
Submitted:
12 June 2023
Posted:
13 June 2023
You are already at the latest version
Abstract
Objectives: Carbapenem-resistant Acinetobacter baumannii (CRAB) isolates are one of the most difficult pathogens to treat. Cefiderocol, a chlorocatechol-substituted siderophore antibiotic, was approved by the U.S. Food and Drug Administration (FDA) in 2019 indicated for the treatment of infections due to CRAB infections. Despite the initial positive treatment outcomes with this antimicrobial, recent studies reported higher than all-cause mortality rate in patients treated with cefiderocol. The cause(s) behind these outcomes remains unconfirmed. A plausible hypothesis is heteroresistance, a phenotype characterized by the survival of a small proportion of cells in a population seemingly isogenic. Recent results have shown that the addition of human fluids to CRAB cultures leads to cefiderocol heteroresistance. Here we describe molecular and phenotypic analyses of CRAB heteroresistant bacterial subpopulations to better understand the nature of the less-than-expected successful outcomes after cefiderocol treatment.
Methods: Isolation of heteroresistant variants of the CRAB strain AMA40 was carried out in cultures supplemented with cefiderocol and human pleural fluid (HPF). Two AMA40 variants, AMA40 IHC1 and IHC2, were subjected to whole genome sequencing and transcriptional analysis to identify mutations and expression changes associated with cefiderocol heteroresistance. The impact of these mutations on the pharmacodynamic activity of cefiderocol was assessed via susceptibility testing, EDTA and boronic acid inhibition analysis, biofilm formation, and static time-kill assays.
Results: Heteroresistant variants AMA40 IHC1 and AMA40 IHC2 have 53 chromosomal mutations, of which 40 are common to both strains. None of the mutations occurred in genes associated with high affinity iron-uptake systems or β-lactam resistance. However, transcriptional analyses showed significant modifications in levels of expression of genes associated with these functions. The blaNDM-1 and blaADC-2, as well as various iron-uptake system genes, were expressed at higher levels than the parental strain. On the other hand, the carO and ompA genes’ expression was reduced. One of the mutations common to both heteroresistant strains mapped within pipA, a gene associated with iron homeostasis in other species. Static time-kill assays showed that supplementing cation-adjusted Mueller-Hinton broth with human serum albumin, the main protein component of HPF, considerably reduced cefiderocol killing activity for all three strains tested. Notably, collateral resistance to amikacin was observed in both variants.
Conclusions: We conclude that exposing CRAB to fluids containing high HSA facilitates the rise of heteroresistance associated with point mutations and upregulation of genes coding for β-lactamases and biofilm formation.
Keywords:
Acinetobacter baumannii
; cefiderocol
; human pleural fluid
; NDM-1
; carbapenem-resistance
Introduction
Acinetobacter baumannii is an opportunistic Gram-negative bacillus that is primarily responsible for causing infections among critically ill patients that may be immunocompromised [1]. The two principal clinical manifestations are pneumonia and bacteremia, followed by complicated urinary tract infections (cUTIs), meningitis, traumatic or post-surgical wound infections, and osteomyelitis [2]. Carbapenem-resistant A. baumannii (CRAB) was recently classified as a critical priority pathogen by the World Health Organization and the Centers for Disease Control and Prevention (CDC) as infections due to this pathogen are challenging to treat given the lack of viable treatment options [3,4]. Additionally, the global emergence and spread of highly resistant A. baumannii highlights the need for new antimicrobial therapies [4]. Despite efforts by several research groups and pharmaceutical companies over the past decade [5-7], the only new novel drug approved by the U.S. Food and Drug Administration (FDA) active against A. baumannii is cefiderocol(https://www.accessdata.fda.gov/drugsatfda_docs/label/2019/209445s000lbl.pdf). Guidance documents from various American and European scientific societies recommend cefiderocol for treating CRAB infections. However, these recommendations are based on in vitro results and only limited clinical trials. Although positive outcomes abound, there are recent reports indicating decreased cefiderocol efficacy against multidrug resistant (MDR) CRAB [8-11] suggestive of increasing cefiderocol resistance [12-14].
Heteroresistance is a phenotype wherein a small fraction of bacteria within a bacterial community develop resistance under antibiotic pressure [15,16]. Heteroresistance can lead to consequential resistance since the resistant subpopulation expands following prolonged antibiotic exposure. Heteroresistance to cefiderocol has been observed among different carbapenem-resistant Gram-negative species [17,18]. CREDIBLE-CR, a randomized, open-label, multicenter phase 3 clinical trial was conducted to evaluate the safety and efficacy of cefiderocol for the treatment of nosocomial pneumonia, blood stream infection, sepsis, or complicated urinary tract infection due to carbapenem-resistant Gram-negative pathogens. [19] Among the 118 patients in the intent-to-treat population, 54 patients were infected with A. baumannii i.e., the most frequent carbapenem-resistant pathogen. Prior randomized trials that included A. baumannii most often focused on colistin based regimens. [8] The all-cause mortality in the cefiderocol group compared to the best available therapy was higher (19/39) particularly in patients with nosocomial pneumonia or bloodstream infection or sepsis with Acinetobacter spp at baseline [19]. Results from more recent studies also reported cefiderocol heteroresistance when A. baumannii was cultured in the presence of human serum albumin (HSA) or human pleural fluids (HPF) [20]. These human fluid components induced modifications in expression levels of genes related to high-affinity iron uptake systems and resistance to β-lactams [21-26]. This is supported by evidence that showed that most of the strains that exhibited heteroresistance, harbored the gene blaPER-7 [27,28]. Choby et al. observed a correlation between amplification of Enterobacterales and A. baumannii ESBLs genes and consequently heteroresistance to cefiderocol [29]. Higher resistance levels were also observed in NDM-producing Enterobacterales isolates and in at least one case increased blaNDM-5 expression was correlated with increased cefiderocol resistance [30,31].
As mentioned before, the addition of human fluids to CRAB cultures can lead to CFDC heteroresistance. In this work, with the aim of gaining a better understanding of the underlying reason for this phenomenon, we carried out molecular and phenotypic analyses of two selected CRAB heteroresistant bacterial subpopulations obtained after exposure to HPF. We observed that the selected heteroresistant variants of the clinical isolate CRAB AMA40 acquired chromosomal mutations that impacted genes coding for numerous functions, one of which could be related to iron metabolism, and produced significant changes in gene expression. Genes coding for β-lactamases, high-affinity iron uptake systems, and functions related to biofilm formation were expressed at higher levels in the heteroresistant variants compared to the AMA40 parental strain. In addition, a decrease in the transcripts of genes coding for outer membrane proteins was observed in the selected mutant variants.
Result
Comparative Whole Genome Sequence Analysis of AMA40 and the IHC1 and IHC2 Heteroresistant Derivatives
The A. baumannii CRAB model strain, AMA40, susceptible to cefiderocol (MIC of 0.5-1 mg/L) harbors the carbapenem resistance gene blaNDM-1 and other relevant β-lactamase coding genes such as blaADC-2 [27,32,33]. However, cefiderocol MIC for AMA40 exposed to HPF were higher by 5 doubling dilutions (> 128 mg/L) [20]. This is indicative of the emergence of cefiderocol resistant colonies within the inhibition ellipse (intracolonies), a response that indicates the presence of heteroresistant derivatives. To better understand some of the factors responsible for this phenomenon, the AMA40 IHC1 and IHC2 isolated strains were subjected to a global genomic comparative analysis.
A total of 53 mutations were identified in the AMA40 IHC1 and IHC2 strains compared to the parental strain. Among these mutations, 39 were seen in both variants,16 of these occurred within intergenic regions. All five mutations unique to AMA40 IHC1 and four out of the nine found in AMA40 IHC2 were intergenic (Table S1). Among the intragenic mutations common to both variants, 1, 24 and 14 were nucleotide insertions, substitutions, or deletions, respectively. The analysis of the nucleotide substitutions revealed nine synonymous and 12 non-synonymous mutations (Table S1). Eleven mutations affected genes coding for hypothetical proteins. In contrast, the rest of the mutations occurred within genes associated with known functions, such as aidA (quorum-quenching), lptA and lptG (outer membrane synthesis), cas3 (CRISPR-associated nuclease/helicase), and others (Table S1). The gene content of AMA40, AMA40 IHC1, and AMA40 IHC2 were identical.
It is well-known that mutations in genes coding for active iron-uptake systems play an important role in cefiderocol resistance. However, the comparative genomic analysis of the AMA40 parental strain and the AMA40 IHC1 and IHC2 heteroresistant derivatives showed no nucleotide changes in genes coding for high-affinity iron acquisition functions, including piuA, fur, tonB3.1, tonB3.2, tonB3.3, pirA, entAB, bauA, and bfnH, among others. Interestingly, the same non-synonymous mutation (S157A) within the ppiA gene of both heteroresistant derivatives was observed, suggesting a link between lack or deficient PpiA function and cefiderocol resistance. Previous work reported a potential correlation between ppiA, which encodes a peptidyl-prolyl cis/trans isomerase (PPIase), and iron uptake regulation [34]. Although the role of PpiA in Acinetobacter remains to be understood, our results indicate that this protein is involved in the decreased cefiderocol susceptibility reported previously [35]. NDM duplication or over-expression has been associated with decreased cefiderocol susceptibility [36,37]. However, that is not the case here as both AMA40 IHC1 and IHC2 variants and the parent strain had an identical single copy of blaNDM-1 present.
