
Submitted:
13 June 2023
Posted:
14 June 2023
You are already at the latest version
Abstract
The problem of increasing life expectancy is solved with the help of many medical and social areas. It has been established that piRNAs and miRNAs can significantly modify the expression of protein-coding genes by suppressing the translation process. The aim of this work was to establish the possibility of binding piRNAs and miRNAs with mRNA of the KLOTHO and FGF23 genes, which promote health and increase life expectancy through participation in key metabolic processes. We used the MirTarget program, which determines the quantitative characteristics of complementary interactions of all piRNAs and miRNAs nucleotides with mRNA of the genes. piR-44682, piR-1940042, piR-3008660, piR-3215034, piR-6885965, piR-7980636 and one miRNA (ID00756.3p-miR) binding to the mRNA of the KLOTHO gene were found in one cluster of binding sites (BSs). piRNA-6890096 interacted with the mRNA of KLOTHO gene in a fully complementary manner using only canonical nucleotides. Among 17494 human genes, target genes interacting with five piRNAs that bind to the mRNA of KLOTHO gene were identified. mRNA of the AFF2, BCL2L11, CPT1A, DAZAP1, NDRG3, SKIDA1, WBP4, ZIC5, ZSWIM6 genes interacted with piR-3215034 and piR-6885965, which formed clusters of BSs located in 5'UTR, CDS and 3'UTR. The piR-576442, piR-1501557, piR-1845735, piR-2069834, and piR-3029987 had BSs in the mRNAs of the FGF23 gene, located only in the 3'UTR. It is proposed to use piRNAs and miRNAs as regulators of the expression of KLOTHO and FGF23 anti-aging genes.
Keywords:
piRNA
; miRNA
; KLOTHO gene
; FGF23 gene
; anti-aging gene
1. Introduction
Chronic kidney disease (CKD) is a set of symptoms resulting from a decrease in the number and function of nephrons. This leads to a violation of the excretory and endocrine activity of the kidneys. As a result, changes in the homeostasis of the internal environment occur, which manifests itself in the violation of many metabolic processes: water-electrolyte, protein, carbohydrate, lipid metabolism. There is an imbalance in the expression of the human genome. As a result, the work of all body systems is disrupted: cardiovascular, respiratory, digestive, hematopoietic and others. All these processes are based on genetically determined changes, therefore, for accurate diagnosis of diseases, it is necessary to study the key molecular genetic foundations of metabolic disorders. Recently, after sequencing of the human genome, it has become possible to establish molecular genetic relationships between various metabolic and physiological processes. Using the example of KLOTHO gene [1], we tried to find out what molecular mechanisms in CKD [2] may be involved in the expression of genes involved in the synthesis of proteins responsible for the manifestation of various diseases. It has been shown that the KLOTHO gene has great potential for future therapeutic purposes in both acute and chronic kidney diseases [3,4,5]. The KLOTHO gene is highly expressed in the kidney (RPKM 80.6) and placenta (RPKM 14.7). A decrease in the synthesis of this protein is observed in patients with CKD, which may underlie diabetes [6,7,8,9]. Mutations in KLOTHO protein are associated with aging and loss of bone mass and phosphorus metabolism [10,11]. Several works are devoted to elucidating the role of KLOTHO in oncological diseases [12,13,14,15]. The KLOTHO protein was initially introduced as an anti-aging molecule [1]. Its deficiency significantly reduces lifespan, and its overexpression extends it and protects against various pathological phenotypes, especially renal disease. Soluble KLOTHO is an anti-aging protein mainly secreted by the kidneys [16] and has been used in diagnostics and therapy [17,18,19,20,21,22].
In recent years, the relationship between KLOTHO and FGF23 proteins has been actively studied. This association is seen in chronic kidney disease and other diseases. The KLOTHO and FGF23 proteins have been studied in renal progression, cardiovascular disorders, and mortality in CKD [23]. The effect of dietary phosphorus restriction on FGF23 and KLOTHO levels in patients with stages 1-2 CKD was studied [24]. It has been established that KLOTHO and FGF23 genes regulate calcium and phosphorus metabolism [25]. The prognostic value of serum KLOTHO and FGF23 proteins was revealed in patients with multiple myeloma [26]. Research was made of the role of KLOTHO and FGF23 genes in cardiovascular disorders in diabetic patients with chronic threatening limb ischemia [27]. The involvement of KLOTHO and FGF23 proteins in phosphorus homeostasis has been established [28]. Data have been obtained on the disruption of the FGF23 and KLOTHO axis in subjects with diabetes mellitus [29]. In recent years, in connection with kidney transplantation, the problem of disturbance of KLOTHO and FGF23 systems has arisen [30]. Small molecule inhibitors of FGF23 signaling have been identified using KLOTHO molecular docking [31]. A correlation has been established between a comparative analysis of FGF23 and cardiovascular diseases in individuals with chronic kidney disease, hypertensive patients, and healthy people [32]. The KLOTHO and FGF23 proteins were used as markers of calcium metabolism [33]. The role of KLOTHO and FGF23 in cardiovascular parameters in diabetic patients with chronic threatening limb ischemia was studied [34]. The FGF23 and KLOTHO proteins have been shown to play an important role in bone and vascular disease in chronic kidney disease [35]. Exercise-mediated activation of the canonical WNT signaling pathway can lead to bone formation and improved levels of the KLOTHO and FGF23 proteins [36]. The KLOTHO has been shown to act either as an obligate FGF23 co-receptor or as a soluble pleiotropic endocrine hormone. With age, human kidney function often deteriorates, and KLOTO levels decrease [20]. FGF23 is a phosphate-regulating protein that is elevated in patients with chronic kidney disease and is associated with cardiovascular mortality, the role of KLOTO has been considered [37]. FGF23 and KLOTHO genes are associated with trabecular bone index but not with bone mineral density in early stages of chronic kidney disease [38].
2. Materials and Methods
The nucleotide (nt) sequence of KLOTHO, FGF23, AFF2, BCL2L11, CPT1A, DAZAP1, NDRG3, SKIDA1, WBP4, ZIC5 and ZSWIM6 genes were downloaded from National Center for Biotechnology Information (NCBI) (https://www.ncbi.nlm.nih.gov, 2022). RPKM of genes were extracted from NCBI. The nucleotide sequences of 480 thousand piRNAs were taken from Wang et al. [39]. The 3707 miRNAs were taken from article Londin et al. [40], the 2567 mature miRNA sequences were taken from miRBase database (http://mirbase.org) and the 1036 miRNAs from article Backes et al. [41]. The piRNA and miRNA binding sites (BSs) in mRNA were predicted using the MirTarget program [42]. This program predicts the following features of piRNA and miRNA binding to mRNA: (a) the initiation of piRNA and miRNA binding to the mRNA from the first nucleotide of the mRNA; (b) the localization of the piRNA and miRNA BSs in the 5′-untranslated region (5′UTR), coding domain sequence (CDS), and 3′-untranslated region (3′UTR) of the mRNAs; (c) the schemes of nucleotide interactions between piRNAs and miRNA with mRNA; (d) the free energy of the interaction between piRNAs, miRNA and the mRNA (ΔG, kJ/mol); and the ratio ΔG/ΔGm (%) is determined for each site. ΔGm equals the free energy of piRNA and miRNA binding with its fully complementary canonical nucleotide sequence. Only piRNAs and miRNA whose nucleotides interacted with mRNA using canonical (G-C and A-U) and noncanonical (G-U and A-C) nucleotides with a given ΔG value were selected from the calculated data. The MirTarget program finds hydrogen bonds between piRNAs and miRNA with mRNA according to the physicochemical characteristics of nucleotide interactions [44,45,46,47,48]. MirTarget differs from other programs in terms of finding piRNA BSs on mRNA in the following: it considers the interaction of piRNA and miRNA with mRNA over the entire piRNA and miRNA sequence; it considers noncanonical pairs G–U and A–C; and it calculates the free energy of the interaction of the piRNAs and miRNA with mRNA. Note that the G, A, C, and U nucleotides, which comprise the RNA structure of microorganisms, plants, and animals, interact identically under equal conditions. Therefore, the physicochemical properties of canonical and noncanonical nucleotide pairs given above do not require additional proof of the previously established physicochemical characteristics of their interaction [43,44,45,46,47].
