
Submitted:
08 June 2023
Posted:
08 June 2023
You are already at the latest version
Abstract
In order to improve the adaptability of broilers to low-temperature environments and their ability to resist acute cold stress (ACS), 240 one-day-old broilers were selected and randomly divided into three groups. The control treatment (CC) was raised at the conventional feeding temperature from 1-43 days (d), the cold stimulation treatments (CS) were kept at 3 °C below the temperature of CC at 1-d intervals for 3 and 6 hours from 15 to 35 d, namely CS3 and CS6, respectively. Then all broilers were kept at 20 °C from 36 to 43 d. ACS was then carried out at 44 d, and the ambient temperature was dropped to 10 °C for 6 hours. The study investigated the production perfor-mance, as well as levels of intestinal barrier genes (including Claudin-1, E-cadherin, Occludin, ZO-1, ZO-2 and Mucin2), secretory IgA in duodenum and jejunum, and immunoglobulins (IgA and IgG) in serum. The results showed that IMCS could increase the daily weight gain and decrease the feed conversion ratio. During the IMCS, the expression levels of intestinal barrier genes were up-regulated and the content of secretory IgA was increased. When IMCS ceased for one week, the level of immunoglobulins in serum stabilized, and the expression levels of Occludin, ZO-2 and Mucin2 still maintained high levels. After ACS, broilers that received IMCS training maintained the high levels of intestinal barrier genes and secretory IgA.
Keywords:
broiler
; duodenum
; jejunum
; production performance
; intestinal barrier
; immunoglobulin
1. Introduction
Animals living in cold regions often suffer from low temperature that can cause damage to various organs, leading to decreased production performance, compromised immune function, and disrupted internal homeostasis[1,2,3]. The intestinal tract not only has a role in digesting and absorbing nutrients but also serves as a barrier for protection. It plays an essential role in the immunity[4,5]. The intestinal mucosal barrier is a protective barrier that prevents the loss of water, electrolytes, and the invasion of microorganisms in the lumen[6,7]. The physical intestinal barrier consists mainly of intestinal epithelial cells and tight junctions (TJ)[8]. TJ plays a crucial role in intercellular connections [9,10]. The absence of TJ complex can impair barrier protection function, affecting the intestinal permeability, which can be used as an indicator to evaluate the integrity of intestinal mucosa[11]. Fanning and Krause et al. discovered that Claudins and ZO-1 proteins can regulate epithelial permeability and help maintain the integrity of intestinal barrier[10,12]. The expression levels of Claudin-1 and Occludin can be used as biomarkers to determine the function of intestinal barrier in broilers[13,14]. Stress can inhibit the expression levels of TJ proteins, increase intestinal permeability, destroy the integrity of the intestine, and result in intestinal barrier dysfunction of broilers[15,16,17]. For instance, heat stress (35 °C) can reduce the levels of TJ proteins and increase the permeability of tight junctions between cells of broilers[18]. Zhao et al. found that low temperature could significantly shorten the villus height of the small intestine of broilers, and seriously damage the intestine[19]. However, appropriate cold or hot stimulation can improve intestinal tightness, thereby enhancing intestinal barrier function. Heat stress (38-39 °C) for 5 days could significantly increase the mRNA levels of TJ proteins in jejunum of broilers [20]. Cold stimulation with an interval of one day, at 3 °C below the conventional temperature could relieve the intestinal damage when were broilers subjected the acute cold stress[21]. Therefore, the impact of environment on organisms is twofold, and reasonable using of environmental stimuli can help to maintain the integrity of intestinal barrier.
Immunoglobulins are crucial effectors of the immune system, and levels of immunoglobulins can vary with changes in the internal and external environment of the body. When environmental changes are excessive, a reduction in the content of immunoglobulin in animal serum can lead to damage to the body. Heat stress (40 °C) for 4 weeks had reduced the IgA and IgG content in serum and affect the development of immune function in broilers[22]. The appropriate cold stimulation can increase the content of immunoglobulin to a certain extent. Mild cold stimulation at 3 °C below the normal feeding temperature can increase the expression of IgA and IgG in the spleen of broilers and activate the innate immune system[23]. After acute cold stress, the content of IgA in the ileum of broilers with cold training was significantly higher than that of broilers without cold training, aiding the body in resisting cold environments[21]. Therefore, the level of immunoglobulin can reflect the immune function of the body.
Low temperature is one of the important factors limiting livestock and poultry industry in northern regions, as cold stress often causes damage to animals. However, reasonable utilization of cold environment can have positive effects. Some studies indicated that proper cold stimulation can enhance the intestinal immunity of broilers[21,23]. It remains unclear whether intermittent and mild cold stimulation (IMCS) can enhance the production performance and intestinal immune function of broilers by enhancing the integrity of intestinal barrier and the levels of immunoglobulins. In the study, broilers were trained with cold stimulation at 3 °C below conventional feeding temperature of every other day, aiming to explore the effect of IMCS on production performance, intestinal barrier and immunity and tried to find out its potential molecular mechanism. We hope to discover a suitable cold stimulation training program to help the body adapt to low-temperature environments, and provide a theoretical basis for the establishment of cold adaptation strategies in poultry production practice in cold regions.
2. Materials and Methods
2.1. Animals and Experimental Design
Two hundred and forty 1-day old Ross 308 broilers were randomly divided into three groups, each with five replicates containing 16 broilers per replicate. All broilers were reared in battery cages located in climate control rooms. The control group (CC) was raised at the standard feeding temperature, which was maintained at 35 °C for the first 3 days (d) then gradually reduced by 0.5 °C every day until it reached 20 °C on day 35. The feeding temperature of the cold stimulation group (CS) was the same as that of the CC group from 1 to 14 d, and the temperature at one-day intervals from 15-35 d was 3 °C below CC group. The duration of cold stimulation was 3 h (CS3) and 6 h (CS6) respectively. From day 36 to 43, broilers in all three experimental groups were kept at a temperature of 20 °C. On day 44, all broilers were subjected to acute cold stress (ACS) of 10 °C for 6 h starting at 07:00 am. The specific feeding temperature is shown in Figure 1. The relative humidity of the room was kept at 60-70% at 1-14 days and 40-50% at 15-44 days. Throughout the study, the birds were given free access to water and feed. The feed supply included: a commercial starter diet was given for 1-3 weeks (metabolizable energy [ME] of 12.1 MJ/kg and crude protein [CP] of 21.0%) and a commercial growing-finishing diet was given for 4-6 weeks (ME of 12.6 MJ/kg and CP of 19.0%) (Baishicheng Animal Husbandry, Harbin, China). The main composition of the commercial diets included corn, soybean meal, vitamin A, vitamin D3, NaCl, copper sulfate, calcium chloride, choline etc. All procedures performed in the present study were approved by the Institutional Animal Care and Use Committee of Northeast Agriculture University (IACUCNEAU20150616).
2.2. Sample Collection
In this study, four broilers in each group were randomly selected and euthanized for blood, duodenum and jejunum at 22, 36, 43 d and after 6 h of ACS. The duodenum and jejunum were taken out and quickly put into liquid nitrogen, and then transferred to - 80 °C until used for RNA extraction. Blood was stewed and centrifuged at 2000×g at 4 °C for 15min. Then the supernatant was taken out and stored at - 20 °C for Elisa detection.
2.3. Evaluation of Broiler Production Performance
Five representative broilers from each sample group were randomly selected, and weight gain was recorded on the first day of weeks 3, 4, 5 and 6. Feed addition and surplus of each cage were counted every day. The average of daily weight gain and daily feed intake was recorded. The number of dead broilers was recorded and excluded from the test, and the average value was measured for the remaining broilers. Feed conversion ratio was calculated using the following formula: daily feed intake/daily weight gain.
2.4. RNA Extraction and Reverse Transcription
Total RNA from duodenum and jejunum was extracted using Rnaiso plus (Bao Bioengineering Co., Ltd., Dalian, China) according to the instructions. RNA concentration and OD260/OD280 ratio were detected and recorded with an ultramicro spectrophotometer (IMPLEN, p330, Germany). RNA concentration was adjusted to 1 μ g/μ l and the OD value was within the range of 1.8-2.1. Complementary DNA (cDNA)was synthesized by PrimeScript® RT reagent Kit with gDNA Eraser (Perfect Real-Time) (TaKaRa, Shiga, Japan). The specific reaction system is shown in Table 1.
2.5. Quantitative Real-Time PCR (qRT-PCR) Analysis
Target genes (Claudin-1, E-cadherin, Occludin, ZO-1, ZO-2, Mucin2) and reference gene (β-actin) were synthesized by Shanghai Sangong Bioengineering Co., Ltd (Shanghai, China). The primer sequence is shown in Table 2. qRT-RCR was performed on AriaMx Real Time PCR instrument (Agilent, USA) using THUNDERBIRDSYBR qPCR Mix (Toyobo, Japan), following the manufacturer's instructions. The reaction procedure was as follows: 95 °C for 60s, followed by 40 cycles of 95 °C for 15s, and 60 °C for 1min. The relative mRNA levels of target gene were calculated by 2 -ΔΔCt method.
