
Submitted:
13 June 2023
Posted:
14 June 2023
You are already at the latest version
Abstract
LOXL2, a copper-dependent amine oxidase, has emerged as a promising therapeutic target in hepatocellular carcinoma (HCC). Increased LOXL2 expression in HCC has been linked with aggressive phenotype and represents a poor prognostic factor. Here, we focus on mechanisms by which LOXL2 orchestrates multiple oncogenic functions in HCC development. We are reviewing current knowledge about the roles of LOXL2 in the modulation of the HCC tumor microenvironment, formation of premetastatic niches, and epithelial-mesenchymal transition. Also, we are highlighting the complex interplay between LOXL2 and hypoxia, angiogenesis, and vasculogenic mimicry in HCC. At the end of the review, we summarize current LOXL2 inhibitors and discuss their potential in HCC precision treatment.
Keywords:
hepatocellular carcinoma (HCC)
; LOXL2
; tumor microenvironment (TME)
; extracellular matrix (ECM)
; LOXL2 inhibitors
; target therapy
1. Introduction
Hepatocellular carcinoma (HCC) is the most common type of primary liver cancer, accounting for more than 80% of cases [1]. With incidence and mortality rates increasing worldwide, it represents a global healthcare concern [1,2]. At the base of HCC is chronic liver damage leading to cirrhosis, the most often caused by chronic viral hepatitis (Hepatitis B and C), alcohol abuse, and non-alcoholic fatty liver disease (NAFLD) [3]. Around 80% of HCC arise in the cirrhotic liver, and among patients with cirrhosis of any etiology, one-third will develop HCC [1,4,5]. The therapeutic options for HCC are dominantly based on TNM tumor stage, degree of background liver damage and patient's performance status [6]. Therapy for an early-stage HCC is liver resection, liver transplantation, or radiofrequency ablation. Transarterial chemoembolization (TACE) and radiation therapy, alone or in combination with systemic therapy, are used in an intermediate-stage HCC, while systemic therapy (chemotherapy, molecular targeted therapy, immunotherapy, and gene therapy) is a treatment option in advanced-stage HCC [6,7]. Unfortunately, due to asymptomatic nature of early-stage HCC, more than 60% are detected in intermediate or advance-stage, where curative therapeutic options are limited [8]. The backbone of systemic therapy for HCC is sorafenib and other multi-kinase inhibitors [6,9,10,11]. Also, ramucirumab, vascular endothelial growth factor (VEGF) inhibitor, and immune checkpoint inhibitors (nivolumab and pembrolizumab), separately or in combination, are being increasingly used [6,12]. Although therapeutic strategies for advanced HCC are rapidly expanding, they provide a variable but still limited extension of survival, cause a wide range of side effects and eventually lead to the development of resistance, which is recognized as one of the biggest problems in the HCC treatment [9]. The mechanism underlying therapy resistance in HCC are multiple and not completely understood, but it has been shown that tumor microenvironment (TME) plays an important role. Modification of TME might serve as a valuable strategy to improve current and develop novel therapeutic options and resensitize resistant tumors to existing therapeutic agents [9,13,14].
In the last fifteen years, lysyl oxidase-like 2 (LOXL2) has emerged as one of the major mediator between tumors cells and TME, involved in every step of tumor progression [15,16,17,18,19,20]. LOXL2 involvement has been reported in the regulation of cancer cell proliferation, epithelial-mesenchymal transition (EMT), migration, extravasation, and creating pre-metastatic niches at distant sites as reviewed by Zhang et al. [21], Lin et al. [22], and Wen et al [23]. The mechanisms by which LOXL2 affects tumor invasiveness can be used as a typical model of solid cancer progression and spreading [24]. In addition, LOXL2 regulates tumor-induced angiogenesis [25,26,27] and mediates the interaction between cancer cells and cancer-associated fibroblasts (CAF) [20,28]. Taken together, LOXL2 represents a multifunctional protein enrolled in almost every step of solid tumor propagation, and in this review, we will summarize the multiple roles of LOXL2 in the progression of HCC and its potential for therapeutic targeting.
2. LOXL2 introduction: LOX family, structure and function of LOXL2
LOXL2 is a secreted and copper-dependent amine oxidase belonging to the lysyl oxidase (LOX) family, which consists of five homologous members: LOX and LOX-like l-4 (LOXL1-4) proteins [29,30,31,32,33,34]. The primary function of the LOX family enzymes is to catalyze the cross-linking of elastin and collagen by oxidation, which is essential for the maintenance of rigidity, stability, and remodeling of the extracellular matrix (ECM) [35,36]. The human LOXL2 gene is positioned on chromosome 8p21 22 and encodes a 774 amino acid protein [24,37]. Structurally, the LOXL2 protein contains a variable N-terminal region and a highly conserved C-terminal region with catalytic activity (Figure 1). The catalytic domains on C-terminus are conserved among LOX family members and consist of the copper-binding domain, lysyl tyrosine quinone (LTQ) element required for cofactor formation, and a cytokine receptor-like (CRL) domain [38,39]. The N-terminal domain of LOXL2 is more variable and includes signal peptide and four scavenger receptor cysteine-rich (SRCR) elements responsible for protein-protein interactions [38,40].
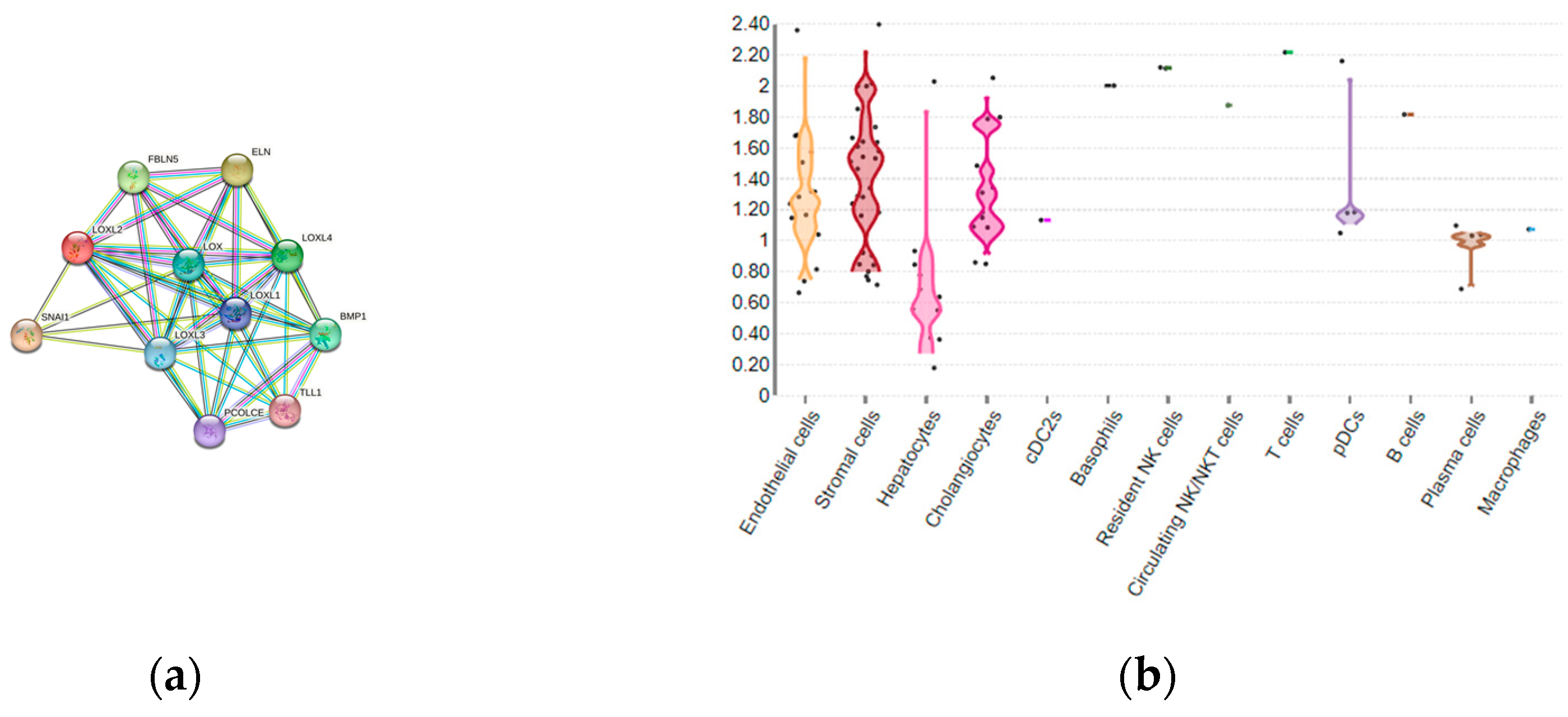
The existence of several different protein domains in the LOX family enzymes connotes their multiple biological functions [24], apart from the ECM sustention role. The LOX and LOXL1-4 proteins enroll in other biological processes, including embryogenesis and development [39,42,43]. Although the LOXL2 is a secretory protein, it is also distributed in the intracellular compartments and within the nucleus and achieves its intracellular and extracellular activities throughout various downstream pathways (Figure 2a) [23]. LOXL2 participates in the structural maintenance of ECM of fibrotic tissues, such as the liver tissue [44,45,46]. LOXL2-specific tissue expression among various liver cell compartments can be estimated using multi-omics datasets presented in Guilliams et al. [47], which detects the overall expression of LOXL2 in 0.7 % of healthy adult liver cells, dominantly in stromal and endothelial cells, as well as in hepatocytes and cholangocytes (Figure 2b) [48].
Altered expression and activation of LOX family members can affect the tissue microenvironment, which is implicated in many pathological conditions, including tissue fibrosis, atherosclerosis, and tumor development [50]. Previous studies indicate that LOXL2 expression is regulated on transcriptional and posttranscriptional levels [51,52,53], as well by alternative splicing [54,55], and by interaction with micro RNA molecules [56,57,58]. Results from several independent studies demonstrate that extracellular factors, such as ECM stiffness or hypoxia, can trigger LOXL2 expression [59,60]. Tumor progression induced by the enhanced LOXL2 expression has been well documented in various digestive system cancers, including HCC [21], where it can be further explored as a potential novel biomarker.
3. LOXL2 expression in HCC and correlation with clinical parameters
It has been shown, both on mRNA and protein levels, that LOXL2 is overexpressed in human HCC tissue compared with healthy liver tissue [15,22,61,62,63,64,65,66]. The LOXL2 protein expression correlated with the amount of fibrosis in the tumor stroma and was more pronounced in the cytoplasm of cancer cells directly adjacent to fibrous stroma, in comparison with centrally located cancer cells [65]. LOXL2 expression positively correlated with direct invasion in adjacent liver tissue [15], more frequent portal vein invasion, poorer tumor differentiation, and more advanced TNM stage [65]. Also, LOXL2 expression in HCC patients, especially high-risk HCCs (HCC tumor size >5 cm, HCCs with portal vein invasion, poor differentiation, and TNM stage II or III) correlated with shorter overall survival (OS), disease-free survival (DFS), disease-specific survival (DSS) and extrahepatic recurrence-free survival (EHRFS) [63,64,65,67]. Hypoxia, chronic inflammation, and fibrosis all induced LOXL2 expression in HCC [13,15]. In human HCC tissue samples, co-expression of LOXL2 and carbon anhydrase IX (CAIX), which is a hypoxia-related biomarker, showed a better correlation with prognostic parameters compared to LOXL2 alone [65]. Interestingly, LOXL2 levels were significantly higher in HCC patients' sera then in sera of non-HCC patients [15], making LOXL2 a promising biomarker for HCC.