Comparative Transcriptional Analysis of AMA40 and the IHC1 and IHC2 Heteroresistant Derivatives
Quantitative RT-PCR (qRT-PCR) analysis showed that the expression of the β-lactamase genes blaNDM-1 and blaADC-2 was significantly increased by 3.5- and 3-fold, respectively, in both heteroresistant strains (AMA40 IHC1 and IHC2) with respect to the parental strain (Figure 1A). Conversely, the porin coding genes carO and ompA were down regulated in the AMA40 IHC1 and IHC2 variants (Figure 1A). Assessment of the expression levels of the iron uptake genes bauA, pirA, piuA, bfnH, exbD, and tonb3 showed that all but bauA were expressed at significantly elevated levels in the AMA40 IHC1 and IHC2 derivatives (Figure 1B).
An increased expression of two β-lactamase genes and a decrease in the ability to penetrate the outer membrane could be key factors contributing to the increased cefiderocol resistance levels expressed by the IHC1 and IHC2 variants. While increased expression of high-affinity iron uptake systems is expected to increase susceptibility to cefiderocol, the balance between increased expression of genes coding for β-lactamases and high affinity iron uptake systems and decreased expression of porin coding genes seems to contribute to increased cefiderocol resistance seen in IHC1 and IHC2 compared to the parental strain.
Susceptibility Assays Suggest the Contribution of Increased Gene Expression of β-Lactamases Resulting in Increased Levels of Cefiderocol Resistance
Intracolonies observed in the inhibition ellipse while determining cefiderocol MIC of AMA40 when exposed to HPF were subcultured and stored at -80°C for further analyses [20]. MIC of cefiderocol determinations using two different methodologies, E-strips, and microdilution assays, showed that both strains have a higher than the parental strain level of resistance (Figure S1 and Table 1). Furthermore, the enhanced resistance phenotype is not lost after subculturing, suggesting that it is a stable trait. Additional susceptibility testing of IHC1 and IHC2 to other antibiotics (meropenem, imipenem, gentamicin, ampicillin/sulbactam, amikacin, ciprofloxacin, levofloxacin, tigecycline, colistin, and trimethoprim-sulfamethoxazole) was performed to determine potential cross-resistance to cefiderocol. As expected, the strains were highly resistant to most of the antibiotic families evaluated (Table 1). However, only a 2- to 3-fold increase in MICs for colistin and amikacin, respectively, was observed in both heteroresistant variants with respect to the parental strain (Table 1). In addition, a 4-fold increase in levofloxacin MIC was seen only for AMA40 IHC2 (Table 1). In addition, MICs of cefiderocol in combination with β-lactamase inhibitors like avibactam, relevactam, or zidebactam were reduced. These results demonstrated the ability of β-lactamase inhibitors in restoring susceptibility to cefiderocol to levels similar to those displayed by the parental strain (Figure S2 and Table S2). Additionally, zinc supplementation was performed to evaluate if the addition of this metal could result in improved cefiderocol susceptibility as NDM-1 activity depends on it [39]. A 3-fold increase in cefiderocol MIC was observed in the wild-type strain when CAMHA was supplemented with 300 mg/L of ZnSO4. However, these changes were not observed with IHC1 and IHC2, where NDM-1 was already overexpressed based on our transcriptional analysis (Table S3).
EDTA and boronic acid assays were performed to evaluate the contribution of metallo-β-lactamases (blaNDM-1) or class C β-lactamases (blaADC-2) towards reducing cefiderocol susceptibility. In both assays, no significant changes were observed for the AMA40 parental strain, however a slight increase in the halo was observed for the heteroresistant strains (Figure S3A and B).
In sum, these results showed an increased resistance to colistin and amikacin in both mutant strains, while increased resistance to levofloxacin was only seen in IHC2. In addition, the role of blaNDM-1 and blaADC-2 in the increase cefiderocol resistance was supported by transcriptional analysis as well as phenotypic assays.
Static Time-Kill Studies Demonstrated Reduced Cefiderocol Killing Activity in the Presence of HSA
Previous studies have shown that the presence of HSA can lead to an increase in cefiderocol MICs [20]. Static time-kill studies were conducted in the presence and absence of a physiologically relevant concentration of 3.5 % HSA [38,39]. In the absence of cefiderocol, all three A. baumannii strains showed a similar growth profile in CAMHB reaching ~8.8-8.9 Log10 CFU/mL by 8 h (Figure 2A-C). In HSA-supplemented CAMHB, the bacterial growth for all three strains by 8 h was between 7.2 and 8.2 Log10 CFU/mL, slightly lower compared to that observed in the absence of HSA (Figure 2D-F). In the absence of HSA, cefiderocol concentrations >1 mg/mL against both AMA40 and AMA40 IHC2, resulted in >2 Log10 CFU/mL reduction in bacterial burden (Figure 2A, 2B and Figure 3A-B) at 8 h. In the presence of 3.5% HSA, cefiderocol activity was considerably reduced against both strains (Figure 4). The addition of 3.5% HSA at 8 h resulted in a 1.1 Log10 CFU/mL with cefiderocol 8 mg/mL against AMA40 alone and none of the cefiderocol concentrations showed any effect against AMA40 IHC2. Notably, none of the cefiderocol concentrations tested were effective against IHC1; the killing activity observed with this derivative was similar to the growth controls either in the presence (Figure 2C) or absence (Figure 2F) of HSA.
Increased Biofilm Formation by the Cefiderocol Heteroresistant CRAB Cells
Biofilms are responsible for persistence of bacterial infections associated with foreign bodies like catheters or prothesis [40]. Hence, determining if there is a change in the ability to form biofilms with heteroresistant AMA40 IHC1 and IHC2 derivatives provides information about their pathogenicity. Both strains produced a significantly greater mass of biofilm than the parental AMA40 strain based on quantification of biofilm production [41] (Figure 4A).
To determine if changes at the biofilm formation phenotype level were correlated with modifications at the transcriptional level, qRT-PCR analysis of biofilm-related genes was carried out. The expression of the genes encoding for the CsuAB fimbrial major subunit and the CsuE component [42,43] as wells as the BfmR response regulator was increased by 2- to 3-fold in the IHC1 and IHC2 when compared with the AMA40 wild-type strain (Figure 4B). The transcriptional expression of csuB and pilT genes was also increase in the IHC1 and IHC2 strain but not significantly.
In sum, these results showed that the heteroresistant cells exhibit an increase biofilm formation with a concomitant increased expression of genes associated with biofilm production. The increased biofilm formation can be contributing to the increased cefiderocol resistance observed in both AMA40 IHC1 and IHC2. An additional interesting observation is the increased expression of bfmR. The two-component regulatory system BfmRS is known to have a role in determining a variety of A. baumannii responses including protection against β-lactam antibiotics [44-46]. This brings up the question if BfmRS can be playing a role in cefiderocol heteroresistance and/or increase in MICs.
Discussion
Cefiderocol has shown to be a promising new option for hard-to-treat infections caused by carbapenem-resistant Gram-negative bacilli, including A. baumannii. However, there have been increasing reports of cefiderocol resistance [8,9,13,14]. In the present study, we studied the emergent heteroresistance AMA40 CRAB cells observed after exposure to HSA-containing human fluids. The genomic, transcriptional, and phenotypic analysis of the two randomly selected isogenic variants indicated that multiple factors may be responsible for the cefiderocol resistance phenotype of IHC1 and IHC2 derivatives, including genomic mutations, increased expression of β-lactamases, reduced expression of porins, and increased biofilm formation. The ppiA mutation is an interesting observation that may be related to the increased cefiderocol resistance of the aforementioned AMA40 derivatives. In Mycobacterium tuberculosis, PpiA is upregulated during heat shock implying that it may be related to stress responses and possibly virulence [47]. The M. tuberculosis ppiA gene was also downregulated during iron depletion, suggesting that its expression could be iron regulated [48]. In other studies, PPIases demonstrated a pivotal role in catalyzing the correct folding of many prokaryotic and eukaryotic proteins involved in diverse biological functions, ranging from cell cycle regulation to bacterial infection [49]. However, in A. baumannii its role in iron homeostasis, antibiotic resistance and virulence has not yet been studied.
It has been reported that one factor that can contribute to cefiderocol r esistance is the increased expression of β-lactamases. Simner et al. [36] reported a case of a transplant recipient infected with an E. coli isolate harboring a blaNDM-5 gene, which progressively lost susceptibility to cefiderocol following treatment. The analysis of different isolates recovered during the course of antibiotic treatment showed an increase in the copy number and expression of blaNDM-5 [36]. A previously reported case of a male patient in his 50s whose initial blood cultures had revealed a susceptible K. pneumoniae, which became resistant to cefiderocol upon completing cefiderocol therapy, provides further evidence about the role of this gene in cefiderocol resistance. The sequencing of this K. pneumoniae isolate identified blaNDM-5, suggesting that the presence of NDM can be implicated in the development of cefiderocol resistance [15]. In addition, Choby et al. [17] , observed the amplification of the ESBLs genes in Enterobacterales and A. baumannii and the consequently development of heteroresistance to cefiderocol. This outcome supports our observation with the AMA40 IHC1 and IHC2 strains identifying blaNDM-1 and blaADC as potential contributors to heteroresistance development [29].