3. Results
Of the entire piRNA database, only seven piRNAs (piR-44682, piR-1940042, piR-3008660, piR-3215034, piR-5194426, piR-6885965, piR-7980636) bound to the mRNA of the KLOTHO gene (Figure 1). Seven piRNAs each had one BSs and piR-6885965 had two BSs. All BSs were located with a partial overlap of nucleotides, forming a cluster of BSs from 33 nt to 71 nt, only 39 nt long. Such an arrangement of BSs piRNAs in mRNA leads to competition between them for binding to mRNA. piR-6890096 interacts with 3′UTR in a completely complementary manner (the value of ΔG/ΔGm is 100%), which, at its concentration comparable to that of mRNA, will lead to inhibition of the translation process. The KLOTHO gene is many times more strongly expressed in the kidneys than in other organs, and a high concentration of piRNA is required to suppress its synthesis. Of the 7310 miRNAs, only ID00756.3p-miR bound to the mRNA of the KLOTHO gene (Figure 1). ID00756.3p-miR can only interact with mRNA of the KCNN2, NOTCH3, and ZNF592 genes. The ID00756.3p-miR was in the BSs cluster of seven piRNAs located in the CDS mRNA KLOTHO gene, therefore, it competed with them.
Out of 8405000 piRNAs, only eight piRNAs were identified, of which piRNA-6885965 has two BSs in the mRNA of KLOTHO gene. piRNA-6890096 interacted with the mRNA of KLOTHO gene in a completely complementary manner using only canonical nucleotides. Out of 7310 miRNAs, only ID00756.3p-miR could bind to the mRNA of KLOTHO gene. The piRNA (except piRNA-6890096) and miRNA bound in a 39 nt region (Figure 1). Here and henceforth, piRNA BSs with overlapping nucleotide sequences will be referred to as BSs clusters. As a result of this arrangement, piRNA BSs will compete with each other for interaction with the target gene mRNA, and as a result, only one piRNA that binds mRNA more strongly than others or is in a significantly higher concentration will be bound for a longer time than other piRNAs. The formation of clusters of piRNA and miRNA BSs in mRNA is a kind of guarantee of the nonrandom association of small RNAs and their target genes.
The results obtained give hope for the possibility of specific regulation of KLOTHO gene expression using piRNAs and miRNAs. However, given that some miRNAs and piRNAs can bind to mRNAs of several or even hundreds of human genes [48,49,50,51], it is necessary to identify human genes that may be affected by piRNAs and miRNAs that act on mRNAs of KLOTHO gene. That is, it is necessary to identify the possible side effect of these piRNA and miRNA on the expression of all human genes if they are used as therapeutic drugs. To this end, we studied the possible binding of ID00756.3p-miR and nine piRNAs to 17484 human genes.
The largest number of target genes was found for piR-3215034 and piR-6885965. In the mRNA of the AFF2 gene, for each of these piRNAs, Eleven and ten BSs were found, respectively, located in one cluster (Figure 2). The beginning of all BSs were located in the 5’UTR through three nucleotides. The free energy of piR-3215034 binding to the mRNA of the AFF2 gene varied from -155 kJ/mol to 161 kJ/mol, and the ΔG/ΔGm value varied from 94% to 97%. During the interaction of piR-3215034 with mRNA, canonical nucleotide pairs were involved in the last two BSs, except for two C-A bonds. These results indicate a high efficiency of piR-3215034 influence on AFF2 gene expression. piR-6885965 is 24 nt long and, therefore, the free energy of interaction with the mRNA of the AFF2 gene was lower. Since the BSs of these piRNAs are in the same cluster, piR-3215034 has the advantage of binding. However, at a significantly higher concentration of piR-6885965, it will have an advantage in binding over piR-3215034. Therefore, when determining the effectiveness of the action of piRNAs on translation, one should take into account the free energy of their interaction with mRNA and the concentration of competing piRNAs.
Since the AFF2 and KLOTHO genes are targets for piR-3215034 and piR-6885965, it is necessary to compare the possible effect of piRNAs on these genes. It should be noted that the AFF2 gene is involved in the development of squamous cell carcinoma [52], sinonasal tract cancer [53], thoracic carcinoma [54], carcinomas of head and neck [55], and in renal cell carcinoma [56,57]. Therefore, by suppressing the oncogenesis caused by the AFF2 gene with piR-3215034 and piR-6885965, the expression of KLOTHO gene will be simultaneously suppressed.
Several publications have established the involvement of the BCL2L11 gene in oncogenesis [58,59,60,61]. For piR-3215034, 12 BSs were identified, which form a cluster of BSs from 55 nt to 114 nt, 60 nt long (Figure 3). piR-6885965 also binds in this BSs cluster, but with a ΔG/ΔGm value of less than 90%, which is below the selection criterion for significant piRNAs.
The CPT1A gene is involved in fatty acid metabolism and manifests its effect during oncogenesis, diabetes, and cardiomyopathy [62,63,64,65,66,67]. Figure 4 shows the interaction schemes of piR-3215034 and piR-6885965 with mRNA of CPT1A gene, which show that their BSs are located in the same cluster from 99 nt to 137 nt. piR-3215034 interacts with the mRNA of CPT1A gene with a ΔG/ΔGm value of 99%, i.e., almost canonical base pairs are formed.
High expression of the DAZAP1 gene is observed in hepatocarcinoma and can serve as a prognostic marker of the disease. Knockdown of DAZAP1 small interfering RNA markedly inhibited proliferation, migration, and invasion of hepatocarcinoma cells [68,69]. piR-3215034 and piR-6885965 can repress DAZAP1 mRNA translation (Figure 5). The results obtained indicate that these piRNAs can significantly influence the expression of the DAZAP1 gene (Figure 5). It should be noted that both piRNAs bind to the mRNA of gene in the same cluster of BSs located in the 5’UTR, i.e., they can stop protein synthesis before the translation process. The DAZAP1 gene is highly expressed in testis (RPKM 19.6), appendix (RPKM 11.8).
A number of publications have shown the involvement of the NDRG3 gene in the development of oncogenesis, and in most cases, increased expression is observed in cancer of various organs [70,71,72,73,74,75,76]. Therefore, it is important to know whether piR-3215034 and piR-6885965 can suppress the expression of the NDRG3 gene. The results shown in Figure 6 indicate that NDRG3 gene expression can be downregulated by these piRNAs.
The RHOT1 gene is involved in the modification of the development of breast cancer [77], pancreatic cancer [78,79], non-small cell lung cancer [80], hepatocellular carcinoma [81], and the risk and occurrence of Parkinson’s disease [82]. Figure 7 shows the interaction schemes of piR-3215034 and piR-6885965 with mRNA of the RHOT1 gene. Both piRNAs have eight BSs in the 5’UTR of the mRNA of the RHOT1 gene located in the same BSs cluster from the first nucleotide to 46 nt. The next BS was located after three nucleotides, and the free energy of interaction was the same in each of the sites for both piRNAs. Therefore, the value of ΔG/ΔGm was the same for each piRNA.
SKIDA1 was significantly overexpressed in all molecular subgroups, except for only two subgroups of acute myeloid leukemia. In validation analyses, SKIDA1 was associated with a higher sensitivity and specificity in acute myeloid leukemia. We highlight that SKIDA1 is one of the promising markers, which has consistent overexpression among several types of acute leukemia. SKIDA1 identified gene in breast cancer [83]. For piR-3215034, 12 BSs were identified in the mRNA of the SKIDA1 gene (Figure 8). At two sites, piR-3215034 interacts with mRNA almost as complementary as possible, since the ΔG/ΔGm value is 99%. Cluster of piR-3215034 BSs with 61 nt in length guarantees the binding of two of these 27 nt piRNAs at once.
The WBP4 gene (synonymous name FBP21) is involved in splicing and therefore affects the maturation of the mRNA of many genes involved in metabolism [84,85,86]. Eight piR-3215034 and six piR-6885965 binds to the mRNA of the WBP4 gene (Figure 9). At position 94 nt, the 5’UTR of piR-3215034 binds only via canonical base pairs. piR-6885965 binds at the 94 nt position, also with a high ΔG/ΔGm value of 97%.
