to Severe Asthenoteratozoospermia Due to Acrosome Hypoplasia and Mitochondrial Sheath Malformations. Reproductive Biology and Endocrinology 2022, 20, 1–15. [Google Scholar] [CrossRef]
- Liu, R.; Yan, Z.; Fan, Y.; Qu, R.; Chen, B.; Li, B.; Wu, L.; Wu, H.; Mu, J.; Zhao, L.; et al. Bi-Allelic Variants in KCNU1 Cause Impaired Acrosome Reactions and Male Infertility. Hum Reprod 2022, 37. [Google Scholar] [CrossRef] [PubMed]
- Chávez, J.C.; Darszon, A.; Treviño, C.L.; Nishigaki, T. Quantitative Intracellular PH Determinations in Single Live Mammalian Spermatozoa Using the Ratiometric Dye SNARF-5F. Front Cell Dev Biol 2020, 7. [Google Scholar] [CrossRef]
- Kaupp, U.B.; Strünker, T. Signaling in Sperm: More Different than Similar. Trends Cell Biol 2017, 27, 101–109. [Google Scholar] [CrossRef]
- Miller, M.R.; Mansell, S.A.; Meyers, S.A.; Lishko, P. V. Flagellar Ion Channels of Sperm: Similarities and Differences between Species. Cell Calcium 2015, 58, 105–113. [Google Scholar] [CrossRef]
- Balestrini, P.A.; Sanchez-Cardenas, C.; Luque, G.M.; Baro Graf, C.; Sierra, J.M.; Hernández-Cruz, A.; Visconti, P.E.; Krapf, D.; Darszon, A.; Buffone, M.G. Membrane Hyperpolarization Abolishes Calcium Oscillations That Prevent Induced Acrosomal Exocytosis in Human Sperm. FASEB Journal 2021, 35, e21478. [Google Scholar] [CrossRef] [PubMed]
- Torrezan-Nitao, E.; Brown, S.G.; Mata-Martínez, E.; Trevi~ No, C.L.; Barratt, C.; Publicover, S. [Ca2+]i Oscillations in Human Sperm Are Triggered in the Flagellum by Membrane Potential Sensitive Activity of CatSper. Human Reproduction 2020. [Google Scholar] [CrossRef] [PubMed]
- Lishko, P. V.; Botchkina, I.L.; Fedorenko, A.; Kirichok, Y. Acid Extrusion from Human Spermatozoa Is Mediated by Flagellar Voltage-Gated Proton Channel. Cell 2010, 140, 327–337. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, Y.; Wu, H.; Zhang, H.; Zhang, H.; Mao, J.; Liu, D.; Zhao, L.; Lin, H.C.; Tang, W.; et al. Sodium-Hydrogen-Exchanger Expression in Human Sperm and Its Relationship with Semen Parameters. J Assist Reprod Genet 2017, 34, 795–801. [Google Scholar] [CrossRef]
- Lishko, P. V.; Kirichok, Y. The Role of Hv1 and CatSper Channels in Sperm Activation. Journal of Physiology 2010, 588, 4667–4672. [Google Scholar] [CrossRef]
- Ramsey, I.S.; Ruchti, E.; Kaczmarek, J.S.; Clapham, D.E. Hv1 Proton Channels Are Required for High-Level NADPH Oxidase-Dependent Superoxide Production during the Phagocyte Respiratory Burst. Proc Natl Acad Sci U S A 2009, 106, 7642–7647. [Google Scholar] [CrossRef]
