
Submitted:
08 June 2023
Posted:
09 June 2023
You are already at the latest version
Abstract
Nowadays, functional foods supplemented with health-promoting microorganisms have attracted the consumers attention due to their health benefits. However, maintaining high cell loads, which consists of an essential requirement for conferring the health effect is a real bottleneck for the food industry, due to viability decline during food processing and storage. Hence, freeze-drying and cell immobilization have been suggested to enhance cell viability. The aim of our study was to assess the effect of freeze-dried immobilized P. acidilactici ORE5 on pistachio nuts on functional regulation of Katiki Domokou-type cheese microbiome. Supplementation of Katiki Domokou-type cheese with free or immobilized P. acidilactici ORE5 culture resulted in cell loads > 8.5 logcfu/g up to 7 days of storage. Both free and immobilized P. acidilactici ORE5 cells suppressed the growth of L. monocytogenes after deliberate inoculation, acting as a protecting shield. SPME GC/MS analysis showed that incorporation of P. acidilactici ORE5 culture in cheese resulted in an improved volatile compounds profile, as verified by the preliminary sensory evaluation. According to Next-Generation Sequencing analysis, a wide range of bacterial diversity was revealed among samples. The most abundant genus was Lactococcus in all samples, while the results showed increased presence of Pediococcus spp. in cheese fortified with P. acidilactici ORE5 culture, highlighting the ability of the strain to survive in the final product. Furthermore, the incorporation of P. acidilactici ORE5 culture in cheese had a significant impact on cheese microbiome composition, as the presence of spoilage bacteria, such as Chryseobacterium, Acinetobacter and Pseudomonas, were significantly lower compared to the control cheese, indicating quality improvement and prolongation of the product’s shelf- life.
Keywords:
Pediococcus acidilactici ORE5
; Katiki Domokou cheese
; probiotic fortification
; biopreservation
1. Introduction
Nowadays, an upsurge of interest in functional foods is witnessed, as consumers have become more conscious of their health and seek for dietary choices that offer additional benefits beyond basic nutrition. Functional foods consist of a variety of components, such as vitamins, essential fatty acids, antioxidants, prebiotics and probiotics. Foods enriched with probiotic microorganisms are considered the main part of functional foods market. Defined by the FAO/WHO as “microorganisms (bacteria or yeasts) which, when administered in adequate concentrations, provide health benefits to the host”, probiotics are gaining increasing scientific and industrial interest [1]. The last years, there have been numerous reports on the biological effects exhibited by probiotics, including their role in digestion, production of immune-modulating bioactive compounds, detoxification of toxins, and influence on the activity of the gut-brain axis [2].
In order to confer the health benefits, probiotic foods should contain high cell able to survive during gastrointestinal tract (GI) passage and colonize the gut epithelium. According to the International Probiotic Association (IPA), the recommended cell levels in the foods are 107 cfu/ g at the time of consumption, in order to achieve a daily dose of 109 cells, considering consumption of 100g product. However, maintaining such high cell loads during food processing and storage consists of a real bottleneck for the food industry. Hence, cell immobilization is recommended as an alternative solution to overcome such obstacles, as it has been reported that it may enhance cell survival rates and ensure cell stability [3,4]. In addition, considering that the food industry favors dried products due to their prolonged shelf life, ease of storage and transportation, and cost-effectiveness resulting from the elimination of refrigeration needs, freeze-drying is commonly employed for delicate ingredients that require special care [5].
The selection of suitable food carrier is of great importance, as it has a significant impact on the adhesion of cells and their ability to colonize the gut effectively [6], along with the sensory properties. Prebiotics, as non-digestible compounds that stimulate the growth or activity of beneficial bacteria in the gut, such as dietary fibers, seem a promising option. Pistachio nuts (Pistacia vera L contain dietary fibers and their administration to diabetic rats resulted in functional modulation of gut microbiota [7] and beneficial alteration in fatty acids profile [8].
Dairy products and especially cheese have been commonly served as food vehicles for probiotics to the human gut, due to their chemical and physical attributes, such as a low pH value and high titratable acidity, contributing to enhanced transit tolerance of bacteria and facilitating the effective delivery of probiotics to the GI tract [9]. Katiki Domokou, also known as Tsalafouti, is a traditional soft cheese manufacturing in Domokos area in Greece from goat’s and ewe’s milk, officially recognized as a Protected Designation of Origin (PDO) since 1994. The process begins by pasteurizing and cooling the milk at temperatures 27 - 28°C. Coagulation is then achieved with or without the addition of rennet, followed by a resting period at temperatures ranging from 20 - 22°C. The resulting curd is subsequently filtered and placed into cloth sacks to allow for drainage. Notably, in the production of the cheese, no commercial starter cultures are usually employed. Instead, the manufacturing process relies mainly on the natural acid fermentation of the milk, facilitated by the microorganisms that are naturally present in the milk. By incorporating fresh product into the same vessel during the cheese-making process, it is possible to cultivate and enhance a particular indigenous microflora that contributes to the milk's acidification during the ripening phase. This practice helps to establish a desired microbial population that promotes the development of specific flavors and textures in the cheese as it matures. The final product retains a high moisture content (approximately 75%), low levels of salt (around 1%) and maintains a pH 4.3 - 4.5. Storage is conducted at temperatures of 4 - 5°C for up to 4 days [10, 11].
The microbiota of dairy products, such as cheese, consist of starter or adjunct cultures, as well as non- starter lactic acid bacteria [12]. The incorporation of functional cultures in dairy products has a profound impact on their microbiome, particularly the composition, functionality, and sensory characteristics. More specifically, lactic acid bacteria ferment lactose and produce lactic acid, creating an acidic environment that inhibits the growth of undesirable microorganisms, while promoting their growth, which are well-adapted to thrive in this environment [12]. Notably, when proper safety measures are neglected during the production process (improper pasteurization, post-pasteurization contamination, etc), cheese constitutes an optimal environment for the proliferation of pathogenic bacteria, including Salmonella spp., Listeria monocytogenes, and Escherichia coli [13]. Moreover, soft and semi-soft cheese products, are susceptible to the growth of molds and bacteria that produce harmful toxins [14]. Microbial hazards have a great economical and legal impact on food industry, intensifying the need of preservatives. Considering the consumers demand for natural alternatives, several studies have suggested the use of lactic acid bacteria as biopreservatives [15]. Additionally, they contribute to the breakdown of proteins and lipids, releasing flavor-enhancing compounds and enzymes that influence the sensory attributes of the cheese. Mechanistically, lactic acid bacteria adhere to the surface of cheese curds, forming biofilms that protect against contamination and provide an environment for microbial interactions. They also produce exopolysaccharides, contributing to the cheese's texture and rheological properties [16]. Hence, the incorporation of probiotics in cheese production not only enhances nutritional value and promotes health benefits but also shapes the microbial dynamics, ultimately influencing the quality, flavor, and texture of the cheese.
Several lactic acid bacteria, such as Lactococcus, Lactobacillus, and Streptococcus species, are commonly employed as starter cultures in cheese production [12]. However, for the selection of a suitable strain for the production of functional food, several criteria should be considered regarding its potential beneficial properties. In this vein, P. acidilactici ORE5, a wild- type strain isolated from kefir, was initially evaluated for potential probiotic characteristics and subsequently, it was tested for the production of a novel functional Katiki Domokou type cheese enriched with immobilized P. acidilactici ORE5 on pistachio nuts, assessing cell survival, along with the physicochemical and microbiome changes during storage. Moreover, potential growth suppression of L. monocytogenes was investigated after deliberate contamination, a food-borne pathogen usually associated with cheese safety.
2. Materials and Methods
2.1. Microbial cultures
Pediococcus acidilactici ORE5 was isolated from kefir grains and species identification was based on 16S rRNA sequencing, according to Li et al [17]. The results were compared with the sequences deposited in the GenBank database using BLAST analysis on the National Center for Biotechnology Information (NCBI).
Lactiplantibacillus plantarum ATCC 14917 was obtained from ATCC (LGC Standards, Middlesex, UK). Both P. acidilactici ORE5 and L. plantarum ATCC 14917 were grown on de Man, Rogosa and Sharpe (MRS) broth (Condalab) at 37°C for 24h. The medium was sterilized at 121°C for 15 minutes prior use.
Listeria monocytogenes NCTC 10527 serotype 4b (kindly provided by the Laboratory of Clinical Microbiology, Sismanoglio General Hospital, Athens, Greece) was grown in Brain Heart Infusion (BHI) broth (Condalab, Madrid, Spain) at 37°C for 24 h. The medium was sterilized at 121°C for 15 minutes before use.
2.2. In vitro screening for probiotic properties
2.2.1. Resistance to low pH, pepsin, pancreatin and tolerance to bile salts
The assessment of resistance of bacterial cells to pH 2.0, 3.0 or 4.0, as well as the tolerance to bile salts, was carried out, according to Argyri et al. [25]. Regarding the resistance of the bacterial cells to pepsin and pancreatin, a method described by Plessas et al. [54] was performed.
2.2.2. Antibiotic susceptibility
The determination of the antibiotic susceptibility was assessed using the M.I.C. Evalu-ator® strips [2018] and expressed as Minimum Inhibitory Concentration (MIC), following the method described by Plessas et al. [54].
2.3. Production of freeze-dried immobilized P. acidilactici ORE5 on pistachio nuts
Cell immobilization on pistachio nuts was carried out as described previously [3]. The immobilized cells were transferred to -80°C overnight and freeze-dried on a BenchTop Pro (Virtis, SP Scientific, Warminster, PA, USA) for 24h at ~30-35 Pa with the condenser temperature at -100°C. For comparison reasons, free freeze-dried Pediococcus acidilactici ORE5 cells were also produced.
2.4. Functional Katiki Domokou type cheese production
Katiki Domokou cheese (“Oreines plagies”, Domokos, Greece) (pasteurized goat and sheep milk, 13% fat, 1% salt) purchased from market was used was enriched with freeze-dried immobilized P. acidilactici ORE5 cells on pistachio nuts pieces (sample KIP), resulting in 8 logcfu/g of cheese. For comparison reasons, Katiki Domokou cheese containing free freeze-dried cells (sample KFP) was also prepared by incorporating in cheese resulting in same levels as KIP sample. Additionally, regular commercial product with no Pediococcus acidilactici ORE5 cells (sample KC) was used as control. All products were stored at refrigeration temperature (4°C) for 14 days, in order to evaluate the product’s self-life. Samples were taken after 3 days and subjected to physicochemical, microbiological and molecular analyses.
2.5. Resistance to spoilage assessment
All products were deliberately inoculated with Listeria monocytogenes (inoculum 105 cfu/g) and stored at refrigeration temperature (4°C) for 14 days in order to monitor cell levels.
2.6. Physicochemical Analysis
2.7. Microbiological Analyses
2.7.1. Monitoring Pediococcus acidilactici ORE5 cell viability
In order to monitor levels of free or immobilized P. acidilactici ORE5 cells, 10g of cheese were blended with 90 mL sterile ¼ Ringer’s solution, decimal diluted, plated on MRS agar and incubated at 37°C for 72h. Cell levels were expressed as logcfu/g of cheese.
2.7.2. Determination of cheese microbiota
Total aerobic counts (TAC), total psychrophilic counts, lactococci, yeasts/ molds, staphylococci, Enterobacteriacae, coliforms and Listeria monocytogenes were determined as described by Nikolaou et al. [4].
2.8. Minor component analysis by HS-SPME GC/MS
Katiki Domokou cheese samples collected at the beginning and after 3 days of storage were analyzed for minor volatiles on an HS-SPME GC/MS system [6890N GC, 5973 NetworkedMS MSD, HP-5MS column (30 m, 0.25 mm i.d., 0.25 μm film thickness), Agilent Technologies, Santa Clara, CA, USA] and semi-quantified using 4-methyl-2-pentanol. Sample analysis was carried out as previously described [19].