2.6. Western Blot Analysis
Western and IP cell lysates (Biosharp, Beijing, China) containing 1% PMSF (SparkJade, Harbin, China) were used to extract proteins from duodenum and jejunum. The protein concentration was determined using a BCA protein detection kit (Biosharp, Beijing, China) and adjusted to 4 mg/mL. The methods of western blot according to our previous reports. Briefly, equal amounts of total protein (40 mg/condition) were separated by 10% SDS-PAGE gel (Beyotime, Shanghai, China), and transferred to nitrocellulose membrane with a semi-dry transfer instrument. The membrane was sealed in 5% skimmed milk for 2 hours, and then washed with 1x PBST for three times. The primary antibodies against Claudin-1(1:500, WanleiBio, Shenyang, China), E-cadherin (1:1000, Sangong, Shanghai, China), Occludin (1:1000, Sangong, Shanghai, China) and β-actin (1:9000, Zenbo, Chengdu, China) were incubated with nitrocellulose membrane at 4 °C overnight. After cleaning the membrane with 1x PBST for 3 times, an HRP goat anti-rabbit IgG (1:9000, ABclonal, Wuhan, China) was added and incubated with membrane for 1 hour. The protein bands were washed with 1x PBST for three times, visualized using ECL chemiluminescence kit (Biosharp, Beijing, China) and scanned with the grayscale scanner (CLINX, Shanghai, China). Finally, Image J software (NIH, Bethesda, MD) was used for gray scale analysis. The relative expression levels of proteins were determined by the ratio of the gray value for the target protein to β-actin.
2.7. Elisa Detection
The levels of immunoglobulin (IgA, IgG and IgM) in serum of broilers at 22, 36 and 43 d and secretory IgA (SIgA) in duodenum and jejunum at day 22, 36, 43 and Y6 were detected by enzyme-linked immunosorbent assay (ELISA), according to the instructions provided by the manufacturer (mlbio, Shanghai, China).
2.8. Statistics and Analysis
All results were analyzed by IBM SPSS 21.0 software (IBM, Armonk, NY, USA). Kolmogorov-Smirnov was used to test the normal distribution of all data. One-way analysis of variance (ANOVA) with Duncan's multiple comparisons was used to analyze the effect of cold stimulation on production performance, the mRNA expression levels of barrier genes in duodenum and jejunum and the content of immunoglobulin in serum, duodenum and jejunum. The results were expressed as mean ± standard deviation (SD), and the significant difference was expressed as P < 0.05.
3. Results
3.1. Production Performance of Broilers
The effect of IMCS on production performance is shown in Table 3. In CS3 group, feed conversion ratio was significantly lower than that in CC and CS6 groups (P < 0.05). Daily weight gain in CS3 group was significantly higher than that in CC and CS6 groups (P < 0.05). No significant difference was observed for daily food intake between CC and CS groups (P > 0.05).
3.2. mRNA Levels of Intestinal Barrier Genes in Duodenum
The results of mRNA expression levels of barrier genes in duodenum are shown in Figure 2. At 22 d, the mRNA expression levels of Claudin-1、E-cadherin and ZO-2 in CS3 were significantly higher than those in CC and CS6 (P < 0.05), but there were no significant differences between CC and CS6 (P > 0.05). Compared to CC, the mRNA levels of Occludin and Mucin2 in CS were increased significantly (P < 0.05), and the levels in CS6 were significantly higher than those in CS3 (P < 0.05). A significant increment in ZO-1 mRNA levels was found in the CC and C3 groups when compared to CC treatment (P < 0.05), but no significant difference was found between CC and CS3 (P > 0.05). At day 36, the mRNA levels of Occludin and Mucin2 in CC and CS3 were significantly higher than those in CS6 (P < 0.05), while no significant difference was observed between CC and CS3 groups (P > 0.05). ZO-2 mRNA level was found to be significantly higher in CS than that in CC (P < 0.05), and that in CS3 was significantly higher than that in CS6 (P < 0.05). A significantly increased in Claudin-1 level was found in CS6 compared with CS3 (P < 0.05). The expression levels of other genes were changed slightly (P > 0.05). At 43 d, the mRNA levels of Occludin, ZO-1, ZO-2 and Mucin2 in CS3 were significantly higher than those in CC and CS6 (P < 0.05), and the Occludin mRNA level in CS6 was higher than that in CC (P < 0.05), while no significant difference was observed in other genes (P > 0.05). Claudin-1 mRNA level in CC group was significantly higher than that in CS groups (P < 0.05), and that in CS3 group was significantly higher than that in CS6 group (P < 0.05). The level of E-cadherin in CS3 group was significantly up-regulated compared with CS6 group (P < 0.05), but there was no significant difference between CC and CS (P > 0.05). After 6 hours of ACS (Y6), the levels of all genes, except for Occludin in CS6, were significantly increased compared to CC and CS3 (P < 0.05), while CC and CS3 groups had no significant difference (P > 0.05). Moreover, the mRNA level of Occludin in CS6 was significantly higher than that in CS3 (P < 0.05).
3.3. Protein Levels of Intestinal Barrier Genes in Duodenum
The results of protein expression levels of Claudin-1, Occludin and E-cadherin in duodenum are shown in Figure 3. At 22 d, the protein expression level of Claudin-1 in CS was significantly higher than that in CC (P < 0.05), but there were no significant differences between CS3 and CS6 (P > 0.05). The level of Occludin in CS3 was significantly higher than that in CS6 (P < 0.05), but no significant difference was found between CC and CS (P > 0.05). Compared to CC, the protein level of E-cadherin in CS was increased significantly (P < 0.05), and that in CS3 was significantly higher than that in CS6 (P < 0.05). At day 36, a significant increment in Claudin-1 and Occludin levels was found in CS6 when compared to CC and CS3 (P < 0.05), and those in CC were significantly higher than those in CS3 (P < 0.05). The E-cadherin level in CS3 was increased significantly compared to CC and CS6 (P < 0.05), and the level of E-cadherin in CC was significantly higher than that in CS6 (P < 0.05). At day 43, compared to CC, the level of Claudin-1 in CS was significantly increased (P < 0.05), but there was no significant difference between CS3 and CS6 (P > 0.05). The protein levels of Occludin and E-cadherin in CS3 were significantly higher than those in CC and CS6 (P < 0.05), while no significant difference was observed between CC and CS groups (P > 0.05). After ACS for 6 h, the levels of Claudin-1 and E-cadherin were significantly increased compared to CC and CS3 (P < 0.05), and the Claudin-1 level in CC was significantly higher than that in CS3 (P < 0.05). The E-cadherin level had no significant difference between CC and CS3 (P > 0.05). A significant increase in Occludin level was observed in CS3 when compared to CC and CS6 (P < 0.05), but there was no significant difference between CC and CS6 (P > 0.05).
3.4. mRNA Levels of Intestinal Barrier Genes in Jejunum
Figure 4 shows the effect of IMCS and ACS on mRNA levels of barrier genes in jejunum. After seven-day IMCS (at 22 d), the level of Mucin2 mRNA in CC was higher than that in CS (P < 0.05), there was no significant difference between CS groups (P > 0.05). Compared to CS6, a significant increase in the mRNA levels of E-cadherin and Occludin and a significant decrease in levels of Claudin-1 and ZO-1 were found in CC and CS3 (P < 0.05), and Claudin-1 in CS3 was significantly higher than that in CC group (P < 0.05). However, no significant difference between CC and CS3 groups was observed in E-cadherin, Occludin and ZO-1 (P > 0.05). The level of ZO-2 was not significantly different among the three groups (P > 0.05). At 36 d, the mRNA level of Occludin in CC was significantly higher than that in CS (P < 0.05), whereas there was no significant difference between CS groups (P > 0.05). Compared with CC and CS6,Mucin2 level in CS3 exhibited a marked increase (P < 0.05), but there was no significant difference between CC and CS6 (P > 0.05). Claudin-1 mRNA level in CS6 was significantly increased compared to CC (P < 0.05). No significant difference was found for other genes (P > 0.05). At 43 d, compared with CC, the mRNA expression levels of Claudin-1 in CS and ZO-1 in CS3 were significantly up-regulated (P < 0.05), while E-cadherin in CS and Mucin2 in CS3 were significantly down-regulated (P < 0.05). There was no significant difference in the levels Claudin-1 and E-cadherin between CS3 and CS6 groups (P > 0.05). After ACS, the mRNA level of Mucin2 in CS was significantly higher than that in CC (P < 0.05), but there was no significant difference in Mucin2 between CS groups (P > 0.05) and other genes among the three groups (P > 0.05).
