Submitted:
07 June 2023
Posted:
07 June 2023
You are already at the latest version
Abstract
The preservative qualities of individual ionic compounds on the infectivity of T4 virions were elucidated. T4 virions were immersed in quasi-pure ionic solutions prior to the adsorption process, and the pfu values of these were measured following the conventional method. In neutral ionic solutions, the minimum and the optimum concentrations for preservative qualities were corresponded with the results obtained from the multi-ionic media/buffers. In acid and alkali solutions, phages show tolerances at a pH range of 5-11 in multi-ionic media/buffers. T4 virions show no tolerance in quasi-pure acid, neutral and weak alkaline conditions. The preservative quality of T4 virions was jumped up in over 10-1 mM OH- solution, equivalent to pH value over 10, which is corresponding to the pKa of the deprotonation of the DNA bases G and T. Infectivity was lost below 10-1 mM OH- and higher than 10 mM OH-. These results implicate the maintaining of infectivity of a virion may need the flexibility of the intra-capsid DNA by deprotonation.
Keywords:
infectivity
; neutral solution
; acidic solution
; alkaline solution
; deprotonation
1. Introduction
Understanding the responses of virions to ambient biological, chemical, and physical factors is critical not only for basic biological viewpoints, but also for defining optimum and/or most harmful conditions for the industrial, medical, and agricultural use of virions [1,2]. Among these factors, chemism has been studied for a long time [3,4,5,6,7,8]. It is generally accepted that divalent cations, such as Ca2+ and Mg2+, affect the lytic cycle of many bacteriophages, acting on either adsorption, penetration, or intracellular development of the virus [2,6,9]. The adsorption on bacterial cells of some phages has been also accelerated by univalent cations, such as Na+ [4,8]. Phages can generally show infection capabilities at a pH range of 5-11 [1,10,11]. The reported results, however, were obtained in solutions of mixed ionic compounds, e.g., media and buffers. Prior to using the multi-ionic conditions for studying the chemical processes of the lytic cycle of bacteriophages, viral responses to individual ion solutions should be elucidated.
To understand the effects of sole-ionic compound solutions on the infectivity of T4 virions, T4 virions need to be immersed in pure ionic solutions. However, the inoculant of virions contains certain number of ionic solutes, because virions will be inactivated immediately in pure water [2, 4, 8, this study]. Therefore, we control the amount of the solutes in the inoculant at the minimum level. This is the reason why we call the sole-ionic compound solutions as quasi-pure.
The pfu spectra indicates the viral infectivity is irreversibly changed by soaking the virions in quasi-pure ionic solutions prior to the adsorption process. Our results generally followed the trends in previous reports in neutral conditions [2,6], e.g., the optimal divalent cation levels ranged between 0.01 and 10 mM, and one order higher for monovalent cations. One of the major findings of our study was the effect of H+ and OH- on phage stability. When T4 virions were directly exposed to acid or alkali solutions, T4 virions were stable in over 10-1 mM OH- solution, equivalent to alkaline pH 10, although infectivity was lost below 10-1 mM OH- and higher than 10 mM OH-. No infectious activity was observed in acid to neutral and even in weak alkaline solutions. Hydrogen ion, H+, clearly shows negative effect on phage stability, while hydroxide ion, OH-, stabilized phages at around 10-1 - 10 mM OH-.
The activation-inactivation is switched on/off at 10-4 N OH-, pH 10 equivalent, which is corresponding to the pKa of the deprotonation of the DNA bases G and T. The results implicate the maintaining of infectivity of a virion may depend on the flexibility of the intra-capsid DNA by deprotonation.
2. Materials and Methods
Strains:
The bacteriophage studied was T4 (ATCC 11303-B4), and its host bacteria were Escherichia coli (ATCC 11303).
Preparation of T4 virions:
Peptone broth was used for culturing the host bacteria, E. coli. T4 suspension was obtained by the plate lysate method and the small-scale liquid culture [12]. In the plate lysate method, the virions were suspended in 2-3 ml of an electrolyte solution including 1.8 mM NaCl, 0.12 mM MgSO4, 0.12 mM MgCl2, 0.034 mM CaCl2 and 0.05 mM KCl. The extracted suspension was sterilized by filtration of the eluent with 0.2 μm filter (Advantec AS020). To exclude the effects of anonymous ions, the suspensions of T4 virions were purified with ultracentrifugation and dialysis. For ultracentrifugation, crude bacteriophage particles were purified by isopycnic centrifugation through CsCl gradients [12, Beckman XPN-90, SW32 rotor, 4℃, 24 h]. Following ultracentrifugation, T4 suspensions were dialysed against T-buffer (modified TM buffer [12] 0.1 M NaCl, 2 mM MgSO4, 0.5 mM phosphate buffer, pH 7.5) for one week replacing the outside T-buffer five times. The chemicals used were special grade products from Wako Pure Chemical Ind. Ltd.
Preparation of suspension of T4 virions in quasi-pure solution:
It is well known that the infectious ability of virions is instantly lost when they are immersed in pure water [2, 4, 8, this study], with the result that the active virions are practically stored in relevant ionic solutions. When the stored virions are used for experiments, aliquots of stored suspensions are inoculated in target solvents. Accordingly, the ions included in the aliquots of the suspensions are added into the target solutions. It is critical to maintain the concentrations of these ions introduced by the inoculations for studying the connections between the ionic concentrations and the viral activity. In these experiments, the ultra-centrifuged T4 virions were dialysed against T-buffer and stored in T-buffer. After the stored virions were appropriately diluted with T-buffer, these T-buffer suspensions were diluted twice. First, dilution with Milli-Q water 100-fold and acclimated to this condition for five minutes. At this point, the ionic concentrations of the viral suspensions were 0.01 T-buffer. Second, this Milli-Q water dilutions were diluted again 100-fold with the test solutions. As a whole, the final concentrations of T-buffer in the test solutions were 0.0001 T-buffer, i.e., 0.01 mM NaCl, 0.2 μM MgSO4, 0.05 μM phosphate buffer. In these diluted ionic conditions, virions gradually lost their infectious ability by the time of their manipulation. The time course measurements of viral activities in these diluted conditions indicating the decrease of the activity were expressed as:
Vt=V0 e-kt,
Where Vt: density of active virions at t, V0: density of active virions at the initial moment, -k: inactivation coefficient, t: time.
For counting plaque forming units, virions were plated after the acclimation at the first dilution for 5min. and followed by incubation in the second dilution for 15 min.
Ionic solutions:
In this paper, we are concerned with the concentration of a solute. To prepare the acid and alkali solutions of the testing concentrations, 1 M solutions of acid and alkali were diluted to the target concentrations. According to the solubilities, the original solution of Ca(OH)2 was 10 mM and it was 0.15 mM for Mg(OH)2. The pH value of the dilution was not measured each time, and pH was not adjusted. However, the pH values measured by Whatman pH-indicator paper were close to the expected values, e.g., the pH value of 1 mM NaOH was ca. pH 11. The pH values reported here, nominal pH, were not the measured values, but the calculated values from concentrations of H+ and OH-. On the other hand, the pH values of the solutions of neutral ionic compounds, i.e., NaCl, Na2SO4, KCl, K2SO4, CaCl2, CaSO4, MgCl2, MgSO4, fell within the range of pH 6 – 6.5.
Plaque forming unit (pfu):
Plaque forming units (pfu) of each case were measured twice independently using two populations of T4, i.e., adjacent two fractions of ultra-centrifuged samples, which were expressed as thicker lines (the heavier fraction) and thinner lines (the lighter fraction). Aliquots of virions were plated with E. coli on 1% agar peptone plates covered with 0.5% agar peptone top agar. Inoculated plates were incubated at 36℃ for 12 h before counting. The numbers of plaques were adjusted to 10 ~ 500 plaques per plate as was possible. The pfu values indicated in figures were % of the pfu values of T4 populations suspended in T-buffer. The pfu values of the stock T4 suspensions were enumerated frequently to confirm the activity of the T4 virion during the experiments.