2.9. DNA Extraction, PCR Amplification and 16S rRNA Sequencing
Total DNA was extracted using the NucleoSpin® Food (MACHEREY-NAGEL GmbH & Co.KG, Germany), following the manufacturer’s instructions. The isolated DNA was quantified using Nanodrop 2000c Spectrophotometer (Thermo Fisher Scientific). A PCR reaction targeting V3 region of the 16S rDNA gene were carried for each sample, using the primers 5’-ACTGAGACACGGTCCAGACT-3’ (F) and 5’-GTATTACCGCGGCTGCTG-3’ ®. For each reaction, 100 ng of DNA were mixed with 10 μL of KAPA HiFi HotStart ReadyMix (Roche), 4 μL of primers (final concentration 0.2 μΜ) and proper volume of Nuclease Free water to a final volume of 20 μL. PCR was carried out under the following conditions - Initial denaturation at 95 oC (5’), followed by 25 cycles of denaturation at 95 oC (30s), annealing at 58 oC (30s) and extension at 72 oC (15s). PCR products were checked in a 2% w/v agarose gel and then purified using magnetic Agencourt AMPure XP beads (Beckman Coulter) according to the manufacturer’s instructions. 75ng of purified PCR products were used for the preparation of the libraries with the Ion Plus Fragment Library Kit (Thermo Fischer Scientific). The libraries were then quantified and prepared for sequencing by the Ion Torrent GeneStudio™ S5 (Thermo Fischer Scientific).
Raw sequencing data were analyzed in order to filter out low-quality reads. Data analysis for amplicon sequencing was performed using Mothur (v.1.45.3) [20]. OTUs (Operational Taxonomical Units) were defined by clustering reads at 3% divergence (97% similarity), using the Greengenes (v.13_8_99) database. The calculation of Shannon’s and Simpson’s indices were performed using the Rhea platform.
2.10. Preliminary sensory evaluation
Katiki Domokou cheese samples were evaluated for their quality characteristics using a previously approved protocol in our laboratory [21]. A mixed panel of 11 random tasters familiar with cheese products were asked to evaluate the samples asking to provide scores on a 0-5 scale (0: unacceptable, 5: exceptional), regarding aroma (fruity, milklike, spirituous, yeasty-breadlike, piquant), texture (watery, creamy, soft, gelatinous, curdy, lumpy, ropy), taste (too acid, acid, bitter, scorched, foreign, sweet, sweet and sour, metallic, salted) and overall quality. During the evaluation, tasters consumed water and crackers between samples. Samples were coded using randomly chosen 3-digit numbers.
2.11. Statistical Analysis
The experiments were conducted in triplicate, and statistical significance was determined at a significance level of P < 0.05. ANOVA and Duncan's multiple range test were employed to analyze the results for statistical significance, and to identify significant differences among the outcomes. The computations for coefficients, ANOVA tables, and significance (P < 0.05) were carried out using Statistica version 10.0 (StatSoft Inc., Tulsa, OK).
3. Results
3.1. Molecular identification
The strain was previously isolated from kefir grains and according to identification based on 16S rRNA sequencing the isolated strain was characterized as Pediococcus acidilactici.
3.2. In vitro screening for probiotic properties
The results of the in vitro assessment for potential probiotic properties of P. acidilactici ORE5 are shown in Table 1. In particular, the resistance to low pH was evaluated and at pH 4, P. acidilactici ORE5 exhibited high survival rates. Yet, a significant reduction of cell viability was noted at pH 2. However, P. acidilactici ORE5 maintained high survival level in high levels (6.4 log cfu/mL). Concerning tolerance to pancreatin, P. acidilactici ORE5 retained its viability at high levels, while resistance to bile salts after 4 h of exposure, was recorded.
3.3. Safety profile - antibiotic susceptibility
The Minimum Inhibitory Concentration (MIC) was estimated based on the diffusion in Mueller-Hinton agar plates, and their mean values ± standard deviation is presented in Table 2. In brief, P. acidilactici ORE5 displayed resistance to clindamycin, erythromycin and tetracycline.
3.4. Physicochemical Characteristics of functional Katiki Domokou type cheese
The physicochemical parameters ranged in usual levels, as published previously (pH 4.3-4.5, moisture 75%) [10, 23] and the results are presented in Table 3. In cheese samples fortified with free or immobilized P. acidilactici ORE5 cells the pH ranged in lower levels (P < 0.05) (4.08-4.15), while lactic acid concentrations were higher (P < 0.05) (0.11-0.15 g lactic acid/ 100 g cheese) compared to the control (pH 4.28-4.33, 0.85-0.95 g lactic acid/ 100 g cheese). Notably, no significant differences in pH and acidity levels were observed in KIP sample compared to KC. After 7 days of storage, a significant increase (P < 0.05) in pH values was observed, whereas acidity was decreased in all samples. The addition of free or immobilized cells had no effect on water activity levels, ranging 0.88-0.92. The moisture content ranged at usual levels (72.7-75.5%) in KC and KFP samples. However, in KIP sample, lower values were observed initially (63.9-65.71%), but increased to similar with KC and KFP samples levels after 2 days of storage. After 7 days of storage a slight increase in moisture content was observed.
3.5. Microbiological Analysis of Functional Katiki Domokou type cheese
The results of the microbiological analysis are presented in Table 4. Lactobacilli were detected at all samples at levels ≥ 6.95 logcfu/g. As expected, cell loads of lactobacilli were significantly (P < 0.05) higher in cheese KFP and KIP samples than in KC product at every timepoint. Up to 3 days of storage, a gradual increase of cell loads was observed at samples with free or immobilized cells, resulting in 8.99 and 9.12 logcfu/g in KFP and KIP sample, respectively. However, storage after the expiration date (4 days after package opening) affected lactobacilli levels, as a significant decline (P < 0.05) in cell viability was observed after 7 days of storage, resulting in 8.57 and 8.96 logcfu/g in KFP and KIP, respectively. Of note, the survival rate of lactobacilli was lower (P < 0.05) in KFP compared to KIP sample. No significant differences were observed in lactococci cell loads (4.80 – 4.82 logcfu/g) among all samples. In accordance with lactobacilli, lactococci were increased gradually up to 3 days of storage, and their numbers were significantly reduced after the expiration date. Yeasts were detected in similar loads (6.35- 6.39 logcfu/g) in all samples at the beginning of storage, but during storage their numbers were significantly (P < 0.05) lower in KFP and KIP samples compared to KC. Importantly, staphylococci were detected at numbers ≤ 3.92 logcfu/g in all cheese samples. Storage affected staphylococci loads, as an increase was observed after the 3rd day of storage, which was significantly (P < 0.05) higher in KC sample compared to KFP and KIP. Significantly coliforms, enterobacteria, Salmonella sp. and L. monocytogenes were below detectable level (≤1 logcfu/g) in all samples.
3.6. Resistance of functional Katiki Domokou type cheese to microbial contamination
Possible resistance to microbial contamination of Katiki Domokou cheese samples fortified with free or immobilized P. acidilactici ORE5 culture after deliberate spiking with L. monocytogenes was investigated and the results are shown in Table 5. Initial cell loads of L. monocytogenes were 5.08-5.11 logcfu/g in all samples and no significant differences were observed among samples up to 2 days of storage. After 3 days and up to 7 days of storage, a gradual increase of L. monocytogenes levels was observed, but higher levels (P < 0.05) were noted in KC sample than KFP and KIP samples. During storage, a significant decline (P < 0.05) in both lactobacilli and lactococci counts was observed in all samples compared to the ones no deliberate spiking with L. monocytogenes. More specifically, up to 3 days of storage lactobacilli cell levels were 8.99 and 9.12 logcfu/g in uninoculated KFP and KIP samples, respectively, while their numbers decreased to 8.45 and 8.54 logcfu/g in KFP and KIP samples deliberately spiking with L. monocytogenes.
3.7. Minor volatiles in functional Katiki Domokou type cheese
Katiki Domokou type cheese samples fortified with free or immobilized P. acidilactici ORE5 cells on pistachio nuts were evaluated regarding their volatile compounds at the beginning and after 3 days of refrigerated storage by HS-SPME GC/MS technique and the results are presented in Table 6. For comparison reasons, Katiki Domokou cheese samples with no P. acidilactici ORE5 cells were also analyzed. Overall, 41 compounds were detected: 14 compounds in KC sample, 27 compounds in KFP sample and 37 compounds in KIP sample. The main classes of volatile compounds identified were esters, acids, alcohols and carbonyl compounds. From a quantitative perspective, the highest (P < 0.05) content of total volatiles was observed in KFP sample (52.3 mg/L), while total volatiles were ranged 31.6 mg/L in KIP sample and 4.8 mg/L in KC sample. More specifically, the incorporation of free P. acidilactici ORE5 cells in cheese resulted in significantly (P < 0.05) higher amounts of total esters, acids and hydrocarbons, while cheese sample fortified with immobilized P. acidilactici ORE5 cells contained higher concentrations of total alcohols (P < 0.05). Regarding to total carbonyl compounds, no significant differences were observed between KFP and KIP samples.
The detected volatile compounds in cheese samples included 7 esters. Among them, ethyl acetate, ethyl hexanoate and 2- phenylethyl acetate were identified in all cheese samples. However, incorporation of free or immobilized P. acidilactici ORE5 cells led to significantly (P < 0.05) increased amounts of these compounds compared to the control sample. Of note, ethyl propanoate, ethyl butyrate, 3- methylbutyl acetate and ethyl decanoate were detected only in KFP and KIP samples.
Among the volatile compounds, 5 acids were detected in cheese samples. Sorbic, octanoic, benzoic and decanoic acid were present in all samples, while hexanoic acid was found in KFP and KIP sample after 3 days of storage. In general, the amounts of acids were higher (P < 0.05) in KFP and KIP samples than in the control cheese (KC).
In total, 6 alcohols were detected in samples. Amounts of 3- methyl- 1- butanol and 2-phenylethanol were identified in all samples, however, higher concentrations were observed in KFP and KIP samples (P < 0.05) compared to control cheese. Of note, 2-methyl-1-propanol and 2-methyl-1-butanol were present only in KFP and KIP, while 1-hexanol and 2-methyl-phenol were detected only in KIP sample. In all cases, the incorporation of immobilized cells in cheese led to the highest concentrations of alcohols (P < 0.05) among the samples.
The carbonyl compounds present in cheese samples included 6 aldehydes and 4 ketones that were more abundant in KFP and KIP samples than in KC. Among them, only hexanal was common in all samples. In KC sample, 2-pentanone and 2-heptanone were the only carbonyl compounds detected, apart from hexanal. 2- nonanone were observed only in KFP sample, while heptanal was present at the beginning of storage in KFP cheese., However, after 3 days of storage it was undetectable. On the other hand, 2-methyl-propanal, 2-methyl-butanal and 3-hydroxy-butanone were present only in KIP sample. Furthermore, 3-methyl-butanal and benzeneacetaldehyde were detected in both KFP and KIP samples. In total, no significant differences were observed between KFP and KIP samples. Remarkably, carbonyl compounds were identified in lower concentrations (P < 0.05) compared to other volatile compounds detected.
Apart from the main classes of volatile compounds, 12 hydrocarbons were identified in our samples. 2-methyl-1,3-pentadiene, octane, styrene, decane, dodecane, 1,3-bis(1,1-dimethylethyl)-benzene and tetradecane were present in both KFP and KIP sample, while 2,4-dimethyl-1-heptane, 2,6-dimethyl-pyrazine and 3-ethyl-2,5-dimethyl-pyrazine were detected in KIP sample. In general, cheese sample fortified with free cells exhibited the highest content of hydrocarbons.
Principal Component Analysis (PCA) results are represented in Figure 1. No significant differences were noted at the beginning of the storage and thus the samples are gathered at the left part of the plot. However, the pistachio addition (KFP samples) correlated positively to PC2. On the other hand, storage (3 days) had a significant effect on both KFP and KIP samples and correlated positively to PC1, as KFP is appeared in the lower right part of diagram and KIP in the upper right part.
3.8. Effect of Immobilized Pediococcus acidilactici ORE5 culture on bacteria microbiome of Katiki Domokou type cheese
Molecular analysis was performed at all cheese samples after 3 days of storage, in order to evaluate the bacteria composition. The bacteria diversity was assessed using Shannon and Simpson indices, as shown in Table 7. The replicates of KIP sample exhibited greater diversity compared to KC and KIP sample (P < 0.05). Furthermore, KC sample replicates were more diverse than KFP sample (P < 0.05).