4. LOXL2 in the regulation of tumor microenvironment and formation of pre-metastatic niches
TME in HCC is a complex and dynamic landscape composed of tumor cells and various host elements, including cells and extracellular ECM components, which surround tumor cells and interact with them, and with each other in multiple ways. Numerous host cellular components are involved in shaping HCC TME, including activated hepatic stellate cells (HSCs), cancer-associated fibroblasts (CAFs), cells of innate and acquired immunity, sinusoidal endothelial cells and bone marrow-derived cells (BMDCs) [68,69]. These cells interact through and with ECM, and an abundant meshwork of proteins, sugars, and water, which is far from the passive network, but a very dynamic environment involved in every step of tumor development and progression [69,70,71].
ECM constituents can be broadly defined as fibers (collagen, elastic and reticular) and ground substance (proteoglycans, glycoproteins, and glycosaminoglycans). The most important factors determining the specificity of the HCC TME are inflammation, hypoxia, and fibrosis [13,68,69]. HCC is the prototype of inflammation-associated carcinoma as around 90% of HCC arise in the settings of chronic inflammation caused by HBV and HCV or NAFLD [68,72]. Hypoxia, described in more detail later in this review, has been traditionally reported as one of the typical features of HCC [59]. As previously mentioned, more than 80% of HCC arises in the settings of pronounced fibrosis or cirrhosis, which significantly influence TME [1,4,5]. The most abundant ECM component in the fibrotic or cirrhotic liver is collagen type I, which is mainly synthesized by activated HSCs and CAFs. Thus, as it has been shown on various HCC cell lines, chronic inflammation (by activating TGF-β/SMAD4 pathway), hypoxia (dominantly via induction of HIFs, but also through induction of TGF-β) and stiff fibrous ECM (via integrin/JNK/c-JUN pathway) in the HCC microenvironment upregulate LOXL2 levels (Sheme I) [15,60].
Secreted LOXL2 catalyzes covalent cross-linking of collagen, which results in the formation of thick collagen fibers [15]. Increased deposition of collagen fibers and LOXL2-mediated collagen cross-linking results in a further increase in ECM stiffness, which changes biochemical and biomechanical ECM properties and, as it has been shown in cell culture experiments, facilitate cancer cell migration and metastasis, EMT, stemness, and proliferation of HCC cells [13,73,74,75] and correlate with poor prognosis [76]. Additionally, it has been demonstrated that LOXL2-mediated increase in ECM stiffness influences the efficacy of chemotherapeutics 5FU and sorafenib through cell adhesion-mediated drug resistance mechanism [13,75].
Migration of cancer cells, which underlies the local invasion and represents the first step in the metastatic cascade, is a complex multistep process. It involves integrin-mediated mechanosignal transduction from the ECM components to the actin cellular network, which results in cytoskeletal remodeling and formation of lamellipodia or invadosomes at the leading edge of cancer cells [77,78]. It has been shown that LOXL2-mediated collagen cross-linking enhanced focal adhesion signaling and the formation of prominent and dynamic cell protrusions [13,15,79] in HCC-cell lines. LOXL2 influences actin remodeling, which is a prerequisite for the formation of dynamic protrusions and cell migration, by inducing integrin activation and activity of the crucial components of the focal adhesions, including FAK, Rho kinase ROCK, paxillin, and vinculin (Scheme 1) [13,15]. It has been shown in vivo animal experiments that LOXL2 enhanced infiltrative tumor growth on the invasive front and thus stimulates local invasion and intrahepatic and distant metastasis [15].
4.1. LOXL2 and CAFs
One of the crucial components of HCC TME, are cancer-associated fibroblasts (CAFs), which morphologically and phenotypically resemble myofibroblasts [80]. Most commonly CAFs are derived from the hepatic stellate cells (HSCs) and to a lesser extent from the resident tissue fibroblasts, mesenchymal stem cells, or endothelial cells [81]. The crosstalk between CAFs and tumor cells is well established. CAFs promote HCC development, growth, angiogenesis, local invasion, and metastasis through the secretion of numerous growth factors and cytokines, interactions with other cellular elements in HCC TME and ECM remodeling [81,82]. On the other hand, tumor cell secretes cytokines, such as TGF-β, that stimulates CAFs transdifferentiation and activity [82]. In chronic liver diseases that precede HCC development, various inflammatory cytokines transform quiescent HSCs and fibroblasts into activated myofibroblasts. Myofibroblasts synthesize different ECM components, including collagen type I, which leads to fibrosis [83,84,85]. In cell culture experiments, it has been shown that CAFs upregulate LOXL2 levels in HCC cells, thus promoting invasion and metastatic capacity of HCC [86]. Also, in human HCC samples and in the cell culture experiments, it has been shown that CAF-derived chemokine CCL5 activates HIF-1α/ZEB1 cascade and promotes metastasis [87]. As previously mentioned, HIF-1α is a potent inducer of LOXL2 expression [15]. On the other hand, it was demonstrated in cell culture experiments that HCC cells by secreting different soluble mediators, upregulate LOXL2 expression in CAFs, thus creating another of many positive feedback loops in the HCC TME, which facilitate invasion and metastatic spread [86].
4.2. LOXL2 and TAMs
Adding further to the complexity of HCC TME, it has been shown in cell culture experiments that high ECM stiffness induces the polarization of macrophages towards the M2 phenotype and increases their LOXL2 expression. In the same study, authors showed that integrin β5-FAK-MEK1/2-ERK1/2 pathway mediate ECM stiffness-induced HIF-1α upregulation, and HIF-1α further stimulated LOXL2 expression in M2 polarized macrophages [61,88]. M2 macrophages phenotypically resemble TAMs (most TAMs show M2 polarization) and are involved in the progression of HCC by secreting various growth- and angiogenic factors, ECM remodeling, immunosuppressive functions [89,90].
4.3. LOXL2 in the formation of pre-metastatic niches
Today, it is known that, before the metastatic spread has occurred, tumors actively induce changes at distant sites that contribute to the formation of permissive microenvironment, which will facilitate survival and proliferation of circulating tumor cells upon arrival. These modified microenvironments are named pre-metastatic niches (PMNs) [91]. The two most commonly described changes in PMNs are bone marrow-derived cells (BMDC) recruitment and ECM remodeling. In cell culture experiments, it has been shown that secreted LOXL2 influence seeding, motility, and invasion of BMDCs. Moreover, secreted LOXL2 induced MMP9 and fibronectin expression in lung fibroblast, which suggests a role in ECM remodeling (Sheme I) [60]. The number of HCC cells that adhered to the LOXL2-exposed lung fibroblast was significantly higher than in the absence of the LOXL2 exposure [60]. Experiments in vivo revealed that mice bearing HCC tumor with LOXL2 inhibition demonstrate significantly reduced frequency of lung metastasis [15,92]. Furthermore, mice with HCC tumors with LOXL2 inhibition showed a significant reduction in collagen-crosslinking and BMDCs infiltration in the lungs, all of which indicated the important role of LOXL2 in PMNs formation [15,92].
5. LOXL2 role in EMT
Another important event in the metastatic spread of carcinomas is the process of epithelial-mesenchymal transition through which the cell partially or completely acquires a mesenchymal phenotype and advantage of migration, which facilitates the detachment of cancer cells from the tissue of origin and further invasion through surrounding tissues, intravasation and extravasation, and distant sites dissemination [93,94]. Previous studies demonstrated the relationship between LOXL2 intracellular activities and EMT induction in different tumor types [95,96,97]. Epithelial-mesenchymal transition usually starts with the downregulation of E-cadherin expression [98], and one of the direct E-cadherin repressors is the Snail transcription factor, which elevated expression was detected in many invasive carcinomas [99]. The mechanism underlying LOXL2-induced EMT includes the inhibition of E-cadherin expression via interaction with the transcription factor Snail (Sheme I) [99]. At first, the study by Peinado et al. [99], revealed that the amine oxidase activity of LOXL2 regulates Snail stability, and subsequently inhibits the expression of E-cadherin. However, further analysis showed that LOXL2 point mutants with inactivated catalytic activity also induce epithelial-mesenchymal transition by physical interaction with Snail1 [100]. Concerning HCC, Park et al. [92] revealed that the FoxM1b transcription factor, which plays an important role in HCC progression, increases LOXL2 expression through direct binding to the promoter of the LOXL2 gene, which consequently activates the AKT-Snail1 pathway and triggers EMT in mouse-derived HCC cell lines and boosts liver fibrosis and HCC metastasis in vivo. In addition, a previous study on HCC human samples confirmed a significant correlation between LOXL2 expression and EMT status determined based on the vimentin to E-cadherin ratio [101].
6. LOXL2 and hypoxia, angiogenesis and vasculogenic mimicry
Oxygen deprivation or hypoxia is a frequent characteristic of TME in solid tumors, including HCC [59]. Moreover, standard palliative HCC treatments impair blood inflow and suppresses cancer growth, simultaneously creating a hypoxic microenvironment [15]. Hypoxic conditions trigger the activation of the hypoxia-inducible factor, HIF-1α, which acts as a transcription activator of numerous genes, including LOXL2, by binding the hypoxia-responsive element (HRE) within the LOXL2 promoter (Sheme I) [102,103]. In the experiments on human HCC cell lines, Wong et al. demonstrated that HIF-1α directly enhanced LOXL2 expression and indirectly via the TGF-B/SMAD4 pathway [15]. Moreover, Tse et al. [104] studied the interaction between HBV viral oncoprotein HBx with transactivator activity and HIF-1α in mice HCC lines, and results revealed that HBx overexpression induces the stabilization of HIF-1α, which further activate LOXL2 expression and HCC metastasis in transgenic HBx mice. Reversely, results published by Fan et al. [105], showed that LOXL2 also acts as an activator of HIF-1α expression in HCC cell lines through the Snail-FBP1 pathway. Collectively, these findings support the significant role of LOXL2 in HCC progression induced by hypoxia.
HCC represents one of the most vascularized solid tumors, the main feature of highly malignant tumors [106,107]. Hypoxic conditions in the TME induced de novo vascularization or angiogenesis, which additionally enhances tumor growth, invasion, and metastases (Sheme I) [108]. The promoter of classical tumor angiogenesis is VEGF, which expression can be regulated by LOXL2 via the PDGFRβ/Akt axis [109]. Standard management of advanced HCC with the first-line anti-angiogenic agents (sorafenib, lenvatinib, regorafenib) represents a promising treatment approach, although patients' response is unsatisfactory [110,111,112]. Anti-angiogenic therapies further cause oxygen and nutrient deprivation within HCC tissue and may induce the alternative method of tumor neovascularization called vascular mimicry (VM) [113]. VM process characterizes the formation of lined vessels consisting of tumor cells supported by the ECM elements [114], and in HCC, VM is associated with tumor aggressiveness and poor clinical outcome [115]. Recent studies provide evidence that LOXL2 is also involved in VM during the progression of HCC [27,61]. Wang et al. [61] investigated mechanisms underlying enhanced levels of LOXL2 in hypoxic TME of HCC cells and noted that LOXL2 activated by HIF-1α also promotes VM, thus affecting HCC progression. Further in vitro and in vivo analysis of VM formation in HCC revealed that LOXL2 induces VM and tumor metastasis through the upregulation of Snail1 and VE-cadherin [27]. A better understanding of LOXL2-mediated VM formation may indicate a new strategy for designing the therapy that could compensate side effects of classical anti-angiogenic drugs in HCC treatment.
