
Submitted:
15 June 2023
Posted:
16 June 2023
You are already at the latest version
Abstract
Lung cancer is the leading cause of global cancer-related mortality resulting in ~1.8 million deaths annually. Systemic, molecular targeted, and immune therapies have provided significant improvements of survival outcomes for patients. However, drug resistance usually arises and there is an urgent need for novel therapy screening and personalized medicine. 3D patient-derived organoid (PDO) models have emerged as a more effective and efficient alternative for ex vivo drug screening than 2D cell culture and patient-derived xenograft (PDX) models. In this review, we performed an extensive search of lung cancer PDO-based ex vivo drug screening studies. Lung cancer PDOs were successfully established from fresh or biobanked sections/biopsy of lung tumors, PDXs, and pleural affusion. PDOs were subject to ex vivo drug screening with chemotherapy, targeted therapy and immune therapy agents. PDOs mainly recapitulated the genomic alterations, transcriptomic landscape and drug sensitivity of primary tumors. Although sample sizes of the previous studies were limited and some technical challenges remained, PDOs showed promise to screen novel therapy drugs. With the technical advance of high throughput, tumor-on-chip, combined microenvironment, and air-liquid interface (ALI), the drug screening using PDOs would serve better for precision care of lung cancer patients.
Keywords:
lung cancer
; organoid
; drug screening
; high throughput
; translational
; pre-clinical
; clinical
; tumor microenvironment
; personalized medicine
1. Introduction
Lung cancer remains the leading cause of cancer-related mortality worldwide accounting for approximately 1.8 million deaths in 2020[1]. Non-small cell lung cancer (NSCLC) is the predominant histologic subtype, accounting for 85% of all lung cancer cases in the United States[2]. In comparison, small cell lung cancer (SCLC) represents 15% of lung cancer cases, occurs almost exclusively in smokers and has the most aggressive clinical course with survival outcomes of 2 to 4 months in untreated patients[3,4]. Systemic therapy is generally indicated for patients who present with advanced disease, including those who present with metastases. With the advent of molecular targeted therapies and immunotherapies, there have been significant improvements in survival outcomes for patients[5,6]. Although these newer therapies have durable responses, drug resistance often occurs with long-term use of these therapies[7]. For these reasons, there is an urgent need to identify novel therapies in patients with treatment-refractory lung cancer of different molecular subtypes who fail standard systemic treatments.
An advanced understanding of lung cancer biology has led to novel therapeutic strategies that target other signal transduction or angiogenesis pathways, as well as leverage the immune system in favor of an anti-tumor response[8,9,10,11,12]. Historically, cancer cell lines have been used to test novel anti-cancer therapies, but they do not accurately predict treatment responses for individual cancer patients due, in part, to interpatient variability and molecular tumor heterogeneity[13,14,15,16]. As a result, ~85% of therapeutic agents that show promising effects in preclinical models fail to demonstrate efficacy in patients[16,17,18]. Though patient-derived xenograft models can mitigate some limitations, poor success rate, intensive resource requirement, high experimental cost, long-term duration of study, limitations owing to statistical power and difficulties in high-throughput expansion have confined their application to preclinical drug screening and demanded the development of alternative models[15,19]. Patient-derived organoids (PDOs), also described as “human tissue in a dish”, can overcome some of these challenges[20,21,22]. Exponentially growing PDO models provide a platform closest to native organs for mapping and exploiting molecular mechanisms of cellular lineage differentiation, organ development, regeneration and diseases[23,24,25,26,27,28]. For malignant progression study, PDOs are 3-D cultures of cancer cells which can be established from a variety of tumor specimens including tissue resections, biopsies, body fluids, circulating tumor cells and pleural effusions from cancers [29,30,31,32,33,34,35,36]. Along with the revolutionary advance, lung organoids were capable of reconstruction of dynamic spatial architecture, molecular identity of cellular components in proximal/distal airways and pathological driver discovery in lung tissue and tumors [37,38,39,40,41,42].
From single stem cells in appropriate Matrigels and culture medium, organoids grow exponentially following unique morphological patterns[24,37,43]. In the presence of small molecule inhibitors or drugs, the growth perturbation efficacy of inhibitors could be assessed based on morphological changes, volume reduction or cell viability of organoids at the endpoints [23]. Thus, PDO models serve as a surrogate of tumors for genomic and transcriptomic profiling, biomarker identification, genetic manipulation and high-throughput drug sensitivity screening. Moreover, they are suitable for long-term expansion or cryopreservation as living organoid biobanks [44,45,46]. An important clinical aspect of PDOs is the potential for genomic and transcriptomic profiling and therapeutic screening as personalized assessment and precision medicine[47,48,49,50,51] (Figure 1). Therefore, PDOs of lung cancer can further our understanding of lung cancer pathophysiology and assess/predict drug efficacy for individual patients in the clinic. Here, we conduct a comprehensive literature review to explore the research to date surrounding ex vivo drug testing using organoids derived from lung cancer patients. We discuss the translational work, challenges and future applications of this promising technology.
2. Methods
This is a comprehensive review of the published literature focusing on research of ex vivo drug testing using organoids from lung cancer patients. The following databases were searched: PubMed, Cochrane Library and Scopus. All full-text, peer-reviewed publications published through March 2023, were included for consideration. The search was restricted to full-text, peer-reviewed articles and reports published in English. Abstract-only publications were excluded, as were any papers that failed to fully elucidate outcomes (editorials, etc.). The search strategy for this review was the result of prior research in the fields. In consultation with an experienced medical librarian, a search strategy was developed. The search terms included (but were not limited to): “patient derived organoids”, “human derived organoids”, and “drug screening”. Further, appropriate MeSH terms and subject headings were also employed. A full search strategy was developed for Ovid Medline and is shown in the Table 1. Lastly, the reference list of identified articles (of the sources that were selected from the full-text review stage and included in the review) were searched for additional sources. The final search results were exported into EndNote and duplicates were removed by the librarian.
Identified titles and abstracts published through March 23, 2022, were screened independently by three reviewers Dr. Josephine Taverna and Dr. Maddison Williams. Disagreements were resolved by discussion until consensus was reached. If consensus could not be reached, a third expert reviewer (Dr. Ryan Williams) was consulted. This process was also used to screen the full text of articles. The reference lists of all included articles were then examined by Dr. Chun-Liang Chen, and additional articles were included in the review until a consensus among the research team was attained. The following information was then abstracted for each included study: primary author, date of publication, country of origin, cancer type, organoid model, methodology, results, conclusions, and limitations.
