
Submitted:
07 June 2023
Posted:
07 June 2023
You are already at the latest version
Abstract
Poro-poro is an important native fruit used in traditional Peruvian medicine with relevant agro-industrial and pharmaceutical potential for its antioxidant capacity for human health. However, to date, there have been few genetic studies. The lack of genomic exploration limits the possibility of expanding our knowledge of its molecular evolution, new molecular pathways, genetic traits, and evolutionary relationships. Here, we report the plastid genome sequence of Passiflora tripartita var. mollissima and the reconstructed phylogenetic tree to infer the phylogenetic relationships among Passiflora species. Our phylogenetic analysis showed that poro-poro is most closely related to Passiflora menispermifolia and Passiflora oerstedii. In summary, our study provides the basis for developing new molecular markers that constitutes a valuable resource for studying molecular evolution and domestication. It also provides a powerful foundation for conservation genetics research and plant breeding programs. To our knowledge, this is the first report on the plastid genome of Passiflora tripartita var. mollissima from Peru.
Keywords:
Chloroplast genome
; Passifloraceae
; Passiflora tripartita
; poro-poro
; Huánuco
; Peru
Introduction
Passiflora tripartita var. mollissima (Kunth) Holms-Niels. & P.M. Jørg [1] previously known as Passiflora mollissima (Kunth) Bailey [2], is a semi-perennial fruit plant [3]. It is a diploid species with a small number of chromosomes (2n = 18) [4], which is placed in the section Elkea of supersection Tacsonia of subgenus Passiflora belonging to the Passifloraceae family [5,6]. Poro-poro is a native fruit of the Andean region [6]. It grows in the Peruvian highlands in the departments of Ancash, Junín, Moquegua, Huancavelica, and Huánuco at altitudes of 1,000–4,000 m.a.s.l. [7,8]. It is widely used in traditional medicine [8] and is considered one of the best Passiflora species based on its organoleptic characteristics [2]. This fruit provides a source of vitamins (A, B3, and C) and minerals (magnesium, potassium, phosphorus, sodium, chlorine, iron, calcium, sulfur, zinc, copper, selenium, cobalt, and nickel) [9,10]. In addition, it has an elevated antioxidant activity and high content of carotenoids (118.8 mg β-carotene), phenols (460.1 mg gallic acid), and flavonoids (1907.6 mg catechin/100 g) [9,10]. Specifically, the high concentration of flavan-3-ols (a group of bioactive compounds) has been associated with beneficial effects on human health, such as cardiovascular protection, neurodegenerative diseases, and as an anti-cancer, anti-microbial, and anti-parasitic agent [11,12].
Plastome sequences of more than 800 sequenced genomes are small in size with high copy numbers and conserved sequences, enabling a significant understanding of plant molecular evolution, structural variations, and evolutionary relationships of plant diversity [13,14]. The plastid genome has a quadripartite structure: a large single-copy (LSC) of 80–90 kilobase pairs (kb), a small single-copy (SSC) of 16–27 kb, and two sets of inverted repeats (IRa and IRb) of 20–28 kb, with 110-130 unique genes, including protein-coding genes, transfer RNA (tRNA), and ribosomal RNA (rRNA) [15,16]. In recent years, declining genome sequencing costs resulted in more than 780 complete plant genomes of different species becoming available [17,18]. Recently, some Passiflora plastid genomes such as Passiflora edulis [19], Passiflora xishuangbannaensis [20], Passiflora caerulea [21], Passiflora serrulata [22], Passiflora foetida [23], and Passiflora arbelaezii [24], became publicly available. However, despite the scarcity of genomic information on underutilized crops [25], we have only begun to investigate the genomics of plants of great importance for plant breeding programs. The aim of the present study was to sequence, assemble, and annotate the plastid genome of poro-poro to contribute to plant breeding programs. In the present study, we report the first plastid genome sequence submitted for an isolate of Passiflora tripartita var. mollissima from Peru, a species with great agro-industrial and pharmaceutical potential because of its beneficial characteristics for human health.
