Submitted:
14 June 2023
Posted:
15 June 2023
You are already at the latest version
Abstract
Diets that restrict the amount of carbohydrates positively affect markers related to metabolic syndrome (MS). However, completely carbohydrate-free diets have been studied in lesser extent, being their effect on biochemical and morphometric parameters in a rat model of MS the main objective of this study. MS was induced by a high-glucose diet for 20 weeks in male Wistar rats with a subsequent exposition to an essentially carbohydrate-free diet (CFD) for 20 weeks. Daily measurements of food consumption and monthly measurements of body weight were made. At week 40, glucose, insulin, lipid profile, ketone bodies, C-reactive protein (CRP), aspartate aminotransferase (AST), alanine aminotransferase (ALT), urea, creatinine, liver and muscle glycogen, and serum, hepatic, renal and pancreatic malondialdehyde (MDA) were assessed. Exposition to CFD showed decrease in caloric intake and body weight. Likewise, the group showed normalization in liver and serum MDA parameters, insulin, lipid profile, ALT, liver glycogen, and creatinine. The CFD decreased glucose concentration and the inflammatory state, also, increased the concentration of ketone bodies and urea. The change towards CFD reverses the metabolic alterations generated with MS except for glycemia and uremia, probably due to the high rate of gluconeogenesis generated by the high protein content in the diet.
Keywords:
carbohydrate-free diet
; metabolic syndrome
; biochemical parameters
; morphometric parameters
; rat model.
1. Introduction
Metabolic syndrome (MS) is a global health problem characterized by a cluster of metabolic abnormalities that include abdominal obesity, systemic hypertension, insulin resistance, and atherogenic dyslipidemia, conferring a higher risk of developing cardiovascular disease (CVD) and type 2 diabetes mellitus (T2DM) [1,2]. Studies published in recent years estimate that from 20 to 30% of the world’s adult population meets the criteria for MS [3]. According to the Center of Disease Control and Prevention (CDC), the prevalence of MS has increased drastically in the last four decades [4,5].
The pathogenesis of MS involves both genetic and acquired factors [6,7]. The rise in the prevalence of MS is largely attributed to the combination of excessive caloric intake and the lack physical activity [8,9]. It has been demonstrated that an increased consumption of carbohydrates is closely associated with the growing prevalence of MS [5]. Access to diets with high fat and carbohydrate contents have contributed to the onset and progression of MS [10,11].
Dietary modifications, particularly carbohydrate restriction has proven to be a more effective therapeutic tool in improving MS than diets that restrict the amount of fat [12,13]. The replacement of carbohydrates with saturated or unsaturated fats (monounsaturated and polyunsaturated fats) and protein has been evidenced to have important effects on glycemic control, weight loss, and dyslipidemia [14].
Carbohydrate-free diets have been evaluated in models with healthy genetically modified animals susceptible to the development of T2DM. Finding an increase in energy expenditures and gluconeogenesis [15], a decrease in body weight and energy intake [13,16], and avoiding the destruction of pancreatic β-cells [17,18]. However, these models lack the complexity of the metabolic abnormalities that occur in MS, showing limited and isolated results. Additionally, the studies evaluating the effects of carbohydrate-free diets have generally been of short duration, and the long-term effects are unclear.
Therefore, the objective of this investigation is to evaluate the effects of a chronic exposure to a carbohydrate-free diet on morphometric and biochemical parameters in a rat model of diet-induced MS, evaluating the characteristic alterations of MS.
2. Materials and Methods
2.1. Animals and Diets
Fifty-three, male Wistar rats weaned at 21 days of age with an average body weight of 61.6 grams were used. The animals, provided by the biotery “Claude Bernard” of the Autonomous University of Zacatecas, were housed in polycarbonate cages with 12-hour day and night lighting cycles, a controlled temperature of 22°C, and 50% humidity.
The animals were randomly divided into two study groups according to the type of diet supplied, the standard diet (SD) group (n=15), fed a balanced commercial diet for rodents (Teklad Lab Diet, Harlan Sprague Dawley Inc., IN, USA; 13% kcal from fat, 34% kcal from protein and 53% kcal from carbohydrates) and purified water ad libitum, and the MS group (n=38), fed a formulated diet (16% kcal from fat, 33% kcal from protein and 51% kcal from carbohydrates) and 25% glucose solution ad libitum for 20 weeks. After the period of exposure to the diets, animals from both groups were randomly selected, the MS state was confirmed by measurement of fasting blood glucose, insulin, lipids profile and C-reactive protein, following the specifications marked later.
MS group’s rats were then divided into two groups, the MS group (n=11) continued feeding with the formulated diet and 25% glucose solution ad libitum for 20 weeks and the MS, subsequent carbohydrate-free diet (MS+CFD) group (n=12) fed a formulated essentially carbohydrate-free diet (52.89% kcal from fat, 47.004% kcal from protein, and 0.105% kcal from carbohydrates) and purified water ad libitum for 20 weeks. The SD group (n=10) continued feeding with the commercial diet and purified water ad libitum for 20 weeks.
The formulated diet consisted of the following ingredients (g/kg): fish flour, 250; bentonite, 50; lard, 3.2; rice, 153.3; oatmeal, 340; corn, 100; coconut oil, 3.5; wheat, 100. The essentially carbohydrate-free diet consisted of (g/kg): fish flour, 450; casein, 200; rice husk, 100; bentonite, 50; lard, 200.
During the experimental period, daily measurements of food and purified water or glucose solution intake were made, caloric intake was calculated using the Atwater factor. Monthly measurements of body weight were also performed.
2.2. Samples Collection
Animals’ blood fasted for 12 hours was obtained by cardiac puncture in the ventricular cavity after previous anesthesia. Blood was centrifugated, and serum samples were stored at -20°C until analysis. The liver, kidney, pancreas, and skeletal muscle were dissected and rinsed in phosphate buffer solution pH 7.
2.3. Determination of Biochemical Parameters
Serum concentrations of glucose, total cholesterol (TC), triglycerides (TRIG), high-density lipoprotein cholesterol (HDL-c), low-density lipoprotein cholesterol (LDL-c), alanine aminotransferase (ALT), aspartate aminotransferase (AST), urea and creatinine were determined by colorimetric enzymatic assays (Spinreact, Girona, Spain). Very low-density lipoprotein cholesterol (VLDL-c) concentration was calculated according to the equation 1:
VLDL-c = TRIG/5,
C-reactive protein (CRP) concentration was determined by a semiquantitative plate agglutination assay (Spinreact, Girona, Spain).
Insulin determination was performed using an ELISA kit, specifically for rats (Spibio, Bertin Pharma, Montigny-le-Bretonneux, France).
β-hydroxybutyrate quantification was determined using reactive strips and a FreeStyle Optium Neo electronic meter (Abbott, Mexico City, Mexico).
2.4. Determination of Malondialdehyde
Damage to lipid membranes in blood serum, liver, kidney, and pancreas was assessed by the formation of malondialdehyde (MDA), using the thiobarbituric acid (TBA) reactive method [19]. Tissues were homogenized at 10% KCl (1.15%) and blood serum in H2SO4 (12 N); the addition of TBA (0.3%) created colorful compounds that were measured photometrically at 534 nm wavelength in a 6715 UV/Vis Jenway spectrophotometer (Cole-Parmer, IL, USA). The MDA level in the samples (expressed as nM/mL in serum and nM/g in tissues) was determined using a calibration curve with 1,1,3,3-tetraethoxypropane (Sigma Chemical, St. Louis, MO, USA).
2.5. Determination of Glycogen
The anthrone reagent method determined glycogen levels in the liver and skeletal muscle [20]. An hydrolyzate of liver and skeletal muscle was made in KOH (30%) in a 1:3 (w/v) ratio; the addition of anthrone solution allowed the coloration of the sample, which was measured photometrically at 620 nm with a 6715 UV/Vis Jenway spectrophotometer. Glycogen concentration was determined using glucose (50%) calibration curve (expressed as µg/mL).
2.6. Statistical Analysis
Experimental results are shown as mean ± SD or median (minimum-maximum). Statistical analysis was performed with SPSS IBM23 (IBM Corp., IL, USA) and GraphPad Prism 5 (GraphPad Software, San Diego, CA, USA) packages. For parametric data, analysis of variance (ANOVA) followed by Tukey’s post hoc analysis was performed. Data not normally distributed were evaluated using the Kruskal-Wallis’ test and Dunn’s test for post hoc multiple comparisons. Differences with a value of p < 0.05 were considered statistically significant.
2.7. Ethical Considerations
The handling and care of the animals, as well as their euthanasia, were carried out in accordance with the regulations and the official standards in force, such as the Official Mexican Norm “NOM-062-ZOO-1999,” which shows the specifications and techniques for the production, care, and use of institutional laboratory animals. Animals’ remains were treated as indicated by the “NOM-087-ECOL-SSA1-2002” for Environmental Protection - Environmental Health – Biologically Infectious Hazardous Waste - Classification and Management Specifications.