3.5. Protein Levels of Intestinal Barrier Genes in Jejunum
Figure 5 shows the effect of IMCS and ACS on protein levels of barrier genes in jejunum. At 22 d, the level of Claudin-1 in CS6 was higher than in CC and CS3 (P < 0.05), and that in CC was significantly higher than that in CS3 (P < 0.05). The Occludin level in CS was significantly higher than that in CC (P < 0.05), there was no significant difference between CS3 and CS6 (P > 0.05). Compared to CC and CS6, a significant increase in the protein levels of E-cadherin was found in CS3 (P < 0.05), and the E-cadherin level in CC was significantly higher than that in CS6 (P < 0.05). At 36 d, the Claudin-1 level in CS6 was significantly higher than that in CC and CS3 (P < 0.05), and no significant difference was found between CC and CS3 (P > 0.05). The levels of Occludin and E-cadherin in CS3 were significantly higher than those in CC and CS6 (P < 0.05), and Occludin level in CS6 was significantly higher than that in CC (P < 0.05), but no significant difference was observed in E-cadherin between CC and CS6 (P > 0.05). At 43 d, the Claudin-1 level in CC and CS3 was significantly higher than that in CS6 (P < 0.05), there was no significant difference between CC and CS3 (P > 0.05). The level of Occludin in CC was significantly increased compared to CS (P < 0.05), and the Occludin level between CS had no significant difference (P > 0.05). The level of E-cadherin in CS6 was significantly higher than that in CC and CS3 (P < 0.05), and that in CC was significantly higher than that in CS3 (P < 0.05). After ACS for 6 h, the levels of Claudin-1 and E-cadherin in CS6 were significantly increased compared to CC and CS3 (P < 0.05), and the E-cadherin level in CC was significantly higher than that in CS3 (P < 0.05), but no significant difference was observed in Claudin-1 between CC and CS3 (P > 0.05). The level of Occludin in CS was significantly increased compared to CC (P < 0.05), and that in CS3 was significantly higher than that in CS6 (P < 0.05).
3.6. Levels of Immunoglobulin in Serum, Duodenum and Jejunum
The effect of IMCS and ACS on the content of immunoglobulin in serum (6A-6C), duodenum and jejunum (6D-6E) is shown in Figure 6. At 22 d, the contents of IgG and IgM in serum in CS6 were significantly higher than those in CC (P < 0.05), and those in CS3 were higher than those in CC, but the difference was not significant (P > 0.05). The level of SIgA in jejunum in CS6 was significantly higher than that in CC and CS3 (P < 0.05), but CC and CS3 had no significant difference (P > 0.05). There was no significant difference in content of IgA in serum and SIgA in duodenum among the three groups (P > 0.05). At 36 d, the content of IgG in CS was significantly higher than that in CC (P < 0.05), but CS3 and CS6 had no significant difference (P > 0.05). The contents of SIgA in jejunum in CC and CS6 were significantly lower than those in CS3 (P < 0.05). There was no significant difference in the contents of IgA, IgM in serum and SIgA in duodenum among the three groups (P > 0.05). At 43 d, the contents of immunoglobulin except for SIgA in jejunum were not significantly different among the three groups (P > 0.05). The content of SIgA in jejunum in CS6 was significantly higher than that in CC and CS3 (P < 0.05). After ACS for 6 h, the content of SIgA in duodenum and jejunum in CS was significantly higher than that in CC (P < 0.05), but there was no significant difference in duodenum among the three groups (P > 0.05).
4. Discussion
Environmental temperature is one of the most important factors affecting broiler production performance, including egg production, food intake, weight gain and ketone body quality[3,24,25]. Low temperature environment leads to significantly decreased body weight gain, pectoral weight and thigh muscle weight of broilers[26]. However, appropriate cold stimulation would prevent adverse effects on production performance. Blahova et al. found that after 20 days of cold stimulation (4-13 °C), no significant difference was found in body weight gain of broilers in the control group and cold stress group[27]. Moreover, Shinder et al. reported that the survival rate of broilers significantly increased after exposure to repeated short-term cold training[28]. The above results were similar to ours. In the present study, the difference in daily feed intake between CC and CS groups was not significant, which may due to the fact that cold stimulation (3 °C below feeding temperature of CC group) was relatively mild. Broilers can adapt to the environment by exerting thermoregulation, without the need for increasing daily feed intake to increase heat production. The daily weight gain of broilers in CS3 group was significantly higher than that of CC and CS6 groups, but no significant difference was observed between CC and CS6 groups; this could be partly explained by the fact that IMCS applied for 3 h increased daily weight gain by increasing fat accumulation, whereas IMCS applied for 6 h would eliminate the accumulation effect observed in broilers submitted to ICMS for 3 h. Feed conversion ratio in broilers of the CS3 group was lower than that of CC and CS6 groups, and no difference was observed in daily feed intake among all groups, although daily weight gain of broilers in CS3 group increased. Collectively, the results showed that appropriate cold stimulation could increase the production performance of broilers.
TJ is the most important connection between intestinal cells. TJ only allows small molecules to pass through and hinders the passage of macromolecules and microorganisms, which plays an important role in maintaining the integrity of mucosa and the stability of barrier function[9,10]. In the present study, at the beginning of CS (22d), the mRNA and protein levels of Claudin-1 and Occludin in duodenum in CS3 and that of Claudin-1 in jejunum in CS6 were up-regulated, to resist the influence of cold stimulation on intestinal mucosa. When the IMCS ended (36d), the protein levels of Claudin-1 in CS6 in duodenum and jejunum and that of Occludin in CS3 in jejunum were increased. The results indicated that the 21-day IMCS training at a temperature of 3 °C below ambient temperature could not cause damage to the intestinal mucosa, and could promote the expression and reasonable distribution of intestinal TJ to a certain extent, thus maintaining the integrity of the mucosa and the stability of the intestinal barrier function. The protein level of Occludin in Caco-2 cells was increased after heat stress (39 or 41 °C)[29]. A study showed that the level of Occludin in jejunum of broilers was significantly increased after cold stimulation at 16 °C for 72 h[30], which were consistent with our results. ZO-1 is the key to the polymerization of Claudins[31]. In the study, one week after IMCS (43d), the mRNA levels of ZO-1 and ZO-2 in the duodenum and jejunum of CS3 group were increased, proving that IMCS could promote Claudins polymerization by increasing the expression levels of ZO-1 and ZO-2, making the intestinal barrier tighter. Consistent with our results, Zhou et al. found that 72 h cold treatment resulted in increased ZO-1 levels. E-cadherin is a calcium dependent transmembrane protein, which distributed in epithelial cells and involved in cell adhesion[32]. Mucin2 is the most abundant mucin in the intestine and is the first line of defense against microbial invasion of the intestine[33]. The study found that the E-cadherin and Mucin2 levels in CS3 in duodenum were higher than those in CC from 22 d to 43 d, indicating that appropriate cold stimulation could up-regulate the E-cadherin and Mucin2 levels, improve the integrity of intestinal epithelial cells and the tightness between cells, protect the intestine from the invasion of susceptible bacteria, and maintain the intestinal barrier function. This is consistent with previous study showing that changes in environmental temperature can affect the expression of TJ genes and thereby alter the intestinal permeability and barrier function[34]. In the study, the expression trend of TJ genes in duodenum and jejunum is slightly different, which may because TJ proteins differentiate and restrict solute channels according to molecular size, and change with different intestinal[35]. Overall, the study suggested that appropriate cold stimulation could promote the expression of TJ genes and improve the intestinal barrier function, thus benefitting the health and production performance of broilers.
As an important effector molecule of immune system, immunoglobulin plays an important role in the process of resisting cold stimulation[36,37]. In this study, at the beginning of IMCS, the levels of IgG and IgM in CS group and SIgA in jejunum in CS6 were increased, and there was no significant difference in immunoglobulin between CC and CS groups at 43 d. This finding suggests that broilers could increase the secretion of immunoglobulin to improve the intestinal immune function and resist the cold environment at the beginning of cold stimulation. Therefore, when broilers adapt to the environment, the content of immunoglobulin returned to the normal level. Increased barrier tightness during cold stimulation may be one reason for the increase of immunoglobulin levels, as tighter barrier prevents pathogens from invading the intestinal tract and improves the immune function. The increase in immunoglobulin content in the serum of broilers and the improvement of immune function reported by Sang Oh Park when exposed to temperatures 8 °C higher than the control group is consistent with our findings[38]. Carr et al. revealed that the levels of IgG and IgM were improved when mice were submitted cold stress. Thaxton et al. found that cold exposure could accelerate the synthesis rate of IgG of chickens and enhance humoral immunity to a certain extent [39]. The above results match with our results. At the same time, the level of immunoglobulin can also affect production performance[40], which is one of the reasons for the improvement of production performance after IMCS training.
Epithelial cells are arranged on the surface of mucosa. These cells establish a selective permeation barrier between the internal and external environment, responsible for digesting food and absorbing nutrients. The study showed that after ACS, the protein levels of Claudin-1 and Occludin, the mRNA levels of ZO-1 and ZO-2 levels in CS6 group were significantly up-regulated, indicating that increasing the expression levels of TJ genes in broilers of the CS group can increase the barrier tightness during the process of ACS and maintain the stability of intestinal function. Yang et al. found that heat stress for I h could enhance the Claudin-1 levels of jejunum in ducks, this match with our results[18]. Liu et al. revealed that broilers which were subjected 21 days of cold stimulation training could significantly up-regulate the Claudin-1 and ZO-2 levels in the ileum after ACS[21]. It indicated that the cold tolerance of the body could be improved by the IMCS in the early stage. In the present study, E-cadherin and Mucin2 levels in duodenum in CS6 and Mucin2 levels in jejunum in CS were up-regulated after ACS. SIgA plays an important role in the intestinal immune response. Immunoglobulin levels can also affect intestinal barrier function[41]. In the present study, after ACS, the level of SIgA in CS was notably higher than that in CC. These results indicated that the expression level of mucin could be increased by IMCS training in the early stage, and the E-cadherin and Mucin2 could be rapidly up-regulated during ACS, activating the body's protective mechanism, improving the intestinal immune function, preventing microorganisms from invading the intestinal tract, and maintaining intestinal health. This finding is consistent with previous research by Liu et al., who reported that the expression level of Mucin2 in the ileum of broilers which had undergone cold training in the early stage did not change significantly when suffered from ACS, and the mRNA level of IgA increased significantly.[21]. In the present study, there was no significant difference in the content of IgA in the serum, indicating that the body could maintain stability of immune function through immunoregulation. According to Varasteh et al., the level of E-cadherin in jejunum and ileum of broilers significantly increased after heat stress (38-39 °C)[20], further supporting the positive effects of appropriate stimulation. In conclusion, the expression level of mucin can be improved to a certain extent after appropriate cold stimulation training in the early stage, which can effectively block the invasion of pathogenic microorganisms, enhance the immune capacity, and thus improving the ability to resist ACS.