Additional factors that could play a role in the increased resistance observed in the AMA40 heteroresistance colonies include the down-regulation of the porin coding genes carO and ompA. CarO allows the permeation of imipenem in A. baumannii [50], while the lack of a functional OmpA is associated with increased susceptibility to different antibiotics such as chloramphenicol, colistin, aztreonam, imipenem, gentamicin and nalidixic acid in this pathogen [51]. Another factor that needs to be considered is the increased expression of biofilm associated genes with the concomitant increase in biofilms formation in both heteroresistant strains. We also observed an increase in the expression of bfmR. There is significant published literature describing the role of the BfmRS two-component system, controlling various A. baumannii cellular processes, including biofilm formation [43,52]. Previous studies have also shown that hyperactive alleles of BfmRS conferred increased resistance and tolerance against an expansive set of antibiotics, including dramatic protection from β-lactam activity [44,46,52,53]. The increased expression of bfmR observed in the heteroresistant cells could be responsible for the increase in colistin and amikacin MICs as reported [52]. Given its role in developing heteroresistance to cefiderocol, further mechanistic studies characterizing the role BfmRS play in cefiderocol resistance are necessary.
Recently, unstable A. baumannii heteroresistance subpopulations were found in 8/10 samples cultured in the presence of high cefiderocol concentrations. Genomic analyses of heteroresistant isolates revealed the presence of PBP3 and TonB3 mutations that were shared by all strains regardless of their resistance phenotype [18]. In contrast, the resistance traits of the AMA40 IHC1 and IHC2 derivatives isolated during our work, which represent subpopulations obtained after the exposure of AMA40 to HSA-containing fluids, were maintained in a stable manner, even in the absence of cefiderocol selection pressure. Furthermore, the genomic analysis of the AMA40 IHC1 and IHC2 derivatives did not reveal a direct and clear connection to the functional expression of high-affinity iron acquisition processes. Taken together, these observations suggests that a combination of different cellular mechanisms are involved in driving the emergence of stable cefiderocol heteroresistance in processes that is affected by the presence of host fluids containing HSA.
Fortunately, several authors reported that the combination of cefiderocol and a diazabicyclooctane (DBO) derivative, like avibactam, relebactam or zidebactam, seems to restore the antibacterial activity of cefiderocol against CRAB, at concentrations that are several times lower than its cefiderocol MIC and limits, in some cases, the emergence of resistance [18,54]. Subpopulations with moderate to high level of resistance to cefiderocol described in this work, recovered susceptibility to cefiderocol regardless its combination with DBO. The mechanism of this synergistic activity of cefiderocol in combination with DBO is not understood especially given that multiple factors are responsible for the emergence of cefiderocol resistant subpopulations. In our work, we observed that even in the case where hyperproduction of β-lactamases that are not inhibited or are unresponsive to DBOs, such as blaNDM and blaADC, the susceptibility to cefiderocol is restored. These results further support the concept that combinatorial therapy is a good option to restore cefiderocol susceptibility while preventing the emergence of heteroresistance or resistant intra-colonies.
The antimicrobial failure and the development of resistance by CRAB and other microbial pathogens was raised during studies that evaluated the efficacy of cefiderocol activity [19,55]. Falcone et al. observed that among patients who experienced medical failure following cefiderocol monotherapy treatment, all had bloodstream infections (30% of Blood Stream Infections patients) [55]. In the presence of HSA, the main serum protein, the killing activity of cefiderocol was reduced against both, susceptible and low-level resistant strains as observed in an in vitro model [32]. Although a reduction in the free fraction of cefiderocol available is expected due to its strong binding to HSA (ca 60%;) [56], the antibiotic concentrations tested by far exceeded the MIC of the parental strain.
In a real scenario, a significant benefit of cefiderocol treatment in patients with CRAB infections was noticed, except in VAP patients [55]. We previously demonstrated that HSA as well HPF modulates the expression of genes associated with iron uptake systems and antibiotic resistance [20,25,57-59].
Concluding Remarks
In the present work, we found that two independent cefiderocol heteroresistant derivatives showed no mutations in genes coding for active iron acquisition or β-lactam resistance functions. However, both derivatives showed the same point mutation in pipA, a gene associated with iron homeostasis in other species. In addition, the blaNDM-1 and blaADC-2, genes were expressed at higher levels in the cefiderocol heteroresistant cells that were associated with a decreased cefiderocol susceptibility. Notably, static time kill assays showed that the cefiderocol killing activity was considerably reduced in the presence of HSA. In sum, HSA-containing fluids result in reduced susceptibility to cefiderocol through mechanism(s) that might include genomic point mutations or phenotypic modifications like increased biofilm formation or changes in gene expression. Further studies focused on understanding the mechanisms through which HSA-rich human fluids elicit antibiotic resistance may provide the basis for designing more effective strategies for treating A. baumannii infections.
Materials and Methods
Bacterial Strains
The carbapenem-resistant clinical A. baumannii AMA40 (blaNDM-1, blaADC-2, and blaOXA-51) strain [27,60] was used in this study. The AMA40 IHC1 and IHC2 cefiderocol heteroresistant strains, which were recovered within the inhibition ellipse zones after exposure of the A. baumannii AMA40 parental strain to HPF, were included in the analysis. Stocks of the IHC1 and IHC2 isolates were kept at -80°C to determine the stability of the increased cefiderocol resistance of these isolates (Figure S1).
Whole Genome Sequencing and Genomic Analysis
Genomic DNA was extracted using the Wizard Promega kit (Promega, Madison, USA) according to manufacturer instructions. The whole genome sequencing was outsourced to SEQCENTER sequencing service (Pittsburgh, PA) and performed using NextSeq 550 Illumina technology. The sequence quality was checked by FASTQ software analysis (https://www.bioinformatics.babraham.ac.uk/projects/fastqc/), and de novo sequence assembly was performed with SPAdes [61] followed by a quality assessment performed with QUAST [62]. Genome annotation was done using PROKKA [63]. Variant calling was performed using the breseq and gdtools software packages [64]. Genes coding for high-affinity iron-uptake systems were identified using the sequences reported by Antunes et al. [65]. The analyses of gain and/or loss of genes were performed using Roary (software version 3.11.2) [66]. The gene expression profiles for the AMA40 wild type strain and the IHC1 and IHC2 derivatives have been deposited in the ArrayExpress database (accession number: E-MTAB-12444).
Transcriptional Analysis Using Quantitative RT-PCR
Overnight cultures of AMA40, IHC1 and IHC2 were diluted 1:10 in iron depleted cation adjusted Mueller Hinton broth (CAMHB) and incubated with agitation for 18 h at 37°C. RNA was extracted from each sample using the Direct-zol RNA Kit (Zymo Research, Irvine, CA, USA) following manufacturer’s instructions. Total RNA extractions were performed using three independent biological replicates for each condition. The extracted DNase-treated RNA was used to synthesize cDNA using the manufacturer’s protocol provided with the iScriptTM Reverse Transcription Supermix for qPCR reagents (Bio-Rad, Hercules, CA, USA). The cDNA concentrations were adjusted to 50 ng/μL and qPCR was conducted using the qPCRBIO SyGreen Blue Mix Lo-ROX following the manufacturer’s protocol (PCR Biosystems,Wayne, PA, USA). The transcriptional analysis of blaADC, blaNDM-1, ompA, carO, pirA, piuA, bauA, bfnH, exbD, tonB3, csuAB, csuB, csuE, bfmR and pilT was done using specific primers (Table S4). At least three independent cDNA replicates were tested in triplicate using the CFX96 TouchTM Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA). Data are presented as NRQ (normalized relative quantities) calculated using the qBASE method [67], with recA and rpoB genes as normalizers [68]. Differences were determined by ANOVA followed by Tukey’s multiple comparison test (P < 0.05) using GraphPad Prism (GraphPad software, San Diego, CA, USA).
Susceptibility Assays
Antibiotic susceptibility assays were performed following the Clinical and Laboratory Standards Institute (CLSI) guidelines [69]. After OD600 adjustment, 100 μL of A. baumannii AMA40, IHC1 and IHC2 cells grown in iron depleted CAMHA were used to perform susceptibility assays. Antimicrobial commercial E-strips (Liofilchem S.r.l., Roseto degli Abruzzi, Italy) for cefiderocol and amikacin were used. In addition, 4 μg/mL of avibactam (Sigma-Aldrich), relevactam (Sigma-Aldrich), or zidebactam (Wockhardt) were added to CAMHA medium when indicated. The plates were incubated at 37ºC for 18 h. CLSI breakpoints were used for data interpretation [69]. Escherichia coli ATCC 25922 was used for quality control purposes. In addition, cefiderocol MICs of AMA40, IHC1 and IHC2 were also performed using microdilution method following CLSI guidelines.
EDTA and Boronic Acid Inhibition Assays
To determine the impact of NDM inhibition on cefiderocol susceptibility, cefiderocol disk diffusion assays with and without EDTA we performed. For this purpose, two 30-μg cefiderocol disks, one supplemented with 10 μL of 0.5 mmol/L EDTA (Sigma-Aldrich), were deposited on the surface of a Mueller-Hinton agar plate inoculated with a lawn of AMA40, AMA40 IHC1, or AMA40 IHC2 cells. The cells were incubated for 18–24 h at 35°C ± 2°C [70]. An increase of the growth inhibition zone >3 mm produced by the addition of EDTA was interpreted as circumstantial evidence that NDM production was contributing to cefiderocol resistance. In addition, to evaluate blaADC (class C β-lactamases) contribution to cefiderocol susceptibility, CAMHA plates containing 300 μg/mL boronic acid (final concentration) were prepared following previously published recommendations [71]. Subsequently, a 30-ug cefiderocol disk was placed on the surface of a CAMHA plate inoculated with a lawn of AMA40, AMA40 IHC1, or AMA40 IHC2 cells and incubated for 18-24 h at 35°C ± 2°C. An increase of the growth inhibition zone >3 mm produced by the addition of boronic acid was interpreted as circumstantial evidence that ADC production was contributing to cefiderocol resistance.
