Submitted:
06 June 2023
Posted:
08 June 2023
You are already at the latest version
Abstract
Biological systematics, exploring the structure of taxonomic diversity (TD), needs a proper conceptualization that would (a) make it complying with certain universal criteria of scientificity and (b) take into account specificity of its studied object. This article outlines a possible conceptual framework for this natural science discipline based on the ideas of contemporary conceptualism. One of the most fundamental of them includes a concept of cognitive situation shaped by three interrelated basic components—ontic, epistemic, and subjective. Its core position is taken by a taxonomic theory (TT) being a conceptual construct resulted from a “summative” effect of all three components and designed for studying TD. The general TT is shown to be composed by a hierarchically arranged array of partial TTs of various levels of generality, and this construct serves as a conceptual justification of taxonomic pluralism. Several kinds of partial TTs are suggested to recognise, viz., ontology- and epistemology-oriented, aspect- and object-based, etc. The aspect-based TTs are of special importance as the conceptual foundations of particular research programs in systematics. The object-based partial TTs are shown to be implemented by concepts of species and homology. A large-scale relation between taxonomic theory and nomenclature is briefly considered.
Keywords:
contemporary conceptualism
; cognitive situation
; biological systematics
; philosophy of systematics
; taxonomic theory
; taxonomic monism & pluralism
; taxonomic nomenclature
1. Introduction
The philosophy of science is vital for the normal development and functioning of science. Understood in the most general sense, it substantiates all aspects of the knowing activity, including rational justification of what (ontology) and how (epistemology) is being cognised in the surrounding world, what kind of knowledge about the latter can be considered scientific (certain criteria of scientificity are presumed), on what grounds and to what extent the world and any of its manifestations (aspects, fragments, etc.) can be considered cognised, what is the role of a cognising subject in the cognition, etc. The rational formulations of these and other relevant questions and the search for possible answers to them shape the most fundamental issues in conceptualization of science, without which its normal development is thought to be impossible [1,2].
It is evident that both the posing and answering such questions are of crucial importance for the elaboration of all the knowledge about Nature, including acknowledging a particular natural science branch and the knowledge it produces scientific or not. With this, the possible ways of their posing and answering can be and indeed are different. This entails, among other things, a serious collision between monism and pluralism in the treatment of the scientific status of the disciplines employing specific exploratory principles. The positivist philosophy of science, having dominated not so long ago, pretended to possessing the only true criteria of scientificity. However, a more recent post-positivist philosophy of science, guided by the ideas of contemporary conceptualism and shaping the so-called non-classical version of science, favours both philosophical and, hence, scientific pluralism. This standpoint presumes that different natural science branches justifiably develop their particular approaches for exploring their specific objects, and these approaches are equally meaningful cognitively, providing that they fit certain general criteria of scientificity.
Biological systematics studies the diversity of organisms and provides the whole of biology with its universal, ordered description in the form of taxonomic classifications. Thus, the latter constitute a primary generalization about organismal diversity that frames, in a specific way, a kind of empirical basis for all biological disciplines. Accordingly, the answers to the above-mentioned fundamental questions, as applied to systematics, are of prime importance for the understanding of the scientific-philosophical status of its generalizations for the rest of biology.
The positivist philosophy of science, especially in its physicalism version that considers scientific only knowledge obtained by and expressed in the physical means, deprives systematics of the status of “real science” and treats it as but a “stamp collecting” [3]. To the contrary, a post-positivist philosophy justifies regarding systematics as a specific, full-fledged scientific discipline, and not a marginal “Cinderella of science” [4]. This non-positivist account of systematics is quite expressly exposed by the zoologist G. Simpson: “All theoretical science is ordering, and if [...] systematics is equated with ordering, then systematics is synonymous with theoretical science” [5, p. 5].
So, contemporary conceptualism seems to provide very important prerequisites for the current development of biological systematics. It causes such a move of the conceptual “historical pendulum” in the most recent development of philosophy of science that gives biological systematics a chance to become an acknowledged full-fledged natural science branch. With this, an anticipated “taxonomic renaissance” [6], when considered conceptually, can be achieved only by an elabouration of a well-developed, consistent theory of systematics based on a clear understanding of its general philosophical-scientific background. This makes the development of theoretical foundations of this discipline on a modern conceptualist basis of paramount importance.
This article, written by a philosophically-minded biologist, addresses both the philosophers of science interested in the theoretical foundations of biological systematics (taxonomy) and the systematicians interested in the philosophy of their discipline. It briefly outlines one of the possible versions of conceptualization of systematics that has been being developed by the author for years. Section 2 presents general ideas about the structure of a cognitive situation in which this scientific discipline operates. In Section 3, the meaning and structure of taxonomic theory (TT), representing a specific form of conceptualization of systematics, is considered in general. Section 4 describes the main research programms of systematics that implement partial aspect-based TTs for the explorations of various aspects of taxonomic diversity (TD). The concepts of species and homology are shown in Section 5 to be implementations of the partiar object-based TTs. A collision between monism and pluralism in the treatments of partial TTs is debated in Section 6. Section 7 discusses a contribution of TT to the development of taxonomic nomenclature as a specific part of the professional language of biological systematics. At last, the Conclusion highlights certain ideas that are considered by the author among the most important for the elaboration of a conceptual framework of contemporary systematics.
2. A Three-Partitioned Cognitive Situation
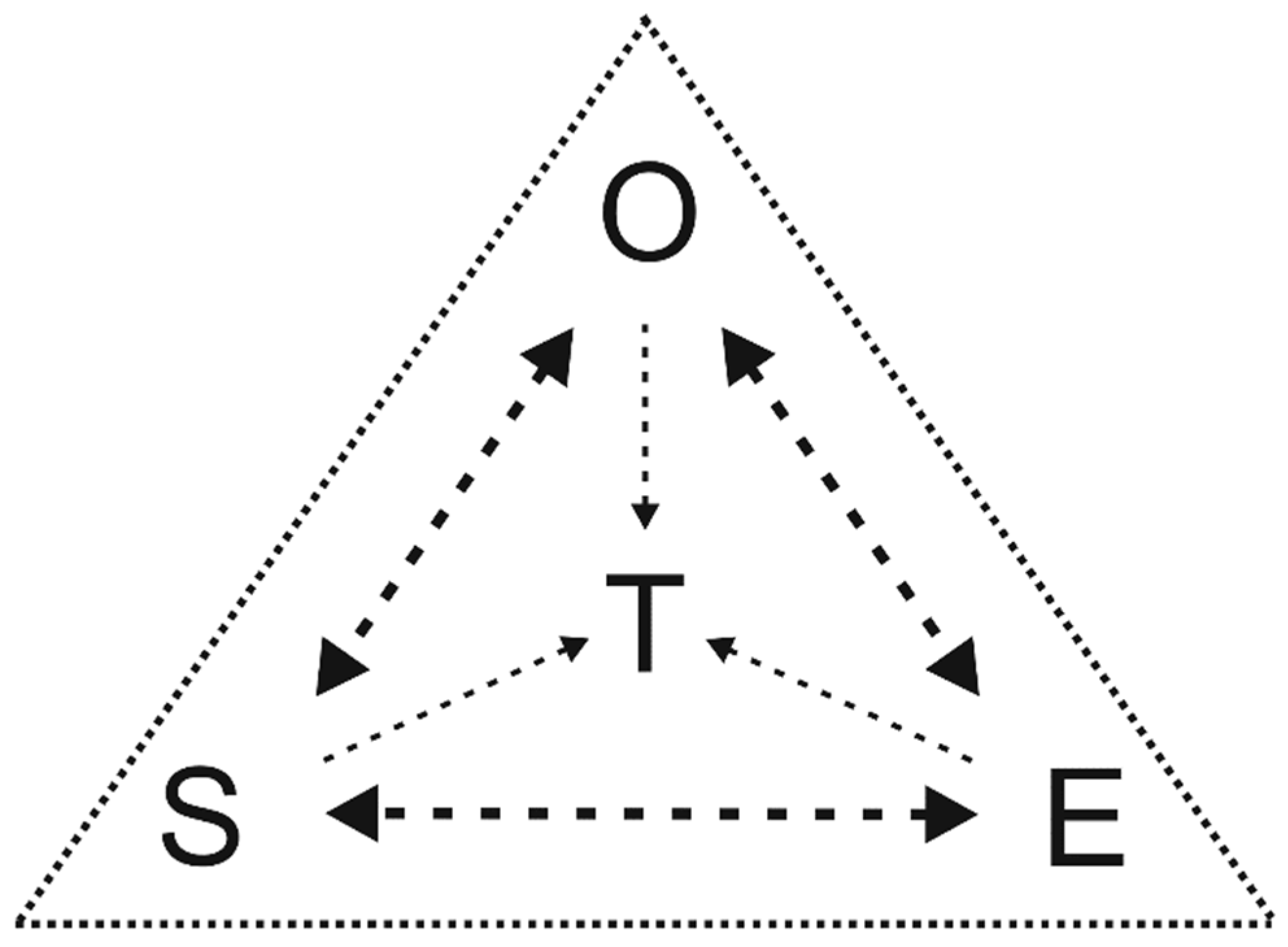
One of the basic provisions of contemporary conceptualism [7] asserts that any knowing activity is carried out in a certain cognitive situation. This fundamental notion refers to certain very basic properties of the process of cognition, so that it has no formalized precise definition, as is usual for such kind of notions, and it occurs rather in the form of an informal and largely intuitive grasping. An essential part of the latter is comprehension of a cognitive situation as being, in general, composed of and structured by three basic components. These are (a) an ontic component corresponding to an object to be cognised, (b) an epistemic component corresponding to certain cognitive aids, and (c) a subjective component corresponding to a cognising subject. They interact with each other in a complex manner, so both their triplicity and interdependence make it convenient to represent the entire cognitive situation metaphorically as a cognitive triangle (Fig. 1). Its vertices correspond to these three components of the cognitive situation, its edges reflect their interrelationships, and its center is occupied by a certain conceptual core, around which the entire cognitive situation is scaffolded. In our case, the core in question corresponds to a TT; and the most important conclusion following from the latter’s conceptualist account outlined here is that the entire content of this “focal” theory is a resultant of the interaction of all three components of the cognitive situation.
Figure 1.
A graphic representation of cognitive triangle visualizing a three-partitioned cognitive situation in biological systematics. Abbreviations: E – epistemic component, O – ontic component, S– subjective component, T – taxonomic theory. See text for explanations.
Figure 1.
A graphic representation of cognitive triangle visualizing a three-partitioned cognitive situation in biological systematics. Abbreviations: E – epistemic component, O – ontic component, S– subjective component, T – taxonomic theory. See text for explanations.
This Section specifies how the general idea of cognitive situation is involved in construing a conceptual framework of biological systematics suggested by the author. For this, its three basic components are characterized, its onto-epistemic background is considered as bounded by an onto-epistemic correspondence between its first two components, and a special emphasize is put on the role of its subjective component shaping that correspondence.
2.1. The Onto-Epistemic Background
The ontic component of a cognitive situation, roughly speaking, refers some way to a cognisable object, which is conveniently designated as a natural phenomenon. From a naturalistic standpoint, it is treated as a certain manifestation of Nature presumably belonging to the objective reality as a totality of all things, events, and processes [8]. However, according to the cognitive science, it is given to a cognising subject not by itself in its “suchness”, but rather it is distinguished and outlined by the very subject guided by a certain cognitive goal and applying certain cognitive means. This is achieved by fixing a set of essential properties, by which a subject differentiates a phenomenon of interest from other manifestations of Nature and, by this, “carves” it out of the latter. Conceptually considered, a cognisable object thus outlined (“carved”) is not a particular natural phenomenon itself but rather its specific representation, and it is pertinent to designate it as a cognitive model of that phenomenon [9,10]. It is actualized in a cognitive situation by a certain conception that refers to the natural phenomenon in question by indicating its essential features fixed by a subject. So, it is a cognitive model (and respective conception), and not a natural phenomenon as such, that constitutes the ontic component of a cognitive situation.
Thus, if a natural phenomenon can be referred to as a part of objective reality, the ontic component of a cognitive situation, given as a respective cognitive model of that phenomenon, should be regarded as a kind of conceptual reality. In the case of biological systematics, the natural phenomenon is aptly considered biological diversity (BD), which is treated as a specific manifestation (aspect) of living matter. The ontic component of a cognitive situation, in which systematics operates, involves a conceptually construed representation of BD referred to as a taxonomic diversity (TD), and it may be conventionally treated as a specific taxonomic reality [11,12] Accordingly, concrete classifications, elabourated by taxonomists to describe particular fragments of TD, can be interpreted as specific cognitive models of the latter.
The epistemic component of a cognitive situation involves a set of normative prescriptions to perform certain cognitive functions, of which the most important is ontic reduction unavoidably inherent in any knowing activity. As said above, it “turns” the objective reality (BD) into a conceptual one (TD) by “carving” a particular studied phenomenon from the totality of Nature and making it a particular cognitive model to become a part of a cognitive situation. Such models are evidently simpler than their “natural prototypes”, so they are separated from the latter by certain ontic gaps that may be more or less extensive depending on degree of reduction.