Results and Discussion
Plastome of Passiflora tripartiva var. mollisima
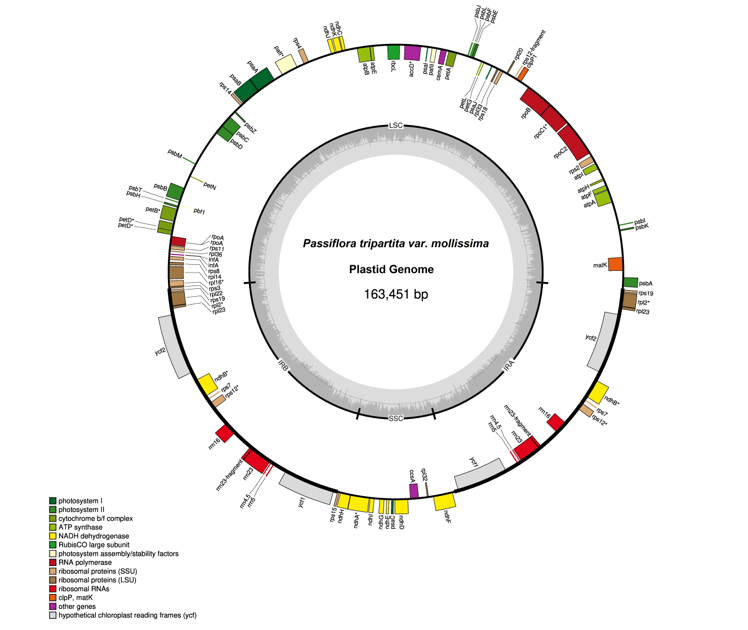
The plastid genome sequences of P. tripartita var. mollissima (poro-poro) (Figure 1) was 163,451 bp in length, with a typical quadripartite structure consisting of a large single-copy (LSC) region of 85,525 bp (52.32% in total) and a small single-copy (SSC) region of 13,518 bp (8.27%), separated by a pair of inverted repeat regions (IRa and IRb) of 32,204 bp (19.70%). The poro-poro plastome is 12,045 bp longer than that of one of the most economically important species, passion fruit (P. edulis) [19], and is only 7,117 bp longer than that of the longest Passiflora plastome reported, i.e., P. arbelaezii [24]. The plastome sequence of poro-poro has a similar quadripartite architecture to other plants [26,27,28]. However, the LSC region is 4,150 bp longer than that of P. xishuangbannaensis but is 98bp, 195 bp, and 1,927 bp shorter than that of P. caerulea, P. edulis, and P. arbelaezii, respectivety. The SSC region is 121 bp, 140 bp, 359 bp, and 754 bp longer than that of P. caerulea, P. edulis, P. xishuangbannaensis, and P. arbelaezii, respectively. The IRs regions are 6,024 bp, 6,050 bp, and 11,600 longer than that of P. caerulea, P. edulis, and P. xishuangbannaensis, respectively; however, it is 2,972 bp shorter than that of P. arbelaezii [19,20,21,24]. The plastome structure of the P. tripartita var. mollissima consisted of A = 30.79%, T(U) = 32.34%, C = 18.67% and G = 18.20%. The overall AT content of the plastid genome was 63.13%, whereas the overall GC content was 36.87% as similar to that of other reported chloroplast genomes from the same family, such as 36.90% in P. arbelaezii [24], 37% in P. edulis and P. serrulata [19,22], 37.03% in P. caerulea [21], and 37.1% in P. xishuangbannaensis [20].
Poro-poro plastid genome annotation identified 128 genes, of which 112 were unique, and 17 were duplicated in the inverted repeat (IR) region. The plastome contained 84 protein-coding genes, 37 transfer RNA (tRNA)-coding genes, 7 ribosomal RNA (rRNA)-coding genes, and 14 genes with introns (12 genes with one intron and 2 genes with two introns), as shown in Table 1. The poro-poro plastid genome contained 112 unique genes, of which there were 29 tRNA genes, 4 rRNA genes, and 79 protein-coding genes. The latter comprised 20 ribosomal subunit genes (9 large subunits and 11 small subunit), 4 DNA-directed RNA polymerase genes, 46 genes were involved in photosynthesis (11 encoded subunits of the NADH oxidoreductase, 7 for photosystem I, 15 for photosystem II, 6 for the cytochrome b6/f complex, 6 for different subunits of ATP synthase, and 1 for the large chain of ribulose biphosphate carboxylase), 8 genes were involved in different functions, and one gene was of unknown function (Table 2).
In the plastid genome, 14 genes contained introns distributed as follows: the LSC, SSC, and IRs regions contained 8 genes (petD, rpl16, rpoC1, trnG-UCC, trnK-UUU, trnL-UAA, trnV-UAC, and ycf3), 1 gene (ndhA), and 5 genes (ndhB, rpl2, rps12, trnA-UGC, and trnI-GAU) respectively. Similarly, these genes included six protein-coding genes, each with a single intron (petD, ndhA, ndhB, rpoC1, rpl2, and rpl16); six tRNA genes, each with a single intron (trnA-UGC, trnG-UCC, trnI-GAU, trnK-UUU, trnL-UAA, and trnV-UAC); and two protein-coding genes with two introns (ycf3 and rps12). Except for 17 genes that were duplicated in the IR region (ndhB, rps19, rpl2, rpl23, rps12, ycf15, rrn5, rrn16, rrn23, trnA-UGC, trnI-CAU, trnI-GAU, trnL-CAA, trnM-CAU, trnN-GUU, trnR-ACG, and trnV-GAC) all genes contained a single copy, as shown in Table 2. The plastome of P. tripartita var. mollissima contained eight genes (ycf1, ycf2, ycf15, rps16, rpl20, rpl22, accD, infA) that were lost or non-functional genes in P. edulis; and compared to P. edulis, it has one absent gene (trnfM-CAU), as previously reported [19]. In this study, the ycf1 sequence encodes a protein essential for plant viability and a vital component of the translocon on the inner chloroplast membrane (TIC) complex [29], and ycf2 is a component of the ATPase motor protein associated with the TIC complex [30].