3. Results
The previous studies of ex vivo drug testing based on patient-derived lung cancer organoids are summarized in Table 2 and organoids culture media with supplements in Table 3. Among the studies, PDOs (n=1-84) were successfully established from resected primary tumors or biopsy of NSCLC (adenocarcinoma, squamous, adenosquamous, and large cell carcinoma) and SCLC, patient-derived xenografts or pleural effusion. Genomic alternations and EGFR/KRAS/BRAF status of PDOs were revealed to be consistent with primary tumors by exome, RNA, targeted sequencing or whole genomic sequencing. The PDOs were subject to ex vivo screening with single or combined antineoplastic drugs (n=1-86), mainly EGFR inhibitors (e.g. Gefitinib, Erlotinib and Afatinib), chemotherapeutic agents (e.g. Cisplatin, Docetaxel and Gemcitabine), immune checkpoint inhibitors and other receptor kinase inhibitors. The contributions and significant discoveries for each study are elaborated separately further in four categories (high throughput, pre-clinical, clinical and mechanistic) as below.
3.1. Organoid technology for high-throughput drug screening
Historically, cancer cell lines have been widely used as preclinical models to evaluate antineoplastic agents. Despite its wide application, the sensitivity and specificity of drug candidates in 2D-cell culture have been widely variable. In order to better predict clinical responses, lung cancer organoid technology was developed for high-throughput drug screening. Li et al. created a living biobank of frozen NSCLC PDOs (n=10) and used high-throughput drug screening assay to demonstrate the anti-tumor effects of three natural compounds -- chelerythrine chloride, cantharidin and harmine[52]. Chen et al. evaluated drug sensitivity of 26 anti-cancer therapeutic agents in organoids derived from 12 patients with locally advanced NSCLC (Stage I-III) which included tumors that harbored EGFR and KRAS G12C mutations[50]. They found that PDOs retained the histological and genetic characteristics of the primary tumors with more than 80% concordance between tumors[50]. Furthermore, they demonstrated PDOs correlated with molecular profiles of primary lung tumors with mutations involving the top 20 NSCLC-related genes[50]. In a separate study, Li et al. tested 24 antineoplastic drugs in PDOs derived from 12 lung adenocarcinoma patients using a high throughput system and found that drug sensitivity also correlated with mutational profiles[53]. Tamura et al. demonstrated the feasibility of using a high throughput system for PDO drug screening from a large compound library comprised of 61 anti-cancer agents (targeted therapies and chemotherapies) [54]. In a similar fashion, Takahashi et al. used their high-throughput assay to evaluate 86 anti-cancer agents (molecular targeted drugs, immune checkpoint inhibitors and cytotoxic chemotherapies) in PDOs developed from lung tumors with squamous and adenosquamous histology[17]. In all these studies, the pharmacogenomic profiles of PDOs were highly comparable to that of tumor tissue indicating that organoid models can better predict individual therapeutic responses on a larger scale to inform therapeutic strategies over a reasonable time frame[55,56,57].
3.2. PDOs in Preclinical Studies
In addition to large-scale screening, PDOs are being employed for personalized drug screening for patients with treatment resistant lung cancer. Jung et al. observed that PDOs derived from SCLC display an inherent drug resistant phenotype to conventional chemotherapy with cisplatin and etoposide[58]. Gmeiner et al. demonstrated that dysregulated pyrimidine biosynthesis contributes to drug resistance in SCLC and they found that the novel fluoropyrimidine polymer (CF10) could overcome drug resistance to conventional chemotherapy using PDOs[59]. In another study, patient derived organoids from NSCLC patient demonstrated the efficacy of an oral potent tubulin destabilizing agent, S-40, in overcoming paclitaxel resistance[60]. These study models highlight the utilization of organoids for assessing drug resistant phenotypes which can be directly translated to the clinic for individual lung cancer patients who develop tumor progression on standard chemotherapy.
The identification of oncogenic activation of tyrosine kinases in NSCLC tumors, most notably mutations in the epidermal growth factor receptor (EGFR) or rearrangements of the anaplastic lymphoma kinase (ALK) gene or c-ROS oncogene 1 (ROS1) gene, has led to the development of targeted agents. Matching a specific targeted drug to the identified driver mutation for an individual patient has resulted in significantly improved survival outcomes, compared with chemotherapy and/or immunotherapy [61,62]. In spite of dramatic tumor shrinkage initially seen in NSCLC patients with driver mutations, drug resistance to targeted therapy often develops. PDOs are currently being used to test small molecular inhibitors with the potential to overcome this secondary resistance. One study conducted by Saraon et al. evaluated the efficacy of EMI66 and other small molecules that inhibit mutant EGFR signaling and alters ER stress response pathway[63]. They found that EMI66 and other derivatives reduced viability of organoids derived from three PDOs with EGFR mutations, suggesting another bypass mechanism to target tumor growth[63]. Hu et al. found that PDOs with the EML4-ALK rearrangement mutation and EGFR activation mutations showed reduced viability and sensitivity to TKI inhibition, compared to wild-type[64]. They demonstrated that lung organoids with TKI sensitive L858R mutation and absent T790 mutation could develop drug resistance early, highlighting the potential role of PDOs to predict drug resistance better than the T790M molecular marker[64]. These collective studies suggests that lung organoids can be used as a treatment prediction model to allow clinicians to prioritize targeted and chemotherapies prospectively by comparing the responses of the individual PDOs to different drug combinations in real-time.
3.3. PDOs in clinical studies - an enhanced model for personalized drug screening
In a large clinical study, 84 organoids were established from patients with oncogene-driven lung adenocarcinoma and found that PDOs recapitulated PFS and ORR of patients receiving clinically approved targeted agents[61]. They found that combination treatment with dabrafenib and trametinib elicited both ex vivo and clinical responses in a NSCLC patient harboring an EGFR exon 19 deletion and a BRAF G464A mutation[61]. They also demonstrated pre-clinical and clinical efficacy of afatinib against PDOs that harbored the rare EGFR L747P mutation. Among ERBB2 targeted therapies (erlotinib, lapatinib, neratinib, and afatinib), poziotinib demonstrated the greatest potency in lung organoids with ERBB2 exon 20 insertions[61]. The preclinical studies led to a phase 2 study (ZENITH20-2 Trial) of poziotinib which yielded a response rate of 28% in 25 of 90 patients with treatment refractory HER2 exon 20 insertion NSCLC. Both the preclinical and clinical studies indicated robust and durable anti-tumor activity of poziotinib in NSCLC patients with ERBB2 exon 20 insertions[61]. Among the multi-kinase inhibitors (vandetanib, cabozantinib, and lenvatinib), pralsetinib demonstrated clinical efficacy in RET-rearranged lung tumors[61]. In a recent phase I/II clinical trial (ARROW trial), robust treatment responses were observed in patients with RET fusion-positive NSCLC (n=233)[65]. In 53 of 87 NSCLC patients previously treated with platinum-based chemotherapy, there was a 61% overall response to therapy including five patients with a complete response[65]. In 19 of 27 treatment naïve NSCLC, a 70% overall response was observed with a complete response in three patients[65]. Overall, these studies highlight the potential of organoid-based drug sensitivity testing to predict clinical outcomes better than molecular markers. This accuracy in treatment prediction offers clinicians the ability to prioritize targeted and chemotherapies prospectively by comparing the individual responses of the generated PDOs to different drugs.