Static Time-Kill Studies
Static time-kill studies were performed to determine bacterial killing kinetics in the absence (growth control) and presence of cefiderocol against AMA40, AMA40 IHC1 and AMA-40 IHC2 strains. Cefiderocol killing activity was evaluated at clinically achievable concentrations (0.5, 1, 4 and 8 µg/mL) [56] with and without 3.5% HSA against an initial inoculum of 5 x 106 CFU/mL. Cefiderocol was added to a log growth phase bacterial suspension. Serial samples obtained at 0, 2, 4, 6, and 8 h following addition of the drug were diluted with normal saline and 50 μL of the appropriate bacterial dilution were spirally plated on CAMHA using an automated spiral plater (Don Whitley WASP Touch, Microbiology International, Frederick, MD) and incubated at 37°C. Following a 24-h incubation period, bacteria were quantified using a ProtoCOL automated colony counter (Symbiosis, Cambridge, United Kingdom). The lower limit of quantification was 1.3 log10 CFU/mL.
Biofilm Assays
Overnight cultures of AMA40, AMA40 IHC1, and AMA40 IHC2 cells grown in fresh LB medium with agitation for 18 h at 37 ºC were used to determine biofilm formation. The optical density at 600 nm (OD600) was adjusted to 0.9-1.1 and placed in a 96-well polystyrene microtiter plate and incubated at 37°C for 24 h without shaking. The next day, the OD600 was measured, using a microplate reader, to determine the total biomass. The wells were emptied, washed three times with 1X phosphate buffered saline (PBS) and stained for 15 min with 1% crystal violet (CV) for 15 min. Excess CV was removed and the CV associated with biofilms was solubilized in ethanol-acetone (80:20) for 30 min. OD580 was measured and the ratio of biofilm to total biomass was determined. Experiments were performed in triplicate, with at least three technical replicates per biological replicate. Statistical significance (P < 0.05) was determined by two-way ANOVA followed by Tukey's multiple comparison test.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
V.M., B.N., J.E., M.R.T., T.S., F.P., G.M.I., R.S., Q.V., R.A.B, M.E.T., G.R. and M.S.R. conceived the study and designed the experiments. B.N., J.E., M.R.T., T.S., V.M., I.M., G.M.I., R.S., Q.V., and M.S.R. performed the experiments and genomics and bioinformatics analyses. G.M.I., M.R.T., T.S., F.P., L.A.A., R.A.B, M.E.T. G.R., and M.S.R. analyzed the data and interpreted the results. R.A.B., M.E.T., G.R., and M.S.R. contributed reagents/materials/analysis tools. G.M.T., M.R.T., T.S., F.P., R.S., R.A.B, LAA, M.E.T., G.R., and M.S.R. wrote and revised the manuscript. All authors read and approved the final manuscript.
Funding
The authors’ work was supported by NIH SC3GM125556 to M.S.R., R01AI100560, R01AI063517, R01AI072219 to R.A.B., and 2R15 AI047115 to M.E.T. This study was supported in part by funds and/or facilities provided by the Cleveland Department of Veterans Affairs, Award Number 1I01BX001974 to RAB from the Biomedical Laboratory Research & Development Service of the VA Office of Research and Development and the Geriatric Research Education and Clinical Center VISN 10 to R.A.B. J.E was supported by grant MHRT 2T37MD001368 from the National Institute on Minority Health and Health Disparities, National Institute of Health. I.M. was supported by Project RAISE, U.S. Department of Education HSI-STEM, award number P031C160152. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health or the Department of Veterans Affairs. M.R.T and T.S are recipient of a postdoctoral fellowship from CONICET.
Acknowledgments
The authors wish to acknowledge the generous support of Wockhardt in supplying Zidebactam for this study.
Conflicts of Interest
The. authors declare no conflict of interest.
References
- Piperaki, E.T.; Tzouvelekis, L.S.; Miriagou, V.; Daikos, G.L. Carbapenem-resistant Acinetobacter baumannii: in pursuit of an effective treatment. Clin Microbiol Infect 2019, 25, 951–957. [Google Scholar] [CrossRef]
- Peleg, A.Y.; Seifert, H.; Paterson, D.L. Acinetobacter baumannii: emergence of a successful pathogen. Clin Microbiol Rev 2008, 21, 538–582. [Google Scholar] [CrossRef]
- (WHO)., W.H.O. (WHO)., W.H.O. Global Priority List of Antibiotic-resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. 2017.
- 4. CDC. Antibiotic Resistance Threats in the United States. Atlanta, GA: U.S. Department of Health and Human Services, CDC; 2019.
- Abdul-Mutakabbir, J.C.; Nguyen, L.; Maassen, P.T.; Stamper, K.C.; Kebriaei, R.; Kaye, K.S.; Castanheira, M.; Rybak, M.J. In Vitro Antibacterial Activity of Cefiderocol against Multidrug-Resistant Acinetobacter baumannii. Antimicrob Agents Chemother 2021, 65, e0264620. [Google Scholar] [CrossRef]
- He, S.; He, H.; Chen, Y.; Chen, Y.; Wang, W.; Yu, D. In vitro and in vivo analysis of antimicrobial agents alone and in combination against multi-drug resistant Acinetobacter baumannii. Front Microbiol 2015, 6, 507. [Google Scholar] [CrossRef]
- Theuretzbacher, U.; Bush, K.; Harbarth, S.; Paul, M.; Rex, J.H.; Tacconelli, E.; Thwaites, G.E. Critical analysis of antibacterial agents in clinical development. Nature reviews. Microbiology 2020, 18, 286–298. [Google Scholar] [CrossRef] [PubMed]
- Choby, J.E.; Ozturk, T.; Satola, S.W.; Jacob, J.T.; Weiss, D.S. Widespread cefiderocol heteroresistance in carbapenem-resistant Gram-negative pathogens. Lancet Infect Dis 2021, 21, 597–598. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Sadek, M.; Nordmann, P. Contribution of PER-type and NDM-type ss-lactamases to cefiderocol resistance in Acinetobacter baumannii. Antimicrob Agents Chemother, 1128; -21. [Google Scholar] [CrossRef]
- Karakonstantis, S.; Rousaki, M.; Kritsotakis, E.I. Cefiderocol: Systematic Review of Mechanisms of Resistance, Heteroresistance and In Vivo Emergence of Resistance. Antibiotics (Basel) 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Streling, A.P.; Al Obaidi, M.M.; Lainhart, W.D.; Zangeneh, T.; Khan, A.; Dinh, A.Q.; Hanson, B.; Arias, C.A.; Miller, W.R. Evolution of Cefiderocol Non-Susceptibility in Pseudomonas aeruginosa in a Patient Without Previous Exposure to the Antibiotic. Clin Infect Dis, 1093. [Google Scholar] [CrossRef]
- Tamma, P.D.; Aitken, S.L.; Bonomo, R.A.; Mathers, A.J.; van Duin, D.; Clancy, C.J. Infectious Diseases Society of America Guidance on the Treatment of Extended-Spectrum beta-lactamase Producing Enterobacterales (ESBL-E), Carbapenem-Resistant Enterobacterales (CRE), and Pseudomonas aeruginosa with Difficult-to-Treat Resistance (DTR-P. aeruginosa). Clin Infect Dis 2021, 72, 1109–1116. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.; Boutin, S.; Kocer, K.; Fiedler, M.O.; Störzinger, D.; Weigand, M.A.; Tan, B.; Richter, D.; Rupp, C.; Mieth, M.; et al. Rapid development of cefiderocol resistance in carbapenem-resistant Enterobacter cloacae during therapy is associated with heterogeneous mutations in the catecholate siderophore receptor cira. Clin Infect Dis, 1093. [Google Scholar] [CrossRef]
- Simner, P.J.; Beisken, S.; Bergman, Y.; Ante, M.; Posch, A.E.; Tamma, P.D. Defining Baseline Mechanisms of Cefiderocol Resistance in the Enterobacterales. Microb Drug Resist, 1089. [Google Scholar] [CrossRef]
- Witt, L.S.; Steed, D.B.; Burd, E.M.; Ozturk, T.; Davis, M.H.; Satola, S.W.; Weiss, D.S.; Jacob, J.T. Bacteraemia with an MBL-producing Klebsiella pneumoniae: treatment and the potential role of cefiderocol heteroresistance. J Antimicrob Chemother 2022, 77, 2569–2571. [Google Scholar] [CrossRef]
- Andersson, D.I.; Nicoloff, H.; Hjort, K. Mechanisms and clinical relevance of bacterial heteroresistance. Nature reviews. Microbiology 2019, 17, 479–496. [Google Scholar] [CrossRef]
- Choby, J.E.; Ozturk, T.; Satola, S.W.; Jacob, J.T.; Weiss, D.S. Does cefiderocol heteroresistance explain the discrepancy between the APEKS-NP and CREDIBLE-CR clinical trial results? Lancet Microbe 2021, 2, e648–e649. [Google Scholar] [CrossRef]