- Martínez-López, P.; Santi, C.M.; Treviño, C.L.; Ocampo-Gutiérrez, A.Y.; Acevedo, J.J.; Alisio, A.; Salkoff, L.B.; Darszon, A. Mouse Sperm K+ Currents Stimulated by PH and CAMP Possibly Coded by Slo3 Channels. Biochem Biophys Res Commun 2009, 381, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Wrighton, D.C.; Muench, S.P.; Lippiat, J.D. Mechanism of Inhibition of Mouse Slo3 (KCa 5.1) Potassium Channels by Quinine, Quinidine and Barium. Br J Pharmacol 2015, 172, 4355–4363. [Google Scholar] [CrossRef] [PubMed]
- Wijerathne, T.D.; Kim, J.; Yang, D.; Lee, K.P. Intracellular Calcium-Dependent Regulation of the Sperm-Specific Calcium-Activated Potassium Channel, HSlo3, by the BKCa Activator LDD175. Korean J Physiol Pharmacol 2017, 21, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.Y.; Zhang, Z.; Xia, O.M.; Lingle, C.J. Block of Mouse Slo1 and Slo3 K+ Channels by CTX, IbTX, TEA, 4-AP and Quinidine. Channels 2010, 4, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Mansell, S.A.; Publicover, S.J.; Barratt, C.L.R.; Wilson, S.M. Patch Clamp Studies of Human Sperm under Physiological Ionic Conditions Reveal Three Functionally and Pharmacologically Distinct Cation Channels. Mol Hum Reprod 2014, 20, 392–408. [Google Scholar] [CrossRef]
- Brenker, C.; Goodwin, N.; Weyand, I.; Kashikar, N.D.; Naruse, M.; Krähling, M.; Müller, A.; Kaupp, U.B.; Strünker, T. The CatSper Channel: A Polymodal Chemosensor in Human Sperm. EMBO J 2012, 31, 1654–1665. [Google Scholar] [CrossRef]
- Lishko, P. V.; Botchkina, I.L.; Kirichok, Y. Progesterone Activates the Principal Ca2+ Channel of Human Sperm. Nature 2011, 471, 387–391. [Google Scholar] [CrossRef]
- Cavarocchi, E.; Whitfield, M.; Saez, F.; Touré, A. Sperm Ion Transporters and Channels in Human Asthenozoospermia: Genetic Etiology, Lessons from Animal Models, and Clinical Perspectives. International Journal of Molecular Sciences 2022, 23, 3926. [Google Scholar] [CrossRef]
Figure 1.
Amino acid sequence homology of mouse, human, and bovine SLO3. Conserved regions are highlighted in blue. Sequence alignment performed using Jalview Version 2 [39,40,41].

Figure 2.
Cartoon of SLO3 potassium channels. SLO3 α-subunits are comprised of seven transmembrane domains with an extracellular N-terminus and a cytoplasmic C-terminus that contains two RCK domains. Four α-subunits come together to form a homotetramer capable of transporting potassium ions through the membrane.
Figure 2.
Cartoon of SLO3 potassium channels. SLO3 α-subunits are comprised of seven transmembrane domains with an extracellular N-terminus and a cytoplasmic C-terminus that contains two RCK domains. Four α-subunits come together to form a homotetramer capable of transporting potassium ions through the membrane.