The results regarding bacterial phyla and genera are presented in Figure 2 and Figure 3, respectively. At phyla level, the most abundant phylum (>93%) was Firmicutes in all samples (P < 0.05), followed by Proteobacteria (0.41-3.79%) and Bacteroidetes (0.22-2.88%). Actinobacteria were present in lower levels (0.13-0.20%), while Saccharibacteria, formerly known as TM7, were found only in KC sample (0.02%). Firmicutes were more abundant in KFP and KIP samples compared to KC (P < 0.05) and conversely Proteobacteria and Bacteroidetes were in lower levels in KFP and KIP than in KC sample (P < 0.05). Regarding Actinobacteria, no significant differences were observed between KC and KIP samples (P > 0.05). However, Actinobacteria were present in lower percentages in KFP sample (P < 0.05).
At genera level, Lactococcus spp. was the most abundant genus (> 78.7%) in all samples (P < 0.05). Notably, lower percentages were observed in KIP sample (78.71%) (P < 0.05) compared to KC and KFP (87.49 and 92.77%, respectively). Pediococcus genus was present, as expected, in cheese samples fortified with free or immobilized P. acidilactici ORE5 with a relative abundance 0.31 and 10.18% in KFP and KIP, respectively, while in KC it was detected in low levels (0.01%). Furthermore, Streptococcus genus was identified in relative abundance ranging 0.74-3.11%, where in KFP and KIP the levels were higher compared to KC sample (P < 0.05). Lactobacillus spp. OTUS were higher in KIP (0.27%) compared to KC and KFP (P < 0.05), while no significant differences were observed between KC and KFP samples (0.10% relative abundance in both samples). The genera Chryseobacterium, Acinetobacter and Pseudomonas were abundant in KC sample (2.44, 1.57 and 1.29%, respectively), while in KFP and KIP were present in significant lower percentages (P < 0.05). Similarly, Staphylococcus, Enhydrobacter and Escherichia were detected in higher levels in KC (P < 0.05) than in KFP and KIP samples. Other genera (Allobaculum, Alloiococcus, Clostridium, Corynebacterium, Enterobacter, Enterococcus, Facklamia, Flavobacterium, Gluconacetobacter, Helicobacter, Janthinobacterium, Jeotgalicoccus, Macrococcus, Mannheimia, Methylobacterium, Oligella, Pedobacter, Prevotella, Propionibacterium, Psychrobacter, , Salmonella, Sphingobacterium and Stenotrophomonas) were identified in low percentages (≤ 0.08%). Of note, the genera Facklamia, Flavobacterium, Salmonella and Sphingobacterium were present in KC sample, while their presence was not detected in KFP and KIP samples. Moreover, Propionibacterium OTUS were higher in KFP and KIP (0.06 and 0.08%, respectively) (P > 0.05) than in KC sample (0.03%).
3.9. Preliminary Sensory Evaluation
The cheese samples fortified with free or immobilized P. acidilactici ORE5 cells (KFP and KIP samples, respectively) were compared with cheese sample without P. acidilactici ORE5 cells (KC sample) and cheese sample with pistachio nuts (KP sample) regarding their sensory characteristics (Table 3). In general, an improvement of overall quality and taste was evident by the addition of pistachio nuts in Katiki Domokou cheese with or without P. acidilactici ORE5 cells. All samples were characterized by a milk-like and yeasty-bread-like aroma, while their texture was mainly curdy and creamy. Furthermore, KC and KFP samples were evaluated as acid and salted, whereas pistachio nuts in KP and KIP samples added also a sweet and sour taste.
4. Discussion
Kefir grains are rich in potentially probiotics microorganisms, and it is established that exhibit noteworthy health-promoting properties [24]. P. acidilactici ORE5, isolated from kefir grains, was initially screened for potential probiotic properties and adequate survival rates at low pH were observed. This is an important finding as a candidate probiotic strain should persist at pH 3.0 or even lower [25]. Furthermore, it exhibited resistance to pancreatin and bile salts. Pancreatin and bile salts are considered as important parameters regarding food digestion. Specifically, pepsin is an enzyme that catalyzes protein digestion, while pancreatin is contributes significantly role in lipid metabolism, On the other hand, bile salts assist to the digestion and absorption of fats. Likewise, all the above are significant requirements for probiotic efficiency [26]. Regarding antibiotic susceptibility, resistance to tetracycline and clindamycin was monitored which is also in agreement with the study of Kastner et al. [27], while resistance to erythromycin has also been reported by Rojo-Bezares et al. [28]. However, these reports are few in the literature and they lack information about the resistance character (intrinsic or acquired).
Meanwhile, the demand of functional foods containing probiotic cells is growing as the consumers awareness about the positive impact of probiotics on health is increasing. Notably, incorporation of probiotics in dairy products is already common, as it has been suggested that milk functions as buffering agent protecting bacteria during the passage through the gastrointestinal tract [29]. In addition, cell immobilization on food carriers may improve this protective effect resulting in high cell viability. Pistachio nuts have been previously evaluated as natural carriers and the positive impact of consumption of pistachio nuts fortified with probiotic cells on lipid profile of diabetic rats was recently reported [3, 8]. In this vein, our strategy involved the incorporation of immobilized P. acidilactici ORE5 cells on pistachio nuts in Katiki Domokou cheese and the evaluation of potential effect of immobilized probiotic cells on cheese microbiome.
Katiki Domokou is a soft cheese made from goat’s and ewe’s milk and owed to its pH (4.3-4.5) may be considered as a suitable food carrier for probiotic cells in order to maintain high cell viability during storage and after consumption. This suggestion has been corroborated by the high cell loads of P. acidilactici ORE5 cells achieved during refrigerated storage of cheese, as lactobacilli levels were >8.5 logcfu/g up to 7 days of storage. The incorporation of immobilized probiotic cells resulted in higher cell viability compared to the cheese sample fortified with free cells, indicating the positive impact of cell immobilization on maintenance of high cell loads. These results are in accordance with previous studies that demonstrated the enhancement of cell survival by cell immobilization [4, 30, 31]. Indeed, the incorporation of immobilized L. casei ATCC 393 on apple pieces and casein in yogurt resulted in cell loads > 7 logcfu/g up to 28 days of storage [30], while Shehata et al. (2022) reported that soft cheese with immobilized L. paracasei cells on wheat bran exhibited high cell loads (> 7.5 logcfu/g) after 30 days of storage [31]. Of note, a significant increase in lactobacilli numbers was observed up to 3 days of storage, which may be attributed to the ability of lactobacilli to adapt well in milk environment [30]. Furthermore, in accordance with the results, Terpou et al. [32] reported that yeast numbers were significantly lower in cheese samples with free and immobilized L. paracasei cells after ripening period compared to control cheese, indicating that L. paracasei strain provided an antifungal effect on cheese. According to Dimitrellou et al. [33] the incorporation of whey cheese with L. casei or L. bulgaricus culture resulted in significantly lower cell loads of staphylococci, coliforms and enterobacteria. It has been proposed that lower pH values and high acidity combined with lower moisture content may have developed an unfavorable environment for spoilage bacteria growth.
The growth of spoilage bacteria in foods consists of a global issue affecting the food industry with subsequent impact on economy and health system. Dairy products are usually susceptible of spoilage due to their high moisture content, nutrient-rich composition, sensitivity to temperature fluctuations, as well as the possibility of contamination during production. A wide range of microorganisms have been indicted for spoilage in dairy products, such as L. monocytogenes, S. aureus, Salmonella spp and Penicillium spp. [34, 35]. Meanwhile, biopreservation by lactic acid bacteria is gaining a lot of scientific interest, as they produce metabolites such as organic acids with antimicrobial activity potential [15, 35]. With the intention of eliminating food spoilage hazard, the resistance of Katiki Domokou cheese samples fortified with P. acidilactici ORE5 culture to deliberately contamination of L. monocytogenes was investigated. The growth rate of L. monocytogenes was significantly limited in cheese samples with free and immobilized P. acidilactici ORE5 culture, indicating its activity against food-borne pathogens. The study of Gonzales-Barron et al. [36] highlighted the anti-listeria effect of intentionally added lactic acid bacteria in artisanal semi-hard cheese, while Afzali et al. [37] reported that L. brevis strains incorporated in yogurt drink exhibited an anti-fungal activity. In addition, Belessi et al. [38] noted that there is a correlation between yeast and Listeria levels, as increased yeast cell loads may favor the viability of Listeria in Feta cheese, by affecting physicochemical characteristics of cheese leading to more favorable pH values for spoilage growth.
Physicochemical characteristics are crucial for the determination of quality of the cheese, as these parameters affect the sensory evaluation, texture and overall acceptability of the cheese. In this vein, pH values, acidity, water activity and moisture content were evaluated. It has been noted that the samples fortified with P. acidilactici ORE5 culture exhibited lower pH values and, subsequently, higher lactic acid concentrations than the control cheese as the storage continued. According to the literature, studies about functional dairy products have reported similar results attributing them to the production of metabolites of probiotic strains [39]. More specifically, post-acidification was observed in yogurts fortified with L. casei ATCC 393 cells after 7 days of storage, according to Dimitrellou et al. [30]. In agreement with moisture content levels detected during storage, Mileriene et al. [40] reported lower moisture values in curd cheese fortified with immobilized L. lactis cells on raisins, indicating that there is a correlation between pH values and moisture content. More specifically, it has been reported that low pH values (< 5.0) in cheese may affect protein to protein interactions resulting in decreased ability of proteins interaction with water and, subsequently, increased syneresis and lower moisture levels [41].
The sensory evaluation of dairy products significantly determines their quality. Subsequently, the development of unique profile of aromatic compounds in cheese consists of a challenge for the food industry. According to previous studies, esters, acids, alcohols and carbonyl compounds are considered the most important volatile compounds contributing to flavor development in cheese samples. Nevertheless, cheese flavor is rather a result of balance between the concentrations of volatile compounds detected in cheese than a contribution of particular components [29]. For the HS-SPME GC/MS analysis, samples at the beginning and after 3 days of storage were tested. The incorporation of free or immobilized cells affected the aroma profile of cheese, in accordance with previous studies on dairy products fortified with lactic acid bacteria [30, 31]. Indeed, regarding esters, Dimitrellou et al. [29] noted that cheese samples fortified with free L. casei cells exhibited higher amounts of ethyl hexanoate and ethyl decanoate compared to control cheese, as well as to cheese sample with immobilized cells on whey protein. Similarly, Shehata et al. [31] reported the detection of ethyl decanoate in soft whey cheese with free and immobilized L. paracasei cells, however, in control cheese it was undetectable. In total, the esters identified in cheese samples are known to produce fruity aroma in foods [32, 42]. The acids found at increased concentrations in KFP and KIP samples were sorbic, octanoic and decanoic acid, that were at higher amounts in KFP samples, results that are in accordance with previous studies in dairy products fortified with probiotics [31, 32]. Among alcohols, the most abundant were 3-methyl-1-butanol and 2-phenylethanol in all samples and are described as exerting malt notes and burnt aroma [4]. Carbonyl compounds detected in cheese samples belong to aldehydes and ketones, but aldehydes were in low concentrations due to their reduction to alcohols [33]. 3-methyl-butanal and hexanal detected only in KFP and KIP samples were similarly identified in cheese samples fortified with free or immobilized lactic acid bacteria, in accordance to previous studies [29]. Of note, 3- methyl- butanal is present also in other cheeses like Camembert and aged Cheddar adding fruity aroma to them [43]. Furthermore, hydrocarbons were also present in cheese, mainly in KFP and KIP samples. In general, hydrocarbons do not directly affect aroma profile of cheese. However, they act as precursors to other volatile compounds [44]. Notably, PCA, which was carried out in order to evaluate the effect of P. acidilactici ORE5 on volatile profile of Katiki Domokou cheese, revealed that the incorporation of free and immobilized cells affected the concentrations of volatile compounds, which can be attributed to their fermentation activity, while similar results have been reported in previous studies [29, 44].