About 2%–3% human epidermal growth factor receptor 2 (HER2, ERBB2) mutation have been identified as one of tumorigenic drivers and observed in of NSCLC[66]. Several HER2-targeted agents (afatinib, dacomitinib, neratinib and trastuzumab) have shown limited clinical activity in patients with HER2-mutant NSCLC[67,68,69,70]. Wang et al. recently studied the anti-tumor effects of pyrotinib, a pan-HER inhibitor, in NSCLC with HER2 exon 20 mutations in both preclinical and clinical models [49]. Pyrotinib showed significant growth inhibition of organoids relative to afatinib[49]. In PDX model, pyrotinib showed a superior antitumor effect compared with afatinib and trastuzumab emtansine (a humanized monoclonal antibody trastuzumab covalently linked to the cytotoxic agent DM1) [49]. Specifically, mice treated with pyrotinib had >50% reduction in their tumor volumes[49]. This anti-tumor drug effect was attributed to the inhibition of HER2 and its downstream signaling pathways, including ERK and Akt. They validated their preclinical findings in a phase 2 study of pyrotinib (400 mg oral daily dose) in patients with advanced stage III-IV NSCLC patients harboring HER2 mutations[71]. In this single arm study, 15 patients with HER2 mutations involving exon 20 (n=11), exon 19 (n=3), and exon 17 (n=1) had reduction in tumor burden with an objective response rate of 19.2%, median duration of 9.9 months and disease control rate of 74.4% suggesting[71]. Collectively study also demonstrated the applicability of lung organoid models to study patient populations with rare mutations.
The fibroblast growth factor/fibroblast growth factor receptor (FGF/FGFR) is a tyrosine kinase signaling pathway plays a critical role in oncogenesis (tumor proliferation, angiogenesis, migration, and survival) via gene amplification, activating mutations, or translocation in lung tumors[72,73]. In this way, the FGFR signaling pathway represents an important target for lung cancer. In vitro studies suggest that combination of MEK and PI3K inhibitors with FGFR inhibitors may be effective in FGFR-aberrant cancers [74]. To test this hypothesis in preclinical models, Shi et al. developed organoids from surgically resected lung tumors derived from 19 patients with lung adenocarcinoma and 15 patients with squamous cell lung carcinoma[74]. Additionally, they processed 16 lung adenocarcinoma PDX and 26 squamous cell lung cancer PDX tumors for organoid establishment[74]. They tested trametinib (MEK inhibitor) and the PI3K inhibitor BKM120 with BGJ398 (inhibits pFGFR and pAkt.) in their FGFR1-amplified organoid models[74]. Strong synergy was observed in the BGJ398 + trametinib combination (combination index < 0.5), whereas weaker synergy (combination index >0.5) was observed in the BGJ398 + BKM120 (PI3K inhibitor) combination[74]. Furthermore, PDOs retained the histologic and molecular features of their parental tumors[74]. The spectrum of mutations was highly concordant between the organoid and matched patient tumor and PDX tissue[74]. Mutational burden in the five long-term established PDOs was similar to parental patient/PDX tumors[74]. This indicates that their organoid culture conditions did not destabilize the cancer genome. Copy number variation (CNV) profiles of the parental tumors were largely preserved during PDO culture[74]. There have been early phase clinical studies in patients with lung cancer (and other solid tumors) evaluating FGFR inhibitors as monotherapy or in combination with existing therapies. As more clinical trials of both selective and non-selective FGFR inhibitors emerge[75], concentration on patient selection as it pertains to predicting response to therapy should be undertaken with patient derived organoids given the accuracy of this model system.
Insertion mutations in exon 20 of the epidermal growth factor receptor (EGFR) gene are the largest class of EGFR mutations in non-small cell lung cancer (NSCLC) for which there are no FDA approved targeted therapies[76]. Yun et al. tested amivantamab (an EGFR-MET bispecific antibody) in tumors derived from organoids and xenograft models harboring diverse Exon 20 insertion mutations and found that the drug showed anti-tumor activity and suppressed EGFR and MET signaling pathways[77]. Furthermore, they established their PDOs from malignant pleural effusions collected from two patients with NSCLC with Exon 20 insertion mutations[77]. In a first-in-human study, amivantamab produced robust tumor responses in two patients with EGFR Exon 20 insertion NSCLC, highlighting the importance of ex vivo drug testing to inform clinical trial design[77].
3.4. Mechanistic studies involving PDOs
PDOs also serve as a valuable model to study tumor evolution and the acquisition of secondary mutations associated with treatment resistance. Banda et al., treated lung tumor organoids with erlotinib, a first generation EGFR-tyrosine kinase inhibitor[78]. Upon subsequent passaging, the organoid cultures developed additional mutations in BRAF, EGFR, KRAS and PIK3CA genes which have been commonly reported in patients who develop resistance against EGFR inhibitors[78]. These observations were supported by a comprehensive mutation profiling analysis (multiplex testing) of tumor specimens collected from 153 NSCLC patients identified the co-existence of EGFR with KRAS (n=29), or BRAF (n=2) or PIK3CA somatic mutations (n=58)[79].