3. Results
3.1. Morphometric Parameters and Calorie Intake
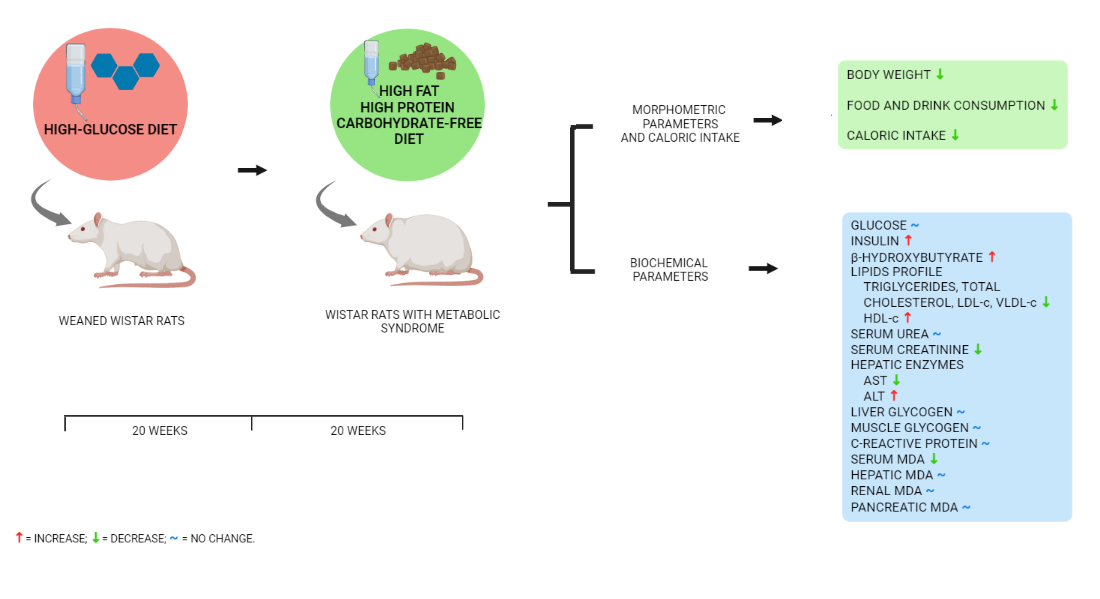
Data for food, drink, caloric intake, and body weight are shown in Figure 1. During the first 20 weeks of dietary exposure, the MS group showed a significant decrease in food and drink consumption concerning the SD group. However, the caloric intake in the MS group since week 20 significantly increased compared to the SD and MS+CFD groups; consequently, the body weight increased in the MS group.
The change to CFD in week 20 in the MS rats’ significant decrements caloric intake due to lower consumption of solid food and drink compared to the SD and MS groups. The CFD also induced a significant loss of body weight.
In the first 20 weeks of experimentation, the SD and MS groups increased their body weight, the SD group showing the highest body weight; from 20 weeks to the end of the examination period, the MS+CFD and MS groups decreased and increased body weight, respectively, being statistically significant concerning to the SD group.
During euthanasia, animals in the MS group exposed to the high-glucose diet for 20 weeks showed a large accumulation of visceral fat relative to the SD group, even though the latter group showed higher body weight.
3.2. Biochemical Parameters
3.2.1. Glucose, Insulin and β-hydroxybutyrate
The glucose concentration and insulin level comparisons between the experimentation groups are shown in Figure 2. At week 20 of exposure to the diets, the MS group showed a statistically significant increase in glucose concentration (p = 0.016) and a decrease in insulin concentration (p = 0.008) compared to the SD group, on the other hand, there were no significant differences between the groups in the β-hydroxybutyrate concentrations.
At week 40 of experimentation, the MS diet produced a statistically significant increase in glucose concentration and a decrease in insulin level compared with the SD group (p = 0.0001, p = 0.023, respectively). When carbohydrates were reduced from the diet (MS+CFD group), a non-significant reduction in glucose level and an increase in insulin concentration were observed compared to the MS group (p = 0.000). The comparison between SD and MS+CFD groups did not show a significant difference in insulin concentration, but there was an increase in glucose concentration (p = 0.004). The MS+CFD group presented the highest levels of β-hydroxybutyrate related to the other groups, this difference was only significant in relation to the MS group (p = 0.001) (Figure 2).
3.2.2. Lipids Profile
At week 20 of diet exposure, the MS group showed significant differences in all lipid profile parameters (TRIG, TC, HDL-c, LDL-c, and VLDL-c) compared to the SD group (p = 0.009, p = 0.009, p = 0.027, p = 0.009, and p = 0.009, respectively), as shown in Figure 3.
At week 40 of experimentation, the MS and the SD groups showed a similar behavior than in the previous evaluation, showing significant differences between both in TRIG, TC, HDL-c, LDL-c, and VLDL-c (p = 0.007, p = 0.000, p = 0.000, p = 0.000, and p = 0.007, respectively), as shown in Figure 3. The MS+CFD group presented a normalization of the lipid profile parameters, showing no significant differences with respect to the SD group.
3.2.3. C-Reactive Protein
Regarding to the CRP concentrations, at week 20 of diet exposure, the MS group showed a significant increase related to the SD group (p = 0.005). At week 40 of experimentation, the MS and MS+CFD groups showed a significant increase compared to the SD group (p = 0.000; p = 0.024, respectively). However, the MS+CFD group showed a non-significant 50% decrease relative to the MS group (Figure 4).
3.2.4. Serum Urea and Creatinine
The MS+CFD group increased the urea concentration by 42.6% in relation to the SD group, this being statistically significant (p = 0.004). The MS group increased by 36.5% in relation to the SD group, showing no significant differences in relation to the MS+CFD group. The MS group showed an increase in creatinine level by 66.3% compared to the SD group and by 112.1% in relation to the MS+CFD group, this difference being significant only in relation to the MS+CFD group (p = 0.000) (Figure 5).
3.2.5. Hepatic Enzymes and Glycogen
The MS+CFD group showed a significant decrease in AST concentration by 47.05% in relation to the SD group and by 40.7% in relation to the MS group, these differences being statistically significant (Table 1). Regarding to ALT concentration, the MS group showed the lowest levels compared to the SD and MS+CFD groups, these differences being statistically significant (Table 1).
The MS group presented the highest concentrations of liver glycogen compared to the SD and the MS+CFD groups, being the difference with respect to the SD group statistically significant; while the MS+CFD group showed a non-significant decrease of 51% compared to the MS group. At the skeletal muscle level, no significant differences were observed between the three groups of study; however, the MS group showed the highest concentrations of muscle glycogen in relation to the SD and MS+CFD groups (Table 1), the SD and MS+CFD groups showed concentrations medians very similar.
3.2.6. Malondialdehyde
The Figure 6 (A) shows the behavior of the serum MDA concentration in the MS group, presenting a statistically significant increase in relation to the SD and MS+CFD groups (p = 0.013 and p = 0.005, respectively) with increases of 40.9 and 39.8% respectively.
At hepatic level, the MS group presented a significant increase by 100.4% in relation to the SD group (p = 0.041), and the MS+CFD group showed a non-significant 30.3% increase in relation to the SD group and a non-significant 35% decrease relative to the MS group (Figure 6, B).
At renal level, no significant differences in MDA concentrations were observed between the three study groups, however, the MS group showed a 26.6% increase in relation to the SD group; the MS+CFD group presented a 41.8% increase relative to the SD group (Figure 6, C).
At pancreatic level, the MS and MS+CFD groups showed a non-significant decrease in MDA concentrations by 35.5% and 20.9%, respectively, in relation to the SD group (Figure 6, D).
4. Discussion
The improvement of the pathophysiology of MS is based on the establishment of pharmacological treatments and lifestyle changes. It is proposed that a healthy eating pattern significantly reduces and controls the risk of developing MS, nevertheless, the adequate nutritional intervention still is controversial. The reduction in the intake of carbohydrates in the diet is one of the main recommendations to reduce body weight, glycemia, and dyslipidemia [21]. However, diets that entirely restrict carbohydrate intake have not been evaluated in an animal model of MS, where the parameters that make up this condition are evaluated jointly.
The diagnostic criteria of MS have not been clearly established for animal models, however, according to the review by Preguiça et al. (2020), the metabolic parameters used to determine the presence of this condition in rodent models can be three of the following five: increased fasting glucose, obesity, increased triglyceride concentration, decreased HDL-c concentration, and increased systolic blood pressure. Exposure to the high-glucose diet for 20 weeks produced significant increases in fasting glucose, triglycerides, HDL-c, and CRP concentrations, as well as a decrease in insulin concentration and accumulation in visceral fat deposits. Previous reports in which diets were used for the induction of MS with and energy density similar to that used in this study demonstrated changes in glucose homeostasis and insulin, as well as oxidative imbalance, low-grade inflammation, and plasma dyslipidemia [22,23].
Increase in visceral fat deposition in diet-induced metabolic syndrome animal models is a determining factor in the establishment of obesity in rodents as it is related to the development and progression of MS [24]. At week 20 the animals in the MS group showed a considerable increase in visceral fat deposits in relation to SD group, coinciding with publications in which exposure to high-carbohydrate diets led to the accumulation of visceral fat [25,26].
Excess in calorie intake is an important factor in the development of metabolic complications related to MS [27]. The decrease in caloric intake observed after exposure to CFD with high protein content is consistent with previous reports in animals. These have been related to a state of satiety and increased energy expenditure produced by high-protein diets [13,28,29]. In this sense, the decrease in caloric intake can be related to the decrease in body weight, possibly, also, through an increase in energy expenditure [30]. Previous studies directly correlate the carbohydrate content in the diet with the increase in body weight and adiposity since carbohydrates are the main secretagogue of insulin, stimulating fat storage [29]. Therefore, the partial elimination of the carbohydrates in the diet promotes the oxidation of lipids at the expense of their deposits, reflected as a reduction in body weight as was observed in the MS+CFD group in relation to the MS group, fed with a high-glucose diet.
According to some reviews about animal models of diet-induced MS, significant increases in the animal's body weight are not always observed in comparison to control groups, even when they do present important metabolic alterations [31,32]. Most of the investigations reported a period of exposure to diets between 6 and 20 weeks, in this study, significant increases in body weight can be observed in animals with MS from week 20 of experimentation compared to the SD group. This verifies the role of chronic exposure to a high-glucose diet in body weight gain in animal models.
