
Submitted:
02 June 2023
Posted:
06 June 2023
You are already at the latest version
Abstract
Management of calcium and phosphate in biofluids is key to maintaining physiological mineral homeostasis (i.e., appropriate mineralization of hard tissues and an absence of mineral deposition in soft tissues). This review describes and contrasts the ways vertebrates manage calcium phosphate in two biological fluids (breast milk and serum) and illustrates the benefits of mineral sequestration by proteins. In milk, phosphoprotein-sequestered calcium magnesium phosphates provide nutritional support, whereas in serum, protein-sequestered calcium phosphates control transport and delivery of calcium and phosphate to tissues for biological function or excretion. In addition, subsets of sequestered phosphates in serum have been identified as culprits underlying ectopic deposition of calcium phosphates and toxicity.

Keywords:
Calcium phosphate
; calciprotein particles
; nanoclusters
; intrinsically disordered proteins
; crystal toxicity
; milk
; serum
Key Points
- Sequestration of nascent clusters of ternary phosphates composed of magnesium, calcium, and phosphate by intrinsically disordered proteins play key roles in physiological calcium phosphate management. Sequestration results in the formation of soluble colloids: large fibrous amorphous calcium magnesium phosphate nanoclusters in milk and small spherical amorphous calcium phosphate nanoclusters in serum.
- Breast milk is composed of large fibrous micelles in which casein embeds amorphous calcium magnesium phosphate nanoclusters. These casein micelles exhibit structural integrity, even in the presence of other proteins. The casein micelles remain remarkably stable during storage as liquids or solids, exposure to heat and cold, drying and reconstitution, and exposure to salts, alcohols, or acids. The phosphates embedded within the casein molecules remain amorphous, a feature which supports their nutritional value as sources of calcium, magnesium and phosphate.
- In healthy individuals, calciprotein nanoparticles, small nanoscale aggregates of amorphous calcium phosphates embedded in binding proteins, promote efficient transport, distribution, and clearance of calcium, magnesium, and phosphate without risk of precipitation. The half-life of these nanoparticles in serum is short, and if not utilized for calcium/phosphate replenishment within minutes of their formation, they are cleared by the kidney, liver and spleen. Some of the nanoparticles may be sufficiently small to pass through renal tissues into urine. As circulating biomaterials, they may couple dietary mineral exposure with endocrine control of mineral metabolism in bone, kidney and intestine. Conversely, when calcium and phosphate homeostasis is disrupted, these particles emerge as mediators of phosphate toxicity throughout the body. If the particles are not scavenged, they increase in size and accumulate a wide variety of organic species typically found in the biofluid environment. The embedded mineral gradually transforms into nanoparticles composed of sequestered mixtures of amorphous and crystalline calcium phosphates. As this ripening occurs, the particles become less soluble in the biofluid and the phosphates become less soluble in acidic media. Ectopic particle deposition on soft tissues is associated with cellular toxicity. Cytotoxicity may be related, at least in part, to the crystallopathy of crystalline calcium phosphates that induce inflammation, injury, and cell death.
1. Introduction
The bodies of vertebrates comprise hard tissues that are heavily mineralized with calcium phosphates beneficially co-existing with soft tissues and biofluids that contain little or none of this mineral. For the hard tissue to stay mineralized, the permeating biofluid must be saturated with the hard tissue mineral. At the same time, both the soft tissues and biofluids must remain free of mineral precipitates. Thus, the permeating biofluids must contain stable forms of the mineral substances in quasi-equilibrium with the mineralized forms found in hard tissues. If stabilization fails, soft tissues can become mineralized and hard tissues can become demineralized, conditions that characterize degenerative, dysfunctional, or diseased states.
A physiological solution to this conundrum likely evolved early and continues to allow physiological calcium phosphate management [1,2,3]. In order to control, regulate, and maintain calcium and phosphate in biological fluids and tissues, a group of secreted, calcium-binding phosphoproteins and a second group of polycarboxylated proteins evolved. Members of these two groups of proteins, both of which are able to sequester calcium phosphate (CaP) in soluble and bioavailable forms, are involved in every aspect of biomineral utilization by vertebrates [4,5,6,7]. Consequentially, numerous studies have explored the mineral composition and protein content of CaP-containing particles derived from human and animal milk and sera, as well as similar particles sourced from other biological fluids (e.g., amniotic fluid, cerebrospinal fluid, urine, ascites and synovial fluid) [4,5,6,7,8,9,10,11,12,13,14,15,16,17,18].
A second enigma arises when one compares the composition of the calciprotein particles (CPPs) that are found in breast milk with the particles that circulate in the sera of normal subjects. Sequestered nanoparticles that have been isolated and analyzed from milk contain magnesium, a non-proteinaceous stabilizing agent that slows maturation of amorphous calcium phosphate (ACP) to more crystalline forms. A review by the EuroSoftCalcNet COST Action consortium emphasizes the central role that calcification inhibitors such as sequestering proteins and pyrophosphate play in preventing vascular calcification, one of the most serious consequences of CaP maturation and deposition in soft tissues [19]. If fetuin-A sequestered ACP is the form that circulates in sera, the apparent absence from the complex of non-proteinaceous ACP-stabilizing agents such as pyrophosphate or magnesium is puzzling. Recent studies by Gelli et al., Babler et al., and Powell et al. suggest that the magnesium ion may constitute an important stabilizing agent for sequestered CaP nanoclusters [6,17,20]. The potential role of such stabilization of CPPs in serum will be discussed in Section 5.8.
2. The Problem: Calcium Phosphate Solubility
Solubility (or its lack) is one of the most important properties of calcium phosphate (CaP) salts. For example, the solubility of these salts determines the course of many chemical reactions, such as dissolution, precipitation, hydrolysis, and phase transition. From a biological perspective, solubility plays a major role in processes such as transport, deposition, and the formation or resorption of hard surfaces. As the data in Table 1 show, solubility products [Ksp; calculated on the basis of ion concentrations or ion activities at specified values of solution pH (column 4 or columns 5 and 6, respectively)] tell us that combining calcium ion and phosphate ion in aqueous solutions at or above neutral pH incurs high probability that CaP will precipitate.
2.1. Reactions of Calcium Ion and Phosphate Ions in Aqueous Solution
Synthetic hydroxyapatite has been studied extensively as an analog of bone and tooth mineral. Recent reviews by Combes and Rey and Edén provide a detailed, description of synthetic reactions providing “hydroxyapatite” and other calcium phosphates and extensively cite and analyze studies that have been reported in the past [24,25].
In general, the reactions of calcium ion and phosphate ions are studied by rapidly mixing an aqueous solution of calcium chloride of known concentration with a second aqueous solution of sodium or potassium phosphate of known concentration. Water of high purity (e.g., “Milli-Q” water) is used as the solvent. Frequently the concentrations of each solution are adjusted to mirror the molar ratios of each ion in a geologic calcium phosphate mineral (e.g., monetite, brushite, hydroxyapatite). The pH of the phosphate solution is adjusted to a known value prior to mixing and then is monitored as reaction takes place. The temperature is maintained within a predetermined range as well. A precipitate forms rapidly under these conditions. At various timepoints, the concentrations of ions remaining in solution are determined using atomic absorption spectroscopy (calcium) and chemical reactions (phosphate). In addition, electron micrography is used to characterize the structures of the calcium phosphate precipitates at the same timepoints. Mathematical calculations (e.g., calculations considering thermodynamics, ion activities, and/or solubility) are typically based on the concentration/activity of the calcium ion and the concentrations/activities of each of the three phosphate ions (H2PO4-, HPO42-, and PO43-) at each value of solution pH.
For example, Christoffersen et al. found that mixing equal volumes of 0.02 M CaCl2 and 0.015 M K2HPO4 (pH 7.4) at 37 °C yielded a solution having a pH of ~6.4 that formed a flocculent precipitate (amorphous calcium phosphate 1; ACP1) within about 1 minute [26]. The precipitate was not stable and converted within 5 minutes to a second, more spherical amorphous calcium phosphate (ACP2) having ten-fold lower solubility (Ks) than ACP1. At the same time the solution pH decreased to about 5.7. Like the first precipitate, the second precipitate was labile and converted within hours to a third, slightly less soluble phosphate, identified as octacalcium phosphate or OCP. A solution pH of 5.4 was observed as the transformation to OCP took place.

Table 2.
Calcium Phosphates Isolated by Precipitation [26].
Table 2.
Calcium Phosphates Isolated by Precipitation [26].
| Calcium Phosphate | Empirical Formula | Molar Ratio Ca/P | -Log Ks | pH |
|---|---|---|---|---|
| Amorphous calcium phosphate 1 | - | 1.35 | 10.6 | ~ 6.4 |
| Amorphous calcium phosphate 2 | - | 1.35 | 11.5 | ~ 5.7 |
| Octacalcium phosphate | Ca8H2(PO4)6 ∙ 5 H2O | 1.35 | 11.7 | ~ 5.4 |
Studies such as these inform us of changes that occur in well-defined solutions similar to those of Christoffersen et al. Although no empirical formulae are provided for ACP1 or ACP2, it is likely that each precipitate is extensively hydrated. Lenton et al. have proposed an empirical formula for ACP of “Ca(PO4)0.52(HPO4)0.22” [27]. If this formula corresponds to ACP2, the pH profile suggests that the transition from ACP1 to ACP2 included loss of protons from some bound HPO42- anions and conversion to bound PO43- anions, a change which would contribute to both the decrease in solution pH that was observed and the significantly lower solubility of ACP2. In addition, investigators have hypothesized an increase in density in conversion of ACP1 to ACP2, an increase which may correspond to loss of bound water from the complex and its components. Finally, it should be noted that small variations in experimental parameters (e.g., stirring rate, particle size, nature of buffer, presence of dissolved carbonate from reagents and air, concentrations of reagents, washing the precipitate with water) have resulted in independent reports of a range of more-or-less acidic ACPs having seemingly distinct empirical formulae and structures. Few of these formulae correspond exactly to the formulae of defined CaP mineral salts [e.g., brushite (CaHPO4 ∙ 2 H2O); octacalcium phosphate (Ca8H2(PO4)6 ∙ 5 H2O); apatite Ca10(OH)(PO4)6].
Both the mechanisms of initial nucleation and the structures of the CaPs that are formed in these reactions remain controversial. Pioneering investigations that were carried out in amine-containing buffers indicated that an amorphous calcium phosphate (ACP) phase formed as a precursor to the precipitation of crystalline hydroxyapatite (HA) [28]. Kinetic analysis suggested that the conversion involved the dissolution of ACP followed by the first-order, autocatalytic propagation of HA crystals at a rate proportional to the amount of crystalline material already present. The sigmoid curves describing this transition exhibited an induction period in which no change in crystallinity was observed, followed by a rapid proliferation period to crystalline HA that gradually tapered to a period of little change. Since many of the conversions are endothermic, additional heating at elevated temperatures alters the characteristics of the final product, as does the addition of alcohols or poly(ethylene) glycols [25,29]. As noted above, detailed, descriptive analyses of many of these investigations are available, and they will not be discussed further herein.
2.2. Maturation
The mineral phase of bone and teeth is typically described as (crystalline) hydroxyapatite, one of the least soluble but most thermodynamically stable forms of calcium phosphate. However, under physiological conditions, calcium phosphate does not precipitate directly in this form [25,30]. It is now generally recognized that the crystallization of many sparingly soluble salts, including calcium phosphates, involves the formation of metastable precursor phases that subsequently and repeatedly redissolve and then reprecipitate as less soluble and more crystalline forms of the salt [31]. During these transitions, the salt passes through several unstable or metastable stages from the most soluble to the least soluble. The kinetics of ripening are dramatically dependent on factors such as pH, temperature, ionic strength, viscosity of the medium, and molecules and ions in solution. The process is an example of the Ostwald Rule of Stages or Ostwald “ripening” [31,32,33,34].
Ostwald ripening is not the only mechanism of maturation that has been considered [35]. A second mechanism involves the reorganization of Posner’s clusters through accretion of other clusters. As the cluster size grows, a simple reorganization involving minimal activation energy could provide the seed crystals of apatite needed for maturation. A third mechanism considers hydrolysis of phosphate on the surface of the agglomerate, resulting in nucleation of apatite, with crystal growth supported by surface ion migration.
What experiments such as those described in the preceding sections do not emphasize is the consequences of maturation of amorphous CaPs into crystalline materials. This change is important to recognize, as a transition to crystallinity reduces solubility and increases physiological toxicity, as discussed below.