5. Conclusions
The IMCS training had a positive effect on the production performance, the function of intestinal barrier and immunity by upregulating the levels of intestinal barrier genes and SIgA. In the process of ACS, the levels of intestinal barrier genes and SIgA in CS group could be rapidly increased to maintain the integrity of intestinal barrier and overall health of broilers.
Author Contributions
Conceptualization, L.X. and J.L.; methodology, L.X., T.L. and H.W.; software, L.X., T.L. and J.B.; validation, T.L. and Y.Z.; formal analysis, L.X. and T.L.; investigation, T.L. and Y.Z.; resources, T.L.; data curation, Y.Z. and J.B.; writing—original draft preparation, L.X..; writing—review and editing, L.X., T.L., Y.Z., J.B., H.W. and J.L.; visualization, L.X.; supervision, H.W. and J.L.; project administration, J.L.; funding acquisition, J.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (Grant No. 32172785).
Acknowledgments
The authors thank the Animal Behavior and Welfare Laboratory for its technical and human support in Northeast Agricultural University.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Aarif, O.; Shergojry, S. A.; Dar, S. A.; Khan, N.; Mir, N. A.; Sheikh, A. A. , Impact of Cold Stress on Blood Biochemical and Immune Status in Male and Female Vanaraja Chickens. Indian Journal of Animal Research 2014, 48, 139–142. [Google Scholar] [CrossRef]
- Su, Y. Y.; Zhang, X.; Xin, H. W.; Li, S.; Li, J. F.; Zhang, R. X.; Li, X.; Li, J. H.; Bao, J. , Effects of prior cold stimulation on inflammatory and immune regulation in ileum of cold-stressed broilers. Poult Sci 2018, 97, 4228–4237. [Google Scholar] [CrossRef] [PubMed]
- Hu, J. Y.; Cheng, H. W. , Warm perches: a novel approach for reducing cold stress effect on production, plasma hormones, and immunity in laying hens. Poult Sci 2021, 100, 101294. [Google Scholar] [CrossRef]
- Hirata, Y.; Broquet, A. H.; Menchén, L.; Kagnoff, M. F. , Activation of Innate Immune Defense Mechanisms by Signaling through RIG-I/IPS-1 in Intestinal Epithelial Cells. The Journal of Immunology 2007, 179, 5425–5432. [Google Scholar] [CrossRef] [PubMed]
- Melo, A. N. F.; Souza, G. T.; Schaffner, D.; Oliveira, T. C. M.; Maciel, J. F.; Souza, E. L.; Magnani, M. , Changes in thermo-tolerance and survival under simulated gastrointestinal conditions of Salmonella Enteritidis PT4 and Salmonella Typhimurium PT4 in chicken breast meat after exposure to sequential stresses. Int J Food Microbiol 2017, 251, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Theerawatanasirikul, S.; Koomkrong, N.; Kayan, A.; Boonkaewwan, C. , Intestinal barrier and mucosal immunity in broilers, Thai Betong, and native Thai Praduhangdum chickens. Turk J Vet Anim Sci 2017, 41, 357–364. [Google Scholar] [CrossRef]
- Brandtzaeg, P. , The gut as communicator between environment and host: Immunological consequences. Eur J Pharmacol 2011, 668, S16–S32. [Google Scholar] [CrossRef] [PubMed]
- Turner, J. R. , Intestinal mucosal barrier function in health and disease. Nat Rev Immunol 2009, 9, 799–809. [Google Scholar] [CrossRef]
- Ulluwishewa, D.; Anderson, R. C.; McNabb, W. C.; Moughan, P. J.; Wells, J. M.; Roy, N. C. , Regulation of tight junction permeability by intestinal bacteria and dietary components. J Nutr 2011, 141, 769–76. [Google Scholar] [CrossRef]
- Fanning, A. S.; Jameson, B. J.; Jesaitis, L. A.; Anderson, J. M. , The Tight Junction Protein ZO-1 Establishes a Link between the Transmembrane Protein Occludin and the Actin Cytoskeleton. Journal of Biological Chemistry 1998, 273, 29745–29753. [Google Scholar] [CrossRef]
- Catalioto, R. M.; Maggi, C. A.; Giuliani, S. , Intestinal epithelial barrier dysfunction in disease and possible therapeutical interventions. Curr Med Chem 2011, 18, 398–426. [Google Scholar] [CrossRef]
- Krause, G.; Winkler, L.; Mueller, S. L.; Haseloff, R. F.; Piontek, J.; Blasig, I. E. , Structure and function of claudins. Biochim Biophys Acta 2008, 1778, 631–45. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tellez, G.; Richards, J. D.; Escobar, J. , Identification of Potential Biomarkers for Gut Barrier Failure in Broiler Chickens. Front Vet Sci 2015, 2, 14. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Guo, Y.; Wang, Z. , beta-1,3/1,6-Glucan alleviated intestinal mucosal barrier impairment of broiler chickens challenged with Salmonella enterica serovar Typhimurium. Poult Sci 2013, 92, 1764–73. [Google Scholar] [CrossRef] [PubMed]
- Quinteiro-Filho, W. M.; Ribeiro, A.; Ferraz-de-Paula, V.; Pinheiro, M. L.; Sakai, M.; Sa, L. R.; Ferreira, A. J.; Palermo-Neto, J. , Heat stress impairs performance parameters, induces intestinal injury, and decreases macrophage activity in broiler chickens. Poult Sci 2010, 89, 1905–14. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y. F.; Chen, Y. P.; Chen, R.; Su, Y.; Zhang, R. Q.; He, Q. F.; Wang, K.; Wen, C.; Zhou, Y. M. , Dietary mannan oligosaccharide ameliorates cyclic heat stress-induced damages on intestinal oxidative status and barrier integrity of broilers. Poult Sci 2019, 98, 4767–4776. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, Y.; Han, H.; Zhang, L.; Wang, T. , Bisdemethoxycurcumin attenuates lipopolysaccharide-induced intestinal damage through improving barrier integrity, suppressing inflammation, and modulating gut microbiota in broilers. J Anim Sci 2021, 99. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Liu, B.; Wang, Y.; Huang, X.; Yan, Z.; Jiang, Q.; Chen, Q. , Ellagic Acid Improves Antioxidant Capacity and Intestinal Barrier Function of Heat-Stressed Broilers via Regulating Gut Microbiota. Animals (Basel) 2022, 12. [Google Scholar] [CrossRef]
- Zhao, F. Q.; Zhang, Z. W.; Yao, H. D.; Wang, L. L.; Liu, T.; Yu, X. Y.; Li, S.; Xu, S. W. , Effects of cold stress on mRNA expression of immunoglobulin and cytokine in the small intestine of broilers. Res Vet Sci 2013, 95, 146–55. [Google Scholar] [CrossRef]