The ZIC5 gene acts as a transcriptional repressor. Increased expression of this gene is observed in various types of human cancer and may contribute to cancer progression [87,88,89,90]. Figure 10 shows the interaction schemes of piR-3215034 and piR-6885965 with mRNA of the ZIC5 gene, which show a high degree of influence of piR-3215034 on translation. At four positions, the ΔG value is -196 kJ/mol and the ΔG/ΔGm ratio is 96% of the maximum value. For piR-6885965, there are only four BSs in the cluster with a ΔG/ΔGm value greater than 90%.
The transcription factor encoded by the ZSWIM6 gene is synthesized in the brain and can affect the expression of a number of genes. Mutations in this gene lead to malformations of the brain [91,92,93]. Figure 11 shows the binding schemes of piR-3215034 and piR-6885965 to the mRNA of the ZSWIM6 gene. piR-3215034 had seven BSs forming a cluster and piR-6885965 had only three BSs in the same cluster. Transcription factors are difficult to study because the product of their activity can be diverse and difficult to control. The importance of their biological role is undoubted.
The FGF23 gene is a member of a large family of fibroblast growth factors [94], which is most associated with the KLOTHO gene in anti-aging processes [95]. A number of publications have examined the relationship of FGF23 and KLOTHO genes in physiological processes and in various diseases [96,97,98,99,100,101,102,103,104]. In this regard, we studied the possible effect of piRNA on the expression of the FGF23 gene. Only piR-576442, piR-1501557, piR-1845735, piR-2069834, piR-3029987 could bind to the mRNA of the FGF23 gene, the interaction schemes of which are shown in Figure 12. All BSs of these piRNAs were located in the 3’UTR at a considerable distance from each other, i.e., BSs did not form BS clusters.
4. Discussion
The piRNA groups that repress KLOTHO expression can repress the expression of oncogenes. Therefore, if the concentration of these piRNAs is reduced, then life expectancy will increase, but this will increase the likelihood of oncogenesis. Therefore, in order to increase lifespan, it is necessary to reduce the concentration of only piRNA, which suppresses the expression of KLOTHO gene with high selectivity. Some authors call KLOTHO protein a hormone, others an anti-inflammatory agent. The latter function has reason to be, because the composition of KLOTHO protein has an increased content of phenylalanine compared to conventional proteins and is comparable to antioxidant proteins.
Groups of piRNAs that inhibit KLOTHO expression can inhibit the expression of oncogenes. Consequently, if the concentration of these piRNAs is reduced, longevity may increase due to increased KLOTHO protein synthesis, but this will increase the likelihood of oncogenesis and other diseases. Common piRNAs for several genes represent a pool of gene expression regulators and maintain the expression homeostasis of these target genes. A change in the expression of any of these genes will cause a redistribution of the degree of influence of the piRNAs on other genes. Therefore, to increase longevity it is necessary to reduce the concentration of only piRNA-6890096 which suppresses the expression of the KLOTHO gene with high selectivity. Some publications have identified an anti-inflammatory effect of the KLOTHO protein. The latter function has reason to be because the KLOTHO protein has an increased phenylalanine content (6%) compared to the phenylalanine content of conventional proteins and comparable to antioxidant proteins.
References
- Kuro-o, M.; Matsumura, Y.; Aizawa, H.; Kawaguchi, H.; Suga, T.; Utsugi, T.; Ohyama, Y.; Kurabayashi, M.; Kaname, T.; Kume, E.; et al. Mutation of the mouse klotho gene leads to a syndrome resembling ageing. Nature 1997, 390, 45–51. [Google Scholar] [CrossRef]
- Li, S.S.; Sheng, M.J.; Sun, Z.Y.; Liang. Y.; Yu, L.X.; Liu, Q.F. Upstream and downstream regulators of Klotho expression in chronic kidney disease. Metabolism 2023, 142, 155530. [Google Scholar] [CrossRef] [PubMed]
- Ni, W.; Zhang, Y.; Yin, Z. The protective mechanism of Klotho gene-modified bone marrow mesenchymal stem cells on acute kidney injury induced by rhabdomyolysis. Regen Ther. 2021, 13, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Jiang, W.; Li, Y.; Gu, W.; Huang, H.; Wei, Q.; Bai, G.; Wang, J.; Rizak, J.D.; Zhou, Z. Evaluation of Klotho gene expression and NGAL levels following acute kidney injury during pregnancy hypertensive disorders. Pregnancy Hypertens 2022, 30, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Cao, W. Epigenetic modifications of Klotho expression in kidney diseases. J Mol Med (Berl) 2021, 99, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Aziz, M.S.; Aamir, A.; Khan, A.; Khan, Z,; Shah, S. Q. et al. Investigation of Klotho G395A and C1818T Polymorphisms and Their Association with Serum Glucose Level and Risk of Type 2 Diabetes Mellitus. Genes (Basel) 2022, 13, 1532. [Google Scholar] [CrossRef] [PubMed]
- Speer, T.; Schunk, S.J. Klotho in diabetic kidney disease: more than dust in the Wnt. Kidney Int 2022, 102, 469–471. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Banchs, P.A.P.; Liu, Y.; Fu, H.; Arena, V.C.; Forno, E.; Libman, I.; Ho, J.; Muzumdar, R. Serum α-KL, a potential early marker of diabetes complications in youth with T1D, is regulated by miRNA 192. Front Endocrinol (Lausanne) 2022, 13, 937093. [Google Scholar] [CrossRef]
- Typiak, M, Piwkowska, A. Antiinflammatory Actions of Klotho: Implications for Therapy of Diabetic Nephropathy. Int J Mol Sci. 2021, 22, 956. [Google Scholar] [CrossRef]
- Iijima, H.; Gilmer, G.; et al. Age-related matrix stiffening epigenetically regulates α-Klotho expression and compromises chondrocyte integrity. Nat Commun 2023, 14, 18. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, J.; Kalantar-Zadeh, K.; Chen, J. Focusing on Phosphorus Loads: From Healthy People to Chronic Kidney Disease. Nutrients 2023, 15, 1236. [Google Scholar] [CrossRef]
- Liu, S.; Yao, W. Prediction of lung cancer using gene expression and deep learning with KL divergence gene selection. BMC Bioinformatics 2022, 23, 175. [Google Scholar] [CrossRef]
- Xie, B.; Chen, J.; Liu, B.; Zhan, J. Klotho acts as a tumor suppressor in cancers. Pathol Oncol Res 2013, 19, 611–617. [Google Scholar] [CrossRef]