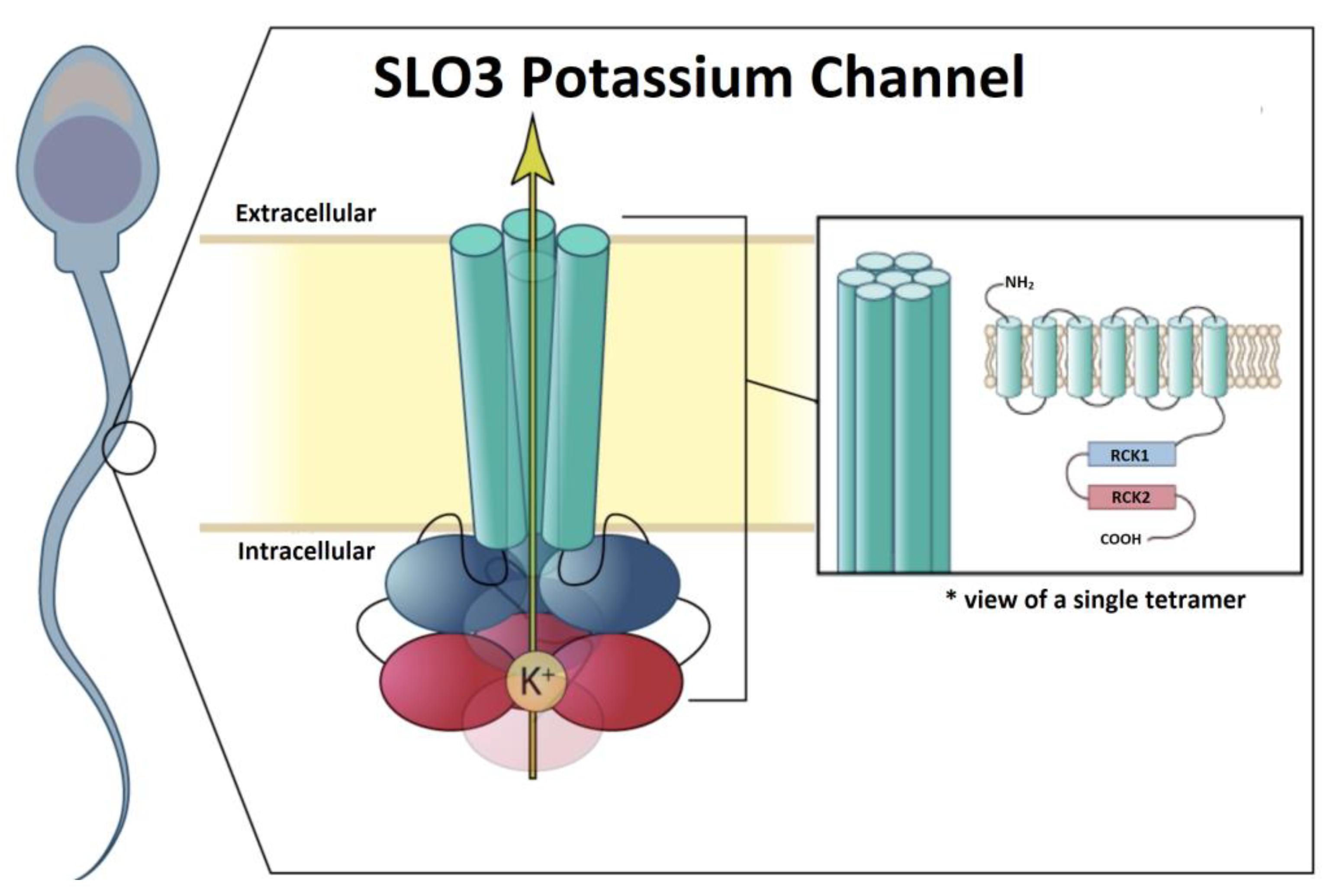
Figure 3.
Models of mouse and human SLO3 activity. (a) Mouse: The exposure to a more alkaline pH and high [HCO3-] concentrations in the female tract contribute to an increase in pHi, potentially through the activation of the sNHE. This rise in pHi leads to the activation of SLO3 channels, resulting in membrane hyperpolarization. This hyperpolarization enhances calcium influx through CatSper channels, possibly through two distinct mechanisms. Firstly, by increasing the inward driving force of calcium. Secondly, it may further activate sNHE to elevate intracellular pH even more. (b) Human: In human sperm, exposure to an elevated external pH could potentially activate the Hv1 channel, resulting in an increase in pHi and contributing to the activation of SLO3 and CatSper channels. However, it is important to note that in humans, SLO3 channels are primarily activated by calcium, while CatSper channels are activated by progesterone. On the other hand, activation of SLO3 leads to membrane hyperpolarization, which has been proposed to remove [Ca2+]i oscillations that inhibit CatSper activation. This raises the question of whether SLO3 is activated upstream or downstream of CatSper channels.
Figure 3.
Models of mouse and human SLO3 activity. (a) Mouse: The exposure to a more alkaline pH and high [HCO3-] concentrations in the female tract contribute to an increase in pHi, potentially through the activation of the sNHE. This rise in pHi leads to the activation of SLO3 channels, resulting in membrane hyperpolarization. This hyperpolarization enhances calcium influx through CatSper channels, possibly through two distinct mechanisms. Firstly, by increasing the inward driving force of calcium. Secondly, it may further activate sNHE to elevate intracellular pH even more. (b) Human: In human sperm, exposure to an elevated external pH could potentially activate the Hv1 channel, resulting in an increase in pHi and contributing to the activation of SLO3 and CatSper channels. However, it is important to note that in humans, SLO3 channels are primarily activated by calcium, while CatSper channels are activated by progesterone. On the other hand, activation of SLO3 leads to membrane hyperpolarization, which has been proposed to remove [Ca2+]i oscillations that inhibit CatSper activation. This raises the question of whether SLO3 is activated upstream or downstream of CatSper channels.