Insulin resistance is one of the main underlying links between excessive carbohydrate consumption and the development of MS [33].
Insulin sensitivity assessment was determined by measuring fasting glucose and insulin at the 40th week of dietary exposure. In the MS group, the high-glucose diet caused deregulation of the carbohydrate metabolism, especially the generation of both hepatic and systemic insulin resistance, observed as a state of hyperglycemia by not suppressing glucose production in the liver. This same group showed a significant decrease in insulin concentration compared to the SD group. Although hyperinsulinemia is considered a significant predictor of the development of MS, the loss of the ability to secrete insulin is strong evidence of pancreatic β-cell deterioration to external stimuli, such as high glucose concentrations for prolonged periods [34].
The CFD (containing high fats and proteins) caused a non-significant decrease in glucose concentration relative to the MS group without reaching a normalization as the SD group. The CFD produced pronounced changes in carbohydrate metabolism due to the strict requirement of glucose by the central nervous system and other tissues, leading to the activation of hepatic gluconeogenesis, coinciding with the results previously reported by other authors [35,36]. The high energy dependence of the gluconeogenesis pathway requires a constant flow of ATP provided by the increase oxidation of fatty acids, thus explaining the weight loss that promotes the increase in this pathway [37]. Likewise, the insulin concentration in the MS+CFD group showed a normalization, observing comparable results to the SD group. There are reports about the role of carbohydrate-restricted diets in reducing oxidative stress and proinflammatory conditions and its positive impact on pancreatic β-cell integrity in early stages of T2DM in animal models [38].
Ketone bodies can substitute for glucose in conditions of fuel and food deficiency. β-hydroxybutyrate is the ketone body found in the highest concentration in blood. It has a fundamental role in the temporary inhibition of lipolysis, inflammation, satiety, and states of atherosclerosis and oxidative stress [39]. The reduction of carbohydrates from the diet promoted the activation of the ketogenesis pathway from free fatty acids as an energy source. It has been reported that a high concentration of β-hydroxybutyrate promotes satiety when a high-protein diet is implemented [28]. Similarly, β-hydroxybutyrate treatment in mice exposed to a high-fat diet has shown to maintain the animals metabolically healthy by influencing pathways related to glucose homeostasis, mitochondrial function, and obesity [40].
Dyslipidemia is an essential alteration of MS and the leading cause of the development of cardiovascular disease. Visceral obesity and insulin resistance have been associated with abnormalities in the lipid profile [41]. MS group exposed to the high-glucose diet showed a state of dyslipidemia with a marked alteration in the concentrations of triglycerides and HDL-c. In contrast, the MS+CFD group presented a normalization of the lipid profile parameters, despite of the increase in the amount of fat in the CFD.
According to Forouhi et al., (2018) lipid markers improve when saturated, monounsaturated, or polyunsaturated fats replace carbohydrates [42]. The role of fats in the diet has an erroneous assumption regarding weight gain, based on the relative energy density of the macronutrients: 9 kcal/g of fat compared to 4 kcal/g of protein or carbohydrates. However, it is common to ignore the role of macronutrients in states of hunger and satiety and in the pathways that regulate energy generation, fat storage, and fatty acid metabolism [43]. The observed results imply that stored and dietary fatty acids were used as the primary fuel in energy generation (gluconeogenesis and ketogenesis) in the absence of carbohydrates [18], having a lower proportion of lipids stored, also reflected in the body weight loss of the animals in the MS+CFD group.
The characteristic visceral obesity of MS causes a state of hypoxia in the adipocytes, generating necrosis and infiltration of macrophages in the adipose tissue, which leads to an overproduction of proinflammatory cytokines, like tumor necrosis factor-alpha (TNF-α) interleukin-6 (IL-6), and CRP, resulting in general systemic inflammation. CRP is an acute phase reactant produced by hepatocytes in response to IL-6 [44]. Exposure to the high-glucose diet in the MS group increased the CRP concentration related to the SD group. According to several reports, elevated CRP levels have been directly correlated with obesity, dyslipidemia, hyperglycemia, and insulin resistance [45,46].
After exposure to the CFD, the animals showed an improvement in the systemic inflammatory state. A direct relationship between dietary carbohydrate restriction and improved inflammation has been described, mediated by adenosine levels, ketone bodies, and gut microbiota [47].
MS, especially obesity, insulin resistance, and inflammation, are associated with alterations in liver function. Several reports indicate that liver function, particularly the levels of total bilirubin, ALT, and AST, are closely related to the incidence of MS, in addition to being predictive factors of hepatic steatosis and non-alcoholic fatty liver disease (NAFLD) [48].
Previous clinical studies that incorporated a partially restricted carbohydrate diet reported no changes in the concentrations of these enzymes in obese patients with T2DM [49,50] and patients with NAFLD [51]. However, in this study the concentration of AST and ALT decreased in the MS+CFD group, this may be because carbohydrate restriction is related to states of ketosis observed in this group. Ketosis is associated with an increase in blood lactate, which causes a rapid consumption of NADH, related to a decrease in the enzymatic activity of AST and ALT [52].
Glycogen is stored in the liver and skeletal muscle, acting as an energy reservoir. Liver glycogen is the main store that maintains blood glucose homeostasis. The levels of this polymer are significantly altered after changes in glucose homeostasis in MS [53].
It has been demonstrated that during the first hours of exposure to a carbohydrate-free and high-protein diet, glycogenolysis is the main route of endogenous glucose generation. However, once glycogen levels decrease, the gluconeogenesis pathway became predominant [15]. In this study, the elimination of carbohydrates in the diet of the MS+CFD group induced a decrease in glucose stored as glycogen in liver and skeletal muscle relative to the MS group, during the activation of glycogenolysis for energy requirement.
MS is associated with an increased risk of developing kidney abnormalities. According to some studies, hypertension, hyperglycemia, dyslipidemia, and obesity play a principal role in promoting kidney damage [54]. Although there were no significant differences between the SD and MS groups, the latter group showed an increased tendency in urea and creatinine concentrations, demonstrating the role of a high-glucose diet generating renal alterations. Potential mechanisms by which MS affects renal physiology include impaired renal hemodynamics, insulin resistance, hyperlipidemia, activation of the renin-angiotensin-aldosterone system, inflammation, and oxidative stress [55].
On the other hand, the high protein content in CFD might have fundamentally impact the metabolic processes regulated by the kidneys and renal function [56]. Increased urea excretion in the MS+CFD group about the SD group was observed; however, creatinine levels were not affected by exposure to CFD, thus indicating that the activation of the gluconeogenesis pathway for energy generation occurred from protein amino acids from the diet and not from the skeletal muscle. These results agree with Ko et al. (2017), who report no changes in the creatinine concentration in animals subjected to a high-protein and low-carbohydrate diet. There is evidence about the relationship between diets low in carbohydrates and high in protein and fat, and the increase in the glomerular filtration rate, which is possibly reflected in kidney damage [57,58]. Nevertheless, previous authors suggest the increase in serum urea as a normal adaptive mechanism that occurs in response to an increase in protein metabolism and does not cause negative effects on renal morphology [59].
Oxidative stress, whose etiology is mainly mitochondrial dysfunction, is related to the development of MS, causing the appearance of inflammation, thrombosis, and atherosclerosis [60]. MDA concentration was evaluated as a marker of oxidative damage (lipoperoxidation) in blood serum, liver, kidney, and pancreas. In the MS group, the high glucose content in the diet probably led to an increase in oxidative activity, causing an overproduction of reactive oxygen species (ROS) in the mitochondria, generating damage to macromolecules [60], reflecting in increases in MDA concentrations at serum and liver levels. Diverse studies have reported increased serum and liver MDA concentrations after exposure to a high-carbohydrate diet, the authors relate this increase with an inflammatory state, especially high levels of CRP [25,61].
In contrast, the MS+CFD group showed a decrease in serum MDA concentration, indicating a decrease in oxidative activity and ROS production in mitochondria [62]. Similarly, decreased lipid peroxidation is related to increased levels of β-hydroxybutyrate, since it inhibits pathways related to the production of ROS and the stimulation of antioxidant enzyme activity [40,63]. The results are consistent with those previously observed, where carbohydrate-restricted diets are incorporated [63].
5. Conclusions
Exposure to CFD in animals with diet-induced MS-related abnormalities, normalized morphometric and biochemical parameters, including dyslipidemia state, insulin resistance, serum and hepatic lipid peroxidation, ALT enzyme activity, liver glycogen and creatinine concentrations, also generating a decrease in the inflammatory state. However, there was an increase in the serum urea concentration and a non-significant decrease in the blood glucose level, probably due to the high protein content in the diet.