Phylogenetic Reconstruction
To identify the evolutionary position of Passiflora tripartita var. mollissima in the Passifloraceae family, phylogenetic relationships based on the OrthoFinder clustering method were used to avoids erroneous rearrangements in phylogenetic tree reconstruction and provides a more reliable evolutionary analysis [31,32]. The phylogenetic tree was constructed based on single-copy orthologous genes [33] and maximum likelihood analysis with the complete annotated protein sequences of 27 plastid genomes, of which 26 were from Passiflora species. One species, Vitis vinifera, was chosen as the outgroup,
Maximum likelihood (ML) bootstrap values ranged from 38%–92% for 7 of the 25 nodes. All nodes except the indicated ones (seven nodes) exhibited bootstrap support (BS) values of 100%. These Passiflora species were divided into four groups: subgenus Passiflora (P. nitida, P. quadrangularis, P. cincinnata, P. caerulea, P. edulis, P. laurifolia, P. vitifolia, P. serratifolia, P. serrulata, P. ligularis, P. serratodigitata, P. actinia, P. menispermifolia and P. oerstedii), subgenus Tetrapathea (P. tetrandra), subgenus Decaloba (P. microstipula, P. xishuangbannaensis, P. biflora, P. lutea, P. jatunsachensis, P. suberosa and P. tenuiloba), and subgenus Deidamoides (P. contracta and P. arbelaezii). The relationships between the four subgenera of Passiflora species (Passiflora, Tetrapathea, Decaloba, and Deidamoides) were congruent and strongly supported by the same patterns as previously reported [34,35]. These results resolved Passiflora tripartita var. mollissima was placed in the subgenus Passiflora, which was closely related to P. menispermifolia and P. oerstedii with 100% BS, and was sister to P. tetrandra (subgenus Tetrapathea), P. biflora (subgenus Decaloba), and P. contracta (subgenus Deidamoides), as shown in the cladogram (Figure 2).
Conclusions
This study provides information on the sequencing, assembly, and annotation of the plastid genome of Passiflora tripartita var. mollissima (poro-poro) from Huánuco, Peru. The plastome structure and gene content were relatively conserved, and the phylogenetic relationships illustrated that this species is placed in the subgenus Passiflora and positioned close to Passiflora menispermifolia and Passiflora oerstedii. This study will open up further avenues for research on plant molecular breeding, molecular markers, evolutionary studies, and conservation genetics of poro-poro.
Materials and Methods
Plant Materials
In November 2022, the fresh leaves of Passiflora tripartita var. mollissima (Supplementary Figure S1) were collected from Raccha Cedrón locality of Quisqui District, Huánuco Province from Peru (9°53’37”S, 76°26’02”W, altitude 2,945 m.a.s.l.). A herbarium voucher specimen (USM<PER>:MHN331530) was deposited in the Herbario San Marcos (USM) of the Museo de Historia Natural (MHN) at the Universidad Nacional Mayor de San Marcos (UNMSM).
DNA Extraction
Total genomic DNA was extracted from approximately 100 mg fresh leaves (from voucher number USM<PER>:MHN331530) using a cetyl-trimethyl ammonium bromide (CTAB) protocol [36]. Genomic DNA quality was assessed using a fluorometry-based Qubit (Thermo Fisher Scientific, USA) coupled to a Broad Range Assay kit. High-quality DNA (230/260 and 260/280 ratios >1.8) was normalized (20 ng/μL) to examine its integrity using 1% (w/v) agarose gel electrophoresis.
Genome Sequencing, Assembly, and Annotation
Qualified DNA was fragmented, and the TruSeq Nano DNA kit (Illumina, San Diego, CA, USA) was used to construct an Illumina paired-end (PE) library. PE sequencing (2 × 150 bp) was performed using the Illumina NovaSeq 6000 platform [37] (Macrogen, Inc., Seoul, Republic of Korea). All adapters and low-quality reads were removed using the FastQC [38] and Cutadapt [39] programs. PE reads (2 × 150 bp) were evaluated for quality using QUAST [40] analysis, and subsequent steps used clean data. Then, clean reads obtained were assembled into a circular contig using NOVOPlasty v.4.3 [41], with P. edulis (NC_034285) as the reference.
Phylogenetic Analysis
We used 26 complete plastome sequences to infer the phylogenetic relationships among Passiflora species, and Vitis vinifera was used as an outgroup (Supplemental Table 1). Single-copy orthologous genes were identified using the Orthofinder version 2.2.6 pipeline [33]. For each gene family, the nucleotide sequences were aligned using the L-INS-i algorithm in MAFFT v7.453 [45]. A phylogenetic tree based on maximum likelihood (ML) was constructed using RAxML v8.2.12 [46] with the GTRCAT model. A phylogenetic ML tree was reconstructed and edited using MEGA 11 [47] with 100 replicates.
Supplementary information
Figure S1: Herbarium specimen voucher of Passiflora tripartita var. mollissima (Kunth) Holms-Niels. & P.M. Jørg (USM<PER>:MHN331530). Table S1: Details of the plastid genome sequences used for phylogenetic analysis.
Author Contributions
Conceptualization, F.A. M.Z.-C. and S.A.V.-Z.; methodology, F.A., M.Z.-C. and S.A.V.-Z.; formal analysis, F.A.; data curation, F.A; writing—original draft preparation, F.A., M.Z.-C. and S.A.V.-Z.; writing—review and editing, F.A., M.Z.-C. and S.A.V.-Z.; visualization, M.Z.-C. and S.A.V.-Z.; supervision, F.A., M.Z.-C. and S.A.V.-Z.; project administration, M.Z.-C. and S.A.V.-Z.; funding acquisition, F.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Plant Science Laboratory E.I.R.L. and Universidad Privada del Norte (Sach’a Ruru grant: RIC-2022-101).