Proper identification of subclones and targeting individual tumor subpopulations with effective targeted therapies can lead to effective elimination of tumor cells and durable treatment responses in lung cancer patients. Taverna et al. used single-cell proteomic profiling to first identify continuous AXL and JAK1-STAT3 (bypass loop) signal activation in lung tumors derived from 11 treatment naïve lung adenocarcinoma patients who underwent tumor resection of their primary tumor[80]. Using single cell profiling, they were able to stratify tumor subpopulations based on their AXL and STAT3 signaling (bypass mechanism) [80]. They found that tumor subpopulations with high AXL/STAT3 expression concordantly expressed high expression of epithelial/ mesenchymal markers (hybrid epithelial-to-mesenchymal phenotype) and cancer stemness proteins, suggesting high tumorigenic and metastatic potential[80]. They subsequently stratified tumor based on high AXL/STAT3 and low AXL/STAT3 expression and found that patient derived organoids (PDO=3) with high AXL/STAT3 expression responded robustly to combination treatment with dubermatinib (AXL inhibitor) and ruxolitnib (JAK inhibitor) as compared with single agents[80]. Conversely, PDOs (n=2) that expressed low AXL/STAT3 expression did not respond to combination treatment and/or single agents[80]. In H2009 and A549 mouse models, they found that combination treatment with AXL and JAK inhibitor was synergistic and reduced tumor volumes significantly compared with single agent treatments (unpublished, manuscript under revision). In this study, they found that PDOs successfully captured the cellular heterogeneity of their primary lung tumor cells and could provide insights into potential combination therapy to overcome drug resistance.
Another major strength for PDOs is that they can be subjected to genetic manipulation, which can be very a very powerful tool understand oncogenic signaling in lung cancer. For example, Dost et al. used organoids model to study transcriptional hallmarks of oncogenic KRAS activation in lung epithelial progenitor cells[81]. They developed organoid systems from primary mouse and human induced pluripotent stem cell-derived lung epithelial cells to model early-stage lung adenocarcinoma, showing organoid approaches can be utilized for uncovering the early consequences of oncogenic KRAS expression.
4. Conclusions and future directions
Lung cancer PDOs have emerged as an effective alternative model for ex vivo drug screening. From the previous studies, lung cancer PDOs showed promise in mechanistic and pre-clinical studies for drugs targeting lung malignancy to achieve potential personalized medicine.
Whether lung organoids can accurately predict treatment responses in clinical trials remains an area of active research. PDOs have become widely used for testing anti-neoplastic agents for therapeutic efficacy in clinical trials. The ability to generate organoid models from individual patients enrolled in clinical trials allows investigation of patient-specific responses to therapeutic drugs in real-time[61]. One research team established a PDO biobank from patients with metastatic gastrointestinal cancer who were actively undergoing treatment in clinical trials and demonstrated that the ex vivo responses in organoid cultures closely mirrored clinical responses with 100% sensitivity, 93% specificity, 100% negative prediction accuracy and 88% positive prediction accuracy[82]. This study provides a strong rationale for designing co-clinical trials with organoid models to better predict drug responses. Some matched studies have also shown that PDOs accurately predicted therapeutic responses of patients and allow comparison of outcomes of different novel monotherapies or combination therapies[77,82] . Organoids have also been adopted to predict treatment response to radiotherapy and immunotherapy[83,84,85]. By simulating cancer behavior ex vivo, organoid technology can integrate molecular biology with the decision-making process of early-phase clinical trials to improve patient selection for particular therapeutic agents.
Based on our review of the literature, organoid models can be successfully established from pleural effusions which is scarce source of lung cancer cells[77,86,87]. Strikingly, organoids derived from pleural fluid aspirate can developed tumor-specific cellular and molecular characteristics and preserved tumor heterogeneity[86,87]. Since pleural effusion aspiration is fairly a non-invasive and routine procedure among lung cancer patients, generation of organoid models from these samples can allow effective disease modeling, monitor disease progression and treatment responses in real-time. Another advantage of using pleural effusion-derived cells is that they constitute both tumor cells and stromal cells such that tumor microenvironment and disease pathophysiology can be preserved and cellular interactions in response to disease progression or anti-cancer agents can be studied simultaneously[87,88]. Co-culturing tumor organoids with immune cells can mimic complex cellular microenvironment and tumor-intrinsic interactions [17]. For example, Takahashi et al. recently demonstrated that lung PDOs exposed to antibody-drug conjugates (trastuzumab, pertuzumab and trastuzumab emtansine) can effectively model complex interactions with immune cells (THP-1 effector cells) and reflect the antibody-dependent cellular cytotoxicity (ADCC) response of the drug[17]. The study also expanded on the use of anti-PD1 monoclonal antibodies (nivolumab and pembrolizumab) to evaluate efficacy of immune checkpoint inhibitors.
Although current methods of generating NSCLC PDOs are robust and straightforward, they are not without limitations. Indeed, some challenges still exist regarding establishing pure NSCLC PDOs. For example, Dijkstra et al. reported that that the majority of organoids derived from intrapulmonary lung cancers are overgrown by normal airway organoids, causing the overall establishment rate of pure lung cancer organoids to be only 17%[89]. Such low establishment rate of pure lung cancer organoids limits their clinical utility. Therefore, innovative methods are still needed to generate more robust and purer NSCLC PDOs.
Although organoid culture systems have several advantages over traditional 2D cell-based models, there are certain drawbacks that need to be overcome. For example, the most common organoid models consist of only the epithelial compartment and lack the stromal signature and further signals from the microenvironment[90]. The contribution of tumor stroma and complex tumor microenvironment in tumor progression has become well recognized and this has led to recent advances in the engineering of co-culture systems. For example, microfluidic organ-on-chips have been developed to model cell-cell and cell-extracellular matrix (ECM) interactions. These novel systems are designed to recapitulate organ-level functionality including physical forces that mimic in vivo cyclic strain and fluid shear stress. Moreover, the microfluidic nature of these systems and microsensors within the microchip sustains longer-term experiments and allows for the collection of real-time data such as barrier function and effluent collection to monitor byproducts as an indirect measure of tissue functionality[91]. As a part of rapidly advancing organ-on-chip field of research, tumor-on-chip technology has emerged as a promising tool to mimic the mechanical and biochemical properties of the tumor microenvironment. This includes factors such as oxygen and nutrient gradients, extracellular matrix stiffness, and cell-cell interactions. In this way, tumor-on-chips create a miniature model of lung tumors on a microfluidic chip allowing researchers to gain insights into how cancer cells directly engage with their environment and allows for the testing of novel therapeutics in a more efficient manner. In lung cancer, there have been several recent advances with tumor-on-chip technology. In a recent study, Liao et al. describe the development of a lung cancer-on-chip model consisting of both tumor and immune cells which recapitulates the complex interactions between these cell types ex vivo[92]. Jiang et al. employed an immunotherapeutic high-throughput observation chamber to test the efficacy of anti-PD-1 antibodies on cancer spheroid (MDA-MB-231) and T cell (Jurkat) cell-cell interactions[93]. The system detected T cell inhibition and activation by measuring IL-2 secretion and measuring tumor infiltration. However, not only cellular make-up of the tumor microenvironment but also mechanical determinants need to be recapitulated on-a-chip to faithfully monitor drug response. For example, Hassell et al. described microfluidic organ chip device complete with endothelium, epithelium and NSCLC cells suitable for studies on cancer growth, invasion, mechanotransduction and drug response. Interestingly, applying breathing-like mechanical motions to such alveolous chip bestowed cancer cells with resistance to a tyrosine kinase inhibitor drug rociletinib[94]. Mechanotransduction is undoubtedly important yet often overlooked aspect of tumorigenesis, tumor spread and drug response. Organ-on-chips studies should facilitate development of transduction-targeting “mechanotherapeutics”[95].