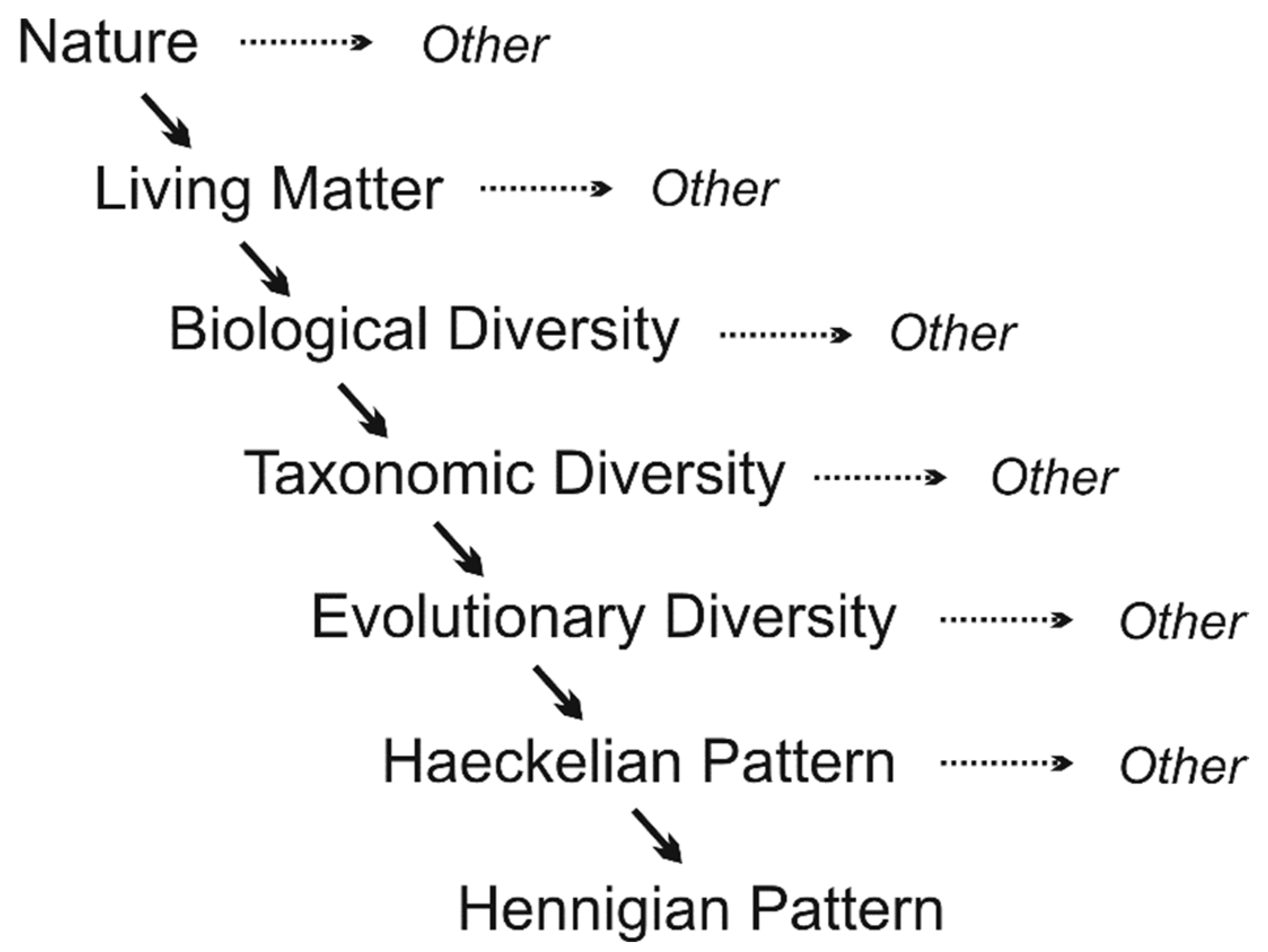
The ontic reduction proceeds as the so-called reduction cascade: it represents a descending stepwise procedure of the sequential fragmentation of the cognitive models of higher levels of generality into those of lower levels. This cascade begins with a model (concept) having the most general meaning, which corresponds to Nature or (in our case) BD. The latter is next fragmented into various manifestations, which are taxonomic, synecological, autecological, physiological, sociological, etc. forms of diversity, being dealt with by various domains of biology, and one of them is systematics. The TD, studied by the latter, is then fragmented into its own particular manifestations (phylogenetic, typological, biomorphological, etc.) as the subject fields of different parts of systematics, and this entire cascade reaches its end point with various operational conceptions that serve for elabourating concrete classifications. So, the principal outcome of such a reduction cascade is an encaptic (inclusive) hierarchy of cognitive models (conceptions) of different levels of generality. Both this cascade and the resulting hierarchy are endowed with some significant features to be kept in mind when reflecting on the conceptualization of biological systematics.
First, the cascade in question provides a kind of deductive argumentation scheme, in which lower-level cognitive models represent partial interpretations of the models of higher levels of generality. This means that each lower-level particular model is inferred within a certain reduction cascade in such a way that makes it acquiring its exact substantial meaning not by itself (ad hoc), but in the context of a more general, or higher-level model (post hoc). This is definitely true for the operational conceptions: they are “substantially empty” (biologically meaningless) if they are elabourated formally without any reference to some biologically meaningful conceptual models. For instance, formal classification methods developed by numerical phenetics do not distinguish between, say, close species and biomorphs, as their recognition is based on certain a priori applied biologically sound criteria [13].
Second, the ontic reduction, as said above, means that any natural phenomenon is more complex than its cognitive model. This is because only some features of the former, considered most essential from a certain standpoint, are included in the elabourating its model, while others are discarded as redundant or irrelevant for resolving the respective cognitive task [9]. Consequently, the longer the reduction cascade leading to lower-level cognitive models, the wider is an ontic gap separating the latter from the “top” phenomenon (BD) (Fig. 2). For instance, the Haeckelian phylogeny is meaningful biologically by referring to the multifold historical development of organisms, including both cladogenetic and anagenetic components of evolution together with their adaptationist explanations and timing. Unlike this, the Hennigian cladistic pattern involves branching phyletic lineages only and discards all other evolutionary features [12,14], so it seems to refer to something biologically meaningless, like a “spherical horse in vacuum”.
Figure 2.
Reduction cascade showing a sequential procedure of transformation of Nature (a part of overall objective reality) into manifestations of taxonomic diversity (TD) as particular conceptual realities of various levels of generality. See text for explanations.
Figure 2.
Reduction cascade showing a sequential procedure of transformation of Nature (a part of overall objective reality) into manifestations of taxonomic diversity (TD) as particular conceptual realities of various levels of generality. See text for explanations.
Third, a multifold natural phenomenon can be represented by (reduced to) several simpler cognitive models, each corresponding to a certain particular manifestation (aspect, fragment, etc.) of its “natural prototype” [9]. As a result, the cascade in question yields something like a conceptual pyramid, in which the number of representations (models) of certain complexly structured natural phenomena gradually increases with the movement from an initial to the final steps of reduction [12]. Indeed, the generally understood taxonomic reality can be conceptualized in various ways to provide a number of different substantive conceptions (as mentioned above), and each of the latter is detailed in different ways to produce an array of simpler operational conceptions. Thus, such a conceptualist interpretation of the reduction cascade presumes plurality of the admissible representations of taxonomic reality. This means that systematics in fact deals not with a unified homogeneous cognitive situation and a corresponding solid conceptual space, but rather with a hierarchically ordered set of local situations and respective (sub)spaces.
It is to be emphasized that the moving from top to bottom of the reduction cascade results in the restructuring the contents of taxonomic realities in such a specific way that their “objective” account successively decreases while “subjective” one increases in the same direction. Recalling the above-provided example with two conceptions of phylogenetic systematics, it is admissible to say that its more complex Haeckelian version is more “objective”, whereas a more reductionist Hennigian one is more “subjective”. Though, the most devoted cladists, who disbelieve in “metaphysics” and prefer to operate with some exactly prescribed algorithms, are sure of the opposite. However, they obviously confuse a natural objectivity (correspondence to the natural phenomena) with an instrumental intersubjectivity, i.e., a repeatability of the results of application of the standardized algorithms to the standardized data [15].
It follows from the preceding that the basic ontological question of the “existence” of something in Nature, to which TD and its particular manifestations supposedly relate one or another way, has no reasonable answer without its epistemic consideration. The latter involves an array of the so-called epistemic values that provide certain criteria of judgment about existence/non-existence of the phenomena under study [16].
2.2. An Effect of the Subjective Component
The place of a subject (in a broad sense) in a cognitive situation is rarely taken into account, and if it is, its role is most usually linked to the so-called non-epistemic values to be applied to the results of knowing [17]. With this, according to the positivist philosophy of science, any possible subjective effects are to be minimized; in systematics, this is especially evident in its numerical school [13]. On the contrary, modern conceptualism, including certain provisions of cognitive science, acknowledges an irremovable influence of a cognising subject on the entire structure of cognitive situation and, consequently, on both the process and the results of knowing [2,7,18]. This is because a subject’s participation in whatever cognitive situation means not a passive reflection of the studied phenomena, but instead an active intellectual interaction with them.
The subjective component is complexly structured by comprising individual researchers, particular scientific groups (schools, labs, etc.), and eventually the entire human society. At a personal level, it is evident in the variety of thinking styles (holistic or elementaristic, intuitive or analytical, etc.) that define how the natural phenomena are perceived. At a higher level, this component manifests itself in the devotion of the local research groups to certain paradigms or other conceptual constructs: they elabourate certain rules that dictate their members, what should be treated as the taxonomic reality and how the latter is to be explored and represented in classifications. At a far higher level, the scientific community is influenced by the governing bodies, whose decisions are often taken on grounds not related to the science itself.
Considering the structure of a conceptual framework of a cognitive situation, the principal function of its subjective component involves the structuring the entire situation in question by regulating both the contents of its two other components and their interrelations. Indeed, it is the cognising subject that formulates all normative onto-epistemic prescriptions, and it regulates by this the entire procedure of ontic reduction, including transformation of BD into its particular manifestations being studied. Proceeding from this ontic reduction, the same subject develops methodologies considered the most appropriate for elabourating particular classifications to make them the most adequate to the TDs resulting from the reduction cascades.
2.3. The Onto-Epistemic Correspondence
A very important outcome of such “integrative” function of a cognising subject is a specific interdependence (albeit partial) of ontic and epistemic components; this is formalized by the principle of onto-epistemic correspondence [11,12]. Due to this, these two components make up together an integrated onto-epistemic basis of a cognitive situation. It shapes a conceptual framework of the latter [19,20], and its elabouration is the principal objective of the whole procedure of conceptualization of biological systematics.
The correspondence in question looks as follows. On the one hand, the studied object is embedded in the cognitive situation, as a part of its ontological component, depending on certain epistemic considerations, before all some fundamental criteria of its knowability by specific research tools. For instance, only observable (albeit instrumentally) and measurable phenomena are regarded cognisable and therefore scientific by positivism and phenomenalism, while any discourses about unobservable (including, say, about biological evolution) are treated as non-scientific metaphysical speculations. On the other hand, some important methodological principles, being a part of epistemic component, are developed based on certain fundamental properties attributed to the studied object. For instance, an antique Pythagorean idea, according to which the whole of Nature is ruled by the Harmony of Numbers, has led to Galilei’s assertion that “The Book of Nature is written in the language of mathematics”, and this inspired Leibniz and Kant to suggest to counting scientific only those judgments about Nature that can be expressed numerically [21], which eventually gave rise to the numerical methods of contemporary systematics.
This two-side interrelation may become predominately one-sided if the most emphasis is put intentionally on either of these two basic components; this is clearly exemplified by two cognitive situations in phylogenetics [14]. In its traditional “handmade” version (Haeckel, Hennig), specific methodologies of phylogenetic reconstructions are inferred from an assumed divergent nature of phylogenesis: here, epistemology follows ontology. In numerical phyletics, phylogeny is suggested to treat as a stochastic process, with this idea being substantiated by the reference to certain statistical methods applied to phylogenetic reconstructions (Felsenstein): this means that ontology follows epistemology.
The principle under consideration may be treated as a specific complement to the general provisions of ontological relativity developed by contemporary conceptualism [18,22]. This “relativity” refers to what was said above concerning the ontic component of a three-partitioned cognitive situation: it represents not the studied natural phenomenon (BD) as such, but its certain intellectual image (TD) fashioned by a subject. These are the epistemic tools that are applied by the latter and therefore are responsible for shaping a significant part of such image and, by this, for the ontological (ontic) relativity. An irremovable effect of the epistemic component inevitably leads to a specific “epistemic pitfall” caused by ignoring or downplaying the above-mentioned ontological gap [23]. Due to this, accordingly to one of the Zen Buddhism aphorisms, “a person is inclined to mistake his finger pointing at the moon for the moon itself”. In our case, when biologists analyse biological diversity—say, more particularly, species diversity—they scarcely realize that it is a particular conception of species (a “finger”) that makes them see in Nature certain phenomena they call the species (the “moon”) [24,25].
3. Basic Properties and Structure of Taxonomic Theory
As the philosophy of science insists, no scientific discipline can function normally without a more or less developed theoretical background, so that the maturity of a discipline is “measured” by its progress with the developing its theory. With this, however, although the notion of scientific theory is one of the key and basic ideas in the philosophy of science, its comprehensive and uniform definition does not exist; moreover, it cannot probably exist because of the qualitative heterogeneity of the entire pool of scientific knowledge [2]. Accordingly, the above-mentioned non-classical philosophy acknowledges that it is normal for particular science branches to develop their own theoretical foundations, presumably most adequate to the nature of the objects they study, providing that they comply with certain most general criteria of scientificity (which in themselves are understood rather ambiguously).