2.3. Calcium and phosphate status in biofluids
Calcium and phosphate ions behave as ideal reactants (i.e., simple ions) only in very dilute aqueous solutions of soluble antecedent salts of non-chelating counterions that are formed and maintained under an inert atmosphere. In stark contrast, however, a biofluid such as serum is a complex environment holding millimolar concentrations of each ion. Moreover, the serum concentrations of either or both ions are susceptible to increases responsive to dietary loading (e.g., after eating or use of supplements) [36]. Thus, the ions do not react as independent entities. Instead, they form soluble ion pairs with each other [e.g., (CaH2PO4)+ and CaHPO4] and with counterions such as citrate and other hydroxyacids having a high affinity for the calcium ion [37,38,39]. The ion pairs convert within a few nanoseconds to thermodynamically unstable nanoclusters of amorphous calcium phosphate (ACPs). If magnesium ion is present, it serves as a pro-aggregant and provides a central ion in the formation of more soluble colloidal nanoclusters of amorphous magnesium calcium phosphate (AMCPs) [40]. Likewise, as the initial phosphate aggregates transform into less soluble calcium phosphate precipitates, carbonate ions may be incorporated into the solid. Sequestration of ACP and/or AMCP nanoclusters before they undergo conversion to less soluble and more crystalline phosphates is key to the physiological solution to calcium phosphate management.
3. The physiological solution to mineral management
Controlled transport and distribution of calcium and phosphate are indispensable for cellular metabolism. Nearly all physiological fluids have the potential to become saturated or locally supersaturated with calcium and phosphate ions. Natural biofluids, therefore, require some means for stabilizing these ions in solution and preventing their inappropriate precipitation and/or crystallization. Current reports point to sequestration by intrinsically disoriented proteins (IDPs) as the predominant mechanism for providing CaP stabilization in biofluids.
Intrinsically disordered proteins (IDPs) comprise a group of soluble proteins that act as sequestration agents [4,27,41,42,43,44]. The biological significance of IDPs has prompted establishment of a curated IDP database (DisProt; url: https://disprot.org) that is growing in parallel with increases in the importance of this vital field of study [45]. Recent reviews by Boskey, Holt, and Lenton describe extracellular IDPs in greater detail than will be provided here [27,41,43]. Another recent review by Wright and Dyson discusses intracellular IDPs and their roles in cellular signaling and regulation [44].
Two characteristics of IDPs illustrate their significance to physiological calcium phosphate management. The first characteristic relates to their flexibility in solution. The second characteristic reflects the loss of flexibility and induced folding upon binding of ligands [46,47]. Thus, IDPs are characterized by a lack of fixed conformation in solution. Their flexibility allows them to rapidly transform through an ensemble of interchangeable structures as well as rapid sampling of a large conformational space. The result is an acceleration of protein-ligand interactions [48]. In the sequestration of ACP or AMCP nanoclusters, this flexibility provides a mechanism through which the protein can rapidly react with nascent aggregating clusters of phosphates to prevent or slow the irreversible formation of more crystalline phases. Slower sequestration resulting from a low affinity of binding, a low concentration of sequestrant or a more rigid conformation would be expected to result in the irreversible formation of crystalline, or poorly crystalline, calcium phosphates exhibiting greater toxicity [49,50].
The loss of flexibility on binding of ligands has several beneficial consequences. For example, sequestration of amorphous calcium phosphate nanoclusters by the IDP inhibits their conversion to less soluble and more crystalline materials. Likewise, upon ligand binding, the structure of the IDP changes in ways that envelop the nanoclusters in a structured protein environment (e.g., a beta sheet in fetuin-A) [51]. Although the overall structure may be described as globular, nanocluster sequestration does not require extensive association of protein segments with the CaP core. In contrast, after sequestration, other polypeptide segments in the IDP may open to interaction with unrelated proteins and soluble moieties in the local milieux, creating a multilayered complex. The resulting nanoparticles are dispersed in biological fluids as soluble colloids.
Inhibition or nucleation properties are not intrinsic to a given IDP. Current data show that an IDP can act in various ways, depending on its concentration, other solution conditions, and its degree and pattern of phosphorylation.
A review by Lenton et al. summarizes knowledge about the structural biology of CaP nanoclusters sequestered by phosphoprotein and phosphopeptide IDPs [27]. In this review, general details about the major proteins that act as sequestrants in milk and serum are presented. Additional details about sequestration in other biofluids and tissues are being defined in a growing body of publications summarizing recent research.
4. IDPs, the physiological sequestration agents in milk
Sequestration of CaP nanoclusters by phosphoproteins and phosphopeptides is widely recognized, and a brief description of a well-studied sequestrant in milk will serve as an example. Investigators have reported and confirmed that milks (including human milk) contain colloidal casein micelles which embed calcium phosphate [41,52,53,54,55,56,57,58]. As a result, milks contain much higher calcium and phosphate concentrations than are permitted by the solubility of inorganic calcium phosphates at the pH of milk (pH 6.5-6.7). Differences in mineral content and casein phosphorylation exist among species [59,60,61]. However, studies of donkey, bovine and human milk provide general information about casein micelle structure and composition as well as mineral distribution in this class of biofluid.
Caseins have been extensively studied in cow’s milk, where they exist as large aggregates of colloidal size known as casein micelles. Although only bovine casein micelles have been studied in detail, their general characteristics may apply to other vertebrate milks as well [41,59,60,61]. For example, a typical bovine milk casein micelle having a 100 nm radius contained about 104 casein molecules (~50 phosphate centers) and 800 nanoclusters of amorphous calcium phosphate [61]. These micelles were not assembled in the “hydrophilic heads out, hydrophobic tails in” typical of amphiphilic molecules in aqueous solutions [62]. Rather, the initial fibrils resulted when the α- and β-casein fractions, which are heavily phosphorylated, interacted and bound strongly with Ca2+ ions, leading to the formation of amorphous nanoclusters of magnesium calcium phosphate embedded in the protein. Then nanocluster-laden caseins aggregate into micelles which are surface-stabilized by κ-casein, which prevents agglomeration. Cryo-transmission electron microscopy (TEM) investigation on casein micelles revealed that the magnesium-containing CaP nanoclusters embedded therein are ∼2.5 nm in size and uniformly distributed in a homogeneous protein network [41]. Small angle scattering investigations revealed a similar cluster size (2 nm) and a correlation length of 0.35 nm-1, which indicates that each AMCP nanocluster is, on average, 18.6 nm apart from the other in bovine casein [41].
The mineral contents in milks from various species have been widely studied as well. As expected, due to variations in maternal nutrition, the mineral content in human milk is widely variable. What is remarkable, however, is the relative invariance of the Ca:P ratio among the species (1.4-1.5) and the consistent presence of millimolar magnesium, an ion known to stabilize sequestered amorphous calcium phosphate nanoclusters.
Table 3.
Mineral content of whole milk.
| Species | Mineral content, mmol/L | Ca:P Ratio | Source | ||
|---|---|---|---|---|---|
| Ca | P | Mg | |||
| Donkey | 13.6 | 9.8 | 1.1 | 1.4 | Malacarne [62] |
| Donkey | 12.3 | 8.2 | 1.1 | 1.5 | Fantuz [63] |
| Cow | 13.0 | 8.8 | 1.1 | 1.5 | Fox [64] |
| Human (Early lactation) | 7.4 | 3.9 | 1.4 | 1.9 | Sanchez [65] |
| Human (Late lactation) | 6.3 | 3.9 | 1.4 | 1.6 | |
| Human (Early lactation) | 6.9 | (3.9) | 1.0 | (1.8) | Li [66] |
| Human (Late lactation) | 6.6 | (3.9) | 0.9 | (1.7) | |
| Human (Established feeding) | 6.7 | (3.9) | 1.5 | (1.7) | |
| Human | 7 | 4.7 | 1.3 | 1.5 | Sanchez [65] |
Note: The values in this table are means and do not reflect the reported standard deviations or the ranges of concentrations found. Phosphate values in parentheses are taken from Sanchez and the Ca:P ratios are calculated on this basis.
In order to determine the mineral distribution, whole milk may be centrifuged to separate fat and obtain skim milk. Skim milk may then be ultracentrifuged to separate a casein pellet from the whey (soluble) fraction. Lastly, the soluble supernatant may be ultrafiltered to obtain an aqueous phase. By this means the distribution of calcium, phosphorus, magnesium and other elements in the various phases may be determined. For example, Fantuz et al. applied the procedure described above to donkey milk samples collected from healthy lactating donkeys [63]. They found that the fat fraction carried very little or none of the investigated elements (calcium, phosphorus, sulfur, magnesium, potassium and sodium). As expected, the majority of calcium (62.9%) and phosphorous (53.1%) was associated with casein. In addition, a significant fraction of magnesium (32.6%) was also associated with the casein fraction. The whey/aqueous phase contained the remainder of these ions.
Table 4.
Percentage of total mineral associated with casein.
| Species | Mineral content in casein micelles (as a percentage of total mineral content in milk) | Reference | ||
|---|---|---|---|---|
| Ca | P | Mg | ||
| Donkey | 69.3% | 63.8% | 31.2% | Malacarne [62] |
| Donkey | 62.9% | 53.1% | 32.6% | Fantuz [63] |
| Cow | 66.5% | 57% | 33% | Fox [64] |
Thus, these data infer that complementary inhibition of ACP maturation by both casein proteins and the magnesium ion bound to the embedded nanoclusters slows the rate of maturation of the amorphous state and prevents CaP crystallization in milks.
Briefly summarized, this description of casein sequestration in milk suggests that the sequestering IDP may contain sequences of phosphorylated serines and/or threonines wherein exposure to protein-bound phosphate facilitates association with and initial binding to nascent ACP nanoclusters [67,68,69,70,71,72]. The resulting complex may contain a large mass of ACP and be quite thermodynamically stable, exhibiting little maturation to crystalline CaPs, even after exposure to heat, cold, drying and reconstitution, exposure to added salt or alcohol, or lengthy storage as liquids or solids.
Among other features of casein-sequestered CaP that are different from the chemical and thermodynamic descriptions of calcium phosphate synthesis reactions in water (Section 2.1) is the role that the magnesium ion plays in the complex. It is well known that an IDP does not act in isolation, and its effects are frequently altered by the presence of other ions and molecules in the biofluid, as well as environmental conditions (e.g., temperature and pH). Known agents that alter the stability of amorphous calcium phosphates embedded in an IDP include magnesium, pyrophosphate, strontium, zinc, citrate, phosphocitrate, ATP and the magnesium ATP complex, carbonate, fluoride, bisphosphonates, microRNAs, and DNA fragments. (This list is not all-inclusive and includes species which alter the growth of crystalline calcium phosphates as well.)
5. CaP Sequestration in serum
Sequestration of ACP nanoclusters by serum proteins provides a stark contrast to the sequestration by caseins that is observed in milk. α2-Heremans-Schmid glycoprotein (fetuin-A or AHSG; accession no. P02765) has been identified as the major sequestrant in serum [73].1 (In the blood of healthy adults, fetuin-A has a concentration of 0.5-1.0 g/L [15]. Its concentration is higher in neonates [74,75].) Fetuin-A is a type 3 cystatin protein family member of about 56 kDa that is secreted by the liver [51]. The protein circulates as a disulfide-bonded protein having three cystatin D domains. Although both in vitro and in vivo studies in mutant mice suggested that fetuin-A sequestered ACP nanoclusters in serum, the absence of phosphorylated sequences in this protein required mutational analysis of the protein to identify the structural features that enabled ACP sequestration [73]. Examination of the structure-function relationship of recombinant forms of fetuin-A with similar proteins from the cystatin superfamily (e.g., fetuin-B, histidine-rich glycoprotein, and kininogen) suggested that sequestration involves acidic amino acid residues in an EF-hand-like sequence on an extended beta-sheet of the cystatin-like fetuin-A domain D1, located near the amino terminus. Upon association with calcium ion in CaP nanoclusters, these residues fold into a defined array that effectively binds calcium without disrupting the nanocluster. Ligand-binding also involves addition of a second fetuin-A molecule, yielding a dimer that is a combination of a heavy- and a light-chain fetuin. Binding results in a structural reorganization that envelops the bound nanoclusters in beta-sheets while opening other parts of the protein to interactions with other moieties in the biofluid. A globular complex having a diameter of about 10 nm results. Heiss et al. suggest that the enveloping protein serves as a diffusion barrier that limits further growth of the ACP nanocluster and its maturation into crystalline CaPs while preventing CaP precipitation [73]. (Interestingly, a substantial fraction of the fetuin-A present in human plasma is serine phosphorylated. Phosphorylation changes the activity of the protein from that of a sequestering agent that controls the transport and distribution of calcium and phosphate to a protein that negatively regulates the insulin receptor tyrosine kinase [76].)