Dairy products, specifically cheeses, are characterized by certain microbiome, which differs between different types of cheese, depending on a variety of factors, such as milk source, the starter and adjunct cultures, the cheese-making process, the geographical region and terroir. Although plate count method provides a quantitative assessment of the microbial composition of the product, it has some limitations due to its sensitivity, as well as its inability sometimes to distinguish between colonies. Hence, it is often recommended to combine the plate count method with other techniques such as molecular methods (e.g., DNA-based techniques) to obtain a more comprehensive understanding of the microbial community [45]. In this study, the effect of P. acidilactici ORE5 cells incorporated in Katiki Domokou cheese on the cheese microbiome was investigated after 16S rRNA sequencing. Concerning the results of Shannon and Simpson indices, the greater bacterial diversity as a result of the incorporation of immobilized cells in cheese sample have also been noted by Mitropoulou et al. [46]. Regarding phyla, Firmicutes was the predominant phylum in all cheese samples, while Bacteroidetes and Proteobacteria were in higher levels in control cheese compared to the samples with P. acidilactici ORE5 culture, in accordance with previous studies [45, 46]. At genera level, Pediococcus were detected, as expected, in higher percentages in samples with free or immobilized P. acidilactici ORE5 cells compared to control. However, lactococci were the most abundant genus in all samples, which may be attributed to the presence of Lactococcus in cheese, as mesophilic lactic acid bacteria, such as Lactococcus, predominate during ripening and storage of cheese [11]. However, their relative abundance values were significantly lower in cheese fortified with immobilized cells. Similarly, Dimitrellou et al. [30] reported a significant decline in the levels of yogurt starter cultures in samples with L. casei culture, due to competitive activity of the probiotic strain. The fact that lactococci were the most abundant genus in all samples contrasts the microbiological analysis results that demonstrated higher counts of lactobacilli compared to lactococci. This may be contributed to the ability of some Lactococcus species to grow on MRS agar, and therefore, not be easily distinguished from other cultures present on petri dishes [47]. Consequently, it is possible some cultures belonging to Lactococcus genus were incorrectly enumerated as lactobacilli. Among other genera found in cheese, Chryseobacterium were detected in control cheese, while in samples with free and immobilized cells was found at levels < 0.05%. Chryseobacterium are considered psychotropic spoilage bacteria and are usually detected in dairy products [48]. Similarly, Pseudomonas, that are considered aerobic psychotropic bacteria, were detectable in higher levels in control cheese in contrast with KFP and KIP samples, where Pseudomonas levels were below 0.02%. It has been reported that Pseudomonas produce enzymes that may lead to spoilage due to their proteolytic activity [49]. Considering that Katiki Domokou cheese is made from goat’s milk, the existence of Acinetobacter in control cheese may be explained, as Acinetobacter is usually found in contaminated raw goat’s milk [50]. Of note, in cheese samples fortified with free and immobilized cells Acinetobacter were found in significant lower levels, which may be attributed to potential antagonistic activity of P. acidilactici ORE5 culture. Higher levels of Staphylococcus were noted in control cheese. Although not all Staphylococcus are considered pathogens, S. aureus is responsible for food poisoning due to production of enterotoxin [51, 52]. Moreover, Enhydrobacter were detected in 0.17% relative abundance in control cheese, while their levels were 0.02% in KFP and KIP samples. The existence of Enhydrobacter is not commonly linked to cheese spoilage, however, some strains may have the potential to cause spoilage under certain conditions [30]. Relative abundance of Sphingobacterium, Enterococcus, Facklamia and Salmonella was recorded only in control cheese and their presence may indicate cheese spoilage. Likewise, Flavobacterium was found only in the control cheese (KC), and it is considered part of natural cheese microflora, while some strains are associated with food spoilage [53]. Overall, NGS technology provides useful information about the microbiome composition of complex ecosystems like cheese, overcoming the limitations of the culture-dependent studies [46].
5. Conclusions
In this study, the effect of free and immobilized P. acidilactici ORE5 cells on pistachio nuts on Katiki Domokou cheese was evaluated. The results demonstrated that Katiki Domokou supplementation with immobilized P. acidilactici ORE5 positively affected the cheese microbiome, while high P. acidilactici cell loads were maintained, alongside with growth suppression of L. monocytogenes after deliberate inoculation. Notably, the use of both free and immobilized P. acidilactici ORE5 cells improved the aroma profile of cheese, while the incorporation of immobilized P. acidilactici ORE5 cells on pistachio nuts enhanced the sensory characteristics of the product. Thus, the development of functional Katiki Domokou type cheese fortified with freeze-dried immobilized P. acidilactici ORE5 cells on pistachio nuts holds market potential. However, more research is required in order to confirm the health- promoting effect of P. acidilactici ORE5 cells in experimental animal models and on humans, as well as to investigate the effectiveness of freeze-dried immobilized P. acidilactici ORE5 cells on pistachio nuts incorporated in Katiki Domokou cheese at industrial scale.
Author Contributions
Conceptualization, Yiannis Kourkoutas; Data curation, Chrysoula Pavlatou, Anastasios Nikolaou, Ioanna Prapa, Konstantinos Tegopoulos and Eugenia Bezirtzoglou; Formal analysis, Chrysoula Pavlatou and Maria E. Grigoriou; Funding acquisition, Yiannis Kourkoutas; Investigation, Chrysoula Pavlatou, Anastasios Nikolaou, Ioanna Prapa, Konstantinos Tegopoulos and Stavros Plessas; Methodology, Anastasios Nikolaou, Ioanna Prapa, Stavros Plessas and Maria E. Grigoriou; Project administration, Yiannis Kourkoutas; Resources, Stavros Plessas, Maria E. Grigoriou, Eugenia Bezirtzoglou and Yiannis Kourkoutas; Software, Eugenia Bezirtzoglou; Supervision, Yiannis Kourkoutas; Validation, Anastasios Nikolaou, Ioanna Prapa, Konstantinos Tegopoulos, Maria E. Grigoriou and Eugenia Bezirtzoglou; Writing – original draft, Chrysoula Pavlatou, Anastasios Nikolaou and Stavros Plessas; Writing – review & editing, Konstantinos Tegopoulos and Yiannis Kourkoutas.
Funding
We acknowledge support of this work by the project “Research Infrastructure on Food Bioprocessing Development and Innovation Exploitation – Food Innovation RI” (MIS 5027222), which is implemented under the Action “Reinforcement of the Research and Innovation Infrastructure”, funded by the Operational Programme “Competitiveness, Entrepreneurship and Innovation” (NSRF 2014–2020) and co-financed by Greece and the European Union (European Regional Development Fund), as well as by the project “Infrastructure of Microbiome Applications in Food Systems-FOODBIOMES” (MIS 5047291), which is implemented under the Action “Regional Excellence in R&D Infrastructures”, funded by the Operational Programme "Competitiveness, Entrepreneurship and Innovation" (NSRF 2014-2020) and co-financed by Greece and the European Union (European Regional Development Fund).
Data Availability Statement
Data is available upon request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Food and Agricultural Organization of the United Nations; World Health Organization. Probiotics in Food: Health and Nutritional Properties and Guidelines for Evaluation. FAO Food Nutr. Paper 2006, 85. Available online: https://www.fao.org/3/a0512e/a0512e.pdf (accessed on 29 January 2023).
- Kvakova, M.; Bertkova, I.; Stofilova, J.; Savidge, T.C. Co-Encapsulated Synbiotics and Immobilized Probiotics in Human Health and Gut Microbiota Modulation. Foods 2021, 10, 1297. [Google Scholar] [CrossRef] [PubMed]
- Prapa, I.; Nikolaou, A.; Panas, P.; Tassou, C.; Kourkoutas, Y. Developing Stable Freeze-Dried Functional Ingredients Containing Wild-Type Presumptive Probiotic Strains for Food Systems. Appl. Sci. 2023, 13, 630. [Google Scholar] [CrossRef]
- Nikolaou, A.; Mitropoulou, G.; Nelios, G.; Kourkoutas, Y. Novel Functional Grape Juices Fortified with Free or Immobilized Lacticaseibacillus rhamnosus OLXAL-1. Microorganisms 2023, 11, 646. [Google Scholar] [CrossRef] [PubMed]
- Oyinloye, T.M.; Yoon, W.B. Effect of Freeze-Drying on Quality and Grinding Process of Food Produce: A Review. Processes 2020, 8, 354. [Google Scholar] [CrossRef]
- Deepika, G.; Charalampopoulos, D. Surface and Adhesion Properties of Lactobacilli. Adv. Appl. Microbiol. 2010, 70, 127–152. [Google Scholar] [CrossRef]
- Yanni AE, Mitropoulou G, Prapa I, Agrogiannis G, Kostomitsopoulos N, Bezirtzoglou E, Kourkoutas Y, Karathanos VT. Functional modulation of gut microbiota in diabetic rats following dietary intervention with pistachio nuts (Pistacia vera L.). Metabol Open 2020, 7, 100040. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Prapa, I.; Yanni, A.E.; Nikolaou, A.; Kostomitsopoulos, N.; Kalogeropoulos, N.; Bezirtzoglou, E.; Karathanos, V.T.; Kourkoutas, Y. Dietary Pistachio (Pistacia vera L.) Beneficially Alters Fatty Acid Profiles in Streptozotocin-Induced Diabetic Rat. Appl. Sci. 2022, 12, 4606. [Google Scholar] [CrossRef]
- Karimi R., Mortazavian A. M., da Cruz A. G. Viability of probiotic microorganisms in cheese during production and storage: a review. Dairy Science and Technology 2011, 91, 283–308. [Google Scholar] [CrossRef]
- Kagkli DM, Iliopoulos V, Stergiou V, Lazaridou A, Nychas GJ. Differential Listeria monocytogenes strain survival and growth in Katiki, a traditional Greek soft cheese, at different storage temperatures. Appl Environ Microbiol. 2009, 75, 3621–6. [Google Scholar] [CrossRef] [PubMed]
- Pappa, E.C.; Kondyli, E. Descriptive Characteristics and Cheesemaking Technology of Greek Cheeses Not Listed in the EU Geographical Indications Registers. Dairy 2023, 4, 43–67. [Google Scholar] [CrossRef]
- Blaya J, Barzideh Z, LaPointe G. Symposium review: Interaction of starter cultures and nonstarter lactic acid bacteria in the cheese environment. J Dairy Sci. 2018, 101, 3611–3629. [Google Scholar] [CrossRef] [PubMed]
- Dimitrellou D, Kourkoutas Y, Koutinas AA, Kanellaki M. Thermally- dried immobilized kefir on casein as starter culture in dried whey cheese production. Food Microbiol. 2009, 26, 809–20. [Google Scholar] [CrossRef] [PubMed]
- Araújo, V.S.; Pagliares, V.A.; Queiroz, M.L.P. and Freitas-Almeida, A.C. Occurrence Staphylococcus and enteropathogens in soft cheese commercialized in the city of Rio de Janeiro, Brazil. Journal of Applied Microbiology 2002, 92, 1172–1177. [Google Scholar] [CrossRef]
- Arena, M. P., Silvain, A., Normanno, G., Grieco, F., Drider, D., Spano, G. and Fiocco, D. Use of Lactobacillus plantarum strains as a bio-control strategy against food-borne pathogenic microorganisms. Frontiers in Microbiology 2016, 7, 464. [Google Scholar] [CrossRef]
- Sherwani, K. F., Bukhari, D., A., A. Probiotics in processed dairy products and their role in gut microbiota health. 2022, 2022. [Google Scholar] [CrossRef]
- Li, C.; Wang, S.; Chen, S.; Wang, X.; Deng, X.; Liu, G.; Chang, W.; Beckers, Y.; Cai, H. Screening and Characterization of Pediococcus acidilactici LC-9-1 toward Selection as a Potential Probiotic for Poultry with Antibacterial and Antioxidative Properties. Antioxidants 2023, 12, 215. [Google Scholar] [CrossRef]
- ISO 5534:2004; Cheese and Processed Cheese. Determination of the Total Solids Content (Reference Method). International Organization for Standardization: Geneva, Switzerland, 2004.