This general non-classical tenet certainly holds true for biological systematics, so the latter’s progress can be measured by the success in the development of its own theory, which is pertinent to name taxonomic theory (TT) [11,12]. The latter is understood herewith as a part of the entire conceptual framework that is construed purposefully for biological systematics, so this notion was suggested keeping in mind the designation of the theoretical branch of the latter as taxonomy since the early 19th century (de Candolle). The basic content and structure of TT, thus considered, corresponds evidently to those characteristic of that framework: it is partitioned into ontic and epistemic components interlinked by onto-epistemic correspondence due to an effect of the subjective component. So it is a particular biological taxonomy, and by this it differs from a “universal” one understood as a “general theory of taxonomy”, representing “a standardized framework to classify and organize items [of any kind] into categories” [26,27].
The attempts to develop theoretical foundations of systematics have been undertaken repeatedly in its long conceptual history. Some of them were aimed at developing certain kinds of universal systems of classification, as the versions of just-above “universal taxonomy”, by applying some elements of the formal languages borrowed from mathematics [28,29,30]. Some others tried elabourating more substantive, biologically meaningful theories, but these served primarily as the foundations of the particular research programms in systematics (phenetics, phylogenetics, evolutionary taxonomy, etc., see also below) [13,31,32]. Thus, we have to state a sorrowful fact that nowadays biological systematics does not possess any well-developed comprehensive TT that would (a) comply with certain most general provisions of theoretical knowledge in natural science, (b) be specific enough to be adequate to the essential properties of an object studied by biological systematics, i.e., BD, and (c) cover by its propositions all research programms (schools) of this discipline.
Any theoretical endeavors of such kind are denied by the empiricists, who believe that systematics, unlike “more fundamental” biological disciplines (e.g., ecology), “has no theory, being merely diagnostics and classification” [33, p. 100]. This standpoint ascends to the folk systematics and preserves its basic features, so it is popular among practical systematicians, who realize their “aspiration for truth without engaging any theoretical reasoning” [34, p. 367]. They usually appeal to the personal intuitive knowledge strengthened by an individual experience, and this position occasionally acquires its philosophical support [35,36]. As follows, the innumerable practical studies on BD, constituting an empirical fundament of all biological knowledge, provide but a vastly fragmented primary information about particular groups of organisms obtained by individual researchers.
From a conceptualist standpoint, the only possible rational basis for integrating a “patchwork” of such particular empirical fragments into a consistent global biological knowledge seems to be a certain TT elabourated as a part of the conceptual framework of systematics. For instance, to “measure” BD by counting the number of species in whatever groups of organisms, it is highly desirable that experts on these groups would operate with (nearly) the same unified, rationally substantiated general species concept covering all of its particular interpretations [37,38]. Thus, the development of a well-founded TT is an urgent need of a proper conceptualization of biological systematics.
This Section summarizes the author’s reflections about one of the possible approaches to develop TT as an essential part of a conceptual framework of systematics [11,12]. Its content should best be considered an attempt to outline one of the possible directions, in which conceptualization of contemporary systematics may proceed to reach a desirable end. Within this task, it is mostly designed to outline more rigidly the basic notions that constitute a conceptual core of TT in its general meaning. Accordingly, it suggests considering TT a quasi-axiomatics, specifies its hierarchical structure, and briefly touches the problem of the definitions of taxonomic objects identified by TT.
3.1. Taxonomic Theory as a Quasi-Axiomatics
The basic conceptual propositions of biological systematics, which are outlined in the advanced (in this respect) manuals, ascending to Linnaeus’ Systema Naturae, are designated as “canons”, “rules”, “axioms” or (more often) “principles”, without delineating them according to their functions. However, in compliance with the general ideas of the structure of a cognitive situation outlined above, these propositions are of two fundamentally different kinds: some of them are about ontology while others about epistemology. To make this imperative difference explicit, it is pertinent to elabourate TT as a quasi-axiomatics, in which two principal categories of basic statements are distinguished [11,12]. Those referring to the ontic component of a cognitive situation could be qualified as quasi-axioms or presumptions, while those referring to its epistemic component could be qualified as inference rules; these are the latter that may appropriately be called principles of taxonomic research.
Defining such theory as a quasi-axiomatics presumes that it is being elabourated not as a system of formal statements: instead, its basic ontic propositions are initially endowed with a certain biological meaning. Indeed, as far as TT is purposefully developed as a specific device for exploring TD, its ontic propositions are substantiated by the reference to the essential properties of TD. Two basic kinds of these propositions are distinguished by their conditionality as follows [11,12,39]. The quasi-axioms are stated in an “absolute” form; their truth is not questioned and is not verified within a specific taxonomic research. The presumptions are adopted in a “relative” form; their truth status is treated probabilistically, and it may change in the course of an exploration. Their conditionality depends on certain background ontic theories, within which these propositions are formulated. For instance, an assumption of evolution as a cause of the diversity of organisms has a quasi-axiomatic status in a phylogeny-based theory, while it is a non-obligatory presumption in a typological theory. The judgments about homologies are always presumptions, as far as they are probabilistic, and their probability assessments depend on the particular substantive contexts.
According to the principle of onto-epistemic correspondence introduced above, the ontic and epistemic premises of the same conceptual framework are interconnected. This means that quasi-axioms/presumptions and inference rules (principles) of a TT should be substantively compatible and accordingly elabourated. For example, if a theory is defined in its ontic part by a certain set of phylogenetic quasi-axioms, then its reference rules should also be phylogenetically interpreted to allow for the classifications to be elabourated as phylogenetically meaningful. This provision entails that if a classification is based on certain phenetic premises, which is usual for molecular studies, its posterior phylogenetic interpretation may appear problematic [14].
Thus interpreted, a TT does not comply with a “standard” theory, as it is understood by physicalist philosophy of science. It does not explain or predict specific facts about TD, but constitutes a part of the conceptual framework of a cognitive situation, in which biological systematics operates. In fact, TT appears as a product of the transformation of a conceptual framework, as it is construed in the general terms, into a more particular (partial) one adapted for the needs of biological systematics. Accordingly, TT can be defined as a coherent set of logically consistent and substantively compatible propositions about TD (ontology) and the principles of its study (epistemology).
3.2. General and Partial Theories in Systematics
As stated above, the basic content and structure of TT correspond to those of the conceptual framework of a cognitive situation, within which it is developed. This means that it is shaped hierarchically like the above-mentioned conceptual pyramid: its top is occupied by a general TT, while its lower levels correspond to the partial TTs of various levels of generality. An ontological component of general TT outlines TD as a whole with all its essential properties (structuredness, possible causality, etc.) defined by respective quasi-axioms/presumptions. Its epistemic component includes basic principles (inference rules), which principal task is to ensure, given the certain data sets, a possibility to “convert” basic ontic premises into particular classifications that would maximally correspond to the structure of TD. Respectively, the partial TTs are derived from the general TT by particular substantive interpretations of the latter’s initial premises.
According to the structure of the hierarchy of TTs thus understood, the general TT does not deal directly with the classificatory tasks, but represents a kind of metatheory that defines the possible ways of formulating and solving these tasks by the partial TTs. Accordingly, these are the latter that are elabourated as specific devices for investigating different manifestations of TD fixed by them. This implies that a general TT makes sense not in itself but as a means of elabourating specific partial TTs, which appear not by themselves but as the particular interpretations of the general TT (see also about reduction cascade above). In a more general sense, this metatheory performs two important functions in the cognitive situation of biological systematics. Its “external” function is to serve a kind of “bridge” between philosophy of science, on the one hand, and partial TTs most close to the empirical issues in systematics, on the other hand. Its “internal” function is to make different partial TTs mutually interpretable by reference to the same basic conceptual framework [12].
By detailing general TT in different ways, partial TTs split the entire conceptual space into isolated portions, each for a particular manifestation of TD fixed by a cognising subject. Keeping in mind the three-partitioned basic structure of that situation, it is reasonable to distinguish three main categories of partial TTs. The ontology-oriented ones define these manifestations by indicating certain essential features of TD; these are, for example, phylogenetic, biomorphological, typological, etc. theories. As far as the general TT outlines TD as a specific conceptual reality, the same function can be ascribed to the partial ontology-oriented TTs, each outlining a particular lower-order conceptual (sub)reality. The epistemology-oriented ones accentuate the specific means, by which these manifestations can be examined; examples are logical and numerical TTs. At last, in the subject-oriented partial TTs, an exceptional emphasis is on the cognitive facilities of the subjects of taxonomic research; for instance, a phenomenological theory of systematics accentuates the personal intuition [35].
The partial TTs of ontic kind can be divided into lower-level subcategories (kinds) according to the more “narrow” manifestations of BD individuated by respective quasi-axioms/presumptions. The aspect-based (aspectual) theories define different aspects of TD (taxonomic or partonomic, processual or structural, etc.) by fixing specific relations (structural similarity, kinship, etc.) between its elements. Taxonomic vs. partonomic partitions of TD represent its most fundamental aspectual accounts: the first involves groups of organisms, the second—groups of their properties (see [12,40,41] on this distinction). The object-based theories deal with various structural units identified within each aspect of TD, which are organisms, particular taxa, partons, and perhaps taxonomic/partonomic ranks in the overall hierarchy of TD. They are mutually complementary, and their interrelation is defined by a quasi-axiom of taxonomic-partonomic correspondence, which means that the taxonomic and partonomic aspects of TD are reflected onto each other in a certain regular, though a fuzzy way [12]. The relational theories consider fundamental properties of the between-elements relations, including their interconnections. At far lower levels of the overall conceptual hierarchy, partial TTs of each of these kinds can also be split by different criteria into certain groups. One of such criteria divides TTs into causal and phenomenological (descriptive) ones, and their contents are evident from their designations. Another criterion yields recognition of processual (evolutionarily interpreted), structural (phenetics, typology), and structural-functional (biomorphics, evo–devo) theories. At last, phylogenetics (in several versions) and biosystematics (in the sense of [42]) are recognised among evolutionarily interpreted theories that consider evolution at macro- and micro-levels, respectively.
Thus, the quasi-axiomatic method of construing a conceptual framework of biological systematics, briefly outlined herewith, allows for analysing explicitly the hierarchical structure of the entire conceptual framework. By this, it makes both the general and partial TTs transparent and comparable regarding their onto-epistemic foundations. This structure can be illustrated by a tree-like scheme that makes it evident how different TTs relate to each other within the general conceptual framework (Fig. 3).
Figure 3.
A tree-like scheme of the relationships among taxonomic theories (TTs) of various levels of generality. See text for the explanations concerning partial TTs.
Figure 3.
A tree-like scheme of the relationships among taxonomic theories (TTs) of various levels of generality. See text for the explanations concerning partial TTs.
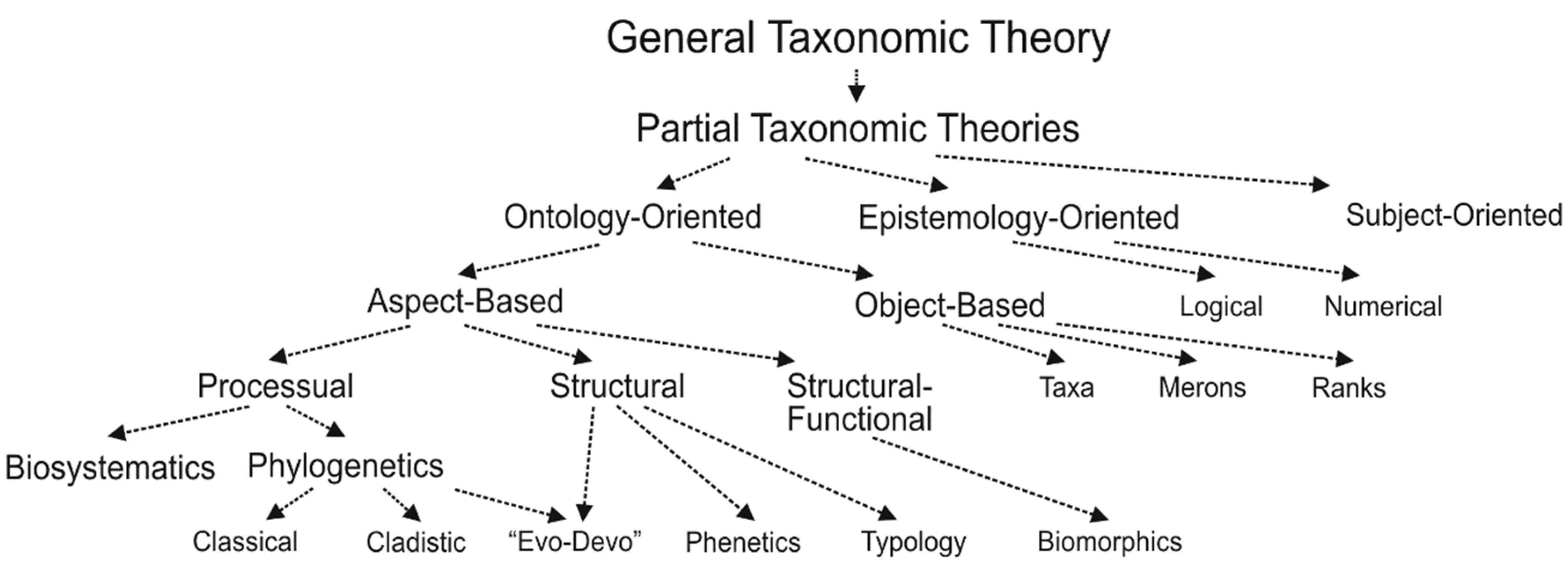
The ontology-oriented TTs specify first, by respective quasi-axioms/presumptions, particular manifestations of TD, such as monophyletic groups, archetypes, biomorphs, etc. recognised within either object- or aspect-based approaches. If these theories are construed as causal, they specify particular causes structuring BD by other quasi-axioms/presumptions—evolutionary (historical) in phylogenetics, structural in typology, functional in biomorphics, etc. If the evolution is adopted as the primary cause of BD, it can be interpreted broadly as an adaptatiogenesis or more narrowly as a cladogenesis, and this yields various evolutionarily interpreted TTs. Within each of the latter it is defined further, which kinds of relations between organisms are presumed—either kinship only or its certain combination with similarity. Based on these initial assumptions about ontology, certain taxonomic principles are developed by respective TTs as a part of their epistemologies. Given the evolutionarily interpreted partial TTs, some of their rules consider homologization criteria, character weighting, similarity assessment, etc., others suggest algorithms for inferring phylogenetic trees, and certain rules are designed for transforming similarity or kinship networks into concrete taxonomic classifications. Such general argumentation scheme is supposed to ensure that the resulting classifications adequately reflect the structure of TD presumed by the initial evolutionary models.