7. LOXL2 and micro-RNAs in HCC
Recent studies revealed that micro-RNAs (miRNAs) may be differentially expressed in HCC, and some of them function as negative epigenetic regulators of LOXL2 and subsequently affect LOXL2 TME-mediated processes. Wong et al. demonstrated that miR-26 and miR-29 suppress LOXL2 in HCC via direct binding to the 3′UTR of LOXL2 mRNA [116]. Specifically, miR29-a has been identified as a negative regulator of hypoxia-responsive genes such as HIF-1α, VEGFa, LOX, and LOXL2 in HCC cell line and HCC tissue functional studies [63,117]. Recent bioinformatics analysis proposed hsa-mir-192-5p as a potential LOXL2 regulator in HCC, however, more in vivo studies are necessary to clarify interactions between LOXL2 and specific micro-RNAs in HCC pathogenesis [118].
8. LOXL2 as potential target for HCC treatment
The well-established treatment options for liver cancer patients have limited clinical efficacy in the late stages of HCC patients due to severe fibrosis and cirrhosis [119]. Therefore, chemoresistant HCC cases require novel combined treatment strategies. Since LOXL2 is unquestionably involved in almost every step of HCC progression and dissemination, this protein fulfills conditions to be considered as a potential target for developing molecular anti-cancer therapy. Considering LOXL2 protein structure, two approaches for targeting were considered in drug development studies. The first was based on the fact that LOXL2 demands copper-ion for enzyme activity, and the second was to target the lysine tyrosylquinone region (LTQ), which possesses cofactor binding function [120].Designing highly selective LOXL2 inhibitors could be promising and beneficial for HCC patients with LOXL2 overexpression since down-regulated LOXL2 expression reduces tumor cell invasiveness and metastasis [121].
The first monoclonal LOXL2 antibody, AB0023, which specifically binds the fourth SRCR domain, was developed in 2010 and showed effectiveness in clinical studies (Table 1) [46,122,123]. Moreover, its efficacy was demonstrated in both tumor xenografts as well as in liver fibrosis models [46,122]. On the other hand, simtuzumab, a humanized LOXL2 antibody (AB0024), hasn’t achieved satisfactory results in clinical trials for fibrotic diseases and several solid tumors [124,125,126]. The lack of clinical effectiveness can be explained by the fact that specific antibody inhibitors probably deactivate only extracellular LOXL2, while intracellular functions of LOXL2 remain due to incomplete antibody internalization or compensation by the activities of other LOX family members [23,74]. In the search for more selective LOXL2 inhibitors, most efforts were oriented toward design a small molecule inhibitors to increase specificity and efficacy, and to minimize side effects [120]. According to the data available on the PHAROS web interface for exploring target/ligand interactions [127], for the query ”LOXL2”, there are 227 active ligands as a result (ChEMBL compounds with an activity cutoff of < 30 nM), but clinical trials focusing on testing LOXL2 based-drugs in HCC are still lacking.
One of the most explored LOX inhibitors is β-aminopropionitrile (BAPN), a small irreversible inhibitor that blocks the catalytic activity of all LOX members and displays specific affinity to LOXL2 [120,128]. Previous studies showed significant tumor suppressive effects of BAPN in several solid tumors and tumor-cell lines [129,130,131,132,133]. Moreover, it has been reported that BAPN affects TME and impedes interaction between cancer-associated fibroblast and gastric cancer cells, resulting in reduced frequency and size of liver metastasis [134]. Considering HCC, it has been demonstrated that BAPN inhibits tumor growth and angiogenesis in vivo and hampers migration and invasion of HCC cell lines [64,101]. According to the study by Liu et al. [135], BAPN also showed an ameliorative effect in the CCl4-induced model of liver fibrosis. The first selective LOXL2 inhibitor, LOXL2-IN-1 hydrochloride, demonstrated potential to be used in HCC treatment since it suppresses Snail1, HIF-1α, and VEGF, the main drivers of HCC progression as mentioned above [67]. In recent years, the second generation of small-molecular-weight haloallylamine-based LOXL2 inhibitors was explored, including PXS-5338, PXS-5382, and PXS-5878, which showed promising results in inhibiting of catalytic activity of LOXL2 [136]. A dual LOXL2/LOXL3 inhibitor, PXS-5153A, designed in 2019 by Schilter et al. [137], also demonstrated ameliorate effects and significantly improved liver fibrosis.
So far, the most promising results in the context of the potential utility of LOXL2 inhibitors in HCC treatment have been published by Gong et al. [13]. The authors demonstrated that a highly selective inhibitor of the LOXL2, namely (2-chloropyridin-4-yl) methanamine [138] in combination with 5FU and sorafenib treatment, significantly decreased the size of a tumor in a mouse model study. Moreover, HCC cells treated with this agent cell lines show a significantly better response to 5FU and sorafenib treatment [13]. These results allude that LOXL2 inhibition has the potential to be more successful in patients with HCC in late fibrotic stages with the concomitant application of 5FU and sorafenib. However, these observations on animal studies have to be validated in further clinical studies.
8. Conclusions
Although treatment strategies for HCC are continuously developing, treatment of intermediate- and advanced-stage HCC remains a major clinical challenge, indicating a great need for novel molecular therapeutic targets. Data summarized in this review emphasize the multiple roles of LOXL2 in crucial processes and factors that regulate the progression of HCC and response to current chemotherapeutics: remodeling of HCC TME, stimulating migration of cancer cells and therefore local invasion, formation of premetastatic niches and metastasis, EMT, angiogenesis and vasculogenic mimicry. All these features make LOXL2 an attractive potential target for innovative therapy design, and further research for clinical-pharmacological solutions of LOXL2-based therapy is warranted. Also, more comprehensive analyses are needed to establish the role of other LOX family members and mutual interactions inside the LOX members in HCC pathogenesis. Additionally, detailed knowledge of multiple LOXL2 interactions with various TME components in HCC and other malignancies, as well as in healthy tissues, is necessary to understand as many effects of potential LOXL2 inhibition. In conclusion, the development of a highly selective LOXL2 inhibitor compatible with concomitant application with standard HCC chemotherapy might be the future approach in HCC personalized medicine.
Author Contributions
Writing—original draft preparation, J.R., B.K, S.D., and I.K.B.; Conceptualization, B.K., S.D. and J.R.; writing—review and editing, S.D., B.K., J.K., and T.V.; visualization, B.V.; supervision, I.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| HCC | Hepatocellular Carcinoma |
| LOXL2 | Lysyl Oxidase-like 2 |
| HBV | Hepatitis B Virus |
| HCV | Hepatitis C Virus |
| NAFLD | Nonalcoholic Fatty Liver Disease |
| TACE | Transarterial Chemoembolization |
| VEGF | Vascular Endothelial Growth Factor |
| TME | Tumor Microenvironment |
| EMT | Epithelial-Mesenchcymal Transition |
| CAF | Cancer-associated Fibroblasts |
| ECM | Extracellular Matrix |
| LTQ | Lysine tyrosylquinone |
| ELN | Elastin |
| FBLN5 | fibulin-5 |
| PCOLCE | procollagen C-endopeptidase enhancer 1 |
| TLL1 | Tolloid-like protein 1 |
| BMP | Bone morphogenic protein 1 |
| OS | Overall Survival |
| DFS | Desease Free Durvival |
| DSS | Disease Specific Survival |
| EHRFS | Extrahepatic Reccurence-free Survival |
| CAIX | Carbon Anhydrase IX |
| HSC | Hepatic Stellate Cells |
| BMDC | Bone Marrow-derived Cells |
| TGF-ß | Transforming Growth Factor Beta |
| HIF-1α | Hypoxia-inducible Factor 1 Alpha |
| JNK-c | Jun N-terminal Kinase |
| 5FU | 5-Fluorouracil |
| FAK | Fokal Adhesion Kinase |
| ROCK | Rho-associated Protein Kinase |
| CCL5 | Chemokine ligand 5 |
| ZEB1 | Zinc finger E-box-binding Homeobox 1 |
| MEK1/2 | Mitogen-activated Protein Kinase 1/2 |
| ERK 1/2 | Extracellular Signal-regulated Kinase 1/2 |
| TAM | Tumor-associated Macrophage |
| PMN | Pre-metastatic Nichae |
| HRE | hypoxia responsive element |
| FBP1 | Fructose-1,6-biphosphatase Protein 1 |
| PDGFRß | Platelt-derived Growth Factor Receptor Beta |
| VM | Vasculogenic Mimicry |
| BAPN | ß-aminopropionitrile. |
References