Data Availability Statement
The plastid genome sequence data have been deposited and openly available in GenBank of NCBI repository under the accession code: OQ910395.
Acknowledgments
We thank the Servicio Nacional Forestal y de Fauna Silvestre (SERFOR) for authorized this research project (AUT-IFL-2022-058). We thank Petr Sklenář (Charles University) and Filip Kolar (Charles University) for their help in the sample collection. We thank curator Julio C. Torres–Martinez (Museo de Historia Natural, Universidad Nacional Mayor de San Marcos) for the taxonomy identification and deposit of the plant specimen. We thank Dr. Rajest Mahato and Dr. Giusseppe D’Auria for the recommendations and bioinformatics support. We thank Mr. Julián Vasquez-Arriaga for administrative support (Plant Science Laboratory).
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- ITIS (2022) Passiflora tripartita var. mollisima (Kunth) Holms-Niels. & P.M. Jørg. Available online: https://www.itis.gov/ (accessed on 01 Jun 2023).
- Primot S, Coppens D’Eeckenbrugge G, Rioux V, Ocampo JA, Garcin F (2005) Variación morfológica de tres especies de curubas (Passiflora tripartita var. mollissima, P. tarminiana y P. mixta) y sus híbridos en el Valle del Cauca (Colombia). Rev Bras Frutic 27: 467–471, doi: 10.1590/S0100-29452005000300030. [CrossRef]
- Mayorga M, Fischer G, Melgarejo LM, Parra-Coronado A (2020) Growth, development and quality of Passiflora tripartita var. mollissima fruits under two environmental tropical conditions. J Appl Bot Food Qual 93: 66–75, doi: 10.5073/JABFQ.2020.093.009. [CrossRef]
- Coppens D’Eeckenbrugge G (2001) Conservación y utilización de recursos geneticos de pasifloras. COLCIENCIAS Informe final 1999-2001. Available online: https://agritrop.cirad.fr/576977 (accessed on 01 Jun 2023).
- egura SD, D’Eeckenbrugge GC, Ocampo CH, Ollitrault P (2005) Isozyme variation in Passiflora subgenus tacsonia: Geographic and interspecific differentiation among the three most common species. Genet Resour Crop Evol 52: 455–463, doi: 10.1007/s10722-005-2255-z. [CrossRef]
- Ocampo J, Coppens d’Eeckenbrugge GC (2017) Morphological characterization in the genus passiflora l.: an approach to understanding its complex variability. Plant Syst Evol 303: 531–558, doi: 10.1007/s00606-017-1390-2. [CrossRef]
- Tapia ME, Fries AM (2007) Guía de campo de los cultivos andinos, 1st ed.; FAO: Roma, Italy, pp. 22–114.
- Ríos-García G (2017) Nivel de aceptabilidad del vino de tumbo serrano (Passiflora mollissima) elaborado con los parámetros tecnológicos óptimos en la ciudad de Huánuco 2015. Master thesis, Universidad Nacional Hermilio Valdizán de Huánuco: Huánuco.
- Leterme P, Buldgen A, Estrada F, Londoño AM (2006) Mineral content of tropical fruits and unconventional foods of the Andes and the rain forest of Colombia. Food Chem 95: 644-652.
- Chaparro-Rojas DC, Maldonado ME, Franco-Londoño MC, Urango-Marchena LA (2014) Características nutricionales y antioxidantes de la fruta curuba larga (Passifora mollissima Bailey). Perspect Nutr Hum 16: 203-212, doi: 10.17533/udea.penh.v16n2a07. [CrossRef]
- Giambanelli E, Gómez-Caravaca AM, Ruiz-Torralba A, Guerra-Hernández EJ, Figueroa-Hurtado JG, García-Villanova B, Verardo V (2020) New advances in the determination of free and bound phenolic compounds of banana passion fruit pulp (Passiflora tripartita, var. mollissima (kunth) l.h. bailey) and their in vitro antioxidant and hypoglycemic capacities. Antioxidants 9: 628, doi: 10.3390/antiox9070628. [CrossRef]
- Luo Y, Jian Y, Liu Y, Jiang S, Muhammad D, Wang W (2022) Flavanols from nature: A phytochemistry and biological activity review. Molecules 27: 719, doi: 10.3390/molecules27030719. [CrossRef]
- Daniell H, Lin CS, Yu M, Chang WJ (2016) Chloroplast genomes: diversity, evolution, and applications in genetic engineering. Genome Biol 17: 134, doi: 10.1186/s13059-016-1004-2. [CrossRef]
- Dobrogojski J, Adamiec M, Lucinski R (2020) The chloroplast genome: A review. Acta Physiol Plant 42: 98, doi: 10.1007/s11738-020-03089-x. [CrossRef]
- 15. Ozeki H, Umesono K, Inokuchi H, Kohchi T, Ohyama K (1989) The chloroplast genome of plants: a unique origin. Genome. 31: 169–174, doi: 10.1139/g89-029. [CrossRef]
- Wang W, Lanfear R (2019) Long-reads reveal that the chloroplast genome exists in two distinct versions in most plants. Genome Biol Evol 11: 3372–3381, doi: 10.1093/gbe/evz256. [CrossRef]
- Marks RA, Hotaling S, Frandsen PB, VanBuren R (2021) Representation and participation across 20 years of plant genome sequencing. Nat Plants 7: 1571–1578, doi: 10.1038/s41477-021-01031-8. [CrossRef]
- Sun Y, Shang L, Zhu QH, Fan L, Guo L (2022) Twenty years of plant genome sequencing: achievements and challenges. Trends Plant Sci 27: 391–401, doi: 10.1016/j.tplants.2021.10.006. [CrossRef]