- Varasteh, S.; Braber, S.; Akbari, P.; Garssen, J.; Fink-Gremmels, J. , Differences in Susceptibility to Heat Stress along the Chicken Intestine and the Protective Effects of Galacto-Oligosaccharides. PLoS One 2015, 10, e0138975. [Google Scholar] [CrossRef]
- Liu, X. T.; Li, S.; Zhao, N.; Xing, L.; Gong, R. X.; Li, T. T.; Zhang, S. J.; Li, J. H.; Bao, J. , Effects of Acute Cold Stress after Intermittent Cold Stimulation on Immune-Related Molecules, Intestinal Barrier Genes, and Heat Shock Proteins in Broiler Ileum. Animals (Basel) 2022, 12. [Google Scholar] [CrossRef]
- Tang, J.; Chen, Z. , The protective effect of gamma-aminobutyric acid on the development of immune function in chickens under heat stress. J Anim Physiol Anim Nutr (Berl) 2016, 100, 768–77. [Google Scholar] [CrossRef]
- Xue, G.; Yin, J.; Zhao, N.; Liu, Y.; Fu, Y.; Zhang, R.; Bao, J.; Li, J. , Intermittent mild cold stimulation improves the immunity and cold resistance of spleens in broilers. Poult Sci 2021, 100, 101492. [Google Scholar] [CrossRef]
- Leeson, S. Nutritional considerations of poultry during heat stress. World's Poultry Science Journal 1986, 42, 69–81. [Google Scholar] [CrossRef]
- Mashaly, M. M.; Hendricks, G. L., 3rd; Kalama, M. A.; Gehad, A. E.; Abbas, A. O.; Patterson, P. H. , Effect of heat stress on production parameters and immune responses of commercial laying hens. Poult Sci 2004, 83, 889–94. [Google Scholar] [CrossRef]
- Sagher, B. M. , The effect of cold stress on muscle growth in young chicks. Growth 1975, 39, 281–8. [Google Scholar] [PubMed]
- Blahová, J.; Dobšíková, R.; Straková, E.; Suchý, P. , Effect of Low Environmental Temperature on Performance and Blood System in Broiler Chickens (Gallus domesticus). Acta Veterinaria Brno 2007, 76, S17–S23. [Google Scholar] [CrossRef]
- Shinder, D. L., D. Rusal, M. Rzepakovsky, V. Bresler, V. Yahav, S., Early age cold conditioning in broiler chickens(Gallus domesticus)_ thermotolerance and growth responses. J Therm Biol 2002, 27, 517–523. [CrossRef]
- Dokladny, K.; Moseley, P. L.; Ma, T. Y. , Physiologically relevant increase in temperature causes an increase in intestinal epithelial tight junction permeability. Am J Physiol Gastrointest Liver Physiol 2006, 290, G204–12. [Google Scholar] [CrossRef]
- Zhou, H. J.; Kong, L. L.; Zhu, L. X.; Hu, X. Y.; Busye, J.; Song, Z. G. , Effects of cold stress on growth performance, serum biochemistry, intestinal barrier molecules, and adenosine monophosphate-activated protein kinase in broilers. Animal 2021, 15, 100138. [Google Scholar] [CrossRef]
- Umeda, K.; Ikenouchi, J.; Katahira-Tayama, S.; Furuse, K.; Sasaki, H.; Nakayama, M.; Matsui, T.; Tsukita, S.; Furuse, M.; Tsukita, S. , ZO-1 and ZO-2 independently determine where claudins are polymerized in tight-junction strand formation. Cell 2006, 126, 741–54. [Google Scholar] [CrossRef] [PubMed]
- Cano, A.; Perez-Moreno, M. A.; Rodrigo, I.; Locascio, A.; Blanco, M. J.; del Barrio, M. G.; Portillo, F.; Nieto, M. A. , The transcription factor snail controls epithelial-mesenchymal transitions by repressing E-cadherin expression. Nat Cell Biol 2000, 2, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Peterson, L. W.; Artis, D. , Intestinal epithelial cells: regulators of barrier function and immune homeostasis. Nat Rev Immunol 2014, 14, 141–53. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Shao, D.; Wu, S.; Song, Z.; Shi, S. , Heat stress-induced intestinal barrier damage and dimethylglycine alleviates via improving the metabolism function of microbiota gut brain axis. Ecotoxicol Environ Saf 2022, 244, 114053. [Google Scholar] [CrossRef] [PubMed]
- Fihn, B. M.; Sjoqvist, A.; Jodal, M. , Permeability of the rat small intestinal epithelium along the villus-crypt axis: effects of glucose transport. Gastroenterology 2000, 119, 1029–36. [Google Scholar] [CrossRef]
- Woof, J. M.; Kerr, M. A. , The function of immunoglobulin A in immunity. J Pathol 2006, 208, 270–82. [Google Scholar] [CrossRef]
- Carr, D. J.; Woolley, T. W.; Blalock, J. E. , Phentolamine but not propranolol blocks the immunopotentiating effect of cold stress on antigen-specific IgM production in mice orally immunized with sheep red blood cells. Brain Behav Immun 1992, 6, 50–63. [Google Scholar] [CrossRef]
- Choi, S.-O. P. J. H. C.-M. R. B.-S. P. Y.-H., Effects of extreme heat stress on growth performance, lymphoid organ, IgG and cecum microflora of broiler chickens. International Journal of Agriculture & Biology 2013, 15, 120.
- Thaxton, P. , Influence of temperature on the immune response of birds. Poult Sci 1978, 57, 1430–40. [Google Scholar] [CrossRef]
- Chen, W.; Yin, C.; Li, J.; Sun, W.; Li, Y.; Wang, C.; Pi, Y.; Cordero, G.; Li, X.; Jiang, X. , Stimbiotics Supplementation Promotes Growth Performance by Improving Plasma Immunoglobulin and IGF-1 Levels and Regulating Gut Microbiota Composition in Weaned Piglets. Biology (Basel) 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Lin, J.; Pan, T.; Li, T.; Jiang, H.; Fang, Y.; Wang, Y.; Wu, F.; Huang, J.; Zhang, H.; Chen, D.; Chen, Y. , Polymeric immunoglobulin receptor deficiency exacerbates autoimmune hepatitis by inducing intestinal dysbiosis and barrier dysfunction. Cell Death Dis 2023, 14, 68. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The specific experimental temperature scheme.
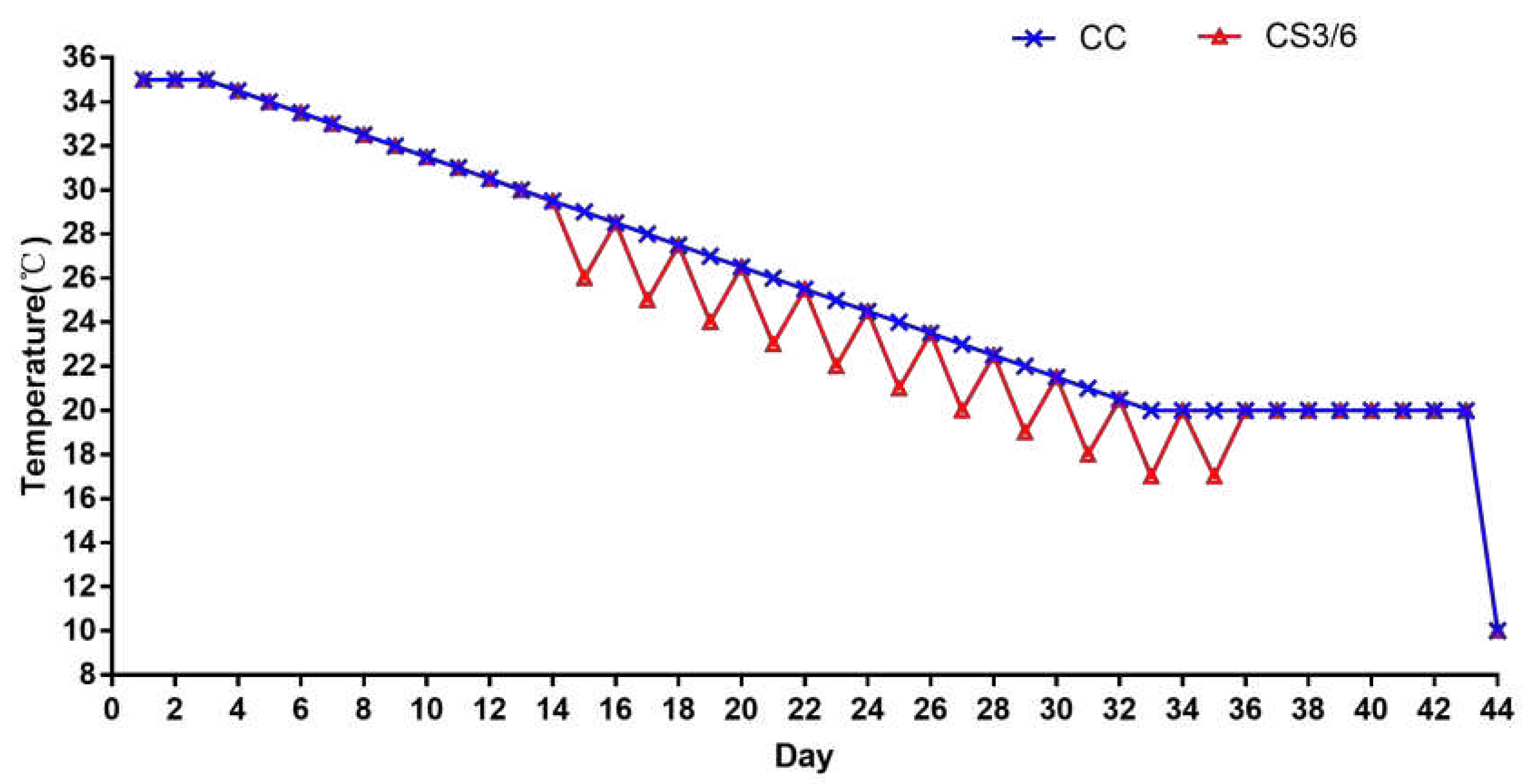
Figure 2.
Effects of cold stimulation on mRNA levels of intestinal barrier genes in duodenum. Values are means ± SD. Different letters indicate significant differences among the treatment (a, b, c) (P < 0.05). P values among groups are respectively: Claudin-1 (22 d: 0.000; 36 d: 0.076; 43 d: 0.001; Y6: 0.002); E-cadherin (22 d: 0.013; 36 d: 0.133; 43 d: 0.043; Y6: 0.008); Occludin (22 d: 0.000; 36 d: 0.009; 43 d: 0.000; Y6: 0.058); ZO-1 (22 d: 0.003; 36 d: 0.556; 43 d: 0.012; Y6: 0.005); ZO-2 (22 d: 0.000; 36 d: 0.001; 43 d: 0.001; Y6: 0.007); Mucin2 (22 d: 0.001; 36 d: 0.005; 43 d: 0.027; Y6: 0.004).