In summary, this description of fetuin-A sequestration of calcium phosphates in serum suggests that exposure of the calcium ion in nascent clusters of amorphous calcium phosphates (i.e., Posner’s clusters) facilitates association with and initial binding to aspartate and glutamate residues in a beta-sheet conformation of cystatin domain D1 (Figure 1). Binding of up to six calcium ions per molecule in the stabilizing dimer does not induce crystallization but increases the size of the resulting calciprotein monomer (CPM) to about 8-10 nm [77]. In contrast to casein-sequestered clusters in milk, the resulting complex contains a small mass of ACP and is quite metastable. Continuing circulation in serum allows maturation to crystalline CaPs.
Among other features of fetuin-sequestered CaP that are different from the chemical and thermodynamic descriptions of casein-sequestered CaP is the role that the magnesium ion plays in the complex. Although in vitro synthetic studies indicate that magnesium and pyrophosphate stabilize the CaP clusters embedded in calciprotein monomers, the inability to detect the CPMs reliably leaves the physiological importance of these stabilizing agents in fetuin-A sequestration an open question.
5.1. Sequestration allows calcium and phosphate transport and distribution throughout the body.
CPMs in serum consist of flexible spherical colloids that are sufficiently small to circulate both in larger vessels of the circulatory system and significantly smaller capillaries, such as those found in the kidney, liver, bone and spleen. The amorphous calcium phosphate mineral phase remains well-hydrated and in a viscous liquid- or gel-like state [78]. The particles readily transfer amorphous calcium phosphate nanoclusters to other templates or are acidified, releasing calcium and phosphate within the lysosomes of endothelial cells and/or scavenging macrophages in the liver and spleen; in proximal tubules of the kidney; or at bone resorption sites.
5.2. Sequestration allows rapid clearance of CaP as calciproteins.
Details of the mechanisms of nanoparticle clearance from serum continue to be delineated. Koeppert et al. synthesized CPM containing fluorescently labeled CaP nanoclusters and found that, in mice, circulating CPM were filtered within 10-20 minutes by the kidney glomeruli into the Bowman space and the proximal tubules [79,80]. This observation correlates with the 60-70 nm diameters of fenestrations in the glomeruli and peritubular renal capillaries that would allow efficient filtration of CPMs but not larger CPPs [81]. Koeppert also noted that uncomplexed fetuin-A was not absorbed at these sites in the kidney, an observation that indicates electrochemical selectivity of absorption by the kidney, since both the surface of glomerular endothelial cells in the kidney and fetuin-A are negatively charged [80].
Larger CPPs that are not filtered by the kidney are rapidly cleared by sinusoidal endothelial cells in the liver and macrophages of the reticuloendothelial system [80,82]. For example, sinusoidal endothelial cells of the liver, which exhibit fenestrations with a flexible diameter of 100-113 nm, would be expected to readily accommodate and clear CPP-1 particles having diameters in this range (50-100 nm) [81]. Indeed, Koeppert et al. reported that liver sinusoidal endothelial cells cleared CPP-1 nanoparticles [80]. Similarly, Herrmann et al., who studied the clearance of (larger) CPP-2 calciprotein particles (120-150 nm) in mice, found that the particles exhibited a t1/2 of about 45 minutes and were cleared by Kupffer cells of the liver and marginal zone macrophages of the spleen [82]. (These cells also clear aged blood cells, LDL particles, cell remnants, platelet fragments, and particulate matter, an observation which may explain why atherosclerotic plaques frequently calcify, especially in patients with perturbed mineral homeostasis.)
5.3. Additional changes during circulation of CPPs
If CPMs are not efficiently cleared, the calcium phosphates enveloped in these particles undergo ripening, a process which is influenced by a number of factors such as the concentrations of fetuin-A, Ca2+, magnesium (Mg2+), phosphate (Pi), as well as the viscosity, temperature and pH of the surrounding microenvironment. Ripening has two physical effects: (1) the particles increase in size and density, changes which reduce their solubility; and (2) the bound CaP becomes more crystalline. These properties may be responsible for the pro-inflammatory nature of CPP-1 particles [79,80].
During circulation, larger CPP nanoparticles continue to sample the seral environment. As a result, analyses of CPP isolated from healthy subjects show the presence of small percentages of albumin, matrix Gla-rich protein, and lipoproteins such as apolipoprotein A1. Closer examination of these larger CPPs shows that these nanoparticles exhibit a prodigious ability to bind and carry organic species. Wu et al. carried out a comprehensive analysis of organic components found in nanoparticles derived from body fluids [83]. Based on biological staining, fluorescent tagging, proteomics and metabolomics, their results indicate that the nanoparticles bind to calcification inhibitors, complement proteins, coagulation factors, glycoproteins, apolipoproteins, protease inhibitors, amino acids, carbohydrates, polysaccharides, phospholipids, lyso-phosphatidyl cholines, mono-acyl glycerols, amides, fatty acids, DNA, and low molecular weight metabolites. In a separate study, Kumon et al. reported that oxidized acidic lipids associated with the sequestered nanoparticles and served as scaffolds for development of carbonate-apatite crystals [84].
5.4. Sequestration enables controlled transfer of calcium and phosphate to other templates.
Current data suggest that IDPs and their sequestered loads may play a role in cell-matrix adhesion and resulting transfer of the sequestered materials. For example, in biomineralization, sequestration may enable: the assembly of protein monomers into supramolecular structures that organize the mineral phase over multiple length scales [85]; crowding-induced modulation of mineralization and confinement-mediated transient stabilization of mineral precursors within pores of organic networks [86]; phase separation of liquid- or gel-like organic phases that modulate mineralization [87,88,89]; mechanical reinforcement of hybrid materials via inter- and intracrystalline occlusion of bioelastomers [90]; and prospective regulatory checkpoints arising from a predisposition of redundant sequences toward effective post-translational alterations [91].
In addition to these interactions, amorphous calcium phosphates, with or without magnesium centers, that are embedded in an IDP retain their solubility and remain in equilibrium with their constituent ions. As a result, both calcium and phosphate retain their ability to circulate in the biofluid and then transfer to other templates or morph into a more crystalline calcium phosphate while continuing to circulate in the biofluid [92]. The nanoclusters dissociate in slightly more acidic environments than serum (pH 7.4), providing calcium and phosphate ions as well as any other species associated with the nanocluster. Moreover, the dissociation may take place at or near a structured cellular macromolecule that supports crystal nucleation (e.g., another IDP or type 1 collagen fibrils) and that macromolecule may concurrently take on regulatory control of the transformation of amorphous calcium phosphate clusters to crystalline salts as well as directing the crystal structure that predominates in the new environment [93,94,95].
5.5. Sequestration provides bioavailable calcium and phosphate supporting numerous physiological functions.
The primary function of sequestration is one of transporting calcium phosphate in a soluble form that can readily be used physiologically without introducing or exacerbating toxicity. That being said, the nanoclusters perform numerous ancillary physiological functions.
For example, Holt and his colleagues have described the unique nutritional value of casein-embedded nanoclusters in milks [41]. According to Holt, the ability of casein to bind AMCP nanoclusters has three-fold biological function: (i) the control of calcium phosphate precipitation in milk through AMCP sequestration, (ii) the suppression of toxic amyloid fibril formation that casein would produce in the absence of AMCP, and (iii) the provision of bioavailable calcium phosphate for nutrition of the neonate. The first two functions are related to the mother and her ability to lactate: pathological calcifications or amyloidosis of the mammary gland would threaten the wellbeing of both the mother and the neonate. The third function provides the neonate with a rich source of calcium and phosphate during periods of extensive skeletal and tooth development. Finally, the observation that sequestered calcium phosphates may carry materials of microbial origin has renewed study into the role of breast milk in the building of an effective immune system in newborns. [5,83,84].
In serum, amorphous calcium phosphate nanoparticles templated on fetuin-A provide means for transportation of calcium and phosphate in soluble form to kidney, bone, skin, and other tissues throughout the body [7,9,77]. CPM nanoparticles having diameters of about 10 nm readily transfer the ions to appropriate sites in kidney or bone, for example, or are cleared within minutes by endothelial cells in the kidney as well as sinusoidal vessels of the liver and spleen and macrophages in these organs [80]. Under normal circumstances, the system works efficiently and effectively to limit the toxicity of insoluble, crystalline material and to prevent sustained disturbances to mineral metabolism that may be injurious (e.g., ectopic calcification and downstream cardiovascular sequelae; organ morbidity; aneurysms; idiopathic preterm birth) [96,97,98,99].
In a series of experiments using human tissue, animal models and advanced imaging techniques, Powell and his colleagues have provided a detailed picture of roles that calcium phosphate nanoparticles play in immune surveillance [20]. The lumen of the small intestine of humans contains as many as 1014 porous nanoparticles composed of amorphous magnesium calcium phosphate [100,101]. The sequestering agents have not been identified. Gelli has speculated that mucin coats the nanoclusters [102]. If so, this glycoprotein provides a viscous environment in which the particles remain stable but exposed to the environment, i.e., a “protein corona”. Once formed, the particles bind other moieties in the intestinal environment, presenting them as sequestered entities for uptake via M cells in Peyer’s patches. Within the cellular membrane, the particles are exposed to antigen-presenting cells (mainly dendritic cells and macrophages), which assess the “self” or “non-self” characteristics of the materials borne by the nanoparticles. The minerals dissolve and are available for transport to the serum.
In addition to their roles as transporters and distributors of calcium, magnesium, and phosphate, CPM may interact with osteocytes in bone to induce release of fibroblast growth factor 23 (FGF23) [103,104,105]. Proof-of-principle studies by Akiyama et al. showed that induction of FGF23 mRNA in bone and subsequent elevations of FGF23 in serum were preceded by a rapid but transient increase in calciprotein particles [103]. The observation that administration of the bisphosphonate alendronate to animals fed a high-phosphate diet also augments total FGF23 concentration infers that small calciprotein particles containing some crystalline phosphate may also serve as strong inducers of FGF23 secretion in vivo [106].
Finally, Martel et al. have noted that similar amorphous calcium phosphate nanoparticles enable disposal of excess minerals that are remnants of calcium homeostasis [107]. Thus, small crystals are often detected in the urine of healthy individuals. Higher numbers of crystals having larger sizes are found in the urine of individuals who are kidney stone formers.
5.6. Changes in Sequestered Entities in Health and Disease
As they circulate, calciprotein particles undergo changes in composition in addition to maturation of CaP. A recent study by Smith et al. provides intriguing information about calciprotein particle transformations in health and disease [108]. These investigators carried out a detailed physiochemical and biochemical comparisons of endogenous CPP isolated from uremic sera with CPP synthesized in vitro from both uremic sera and sera from healthy subjects. Their data showed that calciprotein monomers consist of amorphous calcium hydrogen phosphates and absence of crystallinity. However, as the particles grow in size, crystalline hydroxyapatite features were added to the TEM micrographs. During ripening the particles lose surface charge (reduced zeta potential) and exhibit an increased tendency to aggregate. In a subsequent step, primary CPP may aggregate into colloidal nanoparticles having a more prolate structure and long-axis diameters of 100-250 nm (secondary CPP or CPP-2). The calcium phosphates in CPP-2 transitioned into ordered, more thermodynamically stable crystalline phases having compositions similar to hydroxyapatite. The crystalline mineral may be organized as densely packed, needle-shaped lamellae or as plates. Both crystalline forms are cytotoxic. CPP-2 exhibit pro-inflammatory properties and induce the expression and secretion of TNF-α when exposed to macrophages in vitro. CPP ripening was accompanied by significant enrichment in organic moieties. Quantitative proteomic analysis revealed binding of various proteins from the extracellular space, including transporters, peptidases, mineral-binding proteins (e.g., osteopontin, bone sialoprotein, collagen 1 alpha chains, and osteonectin), as well as enrichment for the smaller soluble/exchangeable apolipoproteins (ApoA1, ApoA4, ApoE, and ApoC3) and components of the complement system. Small amounts of lipids were detected, albeit at much lower concentrations than those in serum. Lipidomic profiling showed a predominance of cholesterol as well as low percentages of long-chain fatty acids. Nucleic acids were not detected in calciprotein monomers, but after ripening and hydroxyapatite formation, both DNA fragments (<200 bp) and small RNA (<200 nt) were detected. Likewise, the larger CPP-II nanoparticles carried microbe-derived components (e.g., hydroxylated fatty acids, peptidoglycan, and bacterial DNA).