3. Results
3.1. Time course of viral activity
Table 1 indicates the values of -k and R2 of formula (1). At selected dilutions of T4 virions in T-buffer, in the first dilution in Milli-Q, when the ion concentration was 0.01 T-buffer, more than 95% of virions were active after 5 min. When the 0.01 T-buffer was diluted into Milli-Q again, the ionic composition was 0.0001 T-buffer. Virions in this condition lost the infectious ability instantly, which indicates that the ionic composition in 0.0001 T-buffer, i.e., 0.01 mM NaCl, 0.2 μM MgSO4, 0.05 μM phosphate buffer, does not support the infectious ability of T4. The testing ionic compound was included in the solution of the second dilution, where even if the remaining T-buffer exists, the remnant of T-buffer does not disturb the infectious process in the testing ionic solution. Therefore, we call this condition “quasi-pure” and can practically measure the action of a sole additional compound on the infectious process.
The incubation period of the test suspension was chosen as 15 min., because the survivals of virions in the test solutions of preferable ion concentrations were higher than 60% after 15 min. (see below), and the differences of the rates of survival between different ion concentrations became sufficiently large.
Survivals of T4 virions in selected sole alkaline solutions were examined in extended time courses. Virions in 3 mM NaOH, in 1 mM Ca(OH)2 and in 0.1 mM Mg(OH)2 solutions maintained 6%, 58% and 98% of infectivity after one week, respectively.
3.2. H+: HCl and H2SO4
The stabilities of T4 in acid solutions, HCl and H2SO4, were measured at the range between 10-2 mM and 103 mM, corresponding to pH 5 and pH 0 (Figure 1). No pfu were observed at suspensions in whole ranges of concentrations of both acids. As outlined below, when the cations are Na+, K+, Ca2+ or Mg2+, the anions Cl- and SO42- do not cause inhibition of pfu formation. It becomes clear the univalent cation, hydrogen ion, H+, inactivates the infectious ability of T4 in the whole spectrum of the concentration. Using the heavier fraction of T4 virions, effects of coexistent ions were examined. Coexistence with other neutral ions, e.g., 0.1 M NaCl or 10 mM CaCl2, protects virions from the inactivation by H+ at the dilute ranges of H+, e.g., lower than 10-4 N or higher than nominal pH 4 (Figure 1, dashed lines).
3.3. Na+: NaOH, NaCl, and Na2SO4
The effects of sodium ion, Na+, in the quasi-pure ionic solutions of NaOH, NaCl, and Na2SO4 were tested (Figure 2). In the alkaline solution of NaOH, ca. 90% of viral activity was maintained at ion levels ranging between 0.1 and 1 mM after 15 min. suspension. No or few pfu were observed below 0.1 mM NaOH (pH 10 equivalent) or higher than 10 mM NaOH (pH 12 equivalent). The pfu values abruptly jumped to 0.1 mM NaOH and over.
In the neutral ionic solutions of NaCl and Na2SO4, the optimal ion levels were 100 mM for NaCl and higher than 1 M for Na2SO4. No pfu were observed below 1 mM Na2SO4 or below 10 mM NaCl. There was a clear difference of the optimal concentrations between the alkaline solution, 0.1 mM, and the neutral solutions, 100 mM or higher. The cation, Na+, was the same in both cases, so, the difference can be attributed to the anions, the optimal concentration of 0.1 mM OH- in alkaline solution and 100 mM or higher Cl- or SO42- in the neutral solutions. In the neutral ionic solutions, NaCl and Na2SO4, the preserving abilities of infectivity of virions by these neutral ions increased with the increase of the ion concentrations higher than 1 mM.
The pfu profiles of virions in mixed solutions consisting of two sodium ionic compounds, NaOH and NaCl, were obtained using the heavier fraction of T4 virions (Figure 2, a dashed line). The concentrations of NaOH were changed, while the concentration of NaCl was constant, 100 mM, which was the concentration maintaining ca. 40% of the viral infectivity in pure condition. The pfu profiles were not simple summations of pfu of 100 mM NaCl plus pfu of NaOH. When the concentrations of NaOH were higher than 0.1 mM, 100 mM NaCl did not modify the pfu profile of NaOH. When the concentrations of NaOH were lower than 0.1 mM, the pfu values of the mixed solutions were more than the sum of NaOH and NaCl at the concentration of NaOH was 0.01 mM, and the pfu values were equivalent to the pfu at 100 mM NaCl when the concentration of NaOH was 0.001 mM.
3.4. K+: KOH, KCl, and K2SO4
The general activities of potassium ion, K+, in the solutions of KOH, KCl, and K2SO4 were similar to that of Na+ (Figure 2 and Figure 3). In the alkaline solution of KOH, ca. 90% of viral activity was maintained as the ion levels ranged between 0.1 and 10 mM after 15 min. suspension. The pfu values were like 0 or 100, or like a switch on/off, when the concentration of KOH was 0.1 mM (pH 10 equivalent). This causes a very wide error range of this point (Figure 3 and Figure 6). No or very few pfu was observed below 0.1 mM KOH or higher than 10 mM KOH (pH 12 equivalent).
In the neutral ionic solutions, KCl and K2SO4, the abilities of preserving the infectivity of virions increased with the increase of ion levels higher than 1 mM. No pfu was observed below 1 mM of KCl or K2SO4. According to the solubility, pfu higher than 100 mM was not tested for the K2SO4 solution. Clear differences of the pfu profiles between the alkaline KOH solution and the neutral KCl and K2SO4 solutions. Like Na+, the cation, K+, was common in these cases, so, the difference can be attributed to the anions, OH- in alkaline solution and Cl- or SO42- in the neutral solutions.
3.5. Mg2+: Mg(OH)2, MgCl2, and MgSO4
In the divalent cation Mg2+, the general profiles of pfu spectra along concentrations were similar to the profiles of the univalent cations, Na+ and K+ (Figure 2, Figure 3 and Figure 4). However, in the alkaline solution of Mg(OH)2, a clear jump of viral activity was observed at 0.05 mM (pH 9.7 equivalent), not 0.1 mM. The highest preservability of the infectivity of T4 virion was observed at 0.15 mM Mg(OH)2, and the preservability was nearly 100% (Figure 4). No or very few pfu were observed below 0.05 mM. According to solubility, concentrations higher than 0.15 mM Mg(OH)2 solution were not examined.
In the neutral ionic solutions, the highest value of the preservability of infectivity was lower than 70% (Figure 4). The preservabilities increased linearly with the increase of the concentrations of MgCl2 and MgSO4 on the semi-log plots, and the ion level of starting the increase was 0.01 mM, which was two orders lower than the cases of univalent cations, Na+ and K+. No pfu was observed below 0.01 mM in both MgCl2 and MgSO4. The highest values of preservation, the initial concentrations of increase the preservation and the slopes of the increases were clearly different between the alkaline solution, Mg(OH)2, and the neutral solutions, MgCl2 and MgSO4 (Figure 4).
3.6. Ca2+: Ca(OH)2, CaCl2, and CaSO4
The general profiles of pfu spectra along Ca(OH)2 concentrations were similar to the profiles of pfu in NaOH or KOH solutions (Figure 2, Figure 3 and Figure 5). In the alkaline solution of Ca(OH)2, 70 - 100% of viral activity was maintained as the ion levels ranged between 0.1 and 10 mM after 15 min. incubation. At the concentration of Ca(OH)2 as 0.1 mM (pH 10 equivalent), the pfu values were like 0 or 100, or like a switch on/off (Figure 5). No or very few pfu were observed below 0.1 mM Ca(OH)2 or higher than 10 mM Ca(OH)2 (pH 12 equivalent).