The epistemology-oriented TTs assume that the classificatory methods are unspecific with respect to the essential properties of various natural phenomena studied by systematics, so any supposed differences in these properties can and should be ignored as irrelevant. Due to this, the ontic premises are minimized to reflect only the most fundamental inherent properties of TD (e.g., axiom of existence), and the onto-epistemic correspondence is simplified accordingly. The primary task becomes an elabouration of certain universal, quite formal (ontology-independent) principles of the analysis of TD, and systematic studies governed by such TTs are designed as mere applications of these fairly formalized inference rules. Such automatization is supposed to produce an “objective” knowledge about TD, but in fact it is intersubjective (see above).
The aspect-based ontic and all epistemic partial TTs are of paramount importance in the structuring cognitive situation of systematics. They serve as conceptual bases for respective research programms implementing them (see Section 4), so some of them are sometimes called “systematic philosophies” [43,44,45,46,47]. The object-based and relational theories are not endowed with such high “philosophical” status, but in fact they are fundamentally significant: these theories interpret the objects and their interrelations that compose the entire TD.
The ontology-oriented partial TTs provide the empirical explorations of TD with a substantive (metaphysical) background knowledge, thus making them biologically meaningful. In this regard, it makes sense to emphasize two important provisions of the proper implication of a conceptualist argumentation scheme. First, according to the post-positivist philosophy of science, metaphysics-free scientific inferences are impossible even at an empirical level [48,49]. Second, such inferences (primary generalizations) are thought of as the “Popperian” hypotheses that are put forward and tested by observing certain rules. In systematics, the empirical generalizations take shape of classifications, so the latter are treated as specific taxonomic hypotheses. An important reservation should be made regarding the latter: the classifications-as-hypotheses are not about individual taxa, but rather about certain multi-taxon fragments of TD [11]. The adepts of a pure empirical treatment of classifications disagree with both these provisions, but their standpoint, as said above, gets systematics back to an initial folk stage of its history.
3.3. On the Definitions of Taxonomic Objects
The most fundamental problem being experienced by systematics is that its knowing activity involves reduction of the studied objective reality (BD) to a conceptual one (TD). The latter is given to a cognising subject by a coherent pool of notions and their definitions: they are “invented” by the very subject, but pretend to reflect in some way certain “properties of interest” of BD. They constitute a basic thesaurus of systematics that factually shapes the ontic component of its cognitive situation. Accordingly, the main issue in its conceptualization involves maximization of a possible correspondence between objective and conceptual realities by a proper construal of the respective thesaurus. However, as presumed by the provisions of ontological relativity specified by the principle of onto-epistemic correspondence (see above), the ontic content of a cognitive situation is robustly, though not evidently, controlled by certain epistemic regulators, which effects cannot be ignored.
One of these regulators is provided by the principle of constructivity [50] that suggests two basic conditions of the sought-for correspondence between objective and conceptual realities: it demands that (a) the thesaurus in question be sufficiently complete in its content and (b) the definitions of notions be sufficiently precise and rigor. These two provisions mean together that only those natural phenomena that are explicitly designated and properly defined can be considered meaningfully represented in a cognitive situation. However, the principle of the inverse relationship between rigor and meaningfulness of notions [50,51] asserts that the more strictly the latter are defined, the less likely there is something in “fuzzy” and “vague” Nature, to which they may exactly correspond. This is because the rigor of definition presumes its formalization, and the latter is the reverse of the richness of content. Such relationship is best exemplified by the mathematical objects: their notions are most precisely defined, but this makes them pure abstractions that are not supposed to relate to something “material” in Nature [52] (recall the above “spherical horse in vacuum”).
This inverse relationship is responsible for another inconspicuous and, with this, inevitable “epistemic pitfall” that puts certain limits to a possible rigor of the definitions of the notions in biological systematics. As far as such notions are context-relative, they can only be fuzzily defined, and this makes them metaphorical to an extent [12,53]. This means that such general notions as taxon, homologue, character, etc. are defined specifically in different conceptual (sub)spaces (“possible worlds” of S. Kripke [54]), therefore they cannot be regarded as rigid designators in the sense of S. Kripke.
With the conceptual (sub)spaces “fuzzily” construed, the fuzzy logic [55,56] and a probabilistic model of the natural science language actualizing it [57] seem to be the most appropriate instruments for elabourating respective linguistic TD pictures by inventing “semi-metaphorical” notions in systematics [11,12]. This means that it is reasonable to fix for each taxonomic notion more or less strictly only its “core” using logical relation “A is B,” while its periphery remains blurry, presuming multiple contextual interpretations by relation “A may be B1, B2, B3, ...”. As a result, each notion thus defined is represented by a probabilistic distribution of an array of its context-dependent interpretative definitions.
These considerations are clearly illustrated by a problematic issue with defining TD, which is evidently central to biological systematics, and yet it has no adequate definition. One of the currently popular ones defines it as an “array of organisms”; however, this is an extensional definition not specific to TD; it is more historically conditioned rather than theoretically outlined, and it is equally applied to any of the manifestations of BD. To develop consistently the notion of TD, we are to begin with taking the latter for a specific conceptual reality, so it cannot be defined otherwise than within a certain higher-order substantial theory (see on conceptual pyramid above). This may be, for instance, a theory of multifaceted structure of biota that can be roughly described by a set of quasi-axioms as follows [12,25]. First, biota is defined as a self-developing non-equilibrium system that is being structured as it develops (evolves) and functions. Second, it is assumed that its structure is determined non-randomly by the actions and interactions of various natural causes. Lastly, it is assumed that the references to these causes make it possible to individuate naturally particular manifestations (aspects, fragments, levels, etc.) of this structure. With this, as far as such causes are conjectural, recognition of the manifestations in question is based factually on specific features and relations of organisms and their aggregations.
Different manifestations of the biotic structure can be fixed (individualized) in two above-mentioned ways, either as objects or aspects. In the first case, particular natural phenomena can be thought of as the real entities existing outside any cognitive situation: examples are organisms, populations, ecosystems, as well as (with some reservations) levels of hierarchical organization of biota. In the second case, these manifestations do not exist on their own as independent natural phenomena; rather, they are epiphenomena individuated by a cognising subject by means of certain concepts. So they all are (let me remind this point once more) conceptual realities, “existing” within the frameworks of certain cognitive situations [12]. What is important is that the references to the supposed natural causes structuring biota can be taken for justification of the naturalness of its aspects being recognised in this way. These are the taxonomic, biogeographic, ecological, etc. aspects of the global structure of biota, which are studied by different biological disciplines.
One of them is TD studied by biological systematics: considered conceptually, it is thought of as aspectually defined by fixing certain relationships among organisms, namely structural (by similarity), historical (by origin), partly autecological, etc. Following our argumentation scheme, it seems reasonable to treat these relationships as the presumed causal natural agents that structure the very TD and individuate particular aspects in it to be fixed by respective partial TTs (see above). With this, the diverse nature of these causal agents yields a fundamental problem with defining TD itself. They appear to be too different to be unified by a certain single higher-level “taxonomic” relationship that would allow for defining TD intensionally and uniformly.
If it is so, then a pretty odd conclusion may be drawn from this consideration: in fact, biological systematics deals with several more or less independent conceptual realities conditioned by essentially different natural mechanisms. Thus, TD turns out to be but an “artificial” conglomerate of these aspectual realities, and the only reason to regard it somehow integrated seems to be a historical legacy. However, there still remains a possibility to define TD both intensionally and “naturally” by appealing to a complex of those diverse causes with reference to the principle of complementarity, which has been formulated in quantum physics and was given afterwards a more broad epistemic meaning [58]. By this, the latter principle appears to be a quite important element of the conceptual framework of contemporary systematics, belonging to the epistemic component of its cognitive situation.
4. Implementation of The Aspect-Based Taxonomic Theories: Research Programms in Systematics
A conceptual framework that “envelopes” biological systematics as a natural science branch can be thought of as a self-developing non-equilibrium information macrosystem [59]. According to one of the basic tenets of synergetics that explores behavior of such systems [60,61], such macrosystems, while developing, immanently differentiates into particular conceptual subsystems with specific theories describing particular manifestations of complexly organized and structured Nature; in systematics these are typology, phylogenetics, etc. Considered conceptually, it is the complexity of Nature—more specifically, BD in our case—that makes impossible comprehension of the latter by a certain unified and therefore the only cognitive model, constituting an all-encompassing “theory of everything”. So, it is this complexity that causes inevitable differentiation of any natural science branch, including systematics, in partial theories, each dealing with a particular manifestation of Nature. This Section considers how this multiplicity of partial theories is implemented in the knowing activity by means of research programms in systematics.
These programms emerge as an outcome of the just-above differentiation of a conceptual framework, and they constitute one of the basic elements of the structure of any cognitive situation, in which a scientific discipline (including systematics) functions and develops [62]. Considered conceptually, such programm functions as an implementation of a partial theory intended to explore a certain natural phenomenon (and eventually epiphenomenon). Being an element of a cognitive situation, it strongly depends on the latter’s subjective component, as it is a particular scientific community that adopts this theory for granted and develops it accordingly. This makes the research programm not so much a purely scientific as also a social phenomenon that is regulated by certain social mechanisms of the actions and interactions of research communities [59].
Regarding biological systematics, three principal research programms, usually called “systematic philosophies”, became acknowledged and most vividly discussed in it since the 1970s, namely, phenetics, cladistics (incorrectly identified with the whole of phylogenetics), and evolutionary taxonomy [43,44,45,46,47]. However, a comprehensive analysis of the diversity of the partial TTs across the whole of biological systematics indicates that such a “three philosophies” viewpoint is based on an oversimplified perception of its theoretical foundations and significantly reduces the real multiplicity of the research programms operating in it [12,63]. This standpoint yields a very distorted representation of the cognitive situation, in which our scientific discipline develops and functions. This prevents a proper comprehension of the real conceptual content of biological systematics, including the historical and scientific-philosophical roots of its research programms, their mutual interactions and influences, and eventually their contribution to the development of both systematics itself and overall biology.
One important feature of the research programms in systematics is that they implement mainly ontology-oriented aspect-based partial TTs, whereas object-based partial TTs are not apparently involved in their individuating (see the next Section). This is because these are the aspect-based partial TTs, just like the very general TT, that deal with the particular conceptual (sub)realities as concrete cognised objects. Thus, the conceptual history of biological systematics may be thought of as the development of research programms following a growing comprehension of a complicated nature of the overall BD, outlined basically by an array of aspect-based partial TTs. Regarding epistemology-oriented TTs, they also may be involved in the forming of research programms in systematics, but only as instrumental add-ons to the ontology-oriented ones. This means, among other things, that there cannot be something like “numerical programm” as such; in fact, numerical methodology in biological systematics is actualized by either phenetic or phyletic ontology-oriented TTs.
Contemporary evolutionary epistemology, supported by non-classical philosophy of science [3,48,64,65], is quite suggestive for an understanding of the historical dynamics of research programms in systematics, including their emergence and vanishing, their diversity, etc. [12]. One of its basic and very significant, though largely metaphoric, assumptions draws a parallel between the historical development of scientific disciplines and biological evolution. According to this metaphor, “conceptiations” represent the main events in the history of science, which are specific analogs of speciation events in the biological evolution. Respectively, two basic trends can be identified in the conceptual history of systematics thus interpreted: anagenesis corresponds to a sequential progression of the taxonomic knowledge from less to more comprehensive, and cladogenesis means its fragmentation into particular theories. Besides, a kind of reticular evolution means exchanges and combinations of the ideas among different theories.
The selecting “environment” for all of this conceptual evolution seems to be the philosophical and scientific ideas (empirical, evolutionary, numerical, etc.) that provide conceptual contexts framing the cognitive situation of systematics [12]. In addition, certain social phenomena should be mentioned, which are responsible for shaping its pragmatic context [59]. In this regard, a biological concept of adaptive zone might provide another suggestive metaphor: it presumes that various context-making ideas act as specific “adaptive zones” stimulating the development of systematics in one direction or another. Accordingly, taxonomic theories and research programms implementing them emerge in “due time” to fill in the new conceptual “adaptive zones” opened by new ideas; and they get “extinct”, when those ideas lose their relevance and respective “adaptive zones” get collapsed.