In a separate study, Aghagolzadeh et al. generated calciprotein particles in vitro and studied their interaction with vascular smooth muscle cells (VSMC) in culture [109]. Their data supported a ripening process in which calciprotein particles transformed into larger particles (CPP-2) exhibiting increased calcium and phosphate content. As the transition proceeded, the number of particles did not change significantly but CCP-1 particles lost their spherical shape and became elongated, spindle-shaped particles with distinctly less solubility in the medium. At the same time, sequestered calcium phosphates became more crystalline, a change which has been shown to drive calcification. Exposure of viable VSMC to CPP-2 particles but not CPP-1 particles led to a pronounced and consistent accumulation of intracellular calcium within one day. Calcified cells increased oxidative stress as evidenced by release of soluble TNF-α, increased extracellular H2O2, and up-regulation of bone morphogenetic protein-2 and Nuclear Factor Kappa-B. Accumulation of CPP-2 particles led to early apoptosis.
5.7. Sequestration protects cells from toxicities associated with CaP crystal deposition.
Crystalline forms of calcium phosphate become two-edged swords: the crystals exhibit reduced solubility and ability to dissolve in acidic environments, as well as the increased toxicity exhibited by crystalline polymorphs of calcium phosphate [49,50,97]. While these changes are beneficial properties of mature bone and tooth enamel, each is a severe drawback when the body attempts to use the crystals for bone restoration, dissolve the crystals in the acidic lumen of renal proximal tubules for calcium and phosphate uptake, or to process crystalline calcium phosphate that enters lysosomes.
Crystal deposition is known to trigger inflammation and injury via three pathways: (1) via the NOD-, LRR- and pyrin-domain-containing protein (NLRP3)-inflammasome and caspase-1-mediated secretion of IL-1β and IL-18, as well as to gasdermin D-mediated pyroptotic cell death [110]; (2) by inducing necroptosis, a receptor-interacting serine-threonine kinase-3 (RIPK3)-dependent and mixed lineage kinase domain-like (MLKL)-dependent form of regulated cell necrosis [111]; and (3) by inducing mitochondrial dysfunction and mitochondrial permeability transition (MPT)-related cell necrosis [112]. Several recent studies have compared the effects of deposition of CaP or calciprotein particles on cells in culture or in tissues after in vivo exposures.
Dautova et al. found that CaP crystals were taken up within minutes of exposure by micropinocytosis, membrane invagination, or plasma membrane damage of vascular smooth muscle cells [113]. Exposure activated cell repair and survival mechanisms, which delayed cell death until 30 minutes after exposure. In contrast, fetuin-A-sequestered CaP particles were taken up more slowly and caused less damage to cellular membranes. Lysosomal dissolution of the particles was also slowed.
Kunishige et al. carried out similar experiments in renal proximal tubular epithelial HK-2 cells to compare the cytotoxicity induced by CaP and CPPs [96]. After cellular uptake, CaP crystals dissolved in the acidic lysosomal environment, dissolution that caused osmotic swelling and lysosomal membrane permeabilization and generation of reactive oxygen species (ROS) as a result of mitochondrial dysfunction. In contrast, CPPs, which were taken up by caveola-mediated endocytosis or macropinocytosis, increased the osmotic pressure inside lysosomes as they dissolved but failed to induce lysosomal membrane permeabilization. An increase in intra-lysosomal pH by ~ 1 unit was observed, which slowed hydrolase activity and inhibited the fusion of autophagosomes with CPP-containing lysosomes, resulting in autophagic flux blockage. Adverse changes in autophagy are known to enhance matrix vesicle release, which in turn, increases the area of tissue subject to damage [114]. Another significant difference reported by Kunishige may be the effect that CPPs had on cholesterol trafficking. As Smith et al. reported earlier, CPPs adsorb cholesterol [108]. Therefore, as the CPPs moved through the plasma membrane, membrane cholesterol content decreased by adsorption to CPPs. Lysosomal processing allowed transfer of the CPP-bound cholesterol to the lysosomal membrane. These changes in cholesterol content may have had differential impact on the signaling induced by lipid raft formation at the two sites, by reducing the repair of defects in the plasma membrane and rendering the cells more vulnerable to damage [115].
Koeppert et al. have studied the pro-inflammatory potency of CPM, CPP-1 and CPP-2 particles [80]. Synthetic fluorescent CPM were injected in mice, and particle clearance was monitored by live two-photon microscopy. Organ sections were analyzed by fluorescence microscopy to assess CPM distribution. Cellular clearance and cytotoxicity were analyzed by flow cytometry and live/dead staining, respectively, in cultured macrophages, liver sinusoidal endothelial cells (LSEC), and human proximal tubule epithelial HK-2 cells. Inflammasome activation was scored in macrophages. Fetuin A monomer and CPM charge were analyzed by ion exchange chromatography. Experimental data showed that CPP-1 predominantly induced NLRP3 inflammasome assembly and subsequent cytokine secretion, while CPP-2 predominantly stimulated immediate secretion of preformed TNF-α. In a subsequent study designed to assess the pro-inflammatory potency of CPM, this group used immortalized ASC-GFP macrophages to study the responses to calcium content-matched (2.5 mM) CPM, CPP-1, and CPP-2. CPP-1 triggered inflammasome assembly within 2 h, while CPP-2 triggered delayed inflammasome assembly after 8 h, as reported earlier [81]. In contrast, CPM containing identical 2.5 mM calcium and 2 mM phosphate did not trigger assembly of the inflammasome at any time during the 24-hr observation period, suggesting high stability of the fetuin-A/calcium-phosphate complex and concomitant low inflammatory potential of CPM compared to Ca, CaP, or CPP-1.
5.8. Fetuin-A Sequestered ACP and Phosphate Toxicity
Phosphate toxicity has long been associated with vascular calcification (VC), ectopic calcification which characterizes the arteriosclerosis of aging, as well as chronic kidney disease, diabetes mellitus, dyslipidemia, and hypertension [116]. Evidence that the sequestered matrix protects from inappropriate calcium phosphate (CaP) deposition and related toxicity in vivo is largely indirect and drawn from studies of vascular calcification in cell cultures or animal models. In many studies, deficiencies in one or more of the sequestrants or inhibitors of crystallization have been implicated in increased risk and incidence of vascular calcification. Shafer et al. were among the first to report that fetuin-A deficiency (and thus a lack of stable CPM and CPP) results in ectopic calcification [18]. In a subsequent report, this group noted that fetuin-A deficiency in mice led to myocardial stiffness, cardiac remodeling, and diastolic dysfunction [117]. Villa-Bellosta and O’Neill summarized knowledge about inhibitors of crystallization such as endogenous pyrophosphate and mechanisms by which this anion acts endogenously to inhibit calcification [118]. The review also highlighted metabolic pathways leading to deficiencies that played a key role in vascular calcification. The benefits of supplementation with inhibitors have also been reported. By way of example, Villa-Bellosta and Sorribas reported the prevention of vascular calcification in rat aorta by addition of extracellular polyphosphates and nucleotides [119].
Herrmann and coworkers elegantly demonstrated the consequences of loss of fetuin-A in a recent study comparing calcification in D2 wildtype mice vis a vis calcification in mutant mouse strain D2,Ahsg-/-, a strain that combines fetuin-A deficiency with the calcification-prone DBA/2 genetic background [98]. The latter exhibit a particularly severe compound phenotype of microvascular and soft tissue calcification. Herrmann et al. analyzed mice longitudinally by echocardiography, X-ray-computed tomography, analytical electron microscopy, histology, mass spectrometry proteomics, and genome-wide microarray-based expression analyses. Fetuin-A-deficient mice had calcified lesions in myocardium, lung, brown adipose tissue, reproductive organs, spleen, pancreas, kidney and the skin, associated with reduced growth, reduced cardiac output and premature death. Importantly, early-stage calcified lesions presented in the lumen of the microvasculature, suggesting precipitation of mineral containing complexes from the fluid phase of blood. Genome-wide expression analysis of calcified lesions and surrounding (not calcified) tissue, together with morphological observations, indicated that the calcification was not associated with osteochondrogenic cell differentiation, but rather with thrombosis and fibrosis. Collectively, these results demonstrate that soft tissue calcification can start by intravascular mineral deposition causing microvasculopathy, which impacts on growth, organ function and survival.
Concurrently, the numbers of clinical investigators who recognize the benefits of adequate serum concentrations of fetuin-A have grown. Bäck et al. have recently published a consensus statement that summarizes current knowledge about a variety of endogenous calcification inhibitors, including fetuin-A, and their benefit in preventing vascular calcification [19]. In addition, Icer and Yildiren have summarized nine clinical studies in which the relationship between serum fetuin-A and coronary artery calcification (CAC) have been assessed [120]. Evaluations included between 42 and 2,505 participants and yielded results that suggested an inverse relationship between serum fetuin-A levels and CAC and/or its severity. Data were more compelling when participants had underlying diabetes or nephropathy. Data from two of the nine studies failed to establish a relationship between the protein and CAC. These authors also discuss human studies evaluating a relationship between serum fetuin-A and kidney disease.
If the description of fetuin-A sequestered ACP corresponds to the entities which circulate in normal sera, what role, if any, do endogenous stabilizing agents play in preventing maturation of the sequestered mineral to crystalline phosphates having lower solubilities and greater toxicity, including increased risks of ectopic deposition?
In 2019, Babler et al. suggested that serum fetuin-A, pyrophosphate, and magnesium constituted the three main calcification inhibitors in mice with a calcification-prone phenotype (i.e., Ahsg-/- mice having a DBA/2 genetic lineage) [17]. These investigators found that intravenous administration of bovine fetuin-A or pyrophosphate, as well as supplementation of chow by doubling or quadrupling its magnesium content, significantly reduced the extent of calcification in brown adipose tissues, kidneys, lungs and heart. Their report renewed interest in manipulating the properties of fetuin-A/ACP complexes to enhance their physiological benefits. We will focus attention on one mineral, the magnesium ion, having anticalcification properties which were highlighted by Babler et al. but not included in the EuroSoftCalcNet consensus statement [19].
Studies of potential stabilization of CaP aggregates by the magnesium ion (Mg2+ or in the discussion that follows, Mg) began concurrently with pioneering studies directed to more detailed analyses of bone structure, formation and maintenance and their synthetic parallels. Investigators recognized quickly that extending the induction period before the rapid transition to less soluble and more crystalline CaPs could be a beneficial property of CaPs in biofluids. For example, Boskey and Posner studied the kinetics of the conversion of ACP to HA at pH 8 and temperatures of 26.0°, 37.5°, and 48.0°C in the presence of different amounts of Mg [121]. Mg/Ca molar ratios in the range from 0.004 to 0.04 were deemed of particular interest, as these corresponded to the molar ratios found in bones and teeth. Kinetic analyses at each temperature provided sigmoid curves in which the induction period was extended incrementally as the molar ratio of Mg/Ca increased up to a molar ratio of 0.04 but the rapid transition to HA which followed displayed superimposable rates. The Mg content incorporated into the amorphous solid was directly proportional to the Mg content of the solution but the average particle size decreased as the Mg content increased. Interestingly, if preformed Mg-containing ACP was added to its parent solution, the induction period was extended but was shorter than the induction period for transition to HA in the parent solution. Since the calcium concentration fell and then increased over the induction period extended by magnesium, the authors suggested that Mg incorporated into the ACP was altering its “solubility.” Studies of solutions containing Mg/Ca ratios of 0.2 failed to show this change, although the induction period was incrementally extended and the average ACP particle size was the smallest of the ACPs that were synthesized. The final HA from all experiments contained very little Mg, and crystallite size was independent of Mg concentration in this range.
Holt et al. prepared amorphous calcium magnesium phosphates by precipitation from moderately supersaturated aqueous solutions at pH 7 [122]. Chemical composition of the precipitates was determined using ion chromatography, and X-ray powder diffraction, scanning electron microscopy, X-ray absorption spectroscopy, and Fourier transform infrared spectroscopy were used to characterize the solids. Two different synthetic approaches were used. Preparation of sample T1 involved mixing solutions of KH2PO4, Ca(NO3)2, and Mg(NO3)2 together at a pH of about 4.5 to yield final concentrations of 10 mM phosphate, 5 mM calcium, and 5 mM magnesium. Ionic strength during was maintained by adding 0.15 M KNO3 as a background electrolyte. Precipitation of CaP was initiated by raising the pH by addition of 0.5 M KOH such that during the formation of the initial gelatinous precipitate, the pH remained constant at 7.0. In contrast to sample T1, samples H1, H2, and H3 were prepared by mixing equal volumes of solution A (100 mM NaH2PO4 and 100 mM Na2HPO4) and solution B (4 mM CaCl2, 135 mM NaCl, and a concentration of 6 mM, 10 mM, or 14 mM MgCl2, respectively). The pH was raised to 7.0 by addition of 0.4 M NaOH, and the initial gelatinous precipitate was allowed to form while maintaining the pH constant at 7.0.