In the neutral ionic solutions, the preservabilities of the infectious abilities increased linearly with the increase of the concentrations of CaCl2 and CaSO4 on the semi-log plots. According to solubility, solution of CaSO4 higher than 10 mM was not examined. The level of concentration at which pfu started to increase was 0.01 mM, which was two orders lower than the cases of univalent cations, Na+ and K+ (Figure 2, Figure 3 and Figure 5). No pfu was observed below 0.01 mM in both CaCl2 and CaSO4 (Figure 5). The highest values of preservation, the initial concentrations of increase the preservation and the slopes of the increases were clearly different between the alkaline solution, Ca(OH)2, and the neutral solutions, CaCl2 and CaSO4 (Figure 5).
The pfu profiles of T4 virions suspended in mixed solutions consisting of two calcium ionic compounds, Ca(OH)2 and CaCl2, were obtained with the heavier fraction of T4 virions (Figure 5, a dashed line). The concentration of CaCl2 was fixed at 100 mM, which maintained ca. 50% of the viral infectivity (Figure 5), while the concentrations of Ca(OH)2 were changed. The pfu profiles were not simple summations of pfu at 10 mM CaCl2 and pfu at Ca(OH)2. When the concentrations of Ca(OH)2 were higher than 0.3 mM, coexistence of 10 mM CaCl2 did not modify the pfu profile of Ca(OH)2. At the concentration of Ca(OH)2 as 0.1 mM, the pfu of the mixed solutions was ca. 100, which was equivalent to the switch on case of Ca(OH)2 (Figure 5). When the concentrations of Ca(OH)2 were lower than 0.1 mM, the pfu values of the mixed solutions were more than the sum of Ca(OH)2 and CaCl2 at the concentration of Ca(OH)2 was 0.01 mM, and the pfu values were equivalent to the pfu of 10 mM CaCl2 when the concentration of Ca(OH)2 was 0.001 mM (Figure 5).
3.7. OH-: NaOH, KOH, Mg(OH)2 and Ca(OH)2
To show the abrupt increase of the infectivity at around 0.1 mN OH-, or pH 10, every count of pfu in alkaline solution in figures 2-5 was plotted against the concentration of alkaline (Figure 6). The sustainability of the infectivity of T4 was investigated in the range of concentration from 0.01 mN to 100 mN. According to the solubilities, the highest concentrations examined were 2 mN and 10 mN for Mg(OH)2 and Ca(OH)2, respectively. Any or little sustainability could be observed in the concentrations of OH- below ca. 0.1 mN. The sustainability of the infectivity of T4 jumps up at around 0.1 mN OH-; 0.1 mN for NaOH, KOH, and Ca(OH)2 and 0.05 mN for Mg(OH)2, corresponding to pH 10 and pH 9.7, respectively. In the higher range concentrations of alkaline solutions, the sustainability was suppressed, and no sustainability was observed over 10 mN OH-, corresponding to pH 12.
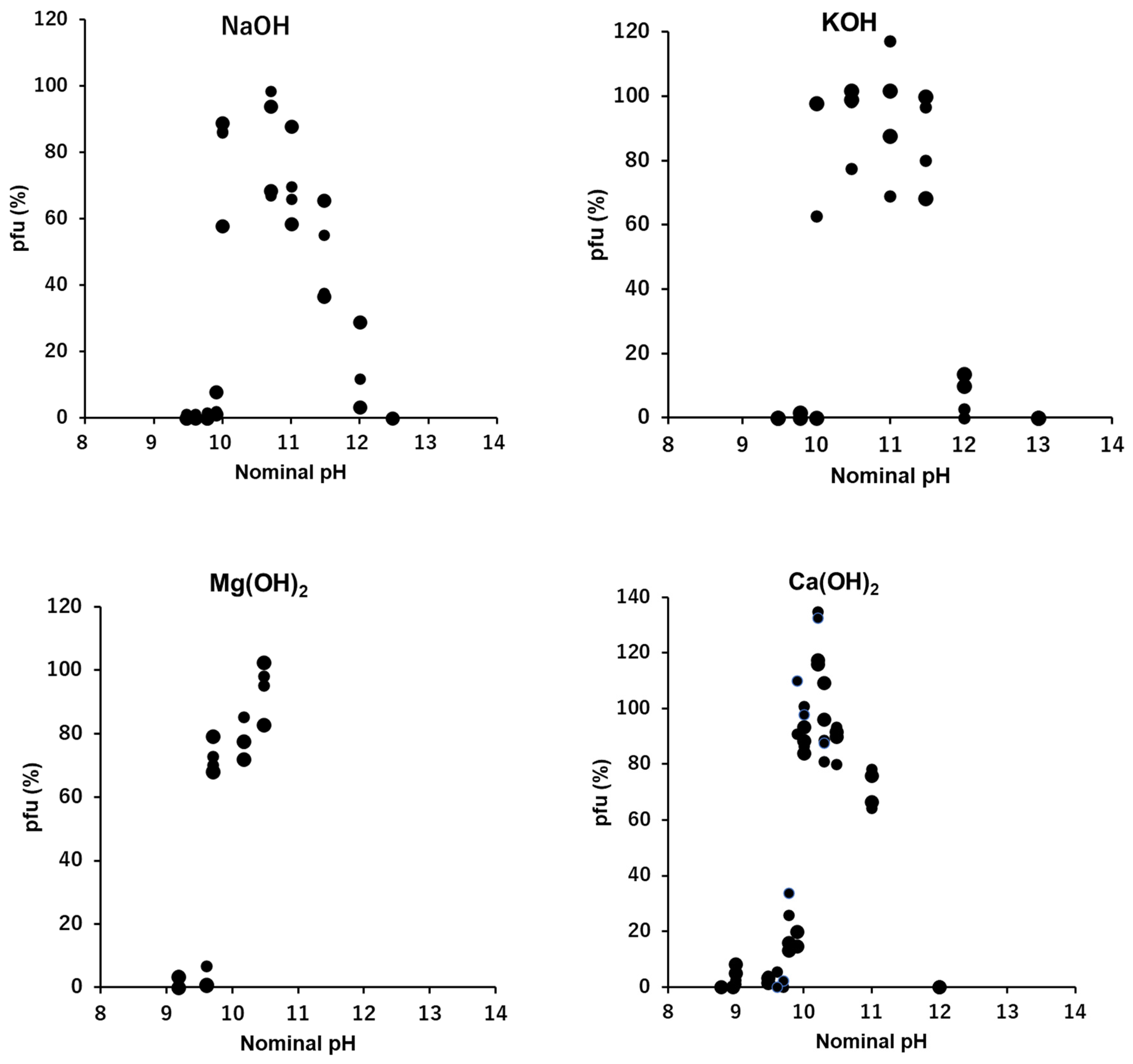
Figure 6.
OH-: The sustainability of the infectivity of T4 in quasi-pure alkaline solutions. The scales of abscissae are shown in nominal pH.
Figure 6.
OH-: The sustainability of the infectivity of T4 in quasi-pure alkaline solutions. The scales of abscissae are shown in nominal pH.