- Stracquadanio, S.; Bonomo, C.; Marino, A.; Bongiorno, D.; Privitera, G.F.; Bivona, D.A.; Mirabile, A.; Bonacci, P.G.; Stefani, S. Acinetobacter baumannii and Cefiderocol, between Cidality and Adaptability. Microbiol Spectr, 1128; -22. [Google Scholar] [CrossRef]
- Bassetti, M.; Echols, R.; Matsunaga, Y.; Ariyasu, M.; Doi, Y.; Ferrer, R.; Lodise, T.P.; Naas, T.; Niki, Y.; Paterson, D.L.; et al. Efficacy and safety of cefiderocol or best available therapy for the treatment of serious infections caused by carbapenem-resistant Gram-negative bacteria (CREDIBLE-CR): a randomised, open-label, multicentre, pathogen-focused, descriptive, phase 3 trial. Lancet Infect Dis 2021, 21, 226–240. [Google Scholar] [CrossRef]
- Le, C.; Pimentel, C.; Pasteran, F.; Tuttobene, M.R.; Subils, T.; Escalante, J.; Nishimura, B.; Arriaga, S.; Carranza, A.; Mezcord, V.; et al. Human Serum Proteins and Susceptibility of Acinetobacter baumannii to Cefiderocol: Role of Iron Transport. Biomedicines 2022, 10, 600. [Google Scholar] [CrossRef]
- Quinn, B.; Rodman, N.; Jara, E.; Fernandez, J.S.; Martinez, J.; Traglia, G.M.; Montana, S.; Cantera, V.; Place, K.; Bonomo, R.A.; et al. Human serum albumin alters specific genes that can play a role in survival and persistence in Acinetobacter baumannii. Sci Rep 2018, 8, 14741. [Google Scholar] [CrossRef]
- Le, C.; Pimentel, C.; Tuttobene, M.R.; Subils, T.; Nishimura, B.; Traglia, G.M.; Perez, F.; Papp-Wallace, K.M.; Bonomo, R.A.; Tolmasky, M.E.; et al. Interplay between meropenem and human serum albumin on expression of carbapenem resistance genes and natural competence in Acinetobacter baumannii. Antimicrob Agents Chemother, 1128; -21. [Google Scholar] [CrossRef]
- Martinez, J.; Fernandez, J.S.; Liu, C.; Hoard, A.; Mendoza, A.; Nakanouchi, J.; Rodman, N.; Courville, R.; Tuttobene, M.R.; Lopez, C.; et al. Human pleural fluid triggers global changes in the transcriptional landscape of Acinetobacter baumannii as an adaptive response to stress. Sci Rep 2019, 9, 17251. [Google Scholar] [CrossRef] [PubMed]
- Rodman Nyah, M.J. , Fung Sammie, Nakanouchi Jun, Myers Amber L., Harris Caitlin M., Dang Emily, Fernandez Jennifer S., Liu Christine, Mendoza Anthony M., Jimenez Veronica, Nikolaidis Nikolas, Brennan Catherine A., Bonomo Robert A., Sieira Rodrigo, Ramirez Maria Soledad. Human Pleural Fluid Elicits Pyruvate and Phenylalanine Metabolism in Acinetobacter baumannii to Enhance Cytotoxicity and Immune Evasion. Frontiers in Microbiology 2019, 10, 1581. [Google Scholar] [CrossRef]
- Pimentel, C.; Le, C.; Tuttobene, M.R.; Subils, T.; Martinez, J.; Sieira, R.; Papp-Wallace, K.M.; Keppetipola, N.; Bonomo, R.A.; Actis, L.A.; et al. Human Pleural Fluid and Human Serum Albumin Modulate the Behavior of a Hypervirulent and Multidrug-Resistant (MDR) Acinetobacter baumannii Representative Strain. Pathogens 2021, 10, 471. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.; Razo-Gutierrez, C.; Le, C.; Courville, R.; Pimentel, C.; Liu, C.; Fung, S.E.; Tuttobene, M.R.; Phan, K.; Vila, A.J.; et al. Cerebrospinal fluid (CSF) augments metabolism and virulence expression factors in Acinetobacter baumannii. Sci Rep 2021, 11, 4737. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.D.; Pasteran, F.; Traglia, G.M.; Martinez, J.; Huang, F.; Liu, C.; Fernandez, J.S.; Lopez, C.; Gonzalez, L.J.; Albornoz, E.; et al. Distinct mechanisms of dissemination of NDM-1 metallo- beta-lactamase in Acinetobacter spp. in Argentina. Antimicrob Agents Chemother, 1128. [Google Scholar] [CrossRef]
- Poirel, L.; Sadek, M.; Nordmann, P. Contribution of PER-Type and NDM-Type beta-Lactamases to Cefiderocol Resistance in Acinetobacter baumannii. Antimicrob Agents Chemother 2021, 65, e0087721. [Google Scholar] [CrossRef]
- Jacob, E. Choby, T.O., Carter N. Abbott, Sarah W. Satola, David S. Weiss. Heteroresistance via beta-lactamase gene amplification threatens the beta-lactam pipeline. bioRxiv 2021.05.19.444899. [CrossRef]
- Kazmierczak, K.M.; Tsuji, M.; Wise, M.G.; Hackel, M.; Yamano, Y.; Echols, R.; Sahm, D.F. In vitro activity of cefiderocol, a siderophore cephalosporin, against a recent collection of clinically relevant carbapenem-non-susceptible Gram-negative bacilli, including serine carbapenemase- and metallo-beta-lactamase-producing isolates (SIDERO-WT-2014 Study). Int J Antimicrob Agents 2019, 53, 177–184. [Google Scholar] [CrossRef]
- Dobias, J.; Dénervaud-Tendon, V.; Poirel, L.; Nordmann, P. Activity of the novel siderophore cephalosporin cefiderocol against multidrug-resistant Gram-negative pathogens. Eur J Clin Microbiol Infect Dis 2017, 36, 2319–2327. [Google Scholar] [CrossRef]
- Le, C.; Pimentel, C.; Pasteran, F.; Tuttobene, M.R.; Subils, T.; Escalante, J.; Nishimura, B.; Arriaga, S.; Carranza, A.; Mezcord, V.; et al. Human Serum Proteins and Susceptibility of Acinetobacter baumannii to Cefiderocol: Role of Iron Transport. Biomedicines 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, B.; Escalante, J.; Tuttobene, M.R.; Subils, T.; Mezcord, V.; Pimentel, C.; Georgeos, N.; Pasteran, F.; Rodriguez, C.; Sieira, R.; et al. Acinetobacter baumannii response to cefiderocol challenge in human urine. Sci Rep 2022, 12, 8763. [Google Scholar] [CrossRef] [PubMed]
- Bhaduri, A.; Misra, R.; Maji, A.; Bhetaria, P.J.; Mishra, S.; Arora, G.; Singh, L.K.; Dhasmana, N.; Dubey, N.; Virdi, J.S.; et al. Mycobacterium tuberculosis cyclophilin A uses novel signal sequence for secretion and mimics eukaryotic cyclophilins for interaction with host protein repertoire. PLoS One 2014, 9, e88090. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Li, B.; Dong, H.; Chen, M.; Yao, S.; Li, J.; Zhang, H.; Liu, X.; Wang, H.; Song, N.; et al. YdiV regulates Escherichia coli ferric uptake by manipulating the DNA-binding ability of Fur in a SlyD-dependent manner. Nucleic Acids Res 2020, 48, 9571–9588. [Google Scholar] [CrossRef]
- Simner, P.J.; Mostafa, H.H.; Bergman, Y.; Ante, M.; Tekle, T.; Adebayo, A.; Beisken, S.; Dzintars, K.; Tamma, P.D. Progressive Development of Cefiderocol Resistance in Escherichia coli During Therapy is Associated With an Increase in blaNDM-5 Copy Number and Gene Expression. Clin Infect Dis 2022, 75, 47–54. [Google Scholar] [CrossRef]
- Frohlich, C.; Sorum, V.; Tokuriki, N.; Johnsen, P.J.; Samuelsen, O. Evolution of beta-lactamase-mediated cefiderocol resistance. J Antimicrob Chemother 2022, 77, 2429–2436. [Google Scholar] [CrossRef]
- Mishra, V.; Heath, R.J. Structural and Biochemical Features of Human Serum Albumin Essential for Eukaryotic Cell Culture. International journal of molecular sciences 2021, 22. [Google Scholar] [CrossRef]
- Merlot, A.M.; Kalinowski, D.S.; Richardson, D.R. Unraveling the mysteries of serum albumin-more than just a serum protein. Front Physiol 2014, 5, 299. [Google Scholar] [CrossRef]
- Høiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic resistance of bacterial biofilms. Int J Antimicrob Agents 2010, 35, 322–332. [Google Scholar] [CrossRef]
- Quinn, B.; Traglia, G.M.; Nguyen, M.; Martinez, J.; Liu, C.; Fernandez, J.S.; Ramirez, M.S. Effect of Host Human Products on Natural Transformation in Acinetobacter baumannii. Curr Microbiol, 1007. [Google Scholar] [CrossRef]
- Tomaras, A.P.; Dorsey, C.W.; Edelmann, R.E.; Actis, L.A. Attachment to and biofilm formation on abiotic surfaces by Acinetobacter baumannii: involvement of a novel chaperone-usher pili assembly system. Microbiology (Reading, England) 2003, 149, 3473–3484. [Google Scholar] [CrossRef] [PubMed]
- Tomaras, A.P.; Flagler, M.J.; Dorsey, C.W.; Gaddy, J.A.; Actis, L.A. Characterization of a two-component regulatory system from Acinetobacter baumannii that controls biofilm formation and cellular morphology. Microbiology (Reading, England) 2008, 154, 3398–3409. [Google Scholar] [CrossRef] [PubMed]
- De Silva, P.M.; Kumar, A. Signal Transduction Proteins in Acinetobacter baumannii: Role in Antibiotic Resistance, Virulence, and Potential as Drug Targets. Front Microbiol 2019, 10, 49. [Google Scholar] [CrossRef] [PubMed]