- Kim, G.; Chung, H.; Lee, S.; Kim, W.H. Reduced Klotho expression and its prognostic significance in canine hepatocellular carcinoma. Vet Comp Oncol 2023, 21, 91–99. [Google Scholar] [CrossRef]
- Chung, H.; Lee, S.; Kim, G.A.; Kim, W.H. Down-expression of klotho in canine mammary gland tumors and its prognostic significance. PLoS One 2022, 17, e0265248. [Google Scholar] [CrossRef]
- Xie, H.; Li, N.; Zhou, G.; Liu, Q.; Wang, H.; Han, J.; Shen, L.; Yu, P.; Chen, J.; Chen, X. Plasma S-Klotho level affects the risk of hyperuricemia in the middle-aged and elderly people. Eur J Med Res 2022, 27, 262. [Google Scholar] [CrossRef]
- Abraham, C.R.; Li, A. Aging-suppressor Klotho: Prospects in diagnostics and therapeuticAgeing Res Rev 2022, 82, 101766. 82. [CrossRef]
- Donate-Correa, J.; Matos-Perdomo, E.; González-Luis, A.; Martín-Olivera, A.; Ortiz, A.; Mora-Fernández, C.; Navarro-González, J.F. The Value of Klotho in Kidney Transplantation. Transplantation 2023, 107, 616–627. [Google Scholar] [CrossRef]
- Topal, M.; Guney, I. The association of soluble Klotho levels with anemia and hemoglobin variability in hemodialysis patients. Semin Dial 2023, 36, 142–146. [Google Scholar] [CrossRef]
- Prud’homme, G.J.; Kurt, M.; Wang, Q. Pathobiology of the Klotho Antiaging Protein and Therapeutic Considerations. Front Aging 2022, 3, 931331. [Google Scholar] [CrossRef]
- Xu, J.P.; Zeng, R.X.; He, M.H.; Lin, S.S.; Guo, L.H.; Zhang, M.Z. Associations Between Serum Soluble α-Klotho and the Prevalence of Specific Cardiovascular Disease. Front Cardiovasc Med 2022, 9, 899307. [Google Scholar] [CrossRef]
- Orces, C.H. The association between metabolic syndrome and the anti-aging humoral factor klotho in middle-aged and older adults. Diabetes Metab Syndr 2022, 16, 102522. [Google Scholar] [CrossRef]
- Kim, H.J; Kim, Y.; Kang, M.; Kim, S.; Park, S.K; Sung, S.; Hyun, Y.Y.; Jung, J.Y.; Ahn, C.; Oh, K.H. Low Klotho/Fibroblast Growth Factor 23 Ratio Is an Independent Risk Factor for Renal Progression in Chronic Kidney Disease: Finding From KNOW-CKD. Front Med (Lausanne) 2022, 9, 904963. [Google Scholar] [CrossRef]
- Saxena, A.; Sachan, T.; Gupta, A.; Kapoor, V. Effect of Dietary Phosphorous Restriction on Fibroblast Growth 2 Factor-23 and sKlotho Levels in Patients with Stages 1-2 Chronic Kidney Disease. Nutrients 2022, 14, 3302. [Google Scholar] [CrossRef]
- Portales-Castillo, I.; Simic, P. PTH, FGF-23, Klotho and Vitamin D as regulators of calcium and phosphorus: Genetics, epigenetics and beyond. Front Endocrinol (Lausanne) 2022, 13, 992666. [Google Scholar] [CrossRef]
- Terzi Demirsoy, E.; Mehtap, O.; Birtas Atesoglu, E.; Tarkun, P.; Gedük, A.; Eren, N.; Hacihanefioglu, A. Prognostic Value of Serum Soluble Klotho and Fibroblast Growth Factor-23 in Multiple Myeloma Patients. Indian J Hematol Blood Transfus 2022, 38, 454–463. [Google Scholar] [CrossRef]
- Biscetti, F.; Rando, M.M.; Cecchini, A.L.; Nicolazzi, M.A.; Rossini, E.; Angelini, F.; Iezzi, R.; Eraso, L.H.; Dimuzio, P.J.; Pitocco, D.; Gasbarrini, A.; Massetti, M.; Flex, A. The role of Klotho and FGF23 in cardiovascular outcomes of diabetic patients with chronic limb threatening ischemia: a prospective study. Sci Rep 2023, 13, 6150. [Google Scholar] [CrossRef]
- Mizuno, Y.; Ishida, T.; Kugimiya, F.; Takai, S.; Nakayama, Y.; Yonemitsu, K.; Harada, E. Deterioration of Phosphate Homeostasis Is a Trigger for Cardiac Afterload- Clinical Importance of Fibroblast Growth Factor 23 for Accelerated. Aging Circ Rep 2022, 5, 4–12. [Google Scholar] [CrossRef]
- Castelblanco, E.; Hernández, M.; Alonso, N.; Ribes-Betriu, A.; Real, J. , Granado-Casas, M.; Rossell, J.; Rojo-López M.I.; Dusso, A.S.; Julve, J.; Mauricio, D. Association of α-klotho with subclinical carotid atherosclerosis in subjects with type 1 diabetes mellitus. Cardiovasc Diabetol 2022, 21, 207. [Google Scholar] [CrossRef]
- Reggiani, F.; Moroni, G.; Ponticelli, C. Cardiovascular Risk after Kidney Transplantation: Causes and Current Approaches to a Relevant Burden J Pers Med 2022, 12, 1200. 12. [CrossRef]
- Liu, S.H.; Xiao, Z.; Mishra, S.K.; Mitchell, J.C.; Smith, J.C.; Quarles, L.D.; Petridis, L. Identification of Small-Molecule Inhibitors of Fibroblast Growth Factor 23 Signaling via In Silico Hot Spot Prediction and Molecular Docking to α-Klotho. J Chem Inf Model 2022, 62, 3627–3637. [Google Scholar] [CrossRef]
- Abiola, B.I.; Raji, Y.R.; Ajayi, S.; Adeoye, A.M.; Salako, B.L.; Arije, A.; Kadiri, S. Comparative analysis of fibroblast growth Factor-23 as a correlate of cardiovascular disease among individuals with chronic kidney disease, hypertensives, and healthy controls. Niger J Clin Pract 2022, 25, 1247–1255. [Google Scholar] [CrossRef]
- Golüke, N.M.S.; Schoffelmeer, M.A.; De Jonghe, A.; Emmelot-Vonk, M.H.; De Jong, P.A.; Koek, H.L. Serum biomarkers for arterial calcification in humans: A systematic review Bone Rep, 2022, 17, 101599. 17. [CrossRef]
- Biscetti, F.; Rando, M.M.; Cecchini, A.L.; Nicolazzi, M.A.; Rossini, E.; Angelini, F.; Iezzi, R.; Eraso, L.H.; Dimuzio, P.J.; Pitocco, D.; Gasbarrini, A.; Massetti, M.; Flex, A. The role of Klotho and FGF23 in cardiovascular outcomes of diabetic patients with chronic limb threatening ischemia: a prospective study. Sci Rep 2023, 13, 6150. [Google Scholar] [CrossRef]
- Wang, Y.P.; Sidibé, A.; Fortier, C.; Desjardins, M.P.; Ung, R.V.; Kremer, R.; Agharazii, M.; Mac-Way, F. Wnt/β-catenin pathway inhibitors, bone metabolism and vascular health in kidney transplant patients. J Nephrol 2023. [Google Scholar] [CrossRef]
- Bishop, N.C.; Burton, J.O.; Graham-Brown, M.P.M.; Stensel, D.J.; Viana, J.L.; Watson, E.L. Exercise and chronic kidney disease: potential mechanisms underlying the physiological benefits. Nat Rev Nephrol 2023, 19, 244–256. [Google Scholar] [CrossRef]
- Yanucil, C.; Kentrup, D.; Campos, I.; Czaya, B.; Heitman. K.; Westbrook, D.; Osis, G.; Grabner, A.; Wende, A.R.; Vallejo, J.; Wacker, M.J.; Navarro-Garcia, J.A.; Ruiz-Hurtado, G.; Zhang, F.; Song, Y.; Linhardt, R.J.; White, K.; Kapiloff, M.S.; Faul, C. Kidney Int 2022, 102, 261–279. [Google Scholar] [CrossRef]
- Kužmová, Z.; Kužma, M.; Gažová, A.; Kovářová, M.; Jackuliak, P.; Killinger, Z.; Kyselovič, J.; Payer, J. Fibroblast Growth Factor 23 and Klotho Are Associated With Trabecular Bone Score but Not Bone Mineral Density in the Early Stages of Chronic Kidney Disease: Results of the Cross-Sectional Study. Physiol Res 2021, 70, S43–S51. [Google Scholar] [CrossRef]
