
Submitted:
11 June 2023
Posted:
12 June 2023
You are already at the latest version
Abstract
MicroRNAs (miRNAs) regulate plant growth and development and their responses to biotic and abiotic stresses. Although extensive studies show that miR159 family members regulate leaf and flower development in Arabidopsis thaliana, the roles of miRNAs in soybean (Glycine max) are poorly understood. Here, we identified six MIR159 genes in soybean, MIR159aMIR159f, and investigate their expression patterns in plants under low-phosphorus (low-P), NaCl, or abscisic acid (ABA) treatments. In soybean leaves, MIR159e and MIR159f expression was induced by low-P treatment, while in roots, MIR159b, MIR159c, MIR159e, and MIR159f expression was upregulated. In flowers, low-P led to upregulation of MIR159a, MIR159b, MIR159c, and MIR159f, but downregulation of MIR159d and MIR159e. In soybean nodules, MIR159b was upregulated but MIR159a, MIR159c, and MIR159d was downregulated under P deficiency. NaCl treatment induced MIR159a, MIR159b, MIR159c, and MIR159e expression in leaves, and MIR159aMIR159f expression in roots. ABA treatment upregulated MIR159a, MIR159b, and MIR159c but downregulated MIR159d, MIR159e, and MIR159f in leaves. These results suggest that miR159 family members function in plant abiotic stress responses. Moreover, total P content in leaves was significantly lower in plants overexpressing MIR159e than in the wild type, suggesting that miR159e may regulate P absorption and transport in soybean.
Keywords:
MiR159
; Soybean
; Phosphorus
; NaCl
; ABA
; Gene function
1. Introduction
MicroRNAs (miRNAs) are non-coding small (20~24 nucleotide) RNAs that are widespread in plants and animals, as well as single-celled algae [1]. MiRNAs inhibit gene expression and translation after transcription [2]. Several conserved miRNA families have been identified. In plants, for example, miR156, miR159, miR164, miR168, miR169, miR171, miR319, miR390, miR393, miR399, and miR827 play crucial roles in growth and development, as well as in responses to biotic and abiotic stress. MiR159, in particular, regulates plant responses to nutrient stress, such as low-phosphate (low-P) stress. MIR159a, MIR319a, MIR396a, MIR389b, and MIR1507a are upregulated under low-P stress [3].
The miR159 family plays important roles in plant growth and development and in plant adaptation to abiotic stress [4]. Eight miRNAs associated with salt tolerance in have been identified in peanut (Arachis hypogea): miR159-1, miR159-2, miR159-3, miR164-2, miR167-3, miR319-1, miR319-2, and miR211-1 [5]. A study of the responses of salt tolerance related miRNAs to high-salt habitats in the mangrove companion plant Sesuvium portulacastrum showed that miR159 is involved in the responses of different tissues to high-salt stress [6]. However, the roles of miR159 family members in soybean (Glycine max) remain poorly understood.
The Arabidopsis (Arabidopsis thaliana) hyponastic leaves 1 (hyl1) mutant, which fails to process primary miRNA transcripts (pri-miRNAs), is sensitive to abscisic acid (ABA) during germination [7], suggesting that ABA signaling pathway might be regulated by miRNAs. Indeed, miR159 responds to ABA and drought treatment [8,9]. Analysis of the upstream regions of three MIR159 genes revealed the presence of ABA-responsive elements (ABREs) and associated stress factors such as AtMYC2 binding sites [10]. In line with this finding, ABRE-like elements were identified in the upstream region of the MIR159a promoter. MYB101 and MYB33 are targets of miR159; these genes encode MYB transcription factors that function in seed germination [11]. However, the responses of soybean MIR159 genes to abiotic stress and their roles in abiotic stress responses are unclear.
In this study, we identified six MIR159 genes in soybean: MIR159a−MIR159f. We then explored the responses of the soybean MIR159 gene family to low-P, NaCl, and ABA treatment. Finally, we overexpressed MIR159e in soybean. Under LP conditions, the total P content of MIR159e-overexpressing transgenic plants was significantly lower than the wild type in leaves but not roots. These results suggest that miR159e modulates the absorption and transport of P in soybean.
2. Materials and Methods
2.1. Plant materials and growth conditions
Seeds of soybean (Glycine max) genotype YC03-3 were sterilized with 10% NaClO, germinated in sand, and transferred to nutrient solution when the cotyledons were open and the apical bud had developed. To explore the responses of soybean miR159 family members to low-P stress, NaCl stress, and ABA treatment, seedlings with the first developed trifoliate leaf were transferred to high phosphate (Pi), high N (HP, 250 µM KH2PO4; HN, 5.3 mM nitrogen), N-deficient (HP, 250 µM KH2PO4; LN, 530 µM nitrogen), or Pi-deficient (LP, 5 µM KH2PO4; HN, 5.3 mM nitrogen; LN, 530 µM nitrogen) medium for 0 days (0 D), 7 days (7 D), and 40 days (40 D), respectively.
The plants were grown in growth chambers under a 16 h light /8 h dark cycle. The nutrient solutions were aerated for 15 min every 3 h and replaced with fresh nutrient solutions every 2 days. The roots, leaves, flowers and nodules were sampled, frozen in liquid nitrogen and stored at − 80℃. After 0 or 6 of ABA treatment, including −ABA (0 μM) or +ABA (300 μM), the treated seedlings were sampled, quickly frozen in liquid nitrogen, and stored at − 80℃. After 0 or 6 h of NaCl treatment, including − NaCl (0 mM) or +NaCl (200 mM), the seedlings were sampled, frozen in liquid nitrogen, and stored at − 80℃.
2.2. Extraction of total RNA, reverse-transcription, and extraction of genomic DNA
Total RNA was extracted from soybean seedlings using the TRIzol method. The RNA was reverse transcribed to cDNA using M-MLV reverse transcriptase. DNA was extracted from the samples using the CTAB method. Other molecular experiments were performed using standard methods as described [12].
2.3. qRT-PCR analysis
To quantify transcript levels in soybean genotype YC03-3 under different P conditions, qRT-PCR was used. The roots and leaves of YC03-3 seedlings that were subjected to low-P stress were quickly frozen in liquid N, and stored at − 80℃. Total RNA extraction and cDNA synthesis were performed using standard methods as described above. Soybean EF1a was used to normalize the PCR data. The expression levels of the six MIR159 genes (MIR159a, MIR159b, MIR159c, MIR159d, MIR159e, MIR159f) were measured in leaves, roots, and nodules. YC03-3 plants grown in soil were sampled and subjected to total RNA extraction using the TRIzol method. cDNAs were obtained using reverse transcriptase. The transcript abundance of the above-mentioned genes was determined via qRT-PCR, and AtEF1a was used as a reference gene to normalize the qRT-PCR results.
2.4. Measurement of fresh weight, soluble phosphate (Pi), and total P contents in soybean seedlings
Samples of soybean plants that were subjected to low-P stress for 40 days as described were weighed to quantify the fresh weight, including the fresh root and leaf. The lengths of roots and shoots were measured, and these values were used to calculate the root-to-shoot ratio. Soybean seedlings were dried at 105℃ for 30 min, oven-dried at 75℃ and then weighed. Soluble Pi and total P concentrations were measured exactly as described [13].
2.5. Data analysis
MEGA 6.0 software [14] was used to construct the phylogenetic tree using the neighbor-joining method. The promoters were analyzed using the TSSP program of Phytozome (http://phytozome.org) and softberry (http://softberry.org); target genes were predicted using psRNA Target (http://psrna target.org) [15].
All data were analyzed with Excel 2010. Student’s t-test was employed to identify significant differences between treatment groups. GraphPad Prism 8 was used to draw figures.
3. Results
3.1. Identification of MIR159 gene family members in soybean
To explore the MIR159 gene family, their target genes, and the evolution of this family in soybean, we obtained sequences of miR159 family members in soybean, rice (Oryza sativa), Arabidopsis, alfalfa (Medicago truncatula), maize (Zea mays) and Brachypodium distachyon from the miRBase website (http://www.mirbase.org). We found that the soybean MIR159 family consists of MIR159a, MIR159b, MIR159c, MIR159d, MIR159e, and MIR159f. MIR159a and MIR159d are located on chromosome 9, MIR159b and MIR159e are on chromosome 7, and MIR159c and MIR159f are on chromosome 16 (Table 1).