- Marrero, J.A.; Kulik, L.M.; Sirlin, C.B.; Zhu, A.X.; Finn, R.S.; Abecassis, M.M.; Roberts, L.R.; Heimbach, J.K. Diagnosis, Staging, and Management of Hepatocellular Carcinoma: 2018 Practice Guidance by the American Association for the Study of Liver Diseases. Hepatol. Baltim. Md 2018, 68, 723–750. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA. Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.D.; Hainaut, P.; Gores, G.J.; Amadou, A.; Plymoth, A.; Roberts, L.R. A Global View of Hepatocellular Carcinoma: Trends, Risk, Prevention and Management. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 589–604. [Google Scholar] [CrossRef]
- Affo, S.; Yu, L.-X.; Schwabe, R.F. The Role of Cancer-Associated Fibroblasts and Fibrosis in Liver Cancer. Annu. Rev. Pathol. 2017, 12, 153–186. [Google Scholar] [CrossRef]
- Sangiovanni, A.; Prati, G.M.; Fasani, P.; Ronchi, G.; Romeo, R.; Manini, M.; Del Ninno, E.; Morabito, A.; Colombo, M. The Natural History of Compensated Cirrhosis Due to Hepatitis C Virus: A 17-Year Cohort Study of 214 Patients. Hepatology 2006, 43, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Wege, H.; Li, J.; Ittrich, H. Treatment Lines in Hepatocellular Carcinoma. Visc. Med. 2019, 35, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Suresh, D.; Srinivas, A.N.; Prashant, A.; Harikumar, K.B.; Kumar, D.P. Therapeutic Options in Hepatocellular Carcinoma: A Comprehensive Review. Clin. Exp. Med. 2023. [Google Scholar] [CrossRef]
- Altekruse, S.F.; McGlynn, K.A.; Reichman, M.E. Hepatocellular Carcinoma Incidence, Mortality, and Survival Trends in the United States From 1975 to 2005. J. Clin. Oncol. 2009, 27, 1485–1491. [Google Scholar] [CrossRef]
- Bao, M.H.-R.; Wong, C.C.-L. Hypoxia, Metabolic Reprogramming, and Drug Resistance in Liver Cancer. Cells 2021, 10, 1715. [Google Scholar] [CrossRef]
- Llovet, J.M.; Ricci, S.; Mazzaferro, V.; Hilgard, P.; Gane, E.; Blanc, J.-F.; De Oliveira, A.C.; Santoro, A.; Raoul, J.-L.; Forner, A.; et al. Sorafenib in Advanced Hepatocellular Carcinoma. N. Engl. J. Med. 2008, 359, 378–390. [Google Scholar] [CrossRef]
- Cheng, A.-L.; Kang, Y.-K.; Chen, Z.; Tsao, C.-J.; Qin, S.; Kim, J.S.; Luo, R.; Feng, J.; Ye, S.; Yang, T.-S.; et al. Efficacy and Safety of Sorafenib in Patients in the Asia-Pacific Region with Advanced Hepatocellular Carcinoma: A Phase III Randomised, Double-Blind, Placebo-Controlled Trial. Lancet Oncol. 2009, 10, 25–34. [Google Scholar] [CrossRef]
- Llovet, J.M.; Kelley, R.K.; Villanueva, A.; Singal, A.G.; Pikarsky, E.; Roayaie, S.; Lencioni, R.; Koike, K.; Zucman-Rossi, J.; Finn, R.S. Hepatocellular Carcinoma. Nat. Rev. Dis. Primer 2021, 7, 6. [Google Scholar] [CrossRef]
- Gong, L.; Zhang, Y.; Yang, Y.; Yan, Q.; Ren, J.; Luo, J.; Tiu, Y.C.; Fang, X.; Liu, B.; Lam, R.H.W.; et al. Inhibition of Lysyl Oxidase-like 2 Overcomes Adhesion-dependent Drug Resistance in the Collagen-enriched Liver Cancer Microenvironment. Hepatol. Commun. 2022, 6, 3194–3211. [Google Scholar] [CrossRef] [PubMed]
- Hernandez–Gea, V.; Toffanin, S.; Friedman, S.L.; Llovet, J.M. Role of the Microenvironment in the Pathogenesis and Treatment of Hepatocellular Carcinoma. Gastroenterology 2013, 144, 512–527. [Google Scholar] [CrossRef]
- Wong, C.C.-L.; Tse, A.P.-W.; Huang, Y.-P.; Zhu, Y.-T.; Chiu, D.K.-C.; Lai, R.K.-H.; Au, S.L.-K.; Kai, A.K.-L.; Lee, J.M.-F.; Wei, L.L.; et al. Lysyl Oxidase-like 2 Is Critical to Tumor Microenvironment and Metastatic Niche Formation in Hepatocellular Carcinoma. Hepatology 2014, 60, 1645–1658. [Google Scholar] [CrossRef]
- Payne, S.L.; Hendrix, M.J.C.; Kirschmann, D.A. Paradoxical Roles for Lysyl Oxidases in Cancer—A Prospect. J. Cell. Biochem. 2007, 101, 1338–1354. [Google Scholar] [CrossRef]
- Zhan, X.; Jiao, J.; Zhang, H.; Li, C.; Zhao, J.; Liao, L.; Wu, J.; Wu, B.; Wu, Z.; Wang, S.; et al. A Three-Gene Signature from Protein-Protein Interaction Network of LOXL2 - and Actin-Related Proteins for Esophageal Squamous Cell Carcinoma Prognosis. Cancer Med. 2017, 6, 1707–1719. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Bueno, G.; Salvador, F.; Martín, A.; Floristán, A.; Cuevas, E.P.; Santos, V.; Montes, A.; Morales, S.; Castilla, M.A.; Rojo-Sebastián, A.; et al. Lysyl Oxidase-like 2 (LOXL2), a New Regulator of Cell Polarity Required for Metastatic Dissemination of Basal-like Breast Carcinomas. EMBO Mol. Med. 2011, 3, 528–544. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Lee, J.; Lee, Y.S.; Kim, J.K.; Dong, S.M.; Yoon, D.S. Emerging Role of LOXL2 in the Promotion of Pancreas Cancer Metastasis. Oncotarget 2106, 7, 42539–42552. [Google Scholar] [CrossRef] [PubMed]
- Torres, S.; Garcia-Palmero, I.; Herrera, M.; Bartolomé, R.A.; Peña, C.; Fernandez-Aceñero, M.J.; Padilla, G.; Peláez-García, A.; Lopez-Lucendo, M.; Rodriguez-Merlo, R.; et al. LOXL2 Is Highly Expressed in Cancer-Associated Fibroblasts and Associates to Poor Colon Cancer Survival. Clin. Cancer Res. 2015, 21, 4892–4902. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, W.; Xu, J. Prognostic Utility and Clinical Significance of Lysyl Oxidase-like 2 Protein Expression in Digestive System Cancers. J. Cell. Physiol. 2019, 234, 20713–20720. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.-Y.; Li, C.-J.; Yang, Y.-L.; Huang, Y.-H.; Hsiau, Y.-T.; Chu, P.-Y. Roles of Lysyl Oxidase Family Members in the Tumor Microenvironment and Progression of Liver Cancer. Int. J. Mol. Sci. 2020, 21, 9751. [Google Scholar] [CrossRef]
- Wen, B.; Xu, L.-Y.; Li, E.-M. LOXL2 in Cancer: Regulation, Downstream Effectors and Novel Roles. Biochim. Biophys. Acta BBA - Rev. Cancer 2020, 1874, 188435. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhu, Y. The Function and Mechanisms of Action of LOXL2 in Cancer (Review). Int. J. Mol. Med. 2015, 36, 1200–1204. [Google Scholar] [CrossRef]
- Peng, T.; Deng, X.; Tian, F.; Li, Z.; Jiang, P.; Zhao, X.; Chen, G.; Chen, Y.; Zheng, P.; Li, D.; et al. The Interaction of LOXL2 with GATA6 Induces VEGFA Expression and Angiogenesis in Cholangiocarcinoma. Int. J. Oncol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Xu, S.; Tian, Y.; Ju, A.; Hou, Q.; Liu, J.; Fu, Y.; Luo, Y. Lysyl Oxidase-Like Protein 2 Promotes Tumor Lymphangiogenesis and Lymph Node Metastasis in Breast Cancer. Neoplasia 2019, 21, 413–427. [Google Scholar] [CrossRef]
- Shao, B.; Zhao, X.; Liu, T.; Zhang, Y.; Sun, R.; Dong, X.; Liu, F.; Zhao, N.; Zhang, D.; Wu, L.; et al. LOXL2 Promotes Vasculogenic Mimicry and Tumour Aggressiveness in Hepatocellular Carcinoma. J. Cell. Mol. Med. 2019, 23, 1363–1374. [Google Scholar] [CrossRef]
- Barker, H.E.; Bird, D.; Lang, G.; Erler, J.T. Tumor-Secreted LOXL2 Activates Fibroblasts through FAK Signaling. Mol. Cancer Res. 2013, 11, 1425–1436. [Google Scholar] [CrossRef]
- Mäki, J.M.; Kivirikko, K.I. Cloning and Characterization of a Fourth Human Lysyl Oxidase Isoenzyme. Biochem. J. 2001, 355, 381–387. [Google Scholar] [CrossRef]
- Molnar, J.; Fong, K.S.K.; He, Q.P.; Hayashi, K.; Kim, Y.; Fong, S.F.T.; Fogelgren, B.; Molnarne Szauter, K.; Mink, M.; Csiszar, K. Structural and Functional Diversity of Lysyl Oxidase and the LOX-like Proteins. Biochim. Biophys. Acta BBA - Proteins Proteomics 2003, 1647, 220–224. [Google Scholar] [CrossRef]
- Kenyon, K.; Modi, W.S.; Contente, S.; Friedman, R.M. A Novel Human CDNA with a Predicted Protein Similar to Lysyl Oxidase Maps to Chromosome 15q24-Q25. J. Biol. Chem. 1993, 268, 18435–18437. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Boyd, C.D.; Csiszar, K. A New Gene with Sequence and Structural Similarity to the Gene Encoding Human Lysyl Oxidase. J. Biol. Chem. 1995, 270, 7176–7182. [Google Scholar] [CrossRef] [PubMed]
- Saito, H.; Papaconstantinou, J.; Sato, H.; Goldstein, S. Regulation of a Novel Gene Encoding a Lysyl Oxidase-Related Protein in Cellular Adhesion and Senescence. J. Biol. Chem. 1997, 272, 8157–8160. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y. Cloning and Characterization of a Human Lysyl Oxidase-like 3 Gene (HLOXL3). Matrix Biol. 2001, 20, 153–157. [Google Scholar] [CrossRef]
- Siegel, R.C.; Pinnell, S.R.; Martin, G.R. Cross-Linking of Collagen and Elastin. Properties of Lysyl Oxidase. Biochemistry 1970, 9, 4486–4492. [Google Scholar] [CrossRef] [PubMed]
- Pinnell, S.R.; Martin, G.R. The Cross-Linking of Collagen and Elastin: Enzymatic Conversion of Lysine in Peptide Linkage to Alpha-Aminoadipic-Delta-Semialdehyde (Allysine) by an Extract from Bone. Proc. Natl. Acad. Sci. 1968, 61, 708–716. [Google Scholar] [CrossRef]
- Jourdan-Le Saux, C.; Le Saux, O.; Donlon, T.; Boyd, C.D.; Csiszar, K. The Human Lysyl Oxidase-Related Gene (LOXL2) Maps between Markers D8S280 and D8S278 on Chromosome 8p21.2–P21.3. Genomics 1998, 51, 305–307. [Google Scholar] [CrossRef]
- Csiszar, K. Lysyl Oxidases: A Novel Multifunctional Amine Oxidase Family. In Progress in Nucleic Acid Research and Molecular Biology; Elsevier, 2001; Vol. 70, pp. 1–32 ISBN 978-0-12-540070-1.