The most significant shifts between particular research programms were usually stipulated by the significant changes in the worldviews and/or basic methodologies, and they marked scientific revolutions in the conceptual history of biological systematics [12]. The first was the scholastic revolution caused by the mastering of the scholastic method of classifying in the 16th century, it shaped the scholastic research programm of the dawn systematics (Cesalpino, etc.). The next was the post-scholastic revolution of the late 18th and early 19th centuries that gave rise to an array of the research programms, namely natural systematics (Adanson, Jussieu, etc.), typology (Cuvier, Baer, Goethe), numerological (MacLeay), and organismic (Oken) ones. The second half of the 19th century experienced the evolutionary revolution in biology and systematics, and it produced two research programms, microevolutionary (Darwin) and phylogenetic (Haeckel). In the early 20th century, positivist philosophy of science became dominant, which resulted in the research programm of numerical phenetics (Sokal & Sneath, etc.). This philosophy lost much of its popularity since the mid-20th century, so it is possible to mark the most recent post-positivist revolution that caused a revitalization of phylogenetics, now in the guises of evolutionary taxonomy (Simpson) and cladistics (Hennig). The latter became most popular among systematicians for some time, and it was therefore designated as a “cladistic revolution” [65]. Now it got transformed into the so-called “new phylogenetics” that combined cladistic methodology, numerical technology, and molecular data [14]; its taxonomic implication was suggested to call genosytematics [67]. Along with this, a new research programm seems to emerge in nowadays systematics, combining Haeckelian phylogenetics and von Baerian epigenetic typology with the most recent developmental genetics to shape “evo–devo systematics” [12,68]; a special emphasizing its ontogenetic component is characteristic of one of its versions called “ontogenetic systematics” [69,70].
It would hardly make sense to provide here a review of the former and currently active research programms in systematics: this has been done comprehensively enough elsewhere [12,63]. Instead, it seems more reasonable to comment briefly on the recent specifics of realization of the dominating trend in the conceptual history of systematics, as the author sees it. This trend marks an anagenetic component of the conceptual history of systematics, and it involves what could be called biologization of this discipline. It sequentially transformed an early “formal” scholastic systematics into a biologically meaningful one by incorporating more and more biological mechanisms (ecological, evolutionary, etc.) in the understanding of the origin and structure of BD. This made systematics multifold and very reach in its biological content by the early 20th century.
However, positivist philosophy of science declared such a substantial interpretation to be a “redundant metaphysics” that should be removed from “real science”; the same was declared regarding any kind of personal knowledge. These declarations were first implemented in systematics by numerical phenetics with its automated classification procedures that resulted in the above-mentioned positivist revolution in systematics; though, given the general trend of biologization, this turn could be perhaps called a “counter-revolution”. The only thing it had then failed to accomplish was complete “elementarization” of characters and automatization of the handling them; however, molecular phylogenetics succeeded in this later. As said above, the latter was effectively incorporated by cladistic methodology with its oversimplification of the process of phylogeny, and all this turned phylogenetic reconstructions into operationalized automated procedures.
Such “instrumentalist” bias is very noticeable in contemporary systematics. Considered conceptually, this yields a fundamental problem of “counter-biologization” of the discipline under consideration. Indeed, such a bias actually moves the latter in a direction that falls out of the “anagenetic” trend dominating so far in its conceptual history. So, it is my feeling that this most recent reductionist development represents something like a “last breath” of positivist philosophy in systematics. In this regard, the above-mentioned research programm of “evo–devo systematics” seems to be the most promising candidate for returning the whole of systematics to its predominant biologization trend to make it a full-fledged biological discipline [12,71,72].
5. Implementation of The Object-Based Taxonomic Theories: The Concepts of Species and Homology as Case Studies
An important idea of the overall structure of general TTs presumes its fragmentation into aspect-based and object-based partial TTs fuzzily tied by a taxonomic-partonomic correspondence. As said above, the former TTs constitute the conceptual bases for the research programms in systematics, whereas the latter do not; therefore, the former are usually considered “systematics philosophies”, whereas the latter are not. With this, however, the aspect-based TTs have no meaning in themselves: in fact, they deal with certain elements of the structure of TD that are defined by the object-based TTs. Accordingly, with recognizing the research programms as the basic immanent elements of the structure of cognitive situation in systematics as a natural science branch, the latter’s conceptualization should involve correct interpretation of interrelationships between these two categories of partial TTs. In turn, this interpretation implies a prior consideration of the object-based TTs in the same manner as the aspect-based ones, that is, how they are implemented in the cognitive situation under consideration. Such a consideration is briefly provided in this Section.
Let us agree at this initial point that the object-based TTs are implemented by respective concepts that function and develop similarly to the research programms. Among the elements of TD structure defined by them, species and homologues are of paramount importance for systematics, so they are proper to briefly consider here as specific case studies. They are recognised within two basic aspects of TD, taxonomic and partonomic, respectively (see above on them). According to this aspectual distinction, species are taxa corresponding to the groups of organisms, whereas homologues are partons corresponding to the complexes of the properties of these organisms. The concepts implementing the underlying partial TTs are treated very controversially, yielding much of uncertainties in all branches of biology dealing with the diversity of organisms and their features. This is reflected in the persistence of the well-known problems of species and homology, discussed in the hundreds of articles and dozens of books. What is of important to our consideration is that their correspondence supposes a certain similarity of their conceptual framing, so they both can be subject to a similar conceptual analysis.
The history of the notion of species was exposed in two recently published books [25,26]. This notion emerged (as an eidos) in the Antique natural philosophy (Aristotle, Boëthius, etc.) to designate (in our terms) both taxonomic and partonomic elements of the diversity of Nature. This universal dual understanding of species became inherited by Medieval natural philosophy, and from the latter by the dawn scholastic systematics. Its basic conceptualization was then based on the Old Testament ontology, according to which “Natura est lex Dei” (Linnaeus), and on the scholastic epistemology with its genus-species deductive argumentation scheme.
A fundamental conceptual shift from a universal dual understanding of species to a biological singular one became a part of the post-scholastic revolution in systematics: its partonomic treatment got discarded in favour of the taxonomic one only. One of the most important premises of this revolution was an acknowledgment of living matter as essentially different in its properties from inert matter, so the species category, in the case of animals and plants, was defined on a solid biological basis, i.e., on the breeding system (Buffon). The next was the evolutionary revolution that suggested a characteristic natural mechanism of the origin of species—not by Divine creation, but by “descent with modification” (Darwinian formula). Such treatment presumed a gradual transformation of local geographic races into species, so it questioned the reality of species as a natural biological unit and yielded the so-called species question.
The recent step in the conceptual development of the notion in question involved two key ideas: (a) species is a fundamental unit of both the biological evolution and, hence, the structure of BD resulting from it, and (b) its essential properties are associated with some biological features of organisms, one of which is their breading systems. The conceptualization of such kind meant a further biologization of the notion of species and made it a unique biological entity. But with this, the second provision gave birth to the so-called species problem that questioned the conceptual universality of the notion in question for all living beings. The emergence and subsequent development of this problem gave rise to an array of the particular species conceptions, and their multiplicity became one of the vividly discussed issues in the contemporary theoretical biology (e.g., [24,25,38,73,74,75,76,77]). The attempts of arranging a chaotic diversity of the particular treatments of species into certain ordered conceptual systems resulted in an idea of their hierarchy according to their levels of generality [25,78,79], and this seems to be a special case of the above-mentioned conceptual pyramid.
The recent reviews of the general concept of homology showed, just as in the case of species, the impossibility of defining it in a unified way [80,81,82,83]. This notion has come into biology first in its idealistic typological interpretations, in which the notion of archetype was central (Goethe, Owen). An essential part of Owen’s concept was distinguishing between homology and analogy, and he additionally divided the former into general, special, serial, etc. forms. Their evolutionary interpretation (Haeckel, Ray Lancaster) became the next significant move towards the contemporary understanding of homology by distinguishing between homophyly/homogeny (inherited from the close ancestor) and homoplasy (resulting from a parallel evolution). Simultaneously, a conception of ontogenetic homology was introduced (Mivart) based on von Baerian epigenetic theory. A more profound comprehension of the basic manifestations of homology provided its structural and transformational conceptions (Naef, etc.), with the latter giving rise subsequently to the process-homology (Hall, etc.). As a result, the number of conceptions of homology raised up to several dozens by the end of the 20th century, so the homology problem emerged similar to that of species, and it was declared “an unsolved problem in biology” [84]. Cladistics made its own contribution to the overall confusion with the conceptions and terms by identifying homology with homophyly/homogeny (Hennig) and thus narrowing the former drastically. The molecular biologists also put their “two cents” into this mess of opinions by adding several special terms (Hillis, etc.). The “evo–devo” approach now emphasizes genetic understanding of homology with reference to the regulator genes (Hox, MADS, etc.) (Holland, Shatalkin, etc.). The attempts to construe some unifying conceptual frames for the plurality of the homology conceptions have led to the same hierarchical idea [85,86,87], as in the case of the species problem.
As easily seen, the two considered object-based partial TTs, within which the concepts of species and homology have been elabourating for centuries, comply with the same general trends in their conceptualization as the aspect-based partial TTs. They both were developing towards their biologization, with the latter yielding complication of the ontic components of their respective conceptual frameworks. This caused fragmentation of these basic concepts into lower-level partial conceptions, and the resulting array of the latter can be ordered in respective conceptual pyramids. By this, the object-based TTs and the concepts implementing them, in parallel to the object-based TTs with their research programs, participate in the structuring of overall conceptual framework of biological systematics in their specific ways [12].
6. Taxonomic Monism vs. Pluralism
As was said in the very beginning of this article, an imminent multiplicity of the ways of posing and answering the questions concerning onto-epistemic foundations of scientific knowledge yields a controversy between their either monistic or pluralistic accounts. This controversy demarcates classical and non-classical philosophies of science [2,3,] that were shown above to be quite relevant to contemporary biological systematics [11,12,63]. Accordingly, an important part of its conceptualization involves consideration of an issue of the relationships between monistic vs. pluralistic treatments of its conceptual framework, which is briefly overviewed in this Section.
The classical philosophy of science is largely monistic. Roughly speaking, it acknowledges that Nature in all its manifestations is organized by a unified global cause, which is a universal Law of Nature. This presumes the unity of the entirety of scientific knowledge in its basic ontic and epistemic premises, and this knowledge should be “objective” and “unified”. The first idea means that it should reflect Nature in its wholeness, which could be achieved, in a company with other matters, by minimization of any subjective impact on both the onto-epistemic foundations and, hence, the results of knowing. The second idea means that the scientific knowledge should be expressed by a single “formula”, representing a “general theory of everything”. Regarded historically, this monistic account is rooted in the Antique and Old Testament onto-epistemologies. They both acknowledge that Nature is ruled by a single major cause, be it the Aristotle’s prime mover or the Biblical plan of creation. With Christianity having become dominant in Europe, this religious monism passed from natural theology into natural science to turn into philosophical-scientific monism [88,89]. Paradoxically enough, although the modern development of natural science largely excludes the Biblical world picture from its ontic background, its basic epistemology preserves a monistic account of scientific knowledge. Accordingly, it is believed that the human knowing steadily moves towards the only possible true knowledge expressed by a certain “final theory” that is considered an ideal of the whole of natural science [90].
The non-classical philosophy of science is pluralistic in its understanding of how both Nature and knowledge about it are arranged. It is acknowledged that Nature is quite complex, and its general organizing principle represents a combination of different causes, local in their actions and not reducible to some unified “cause of everything”. The non-coinciding actions of such causes “decompose” Nature into various particular manifestations—aspects, fragments, levels, etc., each with its own essential properties. Accordingly, this complexly organized Nature can be comprehended by different “local” approaches elabourated especially for its “local” manifestations to produce particular “local” theories about them. Further, an active role of a cognising subject, who “construes” a cognitive situation by certain onto-epistemic premises and elabourates partial theories to individuate and analyse those “local” manifestations of Nature as particular conceptual realities, is acknowledged. Accordingly, scientific knowledge is not theory-free and subject-independent from the very beginning, and it represents a totality of “local” theories, each with its own particular onto-epistemic foundations, methods, and the ways of representation of the results of knowing. However, such scientific pluralism, contrary to a widespread misconception, neither implies the denial of an attainable truth nor it allows for “anything goes”. It means that there is no classical “universal theory of everything” with its “unified truth”, but there is a certain array of “local” theories with their corresponding “local truths”, and the latter in total compound a non-classical “multiple truth” according to the above-mentioned principle of complementarity [58].
A dichotomy of classical vs. non-classical philosophies of science, yielding the controversy of scientific monism vs. pluralism, is evidently reflected in biological systematics [12,63]. Its classical version professes taxonomic monism, according to which (a) the above-mentioned universal Law is given in the form of the single System of Nature and (b) there is the only one Natural Method that allows for comprehending it. Indeed, for C. Linnaeus, “Natura est lex Dei” [91, p. 546], and for M. Adanson, a sought-for method “should be universal, or unified, that is, there should be no exceptions for it” [92, p. civ]. This monistic idea dominated throughout the conceptual history of systematics, and it remains popular nowadays, regardless of the fundamental disagreements about how natural classification and natural method should be understood. So, by a pretension to possessing a certain “universal method” leading to a certain “final theory” of systematics, the contemporary numerical pheneticists or cladists hardly differ principally from those systematicians who have searched for the God-given System of Nature in the 18th century.