Author Contributions
Conceptualization, M.G.-V. and B.P.L.-R.; methodology, D.A.L.-G. and I.J.F.-B.; software, B.P.L.-R.; validation, M.G.-V. and B.P.L.-R.; formal analysis, B.P.L.-R. and D.A.L.-G.; investigation, D.A.L.-G. and I.J.F.-B.; resources, M.G.-V. and B.P.L.-R.; data curation, B.P.L.-R and D.A.L.-G.; writing—original draft preparation, D.A.L.-G.; writing—review and editing, M.G.-V. and B.P.L.-R.; visualization, M.G.-V., B.P.L.-R. and D.A.L.-G.; supervision, M.G.-V. and B.P.L.-R.; project administration, M.G.-V., B.P.L.-R., D.A.L.-G. and I.J.F.-B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The animal study protocol was approved by the Research Ethics Committee of Health Sciences Area of AUTONOMOUS UNIVERSITY OF ZACATECAS (protocol folio 004/CEI-ACS-UAZ/2022, May 4, 2022).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- McCracken: E.; Monaghan, M.; Sreenivasan, S. Pathophysiology of the Metabolic Syndrome. Clin. Dermatol. 2017, 36, 14–20. [CrossRef]
- Rochlani, Y.; Pothineni, N.V.; Kovelamudi, S. Metabolic Syndrome : Pathophysiology, Management, and Modulation by Natural Compounds. Ther Adv Cardiovasc Dis 2017, 11, 215–225. [CrossRef]
- Sherling, D.H.; Perumareddi, P.; Hennekens, C.H. Metabolic Syndrome: Clinical and Policy Implications of the New Silent Killer. J. Cardiovasc. Pharmacol. Ther. 2017, 22, 365–367. [CrossRef]
- Fahed, G.; Aoun, L.; Bou Zerdan, M.; Allam, S.; Bou Zerdan, M.; Bouferraa, Y.; Assi, H.I. Metabolic Syndrome: Updates on Pathophysiology and Management in 2021. Int. J. Mol. Sci. 2022, 23, 786. [CrossRef]
- Soleimani, M.; Barone, S.; Luo, H.; Zahedi, K. Pathogenesis of Hypertension in Metabolic Syndrome: The Role of Fructose and Salt. Int. J. Mol. Sci. 2023, 24, 4294. [CrossRef]
- Kassi, E.; Pervanidou, P.; Kaltsas, G.; Chrousos, G. Metabolic Syndrome: Definitions and Controversies. BMC Med. 2011, 9, 48. [CrossRef]
- Akter, S.; Akhter, H.; Chaudhury, H.S.; Rahman, M.H.; Gorski, A.; Hasan, M.N.; Shin, Y.; Rahman, M.A.; Nguyen, M.N.; Choi, T.G.; et al. Dietary Carbohydrates: Pathogenesis and Potential Therapeutic Targets to Obesity-Associated Metabolic Syndrome. Biofactors 2022, 48, 1036–1059. [CrossRef]
- Xu, H.; Li, X.; Adams, H.; Kubena, K.; Guo, S. Etiology of Metabolic Syndrome and Dietary Intervention. Int. J. Mol. Sci. 2019, 20, 128. [CrossRef]
- Smith, C.J.; Ryckman, K.K. Epigenetic and Developmental Influences on the Risk of Obesity, Diabetes, and Metabolic Syndrome. Diabetes, Metab. Syndr. Obes. Targets Ther. 2015, 8, 295–302. [CrossRef]
- Chan, A.M.L.; Ng, A.M.H.; Mohd Yunus, M.H.; Idrus, R.B.H.; Law, J.X.; Yazid, M.D.; Chin, K.Y.; Shamsuddin, S.A.; Lokanathan, Y. Recent Developments in Rodent Models of High-Fructose Diet-Induced Metabolic Syndrome: A Systematic Review. Nutrients 2021, 13, 1–21. [CrossRef]
- Rodríguez-Monforte, M.; Sánchez, E.; Barrio, F.; Costa, B.; Flores-Mateo, G. Metabolic Syndrome and Dietary Patterns: A Systematic Review and Meta-analysis of Observational Studies. Eur. J. Nutr. 2016, 56, 925–947. [CrossRef]
- Hyde, P.N.; Sapper, T.N.; Crabtree, C.D.; LaFountain, R.A.; Bowling, M.L.; Buga, A.; Fell, B.; McSwiney, F.T.; Dickerson, R.M.; Miller, V.J.; et al. Dietary Carbohydrate Restriction Improves Metabolic Syndrome Independent of Weight Loss. JCI Insight 2019, 4, e128308. [CrossRef]
- Marsset-Baglieri, A.; Fromentin, G.; Tomé, D.; Bensaid, A.; Makkarios, L.; Even, P.C. Increasing the Protein Content in a Carbohydrate-Free Diet Enhances Fat Loss during 35% but Not 75% Energy Restriction in Rats. J. Nutr. 2004, 134, 2646–2652. [CrossRef]
- Volek, J.S.; Feinman, R.D. Carbohydrate Restriction Improves the Features of Metabolic Syndrome. Metabolic Syndrome May Be Defined by the Response to Carbohydrate Restriction. Nutr. Metab. 2005, 2, 1–17. [CrossRef]
- Azzout, B.; Chanez, M.; Bois-Joyeux, B.; Peret, J. Gluconeogenesis from Dihydroxyacetone in Rat Hepatocytes during the Shift from a Low Protein, High Carbohydrate to a High Protein, Carbohydrate-Free Diet. J. Nutr. 1984, 114, 2167–2178. [CrossRef]
- Pichon, L.; Huneu, J.; Fromentin, G.; Tomé, D. A High-Protein, High-Fat, Carbohydrate-Free Diet Reduces Energy Intake, Hepatic Lipogenesis, and Adiposity in Rats. Nutr. Physiol. Metab. Nutr. Interact. 2006, 136. [CrossRef]
- Jürgens, H.S.; Neschen, S.; Ortmann, S.; Scherneck, S.; Schmolz, K.; Schüler, G.; Schmidt, S.; Blüher, M.; Klaus, S.; Perez-Tilve, D.; et al. Development of Diabetes in Obese, Insulin-Resistant Mice: Essential Role of Dietary Carbohydrate in Beta Cell Destruction. Diabetologia 2007, 50, 1481–1489. [CrossRef]
- Mirhashemi, F.; Kluth, O.; Scherneck, S.; Vogel, H.; Schürmann, A.; Susanne, H.J. High-Fat , Carbohydrate-Free Diet Markedly Aggravates Obesity but Prevents β -Cell Loss and Diabetes in the Obese , Diabetes-Susceptible Db / Db Strain. 2008, 292–297. [CrossRef]
- Uchiyama, M.; Mihara, M. Determination of Malonaldehyde Precursor in Tissues by Thiobarbituric Acid Test. Anal. Biochem. 1978, 86, 271–278. [CrossRef]
- Fong, J.; Schaffer, F.L.; Kirk, P.L. The Ultramicrodetermination of Glycogen in Liver. Arch. Biochem. Biophys. 1953, 45, 319–326. [CrossRef]
- O’Neill, B.J. Effect of Low-Carbohydrate Diets on Cardiometabolic Risk, Insulin Resistance, and Metabolic Syndrome. Curr. Opin. Endocrinol. Diabetes. Obes. 2020, 27, 301–307. [CrossRef]
- Korkmaz, O.A.; Sadi, G.; Kocabas, A.; Yildirim, O.G.; Sumlu, E.; Koca, H.B.; Nalbantoglu, B.; Pektas, M.B.; Akar, F. Lactobacillus Helveticus and Lactobacillus Plantarum Modulate Renal Antioxidant Status in a Rat Model of Fructose-Induced Metabolic Syndrome. Arch. Biol. Sci. 2019, 71, 265–273. [CrossRef]
- Kubacka, M.; Kotanska, M.; Szafarz, M.; Pociecha, K.; Waszkielewicz, A.M.; Marona, H.; Filipek, B.; Mogilski, S. Beneficial Effects of Non-Quinazoline A1-Adrenolytics on Hypertension and Altered Metabolism in Fructose-Fed Rats. A Comparison with Prazosin. Nutr. Metab. Cardiovasc. Dis. 2019, 29, 751–760. [CrossRef]
- Neeland, I.J.; Turer, A.T.; Ayers, C.R.; Powell-Wiley, T.M.; Vega, G.L.; Farzaneh-Far, R.; Grundy, S.M.; Khera, A.; McGuire, D.K.; de Lemos, J.A. Dysfunctional Adiposity and the Risk of Prediabetes and Type 2 Diabetes in Obese Adults. JAMA 2013, 308, 1150–1159. [CrossRef]
- Bratoeva, K.; Nikolova, S.; Merdzhanova, A.; Stoyanov, G.S.; Dimitrova, E.; Kashlov, J.; Conev, N.; Radanova, M. Association between Serum CK-18 Levels and the Degree of Liver Damage in Fructose-Induced Metabolic Syndrome. Metab. Syndr. Relat. Disord. 2018, 16, 350–357. [CrossRef]
- Ferreira-Santos, P.; Aparicio, R.; Carrón, R.; Montero, M.J.; Sevilla, M.Á. Lycopene-Supplemented Diet Ameliorates Metabolic Syndrome Induced by Fructose in Rats. J. Funct. Foods 2020, 73, 104098. [CrossRef]
- Preguiça, I.; Alves, A.; Nunes, S.; Fernandes, R.; Gomes, P.; Viana, S.D.; Reis, F. Diet-Induced Rodent Models of Obesity-Related Metabolic Disorders—A Guide to a Translational Perspective. Obes. Rev. 2020, 21, 1–29. [CrossRef]
- Moon, J.; Koh, G. Clinical Evidence and Mechanisms of High-Protein Diet-Induced Weight Loss. J. Obes. Metab. Syndr. 2021, 29, 166–173. [CrossRef]
- Pichon, L.; Huneu, J.; Fromentin, G.; Tomé, D. A High-Protein, High-Fat, Carbohydrate-Free Diet Reduces Energy Intake, Hepatic Lipogenesis, and Adiposity in Rats. Nutr. Physiol. Metab. Nutr. Interact. 2006, 136, 1256–1260. [CrossRef]