- Lucero, H.A.; Kagan, H.M. Lysyl Oxidase: An Oxidative Enzyme and Effector of Cell Function. Cell. Mol. Life Sci. 2006, 63, 2304–2316. [Google Scholar] [CrossRef]
- Xiao, Q.; Ge, G. Lysyl Oxidase, Extracellular Matrix Remodeling and Cancer Metastasis. Cancer Microenviron. 2012, 5, 261–273. [Google Scholar] [CrossRef]
- Meier, A.A.; Kuczera, K.; Mure, M. A 3D–Predicted Structure of the Amine Oxidase Domain of Lysyl Oxidase–Like 2. Int. J. Mol. Sci. 2022, 23, 13385. [Google Scholar] [CrossRef]
- Boufraqech, M.; Zhang, L.; Nilubol, N.; Sadowski, S.M.; Kotian, S.; Quezado, M.; Kebebew, E. Lysyl Oxidase (LOX) Transcriptionally Regulates SNAI2 Expression and TIMP4 Secretion in Human Cancers. Clin. Cancer Res. 2016, 22, 4491–4504. [Google Scholar] [CrossRef] [PubMed]
- Hornstra, I.K.; Birge, S.; Starcher, B.; Bailey, A.J.; Mecham, R.P.; Shapiro, S.D. Lysyl Oxidase Is Required for Vascular and Diaphragmatic Development in Mice. J. Biol. Chem. 2003, 278, 14387–14393. [Google Scholar] [CrossRef] [PubMed]
- Dongiovanni, P.; Meroni, M.; Baselli, G.A.; Bassani, G.A.; Rametta, R.; Pietrelli, A.; Maggioni, M.; Facciotti, F.; Trunzo, V.; Badiali, S.; et al. Insulin Resistance Promotes Lysyl Oxidase Like 2 Induction and Fibrosis Accumulation in Non-Alcoholic Fatty Liver Disease. Clin. Sci. 2017, 131, 1301–1315. [Google Scholar] [CrossRef]
- Zhao, W.; Yang, A.; Chen, W.; Wang, P.; Liu, T.; Cong, M.; Xu, A.; Yan, X.; Jia, J.; You, H. Inhibition of Lysyl Oxidase-like 1 (LOXL1) Expression Arrests Liver Fibrosis Progression in Cirrhosis by Reducing Elastin Crosslinking. Biochim. Biophys. Acta BBA - Mol. Basis Dis. 2018, 1864, 1129–1137. [Google Scholar] [CrossRef] [PubMed]
- Ikenaga, N.; Peng, Z.-W.; Vaid, K.A.; Liu, S.B.; Yoshida, S.; Sverdlov, D.Y.; Mikels-Vigdal, A.; Smith, V.; Schuppan, D.; Popov, Y.V. Selective Targeting of Lysyl Oxidase-like 2 (LOXL2) Suppresses Hepatic Fibrosis Progression and Accelerates Its Reversal. Gut 2017, 66, 1697–1708. [Google Scholar] [CrossRef]
- Guilliams, M.; Bonnardel, J.; Haest, B.; Vanderborght, B.; Wagner, C.; Remmerie, A.; Bujko, A.; Martens, L.; Thoné, T.; Browaeys, R.; et al. Spatial Proteogenomics Reveals Distinct and Evolutionarily Conserved Hepatic Macrophage Niches. Cell 2022, 185, 379–396. [Google Scholar] [CrossRef]
- Liver Cell Atlas. Available online: https://www.livercellatlas.org/umap-humanAll.php (accessed on 10.06.2023).
- String. Available online: https://string-db.org/cgi/network?taskId=b4sTPzQtSCz5&sessionId=bljZKLXOKXys (accessed on 10.06.2023).
- Wang, T.-H.; Hsia, S.-M.; Shieh, T.-M. Lysyl Oxidase and the Tumor Microenvironment. Int. J. Mol. Sci. 2016, 18, 62. [Google Scholar] [CrossRef]
- Xu, L.; Go, E.P.; Finney, J.; Moon, H.; Lantz, M.; Rebecchi, K.; Desaire, H.; Mure, M. Post-Translational Modifications of Recombinant Human Lysyl Oxidase-like 2 (RhLOXL2) Secreted from Drosophila S2 Cells*. J. Biol. Chem. 2013, 288, 5357–5363. [Google Scholar] [CrossRef]
- Luo, W.; Chang, R.; Zhong, J.; Pandey, A.; Semenza, G.L. Histone Demethylase JMJD2C Is a Coactivator for Hypoxia-Inducible Factor 1 That Is Required for Breast Cancer Progression. Proc. Natl. Acad. Sci. 2012, 109. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhu, M.-X.; Zhang, X.-D.; Xu, X.-E.; Wu, Z.-Y.; Liao, L.-D.; Li, L.-Y.; Xie, Y.-M.; Wu, J.-Y.; Zou, H.-Y.; et al. SMYD3 Stimulates EZR and LOXL2 Transcription to Enhance Proliferation, Migration, and Invasion in Esophageal Squamous Cell Carcinoma. Hum. Pathol. 2016, 52, 153–163. [Google Scholar] [CrossRef]
- Liu, C.; Guo, T.; Sakai, A.; Ren, S.; Fukusumi, T.; Ando, M.; Sadat, S.; Saito, Y.; Califano, J.A. A Novel Splice Variant of LOXL2 Promotes Progression of Human Papillomavirus–Negative Head and Neck Squamous Cell Carcinoma. Cancer 2020, 126, 737–748. [Google Scholar] [CrossRef] [PubMed]
- Lv, G.-Q.; Zou, H.-Y.; Liao, L.-D.; Cao, H.-H.; Zeng, F.-M.; Wu, B.-L.; Xie, J.-J.; Fang, W.-K.; Xu, L.-Y.; Li, E.-M. Identification of a Novel Lysyl Oxidase-like 2 Alternative Splicing Isoform, LOXL2 Δe13, in Esophageal Squamous Cell Carcinoma. Biochem. Cell Biol. 2014, 92, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Xiao, H.; Xiao, W.; Xiong, Z.; Hu, W.; Gao, Y.; Ru, Z.; Wang, C.; Bao, L.; Wang, K.; et al. Upregulation of MIAT Regulates LOXL2 Expression by Competitively Binding MiR-29c in Clear Cell Renal Cell Carcinoma. Cell. Physiol. Biochem. 2018, 48, 1075–1087. [Google Scholar] [CrossRef]
- Fukumoto, I.; Kikkawa, N.; Matsushita, R.; Kato, M.; Kurozumi, A.; Nishikawa, R.; Goto, Y.; Koshizuka, K.; Hanazawa, T.; Enokida, H.; et al. Tumor-Suppressive MicroRNAs (MiR-26a/b, MiR-29a/b/c and MiR-218) Concertedly Suppressed Metastasis-Promoting LOXL2 in Head and Neck Squamous Cell Carcinoma. J. Hum. Genet. 2016, 61, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Zhang, J.; Guo, T.; Pan, X. MiR-504 Inhibits Cell Proliferation and Invasion by Targeting LOXL2 in Non Small Cell Lung Cancer. Biomed. Pharmacother. 2018, 97, 1289–1295. [Google Scholar] [CrossRef]
- Gilkes, D.M.; Semenza, G.L.; Wirtz, D. Hypoxia and the Extracellular Matrix: Drivers of Tumour Metastasis. Nat. Rev. Cancer 2014, 14, 430–439. [Google Scholar] [CrossRef]
- Wu, S.; Zheng, Q.; Xing, X.; Dong, Y.; Wang, Y.; You, Y.; Chen, R.; Hu, C.; Chen, J.; Gao, D.; et al. Matrix Stiffness-Upregulated LOXL2 Promotes Fibronectin Production, MMP9 and CXCL12 Expression and BMDCs Recruitment to Assist Pre-Metastatic Niche Formation. J. Exp. Clin. Cancer Res. 2018, 37, 99. [Google Scholar] [CrossRef]
- Wang, M.; Zhao, X.; Zhu, D.; Liu, T.; Liang, X.; Liu, F.; Zhang, Y.; Dong, X.; Sun, B. HIF-1α Promoted Vasculogenic Mimicry Formation in Hepatocellular Carcinoma through LOXL2 up-Regulation in Hypoxic Tumor Microenvironment. J. Exp. Clin. Cancer Res. 2017, 36, 60. [Google Scholar] [CrossRef]
- Zhao, N.; Chen, C.; Guo, Y.; Liu, T.; Che, N.; Zhang, D.; Liang, X.; Zhang, Y.; Zhao, X. LOXL2 Serves as a Prognostic Biomarker for Hepatocellular Carcinoma by Mediating Immune Infiltration and Vasculogenic Mimicry. Dig. Liver Dis. 2023, 55, 661–672. [Google Scholar] [CrossRef]
- Yang, Y.-L.; Tsai, M.-C.; Chang, Y.-H.; Wang, C.-C.; Chu, P.-Y.; Lin, H.-Y.; Huang, Y.-H. MIR29A Impedes Metastatic Behaviors in Hepatocellular Carcinoma via Targeting LOX, LOXL2, and VEGFA. Int. J. Mol. Sci. 2021, 22, 6001. [Google Scholar] [CrossRef]
- Umezaki, N.; Nakagawa, S.; Yamashita, Y.; Kitano, Y.; Arima, K.; Miyata, T.; Hiyoshi, Y.; Okabe, H.; Nitta, H.; Hayashi, H.; et al. Lysyl Oxidase Induces Epithelial-mesenchymal Transition and Predicts Intrahepatic Metastasis of Hepatocellular Carcinoma. Cancer Sci. 2019, cas.14010. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Chung, T.; Rhee, H.; Kim, Y.-J.; Jeon, Y.; Yoo, J.E.; Noh, S.; Han, D.H.; Park, Y.N. Increased Expression of the Matrix-Modifying Enzyme Lysyl Oxidase-Like 2 in Aggressive Hepatocellular Carcinoma with Poor Prognosis. Gut Liver 2019, 13, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhang, Y.; Zhu, Y.; Cong, Q.; Xiang, Y.; Fu, L. The Effect of LOXL2 in Hepatocellular Carcinoma. Mol. Med. Rep. 2016, 14, 1923–1932. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.-Y.; Li, C.-J.; Yang, Y.-L.; Huang, Y.-H.; Hsiau, Y.-T.; Chu, P.-Y. Roles of Lysyl Oxidase Family Members in the Tumor Microenvironment and Progression of Liver Cancer. Int. J. Mol. Sci. 2020, 21, 9751. [Google Scholar] [CrossRef]
- Sas, Z.; Cendrowicz, E.; Weinhäuser, I.; Rygiel, T.P. Tumor Microenvironment of Hepatocellular Carcinoma: Challenges and Opportunities for New Treatment Options. Int. J. Mol. Sci. 2022, 23, 3778. [Google Scholar] [CrossRef] [PubMed]
- Satilmis, B.; Sahin, T.T.; Cicek, E.; Akbulut, S.; Yilmaz, S. Hepatocellular Carcinoma Tumor Microenvironment and Its Implications in Terms of Anti-Tumor Immunity: Future Perspectives for New Therapeutics. J. Gastrointest. Cancer 2021, 52, 1198–1205. [Google Scholar] [CrossRef] [PubMed]
- Guizhen, Z.; Guanchang, J.; Liwen, L.; Huifen, W.; Zhigang, R.; Ranran, S.; Zujiang, Y. The Tumor Microenvironment of Hepatocellular Carcinoma and Its Targeting Strategy by CAR-T Cell Immunotherapy. Front. Endocrinol. 2022, 13, 918869. [Google Scholar] [CrossRef]
- Lu, C.; Rong, D.; Zhang, B.; Zheng, W.; Wang, X.; Chen, Z.; Tang, W. Current Perspectives on the Immunosuppressive Tumor Microenvironment in Hepatocellular Carcinoma: Challenges and Opportunities. Mol. Cancer 2019, 18, 130. [Google Scholar] [CrossRef]