4. Discussion
The effects of a quasi-pure solution of a sole ionic compound on the survival of T4 virions are examined. Previous studies of ionic effects on virion activities were mainly focused on the integrated effects of ions on the virus-host adsorption during pre- and on-adsorption processes in multi-ionic media or buffers [2,4,8,13,14]. In this study, the pre-adsorption processes are separated from on-adsorption processes and the effect of a sole ionic compound on the pre-adsorption process are examined. Virions experienced the sole ionic compound were plated and adsorbed to host bacteria following the conventional plating method. Accordingly, in this study the conditions of host bacteria and the adsorption were practically identical at whole cases and the numbers of pfu were the reflections of the irreversible alternations of virions produced during the 15 min. immersion in solutions of the sole ionic compound prior to the adsorption processes. Virions in our study were not exposed to pure solution of an ionic compound, but quasi-pure solution, i.e., when an aliquot of viral suspension was inoculated, a trace amount of ions included by the inoculants of virion suspensions were introduced to the test solution. However, the inoculated amounts of ions, i.e., 0.0001 T-buffer: 0.01 mM NaCl, 0.2 μM MgSO4, 0.05 μM phosphate buffer, were far below the minimum concentrations of these ions necessary for pfu formation in the conditions tested (Figure 2, Figure 3, Figure 4 and Figure 5). Indeed, if there were no supplementary ions, the virions lost their activity immediately and no pfu was observed at the tested conditions (Table 1). Therefore, even though the solutions tested were quasi-pure, it was practically pure solution from the viewpoint of viral responses to ionic conditions.
The relatively short time exposure to a solution of one ionic compound defines the survivals of following infection and multiplication processes of T4 virions. It has been known that the optimal divalent cation levels ranged between 10-2 and 10 mM [2,6] and phages such as T2 and T4 having a requirement of NaCl for adsorption at the level of 100 mM which is ca. one order of magnitude higher than divalent cations [4,8]. In this study, the results produced in quasi-pure neutral ionic solutions include results equivalent to the previous studies which were gained by controlling the target ions in multiple ionic solutions. The increasing curves of the preservabilities of infectivity along with the increase of the concentrations of quasi-pure neutral ionic solutions were linear to the concentrations of ions in univalent cations (data not shown). However, they were linear in the semi-log plot in divalent cations (Figure 4 and Figure 5) or the preservabilities increase logarithmically for divalent cations, i.e., the slopes of pfu to the concentrations of the divalent cations were in reverse proportion to the concentrations of the ions, i.e., the preservation of the viral infectivity increased rapidly at the threshold concentration of the divalent cation, afterwards, the slope of the increase turned gentle rapidly. The threshold concentration of the divalent cations to preserve infectivity was ca. 10-5 M, which is 10-2 - 10-3 times lower than the univalent cations [4]. This implicates the maintenance of the infectivity, or deprotonation of DNA in the viral head (see below), may require small amounts of divalent cations which may supply the deficient divalent cations to the pre-existed divalent cations in the viral heads.
Phages generally can show infectious abilities in culture media or in buffers the pH ranges of which are adjusted to 5 - 11 [1,10,11,15,16]. In this study, virions were exposed directly to “pure” acid or “pure” alkali solutions. Without additional coexisting media or buffers, T4 virions showed no ability to form pfu in any acid to neutral conditions (Figure 1). Because virions were inactivated instantly in pure water [2, 4, 8, this study], it is not clear the inactivation in the acidic condition was formed by a passive effect derived from no-preservative agent of infectivity or by an active inactivation of acidic ions. Coexistence of neutral ions, e.g., 0.1 M NaCl or 10 mM CaCl2, which had abilities to preserve the activities of virions in ca. 50%, protect virions from the inactivation by H+ only when the concentration of H+ was lower than 10-4 N, or higher than nominal pH 4 (Figure 1). Accordingly, hydrogen ion actively inactivated the T4 virions, and without neutral ions, T4 virions has no tolerance to sole acid to neutral condition to maintain their infectivity. On the contrary, it has been reported that virions were more stable in suitably high enough concentrations of alkaline ranging from pH 7 to pH 11 [10,11,15,16]. In this study, after fifteen minutes incubation, 70 – 90% of viral survival was maintained in 0.1 – 1 mM OH- solutions, that was equivalent to a range between pH 10 and pH 11. This indicates that in quasi-pure alkaline solutions T4 virions cannot sustain their infectivity below pH 10, i.e., T4 virions need alkaline condition, higher than pH 10, to sustain their activity (Figure 2, Figure 3, Figure 4, Figure 5 and Figure 6). Not only in the short time sustainability, the time-course changes of the viral survivals in these solutions indicated the inactivation coefficients, -k in the equation (1), were compatible in 0.1 mM NaOH, 100 mM NaCl and 10 mM CaCl2 and also -k of the sole 0.1 mM Ca(OH)2 and 0.1 mM Mg(OH)2 were equivalent or smaller than the full strength T-buffer (Table 1). Pure relevant alkaline solutions can preserve the viral activity considerably long time. At around pH 10, the bases of DNA, guanine and thymine start the deprotonation, pKa of deprotonation of guanine deoxyribose-5’-phosphate and thymidine-5’-phosphate are 9.7 and 10, respectively [17]. In the environment of pH higher than 12, the viral DNA becomes denatured [18]. This indicates a condition; the bases of the nucleic acids are deprotonated, but the DNA is not denatured; is a required condition for the maintaining of the infectivity of phage virions. The addition of neutral ions, e.g., 0.1 M NaCl to NaOH and 10 mM CaCl2 to Ca(OH)2, expanded the infectivity toward the lower alkaline concentration range. At 0.01 mN of OH-, at which hydroxy ion had no potential to sustain the infectivity, addition of neutral ions showed equivalent sustainability with the infectivity at 0.1 mN hydroxy ion (Figure 2 and Figure 5). This implicates these neutral ions may maintain the pre-existed deprotonation of the DNA bases in a viral head (see below).
In the acidic buffers, virions can maintain the infectivity over wider acidic ranges [1,10,11,19], however, in the quasi-pure acidic condition, T4 virions immediately lose their infectivity (Figure1). Alcohol-based disinfectants appear to have a minimal effect on non-enveloped viruses, while low-pH alcohols exhibit strong virucidal effects against them [19]. Virions tend to irreversibly lose their infectivity in the low-pH range. Contrarily, T4 virions maintain their infectivity in quasi-pure alkaline conditions in the pH range between pH 10 and pH 11 (12) (Figure 2, Figure 3, Figure 4, Figure 5 and Figure 6). When neutral salts coexist in the alkaline solutions, the sustainable range of pH expands wider to the lower alkaline concentrations (Figure 2 and Figure 5). Calcium hydroxide is used as virucide at the pH 13 in farms [20,21]. Our results also indicate the loss of infectivity of T4 virions at the pH range higher than 12. However, alkaline, including calcium hydroxide, show protective actions to the viral infectivity at the dilute ranges (Figure 2, Figure 3, Figure 4, Figure 5 and Figure 6). If other ions coexist, the range of protection can expand wider. Around the area where the alkaline virucidal agent is applied at higher concentration, zones of lower concentration surrounding the area always exist, and some types of virions may be able to maintain their infectivity at these lower concentration zones. While care may be required for using alkaline agents as the virucidal uses, the results of a bacteriophage, T4, cannot directly apply to the pathogenic virions. Understanding the activities of virions in ionic solutions is essential for the viral dynamics [3,4,5,6,7,8,9]. Prior to investigating the behaviors of virions in the solutions of the mixtures of multiple ions, like buffers, effects of individual ions on the viral behavior should be elucidated first. Our methodology provides a tool to elucidate the basic effects of individual ions, as well as mixed and combined ions, on the infectivity of viruses.
