
Submitted:
15 June 2023
Posted:
16 June 2023
You are already at the latest version
Abstract
Cervical cancer is associated with persistent infections by high-risk HPV types that may have nucleotide polymorphisms and, consequently, different oncogenic potentials. Therefore, the objective of this study was to evaluate the genetic variability and structural effects of E7 oncogene of HPV58 in cervical scraping samples from Brazilian women. The study was carried out with patients from hospitals in metropolitan area of Recife, PE, Brazil. The most frequent HPV type was HPV16, 18 and 58, respectively. Phylogenetic analysis showed that the isolates were classified as sublineages A2, C1 and D2. Two positively selected mutations were found in E7: 63G and 64T. The mutations G41R, G63D and T64A in E7 protein were predicted to reduce the stability of the protein structure. Regarding the interaction of the E7 variant of HPV58 with the signaling of the NF-kB pathway, we observed that the variant HPV58/UFPE-54S decreased the activity of the pathway when compared to the prototype and the other variants behaved similarly to the prototype and the prototype increased the activity of the pathway when compared to pcDNA. In this study, it was possible to identify mutations that may interfere in the molecular interaction between the viral oncoproteins and host proteins.
Keywords:
human papillomavirus 58
; E7 oncogene
; variants
; phylogenetic analysis
; gene expression
1. Introduction
It is well established that cervical cancer is caused by the infection of high-risk HPVs (hrHPVs), which include genotypes of high carcinogenic potential [1]. In Brazil, the most frequent hrHPVs are 16, 31, and 58 [2]. Together, they reach the third position in tumor frequency (about 16.340 new cases were reported in 2016) and the fourth in cause of deaths in the female population of this country, constituting a serious public health issue, as it occurs in other developing countries [3]. It was estimated by the World Health Organization (WHO) that 84% of cervical cancer cases occurred in developing countries in 2018, with about 311.000 women deaths. This issue is mainly due to failures in the execution of public health programs for the screening of this cancer [4].
In Northeast of Brazil, HPV58 appears again as one of the most frequent genotypes in cervical lesions (the second most frequent), together with HPVs 16 (1st) and 31 (3rd) [5]. In CIN3 lesions, HPV58 was also the second most frequent and the third in invasive cancer lesions [6]. As hrHPVs 16 and 31, HPV58 is an Alphapapillomavirus, constituting an important cause of cervical cancer and plays a key role in cervical carcinogenesis by the expression of its oncogenes, especially E6 and E7[7].
The impact of different genetics variants on carcinogenesis varies widely, due to their influence in host immune response, cell proliferation and the expression of specific molecules which modify cell signaling and communication as well as proinflammatory and apoptosis activities [8,9]. It is known, for example, that variants established by part of LCR region and entire E6 from HPVs 16 and 18 showed different predispositions for lesion progression to CIN3 [10]. In other study, E6 and E7 oncogenes variations of HPV18 have been associated with different histological forms of cervical cancer [11]. Variants produced by differences in nucleotide sequences of these regions produce effects on viral replication and transcription, and the presence of specific epitopes [2,5]. Therefore, these and other data have established that different variant have different oncogenic potentials.
Regarding HPV58, several studies have found different clinical implications for its oncogene variants, such as E7. For example, a specific variant of this oncogene, T20I/G63S, was associated with an increased immortalizing potential (higher than the prototype), transforming ability, capacity of degrading pRb [12] and risk of cervical cancer due to the induced activation of AKT and K-Ras/ERK signaling pathways [9]. Also, the evaluation of gene variants is important for identification of epitopes for the development more efficient vaccines which would be directed to specific populations [13,14].
In unpublished data, our group has found that different variants of E7 oncogene presented different effects on NF-κB gene expression when compared the variants with each other and with the wild gene. It is well established this transcription factor plays a central role in carcinogenesis of cervical and other cancers due to its effects on transduction signaling (e.g. of TLRs and IFNs), stromal environment, proinflammatory response and immune surveillance [15,16].
In addition, specific variants correlate with specific clinical and epidemiological attributes, with risk of cervical cancer and thus, with disease prognosis [8,10,17]. Therefore, the evaluation of hrHPV oncogenes variants is important to assess disease progression, establish patient prognosis and choose the best therapeutic strategy according to clinical presentation.
In this context, this study aimed to evaluate the genetic variability and structural effects of E7 oncogene of HPV58 in cervical scraping samples from Brazilian women in order to identify the possible impact of these mutations in the carcinogenic process.
2. Materials and Methods
2.1. Study groups and DNA isolation
We enrolled 319 women aging from 18 to 77 years old, with a mean age of 37,5 (s.d. 11.9) attended at the Gynecology Unit at the Clinical Hospital (n=81) and Oswaldo Cruz University Hospital (n=156) at Recife, Pernambuco State, Northeastern Brazil, and at the Center for Integral Attention to Women’s Health (n=82) at Aracaju, Sergipe State, Northeastern Brazil. The patients came from spontaneous demand on routine examinations. Approval of the Ethical Committee (CEP/CCS/UFPE N° 49/11) and informed consent from all patients enrolled for the study were obtained.
The samples selected for analysis came from cervical scraping, collected through a gynecological brush (cytobrush) which reaches the endocervix. The brush containing the collected gynecological material was transferred to a sterile tube containing PBS solution (pH 7.4) to preserve the cells until DNA extraction. Cervical cells were stored at -80°C and the DNA was extracted using DNeasy Blood Tissue Kit (Qiagen) according to the manufacturer manual.
2.2. HPV: detection and genotyping)
The extracted DNA was amplified by Polymerase Chain Reaction (PCR) of human MDM2 gene (forward primer: 5’-GATTTCGGACGGCTCTCGCGGC-3’; reverse primer: 5’-GATTTCGGACGGCTCTCGCGGC-3’) to evaluate the quality of DNA, in order to avoid false negative results [Ma et al., 2006]. The PCR reaction was performed in 20µL volume, containing 2µL (50 ng/µL) of the extracted DNA, 10µL of GoTaq Green PCR Master Mix (Promega), 1.2µL of each primer (12.2 pM/µL) and 5.6µL of Nuclease-Free Water (Promega). The reaction condition was 95ºC for 5 minutes, followed by 35 cycles of 94ºC for 30 seconds, 66.5ºC for 40 seconds, 72ºC for 45 seconds, and then 72ºC for 10 minutes.
HPV DNA was detected by PCR based on the amplification of the viral L1 gene fragment using consensus and degenerate primers MY09 (5’-CGTCCMARRGGAWACTGATC-3’) and MY11 (5’-GCMCAGGGWCATAAYAATGG-3’) [Manos et al., 1989]. The PCR reaction was performed in 25µL volume, containing 2µL (50ng/µL) of the extracted DNA, 12.5µL of GoTaq Green PCR Master Mix (Promega), 1.5µL of each primer (10pM/µL) and 7.5µL of Nuclease-Free Water (Promega). The reaction condition was 94ºC for 1 minute, followed by 35 cycles of 94ºC for 30 seconds, 55ºC for 1 minute, 72ºC for 1 minute, and then 72ºC for 10 minutes.
HPV DNA genotyping was also performed by PCR technique using specific primers for HPV-58 (forward primer: 5’-CCGTTTTGGGTCACATTGTTCATGT-3’ located at positions 7781-7805; reverse primer: 5’-AAGCCTATTTCATCCTCGTCTGAG-3’ located at positions 666-689). The PCR reaction was performed in 25 µL volume, containing 2 µL (50 ng/µL) of the extracted DNA, 12.5 µL of GoTaq Green PCR Master Mix, (Promega), 1.5 µL of each primer (10 pM/µL) and 7.5 µL of Nuclease-Free Water (Promega). The reaction condition was 94ºC for 2 minutes, followed by 35 cycles of 94ºC for 10 seconds, 59ºC for 20 seconds, 72ºC for 1 minute, and then 72ºC for 5 minutes. All samples were amplified in the presence of negative control (milli-Q water). The PCR products obtained were examined on the 2% agarose gel stained with ethidium bromide.
All HPV58 samples were tested for E7 genetic variability using specific primers (E7 forward primer: 5’-ATTTGTCAAAGACAATTGTGTCCAC-3’ located at positions 488-509, E7 reverse primer: 5’-TTAAATCTGTACCACTATCGTCTGC-3’ located at positions 1000-1024). PCR reactions were performed in a final volume of 50 µL, containing 4 µL of the extracted DNA (50 ng/µL), 2 mM MgSO4 (Invitrogen), 0.2 mM dNTP mix (Promega), 10 pM of each primer and 1U Platinum Taq DNA polymerase High Fidelity (Invitrogen). The reaction condition was 94ºC for 2 minutes, followed by 35 cycles of 94ºC for 10 seconds, 58ºC for 20 seconds, 72ºC for 1 minute, and then 72ºC for 5 minutes. The positive fragments were sequenced (both strands) using ABI PRISM BigDye Terminator Cycle Sequencing V.3.1 kit (Applied Biosystems) with the same reverse and forward primers used in the amplification reaction. The obtained sequences were checked for quality and assembled using the Staden package [18]. HPV58 E7 sequences were compared to sequences from GenBank using BLAST [19]. Subsequently, the sequences were aligned with the HPV58 prototype sequence (GenBank: D90400.1) using MEGA (version 7.0) [20].