To decipher the evolutionary history of MIR159 in plants, we compared the MIR159 sequences of soybean with those from the plant species listed above. After aligning them using the CLUSTALW program, we analyzed their sequences using MrBayes3.2, finding that plant MIR159s belong to three subgroups (Figure 1). The soybean MIR159 genes fell into two subgroups: MIR159a, MIR159d, and MIR159e belong to subgroup I, and MIR159b, MIR159c, and MIR159f belong to subgroup II. Subgroup III contains only the MIR159 gene from monocotyledons (Figure 1). These results indicate that MIR159 genes are conserved across plant species, implying that they might play important roles in plant growth and development.
ABREs can be found in the promoter regions of MIR159 genes in Arabidopsis [10], and MIR159e responds to low-P stress in soybean [16]. To identify the cis-elements in these gene promoters, we analyzed the 2000-nucleotide sequence upstream of the start codon (ATG) in all six MIR159 genes (Table 2). As expected, we found phosphorus-responsive elements, NaCl-responsive elements and ABREs in the promoters of all six soybean MIR159 genes (Table 2). Among the phosphorus-responsive elements, we identified three or four TATA-box and W-box binding elements in each of the six MIR159 gene promoters, but no TATA-box-like binding elements, and we found that only MIR159c and MIR159f contain P1BS binding elements. Among the NaCl-responsive elements, all six MIR159 gene promoters contain one ABRE-like and one ACGT sequence binding element, but MIR159b and MIR159c lack RD22 binding elements, and MIR159f lacks AtMYB2 binding elements. Finally, among ABREs, all six MIR159 promoters contain a DPBF binding element, while MIR159c and MIR159f lack ABRE, and only MIR159e contains an RY binding element (Table 2). These results suggest that the transcription of MIR159 genes in soybean seem to be regulated by PHR1, WRKY, and ABF transcriptional regulators.
3.2. Responses of the soybean MIR159 gene family to low-P stress
Various miRNAs are involved in plant responses to low-P (LP) stress. Although MIR159a is upregulated in soybean under LP stress [3], the responses of other MIR159 gene family members to LP stress were not known. To examine this issue, we analyzed the expression of MIR159 genes in soybean roots, leaves, flowers, and nodules after 7 and 40 days of LP stress via quantitative reverse-transcription PCR (qRT-PCR). Compared to those in the high-phosphorus (HP) control, after 7 days of LP stress, MIR159a and MIR159e transcript levels were not significantly altered in leaves (Figure 2A, 2I) but were inhibited in roots (Figure 2B, 2J), whereas MIR159b expression was not significantly changed in leaves but was induced in roots (Figure 2D), MIR159c expression was inhibited in both leaves and roots (Figure 2E, 2F), and MIR159d and MIR159f expression was unaltered. After 40 days of LP stress, however, MIR159a expression was increased in leaves (Figure 2A), MIR159c expression was induced in roots (Figure 2F), and MIR159e and MIR159f expression was induced in both leaves and roots (Figure 2I, 2L). These results suggest that miR159 family members may regulate the responses of soybean to low-P stress in various ways.
MiR159 family members regulate flower development in Arabidopsis [17], and delay flowering under low-P stress [18], but their responses to nutrient deficiency in soybean are unknown. In this study, we analyzed the expression levels of soybean MIR159 gene family members in flowers, after 25 days of LP stress. Compared with that under HP stress, the expression of MIR159a, MIR159b, MIR159c and MIR159f was significantly induced in flowers under LP stress, whereas MIR159d and MIR159e expression was inhibited by this treatment (Figure 3A).
Soybean is a legume crop capable of biological nitrogen fixation; some miR159 family members regulate nodule growth and development in legumes [19]. We therefore cultured soybean plants under high-phosphorus and low-nitrogen (HPLN, control) and low-phosphorus and low-nitrogen (LPLN) conditions for 7 days and examined the samples 33 days after inoculation with rhizobia. We then examined the expression of MIR159 gene family members in nodules. Compared to HPLN, under LPLN conditions, the expression of MIR159a, MIR159c, and MIR159d in nodules was significantly inhibited, MIR159b expression was induced, and MIR159e and MIR159f expression did not significantly change (Figure 3B). These results indicate that MIR159 genes respond to low P stress in nodules.
3.3. Responses of the soybean MIR159 gene family to salt stress
MiR159 responds to high-salt stress in S. portulacastrum [6]. We therefore used qRT-PCR to measure the transcript levels of MIR159 family genes in leaves and roots of soybean plants subjected to short-term (6 h) treatment with 200 mM NaCl. MIR159a (Figure 4A, 4B), MIR159b (Figure 4C, 4D), MIR159c (Figure 4E, 4F), and MIR159e (Figure 4I, 4J) were significantly upregulated in leaves and roots under this treatment; MIR159d was upregulated only in roots (Figure 4G, 4H); and MIR159f was down-regulated in leaves but upregulated in roots (Figure 4K, 4L). These results suggest that the soybean MIR159 gene family plays important roles in plant responses to salt stress.
3.4. Responses of the soybean MIR159 gene family to ABA
To further assess the effects of MIR159 family genes in stress response, we investigated their expression upon exposure to the hormone ABA, a downstream factor in stress response pathways. MIR159a responds to ABA treatment in Arabidopsis [20], but the responses of other soybean MIR159 family genes to ABA remained unclear. In this study, qRT-PCR demonstrated that MIR159 gene expression was induced or inhibited in response to ABA. Compared to the control (−ABA), MIR159a was upregulated in leaves (Figure 5A), but not in roots, after 6 hours of ABA treatment (Figure 5B). In response to ABA treatment, MIR159b was upregulated in leaves and roots (Figure 5C, 5D), MIR159C was upregulated in leaves and downregulated in roots (Figure 5E, 5F), and MIR159d was downregulated in leaves (Figure 5G) and upregulated in roots (Figure 5H). MIR159e was down-regulated in leaves and roots (Figure 5I, 5J), and MIR159f was down-regulated in leaves and upregulated in roots (Figure 5K, 5L). These results suggest that MIR159 family genes play important roles in regulating ABA responses in soybean.
MiR159 transcript levels were quantified in samples from soybean plants of genotype YC03-3 that were transplanted when the first three-emerging compound leaf was fully unfolded and the second three-emerging compound leaf was not fully unfolded, and treated with ABA for 0 h or 6 h. −ABA, 0 μM; +ABA: 300 μM; h, hour. Results are means ± SE from 3 independent experiments. Student’s t-test was used to determine the differences between control and nutrient-deficiency conditions (*, P < 0.05; **, P < 0.01; ***, P < 0.001).
3.5. Overexpressing MIR159e decrease total phosphorus content in soybean leavea under LP conditions
To examine its functions, we overexpressed MIR159e driven by the constitutive CaMV 35S promoter in soybean genotype YC03-3. By screening for Basta resistance conferred by the Bar gene, we identified dozens of transgenic lines. Following confirmation using a chi-squared test, we selected two single-copy T-DNA insertion lines, #2 and #4, for subsequent experiments. After 40 days of growth under HP (250 μM KH2PO4) or LP (5 μM KH2PO4) conditions, the growth performance of lines #2 and #4 was not significantly different from that of wild-type YC03-3 (Supplemental Figure S1). Under HP conditions, the P (Figure 6A) and total nitrogen contents (Figure 6C) in #2 and #4 plants also were not significantly different from YC03-3 in leaves or roots. However, under LP conditions, the total P contents were significantly reduced in leaves of #2 and #4 plants compared to YC03-3 (Figure 6B), although the total nitrogen contents were not significantly different from YC03-3 (Figure 6D). Finally, under both HP and LP conditions, the soluble P concentrations of #2 and #4 plants in old leaves, new leaves, and roots were not significantly different from those of wild-type plants (Figure 6F). These results indicate that overexpressing MIR159e alters total phosphoru contents in soybean.
4. Discussion
Increasing evidence indicates that miRNAs play crucial roles in plant adaptation to nutrient stress. In the past decades, most studies on the roles of miRNAs in nutrient stress have been performed in model plants, such as Arabidopsis and rice, but little is known about their roles in legumes. Soybean is an important leguminous crop with great ability to fix nitrogen from the atmosphere, providing protein and edible oil for human consumption. MiRNAs are a class of non-coding small RNAs that function in the post-transcriptional regulation of their target genes by forming RNA-induced silencing complexes to shear the transcripts of their target genes or to inhibit the translation of these transcripts [21]. Nutrient deficiency in soils inhibits the growth and development of legumes, thereby reducing yields. MiR159 is a highly conserved miRNA family whose members are important in regulating vegetative growth, flowering, anther development, and seed germination in various plants [20,22,23]. However, the roles of the miR159 family in soybean are poorly understood.