The solvent for the thermal denaturation of DNA used in Marmur and Doty (1962) was 0.15 M NaCl plus 0.015 M sodium citrate. The 15 mM of sodium citrate preserved DNA in the coil form, not the compact composition [23]. They emphasized that the solvent of DNA containing the univalent cation as concentration of 0.15 - 0.2 M Na+ ensure the DNA in the coil composition will not to be denatured. This implicates univalent cations may support the infectivity of virions by preventing the denaturation of DNA in the deprotonation condition in the viral head. In addition, Rao and Black (2010) claimed that T4 DNA is packed in the head with ~1000 molecules of imbedded and mobile internal proteins, and the IPI*, the major of them, distributed high density of basic residues on the surfaces pf them that may allow rapid DNA ejection through the portal and tail without unfolding-refolding. If this state of DNA is derived from the deprotonation of the DNA bases and the IPI* is the major agent of the deprotonation in the neutral ambient condition, the neutral ions may support the deprotonation by the IPI*, because without suitable concentrations of neutral ions virions lost the ability of infection [1,2,4,6,8,9,10,11]. Further studies to elucidate the specific roles of neutral ions and chelates on the preservation of the viral infectivity is anticipated.
5. Conclusions
Clear differences were identified in the minimum required concentrations to maintain the infectivity of virions; around 10-1 mM for hydroxide ions, 10-2 mM for divalent cations, and 1 mM for univalent cations; and the pfu-ion concentration curves of viral preservabilities; a switch on/off type for hydroxide ions, direct correlation for univalent cations and logarithmic curve for divalent cations (Figure 2, Figure 3, Figure 4, Figure 5 and Figure 6). The minimum required concentrations and pfu-ion concentration curves may be two facets for one process, while the mechanism how pre-adsorption exposure of virions to ionic solutions causes irreversible changes of infectivity in the following multiplication processes of them is unknown. The viral DNA in capsids maintain their compactions by ions like Ca2+, and immersion of virions in ionic solution will exchange ions between in heads and outer solution [23,25]. This conformational condition of DNA in the head, in addition to the ionic alternation on proteins [2,6], can affect the following multiplication processes of the virions. We proposed here the deprotonation of the bases of DNA, guanine and thymine, is essential for the infectivity of T4 virions. Higher than ca. pH 10 is the critical alkaline condition for the deprotonation [17], which reduces the hydrogen bond energy between the double helix DNA chains [26]. The viral DNA becomes denatured when the environmental pH is higher than 12 [18]. In the pH range between 10 to 12, the DNA is not yet denatured but at the state of lower hydrogen bond energy between the DNA chains. This may be the condition where the viral infectivity is preserved in alkaline solutions. However, the viral infectivity is preserved in the neutral condition when the solvent of the viral suspension contains relevant amounts of neutral ions as discussed above and shown in Figure 2, Figure 3, Figure 4 and Figure 5. These neutral ions are not the acceptor of protons of the bases but may protect the state of the pre-existed deprotonation of DNA. The maintenance of the deprotonation of DNA in the viral head can be another factor which affect the multiplication of the viruses. Further studies are required for elucidating the mechanism of these processes.
Author Contributions
S.H., Conceptualization, analysis and interpretation of data and writing original draft preparation; I.K., Analysis and interpretation of data and funding. All authors have read and agreed to the published version of the manuscript.
Funding
The research was supported by the Cooperative Program (No. 119, 134, 2015) of Atmosphere and Ocean Research Institute, The University of Tokyo to S.H.
Acknowledgments
We thank H. Ogawa and H. Fukuda for funding and supporting experiments. We thank Y. Fu for continuous support and help on our studies. We thank S. Chattarji and J. Mok for help in preparation of the manuscript.
Conflicts of Interest
The authors declare that they have no competing financial and non-financial interests.
References
- Ly-Chatain, M,H. The factors affecting effectiveness of treatment in phages therapy. Front Microbiol 2014, 5, 1–7. [Google Scholar]
- Christi, K.; Elliman, J.; Owens, L. A. Synthesis of the divalent cation requirements for efficient adsorption of bacteriophage on to bacterial cells. In: Harrington, Daniel, (ed.) Bacteriophages: an overview and synthesis of a re-emerging field. Bacteriology Research Developments. Nova Science Publishers Inc, New York, NY, USA, 2017, pp. 43-69.
- Adams, M.H. The stability of bacterial viruses in solutions of salts. J Gen Physiol 1949, 32, 579–594. [Google Scholar] [CrossRef]
- Puck, T.T.; Garen, A.; Cline, J. The mechanism of virus attachment to host cells: I. The role of ions in the primary reaction. J Exp Medic 1951, 93, 65–88. [Google Scholar] [CrossRef]
- Rountree, P.M. The Role of Certain Electrolytes in the Adsorption of Staphylococcal Bacteriophages. J gen Microbiol 1951, 5, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Landry, E.F.; Zsigray, R.M. Effects of Calcium on the Lytic Cycle of Bacillus subtilis Phage 41c. J gen Virol 1980, 51, I25–135. [Google Scholar] [CrossRef] [PubMed]
- Jończyk, E.; Kłak, M.; Międzybrodzki, R.; Górski, A. The influence of external factors on bacteriophages—review. Folia Microbiol 2011, 56, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Zemb, O.; Manefield, M.; Thomas, F.; Jacquet, S. Phage adsorption to bacteria in the light of the electrostatics: A case study using E. coli, T2 and flow cytometry. J Virol Meth 2013, 189, 283–289. [Google Scholar] [CrossRef]
- Luria, S.E.; Steiner, D.I. The role of calcium in the penetration of bacteriophage t5 into its host. J bacteriol 1954, 67, 635. [Google Scholar] [CrossRef]
- Ahmadi, M.; Torshizi, M.A.K.; Rahimi, S.; Dennehy, J.J. Prophylactic Bacteriophage Administration More Effective than Post-infection Administration in Reducing Salmonella enterica serovar Enteritidis Shedding in Quail. Front Microbiol 2016, 7, 1253. [Google Scholar] [CrossRef]
- Phothichaisri, W.; Ounjai. P.; Phetruen, T.; Janvilisri, T.; Khunrae, P.; Singhakaew, S.; Wangroongsarb, P.; Chankhamhaengdecha, S. Characterization of Bacteriophages Infecting Clinical Isolates of Clostridium difficile. Front Microbiol 2018, 9, 1701. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York 2001.
- Rountree, P.M. The Role of Certain Electrolytes in the Adsorption of Staphylococcal Bacteriophages. J gen Microbiol 1951, 5, 673–680. [Google Scholar] [CrossRef]
- Jończyk, E.; Kłak, M.; Międzybrodzki, R.; Górski, A. The influence of external factors on bacteriophages—review. Folia Microbiol 2011, 56, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Brodetsky, A.M.; Romig, W.R. Characterization of Bacillus subtilis bacteriophages. J. Bacteriol 1965, 90, 1655–1663. [Google Scholar] [CrossRef] [PubMed]
- Tran XTT, Tam LD, Hoang HA. 2018 Stability and activity of TG25P phage in control of Aeromonas hydrophila in striped catfish pond water. Sci. Tech. Dev. J.; 21(2):64-70.
- Dawson, R.M.C.; Elliott, D.C.; Elliott, W.H.; Jones, K.M. Data for Biochemical Research (2nd edn), 1969; Oxford, Clarendon Press.