- Cauz-Santos LA, Munhoz CF, Rodde N, Cauet S, Santos AA, Penha HA, Dornelas MC, Varani AM, Oliveira GCX, Gergès H, Vieira MLC (2017) The Chloroplast Genome of Passiflora edulis (Passifloraceae) Assembled from Long Sequence Reads: Structural Organization and Phylogenomic Studies in Malpighiales. Front Plant Sci 8: 334, doi: 10.3389/fpls.2017.00334. [CrossRef]
- Hao C, Wu F (2021) The complete chloroplast genome sequence of Passiflora xishuangbannaensis (Passifloraceae), a vine endemic to Yunnan, China. Mitochondrial DNA B Resour 6: 1763–1764, doi: 10.1080/23802359.2021.1928562. [CrossRef]
- Niu YF, Ni SB, Liu SH, Liu J (2021) The complete chloroplast genome of Passiflora caerulea, a tropical fruit with a distinctive aroma. Mitochondrial DNA B Resour 6: 488–490, doi: 10.1080/23802359.2021.1872442. [CrossRef]
- Mou HF, Huang WH, Liu JY, Wen F, Tian QL, Wu YY, Fu LF, Zhang YJ, Wei YG (2021) Complete chloroplast genome sequence of Passiflora serrulata Jacq. (Passifloraceae). Mitochondrial DNA B Resour 6: 191–193, doi: 10.1080/23802359.2020.1860711. [CrossRef]
- Hopley T, Webber BL, Raghu S, Morin L, Byrne M (2021) Revealing the Introduction History and Phylogenetic Relationships of Passiflora foetida sensu lato in Australia. Front Plant Sci 12: 651805, doi: 10.3389/fpls.2021.651805. [CrossRef]
- Shrestha B, Weng ML, Theriot EC, Gilbert LE, Ruhlman TA, Krosnick SE, Jansen RK (2019) Highly accelerated rates of genomic rearrangements and nucleotide substitutions in plastid genomes of Passiflora subgenus Decaloba. Mol Phylogenet Evol 138: 53–64, doi: 10.1016/j.ympev.2019.05.030. [CrossRef]
- Gioppato HA, da Silva MB, Carrara S, Palermo BRZ, de Souza Moraes T, Dornelas MC (2019) Genomic and transcriptomic approaches to understand Passiflora physiology and to contribute to passionfruit breeding. Theor Exp Plant Physiol 31: 173–181, doi: 10.1007/s40626-018-0134-1. [CrossRef]
- Ohyama K, Fukuzawa H, Kohchi T, Shirai H, Sano T, Sano S, Umesono K, Shiki Y, Takeuchi M, Chang Z, Aota SI, Inokuchi H, Ozeki H (1986) Chloroplast gene organization deduced from complete sequence of liverwort Marchantia polymorpha chloroplast DNA. Nature 322: 572–574, doi: 10.1038/322572a0. [CrossRef]
- Shinozaki K, Ohme M, Tanaka M, Wakasugi T, Hayashida N, Matsubayashi T, Zaita N, Chunwongse J, Obokata J, Yamaguchi-Shinozaki K, Ohto C, Torazawa K, Meng BY, Sugita M, Deno H, Kamogashira T, Yamada K, Kusuda J, Takaiwa F, Kato A, Tohdoh N, Shimada H, Sugiura M (1986) The complete nucleotide sequence of the tobacco chloroplast genome: its gene organization and expression. EMBO J 5: 2043–2049, doi: 10.1002/j.1460-2075.1986.tb04464.x. [CrossRef]
- Nguyen HQ, Nguyen TNL, Doan TN, Nguyen TTN, Phạm MH, Le TL, Sy DT, Chu HH, Chu HM (2021) Complete chloroplast genome of novel Adrinandra megaphylla Hu species: molecular structure, comparative and phylogenetic analysis. Sci Rep 11: 11731, doi: 10.1038/s41598-021-91071-z. [CrossRef]
- 29. Kikuchi S, Bédard J, Hirano M, Hirabayashi Y, Oishi M, Imai M, Takase M, Ide T, Nakai M (2013) Uncovering the protein translocon at the chloroplast inner envelope membrane. Science 339: 571–574, doi: 10.1126/science.1229262. [CrossRef]
- Kikuchi S, Asakura Y, Imai M, Nakahira Y, Kotani Y, Hashiguchi Y, Nakai Y, Takafuji K, Bédard J, Hirabayashi-Ishioka Y, Mori H, Shiina T, Nakai MA (2018) Ycf2-FtsHi Heteromeric AAA-ATPase Complex Is Required for Chloroplast Protein Import. Plant Cell 30: 2677–2703, doi: 10.1105/tpc.18.00357. [CrossRef]
- Gabaldón T. Evolution of proteins and proteomes: a phylogenetics approach. Evol. Bioinform. Online. 2005, 1, 51–61, doi: 10.1177/117693430500100004. [CrossRef]
- Zhang N, Zeng L, Shan H, Ma H (2012) Highly conserved low-copy nuclear genes as effective markers for phylogenetic analyses in angiosperms. New Phytol 195: 923–937, doi: 10.1111/j.1469-8137.2012.04212.x. [CrossRef]
- Emms DM, Kelly S (2019) OrthoFinder: phylogenetic orthology inference for comparative genomics. Genome Biol 20: 238, doi:10.1186/s13059-019-1832-y. [CrossRef]
- Cauz-Santos LA, da Costa ZP, Callot C, Cauet S, Zucchi MI, Bergès H, van den Berg C, Vieira MLC (2020) A Repertory of Rearrangements and the Loss of an Inverted Repeat Region in Passiflora Chloroplast Genomes.Genome Biol Evol12: 1841–1857, doi: 10.1093/gbe/evaa155. [CrossRef]
- Pacheco TG, Lopes AS, Welter JF, Yotoko KSC, Otoni WC, Vieira LDN, Guerra MP, Nodari RO, Balsanelli E, Pedrosa FO, de Souza EM, Rogalski M (2020) Plastome sequences of the subgenus Passiflora reveal highly divergent genes and specific evolutionary features. Plant Mol Biol 104: 21–37, doi: 10.1007/s11103-020-01020-z. [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19,11–15.