Figure 2.
Effects of cold stimulation on mRNA levels of intestinal barrier genes in duodenum. Values are means ± SD. Different letters indicate significant differences among the treatment (a, b, c) (P < 0.05). P values among groups are respectively: Claudin-1 (22 d: 0.000; 36 d: 0.076; 43 d: 0.001; Y6: 0.002); E-cadherin (22 d: 0.013; 36 d: 0.133; 43 d: 0.043; Y6: 0.008); Occludin (22 d: 0.000; 36 d: 0.009; 43 d: 0.000; Y6: 0.058); ZO-1 (22 d: 0.003; 36 d: 0.556; 43 d: 0.012; Y6: 0.005); ZO-2 (22 d: 0.000; 36 d: 0.001; 43 d: 0.001; Y6: 0.007); Mucin2 (22 d: 0.001; 36 d: 0.005; 43 d: 0.027; Y6: 0.004).
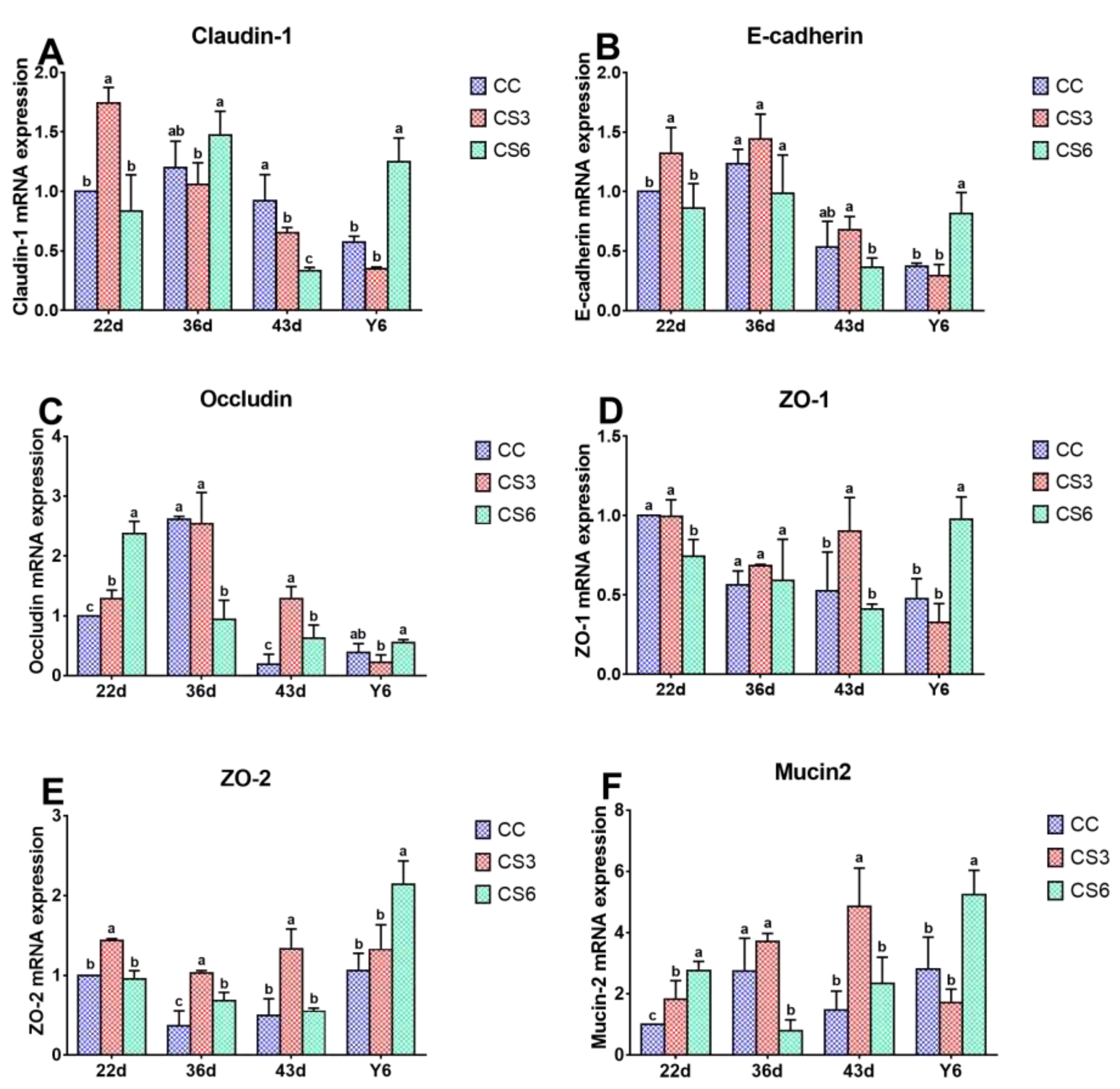
Figure 3.
Effects of cold stimulation on protein levels of intestinal barrier genes in duodenum. Values are means ± SD. Different letters indicate significant differences among the treatment (a, b, c) (P < 0.05). P values among groups are respectively: Claudin-1 (22 d: 0.000; 36 d: 0.000; 43 d: 0.000; Y6: 0.000); E-cadherin (22 d: 0.000; 36 d: 0.000; 43 d: 0.000; Y6: 0.000); Occludin (22 d: 0.069; 36 d: 0.000; 43 d: 0.000; Y6: 0.000).
Figure 3.
Effects of cold stimulation on protein levels of intestinal barrier genes in duodenum. Values are means ± SD. Different letters indicate significant differences among the treatment (a, b, c) (P < 0.05). P values among groups are respectively: Claudin-1 (22 d: 0.000; 36 d: 0.000; 43 d: 0.000; Y6: 0.000); E-cadherin (22 d: 0.000; 36 d: 0.000; 43 d: 0.000; Y6: 0.000); Occludin (22 d: 0.069; 36 d: 0.000; 43 d: 0.000; Y6: 0.000).
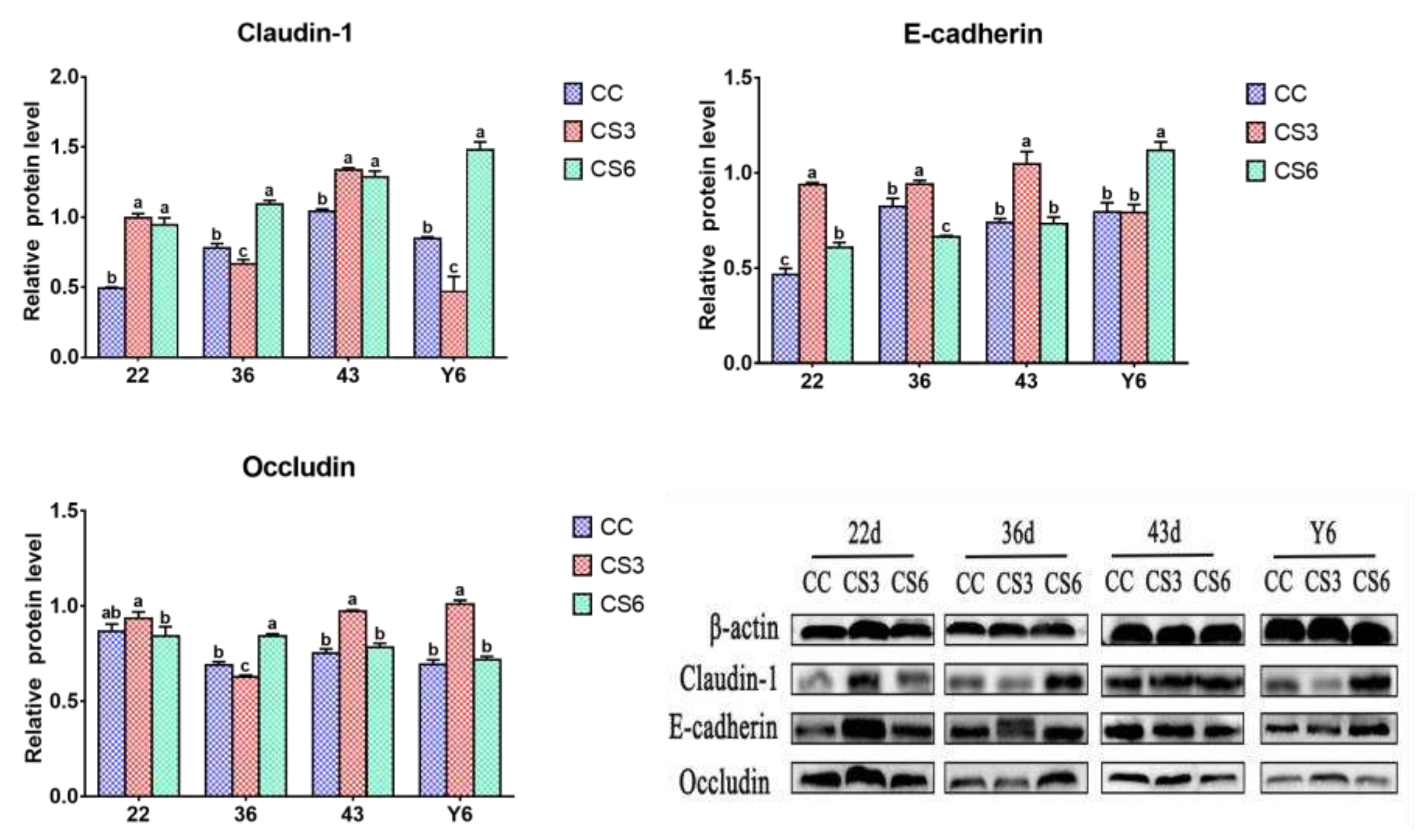
Figure 4.