Here, we identified six MIR159 genes in the soybean genome (Table 1), predicted cis-acting elements in their promoters, and identified candidate miR159 target genes (Table 2). We also examined the expression patterns of MIR159 genes in response to P (Figure 2, Figure 3), NaCl (Figure 4), and ABA (Figure 5) in soybean roots, leaves, flowers, and nodules. The highly conserved miRNA159 family is found in vascular plants and bryophytes. Evolutionary analysis suggested that miR159 shares a common ancestor with miR319, another highly conserved miRNA family [24]. Soybean MIR159 genes are arranged in clusters (Table 1), similar to MIR399 genes [16]. There are 3, 6, 11, 3, 3, and 1 MIR159 genes in alfalfa, maize, rice, Arabidopsis, Brachypodium distachyon, and common bean, respectively. Arabidopsis contains three MIR159 genes, but the soybean genome contains six, suggesting that the roles of soybean miR159 family members are more complex. In addition, the larger size of the soybean MIR159 gene family might be related to the two genome duplication events that occurred during the evolution of the soybean genome [25].
Like structural genes, MIRNA genes are also regulated at the transcriptional level [2]. In this study, three or four TATA-box and W-box binding elements were found in the promoters of all six soybean MIR159 genes. However, MIR159e lacks TATA-box elements, and only MIR159c and MIR159f contain P1BS binding elements (Table 2). MIR159c and MIR159f transcription might be regulated by PHR-type transcriptional regulators in soybean [26]. MIR159a was upregulated in leaves in response to 40 days of low-P stress but was downregulated in roots after 7 days of this treatment (Figure 2A), which is consistent with the results of previous studies [3]. MIR159c expression was inhibited in leaves after 7 days of low-P stress (Figure 2E) but induced in roots after 40 days (Figure 2F). MIR159e was upregulated in both leaves and roots under low-P stress (Figure 2I, 2J), which is consistent with previous reports [16]. MIR159 was upregulated in leaves and roots after 40 days of low-P stress (Figure 2K, 2L). In addition, in flowers, MIR159a, MIR159b, MIR159c, and MIR159f expression was induced, while MIR159d and MIR159e expression was inhibited after 25 days of low-P stress (Figure 3A). Therefore, the transcription of these MIR159 genes is regulated by low-P stress and may be related to the presence of low-P-responsive elements (Table 2). Unlike some other stresses, low -P stress delays flowering [18].
Overexpressing MIR159a in gloxinia (Sinningia speciosa) delays flowering under short-day conditions and downregulates the expression of LEAFY (LFY), AGAMOUS, APETALA1 (AP1) and AP3 genes in flower buds [27]. MIR159 and MYB33 are co-transcribed in the aleurone layer and embryo in germinating Arabidopsis seeds, and the two genes show the same spatiotemporal expression pattern [28]. The Arabidopsis mir159a mir159b double mutant shows plant dwarfing, reduced apical dominance, reduced fertility, and an irregular seed shape [17]. However, whether soybean miR159 is functionally conserved with Arabidopsis miR159 requires further study. Soybean MIR159 is expressed in floral organs and is induced by low-P stress (Figure 3A), but the role of soybean miR159 in regulating flower development remains unclear. In this study, we determined that the expression of MIR159a, MIR159b, MIR159c, and MIR159f increases under low-P conditions. This likely leads to the inhibition of flowering-related genes, such as LFY and AP, thereby delaying flowering under low-P conditions. Of course, further research is needed to confirm or disprove this theory.
Nodules are the sites of symbiosis between leguminous crops and rhizobia. Since nitrogen fixation in nodules requires ATP, maintaining the balance between P and other nutrients is crucial [29,30]. MIR159 is expressed at a relatively high level in alfalfa nodules [31]. Here, we showed that MIR159a, MIR159c, and MIR159d were down-regulated, MIR159b was upregulated, and MIR159e and MIR159f expression was unchanged under low-P stress (Figure 3B). We also noted that MIR159 genes in soybean showed different response patterns to low-P stress in roots (Figure 2) and nodules (Figure 3B), possibly due to different nutritional conditions.
Compared to wild-type soybean (YC03-3), transgenic plants overexpressing MIR159 induced drought resistance, suggesting that miR159 functions in drought resistance and perhaps plant responses to high-salt stress [32]. MIR159 expression is induced in peanut by salt stress, and miR159 regulates the expression of its target genes [33]. In addition, miR159 in different tissues of S. portulacastrum is involved in plant responses to high-salt stress [6]. In the current study, qRT-PCR results showed that MIR159 responds to high-NaCl concentrations in soybean (Figure 4). One ABRE-like and one ACGT sequence binding element were identified in each of the six MIR159 gene promoters in soybean (Table 2). However, MIR159b and MIR159c lack rd22 binding elements, which are found in the other four soybean MIR159 genes. MIR159a−MIR159e contain 2, 1, 2, 3, and 4 AtMYB2 binding elements, respectively, whereas MIR159f did not lack AtMYB2 binding elements (Table 2). Therefore, to understand the functions of soybean miR159s, it will be important to identify the transcriptional regulators of the response of this plant to salt stress and the binding of these regulators to the MIR159 promoters.
Analysis of ABREs showed that all six MIR159 promoters in soybean contain a DPBF-binding element, whereas MIR159c and MIR159f lack ABREs and only MIR159e contains RY-binding elements (Table 2), suggesting that soybean miR159s may respond to ABA signals. In line with this notion, both MIR159a and MIR159b were upregulated by ABA treatment in Arabidopsis [20]. ABA plays an important role in regulating root development, seed maturation and germination, and drought and salt stress tolerance. Several ABA-responsive miRNAs have been identified, such as miR399f in Arabidopsis [34]. Previous studies demonstrated that the Arabidopsis hyl1 mutant is sensitive to ABA during seed germination [7], suggesting that the ABA signaling pathway might be regulated by miRNAs. By isolating and cloning Arabidopsis miRNAs during early germination, Abe et al. (2003) determined that miR159 responds to ABA and drought treatment. Analysis of the upstream regions of the three Arabidopsis MIR159 genes revealed ABREs and binding sites for stress-related transcription factors such as AtMYC2 [10]. The expression of MIR159a and MIR159b is induced by ABA in Arabidopsis at the seed germination stage, but not at the seedling stage [20]. In the current study, the results in Figure 5 suggest that miR159 family members play important roles in regulating plant responses to ABA in soybean. All six MIR159 gene promoters contain DPNF binding elements, whereas MIR159e and MIR159f lack ABREs, and only MIR159e contains RY elements (Table 2), which might help explain the different responses of individual soybean MIIR159 family members to exogenous ABA.
In transgenic soybean plants overexpressing MIR159e and wild-type soybean plants under different P treatments for 40 days, the fresh weights of both transgenic and wild-type plants were lower under low-P vs. high-P treatment (Supplemental Figure S1A), which is consistent with previous findings [35,36]. Under HP and LP conditions, there was no significant difference in fresh weight, primary root length, root area, total P, soluble P, or total nitrogen contents between MIR159e-overexpressing transgenic and wild-type YC03-3 plants (Supplemental Figure S1). However, the primary roots of the Arabidopsis miR159ab double mutant are longer than those of the wild type, and its meristem is enlarged [37]. Under HPLN conditions, MIR159e-overexpressing soybean plants showed reduced total P contents in leaves and roots (Supplemental Figure S2A) and reduced soluble P concentrations in old leaves and nodules compared to the wild type (Supplemental Figure S2C). These results indicate that overexpressing MIR159e affects the uptake and reuse of P in soybean roots under low-nitrogen stress. MIR159e might be involved in nitrogen and P, but the physiological and molecular mechanisms are unclear.