- Dan T, Wang D, Jin RL, Zhang HP, Zhou TT, Sun TS. Characterization of volatile compounds in fermented milk using solid-phase microextraction methods coupled with gas chromatography-mass spectrometry. J Dairy Sci. 2017, 100, 2488–2500. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; Sahl, J.W.; Stres, B.; Thallinger, G.G.; Van Horn, D.J.; Weber, C.F. Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl Environ Microbiol. 2009, 75, 7537–41. [Google Scholar] [CrossRef]
- Nikolaou, A.; Kourkoutas, Y. High-Temperature Semi-Dry and Sweet Low Alcohol Wine-Making Using Immobilized Kefir Culture. Fermentation 2021, 7, 45. [Google Scholar] [CrossRef]
- EFSA Panel on Additives and Products or Substances Used in Animal Feed (FEEDAP). Guidance on the Assessment of Bacterial Susceptibility to Antimicrobials of Human and Veterinary Importance. EFSA J. 2012, 10, 2740. [Google Scholar] [CrossRef]
- Litopoulou-Tzanetaki E, Tzanetakis N. The Microfloras of Traditional Greek Cheeses. Microbiol Spectr. 2014, 2, CM-0009-2012. [Google Scholar] [CrossRef] [PubMed]
- Bengoa, A.A.; Iraporda, C.; Garrote, G.L.; Abraham, A.G. Kefir micro-organisms: their role in grain assembly and health properties of fermented milk. J. Appl. Microbiol. 2019, 126, 686–700. [Google Scholar] [CrossRef]
- Argyri, A.A.; Zoumpopoulou, G.; Karatzas, K.A.; Tsakalidou, E.; Nychas, G.J.; Panagou, E.Z.; Tassou, C.C. Selection of potential probiotic lactic acid bacteria from fermented olives by in vitro tests. Food Microbiol. 2013, 33, 282–291. [Google Scholar] [CrossRef]
- Danielsen, M.; Wind, A. Susceptibility of Lactobacillus spp. to antimicrobial agents. Int. J. Food Microbiol. 2003, 82, 1–11. [Google Scholar] [CrossRef]
- Kastner, S.; Perreten, V.; Bleuler, H.; Hugenschmidt, G.; Lacroix, C.; Meile, L. Antibiotic susceptibility patterns and resistance genes of starter cultures and probiotic bacteria used in food. Syst. Appl. Microbiol. 2006, 29, 145–155. [Google Scholar] [CrossRef]
- Rojo-Bezares, Beatriz, et al. Assessment of antibiotic susceptibility within lactic acid bacteria strains isolated from wine. International journal of food microbiology 2006, 111, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Dimitrellou, Dimitra, et al. Free and immobilized Lactobacillus casei ATCC 393 on whey protein as starter cultures for probiotic Feta-type cheese production. Journal of Dairy Science 2014, 97, 4675–4685. [Google Scholar] [CrossRef] [PubMed]
- Dimitrellou, Dimitra, Panagiotis Kandylis, and Yiannis Kourkoutas. Assessment of freeze-dried immobilized Lactobacillus casei as probiotic adjunct culture in yogurts. Foods 2019, 8, 374. [Google Scholar] [CrossRef]
- Shehata, Mohamed G., et al. Lacticaseibacillus paracasei KC39 immobilized on prebiotic wheat bran to manufacture functional soft white cheese. Fermentation 2022, 8, 496. [Google Scholar] [CrossRef]
- Terpou, Antonia, et al. Growth capacity of a novel potential probiotic Lactobacillus paracasei K5 strain incorporated in industrial white brined cheese as an adjunct culture. Journal of food science 2018, 83, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Dimitrellou, D., Kandylis, P., Kourkoutas, Y., & Kanellaki, M. Novel probiotic whey cheese with immobilized lactobacilli on casein. LWT 2017, 86, 627–634. [Google Scholar] [CrossRef]
- Choi J, Lee SI, Rackerby B, Goddik L, Frojen R, Ha SD, et al. Microbial Communities of a Variety of Cheeses and Comparison between Core and Rnd Region of Cheeses. Journal of Dairy Science 2020, 103, 4026–4042. [Google Scholar] [CrossRef] [PubMed]
- Shi, C., & Maktabdar, M. Lactic acid bacteria as biopreservation against spoilage molds in dairy products–A review. Frontiers in microbiology 2022, 12, 4283. [Google Scholar] [CrossRef]
- Gonzales-Barron U, Campagnollo FB, Schaffner DW, Sant'Ana AS, Cadavez VAP. Behavior of Listeria monocytogenes in the presence or not of intentionally- added lactic acid bacteria during ripening of artisanal Minas semi-hard cheese. Food Microbiol. 2020, 91, 103545. [Google Scholar] [CrossRef] [PubMed]
- Afzali, S., Edalatian Dovom, M. R., Habibi Najafi, M. B., & Mazaheri Tehrani, M. Determination of the anti-yeast activity of Lactobacillus spp. isolated from traditional Iranian cheeses in vitro and in yogurt drink (Doogh). cientific reports 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Belessi, C. I. A., Papanikolaou, S., Drosinos, E. H., & Skandamis, P. N. Survival and acid resistance of Listeria innocua in feta cheese and yogurt, in the presence or absence of fungi. Journal of food protection 2008, 71, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Hong, Q., Wang, G., Zhang, Q., Tian, F., Xiao-Ming, L., Zhao, J., ... & Chen, W. Enhancement of ester formation in Camembert cheese by addition of ethanol. International Journal of Dairy Technology 2017, 70, 220–227. [Google Scholar] [CrossRef]
- Mileriene, J.; Serniene, L.; Kondrotiene, K.; Santarmaki, V.; Kourkoutas, Y.; Vasiliauskaite, A.; Lauciene, L.; Malakauskas, M. Indigenous Lactococcus lactis with Probiotic Properties: Evaluation of Wet, Thermally- and Freeze-Dried Raisins as Supports for Cell Immobilization, Viability and Aromatic Profile in Fresh Curd Cheese. Foods 2022, 11, 1311. [Google Scholar] [CrossRef]
- Pastorino, A.J.; Hansen, C.L.; McMahon, D.J. Effect of pH on the Chemical Composition and Structure-Function Relationships of Cheddar Cheese. J. Dairy Sci. 2003, 86, 2751–2760. [Google Scholar] [CrossRef]
- Hayaloglu, A.A. Comparisons of different single-strain starter cultures for their effects on ripening and grading of Beyaz cheese. International Journal of Food Science & Technology, 2007, 42, 930–938. [Google Scholar] [CrossRef]
- Curioni PM, G.; Bosset, J.O. Key odorants in various cheese types as determined by gas chromatography-olfactometry. International Dairy Journal, 2002, 12, 959–984. [Google Scholar] [CrossRef]
- Bezerra, T.K.A.; de Oliveira Arcanjo, N.M.; de Araújo, A.R.R.; de Queiroz, A.L.M.; de Oliveira, M.E.G.; Gomes, A.M.P.; Madruga, M.S. Volatile profile in goat coalho cheese supplemented with probiotic lactic acid bacteria. LWT-Food Science and Technology, 2017, 76, 209–215. [Google Scholar] [CrossRef]
- Quigley L, O’Sullivan O, Beresford TP, Ros, RP, Fitzgerald GF, Cotter PD. High-throughput sequencing for detection of subpopulations of bacteria not previously associated with artisanal cheeses. [CrossRef]
- Mitropoulou, G., Prapa, I., Nikolaou, A., Tegopoulos, K., Tsirka, T., Chorianopoulos, N., ... & Kourkoutas, Y. Effect of Free or Immobilized Lactiplantibacillus plantarum T571 on Feta-Type Cheese Microbiome. Frontiers in Bioscience-Elite 2022, 14, 31. [Google Scholar] [CrossRef]
- Itoi S, Yuasa K, Washio S, Abe T, Ikuno E, Sugita H. Phenotypic variation in Lactococcus lactis subsp. lactis isolates derived from intestinal tracts of marine and freshwater fish. J Appl Microbiol. 2009, 107, 867–74. [Google Scholar] [CrossRef] [PubMed]
- Papadakis P, Konteles S, Batrinou A, Ouzounis S, Tsironi T, Halvatsiotis P, Tsakali E, Van Impe JFM, Vougiouklaki D, Strati IF, Houhoula D. Characterization of Bacterial Microbiota of P.D.O. Feta Cheese by 16S Metagenomic Analysis. Microorganisms 2021, 9, 2377. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Meng, Z., Zhang, L., Xin, L., Lin, K., Yi, H., & Han, X. Technological characterization of Lactobacillus in semihard artisanal goat cheeses from different Mediterranean areas for potential use as nonstarter lactic acid bacteria. Journal of Dairy Science 2018, 101, 2887–2896. [Google Scholar] [CrossRef]
- Kamilari, E., Anagnostopoulos, D. A., Papademas, P., Kamilaris, A., & Tsaltas, D. Characterizing Halloumi cheese's bacterial communities through metagenomic analysis. Lwt 2020, 126, 109298. [Google Scholar] [CrossRef]
- Kourkoutas, Y., Bosnea, L., Taboukos, S., Baras, C., Lambrou, D., & Kanellaki, M. Probiotic cheese production using Lactobacillus casei cells immobilized on fruit pieces. Journal of dairy science 2006, 89, 1439–1451. [Google Scholar] [CrossRef]
- Crippa, C., Pasquali, F., Lucchi, A., Gambi, L., & De Cesare, A. Investigation on the microbiological hazards in an artisanal soft cheese produced in northern Italy and its production environment in different seasonal periods. Italian Journal of Food Safety 2022, 11. [Google Scholar] [CrossRef]
- Spyrelli, E.D.; Stamatiou, A.; Tassou, C.C.; Nychas, G.J.E.; Doulgeraki, A.I. Microbiological and metagenomic analysis to assess the effect of container material on the microbiota of Feta cheese during ripening. Fermentation. 2020, 6, 12. [Google Scholar] [CrossRef]
Figure 1.
Principal component analysis (PCA) of minor volatiles identified in Katiki Domokou type cheese samples containing free or immobilized P. acidilactici ORE5 cells on pistachio nuts . KC: control samples, KFP: cheese samples containing free P. acidilactici ORE5 cells, KIP: cheese with immobilized P. acidilactici ORE5 cells on pistachio nuts. The storage duration (days) is shown at the end of the sample codes.
Figure 1.
Principal component analysis (PCA) of minor volatiles identified in Katiki Domokou type cheese samples containing free or immobilized P. acidilactici ORE5 cells on pistachio nuts . KC: control samples, KFP: cheese samples containing free P. acidilactici ORE5 cells, KIP: cheese with immobilized P. acidilactici ORE5 cells on pistachio nuts. The storage duration (days) is shown at the end of the sample codes.