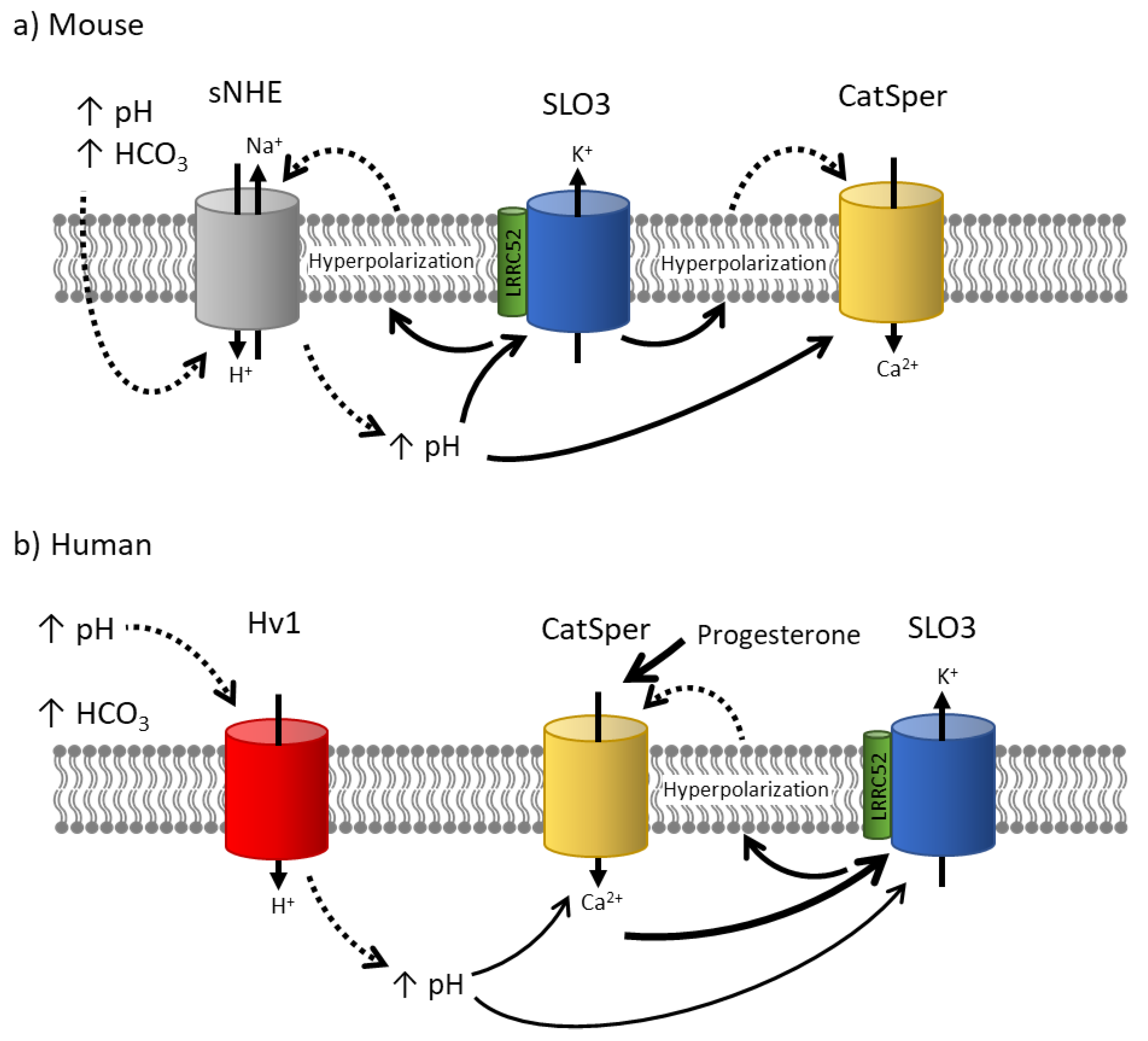

Table 1.