- Yu, L.-X.; Ling, Y.; Wang, H.-Y. Role of Nonresolving Inflammation in Hepatocellular Carcinoma Development and Progression. Npj Precis. Oncol. 2018, 2, 6. [Google Scholar] [CrossRef]
- Lo, C.-M.; Wang, H.-B.; Dembo, M.; Wang, Y. Cell Movement Is Guided by the Rigidity of the Substrate. Biophys. J. 2000, 79, 144–152. [Google Scholar] [CrossRef]
- Liburkin-Dan, T.; Toledano, S.; Neufeld, G. Lysyl Oxidase Family Enzymes and Their Role in Tumor Progression. Int. J. Mol. Sci. 2022, 23, 6249. [Google Scholar] [CrossRef] [PubMed]
- Schrader, J.; Gordon-Walker, T.T.; Aucott, R.L.; Van Deemter, M.; Quaas, A.; Walsh, S.; Benten, D.; Forbes, S.J.; Wells, R.G.; Iredale, J.P. Matrix Stiffness Modulates Proliferation, Chemotherapeutic Response, and Dormancy in Hepatocellular Carcinoma Cells. Hepatology 2011, 53, 1192–1205. [Google Scholar] [CrossRef] [PubMed]
- Tachi, Y.; Hirai, T.; Kojima, Y.; Ishizu, Y.; Honda, T.; Kuzuya, T.; Hayashi, K.; Ishigami, M.; Goto, H. Liver Stiffness Measurement Predicts Hepatocellular Carcinoma Development in Patients Treated with Direct-Acting Antivirals: Liver Stiffness and Cancer Development. JGH Open 2017, 1, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Jiang, J.; Chen, B.; Wang, K.; Tang, Y.; Liang, X. Plasticity of Cancer Cell Invasion: Patterns and Mechanisms. Transl. Oncol. 2021, 14, 100899. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Condeelis, J. Regulation of the Actin Cytoskeleton in Cancer Cell Migration and Invasion. Biochim. Biophys. Acta BBA - Mol. Cell Res. 2007, 1773, 642–652. [Google Scholar] [CrossRef]
- Ezzoukhry, Z.; Henriet, E.; Piquet, L.; Boyé, K.; Bioulac-Sage, P.; Balabaud, C.; Couchy, G.; Zucman-Rossi, J.; Moreau, V.; Saltel, F. TGF-Β1 Promotes Linear Invadosome Formation in Hepatocellular Carcinoma Cells, through DDR1 up-Regulation and Collagen I Cross-Linking. Eur. J. Cell Biol. 2016, 95, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Biffi, G.; Tuveson, D.A. Diversity and Biology of Cancer-Associated Fibroblasts. Physiol. Rev. 2021, 101, 147–176. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Zhu, E.; Zhang, Y. Advances of Cancer-Associated Fibroblasts in Liver Cancer. Biomark. Res. 2022, 10, 59. [Google Scholar] [CrossRef]
- Kubo, N.; Araki, K.; Kuwano, H.; Shirabe, K. Cancer-Associated Fibroblasts in Hepatocellular Carcinoma. World J. Gastroenterol. 2016, 22, 6841. [Google Scholar] [CrossRef]
- Brenner, D.A.; Waterboer, T.; Choi, S.K.; Lindquist, J.N.; Stefanovic, B.; Burchardt, E.; Yamauchi, M.; Gillan, A.; Rippe, R.A. New Aspects of Hepatic Fibrosis. J. Hepatol. 2000, 32, 32–38. [Google Scholar] [CrossRef]
- Török, N.J. Recent Advances in the Pathogenesis and Diagnosis of Liver Fibrosis. J. Gastroenterol. 2008, 43, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Tacke, F.; Luedde, T.; Trautwein, C. Inflammatory Pathways in Liver Homeostasis and Liver Injury. Clin. Rev. Allergy Immunol. 2009, 36, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.-Y.; Chuang, Y.-H.; Chuang, W.-L. Cancer-Associated Fibroblasts up-Regulate CCL2, CCL26, IL6 and LOXL2 Genes Related to Promotion of Cancer Progression in Hepatocellular Carcinoma Cells. Biomed. Pharmacother. 2012, 66, 525–529. [Google Scholar] [CrossRef]
- Xu, H.; Zhao, J.; Li, J.; Zhu, Z.; Cui, Z.; Liu, R.; Lu, R.; Yao, Z.; Xu, Q. Cancer Associated Fibroblast–Derived CCL5 Promotes Hepatocellular Carcinoma Metastasis through Activating HIF1α/ZEB1 Axis. Cell Death Dis. 2022, 13, 478. [Google Scholar] [CrossRef]
- Xing, X.; Wang, Y.; Zhang, X.; Gao, X.; Li, M.; Wu, S.; Zhao, Y.; Chen, J.; Gao, D.; Chen, R.; et al. Matrix Stiffness-mediated Effects on Macrophages Polarization and Their LOXL2 Expression. FEBS J. 2021, 288, 3465–3477. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Shi, Y.; Zhang, M.; Goswami, S.; Afridi, S.; Meng, L.; Ma, J.; Chen, Y.; Lin, Y.; Zhang, J.; et al. Global Immune Characterization of HBV/HCV-Related Hepatocellular Carcinoma Identifies Macrophage and T-Cell Subsets Associated with Disease Progression. Cell Discov. 2020, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Peng, X.; Yang, S.; Li, X.; Huang, M.; Wei, S.; Zhang, S.; He, G.; Liu, J.; Fan, Q.; et al. Targeting Tumor-Associated Macrophages in Hepatocellular Carcinoma: Biology, Strategy, and Immunotherapy. Cell Death Discov. 2023, 9, 65. [Google Scholar] [CrossRef]
- Peinado, H.; Zhang, H.; Matei, I.R.; Costa-Silva, B.; Hoshino, A.; Rodrigues, G.; Psaila, B.; Kaplan, R.N.; Bromberg, J.F.; Kang, Y.; et al. Pre-Metastatic Niches: Organ-Specific Homes for Metastases. Nat. Rev. Cancer 2017, 17, 302–317. [Google Scholar] [CrossRef]
- Park, H.J.; Gusarova, G.; Wang, Z.; Carr, J.R.; Li, J.; Kim, K.; Qiu, J.; Park, Y.; Williamson, P.R.; Hay, N.; et al. Deregulation of FoxM1b Leads to Tumour Metastasis. EMBO Mol. Med. 2011, 3, 21–34. [Google Scholar] [CrossRef]
- Polyak, K.; Weinberg, R.A. Transitions between Epithelial and Mesenchymal States: Acquisition of Malignant and Stem Cell Traits. Nat. Rev. Cancer 2009, 9, 265–273. [Google Scholar] [CrossRef]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.J.; Nieto, M.A. Epithelial-Mesenchymal Transitions in Development and Disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef] [PubMed]
- Mashita, N.; Yamada, S.; Nakayama, G.; Tanaka, C.; Iwata, N.; Kanda, M.; Kobayashi, D.; Fujii, T.; Sugimoto, H.; Koike, M.; et al. Epithelial to Mesenchymal Transition Might Be Induced via CD44 Isoform Switching in Colorectal Cancer: EMT and CD44 in Colorectal Cancer. J. Surg. Oncol. 2014, 110, 745–751. [Google Scholar] [CrossRef]
- Yamada, S.; Fuchs, B.C.; Fujii, T.; Shimoyama, Y.; Sugimoto, H.; Nomoto, S.; Takeda, S.; Tanabe, K.K.; Kodera, Y.; Nakao, A. Epithelial-to-Mesenchymal Transition Predicts Prognosis of Pancreatic Cancer. Surgery 2013, 154, 946–954. [Google Scholar] [CrossRef] [PubMed]
- Murai, T.; Yamada, S.; Fuchs, B.C.; Fujii, T.; Nakayama, G.; Sugimoto, H.; Koike, M.; Fujiwara, M.; Tanabe, K.K.; Kodera, Y. Epithelial-to-Mesenchymal Transition Predicts Prognosis in Clinical Gastric Cancer: EMT in Clinical Gastric Cancer. J. Surg. Oncol. 2014, 109, 684–689. [Google Scholar] [CrossRef] [PubMed]
- Christofori, G.; Semb, H. The Role of the Cell-Adhesion Molecule E-Cadherin as a Tumour-Suppressor Gene. Trends Biochem. Sci. 1999, 24, 73–76. [Google Scholar] [CrossRef]
- Peinado, H.; Del Carmen Iglesias-de La Cruz, M.; Olmeda, D.; Csiszar, K.; Fong, K.S.K.; Vega, S.; Nieto, M.A.; Cano, A.; Portillo, F. A Molecular Role for Lysyl Oxidase-like 2 Enzyme in Snail Regulation and Tumor Progression. EMBO J. 2005, 24, 3446–3458. [Google Scholar] [CrossRef]
- Cuevas, E.P.; Moreno-Bueno, G.; Canesin, G.; Santos, V.; Portillo, F.; Cano, A. LOXL2 Catalytically Inactive Mutants Mediate Epithelial-to-Mesenchymal Transition. Biol. Open 2014, 3, 129–137. [Google Scholar] [CrossRef]
- Ninomiya, G.; Yamada, S.; Hayashi, M.; Takeda, S.; Suenaga, M.; Takami, H.; Kanda, M.; Iwata, N.; Niwa, Y.; Tanaka, C.; et al. Significance of Lysyl Oxidase-like�2 Gene Expression on the Epithelial-mesenchymal Status of Hepatocellular Carcinoma. Oncol. Rep. 2018. [Google Scholar] [CrossRef]
- Postovit, L.-M.; Abbott, D.E.; Payne, S.L.; Wheaton, W.W.; Margaryan, N.V.; Sullivan, R.; Jansen, M.K.; Csiszar, K.; Hendrix, M.J.C.; Kirschmann, D.A. Hypoxia/Reoxygenation: A Dynamic Regulator of Lysyl Oxidase-Facilitated Breast Cancer Migration. J. Cell. Biochem. 2008, 103, 1369–1378. [Google Scholar] [CrossRef]
- Schietke, R.; Warnecke, C.; Wacker, I.; Schödel, J.; Mole, D.R.; Campean, V.; Amann, K.; Goppelt-Struebe, M.; Behrens, J.; Eckardt, K.-U.; et al. The Lysyl Oxidases LOX and LOXL2 Are Necessary and Sufficient to Repress E-Cadherin in Hypoxia. J. Biol. Chem. 2010, 285, 6658–6669. [Google Scholar] [CrossRef]
- Tse, A.P.-W.; Sze, K.M.-F.; Shea, Q.T.-K.; Chiu, E.Y.-T.; Tsang, F.H.-C.; Chiu, D.K.-C.; Zhang, M.S.; Lee, D.; Xu, I.M.-J.; Chan, C.Y.-K.; et al. Hepatitis Transactivator Protein X Promotes Extracellular Matrix Modification through HIF/LOX Pathway in Liver Cancer. Oncogenesis 2018, 7, 44. [Google Scholar] [CrossRef]
- Fan, Z.; Zheng, W.; Li, H.; Wu, W.; Liu, X.; Sun, Z.; Hu, H.; Du, L.; Jia, Q.; Liu, Q. LOXL2 Upregulates Hypoxia-inducible Factor-1α Signaling through Snail-FBP1 Axis in Hepatocellular Carcinoma Cells. Oncol. Rep. 2020. [Google Scholar] [CrossRef]