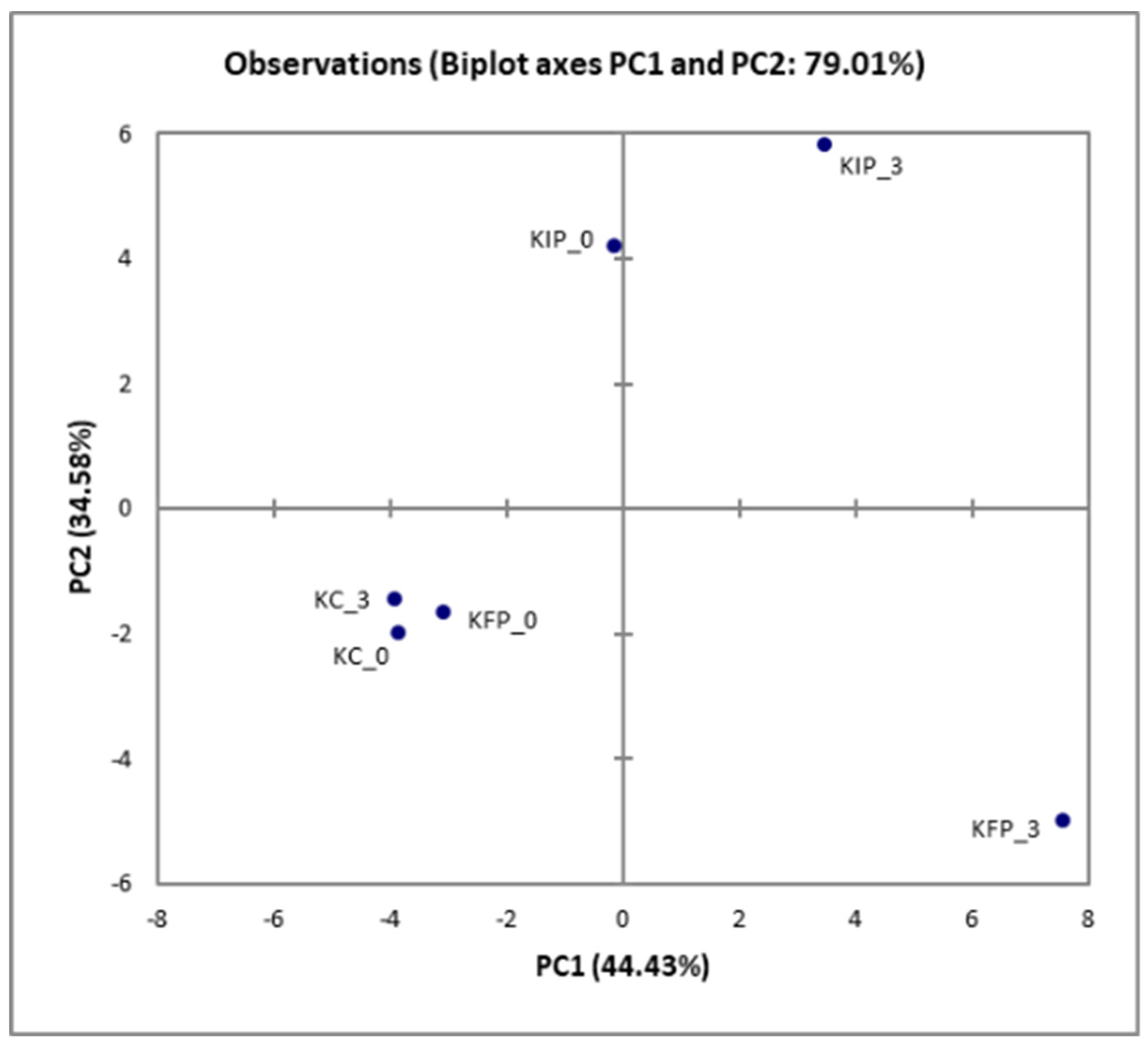
Figure 2.
Relative abundances (%) of different bacteria phyla, after 16S rRNA NGS sequencing in Katiki Domokou type cheese samples. All data represent the mean values from two independent experiments. KC: control samples, KFP: cheese samples containing free P. acidilactici ORE5 cells, KIP: cheese with immobilized P. acidilactici ORE5 cells on pistachio nuts.
Figure 2.
Relative abundances (%) of different bacteria phyla, after 16S rRNA NGS sequencing in Katiki Domokou type cheese samples. All data represent the mean values from two independent experiments. KC: control samples, KFP: cheese samples containing free P. acidilactici ORE5 cells, KIP: cheese with immobilized P. acidilactici ORE5 cells on pistachio nuts.
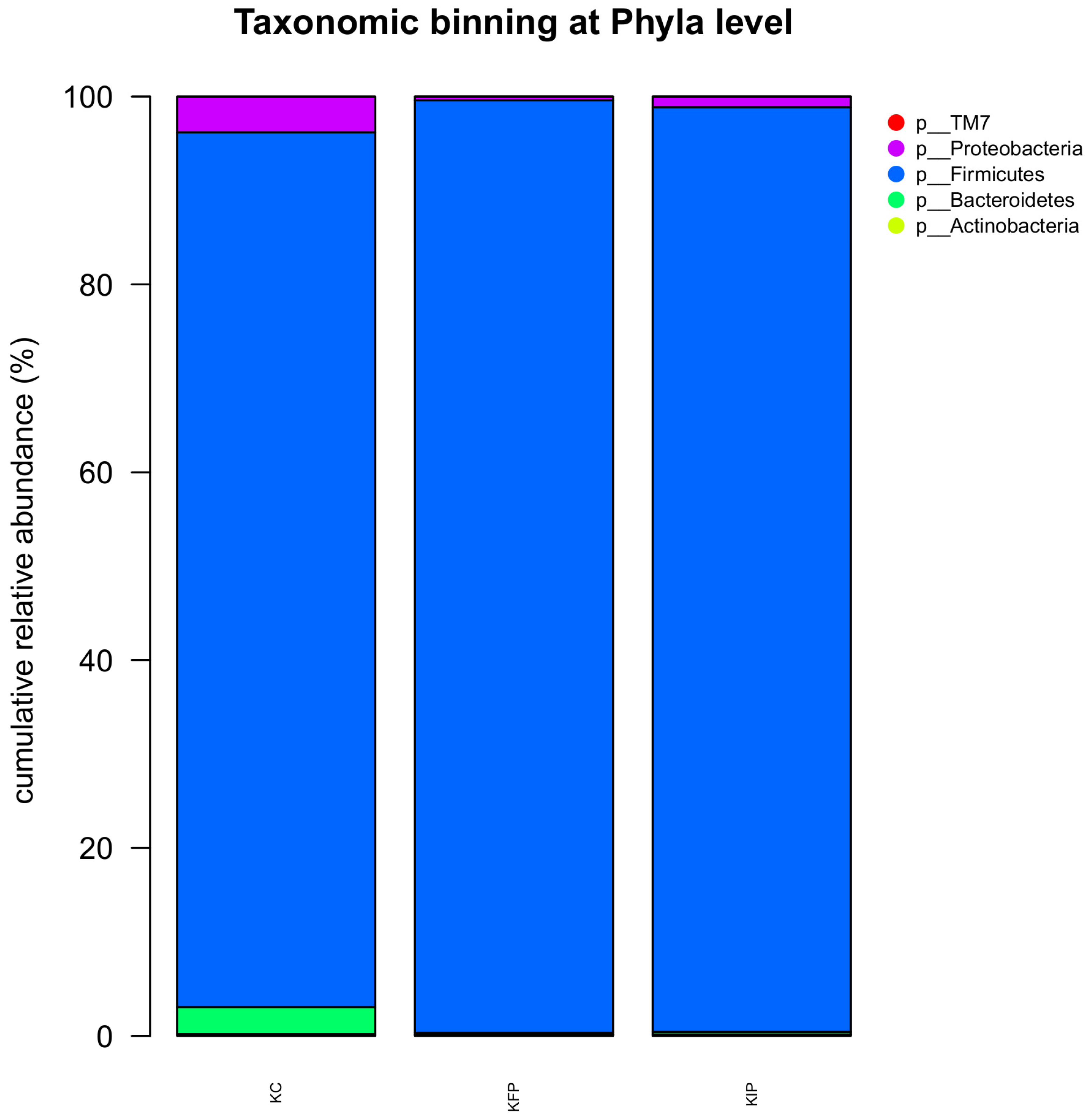
Figure 3.
Relative abundances (%) of different bacterial genera, after 16S rRNA NGS sequencing in Katiki Domokou type cheese samples. All data represent the mean values from two independent experiments. KC: control samples, KFP: cheese samples containing free P. acidilactici ORE5 cells, KIP: cheese with immobilized P. acidilactici ORE5 cells on pistachio nuts.
Figure 3.
Relative abundances (%) of different bacterial genera, after 16S rRNA NGS sequencing in Katiki Domokou type cheese samples. All data represent the mean values from two independent experiments. KC: control samples, KFP: cheese samples containing free P. acidilactici ORE5 cells, KIP: cheese with immobilized P. acidilactici ORE5 cells on pistachio nuts.
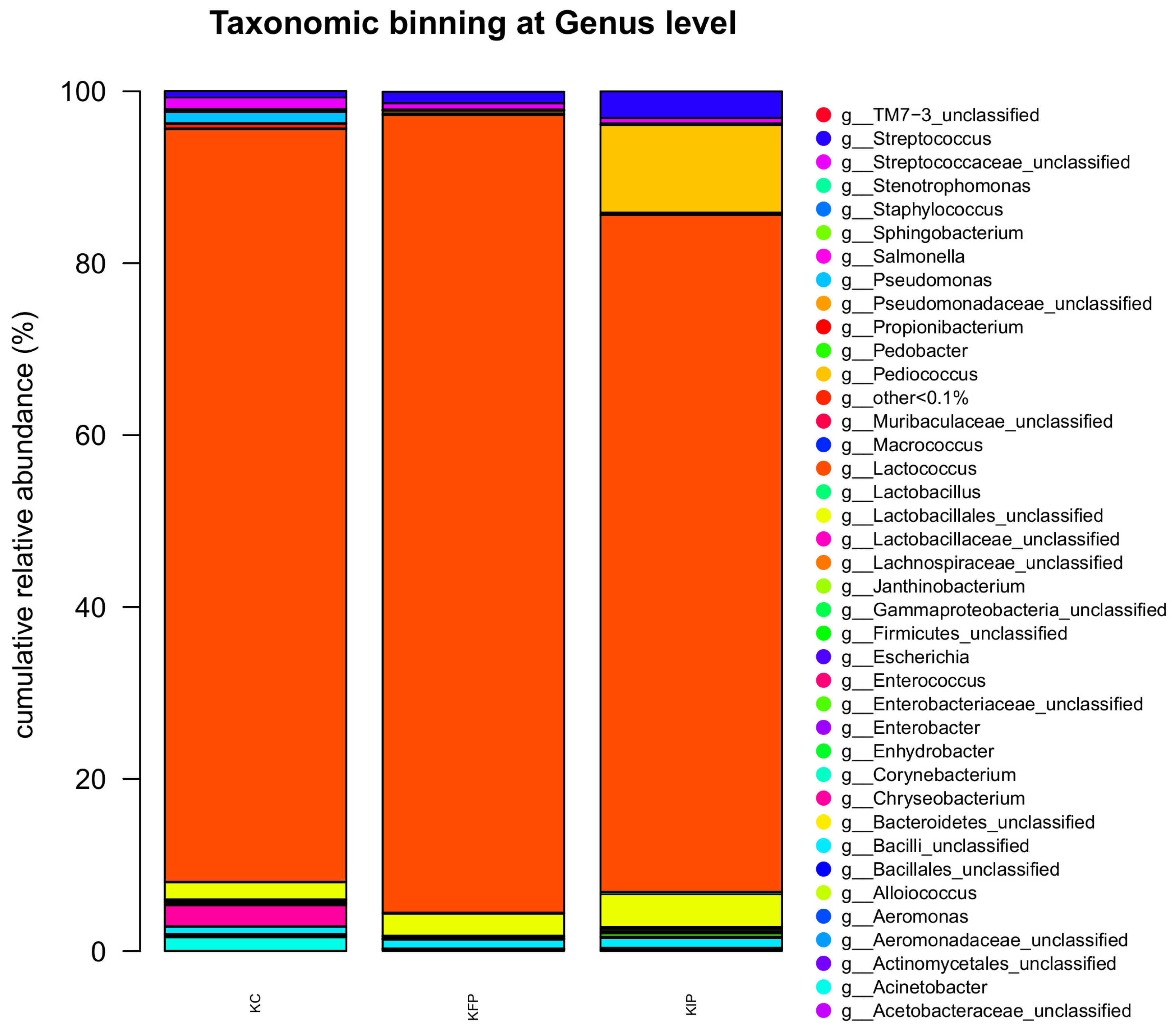

Table 1.
Determination of viability of the isolated strain after exposure to acid environment, bile salts, pepsin and pancreatin. In all tests, the probiotic L. plantarum 14917 served as a reference strain.
Table 1.
Determination of viability of the isolated strain after exposure to acid environment, bile salts, pepsin and pancreatin. In all tests, the probiotic L. plantarum 14917 served as a reference strain.
| Final counts (log cfu/mL) | |||
|---|---|---|---|
| Time (h) |
P. acidilactici ORΕ 5 | L. plantarum ATCC 14971 | |
| Resistance to low pH | 0 | 8.4± 0.21 | 8.3± 0.13 |
| pH=2 | 2 | 6.4± 0.11 | 7.2± 0.07 |
| pH=3 | 2 | 6.9± 0.13 | 7.7± 0.15 |
| pH=4 | 2 | 8.1± 0.07 | 80± 0.12 |
| Pepsin | 0 | 7.6± 0.09 | 7.4± 0.17 |
| 3 | 6.5± 0.19 | 6.8± 0.21 | |
| Pancreatin | 0 | 8.0± 0.37 | 8.1± 0.14 |
| 4 | 7.4± 0.13 | 7.4± 0.21 | |
| Bile salts | 0 | 8.4± 0.22 | 8.5± 0.13 |
| 4 | 8.0± 0.19 | 8.1± 0.08 | |
Table 2.