2.3. Phylogenetic analysis
A Maximum Likelihood phylogenetic tree was reconstructed based on the sequences of E7 gene by using MEGA11[21]. The evolutionary model that best fits the data was used (K80+I), which was selected based on the Bayesian Information Criteria (BIC) using jModelTest 2 [22]. SPR was used as ML heuristic method and moderate branch swap filter. Branch support was assessed with 1000 bootstrap replicates.
The sequences were phylogenetically classified as lineages A, B, C and D, and sublineages A1, A2, A3, B1, B2, C1, D1 and D2. The reference genomes used for this classification were retrieved from PaVE: D90400 (A1), HQ537752 (A2), HQ537758 (A3), HQ537762 (B1), HQ537764 (B2), HQ537774 (C1), HQ537768 (D1) and HQ537770 (D2). Other HPV58 isolate sequences with complete genome were obtained from the NCBI Variant Search link, which was available at PaVE (https://pave.niaid.nih.gov/explore/variants/variant_searches), and they were used in the phylogenetic analysis and displayed in the tree with their GenBank Accession Number.
2.4. Analysis of selective pressures
To evaluate the selective pressure that affects the HPV58 E7 oncogene, estimates of Maximum likelihood were analyzed using the CODEML, incorporated on PAML version 4.9c [Yang, 2007]. The detection of positive selection was performed through the calculation of six codon substitution models parameters, M0, M1, M2, M3, M7 and M8, which uses ω = dN/dS. The basic model uses the ω = dN/dS, that means the ratio of nonsynonymous/synonymous substitution rates; the branch models allow the ω ratio to vary among branches in the phylogeny and are useful for detecting positive selection acting on particular lineages [23,24]; and the site models allow the ω ratio to vary among sites (among codons or amino acids in the protein) [24,25]. The Likelihood Ratio Test (LRT) was used to evaluate which model best fits the data.
2.5. Predicted protein structure modeling
The 3D structure of the reference and mutated E7 protein of HPV58 was in silico modeled. The amino acid sequence of HPV 58 E7 reference protein (BAA31846) was used for the determination of its predicted 3D structure. A blast search was carried out in Protein Data Bank (PDB) using blastp algorithm for template selection. The template search for HPV58 E7 protein showed that there was only the C-terminal of the protein (PDB id: 2F8B) in PDB.
Therefore, the structure model of HPV58 E7 protein was predicted using a combined comparative and ab initio method in Robetta server [26]. The selected models were refined by energy minimization using the 3Drefine server[27]. The predicted models were evaluated by PROCHECK [28], MolProbity [29], QMEAN [30] and TM-align[31]. According to these parameters, the best model was chosen in order to assess the structural variability.
The non-synonymous mutations identified were inserted on the HPV58 E7 reference structure using the Point Mutation service in RosettaBackrub server [32]. The mutated models with the lowest weighted score were selected. The Site Directed Mutator (SDM) method [33] was used to predict the effect of mutations on HPV58 E7 protein structure stability.
2.6. T-cell and B-cell epitopes prediction
2.7. Plasmid constructs
After the analysis of variability in the E7 oncogene sequences (297 bp), three isolates (HPV58/UFPE-54S, HPV58/UFPE-58S and HPV58/UFPE-60M) were chosen because they are more epidemiologically relevant and present more non-synonymous alterations, belonging to variants A, C and D, respectively. These isolates/variants and the prototype (reference gene sequence without changes) of E7 HPV58 were amplified by PCR and cloned into the vector for PCR products (pGEM-T easy - Promega) following the manufacturer's instructions. Subsequently, they were subcloned into a pCDNA 3.1 (+) mammalian cell expression vector. Clones were subjected to automated sequencing using the ABI PRISM BigDyeTM Terminator Cycle Sequencing Kit v3.1 Ready Reaction (Applied Biosystems®) using the ABI Prism 3100 automated DNA sequencer (Applied Biosystem®).
2.8. Transfection of the C33A cells with recombinant expression vectors
After confirming the results by sequencing, the DNA of the recombinant vectors containing the variant sequences of the HPV58 E7 gene and the prototype were isolated by maxi-preparation using the Plasmid Plus Maxi kit (Qiagen) according to instructions. To evaluate the activity of the E7 oncogene using the NF-κB pathway as a model of functionality through luminescence, is was made a co-transfection into keratinocytes resulting from cervical carcinoma (C33A). Transfections were performed in 6-well cell culture plates, where 5x105 cells were plated in 3 ml of culture Dulbecco's ModifiedEagle's Medium (DMEM-Invitrogen®) plus 10% fetal bovine serum (Gibco®); 1% L-Glutamine (Sigma®) - Complete DMEM. Cells were transfected with the recombinant vector constructs using Polyfect transfection reagent (Qiagen). The groups of cells were transfected using the pcDNA3.1 (+) plasmid containing 1.5 µg of the E7 variant, or the prototype (reference gene without any of the studied variations), or even with the empty pcDNA3.1 (+) vector (control negative). Furthermore, all cell groups were co-transfected with an NF-kB dependent firefly luciferase reporter, which is constructed with three kB factor binding sites being Named (kB) 3-Luc (1 μg) (BCCMTM/LMBP, Gent, Belgium) and a plasmid expressing Renilla luciferase (1 ng) as a luminescence normalizer. After 24 hours of transfection, TNFα (10 ng/ml) was added to the plates for 6 hours to stimulate the pathway. All experiments were performed in three experimental replicates in triplicates. Luminescence measurement was performed by the Dual-Luciferase® Reporter Assay System according to the manufacturer's instructions. Relative Luciferase unit readings were performed on the GloMax® 96 Microplate Luminometer w/Dual Injectors (Promega).
2.9. Evaluation of E7 gene expression in transfected C33A cells
Total RNA was obtained from C33A cells transfected with expression vectors for the E7 variants evaluated in this study by the RNeasy mini kit (Qiagen). The cDNA was synthesized using the Improm® Reverse Transcription kit (Promega), according to the manufacturer’s instructions. The qPCR reactions were performed using the QuantiTect SYBR Green ® PCR kit (Qiagen) and the concentrations of primers and cDNA were the same as those described by LEITÃO et al. 2014. Normalization of gene expression adopted the reference genes GAPDH and ACT. The protocol for qPCR was: i) denaturation at 95°C for 5min; ii) 40 cycles of denaturation at 95°C for 10s, annealing at 55°C for 30s (reference genes) or at 60°C (E7, prototype and variants), extension at 72°C for 30s; iii) extension at 72°C for 10 min. Each reaction was performed in three biological replicates, including a negative control for each gene. Gene expression was calculated according to LIVAK; SCHMITTGEN, 2001[37].
2.10. Statistical analysis
Statistical analysis was performed using one-way analysis of variance (ANOVA) followed by Bonferroni correction post-test using GraphPad Prism software. P < 0.05 was considered significant.
3. Results
3.1. HPV detection and typing
Among the 319 collected samples, 227 (71.2 %) were HPV positive. Thereafter the genotyping revealed that 55 (24.2%) samples were positive for HPV16, 36 (15.9%) were positive for HPV31 and 18 (7.9%) were positive for HPV58. Considering the 227 HPV positive samples, 59 samples present a coinfection with more than one HPV type: the results indicated that HPV16 + HPV31 was the most frequent (48, 20.8%), followed by HPV16 + HPV58 (6, 2.6%) and HPV16 + HPV31 + HPV58 (5, 2.2%).
The HPV58 samples were identified as HPV58/UFPE-58S, -30S, -31S, -54S, -88S, -60M, -59M, -1M, -29M, -35P, -45M, -79M, -92M, -28M, -28S and -48S. For these samples the pathological diagnosis was obtained as specified in Table 1. Only samples with a single HPV58 infection were selected for analysis. A total of two samples positive for HPV58 were excluded due to the absence of a pathological diagnosis.
3.2. E7 HPV58 sequence variations
All E7 variants showed nucleotide changes from the prototype, some resulting in codon and amino acid changes. The HPV58/UFPE-54S variant was pathologically classified and the other variants as LSIL.
Nucleotide sequences of the complete E7 oncogene were compared with the HPV58 reference sequence (D90400.1). The comparative analysis of E7 oncogene revealed that sixteen sequences (100%) presented 10 variable sites. Five mutations were missense at codons 41 (G41R), 63 (G63D), 64 (T64A), 74 (T74A) and 76 (D76E) (Table 2). The remaining five mutations were silent (Table 2). The sites that presented the most frequent mutations were 744 (100%, 16 of 16), 761 (93.8%, 15 of 16) and 694 (81.3%, 13 of 16). Other mutations (silent mutations) were T756C (6.3%, 1 of 16), A763G (12.5%, 2 of 16), A793G (18.8%, 3 of 16), C798T (18.8%, 3 of 16), C801A (18.8%, 3 of 16), C840T (18.8%, 3 of 16) and T852C (6.3%, 1 of 16). Nucleotide substitutions resulting in premature stop codons or frameshift changes were not detected.