Whether overexpressing MIR159e affected the expression of a phosphate transporter gene and the abundance of its encoded protein in soybean; this should be further studied under low P or low-nitrogen conditions. Under low-nitrogen conditions, the total nitrogen content was higher in nodules than in leaves and roots, indicating that nodules play a role in nitrogen accumulation and provide nitrogen for plant growth (Supplemental Figure S2). These results are consistent with previous findings [36]. Overexpressing MIR159e did not affect the total nitrogen content in leaves, roots, or nodules under HPHN, LPHN, or HPLN conditions (Figure 6, Supplemental Figure S2). These results suggest that miR159e might not be involved in regulating nutrient balance in plants. Overexpressing MIR159 delayed flowering in ornamental gloxinia (Sinningia speciosa) [27]. However, in the current study, overexpressing MIR159e in soybean did not delay flowering. It will be important to design experiments to determine whether overexpressing MIR159e alters the sensitivity of soybean to ABA and NaCl. In the future, CRISPR-Cas9 or miRNA-STTM technology is needed to knock out or knock down MIR159 expression to further analyze their functions.
5. Conclusions
In this study, we demonstrated that the soybean genome contains six MIR159 family members, which are divided into two subgroups. The promoter regions of soybean MIR159 genes contain phosphorus signal response and NaCl and ABA response elements, and the transcription of various soybean MIR159 genes is regulated by low-P stress, NaCl and ABA, suggesting that the miR159 family plays important roles in abiotic stress in soybean.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplemental Table S1. Predicted miR159 target genes in soybean; Supplemental Figure S1. Effects of MIR159e over expression on growth and development. WT, wild type (YC03-3); OE-2, miR159eOE-2; OE-4, miR159eOE-4; HP, plants treated with a high phosphorus concentration of 250 μM; LP, plants treated with a low-P concentration of 5 μM; treatment time was 40 days. Results are means ± SE from three independent experiments. Student’s t-test was used to compare the differences between Col-0 and transgenic MIR159eOE plants (*P < 0.05); Supplemental Figure S2. Total phosphorus, nitrogen, and soluble phosphorus contents in soybean overexpressing MIR159e under low-nitrogen conditions. WT, wild type (YC03-3); HPLN, high phosphorus and low nitrogen; treatment was performed for 40 days. Results are means ± SE from three independent experiments. Student’s t-test was used to compare the differences between Col-0 and the transgenic line MIR159eOE in the same plant part sampled at the same time point (*P < 0.05); Supplemental Table S2. List of primer pairs used in this study.
Author Contributions
J.W., B.Li. and P.T. conceived and designed the experiments. B. Li., P.T., F.X., and P.H. performed the experiments. B. Li. analyzed the date. J.W. and B. Li wrote the original manuscript. J.W. revised and approved the final version of the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Natural Science Foundation of China (Grant number: 31572184).
Data Availability Statement
All data are provided as figures and table which are included in this paper.
Conflicts of Interest: The authors declare no conflict of interest.
References
- Jonesrhoades, M.W.; Bartel, D.P.; Bartel, B. MicroRNAs and their Regulatory roles in Plants. Annu. Rev. Plant Biol 2006, 57, 19–53. [Google Scholar] [CrossRef]
- Rogers, K.; Chen, X. Biogenesis, turnover, and mode of action of plant microRNAs. Plant Cell 2013, 25, 2383–2399. [Google Scholar] [CrossRef]
- Zeng, H.Q.; Zhu, Y.i.Y.; Huang, S.i.Q.; Yang, Z.M. Analysis of phosphorus-deficient responsive miRNAs and cis-elements from soybean (Glycine max L.). J Plant Physiol 2010, 167, 1289–1297. [Google Scholar] [CrossRef]
- Mallory, A.C.; Vaucheret, H. Functions of microRNAs and related small RNAs in plants. Nat Genet 2006, 38, S31–S36. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xie, M.; Ren, G.; Yu, B. CDC5, a DNA binding protein, positively regulates posttranscriptional processing and/or transcription of primary microRNA transcripts. PNAS 2013, 110, 17588–17593. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.N.; Liu, Q.; Zhang, Y. Response of salt-tolerant mirnas to high-salt habitats in mangrove companion plant hippocampal teeth. MPB 2017, 15, 1137–1142. [Google Scholar]
- Lu, C.; Fedoroff, N. A Mutation in the Arabidopsis HYL1 Gene Encoding a dsRNA Binding Protein Affects Responses to Abscisic Acid, Auxin, and Cytokinin. Plant Cell 2000, 12, 2351–2366. [Google Scholar] [CrossRef]
- Lopez-Molina, L.; Mongrand, S.; Mclachlin, D.T.; Mclachlin, B.T.; Chait, N.-H.C. ABI5 acts downstream of ABI3 to execute an ABA-dependent growth arrest during germination. Plant J 2002, 32, 317–328. [Google Scholar] [CrossRef]
- Zhang, B.H.; Pan, X.P.; Wang, Q.L.; Cobb, G.P.; Anderson, T.A. Identification and characterization of new plant microRNAs using EST analysis. Cell Res 2005, 15, 336–360. [Google Scholar] [CrossRef]
- Abe, H.; Urao, T.; Ito, T.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) Function as Transcriptional Activators in Abscisic Acid Signaling. Plant Cell 2003, 15, 63–78. [Google Scholar] [CrossRef]
- Fujii, H.; Chiou, T.J.; Lin, S.I.; Aung, K. A miRNA Involved in Phosphate-Starvation Response in Arabidopsis. Current Biology CB 2005, 15, 2038–2043. [Google Scholar] [CrossRef] [PubMed]
- Huo, W.G.; Li, B.D.; Kuang, J.B. Functional Characterization of the Steroid Reductase Genes GmDET2 and GmDET2b from Glycine max. Int J Mol Sci 2018, 19, 726. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Jiao, F.; Wu, Z.; Li, Y.; Wang, X.; He, X.; Zhong, W.; Wu, P. OsPHR2 is involved in phosphate-starvation signaling and excessive phosphate accumulation in shoots of plants. Plant Physiol 2008, 146, 1673–1686. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Peterson, D.; Tamura, K. MEGA-CC: Computing core of molecular evolutionary genetics analysis program for automated and iterative data analysis. Bioinformatics 2012, 28, 2685–2686. [Google Scholar] [CrossRef]