- Wang, J.; Shi, Y.; Zhou, H.; Zhang, P.; Song, T.; Ying, Z.; Yu, H.; Li, Y.; Zhao, Y.; Zeng, X.; He, S.; Chen, R. piRBase: integrating piRNA annotation in all aspects. Nucleic Acids Res 2021, 50, 265–272. [Google Scholar] [CrossRef]
- Londin, E.; Loher, P.; Telonis, A.G.; et al. Analysis of 13 cell types reveals evidence for the expression of numerous novel primate- and tissue-specific microRNAs. Proc Natl Acad Sci USA 2015, 112, E1106–E1115. [Google Scholar] [CrossRef]
- Backes, C.; Meder, B.; Hart, M.; Ludwig, N.; Leidinger, P.; Vogel, B.; Galata, V.; Roth, P.; Menegatti, J.; Grasser, F.A.; et al. Prioritizing and selecting likely novel miRNAs from NGS data. Nucleic Acids Res 2015, 44, e53. [Google Scholar] [CrossRef]
- Ivashchenko, A.; Berillo, O.; Pyrkova, A.; Niyazova, R.; Atambayeva, S. MiR-3960 binding sites with mRNA of human genes. Bioinformation 2014, 10, 423–427. [Google Scholar] [CrossRef]
- Friedman RA, Honig BA. Free Energy Analysis of Nucleic Acid Base Stacking in Aqueous Solution. Biophys. J 1995, 69, 1528–1535. [Google Scholar] [CrossRef]
- Garg, A.; Heinemann, U.A. Novel Form of RNA Double Helix Based on G·U and C·A+ Wobble Base Pairing. RNA, 2018, 24, 209–218. [Google Scholar] [CrossRef]
- Leontis, N.B.; Stombaugh, J.; Westhof, E. The Non-watson-crick Base Pairs and Their Associated Isostericity Matrices. Nucleic Acids Res 2002, 30, 3497–3531. [Google Scholar] [CrossRef]
- Kool, E.T. Hydrogen Bonding, Base Stacking, and Steric Effects in DNA Replication. Annu. Rev. Biophys. Biomol. Struct 2001, 30, 1–22. [Google Scholar] [CrossRef]
- Davis, E.; Caiment, F.; Tordoir, X.; Cavaillé, J.; Ferguson-Smith, A.; Cockett, N.; Georges, M.; Charlier, C. RNAi-Mediated Allelic Trans-interaction at the Imprinted Rtl1/ Peg11 Locus. Curr. Biol 2005, 15, 743–749. [Google Scholar] [CrossRef]
- Atambayeva, S.; Niyazova, R.; Ivashchenko, A.; Pyrkova, A.; Pinsky, I.; Akimniyazova, A.; Labeit, S. The Binding Sites of miR-619-5p in the mRNAs of Human and Orthologous Genes. BMC Genom 2017, 18, 428. [Google Scholar] [CrossRef]
- Belkozhayev, A.; Niyazova, R.; Wilson, C.; Jainakbayev, N.; Pyrkova, A.; Ashirbekov, Y.; Akimniyazova, A.; Sharipov, K.; Ivashchenko, A. Bioinformatics Analysis of the Interaction of miRNAs and piRNAs with Human mRNA Genes Having di- and Trinucleotide Repeats. Genes, 2022, 13, 800. [Google Scholar] [CrossRef]
- Kondybayeva, А.М.; Akimniyazova, A.N.; Kamenova, S.U.; Ivashchenko, А.Т. The characteristics of miRNA binding sites in mRNA of ZFHX3 gene and its orthologs. Vavilov journal of genetics and breeding 2018, 22, p–438. [Google Scholar] [CrossRef]
- Kamenova, S.; Sharapkhanova, A.; Akimniyazova, A.; Kuzhybayeva, K.; Kondybayeva, A.; Rakhmetullina, A.; Pyrkova, A.; Ivashchenko, A. piRNA and miRNA Can Suppress the Expression of Multiple Sclerosis Candidate Genes. Nanomaterials (Basel) 2023, 13, 22. [Google Scholar] [CrossRef]
- Kuo, Y.J.; Lewis, J.S.; Zhai, C.; Chen, Y.A.; Chernock, R.D.; Hsieh, M.S.; Lan, M.Y.; Lee, C.K.; Weinreb, I.; Hang, J.F. DEK-AFF2 fusion-associated papillary squamous cell carcinoma of the sinonasal tract: clinicopathologic characterization of seven cases with deceptively bland morphology. Mod Pathol, 2021, 34, 1820–1830. [Google Scholar] [CrossRef]
- Kuo, Y.J.; Lewis, J.S.Jr.; Truong, T.; Yeh, Y.C.; Chernock, R.D.; Zhai, C.; Chen, Y.A.; Hongo, T.; Lee, C.K.; Shi, Q.; Velez Torres, J.M.; Geromes, A.B.; Chu, Y.H.; Hsieh, M.S.; Yamamoto, H.; Weinreb, I.; Hang, J.F. Nuclear expression of AFF2 C-terminus is a sensitive and specific ancillary marker for DEK: AFF2 carcinoma of the sinonasal tract. Mod Pathol 2022, 35, 1587–1595. [Google Scholar] [CrossRef]
- Savari, O.; Chang, J.C.; Bishop, J.A.; Sakthivel, M.K.; Askin, F.B.; Rekhtman, N. First Report of Thoracic Carcinoma With DEK: AFF2 Rearrangement: A Case Report. J Thorac Oncol 2022, 17, 1050–1053. [Google Scholar] [CrossRef]
- Ruangritchankul, K.; Sandison, A. DEK: AFF2 Fusion Carcinomas of Head and Neck. Adv Anat Pathol 2023, 30, 86–94. [Google Scholar] [CrossRef]
- Ji, Z.; Lu, R.; Wu, T.; Chen, Z.; Wen, Z.; Li, Z.; Zheng, X.; Tang, J.; Chen, X.; Yang. Y.; Zheng. Q. Expression profiling of circular RNA reveals a potential miR-145-5p sponge function of circ-AFF2 and circ-ASAP1 in renal cell carcinoma. Am J Transl Res 2023, 15, 82–98. [Google Scholar]
- Ji, Z.; Lu, R.; Wu, T. , Chen, Z., Wen, Z., Li, Z., Zheng, X., Tang, J., Chen, X., Yang, Y., Zheng, Q. Expression profiling of circular RNA reveals a potential miR-145-5p sponge function of circ-AFF2 and circ-ASAP1 in renal cell carcinoma. Am J Transl Res 2023, 15, 82–98. [Google Scholar]
- Taherkhani, A.; Dehto, S.S.; Jamshidi, S.; Shojaei, S. Pathogenesis and prognosis of primary oral squamous cell carcinoma based on microRNAs target genes: a systems biology approach. Genomics Inform 2022, 20, e27. [Google Scholar] [CrossRef]
- Flores, D.; Lopez, A.; Udawant, S.; Gunn, B.; Keniry, M. The FOXO1 inhibitor AS1842856 triggers apoptosis in glioblastoma multiforme and basal-like breast cancer cells. FEBS Open Bio 2023, 13, 352–362. [Google Scholar] [CrossRef]
- Rose, M.M.; Espinoza, V.L.; Hoff, K.J.; Pike, L.A.; Sharma, V.; Hofmann, M.C.; Tan, A.C.; Pozdeyev, N. Schweppe RE. BCL2L11 Induction Mediates Sensitivity to Src and MEK1/2 Inhibition in Thyroid Cancer. Cancers (Basel) 2023, 15, 378. [Google Scholar] [CrossRef]
- Ranapour, S.; Motamed, N. Effect of Silibinin on the Expression of Mir-20b, Bcl2L11, and Erbb2 in Breast Cancer Cell Lines. Mol Biotechnol 2023. [Google Scholar] [CrossRef]
- Li, J.; Zheng, W.; Wu, J.; Zhang, J.; Lv, B.; Li, W.; Liu, J.; Zhang, X.; Huang, T.; Luo, Z. CPT1C-mediated fatty acid oxidation facilitates colorectal cancer cell proliferation and metastasis. Acta Biochim Biophys Sin (Shanghai) 2023. [Google Scholar] [CrossRef]
- Liao, L.; Zhang, F.; Zhuo, Z.; Huang, C.; Zhang, X.; Liu, R.; Gao, B.; Ding, S. Regulation of Fatty Acid Metabolism and Inhibition of Colorectal Cancer Progression by Erchen Decoction Evid Based Complement Alternat Med 2023, 9557720. [CrossRef]
- Xiong, L.; He, T.; Liu, C.; Qin, S.; Xiao, T.; Xin, W.; Wang, Y.; Ran, L.; Zhang, B.; Zhao, J. IL-37 Ameliorates Renal Fibrosis by Restoring CPT1A-Mediated Fatty Acid Oxidation in Diabetic Kidney Disease. Kidney Dis (Basel) 2023, 9, 104–117. [Google Scholar] [CrossRef]