Effects of cold stimulation on mRNA levels of intestinal barrier genes in jejunum. Values are means ± SD. Different letters indicate significant differences among the treatment (a, b, c) (P < 0.05). P values among groups are respectively: Claudin-1 (22 d: 0.001; 36 d: 0.070; 43 d: 0.000; Y6: 0.282); E-cadherin (22 d: 0.002; 36 d: 0.639; 43 d: 0.034; Y6: 0.539); Occludin (22 d: 0.000; 36 d: 0.006; 43 d: 0.122; Y6: 0.759); ZO-1 (22 d: 0.002; 36 d: 0.172; 43 d: 0.039; Y6: 0.188); ZO-2 (22 d: 0.098; 36 d: 0.382; 43 d: 0.159; Y6: 0.453); Mucin2 (22 d: 0.000; 36 d: 0.023; 43 d: 0.111; Y6: 0.032).
Figure 4.
Effects of cold stimulation on mRNA levels of intestinal barrier genes in jejunum. Values are means ± SD. Different letters indicate significant differences among the treatment (a, b, c) (P < 0.05). P values among groups are respectively: Claudin-1 (22 d: 0.001; 36 d: 0.070; 43 d: 0.000; Y6: 0.282); E-cadherin (22 d: 0.002; 36 d: 0.639; 43 d: 0.034; Y6: 0.539); Occludin (22 d: 0.000; 36 d: 0.006; 43 d: 0.122; Y6: 0.759); ZO-1 (22 d: 0.002; 36 d: 0.172; 43 d: 0.039; Y6: 0.188); ZO-2 (22 d: 0.098; 36 d: 0.382; 43 d: 0.159; Y6: 0.453); Mucin2 (22 d: 0.000; 36 d: 0.023; 43 d: 0.111; Y6: 0.032).
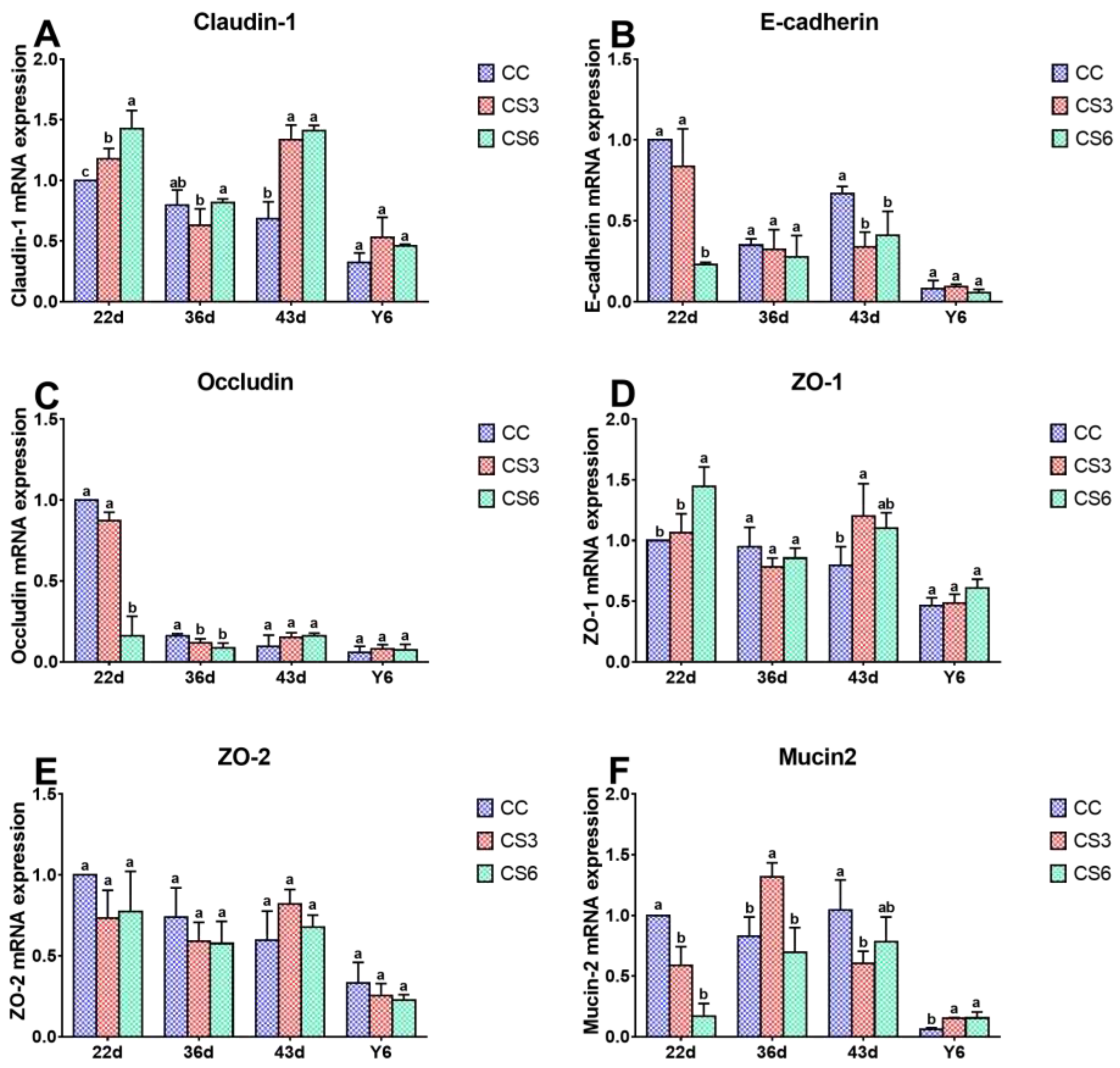
Figure 5.
Effects of cold stimulation on protein levels of intestinal barrier genes in jejunum. Values are means ± SD. Different letters indicate significant differences among the treatment (a, b, c) (P < 0.05). P values among groups are respectively: Claudin-1 (22 d: 0.000; 36 d: 0.004; 43 d: 0.000; Y6: 0.000); E-cadherin (22 d: 0.000; 36 d: 0.000; 43 d: 0.000; Y6: 0.000); Occludin (22 d: 0.006; 36 d: 0.000; 43 d: 0.003; Y6: 0.000).
Figure 5.
Effects of cold stimulation on protein levels of intestinal barrier genes in jejunum. Values are means ± SD. Different letters indicate significant differences among the treatment (a, b, c) (P < 0.05). P values among groups are respectively: Claudin-1 (22 d: 0.000; 36 d: 0.004; 43 d: 0.000; Y6: 0.000); E-cadherin (22 d: 0.000; 36 d: 0.000; 43 d: 0.000; Y6: 0.000); Occludin (22 d: 0.006; 36 d: 0.000; 43 d: 0.003; Y6: 0.000).
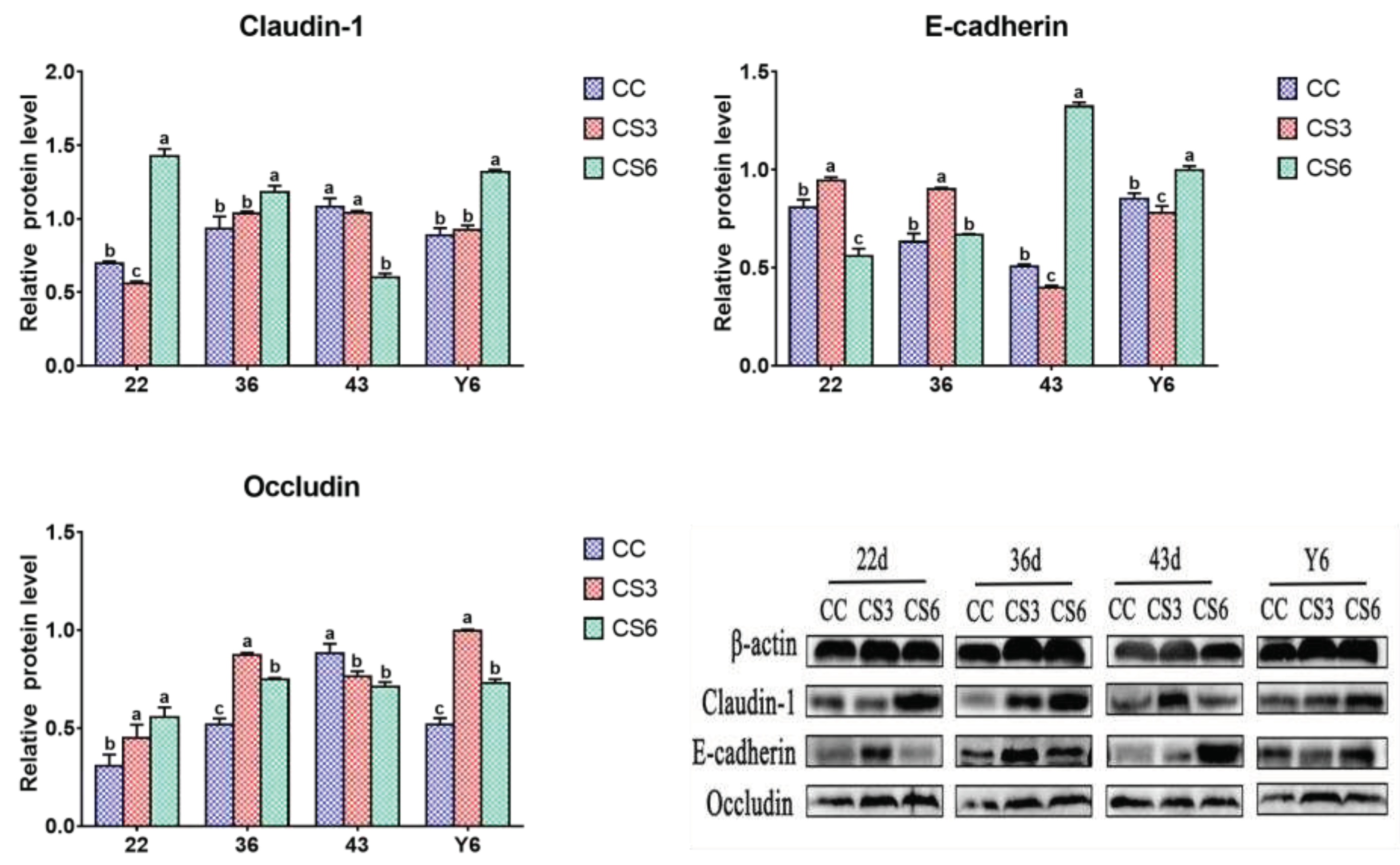
Figure 6.