- Dai, X.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server. Nucleic Acids Research 2011, 39, W155–W159. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Liu, Q.; Chen, L.; Kuang, J.B.; Liao, H. Genome-wide identification of soybean microRNAs and their targets reveals their organ-specificity and responses to phosphate starvation. BMC Genomics 2013, 14, 66. [Google Scholar] [CrossRef]
- Allen, R.S.; Li, J.; Alonsoperal, M.M. MicroR159 regulation of most conserved targets in Arabidopsis has negligible phenotypic effects. Silence 2010, 1, 1–18. [Google Scholar] [CrossRef]
- Nord, E.A.; Lynch, J.P. Delayed reproduction in Arabidopsis thaliana improves fitness in soil with suboptimal phosphorus availability. Plant Cell & Environment 2008, 31, 1432–1441. [Google Scholar]
- Xu, F.; Wang, J.X. Research progress of MicroRNA regulating nutrient stress response of leguminous crops. Plant Nutrition and Fertilizer Journal 2016, 22, 236–244. [Google Scholar]
- Reyes, J.L.; Chua, N. ABA induction of miR159 controls transcript levels of two MYB factors during Arabidopsis seed germination. Plant Journal for Cell & Molecular Biology 2007, 49, 592–606. [Google Scholar]
- Khraiwesh, B.; Zhu, J.K.; Zhu, J. Role of miRNAs and siRNAs in biotic and abiotic stress responses of plants. Biochimica et Biophysica Acta 2012, 1819, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Millar, A.A.; Gubler, F. The Arabidopsis GAMYB-Like Genes, MYB33 and MYB65, Are MicroRNA-Regulated Genes That Redundantly Facilitate Anther Development. Plant Cell 2005, 17, 705–721. [Google Scholar] [CrossRef]
- Allen, R.S.; Li, J.; Stahle, M.I. Genetic analysis reveals functional redundancy and the major target genes of the Arabidopsis miR159 family. PNAS 2007, 104, 16371–16376. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jin, Y.X. Evolution of MIR159/319 MicroRNA Genes and Their Post-transcriptional Regulatory Link to siRNA Pathways. BMC Evol Biol 2011, 11, 122. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.X.; Mitros, T.; Nelson, W.; Hyten, D. Genome sequence of the paleopolyploid soybean (Glycine max (L.) Merr.). Nature 2010, 463, 178–183. [Google Scholar] [CrossRef]
- Bustos, R.; Castrillo, G.; Linhares, F.; Puga, M.I.; Rubio, V.; P’erez-Perez, J.; Solano, R.; Leyva, A.; Paz-Ares, J. A central regulatory system largely controls transcriptional activation and repression responses to phosphate starvation in Arabidopsis. PLOS Genet 2010, 6, e1001102. [Google Scholar] [CrossRef]
- Bian, X.Y.; Song, D.F.; Han, S.Y.; Wang, J.H.; Zhu, M.Y. Flowering time control in ornamental gloxinia (Sinningia speciosa) by manipulation of miR159 expression. Annals of Botany 2013, 111, 791–799. [Google Scholar]
- Csukasi, F.; Donaire, L.; Casañal, A.; Martínez-Priego, L.; Botella, M.A.; Medina-Escobar N,Llave, C.; Valpuesta, V. Two strawberry miR159 family members display developmental-specific expression patterns in the fruit receptacle and cooperatively regulate Fa-GAMYB. New Phytol 2012, 195, 47–57. [Google Scholar] [CrossRef]
- Olivera, M.; Tejera, N.; Lribarne, C.; Ocaña, A.; Lluch, C. Growth, nitrogen fixation and ammonium assimilation in common bean (Phaseolus vulgaris): Effect of phosphorus. Physiol Plantarum 2004, 121, 498–505. [Google Scholar] [CrossRef]
- Schulze, J.; Temple, G.; Temple, S.J.; Beschow, H.; Vance, C.P. Nitrogen fixation by white lupin under phosphorus deficiency. Annals of Botany 2006, 98, 731–740. [Google Scholar] [CrossRef]
- Zucchero, J.C.; Caspi, M.; Dunn, K. ngl9: A third MADS box gene expressed in alfalfa root nodules. Mol Plant Microbe Interact 2001, 14, 1463–1467. [Google Scholar] [CrossRef]
- Li, Y.J.; Alonso-Peral, M.; Wong, G.; Wang, M.B.; Millar, A.A. Ubiquitous miR159 repression of MYB33/65 in Arabidopsis rosettes is robust and is not perturbed by a wide range of stresses. Bmc Plant Biology 2016, 16, 179. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.T.; Yan, C.X.; Zhao, X.B.; Li, C.J.; Shi, C.R.; Yu, J.J.; Chan, S.H.; Li, R.G. Identification and functional characterization of salt tolerance related microRNAs in roots of peanut (Arachis hypogaea L.). Journal of Peanut Science 2016, 45, 36–42. [Google Scholar]
- Panda, S.K.; Sunkar, R. Nutrient- and other stress-responsive microRNAs in plants: Role for thiol-based redox signaling. Plant Signaling & Behavior 2015, 10, e1010916. [Google Scholar]
- Raghothama, K.G. Phosphate Acquisition. Annu Rev Plant Phys 1999, 274, 665–693. [Google Scholar] [CrossRef] [PubMed]
- Reinhart, B.J.; Weinstein, E.G.; Rhoades, M.W.; Bartel, B.; Bartel, D. MicroRNAs in plants. Genes & Development 2002, 16, 1616–1626. [Google Scholar]
- Wang, X.R.; Yan, X.L.; Liao, H. Genetic improvement for phosphorus efficiency in soybean: A radical approach. Ann Bot 2010, 106, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Tao, X.; Liu, Z.; Dai, X.; Xiang, F. Primary root growth in Arabidopsis thaliana is inhibited by the miR159 mediated repression of MYB33, MYB65 and MYB101. Plant Sci 2017, 262, 182–189. [Google Scholar]
Figure 1.
Phylogenetic analysis of the MIR159 family in soybean and other plant species. Note: Abbreviations: gma, Glycine max; ath, Arabidopsis thaliana; osa, Oryza sativa; mtr, Medicago truncatula; zma, Zea mays; bdi, Brachypodium distachyon; pvu, Phaseolus vulgaris.
Figure 1.
Phylogenetic analysis of the MIR159 family in soybean and other plant species. Note: Abbreviations: gma, Glycine max; ath, Arabidopsis thaliana; osa, Oryza sativa; mtr, Medicago truncatula; zma, Zea mays; bdi, Brachypodium distachyon; pvu, Phaseolus vulgaris.
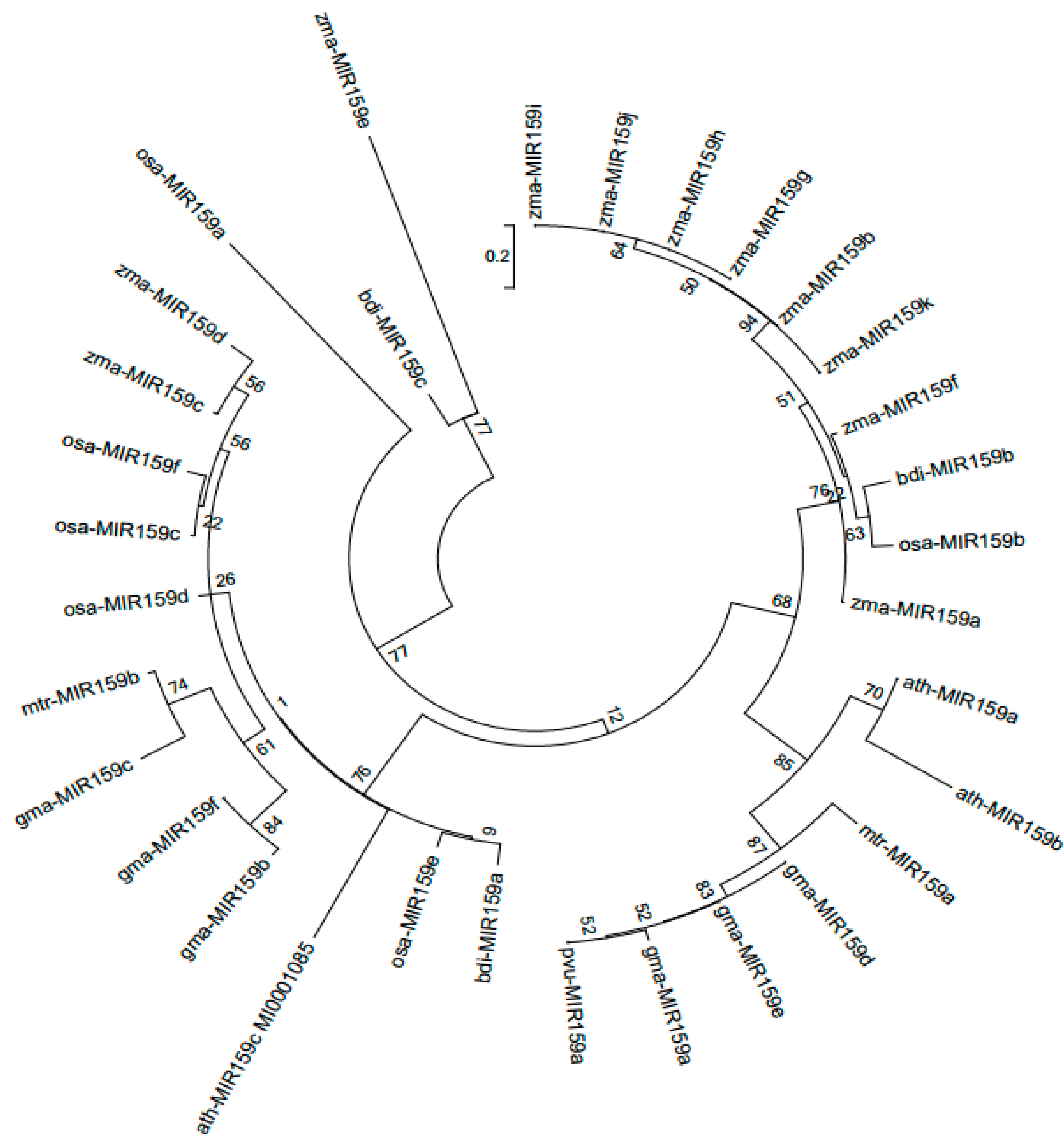
Figure 2.
Responses of the soybean MIR159 gene family to low-P stress in leaves and roots. MIR159 transcript levels were quantified in samples from soybean plants of genotype YC03-3 that were transplanted when the first three-emerging compound leaf was fully unfolded and the second three-emerging compound leaf was not fully unfolded and treated with high and low P (HP, 250 μM KH2PO4; LP, 5 μM KH2PO4) for 7 days (7 D) or 40 days (40 D). Results are means ± SE from 3 independent experiments. Student’s t-test was used to determine the differences between control and nutrient-deficiency conditions (*, P < 0.05; ***, P < 0.001).