- Modi A, Vai S, Caramelli D, Lari M (2021) The illumina sequencing protocol and the novaseq 6000 system. In: Bacterial Pangenomics: Methods and Protocols, 2nd edn; Mengoni, A.; Bacci, G.; Fondi, M., Eds.; Springer: New York, USA, Volume 2242, pp. 15–42, ISBN: 978-1-0716-1099-2.
- Wingett SW, Andrews S (2018) FastQ Screen: A tool for multi-genome mapping and quality control. F1000Res 7: 1338, doi: 10.12688/f1000research.15931.2. [CrossRef]
- Martin M (2011) Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J 17: 10-12, doi: 10.14806/ej.17.1.200. [CrossRef]
- Gurevich A, Saveliev V, Vyahhi N, Tesler G (2013) QUAST: quality assessment tool for genome assemblies. Bioinformatics 29: 1072–1075, doi: 10.1093/bioinformatics/btt086. [CrossRef]
- Dierckxsens N, Mardulyn P, Smits G (2017) NOVOPlasty: de novo assembly of organelle genomes from whole genome data. Nucleic Acids Res45: e18, doi: 10.1093/nar/gkw955. [CrossRef]
- Tillich M, Lehwark P, Pellizzer T, Ulbricht-Jones ES, Fischer A, Bock R, Greiner S (2017) GeSeq - versatile and accurate annotation of organelle genomes. Nucleic Acids Res 45: W6–W11, doi: 10.1093/nar/gkx391. [CrossRef]
- Shi L, Chen H, Jiang M, Wang L, Wu X, Huang L, Liu C (2019) CPGAVAS2, an integrated plastome sequence annotator and analyzer. Nucleic Acids Res 47: W65–W73, doi: 10.1093/nar/gkz345. [CrossRef]
- Greiner S, Lehwark P, Bock R (2019) OrganellarGenomeDRAW (OGDRAW) version 1.3.1: expanded toolkit for the graphical visualization of organellar genomes. Nucleic Acids Res 47: W59–W64, doi: 10.1093/nar/gkz238. [CrossRef]
- Katoh K, Standley DM (2013) MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol 30: 772–780, doi: 10.1093/molbev/mst010. [CrossRef]
- Stamatakis A (2014) RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30: 1312–1313, doi: 10.1093/bioinformatics/btu033. [CrossRef]
- Tamura K, Stecher G, Kumar S (2021) MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol Biol Evol 38: 3022-3027, doi: 10.1093/molbev/msab120. [CrossRef]
Figure 1.
Plastid genome of Passiflora tripartita var. mollissima. The thick lines indicate the IR1 and IR2 regions, which separate the SSC and LSC regions. Genes marked inside the circle are transcribed clockwise, and genes marked outside the circle are transcribed counterclockwise. Genes are color-coded based on their function, shown at the bottom left. The inner circle indicates the inverted boundaries and GC content.
Figure 1.
Plastid genome of Passiflora tripartita var. mollissima. The thick lines indicate the IR1 and IR2 regions, which separate the SSC and LSC regions. Genes marked inside the circle are transcribed clockwise, and genes marked outside the circle are transcribed counterclockwise. Genes are color-coded based on their function, shown at the bottom left. The inner circle indicates the inverted boundaries and GC content.