- Fukazawa, A.; Koike, A.; Karasawa, T.; Tsutsui, M.; Kondo, S.; Terada, S. Effects of a Ketogenic Diet Containing Medium-Chain Triglycerides and Endurance Training on Metabolic Enzyme Adaptations in Rat Skeletal Muscle. Nutrients 2020, 12. [CrossRef]
- Rodríguez-Correa, E.; González-Pérez, I.; Clavel-Pérez, P.I.; Contreras-Vargas, Y.; Carvajal, K. Biochemical and Nutritional Overview of Diet-Induced Metabolic Syndrome Models in Rats: What Is the Best Choice? Nutr. Diabetes 2020, 10, 24. [CrossRef]
- Wong, S.K.; Chin, K.Y.; Suhaimi, F.H.; Fairus, A.; Ima-Nirwana, S. Animal Models of Metabolic Syndrome: A Review. Nutr. Metab. 2016, 13, 1–12. [CrossRef]
- Roberts, C.K.; Hevener, A.L.; Barnard, R.J. Metabolic Syndrome and Insulin Resistance: Underlying Causes and Modification by Exercise Training. Compr. Physiol. 2013, 3, 1–58. [CrossRef]
- Thomas, D.D.; Corkey, B.E.; Istfan, N.W.; Apovian, C.M. Hyperinsulinemia: An Early Indicator of Metabolic Dysfunction. J. Endocr. Soc. 2019, 3, 1727–1747. [CrossRef]
- Brito, S.M.R.C.; Moura, M.A.F.; Kawashita, N.H.; Festuccia, W.T.L.; Garófalo, M.A.R.; Kettelhut, I.C.; Migliorini, R.H. Adaptation to a High Protein, Carbohydrate-Free Diet Induces a Marked Reduction of Fatty Acid Synthesis and Lipogenic Enzymes in Rat Adipose Tissue That Is Rapidly Reverted by a Balanced Diet. Can. J. Physiol. Pharmacol. 2005, 83, 477–482. [CrossRef]
- Veldhorst, M.A.; Westerterp-Plantenga, M.S.; Westerterp, K.R. Gluconeogenesis and Energy Expenditure after a High-Protein, Carbohydrate-Free Diet. Am. J. Clin. Nutr. 2009, 90, 519–526. [CrossRef]
- Sukkar, A.H.; Lett, A.M.; Frost, G.; Chambers, E.S. Regulation of Energy Expenditure and Substrate Oxidation by Short-Chain Fatty Acids. J. Endocrinol. 2019, 242, R1–R8. [CrossRef]
- Locatelli, C.A.A.; Mulvihill, E.E. Islet Health, Hormone Secretion, and Insulin Responsivity with Low-Carbohydrate Feeding in Diabetes. Metabolites 2020, 10, 1–17. [CrossRef]
- Møller, N. Ketone Body, 3-Hydroxybutyrate: Minor Metabolite - Major Medical Manifestations. J. Clin. Endocrinol. Metab. 2020, 105, dgaa370. [CrossRef]
- Newman, J.C.; Verdin, E. β -Hydroxybutyrate: Much More than a Metabolite. Diabetes Res. Clin. Pract. 2014, 106, 173–181. [CrossRef]
- Haile, K.; Haile, A.; Timerga, A. Predictors of Lipid Profile Abnormalities Among Patients with Metabolic Syndrome in Southwest Ethiopia: A Cross-Sectional Study. Vasc. Health Risk Manag. 2021, 17, 461–469. [CrossRef]
- Forouhi, N.G.; Krauss, R.M.; Taubes, G.; Willett, W. Dietary Fat and Cardiometabolic Health: Evidence, Controversies, and Consensus for Guidance. BMJ 2018, 361, k2139. [CrossRef]
- Mansoor, N.; Vinknes, K.J.; Veierød, M.B.; Retterstøl, K. Effects of Low-Carbohydrate Diets v. Low-Fat Diets on Body Weight and Cardiovascular Risk Factors: A Meta-Analysis of Randomised Controlled Trials. Br. J. Nutr. 2016, 115, 466–479. [CrossRef]
- Reddy, P.; Lent-Schochet, D.; Ramakrishnan, N.; Mclaughlin, M.; Jialal, I. Metabolic Syndrome Is an Inflammatory Disorder: A Conspiracy between Adipose Tissue and Phagocytes. Clin. Chim. Acta 2019, 496, 35–44. [CrossRef]
- Kwitek, A.E. Rat Models of Metabolic Syndrome. Methods Mol. Biol. 2019, 2018, 269–285. [CrossRef]
- Monteiro, R.; Azevedo, I. Chronic Inflammation in Obesity and the Metabolic Syndrome. Mediators Inflamm. 2010, 2010. [CrossRef]
- Koh, S.; Dupuis, N.; Cit, S.P.; Debr, R. Ketogenic Diet and Neuroinflammation. Epilepsy Res. 2020, 167, 1–8. [CrossRef]
- Valle-Martos, R.; Valle, M.; Martos, R.; Cañete, R.; Jiménez-Reina, L.; Cañete, M.D. Liver Enzimes Correlate with Metabolic Syndrome, Inflammation, and Endothelial Dysfunction in Prepubertal Children with Obesity. Front. Pediatr. 2021, 9, 629346. [CrossRef]
- Paoli, A.; Bianco, A.; Grimaldi, K.A.; Lodi, A.; Bosco, G. Long Term Successful Weight Loss with a Combination Biphasic Ketogenic Mediterranean Diet and Mediterranean Diet Maintenance Protocol. Nutrients 2013, 5, 5205–5217. [CrossRef]
- Walton, C.M.; Perry, K.; Hart, R.H.; Berry, S.L.; Bikman, B.T. Improvement in Glycemic and Lipid Profiles in Type 2 Diabetics with a 90-Day Ketogenic Diet. 2019, 2019. [CrossRef]
- Luukkonen, P.K.; Dufour, S.; Lyu, K.; Zhang, X.M.; Hakkarainen, A.; Lehtimäki, T.E.; Cline, G.W.; Petersen, K.F.; Shulman, G.I.; Yki-Järvinen, H. Effect of a Ketogenic Diet on Hepatic Steatosis and Hepatic Mitochondrial Metabolism in Nonalcoholic Fatty Liver Disease. Proc. Natl. Acad. Sci. U. S. A. 2020, 117, 7347–7354. [CrossRef]
- Lum, G. Low Activities of Aspartate and Alanine Aminotransferase: Their Significance in Alcoholic Liver Disease. Lab. Med. 1995, 26, 4–7. [CrossRef]
- Roach, P.J.; Depaoli-Roach, A.A.; Hurley, T.D.; Tagliabracci, V.S. Glycogen and Its Metabolism: Some New Developments and Old Themes. Biochem. J. 2016, 441, 763–787. [CrossRef]
- Al-Daghri, N.M.; Al-Attas, O.S.; Wani, K.; Sabico, S.; Alokail, M.S. Serum Uric Acid to Creatinine Ratio and Risk of Metabolic Syndrome in Saudi Type 2 Diabetic Patients. Sci. Rep. 2017, 1–8. [CrossRef]
- Tang, J.; Yan, H.; Zhuang, S. Inflammation and Oxidative Stress in Obesity-Related Glomerulopathy. Int. J. Nephrol. 2012, 2012. [CrossRef]
- Ko, G.J.; Obi, Y.; Tortoricci, A.R.; Kalantar-Zadeh, K. Dietary Protein Intake and Chronic Kidney Disease. Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 77–85. [CrossRef]
- Juraschek, S.P.; Chang, A.R.; Appel, L.J.; Anderson, C.A.M.; Crews, D.C.; Thomas, L.; Charleston, J.; Iii, E.R.M. Effect of Glycemic Index and Carbohydrate Intake on Kidney Function in Healthy Adults. BMC Nephrol. 2016, 17, 70. [CrossRef]
- Kostogrys, R.B.; Franczyk-Żarów, M.; Maślak, E.; Topolska, K. Effect of Low Carbohydrate High Protein (LCHP) Diet on Lipid Metabolism, Liver and Kidney Function in Rats. Environ. Toxicol. Pharmacol. 2015, 39, 713–719. [CrossRef]
- Martin, W.F.; Armstrong, L.E.; Rodriguez, N.R. Dietary Protein Intake and Renal Function. Nutr. Metab. 2005, 2, 25. [CrossRef]
- Onyango, A.N. Lipid Peroxidation as a Link between Unhealthy Diets and the Metabolic Syndrome. In Accenting Lipid Peroxidation; Atukeren, P., Ed.; 2021.
- Panchal, S.K.; Poudyal, H.; Iyer, A.; Nazer, R.; Alam, A.; Diwan, V.; Kauter, K.; Sernia, C.; Campbell, F.; Ward, L.; et al. High-Carbohydrate High-Fat Diet-Induced Metabolic Syndrome and Cardiovascular Remodeling in Rats. J. Cardiovasc. Pharmacol. 2011, 57, 51–64. [CrossRef]
- Avelar, T.M.T.; Storch, A.S.; Castro, L.A.; Azevedo, G.V.M.M.; Ferraz, L.; Lopes, P.F. Oxidative Stress in the Pathophysiology of Metabolic Syndrome : Which Mechanisms Are Involved ? 2015, 231–239. [CrossRef]
- Chimienti, G.; Orlando, A.; Lezza, A.M.S.; D’Attoma, B.; Notarnicola, M.; Gigante, I.; Pesce, V.; Russo, F. The Ketogenic Diet Reduces the Harmful Effects of Stress on Gut Mitochondrial Biogenesis in a Rat Model of Irritable Bowel Syndrome. Int. J. Mol. Sci. 2021, 22, 3498. [CrossRef]
Figure 1.
(A) Food and (B) beverage intake, (C) total caloric intake and (D) body weight in the groups studied throughout the 40 weeks of experimentation. Data are presented as means ± SD. The comparison between groups was performed using an analysis of variance (ANOVA) followed by Tukey’s post hoc adjustment test. It was considered statistically significant when p<0.05, the statistical differences between groups are presented as: a: SD vs MS; b: SD vs MS+CFD; c: MS vs MS+CFD. Abbreviations: SD, standard diet; MS, metabolic syndrome; MS+CFD, metabolic syndrome, subsequent carbohydrate-free diet.
Figure 1.