- Ageno, M.; Dore, E.; Frontali, C. The Alkaline Denaturation of DNA. Biophys J 1969, 1281–1311. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Matsumoto, N.; Hisaie, K.; Uematsu, S. Alcohol abrogates human norovirus infectivity in a pH-dependent manner. Scient Rep 2020, 10, 15878. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Alam, M.S.; Suzuki, M.; Takahashi, S.; Komura, M.; Sangsriratakul, N.; Shoham, D.; Takehara, K. Virucidal activity of a quaternary ammonium compound associated with calcium hydroxide on avian influenza virus, Newcastle disease virus and infectious bursal disease virus. J Vet Med Sci 2018, 80, 574–577. [Google Scholar] [CrossRef]
- Sangsriratanakul, N.; Toyofuku, C.; Suzuki, M.; Komura, M.; Yamada, M.; Alam, M.S.; Ruenphet, S.; Shoham, D.; Sakai, K.; Takehara, K. Virucidal efficacy of food additive grade calcium hydroxide against surrogate of human norovirus. J Virol Meth 2018, 251, 83–87. [Google Scholar] [CrossRef]
- Marmur, J.; Doty, P. Determination of the Base Composition of Deoxyribonucleic Acid from its Thermal Denaturation Temperature. J Mol Biol 1962, 5, 109–118. [Google Scholar] [CrossRef]
- Kuo, T.T.; Huang, T.C.; Wu, R.Y.; Chen, C.P. Specific Dissociation of Phage Xpl2 by Sodium Citrate. J gen Virol 1971, 10, I99–202. [Google Scholar] [CrossRef]
- Rao, V.B.; Black, L.W. Structure and assembly of bacteriophage T4 head. Virol J. 2010, 7:356. [CrossRef]
- Evilevitch, A.; Fang, L.T.; Yoffe, A.M.; Castelnovo, M.; Rau, D.C.; Parsegian, V.A.; Gelbart, W.M.; Knobler, C.M. Effects of Salt Concentrations and Bending Energy on the Extent of Ejection of Phage Genomes. Biophys J 2008, 94, 1110–1120. [Google Scholar] [CrossRef]
- Singer, M.; Berg, P. Genes & Genomes, a Changing Perspective 1991; University Science Books, Mill Valley, California.
Figure 1.
The effects of H+, HCl and H2SO4, on the infectivity of T4. Dashed lines show the cases of coexistence of neutral ions, 0.1 M NaCl or 10 mM CaCl2, in the HCl series.
Figure 1.
The effects of H+, HCl and H2SO4, on the infectivity of T4. Dashed lines show the cases of coexistence of neutral ions, 0.1 M NaCl or 10 mM CaCl2, in the HCl series.
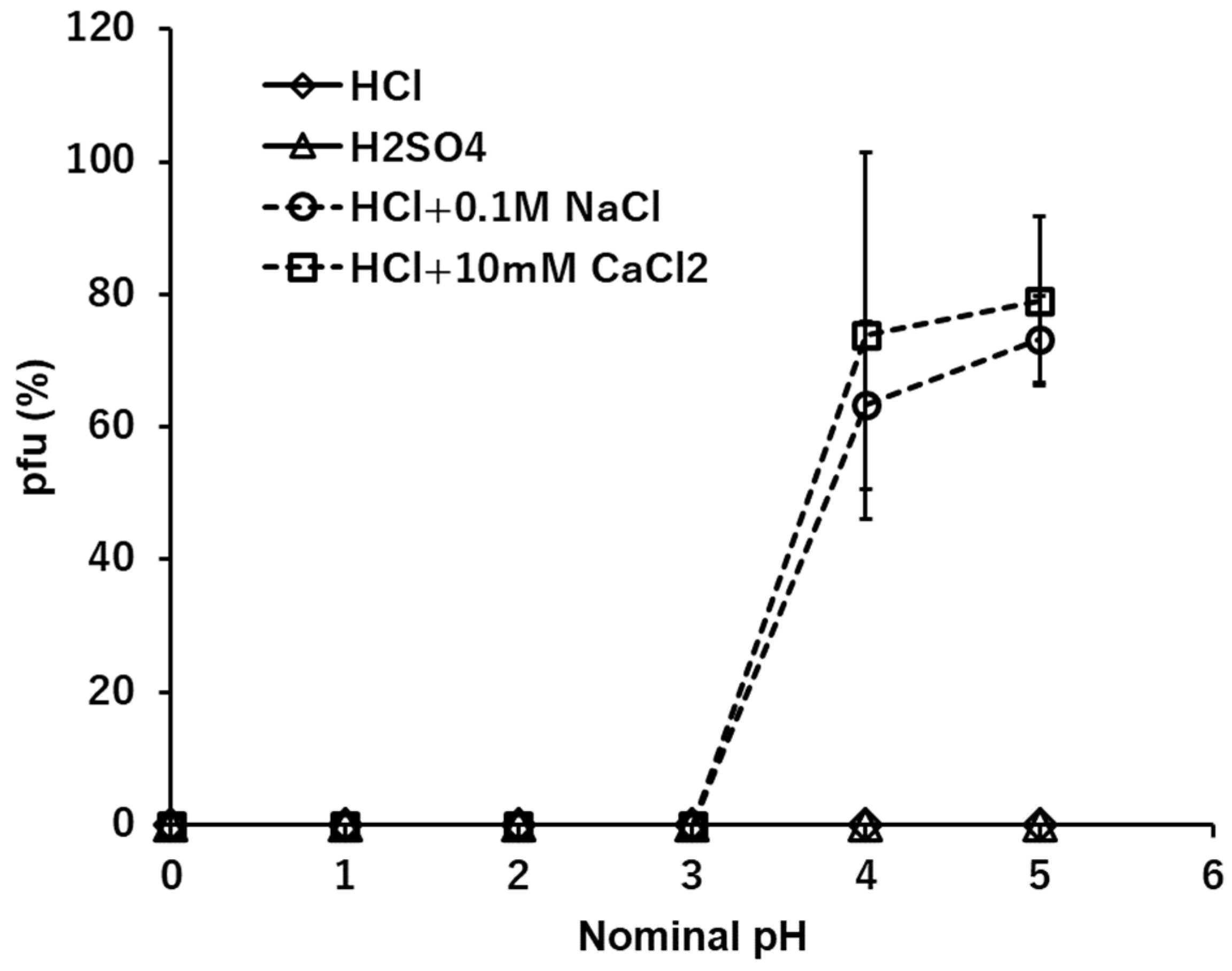
Figure 2.
The effects of the solutions of NaOH, NaCl, and Na2SO4 on the infectivity of T4. Addition of 0.1M NaCl in the NaOH series is shown in a dashed line.
Figure 2.
The effects of the solutions of NaOH, NaCl, and Na2SO4 on the infectivity of T4. Addition of 0.1M NaCl in the NaOH series is shown in a dashed line.