Figure 2.
Responses of the soybean MIR159 gene family to low-P stress in leaves and roots. MIR159 transcript levels were quantified in samples from soybean plants of genotype YC03-3 that were transplanted when the first three-emerging compound leaf was fully unfolded and the second three-emerging compound leaf was not fully unfolded and treated with high and low P (HP, 250 μM KH2PO4; LP, 5 μM KH2PO4) for 7 days (7 D) or 40 days (40 D). Results are means ± SE from 3 independent experiments. Student’s t-test was used to determine the differences between control and nutrient-deficiency conditions (*, P < 0.05; ***, P < 0.001).
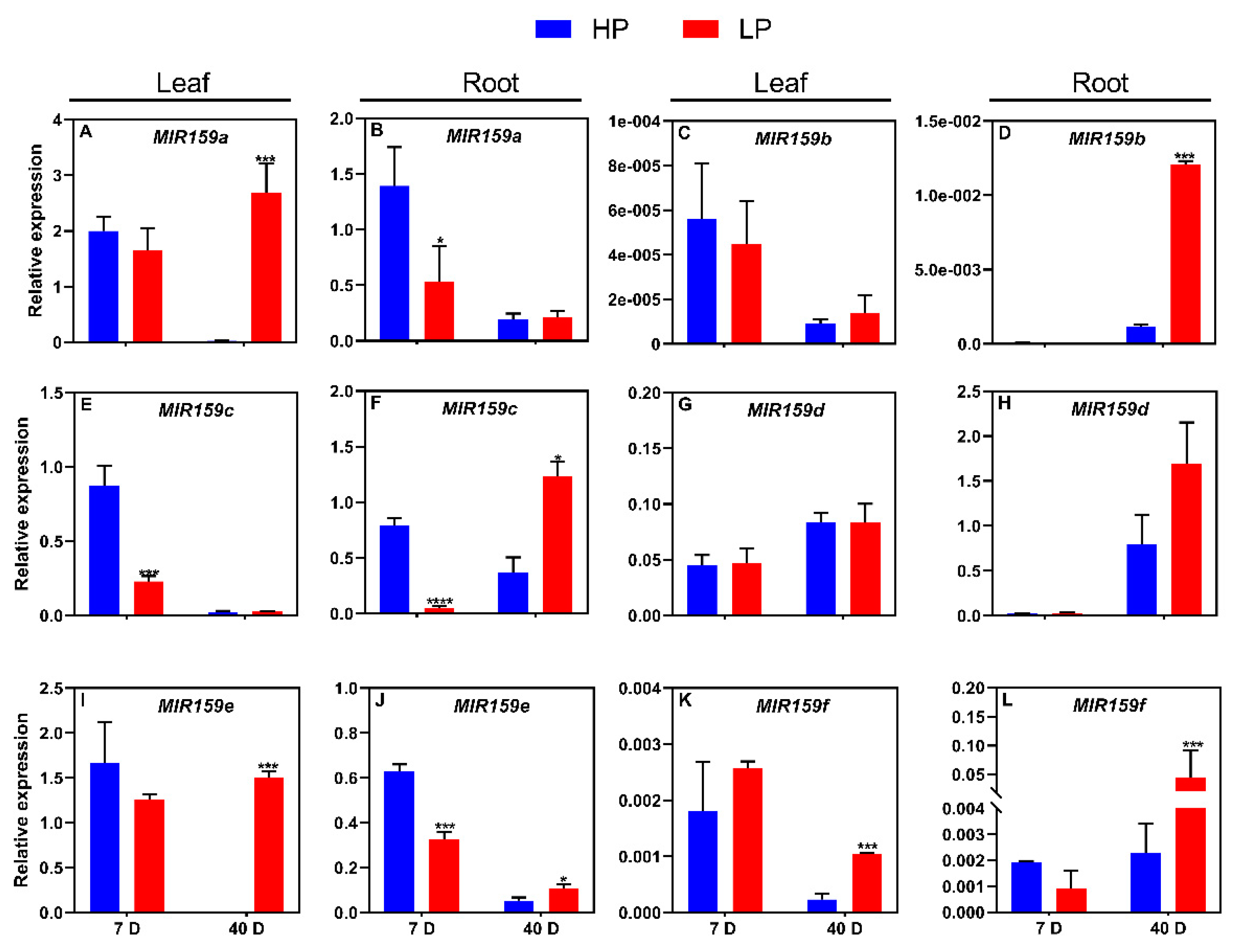
Figure 3.
Responses of the soybean MIR159 gene family to low-P stress in flowers and nodules. (A) MIR159 gene expression in flowers; (B) MIR159 gene expression in nodules. Soybean genotype YC03-3 was transplanted when the first three-emerging compound leaf was fully unfolded and the second three-emerging compound leaf was not fully unfolded, and treated with high and low P (HP, 250 μM KH2PO4; LP, 5 μM KH2PO4) for 25 days. Results are means ± SE from 3 independent experiments. Student’s t-test was used to determine the differences between control and nutrient deficiency conditions (*, P < 0.05; ***, P < 0.001).
Figure 3.
Responses of the soybean MIR159 gene family to low-P stress in flowers and nodules. (A) MIR159 gene expression in flowers; (B) MIR159 gene expression in nodules. Soybean genotype YC03-3 was transplanted when the first three-emerging compound leaf was fully unfolded and the second three-emerging compound leaf was not fully unfolded, and treated with high and low P (HP, 250 μM KH2PO4; LP, 5 μM KH2PO4) for 25 days. Results are means ± SE from 3 independent experiments. Student’s t-test was used to determine the differences between control and nutrient deficiency conditions (*, P < 0.05; ***, P < 0.001).
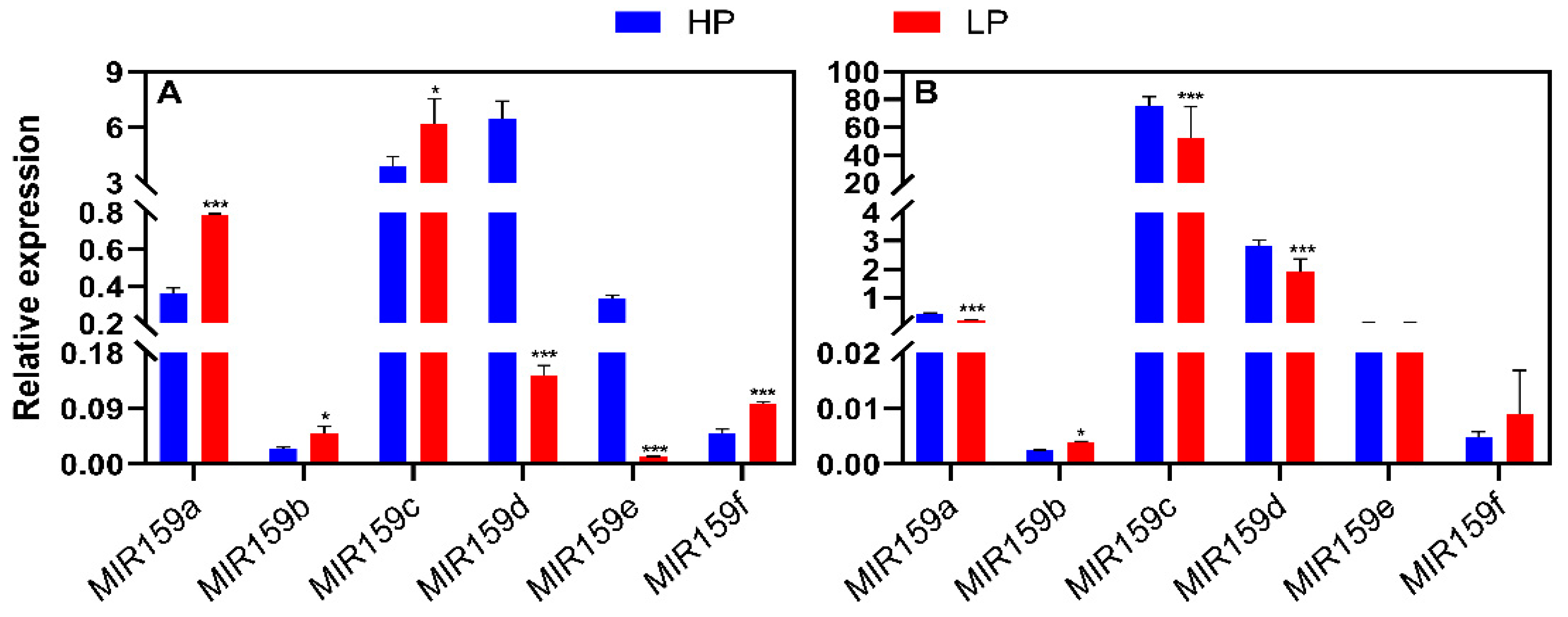
Figure 4.