In its absolute form, this monistic account is most fully manifested in the conviction that there is only one possible way to “carve Nature at its joints” (Plato’s formula) to uncover really “natural” units of diversity [93], and this presumes that there can be only one really natural classification [94]. Moderate monism admits a multiplicity of different partial TTs, but only one of them can and should be ascribed a priority status as “true”. Consequently, there can be many particular classifications, but only one of them is to be considered a priority “general reference” one [31,95]. Quite naturally and expectedly, the ideologists of each partial TT (phenetic, phylogenetic, biosystematic, etc.) attribute such a priority status to the one that they favour.
The non-classical systematics acknowledges taxonomic pluralism that admits a plurality of partial TTs, dealing with particular manifestations of TD [12,63]. Accordingly, they all are ascribed an equal cognitive significance, provided that they are defined by reference to the particular natural causes structuring biota (see above on this). It is this pluralism that yields subsistence of the particular research programms in systematics that implement these TTs. At an ontological level, such a standpoint is justified by acknowledging an irreducible multidimensionality of TD underlying its conceptual “decomposing” into various manifestations. Based on the presumed essential properties of the latter, partial TTs develop both the specific criteria of naturalness of sought-for classifications and the natural methods for construing them. It is to be remembered that a pluralistic account of TD is strongly supported by the above-outlined ideas of reduction cascade and conceptual pyramid that make its monistic account evidently untenable. Among other things, taxonomic pluralism supposes that different lower-order partial TTs may be admissible for the particular groups of organisms according to the specifics of their biological features, including the structure of their TD. This makes a non-classical systematics “biological” in its full sense.
The taxonomic pluralism quite evidently manifests itself in the problems of species and homology discussed in brief above. The so-called “species pluralism” is especially significant in this regard, as the notion of species is sturdily embedded in the thesauruses of many fundamental and applied biological disciplines. Therefore, the inability to reach a uniform understanding and eventually definition of species yields an undesirable uncertainty in the knowledge of the structure of BD. Given this uncertainty, its critics suggest replacing the classical notion of species with some other, mere operational ones lacking a redundant historical-conceptual burden [96,97,98,99]. An extreme expression of such an attitude involves rejection of the species notion altogether that yields another conceptual revolution in systematics by making it, and with it the whole of biology, “speciesless” or “post-species” [100,101,102,103]. However, the so-called “new essentialism” seems to provide a strong argument in favour of considering species a fundamental unit of the diversity of living matter. It is presumed to be endowed with a certain characteristic quiddity informally called specieshood, which is thought of as an integral part of the overall natural history of organisms, so this quiddity evolves together with their particular natural histories and is plural in its manifestations in various groups according to their specific biological organizations [25,104,105,106].
One of the fundamental problems raised with scientific pluralism is the incommensurability of theories elabourated within different conceptual spaces [107,108,109]. They divide an overall cognitive situation of biological systematics into isolated conceptual fragments, making it a kind of patchwork [110], and such fragmentation yields an aspiration to attain a “controlled and manageable” pluralism [111]. To minimize a disagreeable effect of an “uncontrolled pluralism”, it is considered desirable to develop some conceptual translators for the mutual interpretation of the partial TTs that operate with different ontic premises [112,113]. These translators constitute a part of metatheoretical knowledge about multifaceted TD, which once again brings us to acknowledging the fundamental significance of the conceptual pyramid. Another way of ordering the plurality of partial TTs in a united system of the knowledge about TD without eliminating any of them may involve elabouration of something like faceted classification that is organized similarly to the relational databases [114,115]. It would make it possible to combine all “locally natural” particular classifications into a single pool of a faceted classification system that would serve as a “globally natural” representation of overall TD [12].
Thus, the search for an appropriate methodology of elabourating such faceted classification for an array of the partial TTs constitutes an important task of construing of a solid conceptual framework of basically pluralistic contemporary systematics.
7. An All-Encompassing Conceptualization: Taxonomic Theory and Nomenclature
According to the provisions of cognitive linguistics, it is admissible to treat the specific knowledge developed by a scientific discipline as a text written in its professional language [116]. This general consideration seems to be relevant to the professional language of biological systematics, with which it describes the studied TD by elaborating respective taxonomic classifications treated, in a certain simplified account, as ordered sets of the names of taxa [117]. Supposing that a consistently developed conceptualization of systematics is to be all-encompassing by involving all and any of its branches, one of the important tasks should include an analysis of relationships between its TT and its descriptive language. A starting point in this analysis is a presumption that, as far as taxonomic knowledge is conceptually framed and therefore theory-dependent, it is reasonable to suppose that the latter is true for the language of taxonomic descriptions. This starting point is specified and the outcomes of its adoption are considered in the present Section.
An essential part of the professional language of systematics is constituted by nomenclature as an array of the principles and rules designed for handling taxonomic names and summarized by Codes. The nomenclature developed by systematics is usually called biological, but its designation as taxonomic seems more appropriate, as there are several other nomenclatural systems that are parts of the languages of other biological disciplines (biogeography, anatomy, biocaenology, etc.). Most systematic textbooks and manuals in zoology, botany, and microbiology discuss the clauses of their respective nomenclatural Codes, which reflects a great attention paid to the taxonomic nomenclature by systematicians. A discipline that studies nomenclature, including its theoretical foundations, was proposed to recognise under the name onymology or taxonymy [118,119,120]. One of the latter’s key issues is a relation between taxonomic theory and taxonomic nomenclature.
The most of the contemporary Codes expressly emphasize their pure “empirical”, theory-free status, legitimized by the principle of taxonomic freedom. This means that these Codes consider only designations of taxa and ranks and do not regulate concrete taxonomic decisions about them, and more so their theoretical foundations. Conceptually considered, the principle in question indirectly supposes that taxonomic nomenclature is free of any effects of whatever TTs that may underlie such decisions. However, this does not correspond to the actual state of things: in fact, the professional language of biological systematics depends, to a more or less degree and in various ways, on the latter’s conceptual framework [119].
One of the key ideas of the above-mentioned cognitive linguistics is that the structure of a language designed to describe cognised Nature is to be adequate to the latter’s own structure [121,122,123,124]. It is this adequacy that ensures a certain compliance of our descriptions with what we intend to describe. This is a particular manifestation of the above-considered onto-epistemic correspondence, and it provides a specific linguistic world picture that expresses an overall structure of cognised Nature by characteristic linguistic means [122,123,125,126]. In this manner, classifications are admissible to think of as certain “linguistic pictures” of TD studied by systematics [119,127].
As emphasized above, TD is not a part of Nature as objective reality, but it is a conceptual reality outlined by means of TT. So, it is this theory-dependent conceptual reality that is reflected in a particular linguistic world picture. Detailing further this idea, we come to the aspect-based partial TTs, defining certain manifestations of TD as specific taxonomic (sub)realities. Linking cognitive ideas about the latter and linguistic world pictures corresponding to them, one may conclude that each of the manifestations of TD, thus outlined, requires a specific descriptive language advisably most adequate to its structure. As far as these (sub)realities are defined within the framework of partial TTs, the nomenclatural systems, being the parts of respective descriptive languages, seem to indirectly depend on the contents of these theories [119]. So, an accepted view that taxonomic nomenclature is “neutral” regarding any TT [120,128,129] has by no means a serious theoretical justification. As a result, a universal language of taxonomic descriptions becomes fragmented into separate “dialects”, each designed to match as closely as possible a particular taxonomic (sub)reality by developing and applying a specific nomenclatural system.
A large-scale link between taxonomic theory and nomenclature is responsible for their coherent changes in the above-mentioned macro-events of the conceptual history of systematics [119]. This is evident from the below consideration.
Scholastic systematics of the 16th–18th centuries had emerged through the mastering of an essentialist world view of neo-Platonic kind, to which an essentialist nomenclature mostly fit. According to this idea, the names denoting organisms (or rather taxa) should reflect their essences. The French botanist J. Pitton de Tournefort argued that “an idea of character, essentially distinguishing some plants from others, should be invariably associated with the name of each plant” [130, p. 2], and C. Linnaeus later echoed him by stating that “systematic names ought to be essential ones” [131, § 252].
The post-scholastic revolution in systematics at the end of the 18th century has involved significant changes both in the onto-epistemic foundations of systematics and its language. It has led to a transition from essentialist to basically nominalist nomenclature: the names ceased to express the essences of organisms and became simply their “labels” [92,132]. This fundamentally new interpretation of taxonomic nomenclature resulted in a significant simplification of the rules for the formation of taxonomic names.
In the second half of the 19th century, the evolutionary revolution in systematics has led to the recognition of a fundamental importance of infraspecific units, and this gave way to the suggestions to replace the then acknowledged binomial nomenclature with a trinomial one [133,134]. The positivist revolution in systematics of the first half of the 20th century activated some earlier ideas of the rational nomenclature that resulted in the proposal of the so-called “numericlature” [135,136]. Finally, contemporary phylogenetics (cladistics), with declaring its theory to be the most recent revolution in systematics, found it necessary to replace the well-established traditional nomenclature with a new phylogenetic one [137,138,139].
8. Some Considerations in Conclusion: Quo Vadis?
Contemporary biological systematics is by and large treated as being in a state of so-called taxonomic impediment, with the latter’s causes usually discussed at an empirical level, involving the shortages of experts, data, collection curation, financial support, etc. [71,140,141,142]. What is not taken into account is that one of the most significant “hidden” causes of the impediment of systematics is that it is underestimated as a scientific discipline. Such its perception arose more than a hundred years ago with now obsolete positivist philosophy of science, according to which systematics, as a part of natural history, is not a science, but “stamp collection” (see above). Unfortunately, this attitude still reigns in the minds of many leading scientists, philosophers, and decision makers, who are guided by this viewpoint in their selections of the particular departments of natural science to be primarily supported. Therefore, it is hardly surprising that systematics (or taxonomy) is not even mentioned in the basic “List of Life Sciences” [143], although no one of the branches of life science listed in it can do normally without certain results of taxonomic research.
Inspired by positivist philosophy of science, researchers usually believe that contemporary progress in systematics is caused principally by the development of new methods (before all, numerical) and involving new kinds of data (before all, molecular). As a matter of fact, this provides an extraordinary possibility of comparison of all living beings, from microbes to higher plants and animals, on the same standardized methodological and empirical grounds. Due to this, an elabouration of an overall Tree of Life becomes currently possible [144], which may be considered a realization of a centuries-old dream of many philosophically-minded naturalists. However, no matter how significant the success with this molecular tree, it should not be overestimated as an unconditional progress in systematics. Actually, such a tree provides a biased overall picture of TD due to what could be aphoristically called a “microbial worldview”. The most serious problem with the latter is that it does not integrate, for quite a clear reason, those important features of the multicellular organisms, before all their ontogeny, that are not inherent to the prokaryotes. Because of this, the classifications based on this molecular tree do not incorporate the significant differences among multicellulates in their ontogenetic patterns. Meanwhile, emergence of ontogenesis in the evolution of life is one of the latter’s very characteristic features, fundamentally distinguishing (a) living from inert matter and (b) unicellular from multicellular organisms [145]. So, one may question whether such development of contemporary systematics, ignoring one of the most fundamental properties of the living, can actually be treated as a whatsoever “progress”.
As emphasized at the very beginning of this article, the normal development and functioning of biological systematics as a natural science branch cannot proceed without consistent elabouration of its own biologically sound theoretical foundations. Considering the just-above impediment from a conceptualist standpoint, one can suppose that the primary cause of a problematic condition of contemporary systematics seems to be a strong disproportion in the structure of its cognitive situation. It manifests itself in the predominant “technologization” of the current taxonomic research, including its “numericalization” and “molecularization”.
This yields a specific “conceptual” taxonomic impediment that involves “deontologization” and “dehumanization” of systematics, transforming it from a full-fledged natural science branch into a specific technical device for handling bioinformatic resources. This bias is evident from the contemporary theoretical publications on systematics that are devoted mostly to the methodological and methodical issues. Curiously enough, another “new systematics” was recently proclaimed that was stimulated by the new promising possibilities of the computer processing of “big data” [146]. By this, the cognitive situation, in which contemporary systematics develops and functions, loses both the biological content of its ontic component and the creative potential of its subjective component. As a consequence, today's “technological” systematics risks downshifting to the very bottom of the reduction cascade of all of biology to become its “Cinderella” again.
More concretely, the problematic condition of contemporary biological systematics involves a contradiction between most reductionist “new phylogenetics” and most biologically sound “evo–devo”. This is caused basically by the above-mentioned biological nature of microbes making “evo–devo” concept seemingly inapplicable to them. This contradiction promises to continue in the near future to become one of the most serious factors of choosing among possible pathways in the progress of systematics. So, the principal question of the latter’s contemporary conceptual history is of a “to be or not to be” kind: whether systematics will follow the general “anagenetic” trend of its biologization, or will it develop as just a technical device for constructing a kind of “molecular skeleton” without “biological flesh”.
Accordingly, a consistent elabouration of the conceptual framework of contemporary biological systematics, including a profound substantiation of its biologically sound ontology, becomes an urgent task for its proper development aimed at weakening its “theoretical impediment” and making it “great again”. Remaining within a conceptually framed cognitive situation, it is reasonable to highlight three fundamental issues that will impact on the immediate future of our discipline.