Effects of compounds on mouse or human sperm K+ currents. Whole-cell patch clamp studies were performed on of mouse (IKSper) or human (hKSper) sperm and reported the effect of compounds on potassium currents. Effects were categorized as inhibiting (↓) or having no effect (–).
Table 1.
Effects of compounds on mouse or human sperm K+ currents. Whole-cell patch clamp studies were performed on of mouse (IKSper) or human (hKSper) sperm and reported the effect of compounds on potassium currents. Effects were categorized as inhibiting (↓) or having no effect (–).
| Compound | Assay | Concentration | Effect | Study |
|---|---|---|---|---|
| 4-AP | hKSper | 2 mM | – | [109] |
| Bupivacaine | hKSper | 3 mM | ↓ | [109] |
| Charybdotoxin | hKSper | 1 µM | ↓ | [87] |
| Clofilium | hKSper | 50 µM - 5 mM | ↓ | [65,109] |
| Iberiotoxin | hKSper | 100 nM | – | [65] |
| hKSper | 100 nM | ↓ | [87] | |
| Lidocaine | hKSper | 3 mM | ↓ | [109] |
| Paxilline | hKSper | 100 nM | ↓ | [87] |
| Progesterone | hKSper | 0.5 - 30 µM | ↓ | [87,109] |
| Quinidine | hKSper | 300 - 500 µM | ↓ | [65,109] |
| TEA | hKSper | 10 mM | – | [65] |
| VU0546110 | hKSper | 10 µM | ↓ | [90] |
Table 2.
Effects of compounds on mouse, human, and rat SLO3 currents expressed in heterologous systems. Studies were examined that measured the effects on whole-cell SLO3 currents in heterologous expression systems of mouse (mSLO3), human (hSLO3), or rat (rSLO3) channels. These data are only for compounds applied externally. Effects were categorized as activating (↑), inhibiting (↓) or having no effect (–).
Table 2.
Effects of compounds on mouse, human, and rat SLO3 currents expressed in heterologous systems. Studies were examined that measured the effects on whole-cell SLO3 currents in heterologous expression systems of mouse (mSLO3), human (hSLO3), or rat (rSLO3) channels. These data are only for compounds applied externally. Effects were categorized as activating (↑), inhibiting (↓) or having no effect (–).
| Compound | Assay | Concentration | Effect | Study |
|---|---|---|---|---|
| 4-AP | hSLO3 | 25 mM | – | [107] |
| rSLO3 | 100 mM | – | [81] | |
| Ba2+ | hSLO3 | 1 mM | ↓ | [83] |
| mSLO3 | 2 mM | ↓ | [105,106] | |
| Charybdotoxin | hSLO3 | 100 nM | ↓ | [83] |
| Clofilium | hSLO3 | 50 µM | – | [83] |
| mSLO3 | 50 µM | ↓ | [106] | |
| Iberiotoxin | hSLO3 | 100 nM | – | [83,107] |
| hSLO3 | 0.1 - 300 nM | ↓ | [90] | |
| Ketamine | rSLO3 | 25 - 500 µM | ↓ | [81] |
| LDD175 | hSLO3 | 30 µM | ↑ | [107] |
| NS1619 | hSLO3 | 50 µM | ↓ | [107] |
| Paxilline | hSLO3 | 1 - 30 µM | ↓ | [90] |
| Penitrem A | hSLO3 | 100 nM | ↓ | [83] |
| Progesterone | hSLO3 | 30 µM | ↓ | [65,83] |
| Propofol | rSLO3 | 100 - 700 µM | ↓ | [81] |
| Quinidine | hSLO3 | 0.1 - 100 µM | ↓ | [83,90] |
| rSLO3 | 10-500 µM | ↓ | [81] | |
| mSLO3 | 500 µM | ↓ | [106] | |
| Slotoxin | hSLO3 | 100 nM | – | [83] |
| TEA | hSLO3 | 20 mM | ↓ | [107] |
| mSLO3 | 60 mM | ↓ | [105] | |
| mSLO3 | 20 mM | – | [106] | |
| VU0546110 | hSLO3 | 0.3-30 µM | ↓ | [90] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