Responses of the soybean MIR159 gene family to NaCl stress in leaves and roots. MIR159 transcript levels were quantified in samples from soybean plants of genotype YC03-3 that were transplanted when the first three-emerging compound leaf was fully unfolded and the second three-emerging compound leaf was not fully unfolded, and treated NaCl with for 0 or 6 h. −NaCl, 0 mM; +NaCl, 200 mM; h, hours. Results are means ± SE from 3 independent experiments. Student’s t-test was used to determine the differences between control and nutrient deficiency conditions (*, P < 0.05; **, P < 0.01; ***, P < 0.001).
Figure 4.
Responses of the soybean MIR159 gene family to NaCl stress in leaves and roots. MIR159 transcript levels were quantified in samples from soybean plants of genotype YC03-3 that were transplanted when the first three-emerging compound leaf was fully unfolded and the second three-emerging compound leaf was not fully unfolded, and treated NaCl with for 0 or 6 h. −NaCl, 0 mM; +NaCl, 200 mM; h, hours. Results are means ± SE from 3 independent experiments. Student’s t-test was used to determine the differences between control and nutrient deficiency conditions (*, P < 0.05; **, P < 0.01; ***, P < 0.001).
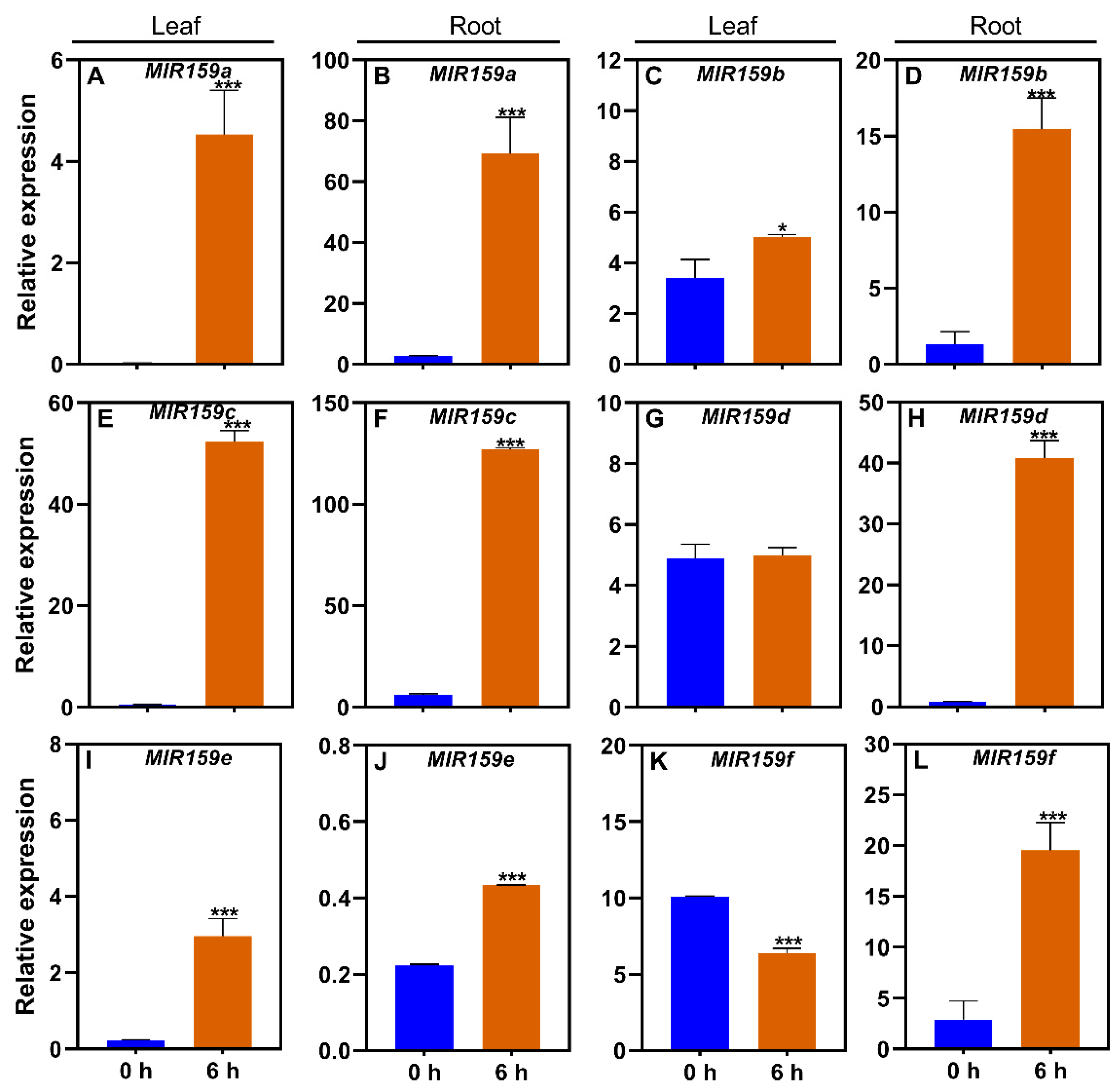
Figure 5.
Responses of the soybean MIR159 gene family to ABA in leaves and roots.
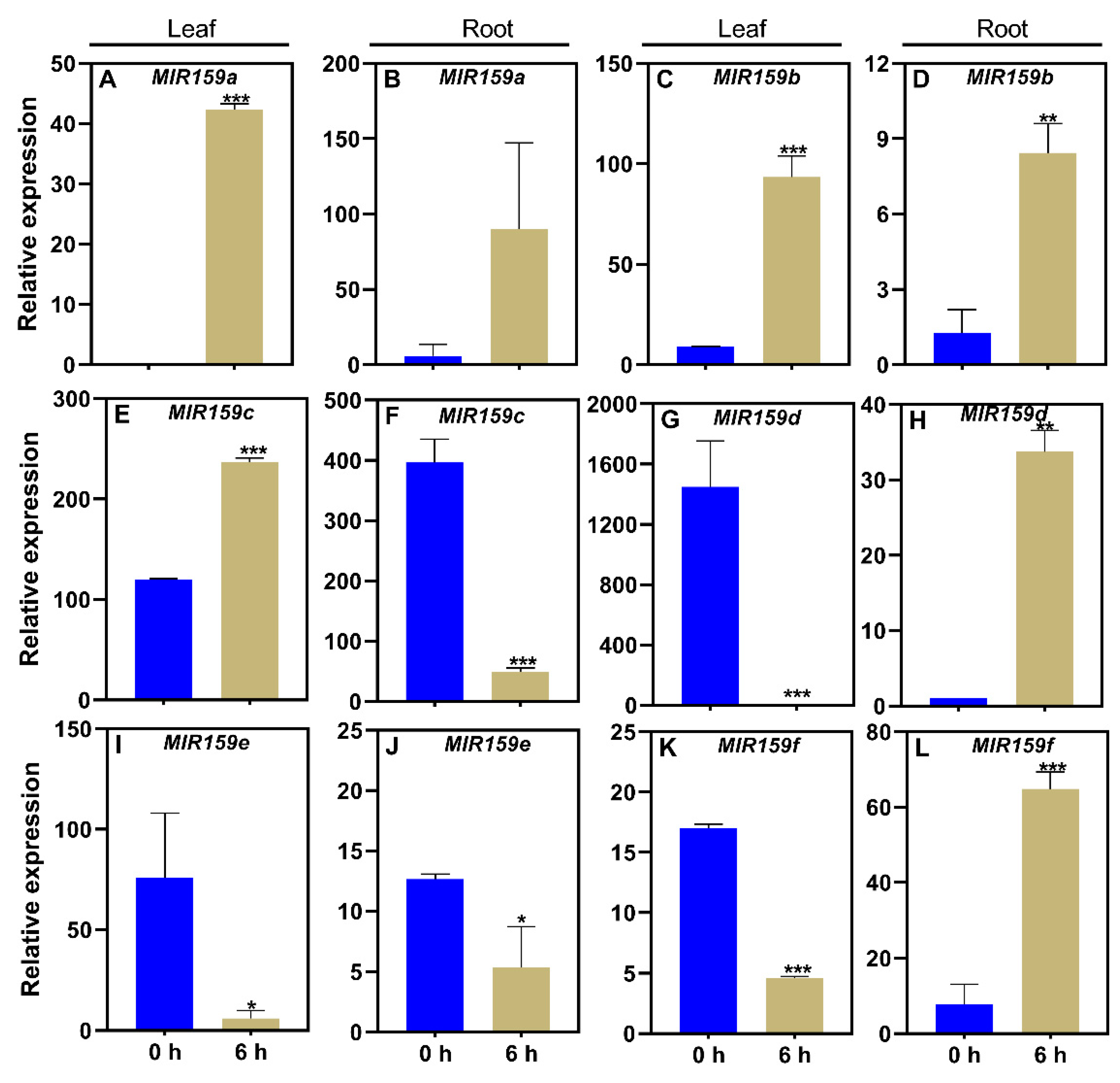
Figure 6.