- Pang, R.W.C.; Joh, J.W.; Johnson, P.J.; Monden, M.; Pawlik, T.M.; Poon, R.T.P. Biology of Hepatocellular Carcinoma. Ann. Surg. Oncol. 2008, 15, 962–971. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Zhou, H.; Zhang, C.; Shang, L.; Zhang, L.; Xu, J.; Zheng, L.; Yuan, Y.; Guo, R.; Jia, W.; et al. A Novel Vascular Pattern Promotes Metastasis of Hepatocellular Carcinoma in an Epithelial–Mesenchymal Transition–Independent Manner. Hepatology 2015, 62, 452–465. [Google Scholar] [CrossRef] [PubMed]
- Kerbel, R.S. Tumor Angiogenesis. N. Engl. J. Med. 2008, 358, 2039–2049. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.-M.; Bird, D.; Welti, J.C.; Gourlaouen, M.; Lang, G.; Murray, G.I.; Reynolds, A.R.; Cox, T.R.; Erler, J.T. Lysyl Oxidase Plays a Critical Role in Endothelial Cell Stimulation to Drive Tumor Angiogenesis. Cancer Res. 2013, 73, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Annese, T.; Ruggieri, S.; Tamma, R.; Crivellato, E. Limitations of Anti-Angiogenic Treatment of Tumors. Transl. Oncol. 2019, 12, 981–986. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.-D.; Sun, H.-C. Emerging Agents and Regimens for Hepatocellular Carcinoma. J. Hematol. Oncol.J Hematol Oncol 2019, 12, 110. [Google Scholar] [CrossRef] [PubMed]
- Bergers, G.; Hanahan, D. Modes of Resistance to Anti-Angiogenic Therapy. Nat. Rev. Cancer 2008, 8, 592–603. [Google Scholar] [CrossRef]
- Zheng, N.; Zhang, S.; Wu, W.; Zhang, N.; Wang, J. Regulatory Mechanisms and Therapeutic Targeting of Vasculogenic Mimicry in Hepatocellular Carcinoma. Pharmacol. Res. 2021, 166, 105507. [Google Scholar] [CrossRef]
- Sun, B.; Zhang, D.; Zhao, N.; Zhao, X. Epithelial-to-Endothelial Transition and Cancer Stem Cells: Two Cornerstones of Vasculogenic Mimicry in Malignant Tumors. Oncotarget 2017, 8, 30502–30510. [Google Scholar] [CrossRef]
- Sun, T.; Sun, B.; Zhao, X.; Zhao, N.; Dong, X.; Che, N.; Yao, Z.; Ma, Y.; Gu, Q.; Zong, W.; et al. Promotion of Tumor Cell Metastasis and Vasculogenic Mimicry by Way of Transcription Coactivation by Bcl-2 and Twist1: A Study of Hepatocellular Carcinoma. Hepatology 2011, 54, 1690–1706. [Google Scholar] [CrossRef]
- Wong, C.C.-L.; Tse, A.P.-W.; Huang, Y.-P.; Zhu, Y.-T.; Chiu, D.K.-C.; Lai, R.K.-H.; Au, S.L.-K.; Kai, A.K.-L.; Lee, J.M.-F.; Wei, L.L.; et al. Lysyl Oxidase-like 2 Is Critical to Tumor Microenvironment and Metastatic Niche Formation in Hepatocellular Carcinoma. Hepatology 2014, 60, 1645–1658. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-H.; Lian, W.-S.; Wang, F.-S.; Wang, P.-W.; Lin, H.-Y.; Tsai, M.-C.; Yang, Y.-L. MiR-29a Curbs Hepatocellular Carcinoma Incidence via Targeting of HIF-1α and ANGPT2. Int. J. Mol. Sci. 2022, 23, 1636. [Google Scholar] [CrossRef]
- Wang, X.; Wu, S.; Yang, Y.; Zhao, J. LncRNA CARMN Affects Hepatocellular Carcinoma Prognosis by Regulating the MiR-192-5p/LOXL2 Axis. Oxid. Med. Cell. Longev. 2022, 2022, 1–24. [Google Scholar] [CrossRef]
- Zhu, Y.; Zheng, B.; Wang, H.; Chen, L. New Knowledge of the Mechanisms of Sorafenib Resistance in Liver Cancer. Acta Pharmacol. Sin. 2017, 38, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Hajdú, I.; Kardos, J.; Major, B.; Fabó, G.; Lőrincz, Z.; Cseh, S.; Dormán, G. Inhibition of the LOX Enzyme Family Members with Old and New Ligands. Selectivity Analysis Revisited. Bioorg. Med. Chem. Lett. 2018, 28, 3113–3118. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, C.; Rodríguez-Sinovas, A.; Martínez-González, J. Lysyl Oxidase as a Potential Therapeutic Target. Drug News Perspect. 2008, 21, 218–224. [Google Scholar] [CrossRef]
- Barry-Hamilton, V.; Spangler, R.; Marshall, D.; McCauley, S.; Rodriguez, H.M.; Oyasu, M.; Mikels, A.; Vaysberg, M.; Ghermazien, H.; Wai, C.; et al. Allosteric Inhibition of Lysyl Oxidase–like-2 Impedes the Development of a Pathologic Microenvironment. Nat. Med. 2010, 16, 1009–1017. [Google Scholar] [CrossRef]
- Rodriguez, H.M.; Vaysberg, M.; Mikels, A.; McCauley, S.; Velayo, A.C.; Garcia, C.; Smith, V. Modulation of Lysyl Oxidase-like 2 Enzymatic Activity by an Allosteric Antibody Inhibitor. J. Biol. Chem. 2010, 285, 20964–20974. [Google Scholar] [CrossRef]
- Verstovsek, S.; Savona, M.R.; Mesa, R.A.; Dong, H.; Maltzman, J.D.; Sharma, S.; Silverman, J.; Oh, S.T.; Gotlib, J. A Phase 2 Study of Simtuzumab in Patients with Primary, Post-Polycythaemia Vera or Post-Essential Thrombocythaemia Myelofibrosis. Br. J. Haematol. 2017, 176, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Harrison, S.A.; Abdelmalek, M.F.; Caldwell, S.; Shiffman, M.L.; Diehl, A.M.; Ghalib, R.; Lawitz, E.J.; Rockey, D.C.; Schall, R.A.; Jia, C.; et al. Simtuzumab Is Ineffective for Patients With Bridging Fibrosis or Compensated Cirrhosis Caused by Nonalcoholic Steatohepatitis. Gastroenterology 2018, 155, 1140–1153. [Google Scholar] [CrossRef] [PubMed]
- Hecht, J.R.; Benson, A.B.; Vyushkov, D.; Yang, Y.; Bendell, J.; Verma, U. A Phase II, Randomized, Double-Blind, Placebo-Controlled Study of Simtuzumab in Combination with FOLFIRI for the Second-Line Treatment of Metastatic KRAS Mutant Colorectal Adenocarcinoma. The Oncologist 2017, 22, 243-e23. [Google Scholar] [CrossRef] [PubMed]
- Pharos. Available online: https://pharos.nih.gov/targets/LOXL2 (accessed on 10.06.2023.).
- Sampath Narayanan, A.; Siegel, R.C.; Martin, G.R. On the Inhibition of Lysyl Oxidase by β-Aminopropionitrile. Biochem. Biophys. Res. Commun. 1972, 46, 745–751. [Google Scholar] [CrossRef]
- Shi, L.; Zhang, N.; Liu, H.; Zhao, L.; Liu, J.; Wan, J.; Wu, W.; Lei, H.; Liu, R.; Han, M. Lysyl Oxidase Inhibition via β-Aminoproprionitrile Hampers Human Umbilical Vein Endothelial Cell Angiogenesis and Migration In�vitro. Mol. Med. Rep. 2018. [Google Scholar] [CrossRef]
- Kirschmann, D.A.; Seftor, E.A.; Fong, S.F.T.; Nieva, D.R.C.; Sullivan, C.M.; Edwards, E.M.; Sommer, P.; Csiszar, K.; Hendrix, M.J.C. A Molecular Role for Lysyl Oxidase in Breast Cancer Invasion. Cancer Res. 2002, 62, 4478–4483. [Google Scholar] [PubMed]
- Abourbih, D.A.; Di Cesare, S.; Orellana, M.E.; Antecka, E.; Martins, C.; Petruccelli, L.A.; Burnier, M.N. Lysyl Oxidase Expression and Inhibition in Uveal Melanoma. Melanoma Res. 2010, 20, 97–106. [Google Scholar] [CrossRef]
- Bondareva, A.; Downey, C.M.; Ayres, F.; Liu, W.; Boyd, S.K.; Hallgrimsson, B.; Jirik, F.R. The Lysyl Oxidase Inhibitor, β-Aminopropionitrile, Diminishes the Metastatic Colonization Potential of Circulating Breast Cancer Cells. PLoS ONE 2009, 4, e5620. [Google Scholar] [CrossRef]
- Yang, X.; Li, S.; Li, W.; Chen, J.; Xiao, X.; Wang, Y.; Yan, G.; Chen, L. Inactivation of Lysyl Oxidase by β-Aminopropionitrile Inhibits Hypoxia-Induced Invasion and Migration of Cervical Cancer Cells. Oncol. Rep. 2013, 29, 541–548. [Google Scholar] [CrossRef]
- Li, Q.; Zhu, C.-C.; Ni, B.; Zhang, Z.-Z.; Jiang, S.-H.; Hu, L.-P.; Wang, X.; Zhang, X.-X.; Huang, P.-Q.; Yang, Q.; et al. Lysyl Oxidase Promotes Liver Metastasis of Gastric Cancer via Facilitating the Reciprocal Interactions between Tumor Cells and Cancer Associated Fibroblasts. eBioMedicine 2019, 49, 157–171. [Google Scholar] [CrossRef]
- Liu, S.B.; Ikenaga, N.; Peng, Z.; Sverdlov, D.Y.; Greenstein, A.; Smith, V.; Schuppan, D.; Popov, Y. Lysyl Oxidase Activity Contributes to Collagen Stabilization during Liver Fibrosis Progression and Limits Spontaneous Fibrosis Reversal in Mice. FASEB J. 2016, 30, 1599–1609. [Google Scholar] [CrossRef] [PubMed]
- Findlay, A.; Turner, C.; Schilter, H.; Deodhar, M.; Zhou, W.; Perryman, L.; Foot, J.; Zahoor, A.; Yao, Y.; Hamilton, R.; et al. An Activity-Based Bioprobe Differentiates a Novel Small Molecule Inhibitor from a LOXL2 Antibody and Provides Renewed Promise for Anti-Fibrotic Therapeutic Strategies. Clin. Transl. Med. 2021, 11, e572. [Google Scholar] [CrossRef] [PubMed]
- Schilter, H.; Findlay, A.D.; Perryman, L.; Yow, T.T.; Moses, J.; Zahoor, A.; Turner, C.I.; Deodhar, M.; Foot, J.S.; Zhou, W.; et al. The Lysyl Oxidase like 2/3 Enzymatic Inhibitor, PXS-5153A, Reduces Crosslinks and Ameliorates Fibrosis. J. Cell. Mol. Med. 2019, 23, 1759–1770. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, J.H.; Rowbottom, M.W.; Lonergan, D.; Darlington, J.; Prodanovich, P.; King, C.D.; Evans, J.F.; Bain, G. Small Molecule Lysyl Oxidase-like 2 (LOXL2) Inhibitors: The Identification of an Inhibitor Selective for LOXL2 over LOX. ACS Med. Chem. Lett. 2017, 8, 423–427. [Google Scholar] [CrossRef]
Figure 1.