3.3. Phylogenetic analysis
The evolutionary relationships among HPV58 E7 sequences were analyzed. The phylogenetic tree has shown a lineage and sublineage based clustering of the isolates, which groups HPV 58 E7 variants into four different lineages and eight sublineages (Figure 1). The phylogenetic tree presents well-supported clusters, making the HPV58 E7 variants genotyping possible. Thirteen isolates could be classified as sublineage A2, one as sublineage C1, and two as sublineage D2.
3.4. Selective pressure analysis
The genomic region that includes the E7 gene of HPV58 presented ω ratios less than 1, which indicates purifying selection, acting to conserve the protein function. The model that best fits the data is the one that shows higher values of log-likelihood (lnL). For this genomic region, the best model is the M3, with ω = 0.87940 (Supplementary Table S1). Although the genomic region E7 of HPV58 presented negative or purifying selection, the LRT test indicated the presence of positively (diversifying) selected protein sites, which represents mutations with high probability of being fixed in the viral population (Table 3). In total, five positively selected sites were identified, 41G, 63G, 64T, 74T and 76D. Of these, four sites (41G, 63G, 74T and 76D) were identified to be positively selected with statistical significance (P > 95% and P > 99%) (Table 3).
3.5. Structural analysis of E7 mutations
The E7 protein was modelled based on a hybrid method, the C-terminal with a comparative method and the A-terminal with an ab initio method. The best HPV58 E7 protein model presented 92.9% residues in most favored regions in the Ramachandran plot, MolProbity Score of 0.79, QMEAN score of -1.16, and TM-align score of 0.61714, which represents a well-resolved structure model. All these quality parameters confirm the reliability of the protein structure models.
The HPV58 E7 structure has a total of 98 amino acid residues, with one zinc-binding domain, consisting of two CXXC motifs separated by 29 amino acid residues. The protein structure presents four alpha helix domains and two beta sheet domains, connected with loop regions (Figure 2). The motif 22LXCXE26 consists in part of an alpha helix and a loop region. No non-synonymous mutations found in this study affect directly these motifs. However, two non-synonymous mutations (G63D and T64A) are located in the loop region right after one CXXC motif (Figure 2). The mutation G41R is located in the second alpha helix region, and the mutations T74A and D76E are located in the third alpha helix. Evaluating the impact of these mutations on the protein stability for the HPV58 E7 protein, we observed that the mutations G41R, G63D and T64A presented negative predicted values of ΔΔG, leading to protein instability and malfunction (Table 4).
3.6. T-cell and B-cell epitopes prediction
We performed B-cell epitope predictions in order to evaluate the impact of these mutations on the immune function of the E7 transforming protein. We observed amino acid changes in sites belonging to T-cell (MHC class-I and class-II) and/or B-cell epitopes as reported in Figure 3 which shows the consensus of T-cell and B-cell epitopes obtained for E7 oncoproteins.
3.7. Effect of E7 polymorphic variants on the NF-kB pathway
The functional study of the HPV-58 E7 oncogene variants were developed using luminescence assays having the NF-kB pathway as a model. Initially, the effect on NF-kB activity of the E7 prototype both HPV-16 and HPV-58 was evaluated (Figure 4A and 4B). The results obtained he showed that the E7 prototype of HPV-16 reduced NF-kB activity the action of each variants on NF-kB activity was verify (Figure 4C). The results showed that all variants induced NF-kB compared to the empty vector. However, when compared to the E7HPV-58 prototype, the 58S and 60M variants were similar NF-kB induction compared to the E7 prototype. with the E7 prototype of HPV-58The studied variants were transfected and their effects on the NF-kB pathway were compared with those found in cells expressing the HPV-58 reference E7 (prototype). The difference between the means of the analyzed groups was significant with P-value: 0.0046. However, there was no statistically significant result when comparing the prototype with each variant. Now, the comparisons between pcDNA and the prototype, as well as pcDNA and the HPV58/UFPE-54S variant, were significant with P-value: 0.001 for all comparisons.
3.8. E7 oncoproteingene expression in C33A transfected cells with HPV-58 E7 variants
The reduction in NF-kB activity caused by the E7-54S variant could be due to a low amount of E7 mRNA. In this way, the E7 expression levels of the different variants were evaluated (Figure 4D). The results showed that the relative expression levels of the HPV58/E7-prototype. However, the relative expression level of the 54S variant was significantly lower than HPV58/e7-prototype. Therefore, this result indicates that the low NF-kB activity presented by the HPV58/E7-54S variants may be due to a low mRNA availability.
4. Discussion
Cervical cancer is a serious public health issue and it is strongly linked to high-risk HPVs infections [1]. In Brazil, HPVs 16, 31 and 58 are the most prevalent high-risk subtypes [2]. E7 is considered one of the main oncogenes and nucleotide variations in this oncogene play a crucial function in cervical cancer due to their ability to induce cellular transformation and immortalization[38]. Studies have investigated HPV16 E7 variants [39,40], however the HPV58 E7 variant, which is considered as a high-risk oncogenic type, have been scarcely studied in Brazil.
Therefore, our study has investigated sequence variations of HPV58 E7 oncogene in cervical scraping samples from Brazilian women to evaluate their structural effects in order to identity the possible impact of these mutations in the carcinogenic process.
In this study, nucleotide changes previously described in HPV58 E7 oncogene were detected at positions G694A[41,42,43], T744G [40,41,42,44], T756C [40], G761A [40,41,42,44], A763G [7], A793G, C798T, C801A, C840T and T852C [41,45].
From the phylogenetic point of view, HPV lineages have been associated with different risks for cervical cancer development as observed for some HPV16 lineages [46,47,48]. With respect to HPV58 phylogeny, sequence variants are associated with the risk for CIN3 (cervical intraepithelial neoplasia grade 3) and invasive cervical cancer [7]. HPV58 consists of four variant lineages (A, B, C, and D) and eight sublineages (A1, A2, A3, B1, B2, C, D1, and D2) [49]. In this study, our analysis on the E7 sequences of HPV58 variants identified the presence of the sublineages A2 (the majority), C1 and D2. It is important to point out that the lineage determination of HPV rely on complete genome sequences and a possible bias might occur. Therefore, in order to confirm the lineage classification in this study, a complete genome sequence comparison and phylogenetic analysis were performed with the reference HPV58 complete genome sequences. Once the lineage classification was confirmed for the reference genome sequences, the phylogenetic analysis performed with the E7 gene sequences (Figure 1) showed a similar topology, which indicates the probable lineage of the isolates.
In a study performed by Chan et al. (2011) an association between oncogenic risk and variant lineage was analysed. Lineage A represented the majority of HSIL/carcinoma samples, however there was no significant association [50].In the present study, most of the A2 line is represented by HSIL samples. Nevertheless, further studies are necessary to evaluate the association of variant lineages and cancer cervical risk.
The analysis indicated that the variant E7 genomic region of HPV58 presented negative or purifying selection. However, evolutionary diversifying selected amino acid residues have been identified in this study. The diversifying selection is associated with mutations with high probability of being fixed in the viral population, being related to the host adaptive response [51]. In this study, we have identified four non-synonymous mutations under diversifying selection in E7 protein: 41G, 63G, 74T and 76D. These mutations are consistently being found in other studies worldwide, which highlights their evolutionary relevance for the virus [52,42,53]. Although more experimental evidences must be found, these findings are relevant because these mutations might be related to adaptation to the human host, increasing the ability of these oncoproteins to interact with the host proteins.
It is widely known that the E7 gene interact with various biological pathways and plays an important role in oncogenesis [54,55,56]. Therefore, genetics variants of E7 may exert impacts in these pathways both by altering protein-protein interaction than affect their stability [57,58]. Some studies showed that E7 oncogene variants suppressed the activity of the NF-kB pathway[59,60]. The HPV16 E7 protein interferes with NF-κB signaling, interacting directly with the IKKα and IKKβ subunits, causing a reduction in the activity related to NF-κB signaling in U2OS cells [60]. Even under TNF-α cytokine stimulation, U2OS cells transfected with the HPV-16 E7 gene were less responsive, exhibiting attenuated NF-κB signaling[61].
In a study by Silva et al. (2020), which evaluated the activity of the NF-kB pathway mediated by variants of the E5 HPV-31 gene, it was reported that all variants increased activity of the pathway [62]. Nucleotide variations present in the HPV E7 oncogene play an important role in the development of cervical cancer, this is due to its ability to inactivate products of important tumor suppressor genes, such as pRb. and malignant transformation of virus-infected cells [63]. The NF-kB pathway and its activation are a frequent event in squamous cell carcinoma, as well as important evidence of epithelial cell modification [64]. Several studies have been carried out investigating variants of HPV 16 E5, for example [65,66,67], while variants of HPV 58 E7 are poorly studied, although HPV -58 is part of the high-risk group of HPVs and is a very prevalent type in some countries of the world, including some regions of Brazil.