(A) Food and (B) beverage intake, (C) total caloric intake and (D) body weight in the groups studied throughout the 40 weeks of experimentation. Data are presented as means ± SD. The comparison between groups was performed using an analysis of variance (ANOVA) followed by Tukey’s post hoc adjustment test. It was considered statistically significant when p<0.05, the statistical differences between groups are presented as: a: SD vs MS; b: SD vs MS+CFD; c: MS vs MS+CFD. Abbreviations: SD, standard diet; MS, metabolic syndrome; MS+CFD, metabolic syndrome, subsequent carbohydrate-free diet.
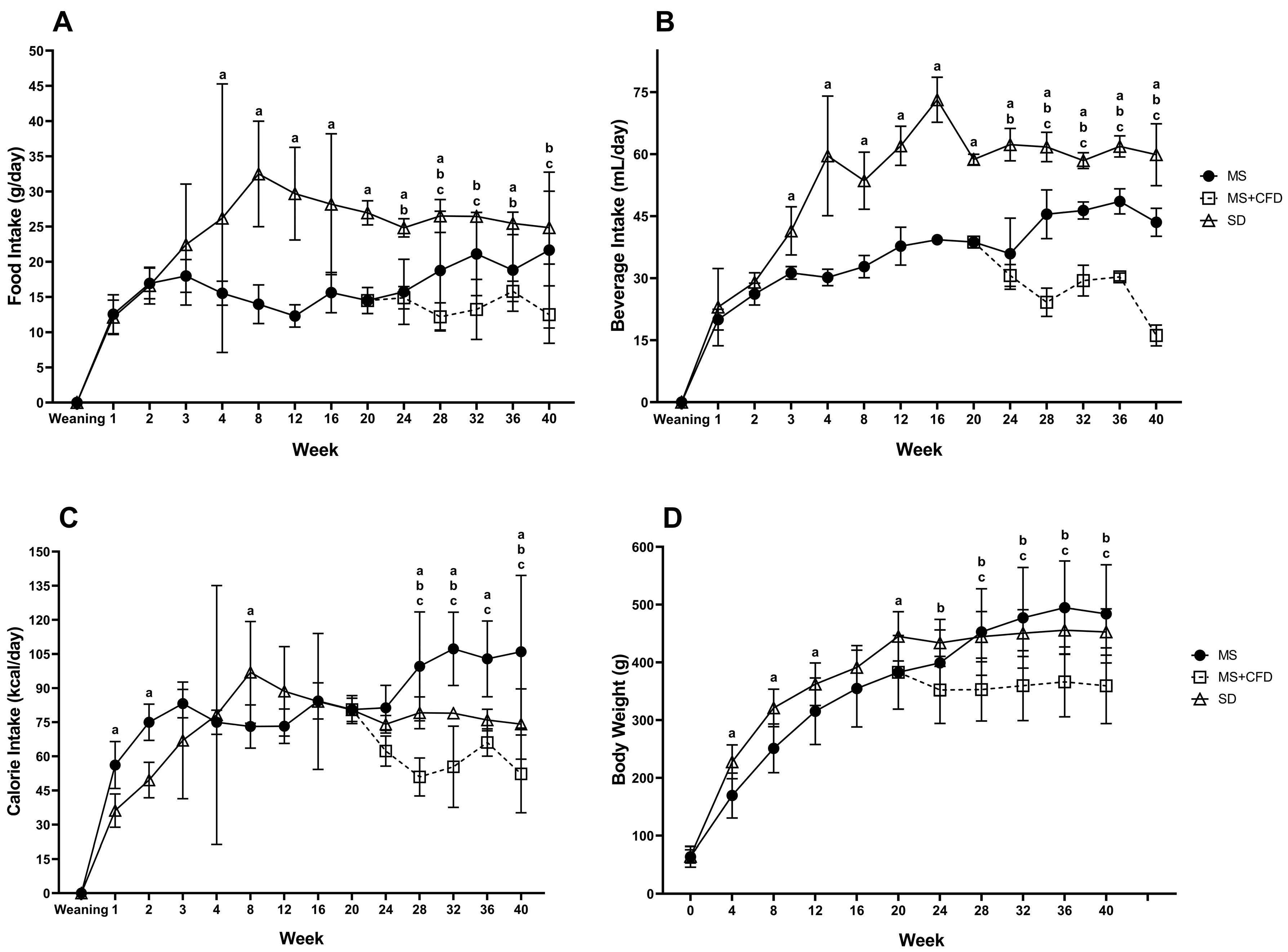
Figure 2.
(A) Glucose, (B) insulin, and (C) β-hydroxybutyrate concentrations at the 40th week of experimentation in the study groups. Data are presented as median (minimum-maximum). A comparison analysis between groups was performed using the Kruskal-Wallis’ test and Dunn’s adjustment test post hoc. It was considered statistically significant when p<0.05. Abbreviations: SD, standard diet; MS, metabolic syndrome; MS+CFD, metabolic syndrome, subsequent carbohydrate-free diet.
Figure 2.
(A) Glucose, (B) insulin, and (C) β-hydroxybutyrate concentrations at the 40th week of experimentation in the study groups. Data are presented as median (minimum-maximum). A comparison analysis between groups was performed using the Kruskal-Wallis’ test and Dunn’s adjustment test post hoc. It was considered statistically significant when p<0.05. Abbreviations: SD, standard diet; MS, metabolic syndrome; MS+CFD, metabolic syndrome, subsequent carbohydrate-free diet.
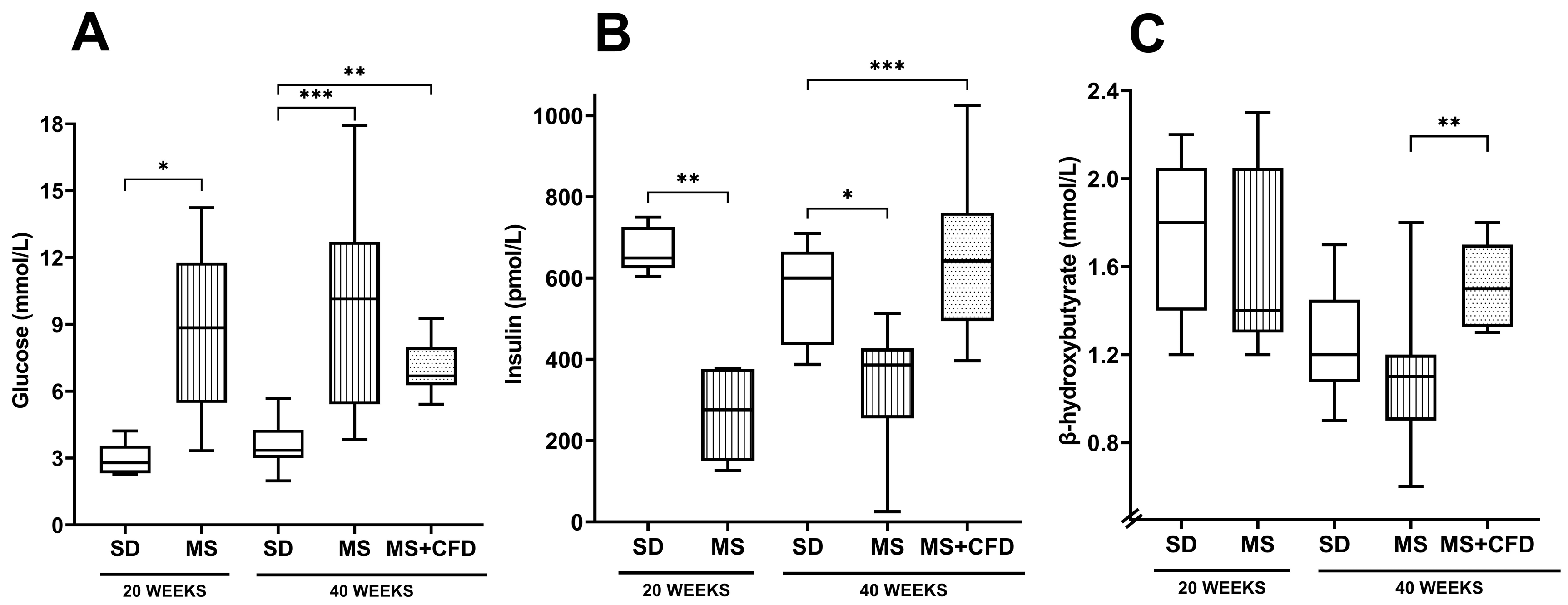
Figure 3.
(A) Triglycerides, (B) total cholesterol, (C) HDL-c, (D) LDL-c, and (E) VLDL-c concentrations at the 40th week of experimentation in the study groups. Data are presented as median (minimum-maximum). A comparison analysis between groups was performed using the Kruskal-Wallis’ test and Dunn’s adjustment test post hoc. It was considered statistically significant when p<0.05. Abbreviations: SD, standard diet; MS, metabolic syndrome; MS+CFD, metabolic syndrome, subsequent carbohydrate-free diet.
Figure 3.
(A) Triglycerides, (B) total cholesterol, (C) HDL-c, (D) LDL-c, and (E) VLDL-c concentrations at the 40th week of experimentation in the study groups. Data are presented as median (minimum-maximum). A comparison analysis between groups was performed using the Kruskal-Wallis’ test and Dunn’s adjustment test post hoc. It was considered statistically significant when p<0.05. Abbreviations: SD, standard diet; MS, metabolic syndrome; MS+CFD, metabolic syndrome, subsequent carbohydrate-free diet.