Summarizing, the future development of PDO on chip technology will revolutionize IO cancer treatment by allowing for personalized treatment reflecting individual patients’ drug responses in their unique tumor microenvironment. These model systems will be able to mimic multiple aspects of tumor microenvironments (tumor cells, stromal fibroblasts, endothelial cells, immune cells, mechanical determinants) and their individual responses (cytokine release, mechanotransduction) to immuno- and other therapies[95].
Tumor-on-chip platform also provides a powerful tool to gain insights into the underlying mechanisms of metastasis. Metastasis is a complex process that involves multiple steps, including the invasion of tumor cells into surrounding tissue, colonization at the secondary site, and intravasation into the blood or lymphatic vessels. Recent studies validated that tumor-on-chip models can be used to simplify and study each of these steps. One example is a study by Zhang et al. where they use tumor-on-chip system to study the interaction of tumor cells with endothelial cells that line blood and lymphatic vessels and demonstrated that an anti-angiogenic drug can inhibit the intravasation and extravasation of breast cancer cells[96]. The microenvironment such as oxygen and nutrient availability is different at secondary site of metastasis and tumor cells must adapt to survive and grow in the new environment. In a recent report, researchers developed microfluidic chips to investigate the colonization of tumor cells at the secondary site during metastasis and showed that hypoxic condition at the secondary site promotes the growth and migration of prostate cancer cells.
Overall, tumor-on-chip technology has proven great promise in drug discovery and development and will likely advance to become more widely used in the future. More specifically, the progress with development of multi-organ-on-chips may allow researchers to test combination therapies that target multiple organs simultaneously and test how drugs affect different organs in the body[91,94,95,97,98,99,100]. This development is crucial to better understand the potential side effects of drugs. In addition, combining advancements in microfluidic technology and automation could enable high-throughput screening of large number of drugs using tumor-on-chip models. This could greatly accelerate the drug discovery process by reducing the need for trial-and-error approaches to treatment, and rapid testing of potential drug candidates in a patient specific manner. Authors should discuss the results and how they can be interpreted from the perspective of previous studies and of the working hypotheses. The findings and their implications should be discussed in the broadest context possible. Future research directions may also be highlighted.
There is a potential drawback of 3D organoid model in that these organoids form 3D structures with apical side of cells facing the lumen. This prevents apical surfaces of cells from air interface and direct exposure to inhaled substances as it occurs in their tissue of origin[46]. The technical downside may be alleviated by The Air-liquid interface (ALI) technique where lung organoids are dissociated and grown as a monolayer may be applied as an alternative to address this issue[101]. Contrary to traditional cell culture methods which grow cells in a liquid medium, in ALI technique, media may be removed from the apical side of cells to expose them directly to the air. This approach not only improves the differentiation process where lung epithelium forms a more mature and functional pseudo-stratified epithelium, it also allows recapitulating the complex physiological conditions of the lung tissue. The validity of ALI as a lung model has been shown by different studies comparing the functionality and genomic profile of harvested cells to in vivo condition[102,103,104]. Although ALI condition has been extensively used for culturing airway epithelial cells, it has not been explored enough for pharmacological assays such as permeability and transport studies. However, the ALI technique not only may improve insights on drug effects compared to the use of submerged cultures, but it also gives access to both apical and basolateral sides of the cells for convenient addition of any stimulus, as well as sample collection. With limited number of studies that used ALI to investigate the effects of smoking or environmental pollutants in lung cancer, the authors demonstrated that the exposure to these factors induces significant changes in cell growth, apoptosis, and inflammation[101,105].
Funding
This research (to CLC) was partially funded by NIH/NCI, grant number 1R21CA264353-01 and U54 CA217297-04. This research (to J.A.T) was supported by the Max and Minnie Tomerlin Voelcker Fund Young Investigator Award, NIH/NCATS grants KL2 TR001118 and KL2 TR002646.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Pikor, L.A.; Ramnarine, V.R.; Lam, S.; Lam, W.L. Genetic alterations defining NSCLC subtypes and their therapeutic implications. Lung Cancer 2013, 82, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Rudin, C.M.; Brambilla, E.; Faivre-Finn, C.; Sage, J. Small-cell lung cancer. Nat Rev Dis Primers 2021, 7, 3. [Google Scholar] [CrossRef]
- Byers, L.A.; Rudin, C.M. Small cell lung cancer: where do we go from here? Cancer 2015, 121, 664–672. [Google Scholar] [CrossRef]