In this study, the E7 prototype was shown to activate the pathway in about 2x when compared to pcDNA. Albeit all the tested variants activate the NF-kB pathways, we observed that the HPV58/UFPE-54S variant seems exhibit a decreased level of activation in comparison to prototype and the other tested variants. It is likely that this difference is due to the lower E7 expression observed in HPV58/UFPE-54S compared both protype and the other tested variants. Although we did not test the stability, bioavailability, or decay rate of the analyzed variant sequences, it is possible that this reduced expression level may be attributed to some of these mechanisms[68,69,70].
Futhermore, in the structural analysis for E7, LXCXE motif is known to be related with the interaction of E7 with pRB. However, the non-synonymous mutations detected in this study did not alter the LXCXE motif. Two mutations (G63D and T64A) are located in the loop right after the CXXC motif, an important zinc-binding motif. G63D has been reported to has a significantly lower risk association with cervical neoplasia[7]. Other mutations detected in this study (G41R, T74A and D76E) are located in the second and third alpha-helices, respectively. In a previous study, G63D and G41R have been reported to show a significantly lower risk association with cervical neoplasia [7]. Moreover, the mutations G41R, G63D and T64A in E7 presented were predicted to lead to protein instability and malfunction. Interestingly, the variant with bellow NF-kB activation (HPV58/UFPE-54S) exhibits two mutations that lead to decreased stability (G41R, G63D).
To the best of our knowledge, this is the first time that these mutations have been structurally characterized. However, the impact of these mutations on the interaction of these HPV oncoprotein and host molecules still needs to be assessed in further experimental studies.
5. Conclusions
In this study, nucleotide changes previously described in HPV58 oncogene E7 were detected. It was observed that these alterations may alter the molecular interaction between viral oncoproteins and host proteins. These amino acid changes in the HPV58 E7 oncoprotein may influence the immune recognition of cells infected by HPV. However, further studies are needed in order to confirm the relationship between these changes and the risk of cervical cancer. The interaction of the E7 oncogene with NF-kB signaling provides important information for a better understanding of how variants can interfere in the carcinogenesis process.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Bárbara S Chagas and Elias T Júnior carried out the main axis of the experiments. Bárbara S Chagas and Ruany C O Silva wrote the manuscript; Ruany C O Silva, Marconi R Barros Junior, Rita C P Lima and Antônio H P Silva Júnior participated in some of the experiments; Maria C V Invenção and Pedro L F Neto contributes with final revision of this manuscript, Jacinto C Silva Neto gave support in obtaining biological samples; Marcus V A Batista performed all bioinformatics analysis and was responsible for by the final revision of this manuscript; Antonio C Freitas was responsible for experimental design, supervision of all experimental steps, and by the final revision of this manuscript.
Funding
This research was funded by Fundação de Amparo à Ciência e Tecnologia do Estado de Pernambuco (FACEPE/PRONEM - APQ-0562-2.02/14; FACEPE/PPSUS - APQ-0748-2.02/17), FAPITEC/SE (00941/2018-5), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPQ 401202/2020-8; CNPq 401305/2014-7; CNPq 447978/2014-4), and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (Estágio Pós-Doutoral do Programa Nacional de Pós-Doutorado/Capes – PNPD/CAPES 2020-2021 – Programa de Pós-Graduação em Biotecnologia; CAPES 23038.010050/2013-04; CAPES Bolsa de Produtividade 2020-2023).
Conflicts of Interest
The authors declare no conflict of interest.
References
- zur Hausen, H. Papillomaviruses in the Causation of Human Cancers - a Brief Historical Account. Virology 2009, 384, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Raiol, T.; Wyant, P.S.; de Amorim, R.M.S.; Cerqueira, D.M.; Milanezi, N. von G.; Brígido, M. de M.; Sichero, L.; Martins, C.R.F. Genetic Variability and Phylogeny of the High-Risk {HPV}-31, -33, -35, -52, and -58 in Central {Brazil}. J. Med. Virol. 2009, 81, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Arbyn, M.; Weiderpass, E.; Bruni, L.; de Sanjosé, S.; Saraiya, M.; Ferlay, J.; Bray, F. Estimates of Incidence and Mortality of Cervical Cancer in 2018: A Worldwide Analysis. Lancet. Glob. Heal. 2020, 8, e191–e203. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: {GLOBOCAN} Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA. Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Gurgel, A.P.A.D.; Chagas, B.S.; do Amaral, C.M.; Nascimento, K.C.G.; Leal, L.R.S.; Silva Neto, J. da C.; Cartaxo Muniz, M.T.; de Freitas, A.C. Prevalence of Human Papillomavirus Variants and Genetic Diversity in the {L1} Gene and Long Control Region of {HPV16}, {HPV31}, and {HPV58} Found in {North}-{East} {Brazil}. Biomed Res. Int. 2015, 2015, 130828. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.; Carvalho, M.; de Fernandes, T.; Araújo, J.; Azevedo, P.; Azevedo, J.; Meissner, R. Prevalence of Human Papillomavirus Type 58 in Women with or without Cervical Lesions in Northeast {Brazil}. Ann. Med. Health Sci. Res. 2013, 3, 504–510. [Google Scholar] [CrossRef]
- Chan, P.K.S.; Zhang, C.; Park, J.-S.; Smith-McCune, K.K.; Palefsky, J.M.; Giovannelli, L.; Coutlée, F.; Hibbitts, S.; Konno, R.; Settheetham-Ishida, W.; et al. Geographical Distribution and Oncogenic Risk Association of Human Papillomavirus Type 58 {E6} and {E7} Sequence Variations. Int. J. Cancer 2013, 132, 2528–2536. [Google Scholar] [CrossRef]
- Chagas, B.S.; Batista, M.V.A.; Guimarães, V.; Balbino, V.Q.; Crovella, S.; Freitas, A.C. New Variants of {E6} and {E7} Oncogenes of Human Papillomavirus Type 31 Identified in {Northeastern} {Brazil}. Gynecol. Oncol. 2011, 123, 284–288. [Google Scholar] [CrossRef]
- Boon, S.S.; Xia, C.; Lim, J.Y.; Chen, Z.; Law, P.T.Y.; Yeung, A.C.M.; Thomas, M.; Banks, L.; Chan, P.K.S. Human {Papillomavirus} 58 {E7} {T20I}/{G63S} {Variant} {Isolated} from an {East} {Asian} {Population} {Possesses} {High} {Oncogenicity}. J. Virol. 2020, 94, e00090–20. [Google Scholar] [CrossRef]
- Xi, L.F.; Koutsky, L.A.; Hildesheim, A.; Galloway, D.A.; Wheeler, C.M.; Winer, R.L.; Ho, J.; Kiviat, N.B. Risk for High-Grade Cervical Intraepithelial Neoplasia Associated with Variants of Human Papillomavirus Types 16 and 18. Cancer Epidemiol. Biomarkers \& Prev. A Publ. Am. Assoc. Cancer Res. Cosponsored by Am. Soc. Prev. Oncol. 2007, 16, 4–10. [Google Scholar] [CrossRef]
- De Boer, M.A.; Peters, L.A.W.; Aziz, M.F.; Siregar, B.; Cornain, S.; Vrede, M.A.; Jordanova, E.S.; Fleuren, G.J. Human Papillomavirus Type 18 Variants: Histopathology and {E6}/{E7} Polymorphisms in Three Countries. Int. J. Cancer 2005, 114, 422–425. [Google Scholar] [CrossRef]