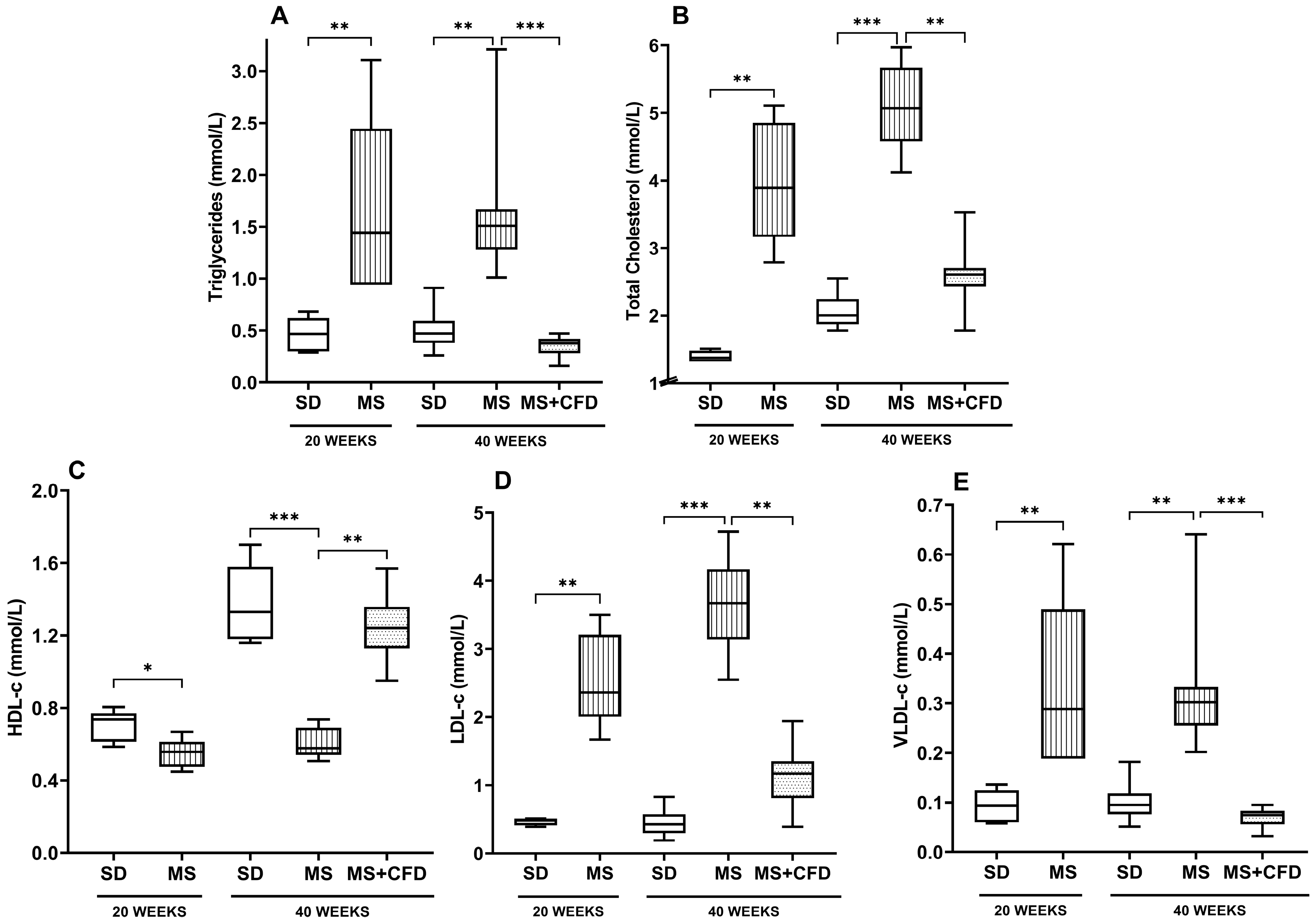
Figure 4.
C-reactive protein concentrations in the study groups at the 40th week of experimentation. Data are presented as median (minimum-maximum). A comparison analysis between groups was performed using the Kruskal-Wallis’ test and Dunn’s adjustment test post hoc. It was considered statistically significant when p<0.05. Abbreviations: SD, standard diet; MS, metabolic syndrome; MS+CFD, metabolic syndrome, subsequent carbohydrate-free diet.
Figure 4.
C-reactive protein concentrations in the study groups at the 40th week of experimentation. Data are presented as median (minimum-maximum). A comparison analysis between groups was performed using the Kruskal-Wallis’ test and Dunn’s adjustment test post hoc. It was considered statistically significant when p<0.05. Abbreviations: SD, standard diet; MS, metabolic syndrome; MS+CFD, metabolic syndrome, subsequent carbohydrate-free diet.
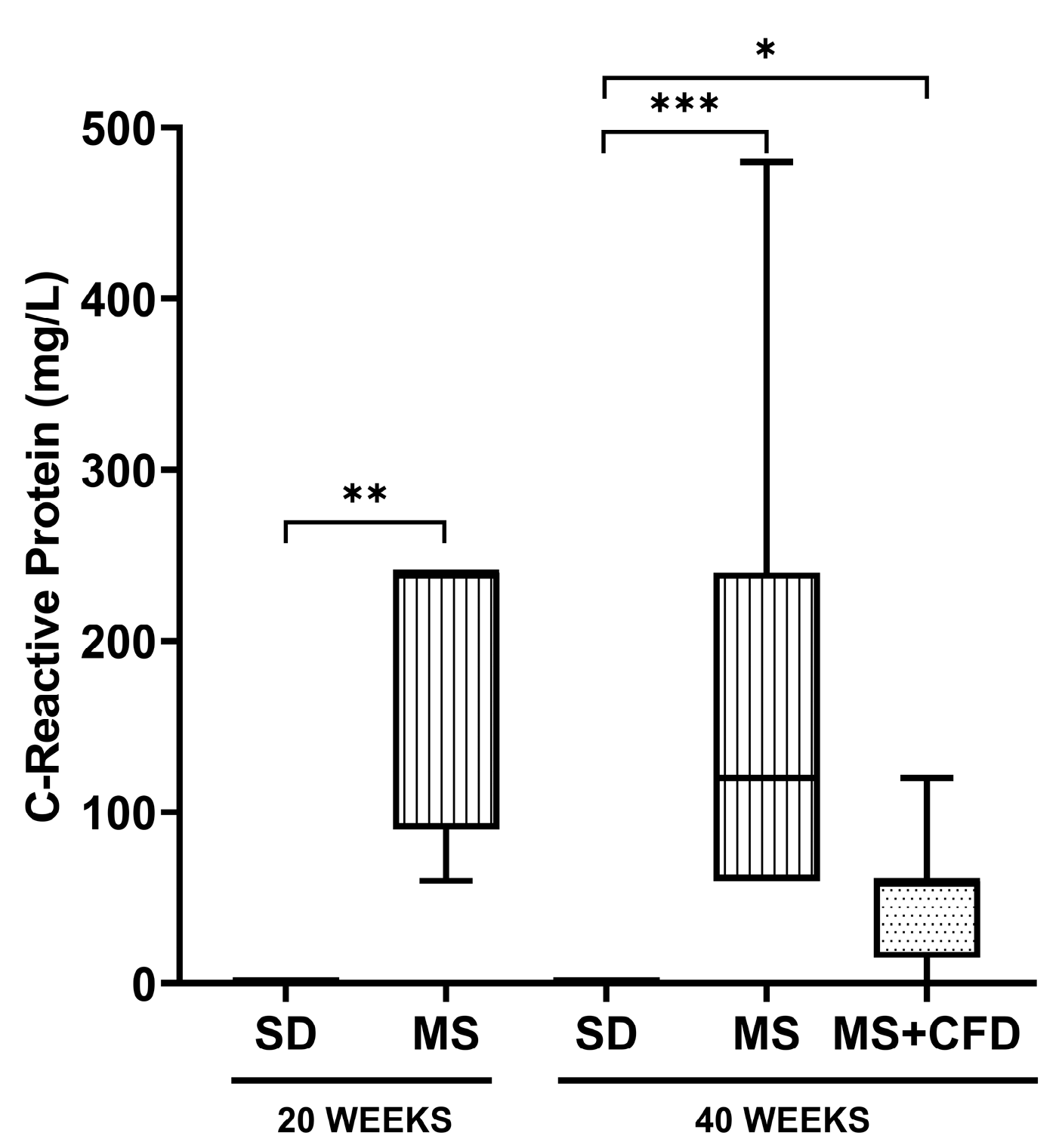
Figure 5.
(A) Serum urea and (B) serum creatinine concentrations at the 40th week of experimentation in the study groups. Data are presented as median (minimum-maximum). A comparison analysis between groups was performed using the Kruskal-Wallis’ test and Dunn’s adjustment test post hoc. It was considered statistically significant when p<0.05. Abbreviations: SD, standard diet; MS, metabolic syndrome; MS+CFD, metabolic syndrome, subsequent carbohydrate-free diet.
Figure 5.
(A) Serum urea and (B) serum creatinine concentrations at the 40th week of experimentation in the study groups. Data are presented as median (minimum-maximum). A comparison analysis between groups was performed using the Kruskal-Wallis’ test and Dunn’s adjustment test post hoc. It was considered statistically significant when p<0.05. Abbreviations: SD, standard diet; MS, metabolic syndrome; MS+CFD, metabolic syndrome, subsequent carbohydrate-free diet.