MIC (μg/mL) of antibiotics for P. acidilactici ORE5 as determined by gradient diffusion using M.I.C. Evaluator® strips.
Table 2.
MIC (μg/mL) of antibiotics for P. acidilactici ORE5 as determined by gradient diffusion using M.I.C. Evaluator® strips.
| Agent | P. acidilactici ORΕ 5 | Cut-Offa |
|---|---|---|
| (MIC μg/mL) | ||
| Amoxycillin | 2.41 ± 0.95 | n.r.b |
| Amoxycillin + Clavulanic acid | 0.25 ± 0.19 | n.r.b |
| Ampicillin | 1.28 ± 0.58 | 4 |
| Clindamycin | 1.91 ± 0.27 | 1 |
| Erythromycin | 1.77 ± 0.19 | 1 |
| Gentamycin | 7.23 ± 2.08 | 16 |
| Metronidazole | 159.2 ± 27.8 | n.r.b |
| Tetracycline | 10.97 ± 1.48 | 8 |
| Tigecycline | 0.49 ± 0.08 | n.r.b |
| Vancomycin | >256 | n.r.b |
aBreakpoints are referred to Pediococcus spp. EFSA breakpoints for other types of LABs are slightly different. Strains with MIC higher than the breakpoints are considered as resistant according to EFSA [22]. bnot required.
Table 3.
Effect of free and immobilized P. acidilactici ORE5 on pistachio nuts on physicochemical characteristics and sensory attributes of Katiki Domokou type cheese.
Table 3.
Effect of free and immobilized P. acidilactici ORE5 on pistachio nuts on physicochemical characteristics and sensory attributes of Katiki Domokou type cheese.
| Cheese1 | Days of storage | Water Activity (aw) |
pH | Acidity (g lactic acid/100g cheese) | % Moisture content | Overall sensory evaluation2 | |
|---|---|---|---|---|---|---|---|
| KC | 0 | 0.889 ± 0.01 | 4.28 ± 0.05 | 0.085 ± 0.02 | 72.71 ± 0.01 | 3.41 ± 0.70 | |
| 1 | 0.892 ± 0.02 | 4.27 ± 0.01 | 0.088 ± 0.04 | 73.32 ± 0.01 | |||
| 2 | 0.889 ± 0.01 | 4.33 ± 0.02 | 0.091 ± 0.05 | 74.20 ± 0.00 | |||
| 3 | 0.899 ± 0.05 | 4.29 ± 0.01 | 0.095 ± 0.03 | 74.71 ± 0.01 | |||
| 7 | 0.906 ± 0.01 | 4.31 ± 0.02 | 0.085 ± 0.01 | 75.51 ± 0.05 | |||
| 10 | 0.908 ± 0.02 | 4.45 ± 0.08 | 0.079 ± 0.02 | 77.82 ± 0.01 | |||
| 14 | 0.888 ± 0.03 | 4.72 ± 0.01 | 0.071 ± 0.01 | 78.90 ± 0.00 | |||
| KFP | 0 | 0.900 ± 0.01 | 4.08 ± 0.01 | 0.11± 0.02 | 73.94 ± 0.00 | 3.11 ± 0.74 | |
| 1 | 0.918 ± 0.02 | 4.11 ± 0.03 | 0.12± 0.07 | 74.11 ± 0.00 | |||
| 2 | 0.922 ± 0.03 | 4.14 ± 0.01 | 0.20± 0.05 | 74.21 ± 0.01 | |||
| 3 | 0.925 ± 0.05 | 4.12 ± 0.01 | 0.29± 0.03 | 74.42 ± 0.05 | |||
| 7 | 0.904 ± 0.07 | 4.35 ± 0.01 | 0.22 ± 0.02 | 74.81 ± 0.01 | |||
| 10 | 0.894 ± 0.06 | 4.38 ± 0.02 | 0.17 ± 0.01 | 75.50 ± 0.02 | |||
| 14 | 0.889 ± 0.01 | 4.45 ± 0.01 | 0.09 ± 0.01 | 78.35 ± 0.04 | |||
| KIP | 0 | 0.910 ± 0.01 | 4.15 ± 0.01 | 0.15 ± 0.05 | 63.91 ± 0.01 | 4.00 ± 0.54 | |
| 1 | 0.912 ± 0.02 | 4.18 ± 0.02 | 0.18 ± 0.05 | 65.72 ± 0.02 | |||
| 2 | 0.915 ± 0.02 | 4.23 ± 0.01 | 0.24 ± 0.01 | 71.51 ± 0.01 | |||
| 3 | 0.919 ± 0.03 | 4.22 ± 0.01 | 0.31 ± 0.02 | 72.11 ± 0.00 | |||
| 7 | 0.915 ± 0.05 | 4.66 ± 0.02 | 0.25 ± 0.03 | 72.91 ± 0.01 | |||
| 10 | 0.929 ± 0.01 | 4.68 ± 0.05 | 0.18 ± 0.01 | 73.51 ± 0.02 | |||
| 14 | 0.908 ± 0.05 | 4.70 ± 0.05 | 0.10 ± 0.01 | 74.13 ± 0.05 | |||
1KC: control samples, KFP: cheese samples containing free P. acidilactici ORE5 cells, KIP: cheese with immobilized P. acidilactici ORE5 cells on pistachio nuts. 2 The ratings ranged from 0 to 5 on a scale (0 = unacceptable; 5 = exceptional).
Table 4.
Effect of free or immobilized P. acidilactici ORE5 on pistachio nuts on total aerobic count, psychrophilic bacteria, lactococci, lactobacilli, yeasts and staphylococci in Katiki Domokou type cheese.
Table 4.
Effect of free or immobilized P. acidilactici ORE5 on pistachio nuts on total aerobic count, psychrophilic bacteria, lactococci, lactobacilli, yeasts and staphylococci in Katiki Domokou type cheese.
| Cheese1 | Days of storage | Total Aerobic Count (TAC) (logcfu/g) |
Psychrophilic bacteria (logcfu/g) |
Lactococci (logcfu/g) |
Lactobacilli (logcfu/g) |
Yeasts (logcfu/g) |
Staphylococci (logcfu/g) |
|---|---|---|---|---|---|---|---|
| KC | 0 | 6.50 ± 0.11 | 4.58 ± 0.01 | 4.80 ± 0.05 | 6.95 ± 0.01 | 6.39 ± 0.03 | 3.92 ± 0.03 |
| 1 | 6.44 ± 0.01 | 5.21 ± 0.03 | 5.00 ± 0.02 | 7.27 ± 0.01 | 6.70 ± 0.07 | 4.19 ± 0.06 | |
| 2 | 6.62 ± 0.02 | 5.38 ± 0.01 | 5,06 ± 0.03 | 7.42 ± 0.03 | 6.89 ± 0.03 | 4.33 ± 0.07 | |
| 3 | 6.74 ± 0.02 | 5.60 ± 0.08 | 5.43 ± 0.10 | 7.52 ± 0.02 | 7.33 ± 0.02 | 4.65 ± 0.01 | |
| 7 | 6.88 ± 0.01 | 5.85 ± 0.14 | 4.46 ± 0.09 | 7.08 ± 0.02 | 7.60 ± 0.08 | 5.35 ± 0.04 | |
| 10 | 6.74 ± 0.05 | 5.62 ± 0.01 | 4.25 ± 0.01 | 6.99 ± 0.01 | 7.99 ± 0.02 | 5.15 ± 0.01 | |
| 14 | 6.53 ± 0.01 | 5.52 ± 0.03 | 4.13 ± 0.07 | 6.78 ± 0.06 | 8.27 ± 0.01 | 5.07 ± 0.01 | |
| KFP | 0 | 8.30 ± 0.13 | 6.92 ± 0.04 | 4.81 ± 0.01 | 8.81 ± 0.01 | 6.38 ± 0.01 | 3.80 ± 0.01 |
| 1 | 8.63 ± 0.07 | 7.42 ± 0.02 | 5.10 ± 0.01 | 8.88 ± 0.07 | 6.50 ± 0.04 | 3.94 ± 0.05 | |
| 2 | 8.85 ± 0.03 | 7.38 ± 0.01 | 5.33 ± 0.02 | 8.91 ± 0.09 | 6.73 ± 0.02 | 4.09 ± 0.05 | |
| 3 | 8.96 ± 0.02 | 7.60 ± 0.08 | 5.76 ± 0.06 | 8.99 ± 0.09 | 7.03 ± 0.01 | 4.30 ± 0.08 | |
| 7 | 8.78 ± 0.05 | 7.85 ± 0.14 | 4.65 ± 0.01 | 8.57 ± 0.02 | 7.21 ± 0.05 | 4.70 ± 0.06 | |
| 10 | 8.63 ± 0.01 | 7.02 ± 0.01 | 4.42 ± 0.01 | 8.22 ± 0.01 | 7.52 ± 0.02 | 4.65 ± 0.02 | |
| 14 | 8.55 ± 0.01 | 6.52 ± 0.03 | 4.37 ± 0.04 | 8.09 ± 0.03 | 7.82 ± 0.11 | 4.57 ± 0.03 | |
| KIP | 0 | 8.88 ± 0.03 | 6.92 ± 0.04 | 4.82 ± 0.01 | 8.89 ± 0.02 | 6.35 ± 0.01 | 3.63 ± 0.02 |
| 1 | 8.95 ± 0.01 | 7.42 ± 0.02 | 5.13 ± 0.01 | 8.99 ± 0.05 | 6.43 ± 0.07 | 3.89 ± 0.00 | |
| 2 | 9.01 ± 0.04 | 7.38 ± 0.01 | 5.44 ± 0.01 | 9.09 ± 0.01 | 6.52 ± 0.03 | 4.02 ± 0.01 | |
| 3 | 9.07 ± 0.03 | 7.60 ± 0.08 | 5.97 ± 0.01 | 9.12 ± 0.02 | 6.97 ± 0.02 | 4.22 ± 0.05 | |
| 7 | 9.12 ± 0.02 | 7.85 ± 0.14 | 4.92 ± 0.08 | 8.96 ± 0.04 | 7.12 ± 0.01 | 4.54 ± 0.00 | |
| 10 | 8.99 ± 0.01 | 7.12 ± 0.02 | 4.72 ± 0.01 | 8.88 ± 0.01 | 7.45 ± 0.03 | 4.42 ± 0.01 | |
| 14 | 8.95 ± 0.02 | 6.52 ± 0.03 | 4.53 ± 0.03 | 8.85 ± 0.03 | 7.67 ± 0.02 | 4.38 ± 0.03 |
1KC: control samples, KFP: cheese samples containing free P. acidilactici ORE5 cells, KIP: cheese with immobilized P. acidilactici ORE5 cells on pistachio nuts.
Table 5.
Effect of free or immobilized P. acidilactici ORE5 on pistachio nuts on total aerobic count, psychrophilic bacteria, lactococci, lactobacilli, yeasts and staphylococci in Katiki Domokou type cheese after deliberate spiking with L. monocytogenes.
Table 5.