Holt’s data provide interesting contrasts to the results obtained by other investigators. None of the samples gave X-ray powder diffraction lines corresponding to crystalline forms of calcium or magnesium phosphates. Sample T1 was found to be amorphous calcium magnesium phosphate(s) overlaid with KNO3 crystals, and careful analysis of samples H1-H3 indicated co-precipitation of NaCl with amorphous calcium magnesium phosphates. In sample T1, about half of the phosphate groups were protonated. Analyses of samples H1-H3 indicated that the degree of protonation increased linearly with the magnesium content of samples and with the ratio of the Mg to calcium ion concentrations in the supernatant. Maturation in the presence of solutions containing Mg ion involved transition from amorphous calcium magnesium phosphates to brushite (CaHPO4 ∙ 2 H2O) rather than OCP.
Kibalczyc et al. provided additional insights into the effects of the Mg ion on CaP precipitation [123]. The experiments were carried out by rapidly mixing equal volumes of 0.02 M CaCl2 and 0.012 M or 0.015 M K2HPO4 having a pH of 7.4 at 30 ± 0.1°C. For experiments where Mg was present, MgCl2 was dissolved in the CaCl2 solution or was added rapidly in the form of 0.100 M MgCl2 solution at various times after mixing. Molar ratios of Mg/Ca ranged from 0 to 0.24 and included a molar ratio of 0.04 (as was studied by Boskey and Posner). By increasing the molar ratio of Mg/Ca from 0 to 0.24, the investigators observed incremental increases in the induction period for conversion of ACP1 to ACP2 from about 4 minutes to about 15 minutes. The pH profiles consistently showed that formation of spherical particles of the first amorphous precipitate (ACP1) was accompanied by a decrease in solution pH from 7.4 to ~ 6.4. The transition from the first amorphous precipitate to the second (ACP2) consistently exhibited a pattern of a small decrease and subsequent increase in pH, followed by a decrease in pH to ~ 5.5 as OCD formed. Both transitions were consistently endothermic.
Kibalczyc’s experiments in which Mg was added after mixing the two component solutions were especially interesting. Delays as short as 1 minute decreased the induction period for ACP2 formation but growth of the succeeding OCP phase took place more slowly. Delays of as little as 3 minutes before addition of Mg reduced the induction period for ACP2 formation to a value almost identical to induction of this phase in the absence of Mg. These data suggest that exposure to Mg ion after conversion to less soluble CaPs will have little effect on their properties.
What, then, are the properties that Mg might confer on fetuin-A sequestered calcium phosphates? Recently Gelli et al. explored the effect of Mg substitution on ACP nanoparticles in systematic experiments in which AMCP particles were synthesized by precipitation of the precursors from aqueous solutions [124]. The particles were analyzed in terms of morphology, crystallinity, and thermal stability. Five different Mg/Ca molar ratios (0.33, 0.5, 1, 1.4, and 2.5), each at three different Mg concentrations (10, 50, and 100 mM), were used. Experiments were carried out at 37 °C and solution pH was adjusted to 7.4 immediately after mixing. They found that at Mg/Ca molar ratios that exceeded 0.5, the synthesized particles were almost completely amorphous. Partially crystalline particles were observed by XRD and FTIR only at lower molar ratios of Mg/Ca. Although the Mg content of the particles increased with increases in the Mg concentration, Mg content in the particles was always lower than that in solution. The difference was most marked in solutions with lower Mg content. Gelli suggested that this reflects the significant differences in solubility of calcium phosphates, a difference which favors precipitation of CaPs.
Since Mg exhibits pleiotropic physiological properties, a significant body of data is available that indicates potential benefit in reducing phosphate toxicity. However, whether the benefit is associated with calciprotein nanoaggregates remains a question.
6. Current Status & Future Perspectives
Despite the significant gains in knowledge about physiological CaP management, important questions remain to be answered, such as the following:
- What is the relationship between changes in the populations of CPMs and CPPs and decreases in renal function? The kidney constitutes the principal organ for absorbing minerals from serum and processing them to recover calcium, magnesium and phosphate in the proximal tubules and/or excreting excesses in the urine [125]. Several groups of investigators have confirmed that CPMs and small CPP-1 nanoparticles are sufficiently small to pass through the renal filtration barrier into the Bowman’s space via the fenestrated endothelium of the glomerular capillaries and the filtration slits of the podocytes. When the kidney is functioning normally, the particles are acid-hydrolyzed into their ionic components, enabling ion recovery or excretion of excesses in urine. It is interesting to note that the concentrations of calcium and phosphate ions within the kidney tubules remain at or above saturation. Deposition of calcium phosphates in the kidney is prevented by adequate concentrations of Mg and citrate ions, as well as sequestration by IDPs such as Tamm-Horsfall protein [125]. However, because current test methods are insufficient to monitor CPMs, few data are available to correlate changes in the populations of CPMs and CPPs with declines in renal function and tissue damage until kidney disease has progressed to organ failure.
- What is the relationship between the sequestered nanoparticles and aberrant deposition, including ectopic deposition in soft tissues? Ectopic calcification is associated with aging as well as chronic diseases such as diabetes, metabolic syndrome, and kidney disease. Each of these chronic conditions is a major cause of blindness, kidney failure, heart attacks, stroke and lower limb amputation, and death. Thus, as populations age, these maladies pose an increasingly heavy economic burden. Therefore, continuing investigations into the relationship between sequestered phosphate nanoparticles and their aberrant deposition in soft tissues are needed to fill knowledge gaps and direct innovation for prevention and treatment.
- What roles do sequestered nanoparticles play in bone remodeling? Despite the observations that skeletal modelling necessarily takes place both in utero and in childhood, and skeletal remodeling continuously repairs defects and supports bone health throughout an individual’s lifetime, detailed mechanisms of bone mineralization remain to be elucidated. Cellular aspects of bone biology are actively being studied and highlight the complex interplay between cells, paracrine and endocrine factors and their balance and communication, as well the actions of cytokines and inflammatory mediators [126,127,128,129]. In general, however, hydroxyapatite formation is described as calcium and phosphate deposition in the holes and pores of collagen fibrils. Classically, deposition of crystalline calcium phosphate or crystalline carboxy calcium phosphate is postulated, wherein the latter gradually loses carbonate content as the crystals mature. In contrast, more recent reports describe deposition of amorphous calcium phosphate, perhaps within collagen matrix vesicles, followed by maturation into crystalline calcium phosphate. Despite the recognition of roles that known IDPs (e.g., osteocalcin, osteonectin, and sclerostin) play in these processes, there is little acknowledgement that these IDPs may act as key sequestration agents, facilitating and regulating maintenance of bone physiology in concordance with the other players.
- Bone dysfunction, particularly the changes in bone resilience associated with osteoporosis, is associated with increased risk and prevalence of fractures. Fractures of hip, spine, vertebrae, wrist or femur are a major cause of morbidity and mortality. The risk of a fracture increases with age and is greatest in women. Looking ahead, the lifetime risk of fractures will increase for all ethnic groups as people live longer. Therefore, increased understanding of the roles that sequestered nanoclusters play in bone remodeling and fracture healing constitutes another area where the knowledge gained through research will significantly impact both prevention and treatment.
- Several years ago, Powell and his colleagues reported extensive constitutive formation of porous amorphous magnesium-substituted calcium phosphate particles with diameters averaging 75-100 nm in both the human and murine gastrointestinal tract [20]. These nanominerals were associated with macromolecules, including bacterial peptidoglycan and dietary proteins. Synthetic analogs of these natural aggregates have been prepared both by Powell and his colleagues and Gelli and her group [100,124]. The apparent roles that calciprotein particles such as these play in immunosurveillance in the gut highlights the need for additional study of the actions of these particles in healthy humans and in those experiencing disruptions in intestinal health.
In summary, current knowledge provides provocative data and supports continuing investigations into the relevance of calciprotein particles in human health and disease.
Author Contributions
Conceptualization, writing—original draft preparation, D.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors acknowledge with thanks editorial review and comments by Dr. Ziad Massy.
Conflicts of Interest
The author is President of BioLink Life Sciences, Inc., a company that is developing mineral salts for use as medical foods and drugs for the treatment of hyperphosphatemia of chronic kidney disease.
| 1 | As a result of differences in material sourcing and differing capabilities for characterization, observers have given these nanoparticles different labels, including fetuin-mineral complexes, nanons, nanobes, bions, calcifying nanoparticles, biomimetic mineralo-protein or mineralo-organic nanoparticles, and nanobacteria. In the discussion that follows, the particles will be identified as calciprotein particles (CPPs). |
References
- Lowenstam HA, Weiner S. Mineralization by organisms and the evolution of biomineralization. In: Biomineralization and Biological Metal Accumulation: Biological and Geological Perspectives, Westbroek P, De Jong EW, Eds., D Reidel Publishing Co., Dordrecht, Holland, 1983.
- Kawasaki K, Weiss KM. Mineralized tissue and vertebrate evolution: the secretory calcium-binding phosphoprotein gene cluster. Proc Natl Acad Sci U S A. 2003 Apr 1; 100(7): 4060-5. [CrossRef]
- Kawasaki K, Suzuki T, Weiss KM. Genetic basis for the evolution of vertebrate mineralized tissue. Proc Natl Acad Sci U S A. 2004 Aug 3; 101(31): 11356-61. [CrossRef]
- Rao A, Drechsler M, Schiller S, Scheffner M, Gebauer D, Cölfen H. Stabilization of mineral precursors by intrinsically disordered proteins. Adv. Funct. Mater. 2018; 28: 1802063. [CrossRef]
- Martel J, Wu CY, Peng HH, Young JD. Mineralo-organic nanoparticles in health and disease: an overview of recent findings. Nanomedicine (Lond). 2018 Jul 1;13(14):1787-1793. [CrossRef] [PubMed]
- Gelli R, Ridi F, Baglioni P. The importance of being amorphous: calcium and magnesium phosphates in the human body. Adv Colloid Interface Sci. 2019 Jul;269:219-235. [CrossRef] [PubMed]
- Smith ER, Hewitson TD, Jahnen-Dechent W. Calciprotein particles: mineral behaving badly? Curr Opin Nephrol Hypertens. 2020 Jul; 29(4): 378-386. [CrossRef] [PubMed]
- Miura Y, Iwazu Y, Shiizaki K, Akimoto T, Kotani K, Kurabayashi M, Kurosu H, Kuro-O M. Identification and quantification of plasma calciprotein particles with distinct physical properties in patients with chronic kidney disease. Sci Rep. 2018 Jan 19; 8(1): 1256. [CrossRef]
- Jahnen-Dechent W, Smith ER. Nature's remedy to phosphate woes: calciprotein particles regulate systemic mineral metabolism. Kidney Int. 2020 Apr; 97(4): 648-651. [CrossRef] [PubMed]
- Wu CY, Martel J, Young JD. Comprehensive organic profiling of biological particles derived from blood. Sci Rep. 2018 Jul 27; 8(1): 11310. [CrossRef]
- Price PA, Nguyen TM, Williamson MK. Biochemical characterization of the serum fetuin-mineral complex. J Biol Chem. 2003 Jun 13; 278(24): 22153-60. [CrossRef] [PubMed]
- Jahnen-Dechent W, Heiss A, Schäfer C, Ketteler M. Fetuin-A regulation of calcified matrix metabolism. Circ Res. 2011 Jun 10; 108(12): 1494-509. [CrossRef]
- Brylka L, Jahnen-Dechent W. The role of fetuin-A in physiological and pathological mineralization. Calcif Tissue Int. 2013 Oct; 93(4): 355-64. [CrossRef]
- Heiss A, Eckert T, Aretz A, Richtering W, van Dorp W, Schäfer C, Jahnen-Dechent W. Hierarchical role of fetuin-A and acidic serum proteins in the formation and stabilization of calcium phosphate particles. J Biol Chem. 2008 May 23; 283(21): 14815-25. [CrossRef] [PubMed]
- Akiyama K, Kimura T, Shiizaki K. Biological and Clinical Effects of Calciprotein Particles on Chronic Kidney Disease-Mineral and Bone Disorder. Int J Endocrinol. 2018 Mar 27; 2018: 5282389. [CrossRef]
- Kutikhin AG, Feenstra L, Kostyunin AE, Yuzhalin AE, Hillebrands JL, Krenning G. Calciprotein Particles: Balancing Mineral Homeostasis and Vascular Pathology. Arterioscler Thromb Vasc Biol. 2021 May 5; 41(5): 1607-1624. [CrossRef]
- Babler A, Schmitz C, Buescher A, Herrmann M, Gremse F, Gorgels T, Floege J, Jahnen-Dechent W. Microvasculopathy and soft tissue calcification in mice are governed by fetuin-A, magnesium and pyrophosphate. PLoS One. 2020 Feb 19; 15(2): e0228938. [CrossRef]
- Schafer C, Heiss A, Schwarz A, Westenfeld R, Ketteler M, Floege J, Muller-Esterl W, Schinke T, Jahnen-Dechent W. The serum protein alpha 2-Heremans-Schmid glycoprotein/fetuin-A is a systemically acting inhibitor of ectopic calcification. J Clin Invest. 2003 Aug; 112(3): 357-66.