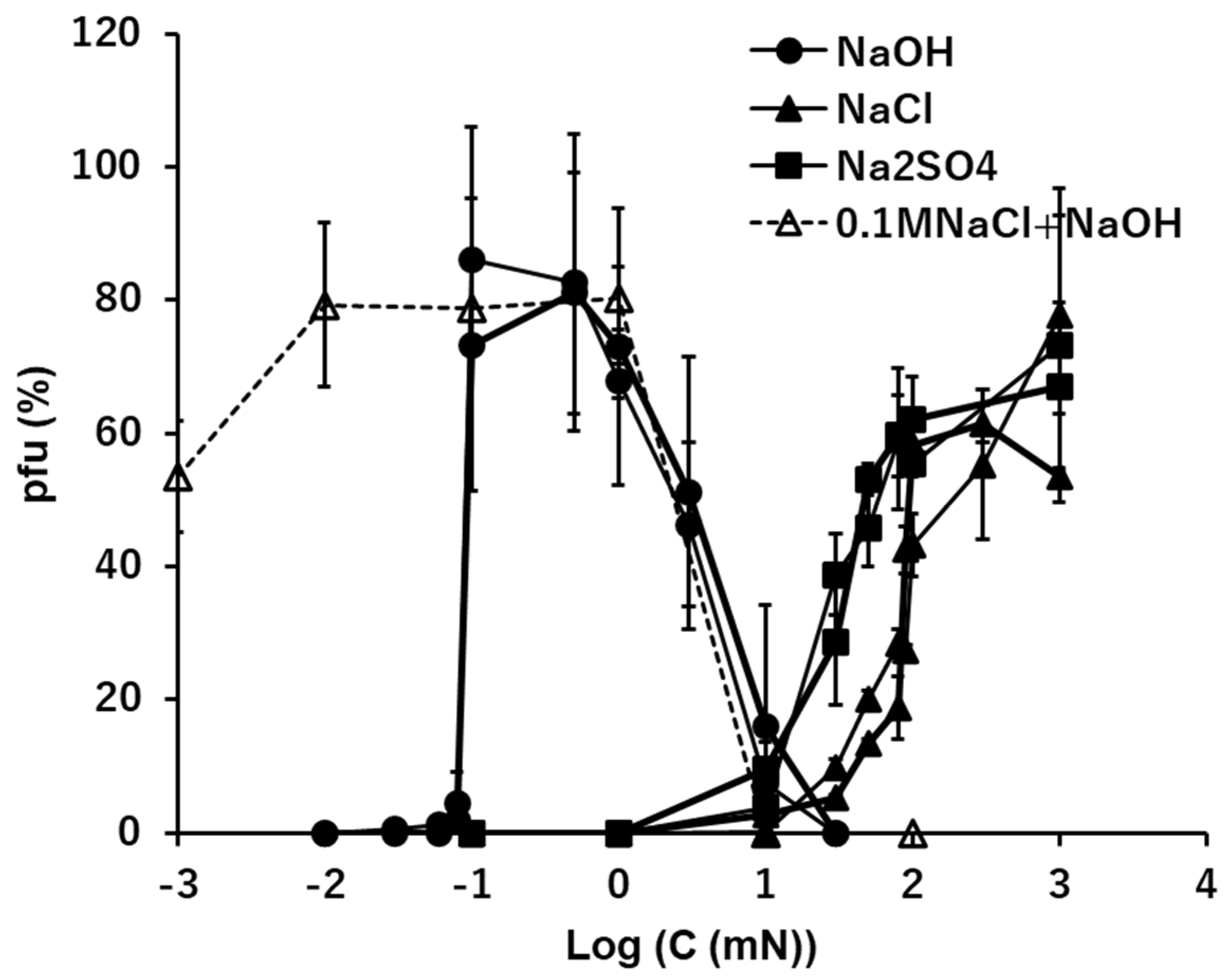
Figure 3.
The effects of the solutions of KOH, KCl, and K2SO4 on the infectivity of T4.
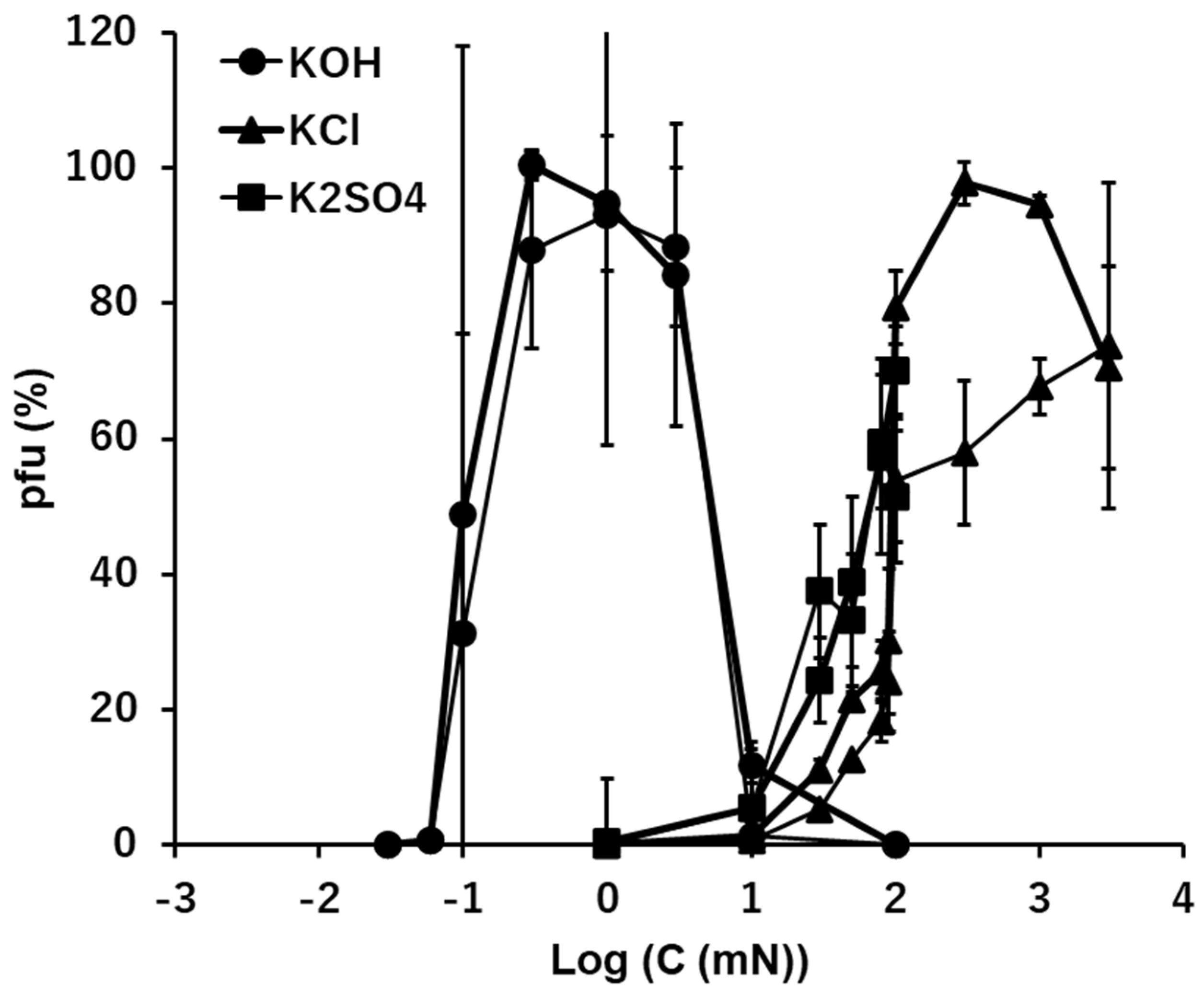
Figure 4.
The effects of the solutions of Mg(OH)2, MgCl2, and MgSO4 on the infectivity of T4.