Effects of IMCS and ACS on the content of immunoglobulin in serum, duodenum and jejunum. Values are means ± SD. Different letters indicate significant differences among the treatment (a, b) (P < 0.05). P values among groups are respectively: IgA (22 d: 0.938; 36 d: 0.316; 43 d: 0.991); IgG (22 d: 0.035; 36 d: 0.022; 43 d: 0.560); IgM (22 d: 0.035; 36 d: 0.770; 43 d: 0.263); SIgA in duodenum (22 d: 0.367; 36 d: 0.106; 43 d: 0.304; Y6: 0.015); SIgA in jejunum (22 d: 0.167; 36 d: 0.019; 43 d: 0.105; Y6: 0.019).
Figure 6.
Effects of IMCS and ACS on the content of immunoglobulin in serum, duodenum and jejunum. Values are means ± SD. Different letters indicate significant differences among the treatment (a, b) (P < 0.05). P values among groups are respectively: IgA (22 d: 0.938; 36 d: 0.316; 43 d: 0.991); IgG (22 d: 0.035; 36 d: 0.022; 43 d: 0.560); IgM (22 d: 0.035; 36 d: 0.770; 43 d: 0.263); SIgA in duodenum (22 d: 0.367; 36 d: 0.106; 43 d: 0.304; Y6: 0.015); SIgA in jejunum (22 d: 0.167; 36 d: 0.019; 43 d: 0.105; Y6: 0.019).
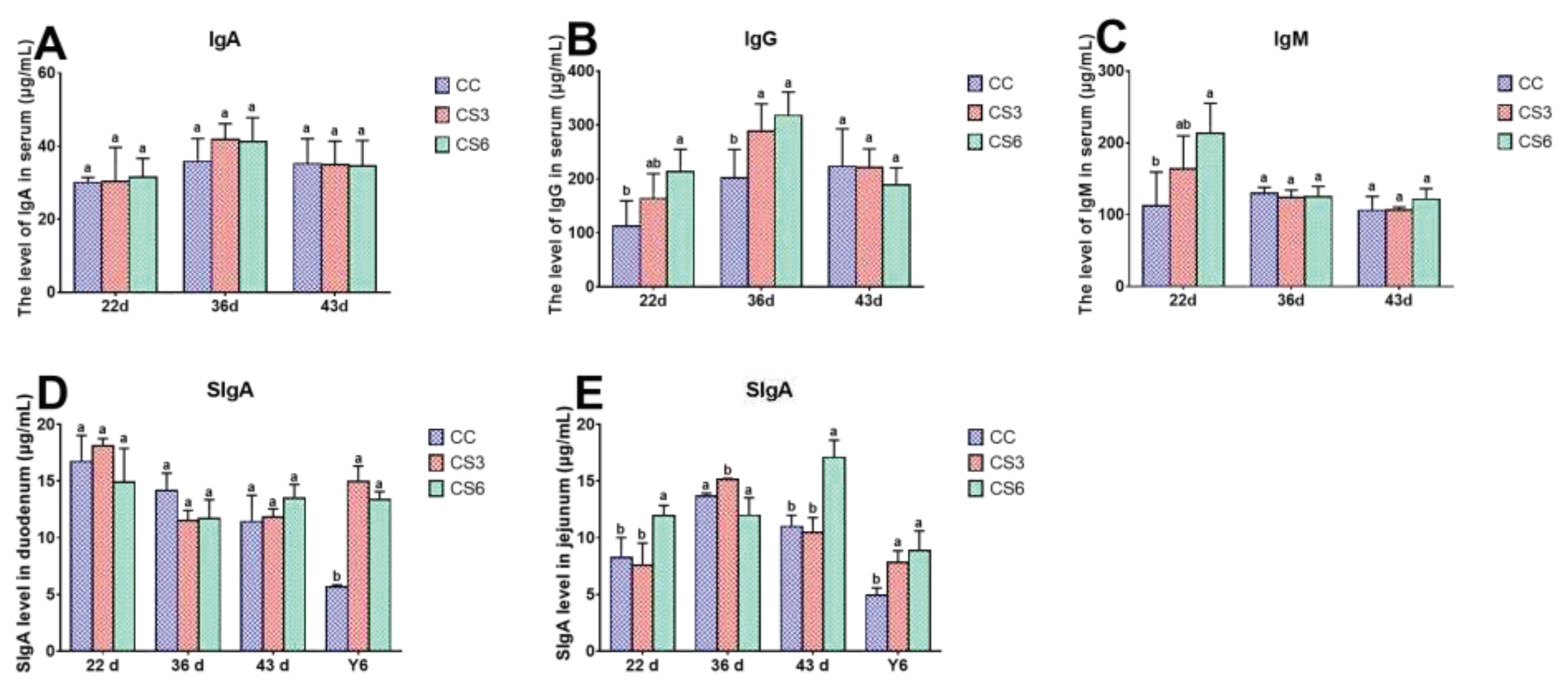

Table 1.
Reverse transcriptional reaction system.
| Reagent | Usage |
| 5×gDNA Eraser Buffer | 2.0µl |
| gDNA Eraser | 1.0µl |
| Total RNA | 1µg |
| RNase Free dH2O up to | 10.0µl |
| 42 °C Water-bath for 2min | |
| 5×PrimeScript Buffer 2 | 4.0µl |
| RT Primer Mix | 1.0µl |
| PrimeScript RT Enzyme Mix I | 1.0µl |
| RNase Free dH2O | 4.0µl |
| 37 °C Water-bath for 15 min, then 85 °C Water-bath for 5 s | |
Table 2.
Gene-special primer sequences used for the study.
| Gene | Gene Reference Sequence | Primer Sequences (5’-3’) |
| β-actin | NM_205518.1 | Forward: CACCACAGCCGAGAGAGAAAT |
| Reverse: TGACCATCAGGGAGTTCATAGC | ||
| Claudin-1 | NM_001013611.2 | Forward: TGGAGGATGACCAGGTGAAGA |
| Reverse: CGAGCCACTCTGTTGCCATA | ||
| E-cadherin | NM 001039258.2 | Forward: GACAGGGACATGAGGCAGAA |
| Reverse: GCCGTGACAATGCCATTCTC | ||
| Occludin | NM 205128.1 | Forward: TCATCGCCTCCATCGTCTAC |
| Reverse: TCTTACTGCGCGTCTTCTGG | ||
| ZO-1 | XM 413773.4 | Forward: TGTAGCCACAGCAAGAGGTG |
| Reverse: CTGGAATGGCTCCTTGTGGT | ||
| ZO-2 | XM_025144669.1 | Forward: CGGCAGCTATCAGACCACTC |
| Reverse: CACAGACCAGCAAGCCTACAG | ||
| Mucin2 | XM_421035 | Forward: CAGCACCAACTTCTCAGTTC |
| Reverse: TCTGCAGCCACACATTCTTT |
Table 3.
Effect of IMCS on production performance of broilers.
| CC | CS3 | CS6 | P value | |
| Feed conversion ratio | 2.01±0.07a | 1.84±0.06b | 1.98±0.07a | 0.000 |
| Daily feed intake (kg) | 1.21±0.37a | 1.21±0.36a | 1.24±0.40a | 0.163 |
| Daily weight gain (kg) | 0.60±0.14b | 0.65±0.15a | 0.62±0.14b | 0.008 |
Values are means ± SD. Values with different lowercase letters are significantly different (P < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