- Li, S.; Liu, M.; Chen, J.; Chen, Y.; Yin, M.; Zhou, Y.; Li. Q.; Xu, F.; Li, Y.; Yan, X.; Xia, Y.; Chen, A.; Lu, D.; Li, C.; Shen, L.; Chen, Z.; Qian, J.; Ge, J. L-carnitine alleviates cardiac microvascular dysfunction in diabetic cardiomyopathy by enhancing PINK1-Parkin-dependent mitophagy through the CPT1a-PHB2-PARL pathways. Acta Physiol (Oxf) 2023, 12, e13975. [Google Scholar] [CrossRef]
- Tian, T.; Lu, Y.; Lin, J.; Chen, M.; Qiu, H.; Zhu, W.; Sun, H.; Huang, J.; Yang, H.; Deng, W. CPT1A promotes anoikis resistance in esophageal squamous cell carcinoma via redox homeostasis. Redox Biol. 2022, 58, 102544. [Google Scholar] [CrossRef]
- Bernard, J.N.; Chinnaiyan, V.; Andl, T.; Le Bras, G.F.; Qureshi, M.N.; Altomare, D.A.; Andl, C.D. Аugmented CPT1A Expression Is Associated with Proliferation and Colony Formation during Barrett’s Tumorigenesis. Int J Mol Sci 2022, 23, 11745. [Google Scholar] [CrossRef]
- Deng, J.J.; Li, G.P.; Lu, W.; Yan, Z.; Wang, Y. DAZAP1 overexpression promotes growth of HCC cell lines: a primary study using CEUS. Clin Transl Oncol 2022, 24, 1168–1176. [Google Scholar] [CrossRef]
- Wang, Q.; Guo, Y.; Wang, W.; Liu, B.; Yang, G.; Xu, Z.; Li, J.; Liu, Z. RNA binding protein DAZAP1 promotes HCC progression and regulates ferroptosis by interacting with SLC7A11 mRNA. Exp Cell Res 2021, 399, 112453. [Google Scholar] [CrossRef]
- Kim, M.C.; Park, M.H.; Kang, S.H.; Bae, Y.K. NDRG3 protein expression is associated with aggressive biologic phenotype and unfavorable outcome in patients with invasive breast cancer. Int J Clin Exp Pathol 2019, 12, 3886–3893. [Google Scholar]
- Liu, Y.; Xia, J.; Zhou, Y.; Shao, S. High expression of NDRG3 correlates with poor prognosis in gastric cancer patients. Rev Esp Enferm Dig 2021, 113, 524–528. [Google Scholar] [CrossRef]
- Ma, W.; Zhao, X.; Xue, N.; Gao, Y.; Xu, Q. The LINC01410/miR-122-5p/NDRG3 axis is involved in the proliferation and migration of osteosarcoma cells. IUBMB Life 2021, 73, 705–717. [Google Scholar] [CrossRef]
- Pappula, A.L.; Rasheed, S.; Mirzaei, G.; Petreaca, R.C.; Bouley, R.A. A Genome-Wide Profiling of Glioma Patients with an IDH1 Mutation Using the Catalogue of Somatic Mutations in Cancer Database. Cancers (Basel) 2021, 13, 4299. [Google Scholar] [CrossRef]
- Yin, X.; Yu, H.; He, X.K.; Yan, S.X. Prognostic and biological role of the N-Myc downstream-regulated gene family in hepatocellular carcinoma. World J Clin Cases 2022, 10, 2072–2086. [Google Scholar] [CrossRef]
- Liu, Y.; Xia, J.; Zheng, R.; Shao, S. High expression of NDRG3 suppresses cell apoptosis and promotes the cell proliferation and migration in gastric cancer. Asian J Surg 2022, 45, 2019–2020. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, J.; Quan, J.; Liu, J.; Tian, L.; Dong, C. Relationship between serum NDRG3 and papillary thyroid carcinoma. Front Endocrinol (Lausanne) 2022, 13, 1091462. [Google Scholar] [CrossRef]
- Zhang, H.; Ge, Z.; Wang, Z.; Gao, Y.; Wang, Y.; Qu, X. Circular RNA RHOT1 promotes progression and inhibits ferroptosis via mir-106a-5p/STAT3 axis in breast cancer. Aging (Albany NY) 2021, 13, 8115–8126. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Yao, L.; Wei, Y.; Geng, S.; He, C.; Jiang, H. Role of RHOT1 on migration and proliferation of pancreatic cancer. Am J Cancer Res 2015, 5, 1460–70. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; He, C.; Geng, S.; Sheng, H.; Shen, X.; Zhang, X.; Li, H.; Zhu, S.; Chen, X.; Yang, C.; Gao, H. RhoT1 and Smad4 are correlated with lymph node metastasis and overall survival in pancreatic cancer. PLoS One. 2012, 7, e42234. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Cheng, F.; Zhang, Z.; Wang, B.; Zhang, X. Propofol suppresses non-small cell lung cancer tumorigenesis by regulation of circ-RHOT1/miR-326/FOXM1 axis. Life Sci. 2021, 119042. [Google Scholar] [CrossRef]
- Wang, L.; Long, H.; Zheng, Q.; Bo, X.; Xiao, X.; Li, B. Circular RNA circRHOT1 promotes hepatocellular carcinoma progression by initiation of NR2F6 expression. Mol Cancer 2019, 18, 119. [Google Scholar] [CrossRef]
- Periñán, M.T.; Gómez-Garre, P.; Blauwendraat, C.; Mir, P.; Bandres-Ciga, S.; International Parkinson’s Disease Genomics Consortium (IPDGC). The role of RHOT1 and RHOT2 genetic variation on Parkinson disease risk and onset. Neurobiol Aging 2021, 97, 144–e1. [Google Scholar] [CrossRef]
- Sun, X.; Luo, Zh.; Gong, L.; Tan, X.; Chen, J.; Liang, X.; Cai, M. Identification of significant genes and therapeutic agents for breast cancer by integrated genomics. Bioengineered 2021, 12, 2140–2154. [Google Scholar] [CrossRef]
- Xing, Q.; Liu, S.; Luan, J.; Wang, Y.; Ma, L. A novel 13 RNA binding proteins (RBPs) signature could predict prostate cancer biochemical recurrence. Pathol Res Pract, 1: 225, 1535. [Google Scholar] [CrossRef]
- Henning, L.M.; Santos, K.F.; Sticht, J.; Jehle, S.; Lee, C.T.; Wittwer, M.; Urlaub, H.; Stelzl, U.; Wahl, M.C.; Freund, C. A new role for FBP21 as regulator of Brr2 helicase activity. Nucleic Acids Res 2017, 45, 7922–7937. [Google Scholar] [CrossRef]
- Henning, L.M.; Bhatia, S.; Bertazzon, M.; Marczynke, M.; Seitz, O.; Volkmer, R.; Haag, R.; Freund, C. Exploring monovalent and multivalent peptides for the inhibition of FBP21-tWW. Beilstein, J. Org Chem. 2015, 11, 701–706. [Google Scholar] [CrossRef] [PubMed]
- Jia, Q.; Song, J.; Xu, T.; Liu, J.; Chai, J.; Yang, Y.; Li, L.; Li, M.; Yang, X. ZIC5 promotes aggressiveness and cancer stemness in cervical squamous cell carcinoma. Pathol Res Pract 2023, 241, 154268. [Google Scholar] [CrossRef] [PubMed]
- Satow, R.; Watanabe, T.; Nomura, M.; Inagaki, S.; Yoneda, A.; Fukami, K. Patulin and LL-Z1640-2 induce apoptosis of cancer cells by decreasing endogenous protein levels of Zic family member 5. J Cell Mol Med 2022, 26, 5680–5689. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.F.; Zhang, Y.; Ge, S.Y.; Zhong, F.; Sun, C.Y.; Xia, G.W. AR-regulated ZIC5 contributes to the aggressiveness of prostate cancer. Cell Death Discov 2022, 8, 393. [Google Scholar] [CrossRef]
- Song, W.; Yu, W.; Li, D.; Cheng, C.; Wu, X.; Chen, J.; Zhang, W. ZIC5 promotes human hepatocellular carcinoma cell proliferation through upregulating COL1A1. J Gastrointest Oncol 2022, 13, 1237–1247. [Google Scholar] [CrossRef]
- Chang, C.C.; Kuo, H.Y.; Chen, S.Y.; Lin, W.T.; Lu, K.M.; Saito, T.; Liu, F.C. Developmental Characterization of Schizophrenia-Associated Gene Zswim6 in Mouse Forebrain. Front Neuroanat 2021, 15, 669631. [Google Scholar] [CrossRef]