- Law, P.T.; Boon, S.S.; Hu, C.; Lung, R.W.; Cheung, G.P.; Ho, W.C.; Chen, Z.; Massimi, P.; Thomas, M.; Pim, D.; et al. Oncogenic Comparison of Human Papillomavirus Type 58 {E7} Variants. J. Cell. Mol. Med. 2019, 23, 1517–1527. [Google Scholar] [CrossRef]
- Ding, T.; Wang, X.; Ye, F.; Cheng, X.; Lu, W.; Xie, X. Distribution of Human Papillomavirus 16 {E6}/{E7} Variants in Cervical Cancer and Intraepithelial Neoplasia in {Chinese} Women. Int. J. Gynecol. Cancer Off. J. Int. Gynecol. Cancer Soc. 2010, 20, 1391–1398. [Google Scholar] [CrossRef]
- He, J.; Yang, Y.; Chen, Z.; Liu, Y.; Bao, S.; Zhao, Y.; Ding, X. Identification of Variants and Therapeutic Epitopes in {HPV}-33/{HPV}-58 {E6} and {E7} in {Southwest} {China}. Virol. J. 2019, 16, 72. [Google Scholar] [CrossRef] [PubMed]
- Barros, M.R.; de Oliveira, T.H.A.; de Melo, C.M.L.; Venuti, A.; de Freitas, A.C. Viral {Modulation} of {TLRs} and {Cytokines} and the {Related} {Immunotherapies} for {HPV}-{Associated} {Cancers}. J. Immunol. Res. 2018, 2018, 2912671. [Google Scholar] [CrossRef]
- Barros, M.R.; de Melo, C.M.L.; Barros, M.L.C.M.G.R.; de Cássia Pereira de Lima, R.; de Freitas, A.C.; Venuti, A. Activities of Stromal and Immune Cells in {HPV}-Related Cancers. J. Exp. \& Clin. cancer Res. CR 2018, 37, 137. [Google Scholar] [CrossRef] [PubMed]
- de Freitas, A.C.; Gurgel, A.P.A.D.; Chagas, B.S.; Coimbra, E.C.; do Amaral, C.M.M. Susceptibility to Cervical Cancer: An Overview. Gynecol. Oncol. 2012, 126, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Staden, R. The {Staden} Sequence Analysis Package. Mol. Biotechnol. 1996, 5, 233–241. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. {MEGA7}: {Molecular} {Evolutionary} {Genetics} {Analysis} {Version} 7.0 for {Bigger} {Datasets}. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. {MEGA11}: {Molecular} {Evolutionary} {Genetics} {Analysis} {Version} 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. {jModelTest} 2: More Models, New Heuristics and Parallel Computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Yang, Z. Likelihood Ratio Tests for Detecting Positive Selection and Application to Primate Lysozyme Evolution. Mol. Biol. Evol. 1998, 15, 568–573. [Google Scholar] [CrossRef]
- Yang, Z.; Nielsen, R. Synonymous and Nonsynonymous Rate Variation in Nuclear Genes of Mammals. J. Mol. Evol. 1998, 46, 409–418. [Google Scholar] [CrossRef]
- Yang, Z. Complexity of the Simplest Phylogenetic Estimation Problem. Proceedings. Biol. Sci. 2000, 267, 109–116. [Google Scholar] [CrossRef]
- Kim, D.E.; Chivian, D.; Baker, D. Protein Structure Prediction and Analysis Using the {Robetta} Server. Nucleic Acids Res. 2004, 32, W526–531. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, D.; Nowotny, J.; Cao, R.; Cheng, J. {3Drefine}: An Interactive Web Server for Efficient Protein Structure Refinement. Nucleic Acids Res. 2016, 44, W406–409. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. {PROCHECK}: A Program to Check the Stereochemical Quality of Protein Structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Chen, V.B.; Arendall, W.B.; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. {MolProbity}: All-Atom Structure Validation for Macromolecular Crystallography. Acta Crystallogr. D. Biol. Crystallogr. 2010, 66, 12–21. [Google Scholar] [CrossRef]
- Benkert, P.; Biasini, M.; Schwede, T. Toward the Estimation of the Absolute Quality of Individual Protein Structure Models. Bioinformatics 2011, 27, 343–350. [Google Scholar] [CrossRef]
- Zhang, Y.; Skolnick, J. {TM}-Align: A Protein Structure Alignment Algorithm Based on the {TM}-Score. Nucleic Acids Res. 2005, 33, 2302–2309. [Google Scholar] [CrossRef] [PubMed]
- Lauck, F.; Smith, C.A.; Friedland, G.F.; Humphris, E.L.; Kortemme, T. {RosettaBackrub}--a Web Server for Flexible Backbone Protein Structure Modeling and Design. Nucleic Acids Res. 2010, 38, W569–575. [Google Scholar] [CrossRef] [PubMed]
- Worth, C.L.; Preissner, R.; Blundell, T.L. {SDM}--a Server for Predicting Effects of Mutations on Protein Stability and Malfunction. Nucleic Acids Res. 2011, 39, W215–222. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Raghava, G.P.S. {ProPred1}: Prediction of Promiscuous {MHC} {Class}-{I} Binding Sites. Bioinformatics 2003, 19, 1009–1014. [Google Scholar] [CrossRef]
- Singh, H.; Raghava, G.P. {ProPred}: Prediction of {HLA}-{DR} Binding Sites. Bioinformatics 2001, 17, 1236–1237. [Google Scholar] [CrossRef]
- Saha, S.; Raghava, G.P.S. {AlgPred}: Prediction of Allergenic Proteins and Mapping of {IgE} Epitopes. Nucleic Acids Res. 2006, 34, W202–209. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Roman, A.; Munger, K. The Papillomavirus {E7} Proteins. Virology 2013, 445, 138–168. [Google Scholar] [CrossRef]
- Ramas, V.; Mirazo, S.; Bonilla, S.; Ruchansky, D.; Arbiza, J. Analysis of Human Papillomavirus 16 {E6}, {E7} Genes and {Long} {Control} {Region} in Cervical Samples from {Uruguayan} Women. Gene 2018, 654, 103–109. [Google Scholar] [CrossRef]
- Yu, J.-H.; Shi, W.-W.; Zhou, M.-Y.; Liu, J.-M.; Han, Q.-Y.; Xu, H.-H. Genetic Variability and Oncogenic Risk Association of Human Papillomavirus Type 58 {E6} and {E7} Genes in {Taizhou} Area, {China}. Gene 2019, 686, 171–176. [Google Scholar] [CrossRef]
- Molano, M.; Buitrago, O.; Moreno-Acosta, P.; Garland, S.M.; Morales, N.; Huertas, A.; Martinez, T.; Gamboa, O.; Tabrizi, S.N.; Cornall, A.; et al. Follow-up Study of {HPV58} Variants in Women with Incident {HPV58} Infection from a {Colombian} Cohort. Oncol. Lett. 2018, 16, 2511–2516. [Google Scholar] [CrossRef] [PubMed]
- Xi, J.; Chen, J.; Xu, M.; Yang, H.; Wen, S.; Pan, Y.; Wang, X.; Ye, C.; Qiu, L.; Sun, Q. The Polymorphisms of {LCR}, {E6}, and {E7} of {HPV}-58 Isolates in {Yunnan}, {Southwest} {China}. Virol. J. 2018, 15, 76. [Google Scholar] [CrossRef]
- Zhou, Z.; Yang, H.; Yang, L.; Yao, Y.; Dai, S.; Shi, L.; Li, C.; Yang, L.; Yan, Z.; Yao, Y. Human Papillomavirus Type 16 {E6} and {E7} Gene Variations Associated with Cervical Cancer in a {Han} {Chinese} Population. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2019, 73, 13–20. [Google Scholar] [CrossRef]
- Conde-Ferraez, L.; Pacheco-Arjona, R.; Novelo Canul, C.; Gomez-Carballo, J.; Ramirez-Prado, J.H.; Ayora-Talavera, G.; González-Losa, M.D.R. Genetic {Variability} in {E6} and {E7} {Oncogenes} from {Human} {Papillomavirus} {Type} 58 in {Mexican} {Women}. Intervirology 2017, 60, 235–246. [Google Scholar] [CrossRef]
- Tenjimbayashi, Y.; Onuki, M.; Hirose, Y.; Mori, S.; Ishii, Y.; Takeuchi, T.; Tasaka, N.; Satoh, T.; Morisada, T.; Iwata, T.; et al. Whole-Genome Analysis of Human Papillomavirus Genotypes 52 and 58 Isolated from {Japanese} Women with Cervical Intraepithelial Neoplasia and Invasive Cervical Cancer. Infect. Agent. Cancer 2017, 12, 44. [Google Scholar] [CrossRef] [PubMed]
- Berumen, J.; Ordoñez, R.M.; Lazcano, E.; Salmeron, J.; Galvan, S.C.; Estrada, R.A.; Yunes, E.; Garcia-Carranca, A.; Gonzalez-Lira, G.; la Campa, A. Asian-{American} Variants of Human Papillomavirus 16 and Risk for Cervical Cancer: A Case-Control Study. J. Natl. Cancer Inst. 2001, 93, 1325–1330. [Google Scholar] [CrossRef] [PubMed]