- Bäck M, Aranyi T, Cancela ML, Carracedo M, Conceição N, Leftheriotis G, Macrae V, Martin L, Nitschke Y, Pasch A, Quaglino D, Rutsch F, Shanahan C, Sorribas V, Szeri F, Valdivielso P, Vanakker O, Kempf H. Endogenous Calcification Inhibitors in the Prevention of Vascular Calcification: A Consensus Statement From the COST Action EuroSoftCalcNet. Front Cardiovasc Med. 2019 Jan 18; 5: 196. [CrossRef]
- Powell JJ, Thomas-McKay E, Thoree V, Robertson J, Hewitt RE, Skepper JN, Brown A, Hernandez-Garrido JC, Midgley PA, Gomez-Morilla I, Grime GW, Kirkby KJ, Mabbott NA, Donaldson DS, Williams IR, Rios D, Girardin SE, Haas CT, Bruggraber SF, Laman JD, Tanriver Y, Lombardi G, Lechler R, Thompson RP, Pele LC. An endogenous nanomineral chaperones luminal antigen and peptidoglycan to intestinal immune cells. Nat Nanotechnol. 2015 Apr; 10(4): 361-9. [CrossRef]
- Chow, LC. Solubility of calcium phosphates. Monogr Oral Sci. 2001; l13: 94-111.
- Tung MS, Eidelman N, Sieck B, Brown WE. Octacalcium phosphate solubility product from 4 to 37 °C. J Res Nat Bur Stand 1988; 93(5): 613-624.
- Millán Á, Lanzer P, Sorribas V. The Thermodynamics of Medial Vascular Calcification. Front Cell Dev Biol. 2021 Apr 14; 9: 633465. [CrossRef]
- Combes C, Rey C. Amorphous calcium phosphates: synthesis, properties and uses in biomaterials. Acta Biomater. 2010 Sep; 6(9): 3362-78. [CrossRef]
- Edén, M. Structure and formation of amorphous calcium phosphate and its role as surface layer of nanocrystalline apatite: Implications for bone mineralization. Materialia 2021 April; 17(52–54): 101107. [CrossRef]
- Christoffersen MR, Christoffersen J, Kibalczyc W. Apparent solubilities of two amorphous calcium phosphates and of octacalcium phosphate in the temperature range 30-42 °C. J Cryst Growth 1990; 106: 349-354.
- Lenton S, Wang Q, Nylander T, Teixeira S, Holt C. Structural biology of calcium phosphate nanoclusters sequestered by phosphoproteins. Crystals. 2020; 10(9): 755. [CrossRef]
- Boskey AL, Posner AS. Conversion of amorphous calcium phosphate to microcrystalline hydroxyapatite. A pH-dependent, solution-mediated, solid-solid conversion. J Phys Chem 1973; 77: 2313-2317.
- De Yoreo JJ, Gilbert PU, Sommerdijk NA, Penn RL, Whitelam S, Joester D, Zhang H, Rimer JD, Navrotsky A, Banfield JF, Wallace AF, Michel FM, Meldrum FC, Cölfen H, Dove PM. Crystal Growth. Crystallization by particle attachment in synthetic, biogenic, and geologic environments. Science. 2015 Jul 31; 349(6247): aaa6760. [CrossRef] [PubMed]
- Querido W, Shanas N, Bookbinder S, Oliveira-Nunes MC, Krynska B, Pleshko N. Fourier transform infrared spectroscopy of developing bone mineral: from amorphous precursor to mature crystal. Analyst. 2020 Feb 3; 145(3): 764-776. [CrossRef]
- Nývlt, J. The Ostwald Rule of Stages. Cryst. Res. Technol. 1995; 30: 443-449. [CrossRef]
- Levin A, Mason TO, Adler-Abramovich L, Buell AK, Meisl G, Galvagnion C, Bram Y, Stratford SA, Dobson CM, Knowles TP, Gazit E. Ostwald's rule of stages governs structural transitions and morphology of dipeptide supramolecular polymers. Nat Commun. 2014 Nov 13; 5: 5219. [CrossRef] [PubMed]
- Ostwald,W. Studien über die bildung und umwandlung fester körper. Z. Phys. Chem. 1897; 22: 289.
- Threlfall, T. Structural and thermodynamic explanations of Ostwald’s Rule. Org. Process Res. Dev. 2003; 7(6): 1017–1027. [CrossRef]
- Sharma V, Srinivasan A, Nikolajeff F, Kumar S. Biomineralization process in hard tissues: The interaction complexity within protein and inorganic counterparts. Acta Biomater. 2021 Jan 15; 120: 20-37. [CrossRef] [PubMed]
- Yamada H, Kuro-O M, Ishikawa SE, Funazaki S, Kusaka I, Kakei M, Hara K. Daily variability in serum levels of calciprotein particles and their association with mineral metabolism parameters: A cross-sectional pilot study. Nephrology (Carlton). 2018 Mar; 23(3): 226-230. [CrossRef] [PubMed]
- Moreno EC, Gregory TM, Brown WE. Solubility of CaHPO4 · 2 H2O and formation of ion pairs in the system Ca(OH)2 - H3PO4 - H2O at 37.5 °C. J Res Natl Bur Stand A Phys Chem. 1966 Nov-Dec; 70A(6): 545-552. [CrossRef]
- Sun L, Chow LC, Frukhtbeyn SA, Bonevich JE. Preparation and properties of nanoparticles of calcium phosphates with various Ca/P ratios. J Res Natl Inst Stand Technol. 2010 Jul-Aug; 115(4): 243-255. [CrossRef]
- Garcia AC, Vavrusova M, Skibsted LH. Supersaturation of calcium citrate as a mechanism behind enhanced availability of calcium phosphates by presence of citrate. Food Res Int. 2018 May; 107: 195-205. [CrossRef] [PubMed]
- Holt LE, Pierce JA, Kajdi CN. The solubility of the phosphates of strontium, barium, and magnesium and their relation to the problem of calcification. J Colloid Sci 1954; 9(5): 409-426. [CrossRef]
- Lenton S, Nylander T, Teixeira SC, Holt C. A review of the biology of calcium phosphate sequestration with special reference to milk. Dairy Sci Technol. 2015; 95: 3-14. [CrossRef]
- Holt, C, Raynes, JK, Carver, JA. Sequence characteristics responsible for protein-protein interactions in the intrinsically disordered regions of caseins, amelogenins, and small heat-shock proteins. Biopolymers 2019; 110: e23319. [CrossRef]
- Boskey AL, Villarreal-Ramirez E. Intrinsically disordered proteins and biomineralization. Matrix Biol. 2016 May-Jul; 52-54: 43-59. [CrossRef]
- Wright PE, Dyson HJ. Intrinsically disordered proteins in cellular signalling and regulation. Nat Rev Mol Cell Biol. 2015 Jan; 16(1): 18-29. [CrossRef]
- Piovesan D, Tabaro F, Mičetić I, Necci M, Quaglia F, Oldfield CJ, Aspromonte MC, Davey NE, Davidović R, Dosztányi Z, Elofsson A, Gasparini A, Hatos A, Kajava AV, Kalmar L, Leonardi E, Lazar T, Macedo-Ribeiro S, Macossay-Castillo M, Meszaros A, Minervini G, Murvai N, Pujols J, Roche DB, Salladini E, Schad E, Schramm A, Szabo B, Tantos A, Tonello F, Tsirigos KD, Veljković N, Ventura S, Vranken W, Warholm P, Uversky VN, Dunker AK, Longhi S, Tompa P, Tosatto SC. DisProt 7.0: a major update of the database of disordered proteins. Nucleic Acids Res. 2017 Jan 4; 45(D1): D219-D227. [CrossRef]
- Sugase K, Dyson HJ, Wright PE. Mechanism of coupled folding and binding of an intrinsically disordered protein. Nature. 2007 Jun 21; 447(7147): 1021-5. [CrossRef] [PubMed]
- Arai M, Sugase K, Dyson HJ, Wright PE. Conformational propensities of intrinsically disordered proteins influence the mechanism of binding and folding. Proc Natl Acad Sci U S A. 2015 Aug 4; 112(31): 9614-9.
- Shoemaker BA, Portman JJ, Wolynes PG. Speeding molecular recognition by using the folding funnel: the fly-casting mechanism. Proc Natl Acad Sci U S A. 2000 Aug 1; 97(16): 8868-73. [CrossRef]
- Huang LH, Han J, Ouyang JM, Gui BS. Shape-dependent adhesion and endocytosis of hydroxyapatite nanoparticles on A7R5 aortic smooth muscle cells. J Cell Physiol. 2020 Jan; 235(1): 465-479. [CrossRef] [PubMed]
- Huang LH, Sun XY, Ouyang JM. Shape-dependent toxicity and mineralization of hydroxyapatite nanoparticles in A7R5 aortic smooth muscle cells. Sci Rep. 2019 Dec 12; 9(1): 18979. [CrossRef]
- Cai MM, Smith ER, Holt SG. The role of fetuin-A in mineral trafficking and deposition. Bonekey Rep. 2015 May 6; 4: 672. [CrossRef]
- de Kruif CG, Huppertz T, Urban VS, Petukhov AV. Casein micelles and their internal structure. Adv Colloid Interface Sci. 2012 Mar-Apr; 171-172: 36-52. [CrossRef] [PubMed]
- Holt, C. The Milk Salts and Their Interaction with Casein. In Advanced Dairy Chemistry; Fox PF, Ed.; Chapman and Hall: London, UK, 1997; pp. 233–254. [Google Scholar]
- Ingham B, Erlangga GD, Smialowska A, Kirby NM, Wang C, Matia-Merino L, Haverkamp RG, Carr AJ. Solving the mystery of the internal structure of casein micelles. Soft Matter. 2015 Apr 14; 11(14): 2723-5. [CrossRef] [PubMed]
- Kamigaki T, Ito Y, Nishino Y, Miyazawa A. Microstructural observation of casein micelles in milk by cryo-electron microscopy of vitreous sections (CEMOVIS). Microscopy (Oxf). 2018 Jun 1; 67(3): 164-170. [CrossRef] [PubMed]
- Hettiarachchi CA, Swulius MT, Harte FM. Assessing constituent volumes and morphology of bovine casein micelles using cryo-electron tomography. J Dairy Sci. 2020 May; 103(5): 3971-3979. [CrossRef] [PubMed]
- McMahon DJ, Oommen BS. Supramolecular structure of the casein micelle. J Dairy Sci. 2008 May; 91(5): 1709-21. [CrossRef] [PubMed]
- Jenness, R.; Sloan, R.E. The Composition of Milk of Various Species: A Review. Dairy Sci. Abstr. 1970, 32, 599–612. [Google Scholar]
- Eisert, R.; Oftedal, O.T. Capital expenditure and income (foraging) during pinniped lactation: The example of the weddell seal (Leptonychotes weddellii). In Smithsonian at the Poles: Contributions to International Polar Year Science; Lang, M.A., Miller, S.E., Krupnik, I., Eds.; Smithsonian Institute: Washington, DC, USA, 2009; pp. 335–346. [Google Scholar]
- Jenness R, Holt C. Casein and lactose concentrations in milk of 31 species are negatively correlated. Experientia. 1987 Sep 15; 43(9): 1015-8. [CrossRef] [PubMed]
- Aoki T, Yamada N, Tomita I, Kako Y, Imamura T. Caseins are cross-linked through their ester phosphate groups by colloidal calcium phosphate. Biochim Biophys Acta. 1987 Jan 30; 911(2): 238-43. [CrossRef] [PubMed]
- Malacarne M, Criscione A, Franceschi P, Bordonaro S, Formaggioni P, Marletta D, Summer A. New Insights into Chemical and Mineral Composition of Donkey Milk throughout Nine Months of Lactation. Animals (Basel). 2019 Dec 17; 9(12): 1161. [CrossRef]
- Fantuz F, Ferraro S, Todini L, Cimarelli L, Fatica A, Marcantoni F, Salimei E. Distribution of calcium, phosphorus, sulfur, magnesium, potassium, and sodium in major fractions of donkey milk. J Dairy Sci. 2020 Oct; 103(10): 8741-8749. [CrossRef] [PubMed]
- Fox, P.F. , Uniacke-Lowe, T., McSweeney, P.L.H., O'Mahony, J.A. Dairy Chemistry and Biochemistry. 2nd Ed., 2015. Springer International Publishing Switzerland. Chem, Switzerland.