- Yanagishita, T.; Eto, K.; Yamamoto-Shimojima, K.; Segawa, O.; Nagata, M.; Ishihara, Y.; Miyashita, Y.; Asano, Y.; Sakata, Y.; Nagata, S.; Yamamoto, T. A recurrent de novo ZSWIM6 variant in a Japanese patient with severe neurodevelopmental delay and frequent vomiting. Hum Genome Var 2021, 8, 16. [Google Scholar] [CrossRef]
- Tischfield, D.J.; Saraswat, D.K.; Furash, A.; Fowler, S.C.; Fuccillo, M.V.; Anderson, S.A. Loss of the neurodevelopmental gene Zswim6 alters striatal morphology and motor regulation. Neurobiol Dis 2017, 103, 174–183. [Google Scholar] [CrossRef]
- Bogdanova, E.; Sadykov, A.; Ivanova, G.; Zubina, I.; Beresneva, O.; Semenova, N.; Galkina, O.; Parastaeva, M.; Sharoyko, V.; Dobronravov, V. Mild Chronic Kidney Disease Associated with Low Bone Formation and Decrease in Phosphate Transporters and Signaling Pathways Gene Expression. Int J Mol Sci 2023, 24, 7270. [Google Scholar] [CrossRef]
- Biscetti, F.; Rando, M.M.; Cecchini, A.L.; Nicolazzi, M.A.; Rossini, E.; Angelini, F.; Iezzi, R.; Eraso, L.H.; Dimuzio, P.J.; Pitocco, D.; Gasbarrini, A.; Massetti, M.; Flex, A. The role of Klotho and FGF23 in cardiovascular outcomes of diabetic patients with chronic limb threatening ischemia: a prospective study. Sci Rep 2023, 13, 6150. [Google Scholar] [CrossRef]
- Kubota, M.; Hamasaki, Y.; Hashimoto, J.; Aoki, Y.; Kawamura, T.; Saito, A.; Yuasa, R.; Muramatsu, M.; Komaba, H.; Toyoda, M.; Fukagawa, M.; Shishido, S.; Sakai, K. Fibroblast growth factor 23-Klotho and mineral metabolism in the first year after pediatric kidney transplantation: A single-center prospective study. Pediatr Transplant 2023, 27, e14440. [Google Scholar] [CrossRef] [PubMed]
- Balcázar-Hernández, L.; Manuel-Apolinar, L.; Vargas Ortega, G.; González-Virla, B.; Reza-Albarrán, A.A.; Jiménez Martínez, M.D.C.; Martínez Ordaz, J.L.; Mendoza-Zubieta, V.; Basurto, L. Vitamin D and its positive effect on the PTH/vitamin D/calcium-FGF23/klotho/phosphorus axis in kidney transplant recipients. Nutr Hosp 2023, 40, 428–435. [Google Scholar] [CrossRef]
- Nakano, T.; Kishimoto, H.; Tokumoto, M. Direct and indirect effects of fibroblast growth factor 23 on the heart. Front Endocrinol (Lausanne) 2023, 14, 1059179. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, J.; Kalantar-Zadeh, K.; Chen, J. Focusing on Phosphorus Loads: From Healthy People to Chronic Kidney Disease. Nutrients 2023, 15, 1236. [Google Scholar] [CrossRef]
- Karava, V.; Dotis, J.; Kondou, A.; Christoforidis, A.; Taparkou, A.; Farmaki, E.; Economou, M.; Printza, N. Fibroblast growth-factor 23 and vitamin D are associated with iron deficiency and anemia in children with chronic kidney disease. Pediatr Nephrol 2023. [Google Scholar] [CrossRef]
- Kaplan, J.; Tommasini, S.; Yao, G.Q.; Zhu, M.; Nishimura, S.; Ghazarian, S.; Louvi, A.; Insogna, K. Altered Expression of Several Molecular Mediators of Cerebrospinal Fluid Production in Hyp Mice. J Endocr Soc 2023, 7, bvad022. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.P.; Sidibé, A.; Fortier, C.; Desjardins, M.P.; Ung, R.V.; Kremer, R.; Agharazii, M.; Mac-Way, F. Wnt/β-catenin pathway inhibitors, bone metabolism and vascular health in kidney transplant patients. J Nephrol 2023. [Google Scholar] [CrossRef] [PubMed]
- Bishop, N.C.; Burton, J.O.; Graham-Brown, M.P.M.; Stensel, D.J.; Viana, J.L.; Watson, E.L. Exercise and chronic kidney disease: potential mechanisms underlying the physiological benefits. Nat Rev Nephrol 2023, 19, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, Y.; Ishida, T.; Kugimiya, F.; Takai, S.; Nakayama, Y.; Yonemitsu, K.; Harada, E. Deterioration of Phosphate Homeostasis Is a Trigger for Cardiac Afterload- Clinical Importance of Fibroblast Growth Factor 23 for Accelerated Aging. Circ Rep. 2022, 5, 4–12. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schemes of interaction of piRNA and miRNA with mRNA of the KLOTHO gene. Note: The mRNA nucleotides are highlighted in red. The piRNA nucleotides that form canonical pairs with gRNA are highlighted in violet and noncanonical pairs are highlighted in green.
Figure 1.
Schemes of interaction of piRNA and miRNA with mRNA of the KLOTHO gene. Note: The mRNA nucleotides are highlighted in red. The piRNA nucleotides that form canonical pairs with gRNA are highlighted in violet and noncanonical pairs are highlighted in green.

Figure 2.
Schemes of interaction of piR-3215034 and piR-6885965 with the mRNA AFF2 gene. Note: The mRNA nucleotides are highlighted in red. The piRNA nucleotides that form canonical pairs with gRNA are highlighted in violet and noncanonical pairs are highlighted in green.
Figure 2.
Schemes of interaction of piR-3215034 and piR-6885965 with the mRNA AFF2 gene. Note: The mRNA nucleotides are highlighted in red. The piRNA nucleotides that form canonical pairs with gRNA are highlighted in violet and noncanonical pairs are highlighted in green.

Figure 3.
Schemes of interaction of piR-3215034 with the mRNA of the BCL2L11 gene. Note: The mRNA nucleotides are highlighted in red. The piRNA nucleotides that form canonical pairs with gRNA are highlighted in violet and noncanonical pairs are highlighted in green.
Figure 3.
Schemes of interaction of piR-3215034 with the mRNA of the BCL2L11 gene. Note: The mRNA nucleotides are highlighted in red. The piRNA nucleotides that form canonical pairs with gRNA are highlighted in violet and noncanonical pairs are highlighted in green.

Figure 4.
Interaction schemes of piR-3215034 and piR-6885965 with mRNA of the CPT1A gene. Note: The mRNA nucleotides are highlighted in red. The piRNA nucleotides that form canonical pairs with gRNA are highlighted in violet and noncanonical pairs are highlighted in green.
Figure 4.
Interaction schemes of piR-3215034 and piR-6885965 with mRNA of the CPT1A gene. Note: The mRNA nucleotides are highlighted in red. The piRNA nucleotides that form canonical pairs with gRNA are highlighted in violet and noncanonical pairs are highlighted in green.

Figure 5.
Schemes of interaction of piR-3215034 and piR-6885965 with mRNA of the DAZAP1 gene. Note: The mRNA nucleotides are highlighted in red. The piRNA nucleotides that form canonical pairs with gRNA are highlighted in violet and noncanonical pairs are highlighted in green.
Figure 5.