LOXL2 protein structure. Arrows indicate variable N-terminal domain and conserved C-terminal domain consisted of several different functional domains represented by boxes. Si, signal peptide; SRCR, scavenger receptor cysteine-rich domain; Cu2+, copper binding domain; LTQ, lysine tyrosylquinone cofactor residues; CF, cofactor formation; CRL, cytokine receptor-like domain. The figure is a not-to-scale drawing (for detailed LOXL2 structure, see Meier et al. [41]).
Figure 1.
LOXL2 protein structure. Arrows indicate variable N-terminal domain and conserved C-terminal domain consisted of several different functional domains represented by boxes. Si, signal peptide; SRCR, scavenger receptor cysteine-rich domain; Cu2+, copper binding domain; LTQ, lysine tyrosylquinone cofactor residues; CF, cofactor formation; CRL, cytokine receptor-like domain. The figure is a not-to-scale drawing (for detailed LOXL2 structure, see Meier et al. [41]).
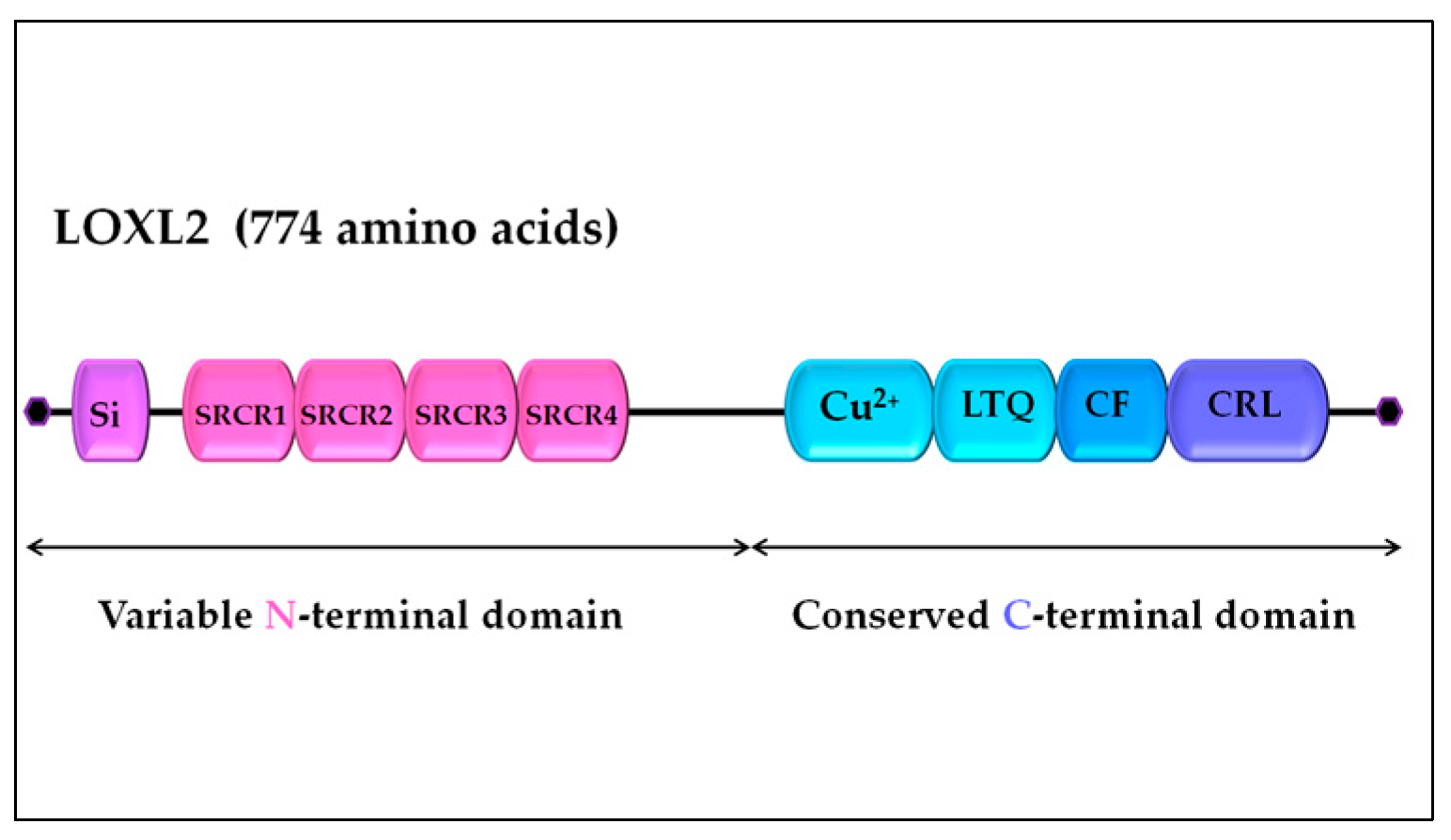
Figure 2.
(a) LOXL2 protein interactome. Available on String platform [49]. SNAI1, zinc finger protein involved in induction of the epithelial-mesenchymal transition (EMT); ELN, elastin; FBLN5, fibulin-5,essential for elastic fiber formation; PCOLCE, procollagen C-endopeptidase enhancer 1, binds to the C-terminal propeptide of type I procollagen; TLL1, Tolloid-like protein 1, protease which processes procollagen C-propeptides; BMP1, Bone morphogenetic protein 1, cleaves the C-terminal propeptides of procollagen I, II and III. (b) LOXL2 spatial expression in healthy liver cells. Available from Liver Cell Atlas online dataset [48].
Figure 2.
(a) LOXL2 protein interactome. Available on String platform [49]. SNAI1, zinc finger protein involved in induction of the epithelial-mesenchymal transition (EMT); ELN, elastin; FBLN5, fibulin-5,essential for elastic fiber formation; PCOLCE, procollagen C-endopeptidase enhancer 1, binds to the C-terminal propeptide of type I procollagen; TLL1, Tolloid-like protein 1, protease which processes procollagen C-propeptides; BMP1, Bone morphogenetic protein 1, cleaves the C-terminal propeptides of procollagen I, II and III. (b) LOXL2 spatial expression in healthy liver cells. Available from Liver Cell Atlas online dataset [48].
Scheme 1.
LOXL2 interaction with HCC tumor microenvironment and roles in HCC progression. ECM, extracellular matrix; EMT, epithelial-mesenchymal transition; VM, vasculogenic mimicry; PMN, pre-metastatic niches.
Scheme 1.
LOXL2 interaction with HCC tumor microenvironment and roles in HCC progression. ECM, extracellular matrix; EMT, epithelial-mesenchymal transition; VM, vasculogenic mimicry; PMN, pre-metastatic niches.
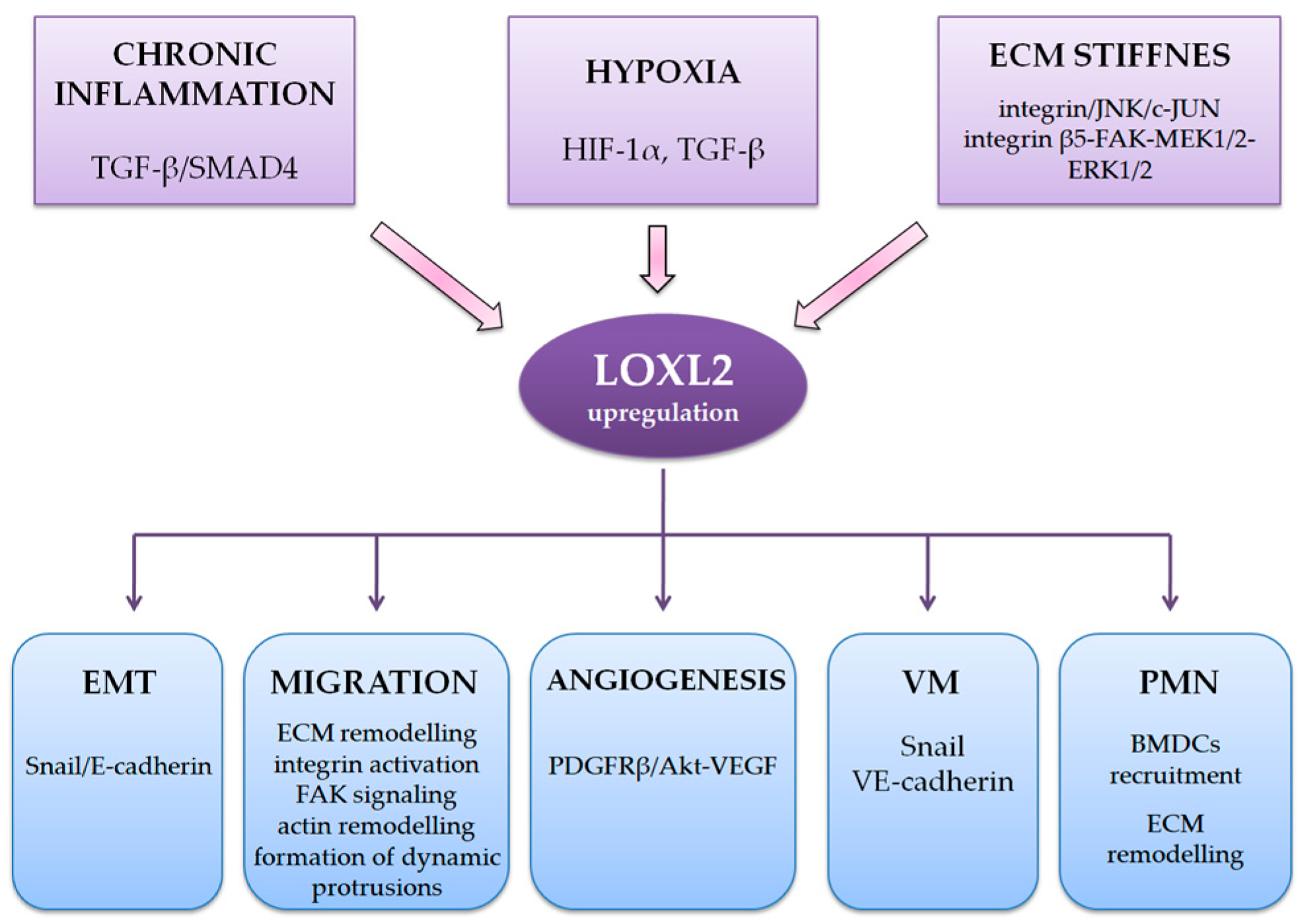

Table 1.
LOXL2 inhibitors in HCC and liver fibrosis.
| Type | Agent | Target | References |
|---|---|---|---|
| monoclonal antibody |
AB0023 | LOXL2 | [46,122,123] |
| AB0024 | LOXL2 | [124,125,126] | |
| small-molecule inhibitor |
BAPN | LOX/LOXL1-4 | [101,134,135] |
| LOXL2-IN-1 | LOXL2 | [105] | |
| PXS-5338 | LOXL2 | [136] | |
| PXS-5382 | LOXL2 | [136] | |
| PXS-5878 | LOXL2 | [136] | |
| PXS-5153A | LOXL2/LOXL3 | [137] | |
| (2-chloropyridin-4-yl) methanamine | LOXL2 | [138] |
* BAPN, beta-aminopropionitrile; PXS, (2S)-propane-1,2-diyl dihexadecanoate.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