- Sánchez C, Fente C, Barreiro R, López-Racamonde O, Cepeda A, Regal P. Association between Breast Milk Mineral Content and Maternal Adherence to Healthy Dietary Patterns in Spain: A Transversal Study. Foods. 2020 May 20; 9(5): 659. [CrossRef]
- Li C, Solomons NW, Scott ME, Koski KG. Minerals and Trace Elements in Human Breast Milk Are Associated with Guatemalan Infant Anthropometric Outcomes within the First 6 Months. J Nutr. 2016 Oct; 146(10): 2067-2074. [CrossRef] [PubMed]
- Dalgleish DG, Parker TG. Binding of calcium ions to bovine alpha-S1-casein and precipitability of the protein-calcium ion complexes. J. Dairy Res. 1980; 47(1): 113–122. [CrossRef]
- Parker TG, Dalgleish DG. Binding of calcium ions to bovine beta-casein. J Dairy Res. 1981 Feb; 48(1): 71-6. [CrossRef] [PubMed]
- Sleigh RW, Mackinlay AG, Pope JM. NMR studies of the phosphoserine regions of bovine alpha s1- and beta-casein. Assignment of 31P resonances to specific phosphoserines and cation binding studied by measurement of enhancement of 1H relaxation rate. Biochim Biophys Acta. 1983 Jan 12; 742(1): 175-83. [CrossRef] [PubMed]
- Sleigh RW, Sculley TB, Mackinlay AG. The binding of beta-casein to hydroxyapatite: the effect of phosphate content and location. J Dairy Res. 1979 Apr; 46(2): 337-42. [CrossRef] [PubMed]
- Bijl E, Huppertz T, van Valenberg H, Holt C. A quantitative model of the bovine casein micelle: ion equilibria and calcium phosphate sequestration by individual caseins in bovine milk. Eur Biophys J. 2019 Jan; 48(1): 45-59. [CrossRef] [PubMed]
- Holt C, Sawyer L. Caseins as rheomorphic proteins - Interpretation of primary and secondary structures of the alpha-S1-caseins, beta-caseins and kappa-caseins. J Chem Soc. Faraday Trans. 1993; 89: 2683–2692. [CrossRef]
- Heiss A, DuChesne A, Denecke B, Grötzinger J, Yamamoto K, Renné T, Jahnen-Dechent W. Structural basis of calcification inhibition by alpha 2-HS glycoprotein/fetuin-A. Formation of colloidal calciprotein particles. J Biol Chem. 2003 Apr 11; 278(15): 13333-41. [CrossRef] [PubMed]
- Häusler M, Schäfer C, Osterwinter C, Jahnen-Dechent W. The physiologic development of fetuin-a serum concentrations in children. Pediatr Res. 2009 Dec; 66(6): 660-4. [CrossRef] [PubMed]
- Šimják P, Cinkajzlová A, Anderlová K, Kloučková J, Kratochvílová H, Lacinová Z, Kaválková P, Krejčí H, Mráz M, Pařízek A, Kršek M, Haluzík M. Changes in plasma concentrations and mRNA expression of hepatokines fetuin A, fetuin B and FGF21 in physiological pregnancy and gestational diabetes mellitus. Physiol Res. 2018 Nov 28; 67(Suppl 3): S531-S542. [CrossRef] [PubMed]
- Ren G, Kim T, Papizan JB, Okerberg CK, Kothari VM, Zaid H, Bilan PJ, Araya-Ramirez F, Littlefield LA, Bowers RL, Mahurin AJ, Nickles MM, Ludvigsen R, He X, Grandjean PW, Mathews ST. Phosphorylation status of fetuin-A is critical for inhibition of insulin action and is correlated with obesity and insulin resistance. Am J Physiol Endocrinol Metab. 2019 Aug 1; 317(2): E250-E260. [CrossRef] [PubMed]
- Jahnen-Dechent W, Büscher A, Köppert S, Heiss A, Kuro-O M, Smith ER. Mud in the blood: the role of protein-mineral complexes and extracellular vesicles in biomineralisation and calcification. J Struct Biol. 2020 Oct 1; 212(1): 107577. [CrossRef] [PubMed]
- Gebauer D, Kellermeier M, Gale JD, Bergström L, Cölfen H. Pre-nucleation clusters as solute precursors in crystallisation. Chem Soc Rev. 2014 Apr 7; 43(7): 2348-71. [CrossRef] [PubMed]
- Köppert S, Büscher A, Babler A, Ghallab A, Buhl EM, Latz E, Hengstler JG, Smith ER, Jahnen-Dechent W. Cellular Clearance and Biological Activity of Calciprotein Particles Depend on Their Maturation State and Crystallinity. Front Immunol. 2018 Sep 4; 9: 1991. [CrossRef]
- Koeppert S, Ghallab A, Peglow S, Winkler CF, Graeber S, Büscher A, Hengstler JG, Jahnen-Dechent W. Live Imaging of Calciprotein Particle Clearance and Receptor Mediated Uptake: Role of Calciprotein Monomers. Front Cell Dev Biol. 2021 Apr 29;9:633925. [CrossRef]
- Satchell SC, Braet F. Glomerular endothelial cell fenestrations: an integral component of the glomerular filtration barrier. Am J Physiol Renal Physiol. 2009 May; 296(5): F947-56.
- Herrmann M, Schäfer C, Heiss A, Gräber S, Kinkeldey A, Büscher A, Schmitt MM, Bornemann J, Nimmerjahn F, Herrmann M, Helming L, Gordon S, Jahnen-Dechent W. Clearance of fetuin-A-containing calciprotein particles is mediated by scavenger receptor-A. Circ Res. 2012 Aug 17; 111(5): 575-84. [CrossRef] [PubMed]
- Wu CY, Martel J, Young JD. Ectopic calcification and formation of mineralo-organic particles in arteries of diabetic subjects. Sci Rep. 2020 May 22; 10(1): 8545. [CrossRef]
- Kumon H, Matsuura E, Nagaoka N, Yamamoto T, Uehara S, Araki M, Matsunami Y, Kobayashi K, Matsumoto A. Ectopic calcification: importance of common nanoparticle scaffolds containing oxidized acidic lipids. Nanomedicine. 2014 Feb; 10(2): 441-50. [CrossRef] [PubMed]
- Perovic I, Chang EP, Lui M, Rao A, Cölfen H, Evans JS. A nacre protein, n16.3, self-assembles to form protein oligomers that dimensionally limit and organize mineral deposits. Biochemistry. 2014 Apr 29;53(16):2739-48. [CrossRef] [PubMed]
- Szasz CS, Alexa A, Toth K, Rakacs M, Langowski J, Tompa P. Protein disorder prevails under crowded conditions. Biochemistry. 2011 Jul 5; 50(26): 5834-44. [CrossRef] [PubMed]
- Stawski TM, Roncal-Herrero T, Fernandez-Martinez A, Matamoros-Veloza A, Kröger R, Benning LG. "On demand" triggered crystallization of CaCO3 from solute precursor species stabilized by the water-in-oil microemulsion. Phys Chem Chem Phys. 2018 May 23; 20(20): 13825-13835. [CrossRef] [PubMed]
- Jain G, Pendola M, Rao A, Cölfen H, Evans JS. A Model Sea Urchin Spicule Matrix Protein Self-Associates To Form Mineral-Modifying Protein Hydrogels. Biochemistry. 2016 Aug 9; 55(31): 4410-21. [CrossRef] [PubMed]
- Perovic I, Davidyants A, Evans JS. Aragonite-Associated Mollusk Shell Protein Aggregates To Form Mesoscale "Smart" Hydrogels. ACS Omega. 2016 Nov 30; 1(5): 886-893. [CrossRef]
- Pendola M, Jain G, Evans JS. Skeletal development in the sea urchin relies upon protein families that contain intrinsic disorder, aggregation-prone, and conserved globular interactive domains. PLoS One. 2019 Oct 1; 14(10): e0222068. [CrossRef]
- Hołubowicz R, Wojtas M, Taube M, Kozak M, Ożyhar A, Dobryszycki P. Effect of calcium ions on structure and stability of the C1q-like domain of otolin-1 from human and zebrafish. FEBS J. 2017 Dec; 284(24): 4278-4297. [CrossRef] [PubMed]
- George A, Veis A. Phosphorylated proteins and control over apatite nucleation, crystal growth, and inhibition. Chem Rev. 2008 Nov; 108(11): 4670-93. [CrossRef]
- Farvadi F, Ghahremani MH, Hashemi F, Reza Hormozi-Nezhad M, Raoufi M, Zanganeh S, Atyabi F, Dinarvand R, Mahmoudi M. Cell shape affects nanoparticle uptake and toxicity: An overlooked factor at the nanobio interfaces. J Colloid Interface Sci. 2018 Dec 1; 531: 245-252. [CrossRef] [PubMed]
- Moreland KT, Hong M, Lu W, Rowley CW, Ornitz DM, De Yoreo JJ, Thalmann R. In vitro calcite crystal morphology is modulated by otoconial proteins otolin-1 and otoconin-90. PLoS One. 2014 Apr 18; 9(4): e95333. [CrossRef]
- Poundarik AA, Boskey A, Gundberg C, Vashishth D. Biomolecular regulation, composition and nanoarchitecture of bone mineral. Sci Rep. 2018 Jan 19; 8(1): 1191. [CrossRef]
- Kunishige R, Mizoguchi M, Tsubouchi A, Hanaoka K, Miura Y, Kurosu H, Urano Y, Kuro-O M, Murata M. Calciprotein particle-induced cytotoxicity via lysosomal dysfunction and altered cholesterol distribution in renal epithelial HK-2 cells. Sci Rep. 2020 Nov 18; 10(1): 20125. [CrossRef]
- Ewence AE, Bootman M, Roderick HL, Skepper JN, McCarthy G, Epple M, Neumann M, Shanahan CM, Proudfoot D. Calcium phosphate crystals induce cell death in human vascular smooth muscle cells: a potential mechanism in atherosclerotic plaque destabilization. Circ Res. 2008 Aug 29; 103(5): e28-34. [CrossRef] [PubMed]
- Herrmann M, Babler A, Moshkova I, Gremse F, Kiessling F, Kusebauch U, Nelea V, Kramann R, Moritz RL, McKee MD, Jahnen-Dechent W. Lumenal calcification and microvasculopathy in fetuin-A-deficient mice lead to multiple organ morbidity. PLoS One. 2020 Feb 19; 15(2): e0228503. [CrossRef]