Overexpressing MIR159e affects total phosphorus, total nitrogen, and soluble phosphorus contents in soybean. WT, wild type (YC03-3); OL, old leaves; YL, young leaves. Soybean genotype YC03-3 was transplanted when the first emerging compound leaf was fully unfolded and the second emerging compound leaf was not fully unfolded, and treated with high and low P (HP, 250 μM KH2PO4; LP, 5 μM KH2PO4) for 40 days (40D). Results are means ± SE from 3 independent experiments. Student’s t-test was used to determine the differences between control and nutrient deficiency conditions (**, P < 0.01; ***, P < 0.001).
Figure 6.
Overexpressing MIR159e affects total phosphorus, total nitrogen, and soluble phosphorus contents in soybean. WT, wild type (YC03-3); OL, old leaves; YL, young leaves. Soybean genotype YC03-3 was transplanted when the first emerging compound leaf was fully unfolded and the second emerging compound leaf was not fully unfolded, and treated with high and low P (HP, 250 μM KH2PO4; LP, 5 μM KH2PO4) for 40 days (40D). Results are means ± SE from 3 independent experiments. Student’s t-test was used to determine the differences between control and nutrient deficiency conditions (**, P < 0.01; ***, P < 0.001).
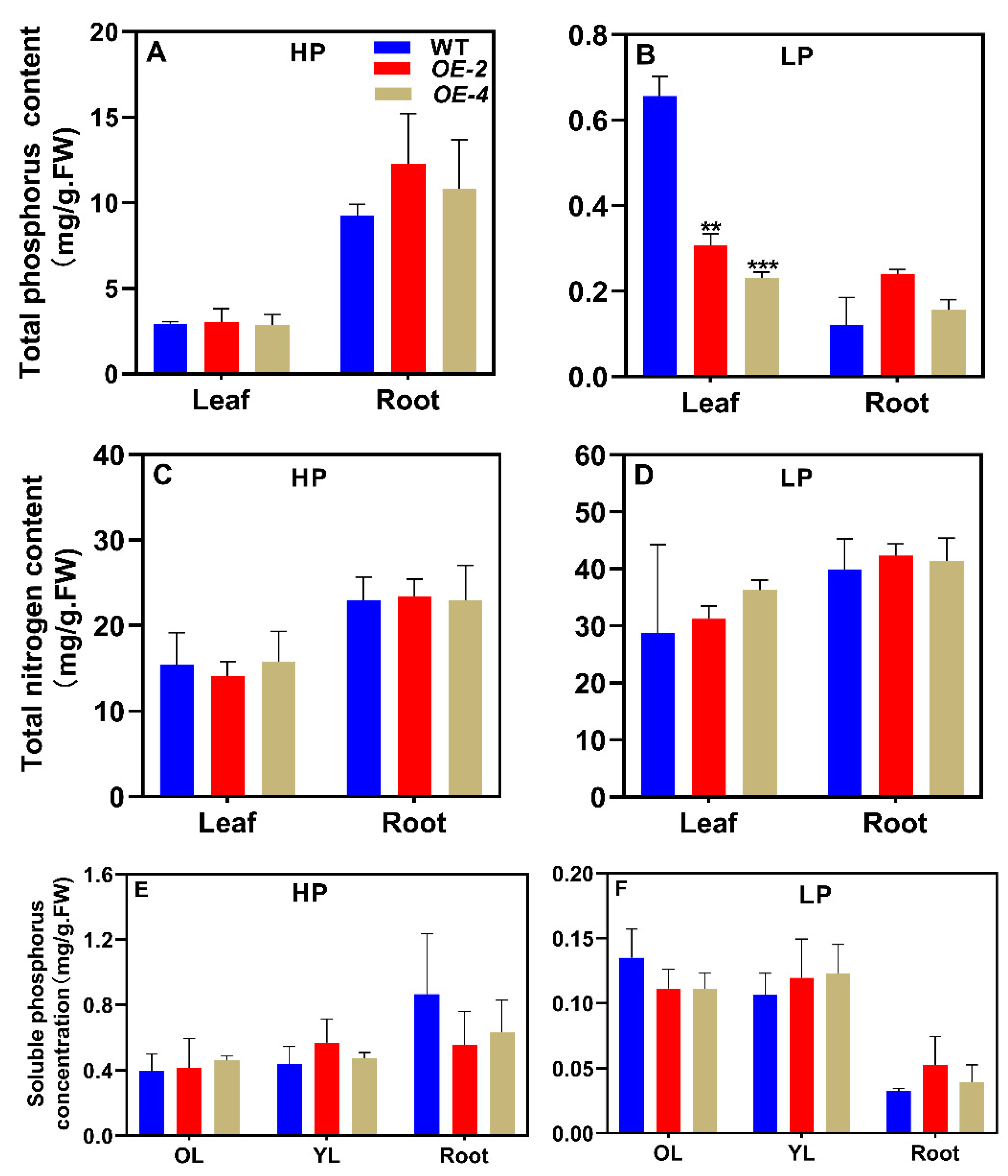

Table 1.
The MIR159 family in soybean.
| Name | Mature miRNA sequence | Position on chromosome |
|---|---|---|
| MIR159a | GAGCUCCUUGAAGUCCAAUUG | Gm09: 40266722-40266935 + |
| MIR159b | GAGUUCCCUGCACUCCAAGUC | Gm07: 5424789-5424974 − |
| MIR159c | AUUGGAGUGAAGGGAGCUCCG | Gm16: 2830034-2830218 − |
| MIR159d | AGCUGCUUAGCUAUGGAUCCC | Gm09:40267077-40267097+ |
| MIR159e | GAGCUCCUUGAAGUCCAAUU | Gm07: 9561934-9562144 − |
| MIR159f | GAGUUCCCUGCACUCCAAGUC | Gm16: 2819636-2819815 − |
Note: Gm, Glycine max; +, sense strand; −, antisense strand.
Table 2.
Analysis of elements in the MIR159 promoters in soybean.
| Low-P Responsive Elements | NaCl-Responsive Elements | ABA-Responsive Elements | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TATA -Box | TATA -Box Like | W-Box | PHR1 Element | ABRE-like | ACGT Sequence | rd22 | AtMYB2 | MYC2 | ABRE | DPBF | RY Elements |
|
| MIR159a | 4 | 1 | 4 | 1 | 1 | 1 | 2 | 1 | 1 | 1 | ||
| MIR159b | 4 | 1 | 3 | 1 | 1 | 1 | 1 | 1 | 1 | |||
| MIR159c | 4 | 1 | 4 | 1 | 1 | 1 | 2 | 2 | 1 | |||
| MIR159d | 4 | 1 | 4 | 1 | 1 | 1 | 3 | 1 | 1 | 1 | ||
| MIR159e | 4 | 4 | 1 | 1 | 1 | 4 | 3 | 1 | 1 | 1 | ||
| MIR159f | 4 | 1 | 3 | 1 | 1 | 1 | 1 | 1 | 1 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