- Hirsch, F.R.; Scagliotti, G.V.; Mulshine, J.L.; Kwon, R.; Curran, W.J., Jr.; Wu, Y.L.; Paz-Ares, L. Lung cancer: current therapies and new targeted treatments. Lancet 2017, 389, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Reck, M.; Rodriguez-Abreu, D.; Robinson, A.G.; Hui, R.; Csoszi, T.; Fulop, A.; Gottfried, M.; Peled, N.; Tafreshi, A.; Cuffe, S.; et al. Pembrolizumab versus Chemotherapy for PD-L1-Positive Non-Small-Cell Lung Cancer. N Engl J Med 2016, 375, 1823–1833. [Google Scholar] [CrossRef]
- Lim, Z.F.; Ma, P.C. Emerging insights of tumor heterogeneity and drug resistance mechanisms in lung cancer targeted therapy. J Hematol Oncol 2019, 12, 134. [Google Scholar] [CrossRef]
- Yuan, M.; Zhao, Y.; Arkenau, H.T.; Lao, T.; Chu, L.; Xu, Q. Signal pathways and precision therapy of small-cell lung cancer. Signal Transduct Target Ther 2022, 7, 187. [Google Scholar] [CrossRef]
- Niu, Z.; Jin, R.; Zhang, Y.; Li, H. Signaling pathways and targeted therapies in lung squamous cell carcinoma: mechanisms and clinical trials. Signal Transduct Target Ther 2022, 7, 353. [Google Scholar] [CrossRef]
- Rotow, J.; Bivona, T.G. Understanding and targeting resistance mechanisms in NSCLC. Nat Rev Cancer 2017, 17, 637–658. [Google Scholar] [CrossRef]
- Altorki, N.K.; Markowitz, G.J.; Gao, D.; Port, J.L.; Saxena, A.; Stiles, B.; McGraw, T.; Mittal, V. The lung microenvironment: an important regulator of tumour growth and metastasis. Nat Rev Cancer 2019, 19, 9–31. [Google Scholar] [CrossRef] [PubMed]
- Hiam-Galvez, K.J.; Allen, B.M.; Spitzer, M.H. Systemic immunity in cancer. Nat Rev Cancer 2021, 21, 345–359. [Google Scholar] [CrossRef]
- Douglas, W.H.; McAteer, J.A.; Smith, J.R.; Braunschweiger, W.R. Type II alveolar pneumonocytes in vitro. Int Rev Cytol Suppl 1979, 45–65. [Google Scholar] [CrossRef]
- Choi, J.; Iich, E.; Lee, J.H. Organogenesis of adult lung in a dish: Differentiation, disease and therapy. Dev Biol 2016, 420, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Bleijs, M.; van de Wetering, M.; Clevers, H.; Drost, J. Xenograft and organoid model systems in cancer research. Embo J 2019, 38, e101654. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Mun, H.; Sung, C.O.; Cho, E.J.; Jeon, H.J.; Chun, S.M.; Jung, D.J.; Shin, T.H.; Jeong, G.S.; Kim, D.K.; et al. Patient-derived lung cancer organoids as in vitro cancer models for therapeutic screening. Nat Commun 2019, 10, 3991. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Hoshi, H.; Higa, A.; Hiyama, G.; Tamura, H.; Ogawa, M.; Takagi, K.; Goda, K.; Okabe, N.; Muto, S.; et al. An In Vitro System for Evaluating Molecular Targeted Drugs Using Lung Patient-Derived Tumor Organoids. Cells 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- DiMasi, J.A.; Reichert, J.M.; Feldman, L.; Malins, A. Clinical approval success rates for investigational cancer drugs. Clin Pharmacol Ther 2013, 94, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Morgan, K.M.; Riedlinger, G.M.; Rosenfeld, J.; Ganesan, S.; Pine, S.R. Patient-Derived Xenograft Models of Non-Small Cell Lung Cancer and Their Potential Utility in Personalized Medicine. Front Oncol 2017, 7, 2. [Google Scholar] [CrossRef] [PubMed]
- Bredenoord, A.L.; Clevers, H.; Knoblich, J.A. Human tissues in a dish: The research and ethical implications of organoid technology. Science 2017, 355, 20. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H.C. Organoids: Avatars for Personalized Medicine. Keio J Med 2019, 68, 95. [Google Scholar] [CrossRef]
- Iakobachvili, N.; Peters, P.J. Humans in a Dish: The Potential of Organoids in Modeling Immunity and Infectious Diseases. Front Microbiol 2017, 8, 2402. [Google Scholar] [CrossRef]
- Chen, C.L.; Yu, X.; James, I.O.; Zhang, H.Y.; Yang, J.; Radulescu, A.; Zhou, Y.; Besner, G.E. Heparin-binding EGF-like growth factor protects intestinal stem cells from injury in a rat model of necrotizing enterocolitis. Lab Invest 2012, 92, 331–344. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Vries, R.G.; Snippert, H.J.; van de Wetering, M.; Barker, N.; Stange, D.E.; van Es, J.H.; Abo, A.; Kujala, P.; Peters, P.J.; et al. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature 2009, 459, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, M.A.; Knoblich, J.A. Generation of cerebral organoids from human pluripotent stem cells. Nat Protoc 2014, 9, 2329–2340. [Google Scholar] [CrossRef]
- Rossi, G.; Manfrin, A.; Lutolf, M.P. Progress and potential in organoid research. Nat Rev Genet 2018, 19, 671–687. [Google Scholar] [CrossRef] [PubMed]
- Velasco, S.; Kedaigle, A.J.; Simmons, S.K.; Nash, A.; Rocha, M.; Quadrato, G.; Paulsen, B.; Nguyen, L.; Adiconis, X.; Regev, A.; et al. Individual brain organoids reproducibly form cell diversity of the human cerebral cortex. Nature 2019, 570, 523–527. [Google Scholar] [CrossRef]
- Lee, S.-E.; Shin, N.; Kook, M.G.; Kong, D.; Kim, N.G.; Choi, S.W.; Kang, K.-S. Human iNSC-derived brain organoid model of lysosomal storage disorder in Niemann-Pick disease type C. Cell death & disease 2020, 11, 1059. [Google Scholar] [CrossRef]
- Gao, D.; Vela, I.; Sboner, A.; Iaquinta, P.J.; Karthaus, W.R.; Gopalan, A.; Dowling, C.; Wanjala, J.N.; Undvall, E.A.; Arora, V.K.; et al. Organoid cultures derived from patients with advanced prostate cancer. Cell 2014, 159, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. Modeling Development and Disease with Organoids. Cell 2016, 165, 1586–1597. [Google Scholar] [CrossRef] [PubMed]