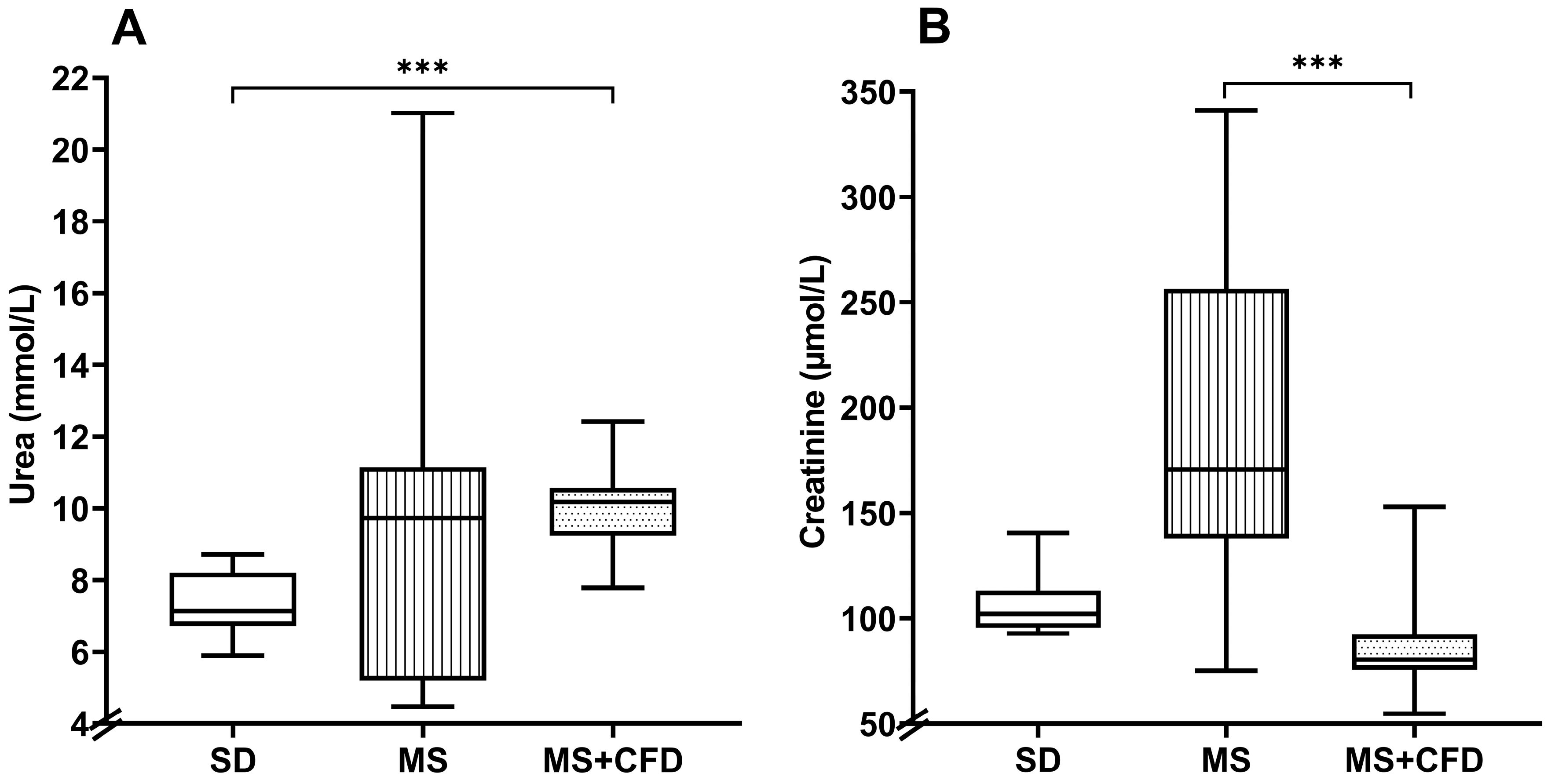
Figure 6.
MDA concentrations in (A) blood serum, (B) liver, (C) kidney and (D) pancreas in the study groups at the 40th week of experimentation. Data are presented as median (minimum-maximum). A comparison analysis between groups was performed using the Kruskal-Wallis’ test and Dunn’s adjustment test post hoc. It was considered statistically significant when p<0.05. Abbreviations: SD, standard diet; MS, metabolic syndrome; MS+CFD, metabolic syndrome, subsequent carbohydrate-free diet; MDA, malondialdehyde.
Figure 6.
MDA concentrations in (A) blood serum, (B) liver, (C) kidney and (D) pancreas in the study groups at the 40th week of experimentation. Data are presented as median (minimum-maximum). A comparison analysis between groups was performed using the Kruskal-Wallis’ test and Dunn’s adjustment test post hoc. It was considered statistically significant when p<0.05. Abbreviations: SD, standard diet; MS, metabolic syndrome; MS+CFD, metabolic syndrome, subsequent carbohydrate-free diet; MDA, malondialdehyde.
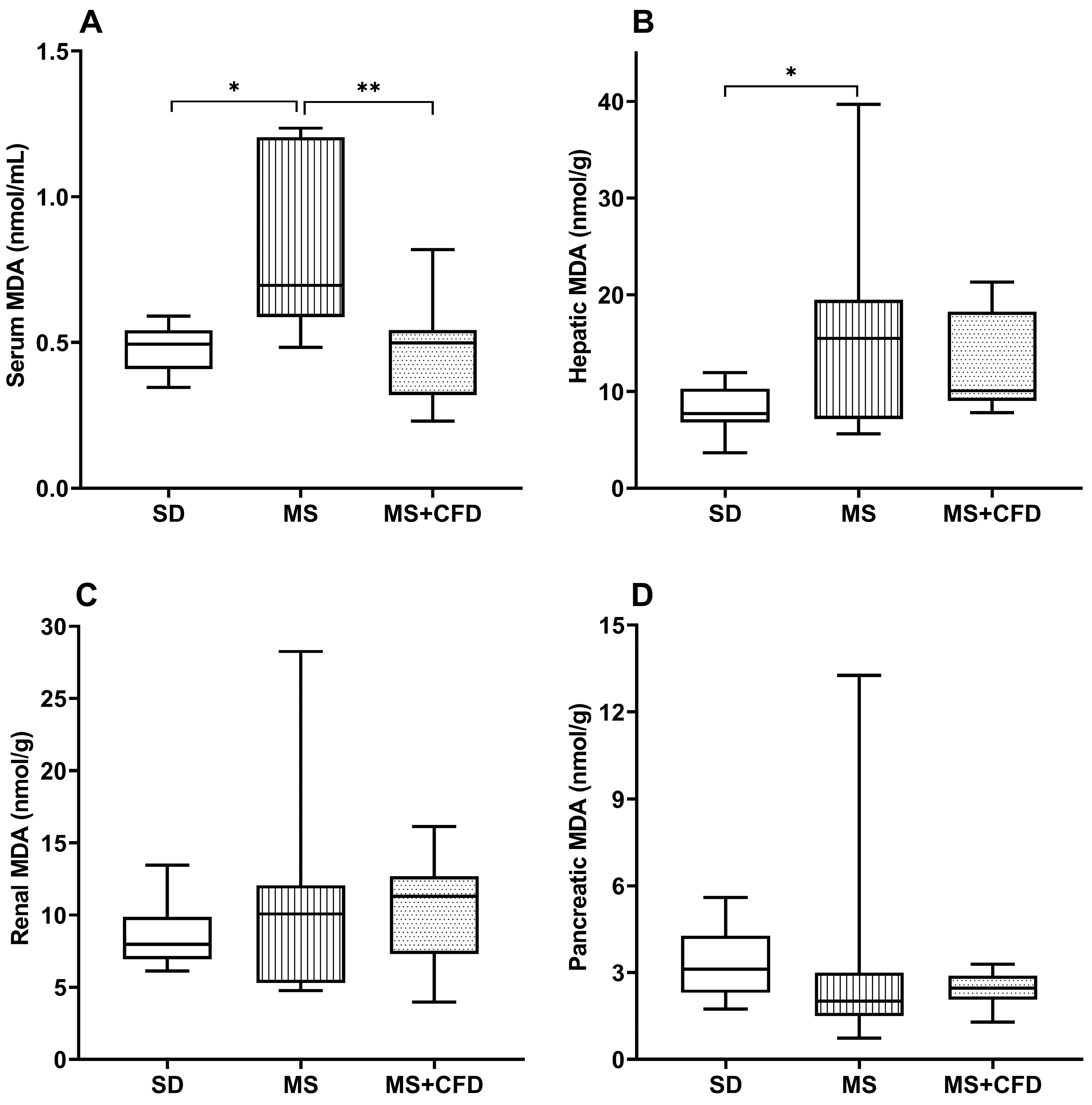

Table 1.
Hepatic enzymes and, liver and muscle glycogen concentrations.
| Biochemical Parameter | SD Group | MS Group | MS+CFD Group | p Value |
|---|---|---|---|---|
| AST (U/L) | 84.29 (72.92-214.08) | 75.25 (54.83-155.75) | 44.63 (11.66-88.08) | 0.00082 0.0133 |
| ALT (U/L) | 49.88 (39.08-99.17) | 16.33 (13.42-24.50) | 31.79 (7.00-66.50) | 0.0001 0.0213 |
| Liver Glycogen (µmol/L) | 1.82 (1.41-2.85) | 6.47 (2.69-22.33) | 3.13 (0.14-5.23) | 0.0001 |
| Muscle Glycogen (µmol/L) | 2.64 (1.51-6.0) | 4.23 (1.84-5.75) | 2.85 (0.22-5.49) | ns |
Data are presented as median (minimum-maximum). A comparison analysis between groups was performed using the Kruskal-Wallis’ test and Dunn’s adjustment test post hoc. It was considered statistically significant when p<0.05. The comparisons were: 1: ND vs MS; 2: ND vs MS+CFD; 3: MS vs MS+CFD. Abbreviations: SD, standard diet; MS, metabolic syndrome; MS+CFD, metabolic syndrome, subsequent carbohydrate-free diet; ALT, alanine aminotransferase; AST, aspartate aminotransferase.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