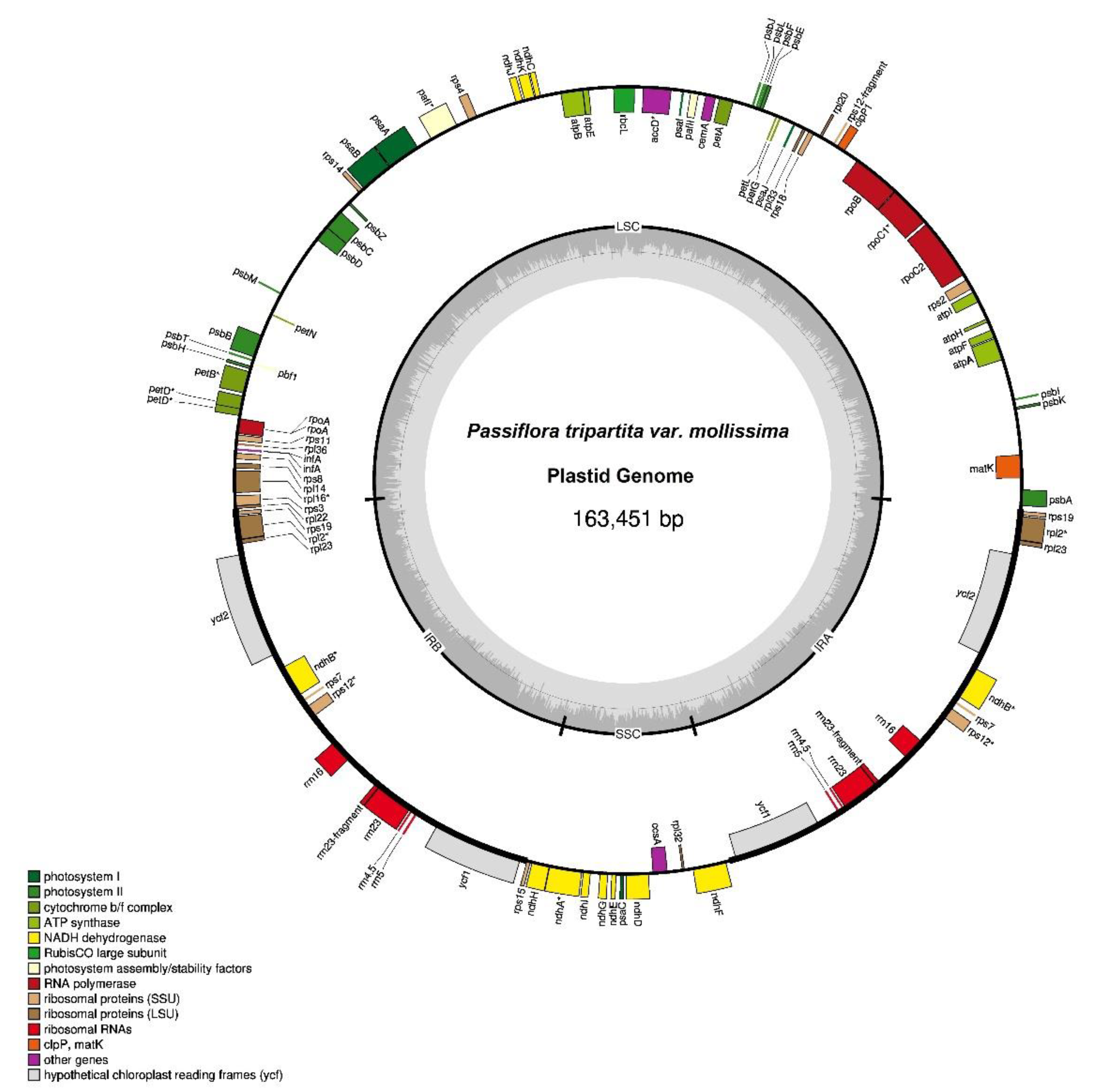
Figure 2.
Phylogenetic tree of 27 plastid genomes using maximum likelihood analysis based on single-copy orthologous protein. Bootstrap values on the branches were calculated from 100 replicates.
Figure 2.
Phylogenetic tree of 27 plastid genomes using maximum likelihood analysis based on single-copy orthologous protein. Bootstrap values on the branches were calculated from 100 replicates.
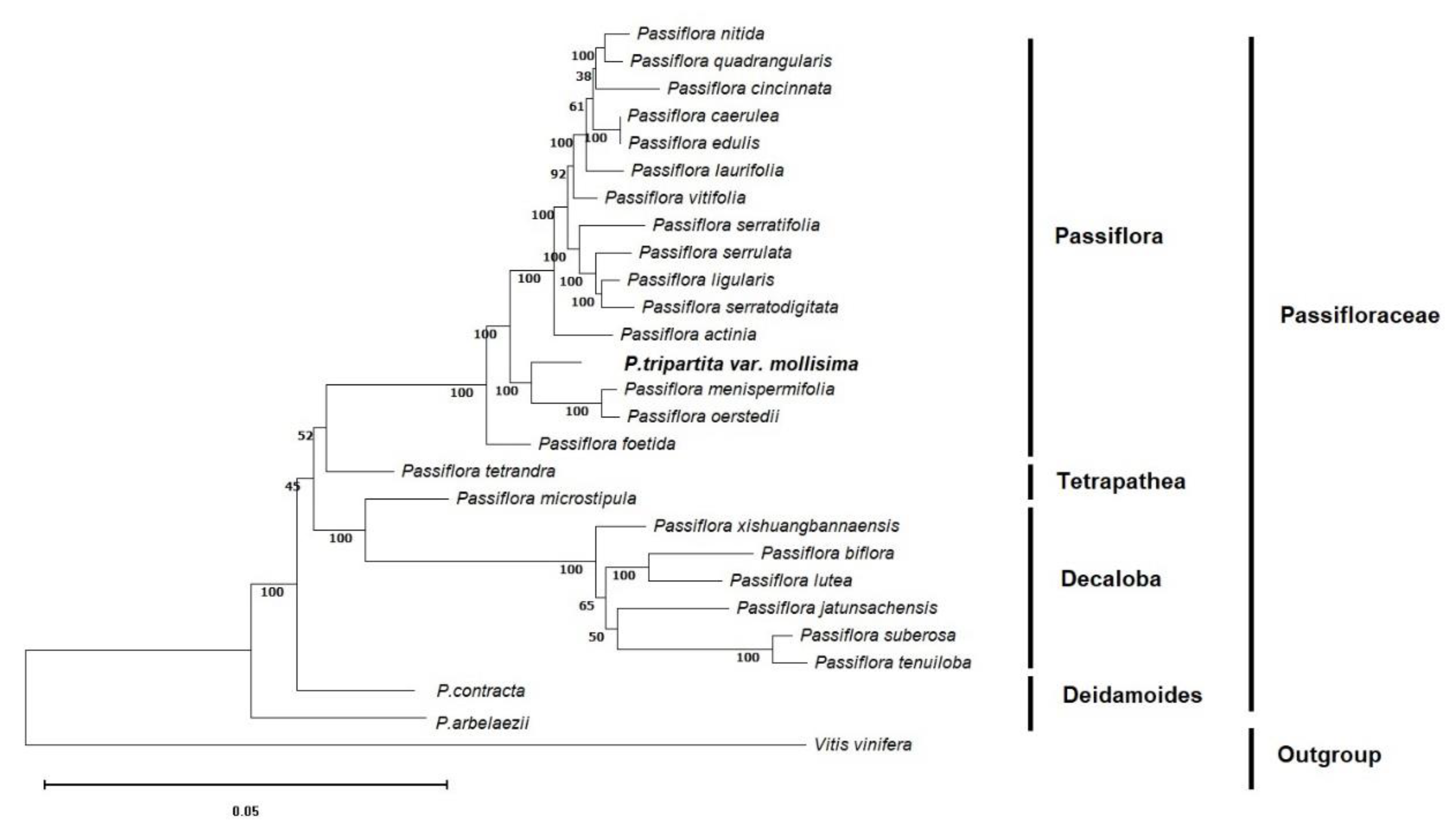

Table 1.
Plastid genome features of the P. tripartita var. mollissima.
| Features | Poro-poro 1 |
|---|---|
| Genome size (bp) | 163,451 |
| LSC length (bp) | 85, 525 |
| SSC length (bp) | 13,518 |
| IR length (bp) | 32,204 |
| Total GC content (%) | 36.87 |
| A content (%) | 30.79 |
| T(U) content (%) | 32.34 |
| G content (%) | 18.20 |
| C content (%) | 18.67 |
| Total number of genes | 128 |
| Protein-coding genes | 84 |
| rRNA coding genes | 7 |
| tRNA coding genes | 37 |
| Genes duplicated in IR regions | 17 |
| Total introns | 14 |
| Single introns (gene) | 12 |
| Double introns (gene) | 2 |
Table 2.
Genes present in the plastid genome of P. tripartita var. mollissima.
| Group of genes | Gene names |
|---|---|
| Photosystem I | psaA, psaB, psaC, psaI, psaJ, ycf3 **, ycf4 |
| Photosystem II | psbA, psbB, psbC, psbD, psbE, psbF, psbH, psbI, psbJ, psbK, psbL, psbM, psbN, psbT, psbZ |