One of them involves the concretization of general ideas about construing a conceptual framework of systematics as a quasi-axiomatics by elabouration of respective pools of quasi-axioms/presumptions and inference rules for both the general and partial TTs. The conceptual history of mathematics indicates that this is not an easy task, requiring a lot of effort and time [52]. However, a persevering movement in this direction seems to be worthwhile, as it promises to drive biological systematics towards the status of a respected full-fledged natural science branch.
Another issue, closely related to the previous one, concerns a more profound elabouration of the onto-epistemic background of the cognitive situation of systematics. This would involve the development of a biologically sound metatheory as a basis for construing a conceptual pyramid of the ontology-oriented partial TTs of various levels of generality. Its metalanguage would hopefully make both these theories and particular classifications conjectured with them mutually interpretable. With this, such a pyramid of partial TTs would positively resolve the “monism vs. pluralism” problem by turning an “uncontrolled pluralism” into a “moderate” one.
Lastly, a currently arisen controversy between microbial and multicellular “worldviews” due to the incommensurability of some of their fundamental biological features is waiting for its meaningful decision. Operationally, this controversy is a result of incompatibility of two fundamentally different kinds of characters in construing an overall System of Living Nature. One of them includes molecular sequences, which diversity provides a great resolution of the branching of the Tree of Life at its microbial root. Another includes ontogenetic patterns, which diversity is relevant to an analysis of the branching of the same Tree at its crown encompassing plants and animals. Accordingly, this aim could be achieved by elabouration of a biologically meaningful system of “weighing” of those two kinds of characters to make commensurable both themselves and the ranks of groups identified by them.
Funding
This research was supported by The Governmental Theme No. AAAA-A16-116021660077-3.3, implemented by The Research Zoological Museum at Lomonosov Moscow State University.
Acknowledgments
Ms. Joey Jin (MDPI Philosophies Editorial Office) is acknowledged for her having invited me to contribute to The Philosophies with this article. I am highly indebted to Drs Catherine Kendig (Michigan State University), Alexander Pozdnyakov (Institute of Animal Systematics and Ecology), and Maxim Vinarski (Saint-Petersburg State University) for their useful comments on an early draft of the article, as well as to my friend, Prof. Douglas Wiener (University of Arizona) for his improving my English.
Conflicts of Interest
The author declares no conflict of interest.
References
- Heisenberg, W. Physics and philosophy. The revolution in modern science; London: George Allen & Unwin, 1959. [Google Scholar]
- Stepin, V.S. Theoretical knowledge; Springer: Heidelberg, 2005. [Google Scholar]
- Johnson, K. Natural history as stamp collecting: A brief history. Arch. Nat. Hist. 2007, 34, 244–258. [Google Scholar] [CrossRef]
- Rosenberg, A. The structure of biological science. Cambridge Univ. Press: New York, US, 1985.
- Simpson, G.G. Principles of Animal Taxonomy; Columbia Univ. Press: New York, US, 1961. [Google Scholar]
- Wheeler, Q. A taxonomic renaissance in three acts. Megataxa 2020, 1, 4–8. [Google Scholar] [CrossRef]
- Swoyer, C. Conceptualism. In Universals, Concepts, and Qualities: New Essays on the Meaning of Predicates; Trawson, E.S., Chakrabarti, A., Eds.; CRC Press: Routledge, UK, 2006; pp. 127–154. [Google Scholar]
- Stroud, B. The cahrm of naturalism. In Naturalism in question; De Caro, M., Macarthus, D., Eds.; Harvard Univ. Press: Cambridge, US, 2004; pp. 21–35. [Google Scholar]
- Wartofsky, M.W. Models: Representation and Scientific Understanding; Reidel: Dordrecht, The Netherlands, 1979. [Google Scholar]
- Contessa, G. Scientific models and representation. In The Bloomsbury Companion to the Philosophy of Science ; French, S., Saatsi, J., Eds.; Bloomsbury Acad.: New York, US, 2011; pp. 120–137. [Google Scholar]
- Pavlinov, I.Ya. [How it is possible to build a taxonomic theory?]. Zool. Issled. 2011, 10, 45–100. (in Russian).
- Pavlinov, I.Ya. Biological Systematics. History and Theory; CRC Press: Boca Raton, US, 2021. [Google Scholar]
- Sneath, R.H.A.; Sokal, R.R. Numercial Taxonomy. The Principles and Methods of Numerical Classification; W.H. Freeman & Co.: San Francisco, US, 1973. [Google Scholar]
- Pavlinov, I.Ya. [Introduction to Contemporary Phylogenetics]; KMK Sci. Press: Moscow, RF, 2005. (In Russian) [Google Scholar]
- Smaling, A. Varieties of methodological intersubjectivity: The relations with qualitative and quantitative research and with objectivity. Qual. Quant. 1992, 26, 169–180. [Google Scholar] [CrossRef]
- Rooney, P. On Values in science: Is the epistemic/non-epistemic distinction useful? Proc. Bienn. Meet. Phil. Sci. Assoc., vol. 1; Univ. Chicago Press: Chicago, US, 1992; pp. 13–22. [Google Scholar]
- Elliott, K.C.; McKaughan, D.J. Nonepistemic values and the multiple goals of science. Phil. Sci. 2014, 81, 1–21. [Google Scholar] [CrossRef]
- Quine, W.V. Ontological Relativity & Other Essays; Columbia Univ. Press: New York, US, 1969. [Google Scholar]
- Botha, M.E. Theory development in perspective: The role of conceptual frameworks and models in theory development. J. Adv. Nurs. 1989, 14, 49–55. [Google Scholar] [CrossRef]
- Gärdenfors, P. Conceptual spaces as a framework for knowledge representation. Mind & Matter 2004, 2, 9–27.
- de Castro, M.A.B.; Some Antecedents of Leibniz’s Principles, Thes. DrPh; Univ. New South Wales, Sydney, AU, 2008. Available online: https://philarchive.org/archive/DECSAO-4 (accessed on 14 April, 2023).
- Dupré, J. The disorder of things. Metaphysical foundations of the disunity of science; Harvard Univ. Press: Cambridge, US, 1993. [Google Scholar]
- Williamson, T. Knowledge and its Limits, Oxford Univ. Press: Oxford, UK, 2000.
- Bartlett, S.J. The species problem and its logic, 2015. Available online: http://philsci-archive.pitt.edu/id/eprint/11655 (accessed on 01 January, 2015).
- Pavlinov, I.Ya. The Species Problem. A Conceptual History; CRC Press: Boca Raton, US, 2023. [Google Scholar]
- Wilkins, J.S. Species: A History of the Idea; University California Press: Berkely, US, 2010. [Google Scholar]
- D'Amico, G.; Rabadan, R.; Kleban, M. A Theory of taxonomy. arXiv:1611.03890 [physics.soc-ph], 2016. Available online: https://arxiv.org/abs/1611.03890 (accessed on 14 April, 2023).
- Woodger, J.H. The Axiomatic Method in Biology; Cambridge Univ. Press: Cambridge, UK, 1937. [Google Scholar]
- Gregg, J.R. The Language of Taxonomy; Columbia Univ. Press: New York, US, 1954. [Google Scholar]
- Mahner, M.; Bunge, M. Foundations of Biophilosophy; Springer: Frankfurt, Germany, 1997. [Google Scholar]
- Hennig, W. Phylogenetic Systematics; Univ. Illinois Press: Urbana, US, 1966. [Google Scholar]
- Wiley, E.O. Phylogenetics: The Theory and Practice of Phylogenetic Systematics; JohnWiley & Sons: New York, US, 1981. [Google Scholar]
- Wheeler, W.M. Present tendencies in biological theory. Sci. Month. 1929, 28, 97–109. [Google Scholar]
- Stekolnikov, A. A. [The problem of truth in biological systematics]. Zhurn. Obsh. Biol. 2003, 64, 357– 68. (in Russian).
- Oskolski, А. The taxon as an ontological problem. Biosemiotics 2011, 4, 201–222. [Google Scholar] [CrossRef]
- Wilkins, J.S. The reality of species: Real phenomena not theoretical objects. In Routledge Handbook of Evolution and Philosophy; Joyce, R., Ed.; Routledge: Abingdon, UK, 2018; pp. 167–181. [Google Scholar]
- Sarkar, S. Biodiversity and Environmental Philosophy: An Introduction; Cambridge Univ. Press: New York, US, 2005. [Google Scholar]
- Richards, R.A. The Species Problem: A Philosophical Analysis; Cambridge Univ. Press: New York, US, 2010. [Google Scholar]
- Rasnitsyn, A.P. Conceptual issues in phylogeny, taxonomy, and nomenclature. Contribut. Zool. 1996, 66, 3–41. [Google Scholar] [CrossRef]
- Meyen, S.V. [The main aspects of typology of organisms]. Zh. Obshch. Biol. 1978, 39, 495–508. (In Russian).
- Tversky, B. Parts, partonomies, and taxonomies. Develop. Psychol 1989, 25, 983–995. [Google Scholar] [CrossRef]
- Stace, C.A. Plant taxonomy and biosystematics, 2nd ed. Cambridge Univ. Press: London, UK, 1989.
- Hull, D.L. Contemporary systematic philosophies. Ann. Rev. Ecol. Evol. Syst. 1970, 1, 19–54. [Google Scholar] [CrossRef]
- Mayr, E. The growth of biological thought: Diversity, evolution, and inheritance; Belknap Press: Cambridge, US, 1982. [Google Scholar]
- Mayr, E. Toward a New Philosophy of Biology; Cambridge University Press: New York, US, 1988. [Google Scholar]
- Minelli, A. Biological systematics. The state of the art; Chapman & Hall: London, UK, 1994. [Google Scholar]
- Ereshefsky, M. Philosophy of biological classification. In Encyclopedia of life sciences, 2001. Available online: http://mrw.interscience.wiley.com/emrw/9780470015902/els/article/a0003447/current/abstract (accessed on 01 January, 2015).
- Popper, K.R. Objective Knowledge: An Evolutionary Approach; Oxford Univ. Press: Oxford, UK, 1975. [Google Scholar]
- Popper, K.R. The Logic of Scientific Discovery, 2d ed. Routledge: New York, US, 2002.
- Kuraev, V.I.; Lazarev, F.V. [Precision, truth, and growth of knowledge]; Nauka: Moscow, RF, 1988. (in Russian) [Google Scholar]
- Voyshvillo, E.K. [Concept as a Form of Thinking: Logical and Epistemological Analysis]; Moscow State Univ. Publ.: Moscow, RF, 1989. (In Russian) [Google Scholar]
- Perminov, V.Ya. [Philosophy and foundations of mathematics]. Progress-Traditsia: Moscow, RF, 2001. (in Russian).
- Murphy, G.L. The big book of concepts; MIT Press: Cambridge, US, 2002. [Google Scholar]
- Kripke, S.A. Naming and necessity; Harvard Univ. Press: Cambridge, US, 1972. [Google Scholar]
- Zadeh, L.A. Knowledge representation in fuzzy logic. In An introduction to fuzzy logic applications in intelligent systems; Ygaer R.R.; Zadeh L.A. Eds.; Springer Science + Business Media: New York, 1992; pp. 1–25. [Google Scholar]
- Kosko, B. Fuzzy Thinking: The New Science of Fuzzy Logic; Hyperion: New York, US, 1993. [Google Scholar]
- Nalimov, V.V. [A probabilistic model of language. On relation between natural and artificial languages]; Nauka: Moscow, RF, 1979. (in Russian) [Google Scholar]
- Armand, A.D. [Two in one: The law of complementarity]; LKI: Moscow, RF, 2008. (in Russian) [Google Scholar]
- Hull, D.L. Science as a Process; Univ. Chicago Press: Chicago, US, 1988. [Google Scholar]
- Prigogine, I.; Stengers, I. Order out of Chaos. Man’s New Dialogue with Nature; Heinemann: London, 1984. [Google Scholar]
- Barantsev, R.G. [Synergetics in modern natural science]; Editorial URSS: Moscow, RF, 2003. (in Russian) [Google Scholar]
- Lakatos, I. The Methodology of Scientific Research Programmes; Cambridge Univ. Press: New York, US, 1995. [Google Scholar]
- Pavlinov, I.Ya. Multiplicity of research programs in the biological systematics: A case for scientific pluralism. Philosophies 2020, 5, 7. [Google Scholar] [CrossRef]
- Radnitzky, G.; Bartley, W.W. (Eds.) . Evolutionary epistemology, rationality, and the sociology of knowledge; Open Court Publ.: La Salle, US, 1993. [Google Scholar]
- Merkulov, I.P. (Ed.) . [Evolutionary epistemology: Problems and Perspectives]; ROSSPEN: Moscow, RF, 1996. (in Russian) [Google Scholar]
- Giangrande, A.; Rouse, G.W.; Plejie, F.J.J.; et al. The cladistic revolution, new taxonomic instruments and implications for ecologists. Bioi. Mar. Medit. 2005, 12, 23–30. [Google Scholar]
- Antonov, A.S. [Genosystematics of plants]; Akademkniga: Moscow, RF, 2006. (in Russian) [Google Scholar]
- Minelli, A. Biological systematics in the evo–devo era. Europ. J. Taxon. 2015, 125, 1–23. [Google Scholar] [CrossRef]
- Martynov, A.V. [Ontogenetic Systematics, and a New Model of Bilaterian Evoluiton]; KMK Sci. Press: Moscow, RF, 2011. (In Russian) [Google Scholar]
- Martynov, A.; Lundin, K.; Korshunova, T. Ontogeny, phylotypic periods, paedomorphosis, and ontogenetic systematics. Front. Ecol. Evol. 2022, 10, 806414. [Google Scholar] [CrossRef]
- Wheeler, Q.D. (Ed.) The New Taxonomy; CRC Press: Boca Raton, US, 2008. [Google Scholar]
- Zander, R.H. A Framework for Post-Phylogenetic Systematics; Zetetic Publ.: St. Louis, 2013. [Google Scholar]
- Stamos, D.N. The Species Problem. Biological Species, Ontology, and the Metaphysics of Biology; Lexington Books: Oxford, UK, 2003. [Google Scholar]
- Amitani, Y.; The persistence question of the species problem. Thes. DrPh. Univ. British Columbia: Vancouver, US, 2010. Available online: https://open.library.ubc.ca/media/stream/pdf/24/1.0071552/1 (accessed on 01 January, 2015).