Schemes of interaction of piR-3215034 and piR-6885965 with mRNA of the DAZAP1 gene. Note: The mRNA nucleotides are highlighted in red. The piRNA nucleotides that form canonical pairs with gRNA are highlighted in violet and noncanonical pairs are highlighted in green.

Figure 6.
Schemes of interaction of piR-3215034 and piR-6885965 with mRNA of the NDRG3 gene. Note: The mRNA nucleotides are highlighted in red. The piRNA nucleotides that form canonical pairs with gRNA are highlighted in violet and noncanonical pairs are highlighted in green.
Figure 6.
Schemes of interaction of piR-3215034 and piR-6885965 with mRNA of the NDRG3 gene. Note: The mRNA nucleotides are highlighted in red. The piRNA nucleotides that form canonical pairs with gRNA are highlighted in violet and noncanonical pairs are highlighted in green.

Figure 7.
Interaction schemes of piR-3215034 and piR-6885965 with mRNA of the RHOT1 gene. Note: The mRNA nucleotides are highlighted in red. The piRNA nucleotides that form canonical pairs with gRNA are highlighted in violet and noncanonical pairs are highlighted in green.
Figure 7.
Interaction schemes of piR-3215034 and piR-6885965 with mRNA of the RHOT1 gene. Note: The mRNA nucleotides are highlighted in red. The piRNA nucleotides that form canonical pairs with gRNA are highlighted in violet and noncanonical pairs are highlighted in green.
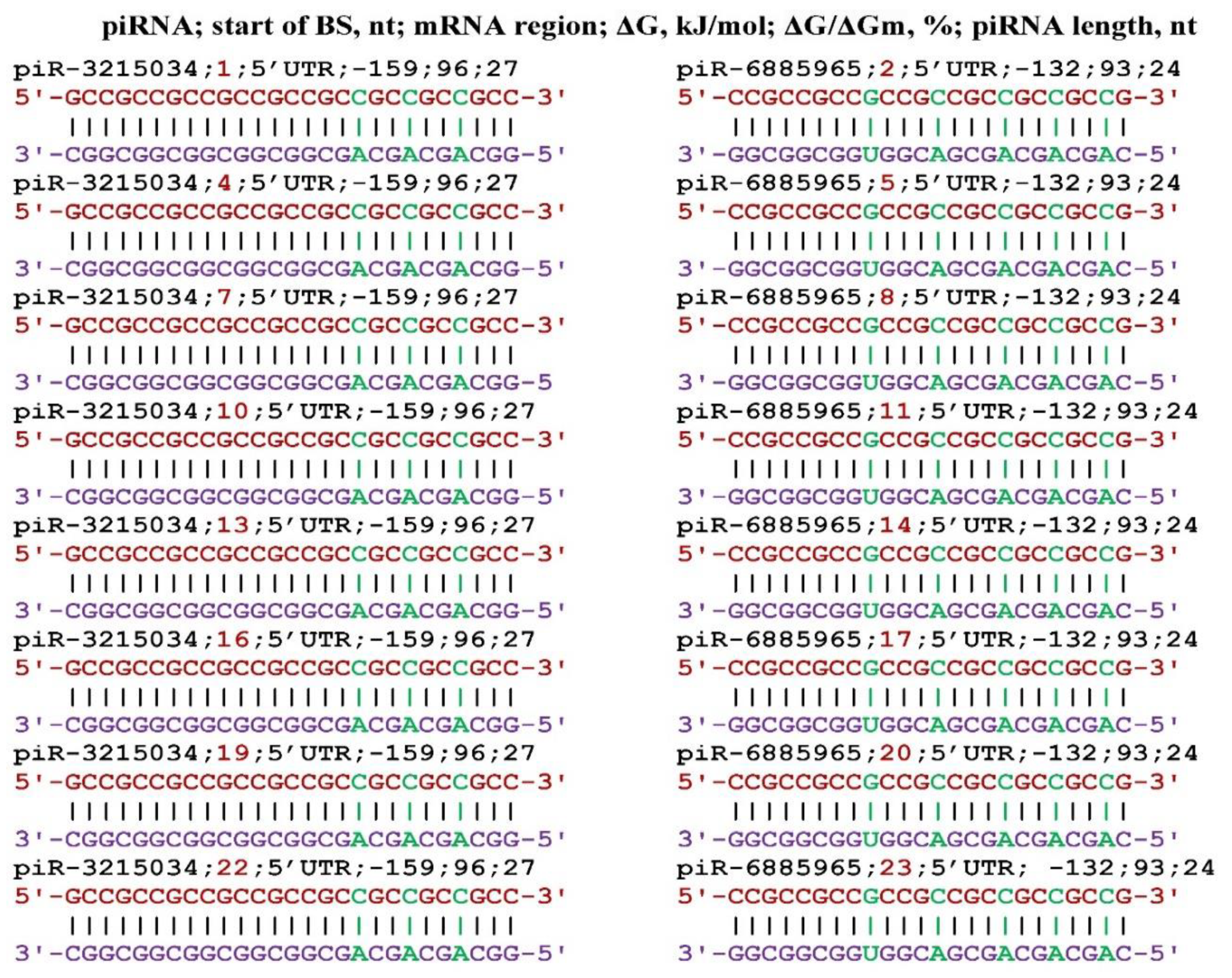
Figure 8.
Schemes of interaction of piR-3215034 with the mRNA of the SKIDA1 gene. Note: The mRNA nucleotides are highlighted in red. The piRNA nucleotides that form canonical pairs with gRNA are highlighted in violet and noncanonical pairs are highlighted in green.
Figure 8.
Schemes of interaction of piR-3215034 with the mRNA of the SKIDA1 gene. Note: The mRNA nucleotides are highlighted in red. The piRNA nucleotides that form canonical pairs with gRNA are highlighted in violet and noncanonical pairs are highlighted in green.

Figure 9.
Schemes of the interaction of piR-3215034 and piR-6885965 with mRNA of the WBP4 gene. Note: The mRNA nucleotides are highlighted in red. The piRNA nucleotides that form canonical pairs with gRNA are highlighted in violet and noncanonical pairs are highlighted in green.
Figure 9.
Schemes of the interaction of piR-3215034 and piR-6885965 with mRNA of the WBP4 gene. Note: The mRNA nucleotides are highlighted in red. The piRNA nucleotides that form canonical pairs with gRNA are highlighted in violet and noncanonical pairs are highlighted in green.

Figure 10.
Schemes of interaction of piR-3215034 and piR-6885965 with the mRNA of the ZIC5 gene. Note: The mRNA nucleotides are highlighted in red. The piRNA nucleotides that form canonical pairs with gRNA are highlighted in violet and noncanonical pairs are highlighted in green.
Figure 10.
Schemes of interaction of piR-3215034 and piR-6885965 with the mRNA of the ZIC5 gene. Note: The mRNA nucleotides are highlighted in red. The piRNA nucleotides that form canonical pairs with gRNA are highlighted in violet and noncanonical pairs are highlighted in green.

Figure 11.
Schemes of interaction of piR-3215034 and piR-6885965 with the mRNA of the ZSWIM6 gene. Note: The mRNA nucleotides are highlighted in red. The piRNA nucleotides that form canonical pairs with gRNA are highlighted in violet and noncanonical pairs are highlighted in green.
Figure 11.
Schemes of interaction of piR-3215034 and piR-6885965 with the mRNA of the ZSWIM6 gene. Note: The mRNA nucleotides are highlighted in red. The piRNA nucleotides that form canonical pairs with gRNA are highlighted in violet and noncanonical pairs are highlighted in green.

Figure 12.
Schemes of piRNA interaction with the mRNA of the FGF23 gene. Note: The mRNA of nucleotides are highlighted in red. The piRNA nucleotides that form canonical pairs with gRNA are highlighted in violet and noncanonical pairs are highlighted in green.
Figure 12.
Schemes of piRNA interaction with the mRNA of the FGF23 gene. Note: The mRNA of nucleotides are highlighted in red. The piRNA nucleotides that form canonical pairs with gRNA are highlighted in violet and noncanonical pairs are highlighted in green.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