- Drost, J.; Clevers, H. Organoids in cancer research. Nat Rev Cancer 2018, 18, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Sachs, N.; de Ligt, J.; Kopper, O.; Gogola, E.; Bounova, G.; Weeber, F.; Balgobind, A.V.; Wind, K.; Gracanin, A.; Begthel, H.; et al. A Living Biobank of Breast Cancer Organoids Captures Disease Heterogeneity. Cell 2018, 172, 373–386. [Google Scholar] [CrossRef]
- Driehuis, E.; Kretzschmar, K.; Clevers, H. Establishment of patient-derived cancer organoids for drug-screening applications. Nature protocols 2020, 15, 3380–3409. [Google Scholar] [CrossRef]
- Yu, J.H.; Ma, S. Organoids as research models for hepatocellular carcinoma. Experimental cell research 2022, 411, 112987. [Google Scholar] [CrossRef]
- Baiao, A.; Dias, S.; Soares, A.F.; Pereira, C.L.; Oliveira, C.; Sarmento, B. Advances in the use of 3D colorectal cancer models for novel drug discovery. Expert opinion on drug discovery 2022, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Doffo, J.; Bamopoulos, S.A.; Kose, H.; Orben, F.; Zang, C.; Pons, M.; den Dekker, A.T.; Brouwer, R.W.W.; Baluapuri, A.; Habringer, S.; et al. NOXA expression drives synthetic lethality to RUNX1 inhibition in pancreatic cancer. Proceedings of the National Academy of Sciences of the United States of America 2022, 119. [Google Scholar] [CrossRef]
- Rock, J.R.; Onaitis, M.W.; Rawlins, E.L.; Lu, Y.; Clark, C.P.; Xue, Y.; Randell, S.H.; Hogan, B.L. Basal cells as stem cells of the mouse trachea and human airway epithelium. Proc Natl Acad Sci U S A 2009, 106, 12771–12775. [Google Scholar] [CrossRef]
- Hild, M.; Jaffe, A.B. Production of 3-D Airway Organoids From Primary Human Airway Basal Cells and Their Use in High-Throughput Screening. Curr Protoc Stem Cell Biol 2016, 37, IE 9 1–IE 9 15. [Google Scholar] [CrossRef]
- Nadkarni, R.R.; Abed, S.; Draper, J.S. Organoids as a model system for studying human lung development and disease. Biochem Biophys Res Commun 2016, 473, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.W.; Huang, S.X.; de Carvalho, A.; Ho, S.H.; Islam, M.N.; Volpi, S.; Notarangelo, L.D.; Ciancanelli, M.; Casanova, J.L.; Bhattacharya, J.; et al. A three-dimensional model of human lung development and disease from pluripotent stem cells. Nat Cell Biol 2017, 19, 542–549. [Google Scholar] [CrossRef]
- Han, Y.; Duan, X.; Yang, L.; Nilsson-Payant, B.E.; Wang, P.; Duan, F.; Tang, X.; Yaron, T.M.; Zhang, T.; Uhl, S.; et al. Identification of SARS-CoV-2 inhibitors using lung and colonic organoids. Nature 2021, 589, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Kadur Lakshminarasimha Murthy, P.; Sontake, V.; Tata, A.; Kobayashi, Y.; Macadlo, L.; Okuda, K.; Conchola, A.S.; Nakano, S.; Gregory, S.; Miller, L.A.; et al. Human distal lung maps and lineage hierarchies reveal a bipotent progenitor. Nature 2022, 604, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, J. Patient-Derived Organoids Predict Cancer Treatment Response. Jama 2018, 319, 1427. [Google Scholar] [CrossRef]
- Clevers, H.; Bender, E. Q&A: Hans Clevers. Banking on organoids. Nature 2015, 521, S15. [Google Scholar] [CrossRef]
- Weeber, F.; van de Wetering, M.; Hoogstraat, M.; Dijkstra, K.K.; Krijgsman, O.; Kuilman, T.; Gadellaa-van Hooijdonk, C.G.; van der Velden, D.L.; Peeper, D.S.; Cuppen, E.P.; et al. Preserved genetic diversity in organoids cultured from biopsies of human colorectal cancer metastases. Proc Natl Acad Sci U S A 2015, 112, 13308–13311. [Google Scholar] [CrossRef] [PubMed]
- Sachs, N.; Papaspyropoulos, A.; Zomer-van Ommen, D.D.; Heo, I.; Bottinger, L.; Klay, D.; Weeber, F.; Huelsz-Prince, G.; Iakobachvili, N.; Amatngalim, G.D.; et al. Long-term expanding human airway organoids for disease modeling. Embo J 2019, 38, 15. [Google Scholar] [CrossRef]
- Verissimo, C.S.; Overmeer, R.M.; Ponsioen, B.; Drost, J.; Mertens, S.; Verlaan-Klink, I.; Gerwen, B.V.; van der Ven, M.; Wetering, M.V.; Egan, D.A.; et al. Targeting mutant RAS in patient-derived colorectal cancer organoids by combinatorial drug screening. Elife 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Pauli, C.; Hopkins, B.D.; Prandi, D.; Shaw, R.; Fedrizzi, T.; Sboner, A.; Sailer, V.; Augello, M.; Puca, L.; Rosati, R.; et al. Personalized In Vitro and In Vivo Cancer Models to Guide Precision Medicine. Cancer Discov 2017, 7, 462–477. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, T.; Qin, Z.; Jiang, J.; Wang, Q.; Yang, S.; Rivard, C.; Gao, G.; Ng, T.L.; Tu, M.M.; et al. HER2 exon 20 insertions in non-small-cell lung cancer are sensitive to the irreversible pan-HER receptor tyrosine kinase inhibitor pyrotinib. Ann Oncol 2019, 30, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.H.; Chu, X.P.; Zhang, J.T.; Nie, Q.; Tang, W.F.; Su, J.; Yan, H.H.; Zheng, H.P.; Chen, Z.X.; Chen, X.; et al. Genomic characteristics and drug screening among organoids derived from non-small cell lung cancer patients. Thorac Cancer 2020, 11, 2279–2290. [Google Scholar] [CrossRef]
- Zhang, Z.; Karthaus, W.R.; Lee, Y.S.; Gao, V.R.; Wu, C.; Russo, J.W.; Liu, M.; Mota, J.M.; Abida, W.; Linton, E.; et al. Tumor Microenvironment-Derived NRG1 Promotes Antiandrogen Resistance in Prostate Cancer. Cancer Cell 2020, 38, 279–296 e279. [Google Scholar] [CrossRef]
- Li, Y.F.; Gao, Y.; Liang, B.W.; Cao, X.Q.; Sun, Z.J.; Yu, J.H.; Liu, Z.D.; Han, Y. Patient-derived organoids of non-small cells lung cancer and their application for drug screening. Neoplasma 2020, 67, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Qian, Y.; Li, W.; Liu, L.; Yu, L.; Liu, X.; Wu, G.; Wang, Y.; Luo, W.; Fang, F.; et al. Human Lung Adenocarcinoma-Derived Organoid Models for Drug Screening. iScience 2020, 23, 101411. [Google Scholar] [CrossRef]