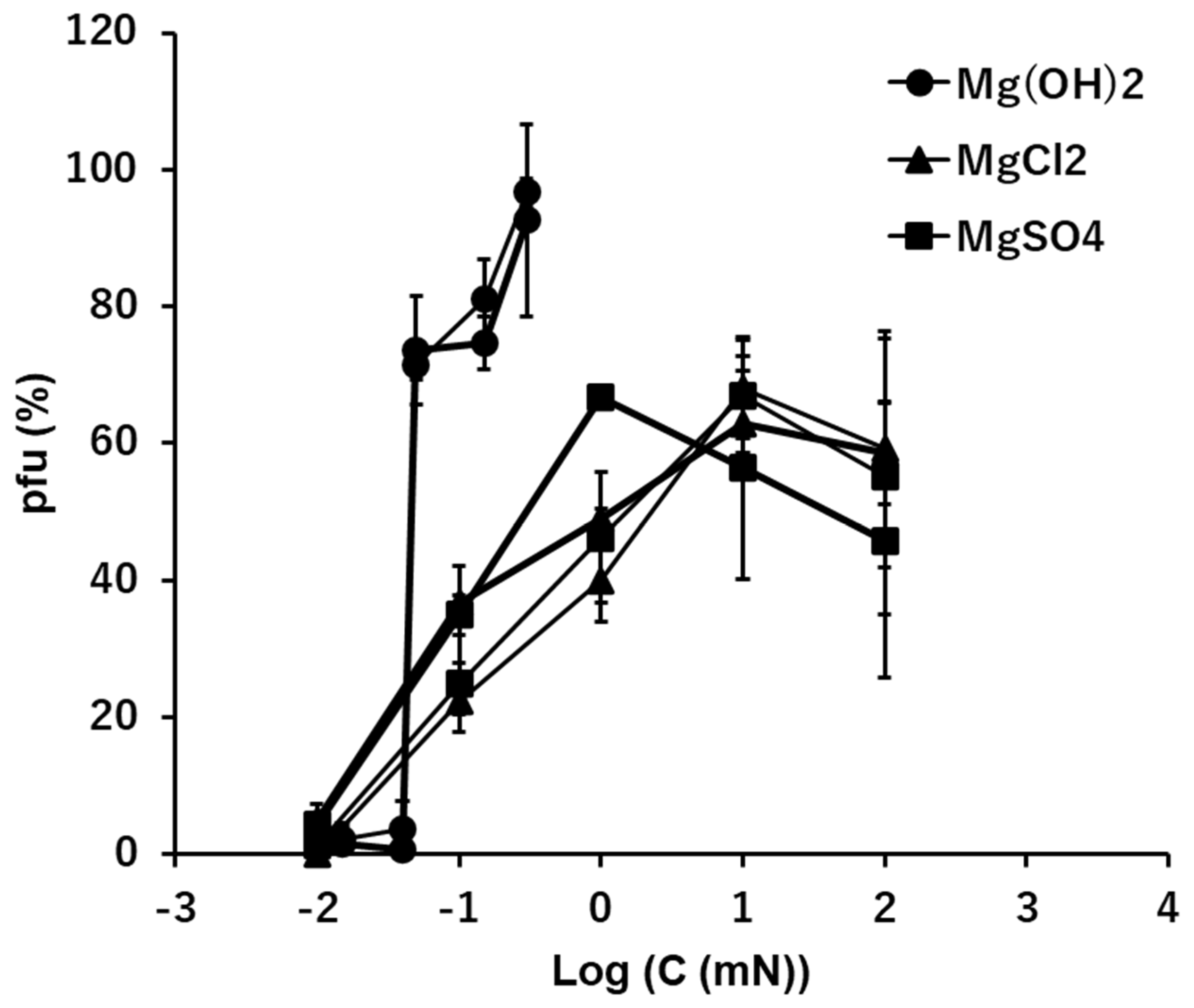
Figure 5.
The effects of the solutions of Ca(OH)2, CaCl2, and CaSO4 on the infectivity of T4. Addition of 10 mM CaCl2 in the Ca(OH)2 series is shown in a dashed line.
Figure 5.
The effects of the solutions of Ca(OH)2, CaCl2, and CaSO4 on the infectivity of T4. Addition of 10 mM CaCl2 in the Ca(OH)2 series is shown in a dashed line.
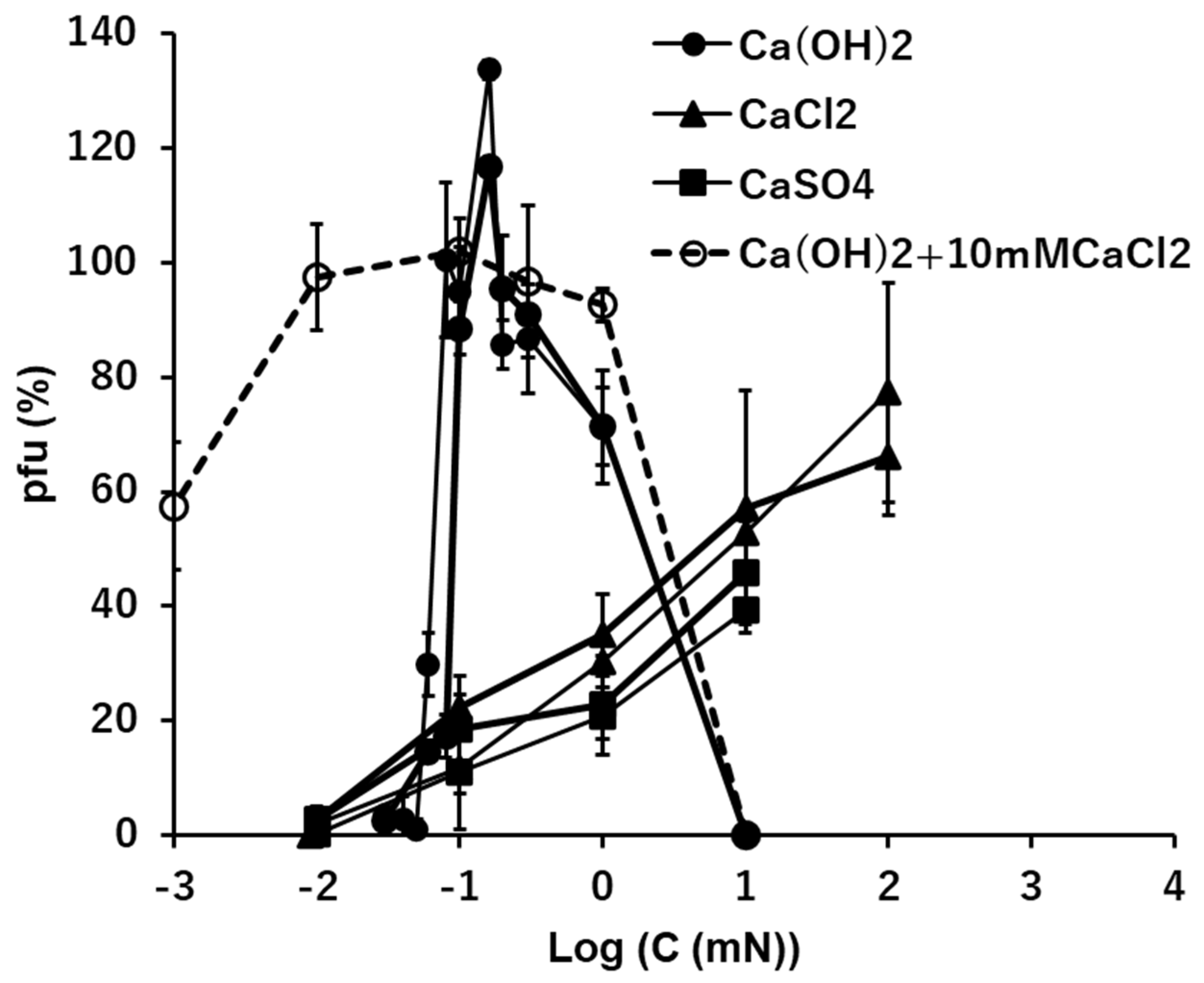

Table 1.
The inactivation coefficients in diluted T-buffers and quasi-pure ionic solutions. At 0.0001-fold dilution, virions were inactivated before plating and no pfu were counted. Slope is -k in the equation (1).
Table 1.
The inactivation coefficients in diluted T-buffers and quasi-pure ionic solutions. At 0.0001-fold dilution, virions were inactivated before plating and no pfu were counted. Slope is -k in the equation (1).
| Slope -k (min-1) | R2 | |
|---|---|---|
| 0.0001 T-buffer | † | † |
| 0.001 T-buffer | -0.38 | 1.00 |
| 0.01 T-buffer | -0.0095 | 0.72 |
| 1 T-buffer | -1.5x10-5 | 0.84 |
| 100 mM NaCl | -0.0014 | 0.99 |
| 10 mM CaCl2 | -0.0023 | 0.98 |
| 0.1 mM NaOH | -0.00064 | 0.59 |
| 0.1 mM Ca(OH)2 | -5.5x10-5 | 0.86 |
| 0.1 mM Mg(OH)2 | -2.1x10-6 | 0.26 |
| †: no pfu-value | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