| Cytochrome b/f complex | petA, petB, petD *, petG, petL, petN |
| ATP synthase | atpA, atpB, atpE, atpF, atpH, atpI |
| NADH dehydrogenase | ndhA*, ndhB * (X2), ndhC, ndhD, ndhE, ndhF, ndhG, ndhH, ndhI, ndhJ, ndhK |
| RubisCO large subunit | rbcL |
| DNA-dependent RNA polymerase | rpoA, rpoB, rpoC1 *, rpoC2 |
| Ribosomal proteins (SSU) | rps2, rps3, rps4, rps8, rps11, rps12 ** (X2), rps14, rps15, rps16, rps18, rps19 (X2) |
| Ribosomal proteins (LSU) | rpl2 * (X2), rpl14, rpl16 *, rpl20, rpl22, rpl23 (X2), rpl32, rpl33, rpl36 |
| Acetyl-CoA carboxylase | accD |
| C-type cytochrome synthesis | ccsA |
| Envelope membrane protein | cemA |
| Protease | clpP |
| Translational initiation factor IF-1 | infA |
| Maturase | matK |
| Component of TIC complex | yct1, ycf2 |
| Unknown function protein-coding | ycf15 (X2) |
| Ribosomal RNAs | rrn4.5, rrn5 (X2), rrn16 (X2), rrn23 (X2) |
| Transfer RNAs | trnA-UGC * (X2), trnC-GCA, trnD-GUC, trnE-UUC, trnF-GAA, trnG-GCC, trnG-UCC *, trnH-GUG, trnI-CAU (X2), trnI-GAU * (X2), trnK-UUU *, trnL-CAA (X2), trnL-UAA *, trnL-UAG, trnM-CAU (X2), trnN-GUU (X2), trnP-UGG, trnQ-UUG, trnR-ACG (X2), trnR-UCU, trnS-GCU, trnS-GGA, trnS-UGA, trnT-GGU, trnT-UGU, trnV-GAC (X2), trnV-UAC *, trnW-CCA, trnY-GUA |
* Gene contains one intron; ** gene contains two introns; (X2) indicates two gene copies in IRs.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