- Chambers, G. The species problem: Seeking new solutions for philosophers and biologists. Biol. Philos. 2012, 27, 755–765. [Google Scholar] [CrossRef]
- Pavlinov, I.Y. (Ed.) The Species Problem: Ongoing Issues; InTech Open Access Publ.: Rijeka, Croatia, 2013. [Google Scholar]
- Wilkins, J.S.; Zachos, F.E.; Pavlinov, I.Ya. (Eds.) . Species Problems and Beyond. Contemporary Issues in Philosophy and Practice. CRC Press: Boca Raton, US, 2022.
- Mayden, R.L. A hierarchy of species concepts: The denouement in the saga of the species problem. In Species. The units of biodiversity, Claridge M.F.; Dawah, A.H., Wilson, M.R., Eds.; Chapman & Hall: London, 1997, pp. 381– 424. [Google Scholar]
- Naomi, S.-I. On the integrated frameworks of species concepts: Mayden’s hierarchy of species concepts and de Queiroz’s unified concept of species. J. Zool. Syst. Evol. Res. 2011, 49, 177–184. [Google Scholar] [CrossRef]
- Hall, B.K. (Ed.) . Homology, the hierarchical basis of comparative biology; Acad. Press: San Diego, US, 1994. [Google Scholar]
- Bock, G.R.; Cardew, G. (Eds.) Homology; John Wiley & Sons: Chichester, UK, 1999. [Google Scholar]
- Pavlinov, I.Y. The contemporary concepts of homology in biology: A theoretical review. Biol. Bull. Rev. 2012, 2, 36–54. [Google Scholar] [CrossRef]
- Wagner, G.P. Homology, genes, and evolutionary innovation. Princeton Univ. Press: Princeton, US, 2014.
- de Beer, G. Homology, an Unsolved Problem; Oxford University Press: Oxford, UK, 1971. [Google Scholar]
- Scotland, R.W. Deep homology: A view from systematics. Bioessays 2010, 32, 438–449. [Google Scholar] [CrossRef] [PubMed]
- Minelli, A.; Fusco, G. Homology. In The Philosophy of Biology: A companion for Educators; Kampourakis, K., Ed.; Springer Science + Business Media: Dordrecht, 2013; pp. 289–322. [Google Scholar]
- Minelli, A. Tracing homologies in an ever-changing world. Riv. Estet. 2016, 62, 40–55. [Google Scholar] [CrossRef]
- Hempel, G. Aspects of Scientific Explanation and Other Essays in the Philosophy of Science; Free Press: New York, US, 1965. [Google Scholar]
- Gaydenko, P.P. Scientific Rationality and Philosophical Mind; Progress-Traditsia: Moscow, RF, 2003. (In Russian) [Google Scholar]
- Weinberg, S. Dreams of a Final Theory: The Scientist’s Search for the Ultimate Laws of Nature; Pantheon: New York, 1992. [Google Scholar]
- Linnaeus, C. Amoenitas Academiae [...]; Godofredum Kiesewetter: Holmiae et Lipsiae, Germany, 1749. [Google Scholar]
- Adanson, M. Familles des Plantes; Vincent: Paris, France, 1763. [Google Scholar]
- Lam, K.H. The realism of taxonomic pluralism. Metaphysics 2020, 3, 1–16. [Google Scholar] [CrossRef]
- Rozov, M. A. [Classification and theory as knowledge systems]. In [On the way to the theory of classification], Rozova S.S. Ed.; Novosibirsk Univ. Publ.: Novosibirsk, RF, 1995; pp. 81–127. (in Russian) [Google Scholar]
- Gilmour, J.S.L. Taxonomy and philosophy. In The New Systematics; Huxley, J., Ed.; Oxford University Press: Oxford, UK, 1940; pp. 461–474. [Google Scholar]
- Doyen, J.T.; Slobodchikoff, C.N. An operational approach to species classification. Syst. Biol. 1974, 23, 239–247. [Google Scholar] [CrossRef]
- Pleijel, F.; Rouse, G.W. Least-inclusive taxonomic unit: A new taxonomic concept for biology. Proc. Roy. Soc. B Biol. Sci. 2000, 267, 627–630. [Google Scholar] [CrossRef]
- Sarkar, S.; Margules, C. Operationalizing biodiversity for conservation planning. J. Biosci. 2001, 27, Suppl. 2, 299–308. [Google Scholar] [CrossRef]
- Sites, J.W.; Marshall, J.C. Operational criteria for delimiting species. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 199–227. [Google Scholar] [CrossRef]
- Riddle, B.R.; Hafner, D.J. Species as units of analysis in ecology and biogeography: Time to take the blinders off. Global Ecol. Biogeogr. 1999, 8, 433–441. [Google Scholar] [CrossRef]
- Hendry, A.P.; Vamosi, S.M.; Latham, S.J.; et al. Questioning species realities. Conserv. Genet. 2000, 1, 67–76. [Google Scholar] [CrossRef]
- Kober, G. 2010. Biology without species: A solution to the species problem, PhD Diss.; Boston Univ., US. Available online: https://philpapers.org/rec/KOBBWS (accessed on 01 January, 2015).
- Mishler, B.D. What, if anything, are species? CRC Press: Boca Raton, US, 2021.
- Brooks, D.R.; McLennan, D.A. Species: Turning a conundrum into a research program. J. Nematol. 1999, 31, 117–133. [Google Scholar] [PubMed]
- Kendig, C. Towards a multidimensional metaconception of species. Ratio 2014, 27, 155–172. [Google Scholar] [CrossRef]
- LaPorte, J. Modern essentialism for species and its animadversions. In The Routledge handbook of evolution and philosophy; Joyce R., Ed., Routledge: New York, US, 2017; pp. 182–193. [Google Scholar]
- Kuhn, T.S. The structure of scientific revolutions. Univ. Chicago Press: Chicago, US, 1962.
- Laudan, L. A confutation of convergent realism. Phil. Sci. 1981, 48, 19–49. [Google Scholar] [CrossRef]
- Hacking, I. Representing and intervening, Introductory topics in the philosophy of natural science; Cambridge Univ. Press: Cambridge, UK, 1983. [Google Scholar]
- Novick, A.; Doolittle, W.F. ‘Species’ without species. Stud. Hist. Philos. Sci. 2021, 87, 72–80. [Google Scholar] [CrossRef]
- Minelli, A. Taxonomy needs pluralism, but a controlled and manageable one. Megataxa 2020, 1, 9–18. [Google Scholar] [CrossRef]
- Wang, X. Taxonomy, truth-value gaps and incommensurability: A reconstruction of Kuhn’s taxonomic interpretation of incommensurability. Stud. Hist. Phil. Sci. 2002, 33, 465–485. [Google Scholar] [CrossRef]
- Kellert, S.H.; Longino, H.E.; Waters, C.K. (Eds.) . Scientific Pluralism; Univ. Minnesota Press: Minneapolis, US, 2006. [Google Scholar]
- Broughton, V. The need for a faceted classification as the basis of all methods of information retrieval. New Inform. Persp. 2006, 58, 49–72. [Google Scholar] [CrossRef]
- Chan, L.M.; Salaba, A. Cataloging and classification: An introduction, 4th ed.; Rowman & Littlefield Publ.: Lanham, US, 2016. [Google Scholar]
- Croft, W.; Cruse, A. Cognitive linguistics; Cambridge Univ. Press: Cambridge, UK, 2004. [Google Scholar]
- Chebanov, S.V.; Martynenko, G.Ya. [From the history of typological notions.] [Structural and applied linguistics] 2008, 6, 328–390. (in Russian).
- Dubois, A. Bionomina, a forum for the discussion of nomenclatural and terminological issues in biology. Bionomina 2010, 1, 1–10. [Google Scholar] [CrossRef]
- Pavlinov, I.Ya. Taxonomic Nomenclature. What’s in a Name—Theory and History; CRC Press: Boca Raton, US, 2022. [Google Scholar]
- Aescht, E. Reflecting on a theory of biological nomenclature (onymology). Alytes 2018, 36, 212–237. [Google Scholar]
- Chomsky, N. Language and problems of knowledge; MIT Press: Cambridge, US, 1987. [Google Scholar]
- Hill, J.H.; Mannheim, B. Language and world view. Annu. Rev. Anthrop. 1992, 21, 381–406. [Google Scholar] [CrossRef]
- Devitt, M.; Sterelny, K. Language and reality. An introduction to the philosophy of language, 2nd ed.; MIT Press: Cambridge, US, 1999. [Google Scholar]
- Allard-Kropp, M. Languages and worldview; Univ. Missouri: St. Louis, US, 2020. [Google Scholar]
- Uryson, E.V. [Research problems of the linguistic world picture. Analogy in semantics]; YaSK: Moscow, RF, 2003. (in Russian) [Google Scholar]
- Kubryakova, E.S. [Language and knowledge. The role of language in the knowing the world]; Inst. of Language RAS: Moscow, RF, 2004. (in Russian) [Google Scholar]
- Slaughter, M. Universal languages and scientific taxonomy in the seventeenth century; Cambridge Univ. Press: Cambridge, UK, 1982. [Google Scholar]
- Dubois, A. Proposed rules for the incorporation of nomina of higher- ranked zoological taxa in the International Code of Zoological Nomenclature. 1. Some general questions, concepts and terms of biological nomenclature. Zoosystema 2005, 27, 365– 426.
- Dubois, A.; Bauer, A.M. , Ceríaco, L.M.P.; et al. The Linz ZooCode project: A set of new proposals regarding the terminology, the principles and rules of zoological nomenclature [...]. Bionomina 2019, 17, 1–111. [Google Scholar] [CrossRef]
- Tournefort, J. Pitton de. Élémens de Botanique, ou Méthode pour Connoître les Plantes, vol. 1; De l’Imprim. Roy.: Paris, France, 1694. [Google Scholar]
- Linnaeus, C. Philosophia botanica in qua explicantur fundamenta botanica [...]; Godofr. Kiesewetter: Stockholm, Sweeden, 1751. [Google Scholar]
- Fabricius, I.C. Systema eleutheratorum secundum ordines, genera, species [...], vol. 1; Bibliopol. Acad. Novi: Kiliae, Germany, 1801. [Google Scholar]
- de Candolle, A. Nouvelles remarques sur la nomenclature botanique; H. Georg: Geneva, Swiss, 1883. [Google Scholar]
- Coues, E.; Allen, J.A.; Ridgway, R.; et al. The Code of Nomenclature and Check-List of North American Birds [...]. American Ornithologists’ Union: New York, US, 1886.
- Michener, C.D. Some future developments in taxonomy. Syst. Zool. 1963, 12, 151–172. [Google Scholar] [CrossRef]
- Hull, D.L. The syntax of numericlature. Syst. Zool. 1968, 17, 472–274. [Google Scholar] [CrossRef]
- de Queiroz, К.; Gauthier, J. Toward a phylogenetic system of biological nomenclature. Trends Ecol. Evol. 1994, 9, 27–31. [CrossRef] [PubMed]
- de Queiroz, K.; Cantino, P.D. Phylogenetic nomenclature and the PhyloCode. Bull. Zool. Nomencl. 2001, 58, 254–271. [Google Scholar]
- de Queiroz, К. Biological nomenclature from Linnaeus to the PhyloCode. Bibl. Herpet. 2012, 9, 135–145. [Google Scholar]
- Wheeler, Q.D.; Raven, P.H.; Wilson, E.O. Taxonomy: impediment or expedient? Science 2004, 303: 285.
- de Carvalho, M.R.; Bockmann, F.A.; Amorim, D.S.; et al. Taxonomic impediment or impediment to taxonomy? A commentary on systematics and the cybertaxonomic-automation paradigm. Evol. Biol. 2007, 34, 140–143. [Google Scholar] [CrossRef]
- Engel, M.S.; Ceríaco, L.M.P.; Daniel, G. M.; et al. The taxonomic impediment: a shortage of taxonomists, not the lack of technical approaches. Zool. J. Linn. Soc. 2021, 193, 381–387. [Google Scholar] [CrossRef]
- List of Life Sciences, 2023. Available online: https://en.wikipedia.org/wiki/List_of_life_sciences (accessed on 10 June, 2023).
- Hedges, S.B.; Kumar, S. (Eds.) . The Timetree of Life; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Pozdnyakov, A.A. [Philosophical foundations of contemporary biology. An introduction to organic biology]. URSS: Moscow, RF, 2018. (in Russian).
- Schram, F.R. The truly new systematics—megascience in the information age. Hydrobiologie 2004, 519, 1– 7. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