- Zuna, R.E.; Moore, W.E.; Shanesmith, R.P.; Dunn, S.T.; Wang, S.S.; Schiffman, M.; Blakey, G.L.; Teel, T. Association of {HPV16} {E6} Variants with Diagnostic Severity in Cervical Cytology Samples of 354 Women in a {US} Population. Int. J. Cancer 2009, 125, 2609–2613. [Google Scholar] [CrossRef]
- Cornet, I.; Gheit, T.; Iannacone, M.R.; Vignat, J.; Sylla, B.S.; Del Mistro, A.; Franceschi, S.; Tommasino, M.; Clifford, G.M. {HPV16} Genetic Variation and the Development of Cervical Cancer Worldwide. Br. J. Cancer 2013, 108, 240–244. [Google Scholar] [CrossRef]
- Burk, R.D.; Harari, A.; Chen, Z. Human Papillomavirus Genome Variants. Virology 2013, 445, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.K.S.; Luk, A.C.S.; Park, J.-S.; Smith-McCune, K.K.; Palefsky, J.M.; Konno, R.; Giovannelli, L.; Coutlée, F.; Hibbitts, S.; Chu, T.-Y.; et al. Identification of Human Papillomavirus Type 58 Lineages and the Distribution Worldwide. J. Infect. Dis. 2011, 203, 1565–1573. [Google Scholar] [CrossRef]
- Dearlove, B.; Lewitus, E.; Bai, H.; Li, Y.; Reeves, D.B.; Joyce, M.G.; Scott, P.T.; Amare, M.F.; Vasan, S.; Michael, N.L.; et al. A {SARS}-{CoV}-2 Vaccine Candidate Would Likely Match All Currently Circulating Variants. Proc. Natl. Acad. Sci. U. S. A. 2020, 117, 23652–23662. [Google Scholar] [CrossRef]
- Bao, E.L.; Cheng, A.N.; Sankaran, V.G. The Genetics of Human Hematopoiesis and Its Disruption in Disease. EMBO Mol. Med. 2019, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhang, C.; Luo, P.; Ye, M.; Gong, Q.; Mei, B. Genetic Variability of {E6} and {E7} Genes of Human Papillomavirus Type 58 in {Jingzhou}, {Hubei} {Province} of Central {China}. Virol. J. 2022, 19, 71. [Google Scholar] [CrossRef] [PubMed]
- Bodily, J.M.; Mehta, K.P.M.; Laimins, L.A. Human Papillomavirus {E7} Enhances Hypoxia-Inducible Factor 1-Mediated Transcription by Inhibiting Binding of Histone Deacetylases. Cancer Res. 2011, 71, 1187–1195. [Google Scholar] [CrossRef] [PubMed]
- Honegger, A.; Schilling, D.; Bastian, S.; Sponagel, J.; Kuryshev, V.; Sültmann, H.; Scheffner, M.; Hoppe-Seyler, K.; Hoppe-Seyler, F. Dependence of Intracellular and Exosomal {microRNAs} on Viral {E6}/{E7} Oncogene Expression in {HPV}-Positive Tumor Cells. PLoS Pathog. 2015, 11, e1004712. [Google Scholar] [CrossRef] [PubMed]
- White, E.A.; Sowa, M.E.; Tan, M.J.A.; Jeudy, S.; Hayes, S.D.; Santha, S.; Münger, K.; Harper, J.W.; Howley, P.M. Systematic Identification of Interactions between Host Cell Proteins and {E7} Oncoproteins from Diverse Human Papillomaviruses. Proc. Natl. Acad. Sci. U. S. A. 2012, 109, E260–267. [Google Scholar] [CrossRef]
- Lou, H.; Boland, J.F.; Li, H.; Burk, R.; Yeager, M.; Anderson, S.K.; Wentzensen, N.; Schiffman, M.; Mirabello, L.; Dean, M. {HPV16} {E7} {Nucleotide} {Variants} {Found} in {Cancer}-{Free} {Subjects} {Affect} {E7} {Protein} {Expression} and {Transformation}. Cancers (Basel). 2022, 14, 4895. [Google Scholar] [CrossRef]
- Nunes, E.M.; Talpe-Nunes, V.; Sobrinho, J.S.; Ferreira, S.; Lino, V. de S.; Termini, L.; Silva, G.Á.F.; Boccardo, E.; Villa, L.L.; Sichero, L. E6/{E7} {Functional} {Differences} among {Two} {Natural} {Human} {Papillomavirus} 18 {Variants} in {Human} {Keratinocytes}. Viruses 2021, 13, 1114. [Google Scholar] [CrossRef]
- Vandermark, E.R.; Deluca, K.A.; Gardner, C.R.; Marker, D.F.; Schreiner, C.N.; Strickland, D.A.; Wilton, K.M.; Mondal, S.; Woodworth, C.D. Human Papillomavirus Type 16 {E6} and {E} 7 Proteins Alter {NF}-{kB} in Cultured Cervical Epithelial Cells and Inhibition of {NF}-{kB} Promotes Cell Growth and Immortalization. Virology 2012, 425, 53–60. [Google Scholar] [CrossRef]
- Spitkovsky, D.; Hehner, S.P.; Hofmann, T.G.; Möller, A.; Schmitz, M.L. The Human Papillomavirus Oncoprotein {E7} Attenuates {NF}-Kappa {B} Activation by Targeting the {Ikappa} {B} Kinase Complex. J. Biol. Chem. 2002, 277, 25576–25582. [Google Scholar] [CrossRef]
- Byg, L.M.; Vidlund, J.; Vasiljevic, N.; Clausen, D.; Forslund, O.; Norrild, B. {NF}-Κ{B} Signalling Is Attenuated by the {E7} Protein from Cutaneous Human Papillomaviruses. Virus Res. 2012, 169, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Silva, R.C. de O.; da Silva Júnior, A.H.P.; Gurgel, A.P.A.D.; Barros Junior, M.R.; Santos, D.L.; de Lima, R. de C.P.; Batista, M.V.A.; Pena, L.J.; Chagas, B.S.; Freitas, A.C. Structural and Functional Impacts of {E5} Genetic Variants of Human Papillomavirus Type 31. Virus Res. 2020, 290, 198143. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin-Drubin, M.E.; Münger, K. Oncogenic Activities of Human Papillomaviruses. Virus Res. 2009, 143, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Huber, M.A.; Azoitei, N.; Baumann, B.; Grünert, S.; Sommer, A.; Pehamberger, H.; Kraut, N.; Beug, H.; Wirth, T. {NF}-{kappaB} Is Essential for Epithelial-Mesenchymal Transition and Metastasis in a Model of Breast Cancer Progression. J. Clin. Invest. 2004, 114, 569–581. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Pang, T.; Guo, Z.; Pontén, J.; Nistér, M.; Bernard Afink, G. Oncogene Lineages of Human Papillomavirus Type 16 {E6}, {E7} and {E5} in Preinvasive and Invasive Cervical Squamous Cell Carcinoma. J. Pathol. 2001, 195, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, B.; Mandal, N.R.; Roy, S.; Sengupta, S. Characterization of Sequence Variations within {HPV16} Isolates among {Indian} Women: Prediction of Causal Role of Rare Non-Synonymous Variations within Intact Isolates in Cervical Cancer Pathogenesis. Virology 2008, 377, 143–150. [Google Scholar] [CrossRef]
- Makowsky, R.; Lhaki, P.; Wiener, H.W.; Bhatta, M.P.; Cullen, M.; Johnson, D.C.; Perry, R.T.; Lama, M.; Boland, J.F.; Yeager, M.; et al. Genomic Diversity and Phylogenetic Relationships of Human Papillomavirus 16 ({HPV16}) in {Nepal}. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2016, 46, 7–11. [Google Scholar] [CrossRef]
- Bicknell, A.A.; Ricci, E.P. When {mRNA} Translation Meets Decay. Biochem. Soc. Trans. 2017, 45, 339–351. [Google Scholar] [CrossRef]
- Hanson, G.; Coller, J. Codon Optimality, Bias and Usage in Translation and {mRNA} Decay. Nat. Rev. Mol. Cell Biol. 2018, 19, 20–30. [Google Scholar] [CrossRef]
- Jia, L.; Mao, Y.; Ji, Q.; Dersh, D.; Yewdell, J.W.; Qian, S.-B. Decoding {mRNA} Translatability and Stability from the 5′ {UTR}. Nat. Struct. \& Mol. Biol. 2020, 27, 814–821. [Google Scholar] [CrossRef]
Figure 1.
Midpoint-rooted Maximum Likelihood phylogenetic tree of HPV 58 strains based on E7 gene sequences. Four clusters were identified as lineages A, B, C and D, and eight sub-clusters are identified as sublineages. Bootstrap replicates are represented as branch support.
Figure 1.
Midpoint-rooted Maximum Likelihood phylogenetic tree of HPV 58 strains based on E7 gene sequences. Four clusters were identified as lineages A, B, C and D, and eight sub-clusters are identified as sublineages. Bootstrap replicates are represented as branch support.