- Kroger, C.; Kary, S.C.; Schauer, K.; Cameron, A.D. Genetic Regulation of Virulence and Antibiotic Resistance in Acinetobacter baumannii. Genes (Basel) 2016, 8. [Google Scholar] [CrossRef]
- Russo, T.A.; Manohar, A.; Beanan, J.M.; Olson, R.; MacDonald, U.; Graham, J.; Umland, T.C. The Response Regulator BfmR Is a Potential Drug Target for Acinetobacter baumannii. mSphere 2016, 1. [Google Scholar] [CrossRef] [PubMed]
- Stewart, G.R.; Wernisch, L.; Stabler, R.; Mangan, J.A.; Hinds, J.; Laing, K.G.; Young, D.B.; Butcher, P.D. Dissection of the heat-shock response in Mycobacterium tuberculosis using mutants and microarrays. Microbiology (Reading) 2002, 148, 3129–3138. [Google Scholar] [CrossRef]
- Wong, D.K.; Lee, B.Y.; Horwitz, M.A.; Gibson, B.W. Identification of fur, aconitase, and other proteins expressed by Mycobacterium tuberculosis under conditions of low and high concentrations of iron by combined two-dimensional gel electrophoresis and mass spectrometry. Infect Immun 1999, 67, 327–336. [Google Scholar] [CrossRef]
- Gavini, N.; Tungtur, S.; Pulakat, L. Peptidyl-prolyl cis/trans isomerase-independent functional NifH mutant of Azotobacter vinelandii. J Bacteriol 2006, 188, 6020–6025. [Google Scholar] [CrossRef]
- Mussi, M.A.; Limansky, A.S.; Viale, A.M. Acquisition of resistance to carbapenems in multidrug-resistant clinical strains of Acinetobacter baumannii: natural insertional inactivation of a gene encoding a member of a novel family of beta-barrel outer membrane proteins. Antimicrob Agents Chemother 2005, 49, 1432–1440. [Google Scholar] [CrossRef]
- Smani, Y.; Fabrega, A.; Roca, I.; Sanchez-Encinales, V.; Vila, J.; Pachon, J. Role of OmpA in the multidrug resistance phenotype of Acinetobacter baumannii. Antimicrob Agents Chemother 2014, 58, 1806–1808. [Google Scholar] [CrossRef]
- Geisinger, E.; Mortman, N.J.; Vargas-Cuebas, G.; Tai, A.K.; Isberg, R.R. A global regulatory system links virulence and antibiotic resistance to envelope homeostasis in Acinetobacter baumannii. PLoS Pathog 2018, 14, e1007030. [Google Scholar] [CrossRef]
- Farrow, J.M., 3rd; Wells, G.; Pesci, E.C. Desiccation tolerance in Acinetobacter baumannii is mediated by the two-component response regulator BfmR. PLoS One 2018, 13, e0205638. [Google Scholar] [CrossRef]
- Mezcord, V.; Wong, O.; Pasteran, F.; Corso, A.; Tolmasky, M.E.; Bonomo, R.A.; Ramirez, M.S. Role of beta-lactamase inhibitors on cefiderocol activity against carbapenem-resistant Acinetobacter species. Int J Antimicrob Agents, 1016. [Google Scholar] [CrossRef]
- Falcone, M.; Tiseo, G.; Leonildi, A.; Della Sala, L.; Vecchione, A.; Barnini, S.; Farcomeni, A.; Menichetti, F. Cefiderocol- Compared to Colistin-Based Regimens for the Treatment of Severe Infections Caused by Carbapenem-Resistant Acinetobacter baumannii. Antimicrob Agents Chemother 2022, 66, e0214221. [Google Scholar] [CrossRef]
- Katsube, T.; Echols, R.; Wajima, T. Pharmacokinetic and Pharmacodynamic Profiles of Cefiderocol, a Novel Siderophore Cephalosporin. Clin Infect Dis 2019, 69, S552–S558. [Google Scholar] [CrossRef]
- Le, C.; Pimentel, C.; Tuttobene, M.R.; Subils, T.; Papp-Wallace, K.M.; Bonomo, R.A.; Actis, L.A.; Tolmasky, M.E.; Ramirez, M.S. Effect of Serum Albumin, a Component of Human Pleural Fluid, on Transcriptional and Phenotypic Changes on Acinetobacter baumannii A118. Curr Microbiol 2021, 78, 3829–3834. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, C.; Le, C.; Tuttobene, M.R.; Subils, T.; Papp-Wallace, K.M.; Bonomo, R.A.; Tolmasky, M.E.; Ramirez, M.S. Interaction of Acinetobacter baumannii with Human Serum Albumin: Does the Host Determine the Outcome? Antibiotics (Basel) 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Le, C.; Pimentel, C.; Tuttobene, M.R.; Subils, T.; Nishimura, B.; Traglia, G.M.; Perez, F.; Papp-Wallace, K.M.; Bonomo, R.A.; Tolmasky, M.E.; et al. Interplay between Meropenem and Human Serum Albumin on Expression of Carbapenem Resistance Genes and Natural Competence in Acinetobacter baumannii. Antimicrob Agents Chemother 2021, 65, e0101921. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, D.; Pasteran, F.; Calderon, M.; Jaber, S.; Traglia, G.M.; Albornoz, E.; Corso, A.; Vila, A.J.; Bonomo, R.A.; Adams, M.D.; et al. Characterisation of ST25 NDM-1-producing Acinetobacter spp. strains leading the increase in NDM-1 emergence in Argentina. J Glob Antimicrob Resist 2020, 23, 108–110. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J Comput Biol 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Deatherage, D.E.; Barrick, J.E. Identification of mutations in laboratory-evolved microbes from next-generation sequencing data using breseq. Methods Mol Biol 2014, 1151, 165–188. [Google Scholar] [CrossRef] [PubMed]
- Antunes, L.C.; Imperi, F.; Towner, K.J.; Visca, P. Genome-assisted identification of putative iron-utilization genes in Acinetobacter baumannii and their distribution among a genotypically diverse collection of clinical isolates. Res Microbiol 2011, 162, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef] [PubMed]
- Hellemans, J.; Mortier, G.; De Paepe, A.; Speleman, F.; Vandesompele, J. qBase relative quantification framework and software for management and automated analysis of real-time quantitative PCR data. Genome Biol 2007, 8, R19. [Google Scholar] [CrossRef] [PubMed]
- Escalante, J.; Nishimura, B.; Tuttobene, M.R.; Subils, T.; Pimentel, C.; Georgeos, N.; Sieira, R.; Bonomo, R.A.; Tolmasky, M.E.; Ramirez, M.S. Human serum albumin (HSA) regulates the expression of histone-like nucleoid structure protein (H-NS) in Acinetobacter baumannii. Sci Rep 2022, 12, 14644. [Google Scholar] [CrossRef]
- (CLSI), C.L.S.I. Performance standards for antimicrobial susceptibility testing: Thirty Edition informational supplement. CLSI Document M100-S30:2020. Clinical Lab Standards Institute.
- Lee, K.; Lim, Y.S.; Yong, D.; Yum, J.H.; Chong, Y. Evaluation of the Hodge test and the imipenem-EDTA double-disk synergy test for differentiating metallo-beta-lactamase-producing isolates of Pseudomonas spp. and Acinetobacter spp. J Clin Microbiol 2003, 41, 4623–4629. [Google Scholar] [CrossRef]
- Pasteran, F.; Mendez, T.; Guerriero, L.; Rapoport, M.; Corso, A. Sensitive screening tests for suspected class A carbapenemase production in species of Enterobacteriaceae. J Clin Microbiol 2009, 47, 1631–1639. [Google Scholar] [CrossRef]
Figure 1.
Expression of genes coding for β-lactamases, outer membrane proteins, and iron uptake functions in the AMA40, AMA40 IHC1, and AMA40 IHC2 strains. qRT-PCR of bla genes (blaADC, and blaNDM-1), genes coding for the outer membrane proteins OmpA, and CarO (A) and (iron uptake-related proteins receptors PirA, PiuA, BauA, BfnH, ExbD and TonB3 (B) expressed in CAMHB. The data shown are the mean ± SD of normalized relative quantities (NRQ) obtained from transcript levels. At least three independent biological samples were tested using four technical replicates for each sample. Statistical significance (P < 0.05) was determined by two-way ANOVA followed by Tukey's multiple comparison test. Significance was indicated by: one asterisk P < 0.05; two asterisks: P < 0.01; three asterisks: P < 0.001; four asterisks: P < 0.0001.
Figure 1.
Expression of genes coding for β-lactamases, outer membrane proteins, and iron uptake functions in the AMA40, AMA40 IHC1, and AMA40 IHC2 strains. qRT-PCR of bla genes (blaADC, and blaNDM-1), genes coding for the outer membrane proteins OmpA, and CarO (A) and (iron uptake-related proteins receptors PirA, PiuA, BauA, BfnH, ExbD and TonB3 (B) expressed in CAMHB. The data shown are the mean ± SD of normalized relative quantities (NRQ) obtained from transcript levels. At least three independent biological samples were tested using four technical replicates for each sample. Statistical significance (P < 0.05) was determined by two-way ANOVA followed by Tukey's multiple comparison test. Significance was indicated by: one asterisk P < 0.05; two asterisks: P < 0.01; three asterisks: P < 0.001; four asterisks: P < 0.0001.