Effect of free or immobilized P. acidilactici ORE5 on pistachio nuts on total aerobic count, psychrophilic bacteria, lactococci, lactobacilli, yeasts and staphylococci in Katiki Domokou type cheese after deliberate spiking with L. monocytogenes.
| Cheese1 | Days of storage | Total Aerobic Count (TAC) (logcfu/g) |
Psychrophilic bacteria (logcfu/g) |
Lactococci (logcfu/g) |
Lactobacilli (logcfu/g) |
Yeasts (logcfu/g) |
Staphylococci (logcfu/g) |
L. monocytogenes (logcfu/g) |
|
|---|---|---|---|---|---|---|---|---|---|
| KC | 0 | 7.52 ± 0.04 | 4.69 ± 0.01 | 4.77 ± 0.03 | 6.89 ± 0.00 | 6.42 ± 0.03 | 3.99 ± 0.01 | 5.11 ± 0.01 | |
| 1 | 7.64 ± 0.06 | 5.42 ± 0.02 | 4.56 ± 0.02 | 6.93 ± 0.02 | 6.65 ± 0.01 | 4.11 ± 0.04 | 5.12 ± 0.01 | ||
| 2 | 8.23 ± 0.01 | 5.53 ± 0.01 | 4.44 ± 0.01 | 7.21 ± 0.05 | 6.95 ± 0.01 | 4.42 ± 0.01 | 5.15 ± 0.02 | ||
| 3 | 8.52 ± 0.03 | 6.28 ± 0.06 | 4.33 ± 0.04 | 7.36 ± 0.02 | 7.48 ± 0.02 | 4.65 ± 0.01 | 6.34 ± 0.06 | ||
| 7 | 8.96 ± 0.02 | 6.01 ± 0.04 | 4.11 ± 0.02 | 6.74 ± 0.02 | 7.68 ± 0.03 | 5.25 ± 0.04 | 6.28 ± 0.02 | ||
| 10 | 8.81 ± 0.01 | 5.89 ± 0.01 | 3.98 ± 0.01 | 6.65 ± 0.01 | 8.07 ± 0.05 | 4.79 ± 0.01 | 6.25 ± 0.05 | ||
| 14 | 8.77 ± 0.04 | 5.7 ± 0.06 | 3.66 ± 0.05 | 6.56 ± 0.02 | 8.25 ± 0.06 | 4.72 ± 0.03 | 6.22 ± 0.04 | ||
| KFP | 0 | 8.13 ± 0.06 | 6.69 ± 0.01 | 4.89 ± 0.05 | 8.32 ± 0.01 | 6.43 ± 0.01 | 3.78 ± 0.01 | 5.09 ± 0.06 | |
| 1 | 8.54 ± 0.01 | 7.21 ± 0.03 | 5.10 ± 0.01 | 8.38 ± 0.04 | 6.54 ± 0.07 | 3.89 ± 0.08 | 5.12 ± 0.01 | ||
| 2 | 8.57 ± 0.01 | 7.14 ± 0.02 | 5.33 ± 0.02 | 8.42 ± 0.07 | 6.85 ± 0.01 | 4.02 ± 0.02 | 5.15 ± 0.02 | ||
| 3 | 8.62 ± 0.04 | 7.54 ± 0.01 | 5.46 ± 0.06 | 8.45 ± 0.02 | 7.11 ± 0.04 | 4.28 ± 0.01 | 5.27 ± 0.02 | ||
| 7 | 8.56 ± 0.01 | 7.43 ± 0.04 | 4.65 ± 0.01 | 7.97 ± 0.03 | 7.32 ± 0.01 | 4.22 ± 0.01 | 5.43 ± 0.01 | ||
| 10 | 8.28 ± 0.02 | 6.89 ± 0.01 | 4.45 ± 0.01 | 7.88 ± 0.04 | 7.68 ± 0.05 | 4.15 ± 0.02 | 5.69 ± 0.05 | ||
| 14 | 8.18 ± 0.01 | 6.11 ± 0.07 | 4.37 ± 0.04 | 7.78 ± 0.04 | 7.96 ± 0.01 | 4.02 ± 0.01 | 5.49 ± 0.02 | ||
| KIP | 0 | 8.71 ± 0.02 | 6.77 ± 0.01 | 4.89 ± 0.01 | 8.42 ± 0.02 | 6.37 ± 0.01 | 3.70 ± 0.01 | 5.08 ± 0.04 | |
| 1 | 8.65 ± 0.05 | 7.24 ± 0.03 | 5.09 ± 0.01 | 8.49 ± 0.01 | 6.43 ± 0.07 | 3.83 ± 0.04 | 5.11 ± 0.04 | ||
| 2 | 8.67 ± 0.03 | 7.17 ± 0.02 | 5.31 ± 0.03 | 8.51 ± 0.01 | 6.59 ± 0.03 | 3.97 ± 0.03 | 5.14 ± 0.01 | ||
| 3 | 8.97 ± 0.01 | 7.57 ± 0.00 | 5.79 ± 0.05 | 8.54 ± 0.01 | 7.05 ± 0.02 | 4.27 ± 0.01 | 5.23 ± 0.01 | ||
| 7 | 8.76 ± 0.01 | 7.46 ± 0.02 | 4.69 ± 0.05 | 8.28 ± 0.01 | 7.20 ± 0.01 | 4.15 ± 0.01 | 5.55 ± 0.01 | ||
| 10 | 8.61 ± 0.02 | 6.94 ± 0.01 | 4.52 ± 0.01 | 8.11 ± 0.02 | 7.35 ± 0.01 | 4.12 ± 0.02 | 5.62 ± 0.04 | ||
| 14 | 8.54 ± 0.04 | 6.14 ± 0.07 | 4.39 ± 0.04 | 8.06 ± 0.04 | 7.72 ± 0.02 | 4.10 ± 0.01 | 5.45 ± 0.02 | ||
1KC: control samples, KFP: cheese samples containing free P. acidilactici ORE5 cells, KIP: cheese with immobilized P. acidilactici ORE5 cells on pistachio nuts.
Table 6.
Effect of free or immobilized P. acidilactici ORE5 on pistachio nuts on minor volatile compounds (μg/g) of Katiki Domokou type cheese after production (day 0) and after 3 days of storage.
Table 6.
Effect of free or immobilized P. acidilactici ORE5 on pistachio nuts on minor volatile compounds (μg/g) of Katiki Domokou type cheese after production (day 0) and after 3 days of storage.
| Compound | Identification Method | KC | KFP | KIP | |||
|---|---|---|---|---|---|---|---|
| d0 | d3 | d0 | d3 | d0 | d3 | ||
| Esters | |||||||
| Ethyl acetate | KI | N. D. | 0.1 | 0.6 | 13.8 | 0.5 | 6.4 |
| Ethyl propanoate | KI | N. D. | N. D. | N. D. | 0.1 | 0.0 | 0.1 |
| Ethyl butyrate | KI | N. D. | N. D. | 0.1 | 0.2 | 0.2 | 0.1 |
| 3- methylbutyl acetate | KI | N. D. | N. D. | N. D. | 0.2 | N. D. | 0.2 |
| Ethyl hexanoate | KI | N. D. | 0.2 | 0.8 | 1.8 | 0.6 | 0.6 |
| 2- phenylethyl acetate | KI | N. D. | 0.2 | 1.6 | 8.0 | 1.0 | 3.0 |
| Ethyl decanoate | KI | N. D. | N. D. | 0.1 | 1.8 | 0.2 | 0.2 |
| Organic acids | |||||||
| Hexanoic acid | KI | 0.1 | N. D. | N. D. | 0.8 | 0.2 | 0.4 |
| Sorbic acid | KI | 0.5 | 0.5 | 1.0 | 1.5 | 0.4 | 0.2 |
| Octanoic acid | KI | 0.1 | 0.4 | 0.1 | 3.5 | 0.1 | 0.1 |
| Benzoic acid | KI | 0.3 | 0.6 | 0.1 | 1.0 | 0.1 | 0.1 |
| Decanoic acid | KI | N. D. | 0.1 | 0.1 | 1.9 | 0.3 | 0.1 |
| Alcohols | KI | ||||||
| 2- methyl-1 propanol | KI | N. D. | N. D. | 0.1 | 0.2 | 0.3 | 0.5 |
| 3- methyl- 1 butanol | KI | 0.2 | 1.0 | 1.4 | 3.1 | 2.4 | 9.4 |
| 2- methyl- 1 butanol | KI | N. D. | N. D. | 0.3 | 0.4 | 0.4 | 1.4 |
| 1-hexanol | KI | N. D. | N. D. | N. D. | N. D. | 0.1 | 0.1 |
| 2- methyl- phenol | KI | N. D. | N. D. | N. D. | N. D. | 0.1 | 0.1 |
| 2- phenylethanol | KI | 0.2 | 1.3 | 0.9 | 1.3 | 1.1 | 3.1 |
| Carbonyl compounds | |||||||
| 2- methyl- propanal | KI | N. D, | N. D, | N. D. | N. D. | N.D. | 0.1 |
| Butanal | KI | N. D, | N. D. | N. D. | N. D. | N. D. | N. D. |
| 3- methyl- butanal | KI | N. D, | N. D. | N. D. | 0.4 | 0.1 | 0.2 |
| 2- methyl- butanal | KI | N. D, | N. D. | N. D. | N. D. | 0.1 | 0.1 |
| 2- pentanone | KI | N. D, | 0.1 | N. D. | N. D. | N. D. | N. D. |
| 3- hydroxy- butanone | KI | N. D, | N. D. | N. D. | N. D. | 0.3 | 0.4 |
| Hexanal | KI | N. D, | 0.1 | 0.2 | 0.1 | 0.1 | 0.1 |
| 2- heptanone | KI | N. D, | 0.1 | N. D. | N. D. | 0.1 | N. D. |
| Heptanal | KI | N. D, | N. D. | 0.1 | N. D. | N. D. | N. D. |
| Benzeneacetaldehyde | KI | N. D, | N. D. | N. D. | 0.1 | 0.1 | 0.1 |
| 2- nonanone | KI | N. D, | N. D. | N. D. | 0.1 | N. D. | N. D. |
| Miscellaneous Compounds | |||||||
| 2- methyl- 1,3- pentadiene | KI | N. D. | N. D. | N. D. | 0.1 | 0.1 | 0.2 |
| Octane | KI | N. D. | N. D. | 0.1 | 0.1 | 0.1 | 0.1 |
| 2,4- dimethyl- 1- heptane | KI | N. D. | N. D. | N. D. | N. D. | 0.3 | 0.4 |
| Styrene | KI | N. D. | N. D. | N. D. | 0.1 | N. D. | 0.1 |
| 2,6- dimethyl- pyrazine | KI | N. D. | N. D. | N. D. | N. D. | 0.2 | 0.1 |
| a- pinene | KI | N. D. | 0.1 | N. D. | N. D. | 0.2 | 0.1 |
| Decane | KI | N. D. | N. D. | 0.1 | 1.5 | 0.7 | 0.9 |
| Limonene | KI | N. D. | 0.1 | N. D. | N. D. | 0.3 | 0.2 |
| 3-ethyl-2,5-dimethyl-pyrazine | KI | N. D. | N. D. | N. D. | N. D. | 0.2 | 0.4 |
| Dodecane | KI | N. D. | N. D. | N. D. | 8.0 | 1.2 | 1.6 |
| 1,3-bis(1,1-dimethylethyl)-benzene | KI | N. D. | N. D. | 0.1 | 1.2 | 0.3 | 0.4 |
| Tetradecane | KI | N. D. | N. D. | N. D. | 1.0 | 0.1 | 0.1 |
KI: Tentative identification by kovats' retention index compared to the literature. N. D.: Not detected. KC: control samples, KFP: cheese samples containing free P. acidilactici ORE5 cells, KIP: cheese with immobilized P. acidilactici ORE5 cells on pistachio nuts.
Table 7.
Bacterial diversity indices Shannon’s and Simpson’s after 16S rRNA NGS sequencing in Katiki Domokou type cheese samples, calculated using Rhea platform and a-diversity script.
Table 7.
Bacterial diversity indices Shannon’s and Simpson’s after 16S rRNA NGS sequencing in Katiki Domokou type cheese samples, calculated using Rhea platform and a-diversity script.
| 16S rRNA OTUs | KC | KFP | KIP |
|---|---|---|---|
| Shannon’s Index | 0.71 ± 0.05b,c | 0.50 ± 0.03a,c | 0.89 ± 0.01 a,b |
| Simpson’s Index | 0.23 ± 0.02b, c | 0.14 ± 0.05a | 0.17 ± 0.02a |
KC: control sample, KFP: cheese sample containing free P. acidilactici ORE5 cells, KIP: cheese sample with immobilized P. acidilactici ORE5 cells on pistachio nuts. All data represent the mean ± standard deviation (SD) from two independent experiments. Significant differences: a (P < 0.05 vs KC), b (P < 0.05 vs KFP), c (P < 0.05 vs KIP).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