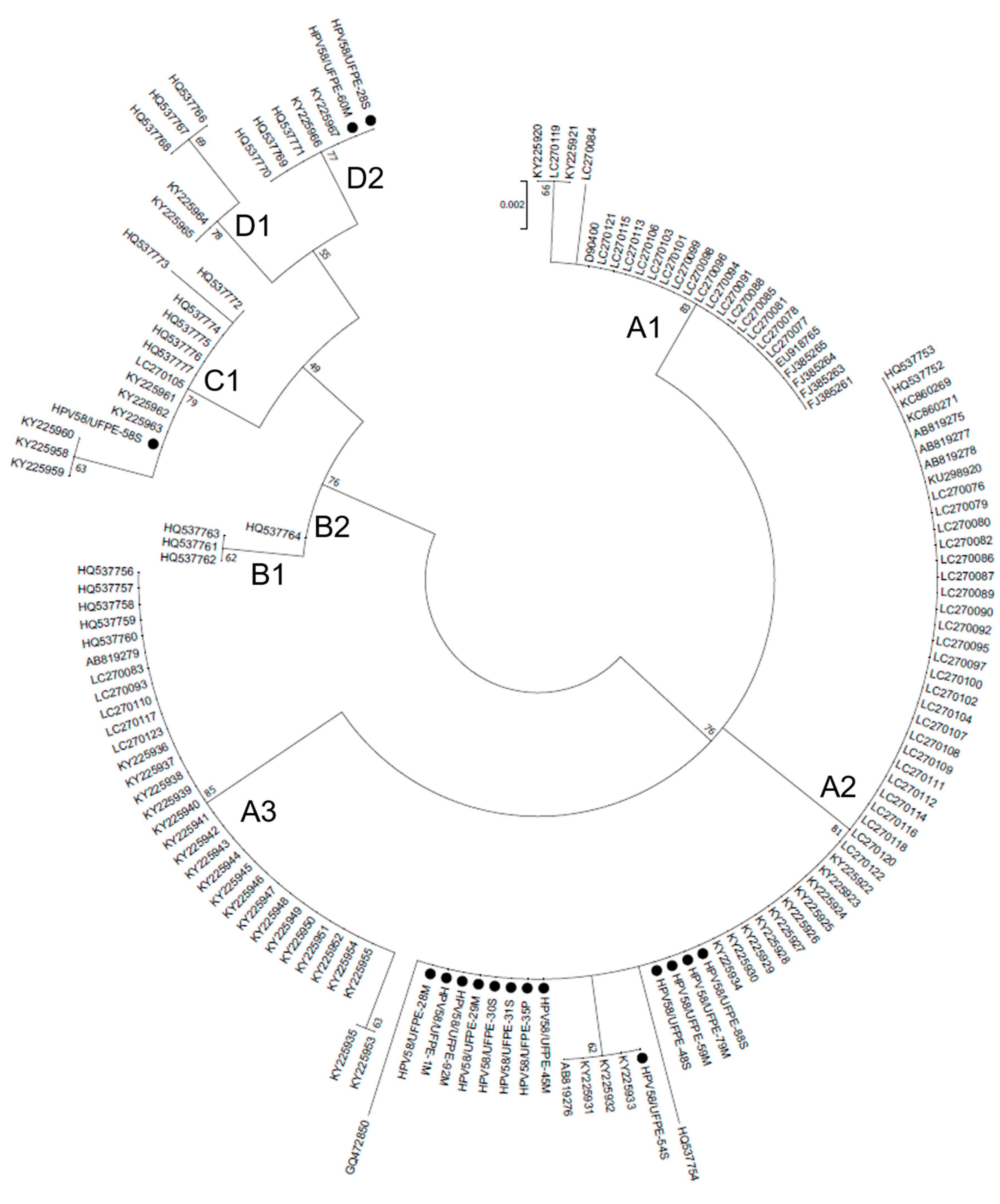
Figure 2.
Representation of HPV58 E7 protein model as displayed in Discovery Studio Visualizer. Non-synonymous mutations found in this study are represented in the structure. Four alpha helices are represented in red, beta sheets in blue, and loops in gray and green.
Figure 2.
Representation of HPV58 E7 protein model as displayed in Discovery Studio Visualizer. Non-synonymous mutations found in this study are represented in the structure. Four alpha helices are represented in red, beta sheets in blue, and loops in gray and green.
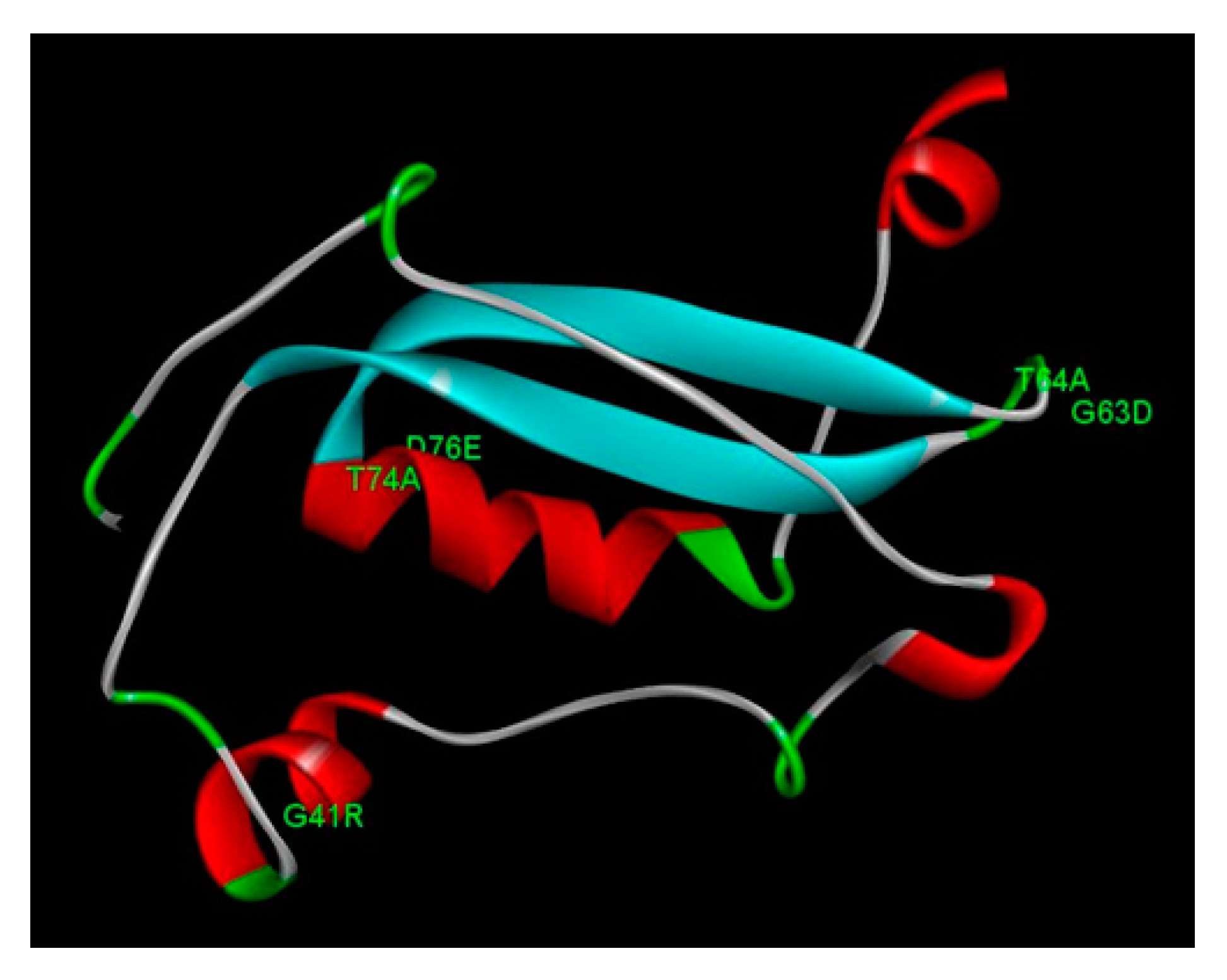
Figure 3.
Predicted T-cell and B-cell epitopes. Epitopes within the E7 oncoprotein of HPV58 variants. In blue, a consensus of the predicted T-cell epitopes. In green, a consensus of the predicted B-cell epitopes. Red arrows indicate mutations sites presented in E7 HPV58 isolates.
Figure 3.
Predicted T-cell and B-cell epitopes. Epitopes within the E7 oncoprotein of HPV58 variants. In blue, a consensus of the predicted T-cell epitopes. In green, a consensus of the predicted B-cell epitopes. Red arrows indicate mutations sites presented in E7 HPV58 isolates.

Figure 4.
Effect of the E7 oncoprotein from HPV 16 (A) and HPV 58 (B) on NF-kB activity. The E7 oncoprotein from HPV16 reduces NF-kB activity, while the E7 from HPV58 increases it. (C) Effect of the HPV58 prototype and variants on NF-kB activity. The 58S and 60M variants do not exhibit a statistically significant difference compared to the prototype. However, the 54S variant, while lacking statistical significance, appears to demonstrate a lower activity in the NF-kB pathway. (D) Comparison of gene expression among different HPV58 variants and the prototype. While the 58S and 60M variants did not exhibit a significant difference in expression compared to the prototype, the 54S variant showed a notably reduced expression level.
Figure 4.
Effect of the E7 oncoprotein from HPV 16 (A) and HPV 58 (B) on NF-kB activity. The E7 oncoprotein from HPV16 reduces NF-kB activity, while the E7 from HPV58 increases it. (C) Effect of the HPV58 prototype and variants on NF-kB activity. The 58S and 60M variants do not exhibit a statistically significant difference compared to the prototype. However, the 54S variant, while lacking statistical significance, appears to demonstrate a lower activity in the NF-kB pathway. (D) Comparison of gene expression among different HPV58 variants and the prototype. While the 58S and 60M variants did not exhibit a significant difference in expression compared to the prototype, the 54S variant showed a notably reduced expression level.