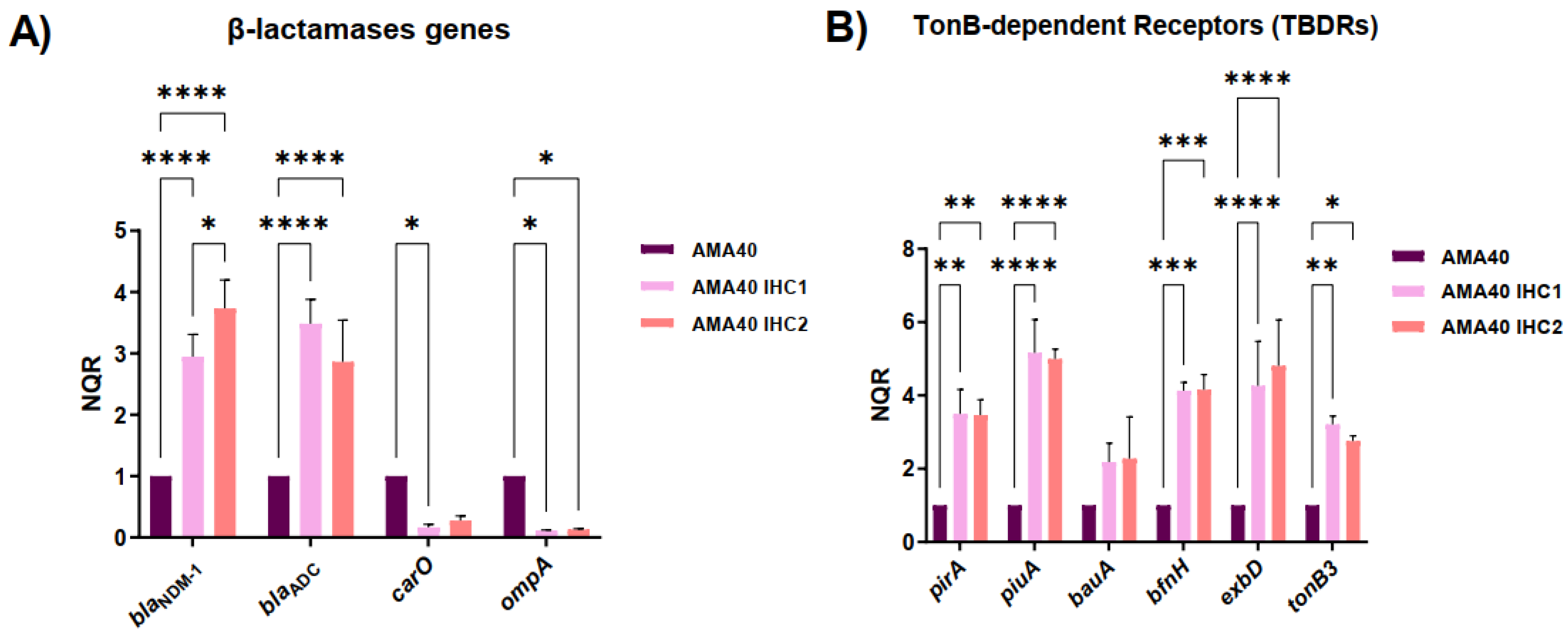
Figure 2.
Static time-kill kinetics of cefiderocol monotherapy (0.5, 1, 4 and 8 mg/L) against an initial inoculum of 5 x 106 CFU/mL of A. baumannii isolates AMA40 (A, D), AMA40 IHC2 (B, E) and AMA40 IHC1 (C, F) in CAMHB (A-C) or CAMHB supplemented with 3.5% HSA (D-F) over 8 h of incubation at 37°C. The black dashed line represents the limit of quantification for CFU/mL bacterial count (1.3 Log10 CFU/mL).
Figure 2.
Static time-kill kinetics of cefiderocol monotherapy (0.5, 1, 4 and 8 mg/L) against an initial inoculum of 5 x 106 CFU/mL of A. baumannii isolates AMA40 (A, D), AMA40 IHC2 (B, E) and AMA40 IHC1 (C, F) in CAMHB (A-C) or CAMHB supplemented with 3.5% HSA (D-F) over 8 h of incubation at 37°C. The black dashed line represents the limit of quantification for CFU/mL bacterial count (1.3 Log10 CFU/mL).
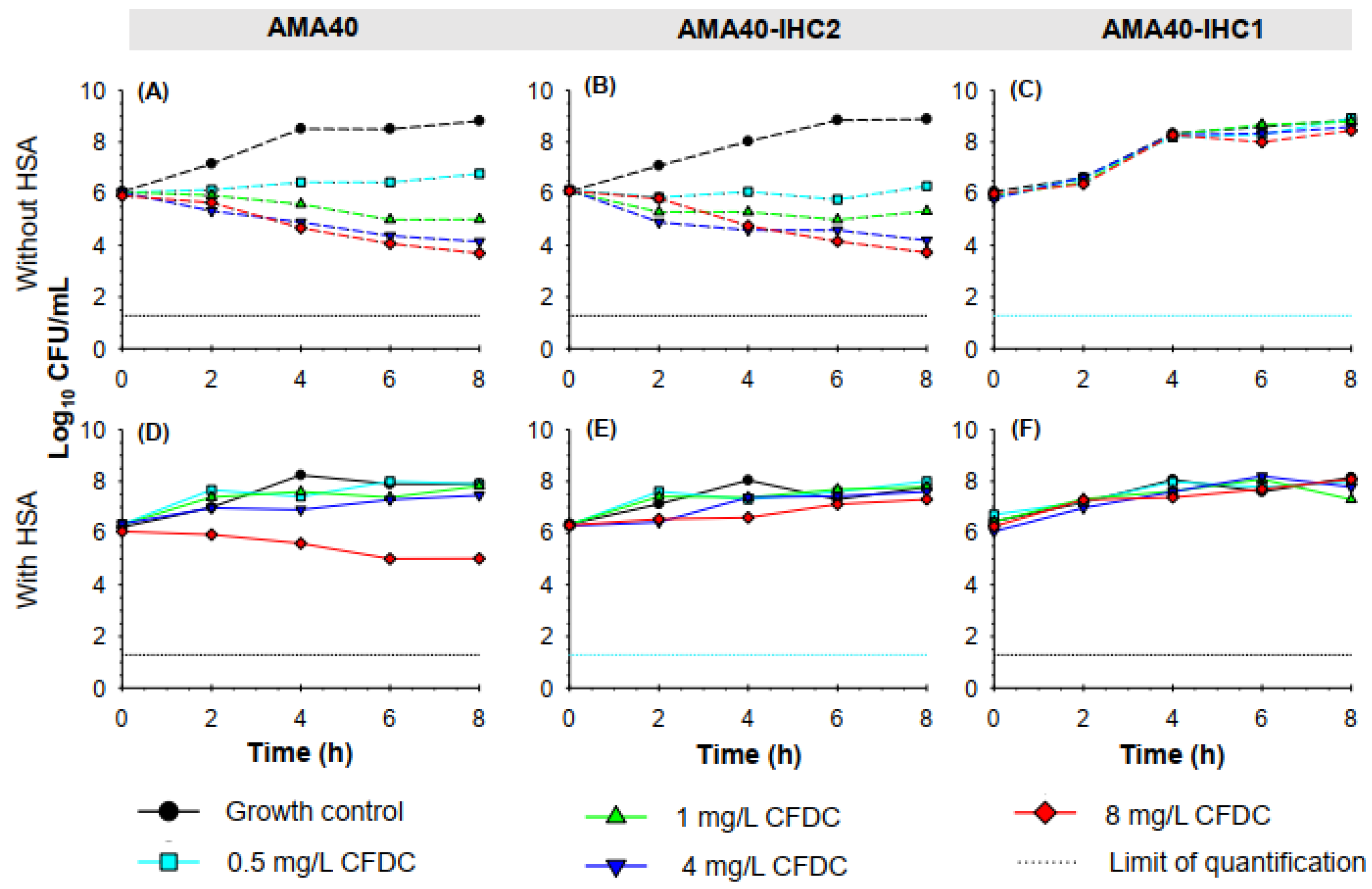
Figure 3.
Reduction or increase of bacterial counts (Log10 CFU/mL) of (A) A. baumannii AMA40, (B) AMA40 IHC2 and (C) AMA40 IHC1 cultured in CAMHB (dashed bars) or CAMHB + 3.5% HSA (solid bars) for the different cefiderocol concentrations evaluated.
Figure 3.
Reduction or increase of bacterial counts (Log10 CFU/mL) of (A) A. baumannii AMA40, (B) AMA40 IHC2 and (C) AMA40 IHC1 cultured in CAMHB (dashed bars) or CAMHB + 3.5% HSA (solid bars) for the different cefiderocol concentrations evaluated.