- Shook LL, Buhimschi CS, Dulay AT, McCarthy ME, Hardy JT, Duzyj Buniak CM, Zhao G, Buhimschi IA. Calciprotein particles as potential etiologic agents of idiopathic preterm birth. Sci Transl Med. 2016 Nov 9;8(364):364ra154. [CrossRef] [PubMed]
- Pele LC, Haas CT, Hewitt RE, Robertson J, Skepper J, Brown A, Hernandez-Garrido JC, Midgley PA, Faria N, Chappell H, Powell JJ. Synthetic mimetics of the endogenous gastrointestinal nanomineral: Silent constructs that trap macromolecules for intracellular delivery. Nanomedicine. 2017 Feb; 13(2): 619-630. [CrossRef]
- Gelli R, Tempesti P, Ridi F, Baglioni P. Formation and properties of amorphous magnesium-calcium phosphate particles in a simulated intestinal fluid. J Colloid Interface Sci. 2019 Jun 15;546:130-138. [CrossRef] [PubMed]
- Gelli R, Martini F, Geppi M, Borsacchi S, Ridi F, Baglioni P. Exploring the interplay of mucin with biologically-relevant amorphous magnesium-calcium phosphate nanoparticles. J Colloid Interface Sci. 2021 Mar 16; 594: 802-811. [CrossRef] [PubMed]
- Akiyama KI, Miura Y, Hayashi H, Sakata A, Matsumura Y, Kojima M, Tsuchiya K, Nitta K, Shiizaki K, Kurosu H, Kuro-O M. Calciprotein particles regulate fibroblast growth factor-23 expression in osteoblasts. Kidney Int. 2020 Apr; 97(4): 702-712. [CrossRef] [PubMed]
- Kuro-O, M. Klotho and endocrine fibroblast growth factors: markers of chronic kidney disease progression and cardiovascular complications? Nephrol Dial Transplant. 2019 Jan 1; 34(1): 15-21. [CrossRef] [PubMed]
- Kuro-O, M. Aging and FGF23-klotho system. Vitam Horm. 2021; 115: 317-332. [CrossRef] [PubMed]
- Price PA, Thomas GR, Pardini AW, Figueira WF, Caputo JM, Williamson MK. Discovery of a high molecular weight complex of calcium, phosphate, fetuin, and matrix gamma-carboxyglutamic acid protein in the serum of etidronate-treated rats. J Biol Chem. 2002 Feb 8; 277(6): 3926-34. [CrossRef] [PubMed]
- Wong TY, Wu CY, Martel J, Lin CW, Hsu FY, Ojcius DM, Lin PY, Young JD. Detection and characterization of mineralo-organic nanoparticles in human kidneys. Sci Rep. 2015 Oct 26; 5: 15272. [CrossRef]
- Smith ER, Hewitson TD, Hanssen E, Holt SG. Biochemical transformation of calciprotein particles in uraemia. Bone. 2018 May; 110: 355-367. [CrossRef] [PubMed]
- Aghagolzadeh P, Bachtler M, Bijarnia R, Jackson C, Smith ER, Odermatt A, Radpour R, Pasch A. Calcification of vascular smooth muscle cells is induced by secondary calciprotein particles and enhanced by tumor necrosis factor-α. Atherosclerosis. 2016 Aug; 251: 404-414. [CrossRef] [PubMed]
- Swanson KV, Deng M, Ting JP. The NLRP3 inflammasome: molecular activation and regulation to therapeutics. Nat Rev Immunol. 2019 Aug; 19(8): 477-489. [CrossRef]
- Honarpisheh M, Foresto-Neto O, Desai J, Steiger S, Gómez LA, Popper B, Boor P, Anders HJ, Mulay SR. Phagocytosis of environmental or metabolic crystalline particles induces cytotoxicity by triggering necroptosis across a broad range of particle size and shape. Sci Rep. 2017 Nov 14;7(1):15523. [CrossRef]
- Mulay SR, Honarpisheh MM, Foresto-Neto O, Shi C, Desai J, Zhao ZB, Marschner JA, Popper B, Buhl EM, Boor P, Linkermann A, Liapis H, Bilyy R, Herrmann M, Romagnani P, Belevich I, Jokitalo E, Becker JU, Anders HJ. Mitochondria Permeability Transition versus Necroptosis in Oxalate-Induced AKI. J Am Soc Nephrol. 2019 Oct; 30(10): 1857-1869. [CrossRef]
- Dautova Y, Kozlova D, Skepper JN, Epple M, Bootman MD, Proudfoot D. Fetuin-A and albumin alter cytotoxic effects of calcium phosphate nanoparticles on human vascular smooth muscle cells. PLoS One. 2014 May 21; 9(5): e97565. [CrossRef]
- Dai XY, Zhao MM, Cai Y, Guan QC, Zhao Y, Guan Y, Kong W, Zhu WG, Xu MJ, Wang X. Phosphate-induced autophagy counteracts vascular calcification by reducing matrix vesicle release. Kidney Int. 2013 Jun; 83(6): 1042-51. [CrossRef] [PubMed]
- Wåhlén E, Olsson F, Söderberg O, Lennartsson J, Heldin J. Differential impact of lipid raft depletion on platelet-derived growth factor (PDGF)-induced ERK1/2 MAP-kinase, SRC 22and AKT signaling. Cell Signal. 2022 Aug; 96: 110356. Erratum in: Cell Signal. 2022 Jul 22;98:110411. [CrossRef] [PubMed]
- Brown, RB. Diabetes, Diabetic Complications, and Phosphate Toxicity: A Scoping Review. Curr Diabetes Rev. 2020; 16(7): 674-689. [CrossRef] [PubMed]
- Merx MW, Schäfer C, Westenfeld R, Brandenburg V, Hidajat S, Weber C, Ketteler M, Jahnen-Dechent W. Myocardial stiffness, cardiac remodeling, and diastolic dysfunction in calcification-prone fetuin-A-deficient mice. J Am Soc Nephrol. 2005 Nov; 16(11): 3357-64. [PubMed]
- Villa-Bellosta R, O'Neill WC. Pyrophosphate deficiency in vascular calcification. Kidney Int. 2018 Jun; 93(6): 1293-1297. [CrossRef] [PubMed]
- Villa-Bellosta R, Sorribas V. Prevention of vascular calcification by polyphosphates and nucleotides- role of ATP. Circ J. 2013; 77(8): 2145-51. [CrossRef] [PubMed]
- Icer MA, Yıldıran H. Effects of fetuin-A with diverse functions and multiple mechanisms on human health. Clin Biochem. 2021 Feb;88:1-10. [CrossRef] [PubMed]
- Boskey AL, Posner AS. Magnesium stabilization of amorphous calcium phosphate: A kinetic study. Mat Res Bull 1974; 9: 907-916.
- Holt C, Van Kemenade MJJM, Harries JE, Nelson LS, Bailey RT, Hukins DWL, Hasnain SS, De Bruyn PL. Preparation of amorphous calcium magnesium phosphates at pH 7 and characterization by X-ray absorption and Fourier transform infrared spectroscopy. J Cryst Growth 1988; 92: 239-252.
- Kibalczyc W, Christoffersen J, Christoffersen MR, Zielenkiewicz A, Zielenkiewicz W. The effect of magnesium ions on the precipitation of calcium phosphates. J Cryst Growth 1990; 106: 355-366.
- Gelli R, Scudero M, Gigli L, Severi M, Bonini M, Ridi F, Baglioni P. Effect of pH and Mg2+ on Amorphous Magnesium-Calcium Phosphate (AMCP) stability. J Colloid Interface Sci. 2018 Dec 1; 531: 681-692. [CrossRef] [PubMed]
- Rodelo-Haad C, Pendón-Ruiz de Mier MV, Díaz-Tocados JM, Martin-Malo A, Santamaria R, Muñoz-Castañeda JR, Rodríguez M. The Role of Disturbed Mg Homeostasis in Chronic Kidney Disease Comorbidities. Front Cell Dev Biol. 2020 Nov 12; 8: 543099. [CrossRef]
- Diaz-Tocados JM, Peralta-Ramirez A, Rodríguez-Ortiz ME, Raya AI, Lopez I, Pineda C, Herencia C, Montes de Oca A, Vergara N, Steppan S, Pendon-Ruiz de Mier MV, Buendía P, Carmona A, Carracedo J, Alcalá-Díaz JF, Frazao J, Martínez-Moreno JM, Canalejo A, Felsenfeld A, Rodriguez M, Aguilera-Tejero E, Almadén Y, Muñoz-Castañeda JR. Dietary magnesium supplementation prevents and reverses vascular and soft tissue calcifications in uremic rats. Kidney Int. 2017 Nov;92(5):1084-1099. [CrossRef] [PubMed]
- Ter Braake AD, Smit AE, Bos C, van Herwaarden AE, Alkema W, van Essen HW, Bravenboer N, Vervloet MG, Hoenderop JGJ, de Baaij JHF. Magnesium prevents vascular calcification in Klotho deficiency. Kidney Int. 2020 Mar; 97(3): 487-501. [CrossRef] [PubMed]
- ter Braake AD, Eelderink C, Zeper LW, Pasch A, Bakker SJL, de Borst MH, Hoenderop JGJ, de Baaij JHF. Calciprotein particle inhibition explains magnesium-mediated protection against vascular calcification. Nephrol Dial Transplant. 2020 May 1;35(5):765-773. [CrossRef]
Figure 1.
A graphic presentation of the mechanism of formation and properties associated with calciprotein particles.
Figure 1.
A graphic presentation of the mechanism of formation and properties associated with calciprotein particles.
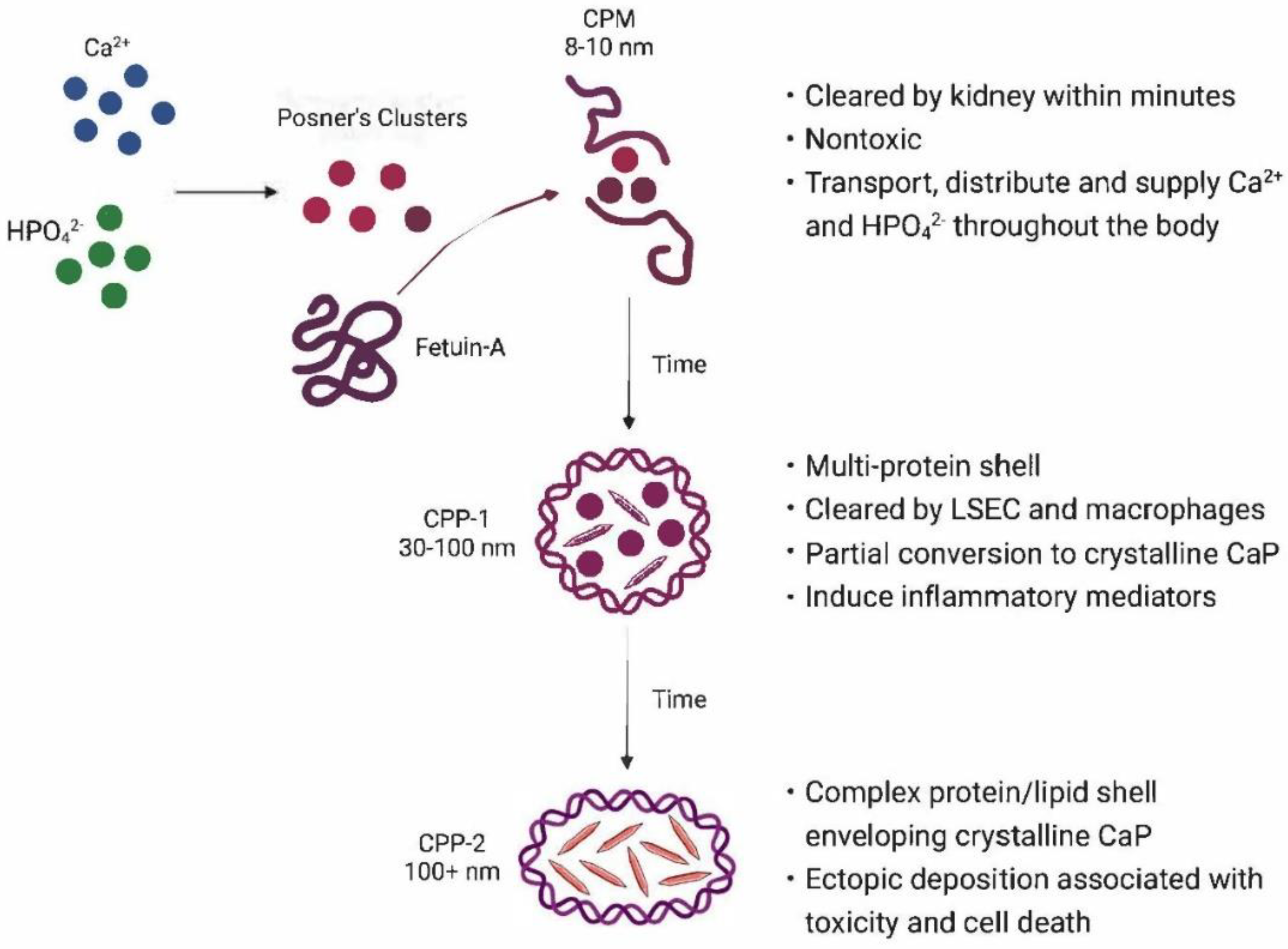
Table 1.
Solubilities of representative calcium phosphates.
| Salt | Formula | Ca/P | -log Ksp) at 25°C1 | -log (Ksp) at 37°C2 | pH |
|---|---|---|---|---|---|
| Monocalcium phosphate monohydrate | Ca(H2PO4)2 ∙ H2O | 0.5 | Highly soluble | Soluble | < 2 |
| Dicalcium phosphate dihydrate | Ca2(HPO4)2 ∙ 2 H2O | 1.0 | 6.59 | 6.66 | 3.5 – 6.8 |
| Octacalcium phosphate | Ca8H2(PO4)6 ∙ 5 H2O | 1.33 | 48.4 | 48.7 | ~ 6 |
| α-Tricalcium phosphate | α-Ca3(PO4)2 | 1.5 | 25.5 | ||
| β-Tricalcium phosphate | β-Ca3(PO4)2 | 1.5 | 28.9 | ||
| Hydroxyapatite | Ca10(PO4)6(OH)2 | 1.67 | 58.4 | 117.3 | 9.5 – 12 |
| Fluoroapatite | Ca5(PO4)3F | 1.67 | 60.5 | ||
| Tetracalcium phosphate | Ca4(PO4)2O | 2.0 | 38.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