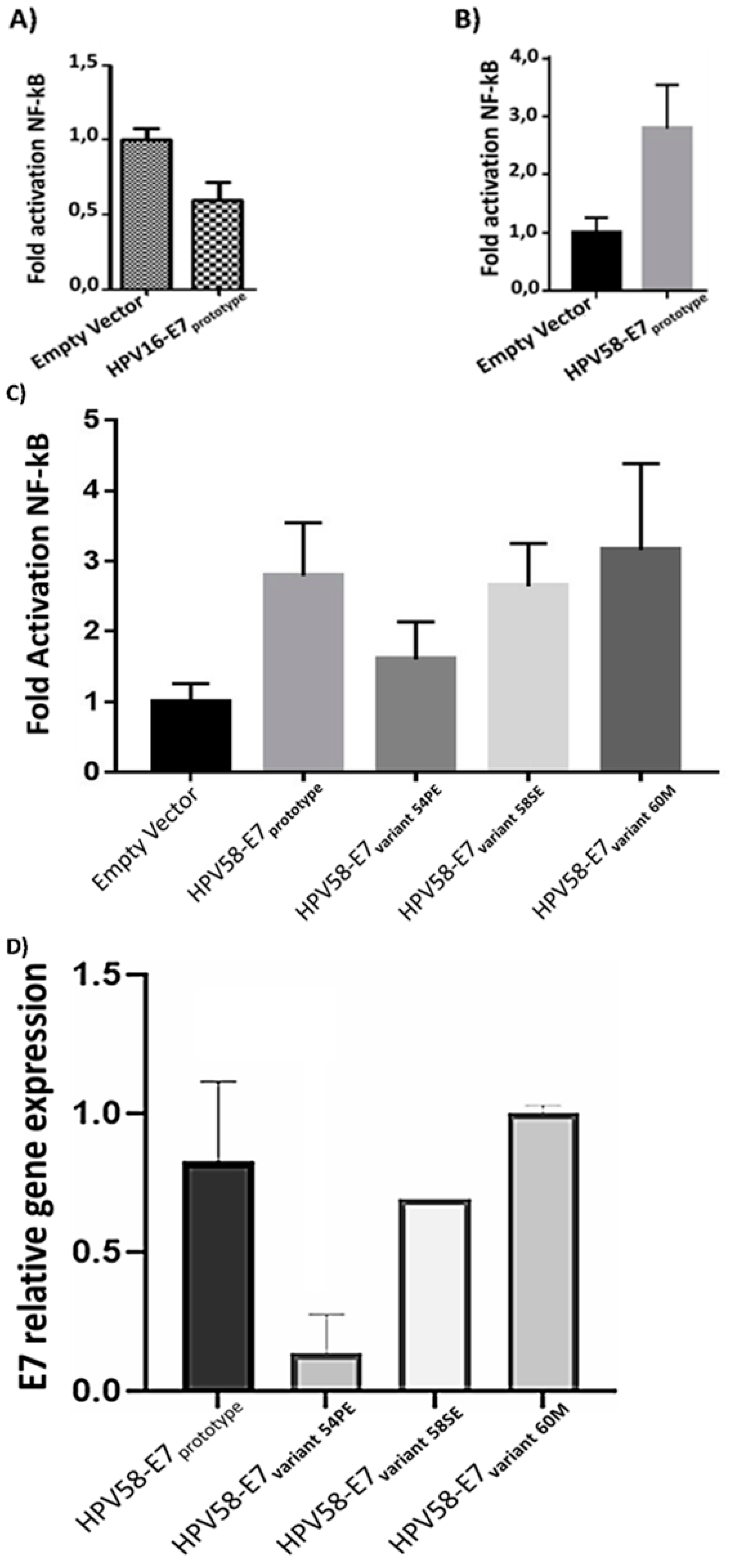

Table 1.
Pathological diagnosis of HPV58 positive samples.
| Samples | Pathological diagnosis |
|---|---|
| HPV58/UFPE-58S | LSIL |
| HPV58/UFPE-30S | HSIL |
| HPV58/UFPE-31S | HSIL |
| HPV58/UFPE-54S | HSIL |
| HPV58/UFPE-88S | LSIL |
| HPV58/UFPE-60M | LSIL |
| HPV58/UFPE-59M | HSIL |
| HPV58/UFPE-1M | HSIL |
| HPV58/UFPE-29M | Without lesion |
| HPV58/UFPE-35P | Without lesion |
| HPV58/UFPE-45M | HSIL |
| HPV58/UFPE-79M | HSIL |
| HPV58/UFPE-92M | HSIL |
| HPV58/UFPE-28M | LSIL |
| HPV58/UFPE-28S | HSIL |
| HPV58/UFPE-48S | HSIL |
LSIL: low-grade squamous intraepithelial lesion. HSIL: high-grade squamous intraepithelial lesion.
Table 2.
Nucleotide and amino acid variability of HPV58 E7 region and the putative biological function affected. Conservative nucleotides with respect to the reference sequence (D90400.1) are marked with a dot (.). Reference: prototype sequence of HPV58.
Table 2.
Nucleotide and amino acid variability of HPV58 E7 region and the putative biological function affected. Conservative nucleotides with respect to the reference sequence (D90400.1) are marked with a dot (.). Reference: prototype sequence of HPV58.
| E7 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Nt | 694 | 744 | 756 | 761 | 763 | 793 | 798 | 801 | 840 | 852 |
| Reference | G | T | T | G | A | A | C | C | C | T |
| HPV58/UFPE-58S | . | G | . | . | . | G | T | A | T | C |
| HPV58/UFPE-30S | A | G | . | A | . | . | . | . | . | . |
| HPV58/UFPE-31S | A | G | . | A | . | . | . | . | . | . |
| HPV58/UFPE-54S | A | G | C | A | . | . | . | . | . | . |
| HPV58/UFPE-88S | A | G | . | A | . | . | . | . | . | . |
| HPV58/UFPE-60M | . | G | . | A | G | G | T | A | T | . |
| HPV58/UFPE-59M | A | G | . | A | . | . | . | . | . | . |
| HPV58/UFPE-1M | A | G | . | A | . | . | . | . | . | . |
| HPV58/UFPE-29M | A | G | . | A | . | . | . | . | . | . |
| HPV58/UFPE-35P | A | G | . | A | . | . | . | . | . | . |
| HPV58/UFPE-45M | A | G | . | A | . | . | . | . | . | . |
| HPV58/UFPE-79M | A | G | . | A | . | . | . | . | . | . |
| HPV58/UFPE-92M | A | G | . | A | . | . | . | . | . | . |
| HPV58/UFPE-28M | A | G | . | A | . | . | . | . | . | . |
| HPV58/UFPE-28S | . | G | . | A | G | G | T | A | T | . |
| HPV58/UFPE-48S | A | G | . | A | . | . | . | . | . | . |
| Reference amino acid | G | G | T | T | D | |||||
| Amino acid position | 41 | 63 | 64 | 74 | 76 | |||||
| Amino changed | R | D | A | A | E | |||||
Nt: Nucleotide. T: threonine, D: Aspartic acid, E: glutamate, R: arginine, G: glycine, A: alanine.
Table 3.
LRT test results for the detection of sites under positive (diversifying) selection. Values in between parenthesis indicate the posterior probability of the site to be positively selected. * indicates P > 95%, and ** indicates P > 99%.
Table 3.
LRT test results for the detection of sites under positive (diversifying) selection. Values in between parenthesis indicate the posterior probability of the site to be positively selected. * indicates P > 95%, and ** indicates P > 99%.
| Model 1 | Model 2 | LRT | Positively selected sites |
|---|---|---|---|
| M0 | M3 | 48.63069 | - |
| M1 |
M2 |
31.954069 |
41G (0.979*) 63G (1.000**) 64T (0.826) 74T (0.976*) 76D (0.997**) |
| M1 |
M3 |
31.954662 |
41G (0.997**) 63G (1.000**) 64T (0.893) 74T (0.997**) 76D (1.000**) |
| M7 |
M8 |
32.308663 |
41G (0.987*) 63G (1.000**) 64T (0.857) 74T (0.985*) 76D (0.999**) |
Table 4.
Predicted impact of mutations on the structural stability of HPV58 E7 protein, as determined by Site Directed Mutator. Positive values of ΔΔG are associated with increased protein stability. Negative values of ΔΔG are associated with decreased protein stability.
Table 4.
Predicted impact of mutations on the structural stability of HPV58 E7 protein, as determined by Site Directed Mutator. Positive values of ΔΔG are associated with increased protein stability. Negative values of ΔΔG are associated with decreased protein stability.
| Protein | Mutation | Predicted ΔΔG | Outcome |
|---|---|---|---|
| E7 | G41R | -0.14 | Reduced stability |
| G63D | -2.48 | Reduced stability | |
| T64A | -0.12 | Reduced stability | |
| T74A | 1.26 | Increased stability | |
| D76E | 1.25 | Increased stability |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