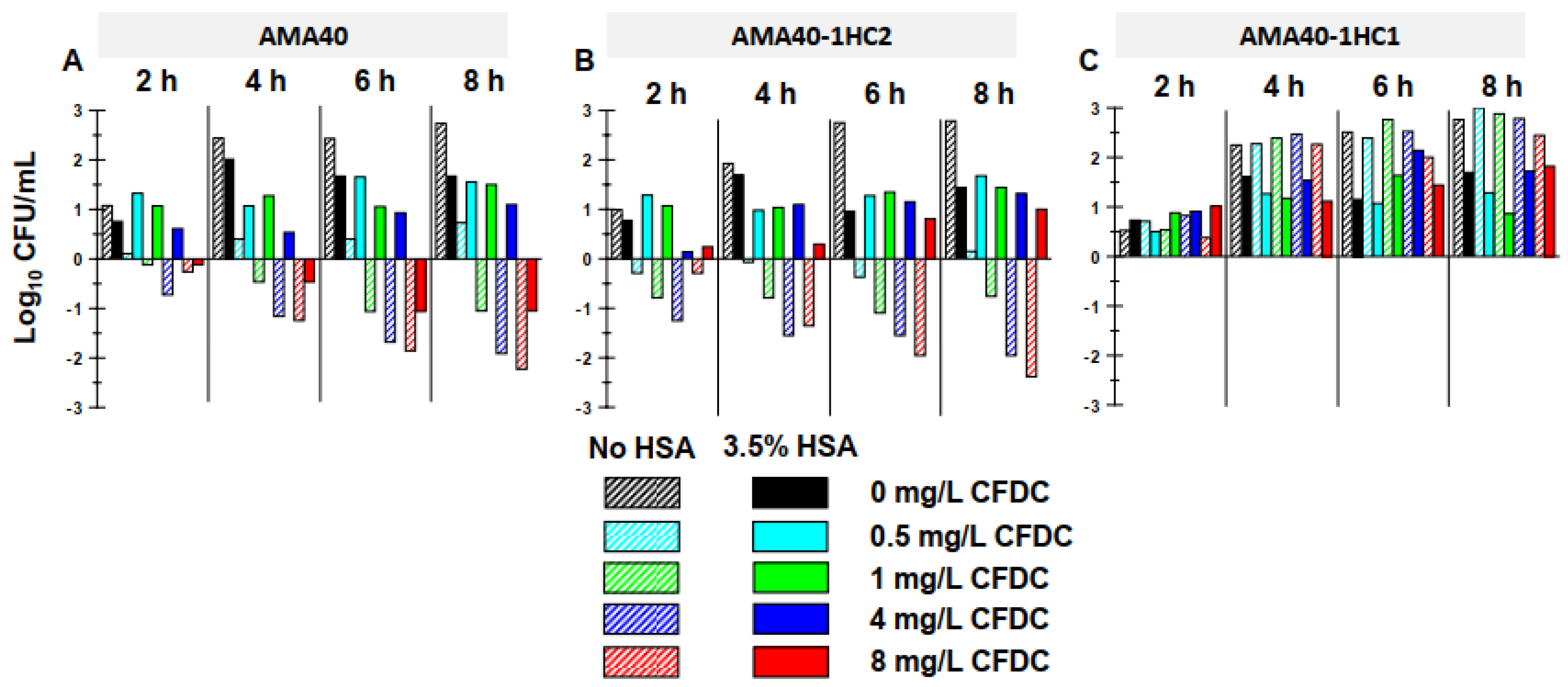
Figure 4.
Genetic and phenotypic analysis of biofilm production. A) Biofilm assays performed with A. baumannii AMA40, AMA40 IHC1 and AMA40 IHC2 represented by the OD580/OD600 ration. Statistical significance (P < 0.05) was determined by two-way ANOVA followed by Tukey's multiple comparison test. Significance was indicated by: P < 0.0001. B) qRT-PCR of csuAB, csuB, csuE, bfmR and pilT genes expressed in CAMHB. The data shown are mean ± SD of normalized relative quantities (NRQ) obtained from transcript levels. At least three independent biological samples were tested using four technical replicates. Statistical significance (P < 0.05) was determined by two-way ANOVA followed by Tukey's multiple comparison test. Significance was indicated by: one asterisk P < 0.05; two asterisks: P < 0.01; and three asterisks: P < 0.001.
Figure 4.
Genetic and phenotypic analysis of biofilm production. A) Biofilm assays performed with A. baumannii AMA40, AMA40 IHC1 and AMA40 IHC2 represented by the OD580/OD600 ration. Statistical significance (P < 0.05) was determined by two-way ANOVA followed by Tukey's multiple comparison test. Significance was indicated by: P < 0.0001. B) qRT-PCR of csuAB, csuB, csuE, bfmR and pilT genes expressed in CAMHB. The data shown are mean ± SD of normalized relative quantities (NRQ) obtained from transcript levels. At least three independent biological samples were tested using four technical replicates. Statistical significance (P < 0.05) was determined by two-way ANOVA followed by Tukey's multiple comparison test. Significance was indicated by: one asterisk P < 0.05; two asterisks: P < 0.01; and three asterisks: P < 0.001.
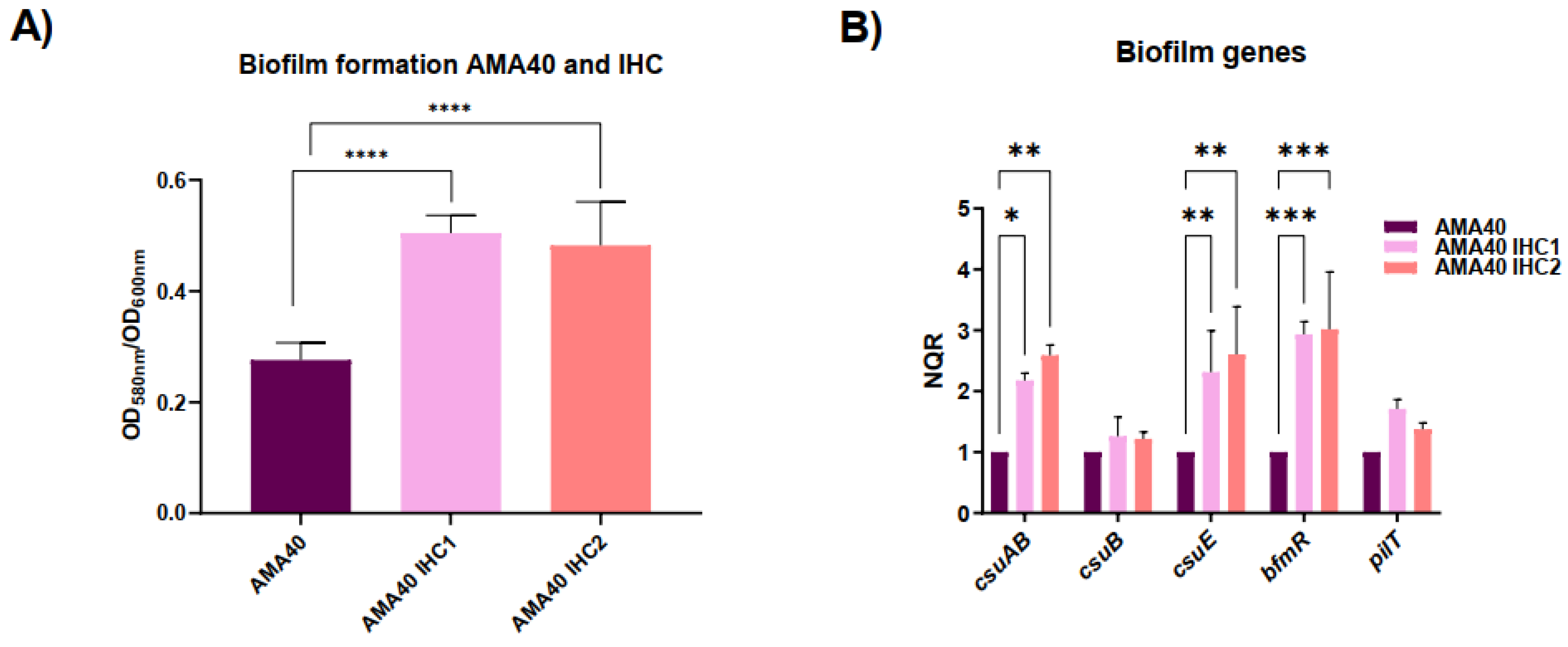

Table 1.
Minimal Inhibitory Concentrations (MICs) of the CRAB AMA40 and heteroresistant strains, performed using MTS strips (Liofilchem S.r.l., Italy) on Cation Adjusted Mueller Hinton Agar.
Table 1.
Minimal Inhibitory Concentrations (MICs) of the CRAB AMA40 and heteroresistant strains, performed using MTS strips (Liofilchem S.r.l., Italy) on Cation Adjusted Mueller Hinton Agar.
| MICs (mg/L) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Strain | CFDC | MEM | GEN | AK | AMS | CIP | M/V | CZA | CX | SXT | I/R | CO | TGC | LEV |
| AMA40 | 0.5 | >256 | >256 | 32 | >256 | >256 | 32 | >256 | >256 | >32 | >32 | 0.125 | 0.125 | 4 |
| AMA40 IHC1 | >256 | >256 | >256 | >256 | >256 | >256 | 64 | >256 | >256 | >32 | >32 | 0.50 | 0.125 | 4 |
| AMA40 IHC2 | 8 | >256 | >256 | >256 | >256 | >256 | 64 | >256 | >256 | >32 | >32 | 0.50 | 0.19 | >32 |
CFDC: cefiderocol, MEM: meropenem, GEN: gengtamicin, AK: amikacin, AMS: ampicillin-sulbactman, CIP: ciprofloxacin, M/V: meropenem-vaborbactam, CZA: ceftazidime-avibactam, CX: cloxacillin, SXT: trimethoprim-sulfamethoxazole, I/R: imipenem-relebactam, CO: colistin, TGC: tigecycline, and LEV: levofloxacin.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