- Tamura, H.; Higa, A.; Hoshi, H.; Hiyama, G.; Takahashi, N.; Ryufuku, M.; Morisawa, G.; Yanagisawa, Y.; Ito, E.; Imai, J.I.; et al. Evaluation of anticancer agents using patient-derived tumor organoids characteristically similar to source tissues. Oncol Rep 2018, 40, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Mun, H.; Sung, C.O.; Cho, E.J.; Jeon, H.J.; Chun, S.M.; Jung, D.J.; Shin, T.H.; Jeong, G.S.; Kim, D.K.; et al. Patient-derived lung cancer organoids as in vitro cancer models for therapeutic screening. Nat Commun 2019, 10, 3991. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-Y.; Kim, S.-M.; Lim, S.; Lee, J.Y.; Choi, S.-J.; Yang, S.-D.; Yun, M.R.; Kim, C.G.; Gu, S.R.; Park, C.; et al. Modeling Clinical Responses to Targeted Therapies by Patient-Derived Organoids of Advanced Lung Adenocarcinoma. Clinical cancer research : an official journal of the American Association for Cancer Research 2021, 27, 4397–4409. [Google Scholar] [CrossRef] [PubMed]
- Pomerenke, A. Organotypic Models of Lung Cancer. Curr Top Microbiol Immunol 2018, 09, 09. [Google Scholar] [CrossRef]
- Jung, D.J.; Shin, T.H.; Kim, M.; Sung, C.O.; Jang, S.J.; Jeong, G.S. A one-stop microfluidic-based lung cancer organoid culture platform for testing drug sensitivity. Lab chip 2019, 19, 2854–2865. [Google Scholar] [CrossRef] [PubMed]
- Gmeiner, W.H.; Miller, L.D.; Chou, J.W.; Dominijanni, A.; Mutkus, L.; Marini, F.; Ruiz, J.; Dotson, T.; Thomas, K.W.; Parks, G.; et al. Dysregulated Pyrimidine Biosynthesis Contributes to 5-FU Resistance in SCLC Patient-Derived Organoids but Response to a Novel Polymeric Fluoropyrimidine, CF10. Cancers (Basel) 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Du, T.; Lin, S.; Ji, M.; Xue, N.; Liu, Y.; Zhang, Z.; Zhang, K.; Zhang, J.; Zhang, Y.; Wang, Q.; et al. A novel orally active microtubule destabilizing agent S-40 targets the colchicine-binding site and shows potent antitumor activity. Cancer Letters 2020, 495, 22–32. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kim, S.M.; Lim, S.; Lee, J.Y.; Choi, S.J.; Yang, S.D.; Yun, M.R.; Kim, C.G.; Gu, S.R.; Park, C.; et al. Modeling Clinical Responses to Targeted Therapies by Patient-Derived Organoids of Advanced Lung Adenocarcinoma. Clin Cancer Res 2021, 27, 4397–4409. [Google Scholar] [CrossRef]
- Verduin, M.; Hoeben, A.; De Ruysscher, D.; Vooijs, M. Patient-Derived Cancer Organoids as Predictors of Treatment Response. Frontiers in oncology 2021, 11, 641980. [Google Scholar] [CrossRef]
- Saraon, P.; Snider, J.; Schormann, W.; Rai, A.; Radulovich, N.; Sánchez-Osuna, M.; Coulombe-Huntington, J.; Huard, C.; Mohammed, M.; Lima-Fernandes, E.; et al. Chemical Genetics Screen Identifies COPB2 Tool Compounds That Alters ER Stress Response and Induces RTK Dysregulation in Lung Cancer Cells. J Mol Biol 2021, 433, 167294. [Google Scholar] [CrossRef]
- Hu, Y.; Sui, X.; Song, F.; Li, Y.; Li, K.; Chen, Z.; Yang, F.; Chen, X.; Zhang, Y.; Wang, X.; et al. Lung cancer organoids analyzed on microwell arrays predict drug responses of patients within a week. Nat Commun 2021, 12, 2581. [Google Scholar] [CrossRef] [PubMed]
- Gainor, J.F.; Curigliano, G.; Kim, D.W.; Lee, D.H.; Besse, B.; Baik, C.S.; Doebele, R.C.; Cassier, P.A.; Lopes, G.; Tan, D.S.W.; et al. Pralsetinib for RET fusion-positive non-small-cell lung cancer (ARROW): a multi-cohort, open-label, phase 1/2 study. Lancet Oncol 2021, 22, 959–969. [Google Scholar] [CrossRef] [PubMed]
- Shigematsu, H.; Takahashi, T.; Nomura, M.; Majmudar, K.; Suzuki, M.; Lee, H.; Wistuba, II; Fong, K. M.; Toyooka, S.; Shimizu, N.; et al. Somatic mutations of the HER2 kinase domain in lung adenocarcinomas. Cancer Res 2005, 65, 1642–1646. [Google Scholar] [CrossRef] [PubMed]
- De Greve, J.; Teugels, E.; Geers, C.; Decoster, L.; Galdermans, D.; De Mey, J.; Everaert, H.; Umelo, I.; In't Veld, P.; Schallier, D. Clinical activity of afatinib (BIBW 2992) in patients with lung adenocarcinoma with mutations in the kinase domain of HER2/neu. Lung Cancer 2012, 76, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Kris, M.G.; Camidge, D.R.; Giaccone, G.; Hida, T.; Li, B.T.; O'Connell, J.; Taylor, I.; Zhang, H.; Arcila, M.E.; Goldberg, Z.; et al. Targeting HER2 aberrations as actionable drivers in lung cancers: phase II trial of the pan-HER tyrosine kinase inhibitor dacomitinib in patients with HER2-mutant or amplified tumors. Ann Oncol 2015, 26, 1421–1427. [Google Scholar] [CrossRef] [PubMed]
- Mazieres, J.; Barlesi, F.; Filleron, T.; Besse, B.; Monnet, I.; Beau-Faller, M.; Peters, S.; Dansin, E.; Fruh, M.; Pless, M.; et al. Lung cancer patients with HER2 mutations treated with chemotherapy and HER2-targeted drugs: results from the European EUHER2 cohort. Ann Oncol 2016, 27, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Hyman, D.M.; Piha-Paul, S.A.; Won, H.; Rodon, J.; Saura, C.; Shapiro, G.I.; Juric, D.; Quinn, D.I.; Moreno, V.; Doger, B.; et al. HER kinase inhibition in patients with HER2- and HER3-mutant cancers. Nature 2018, 554, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Li, Y.; Chen, S.; Ying, S.; Xu, S.; Huang, J.; Wu, D.; Lv, D.; Bei, T.; Liu, S.; et al. Efficacy and safety of pyrotinib in advanced lung adenocarcinoma with HER2 mutations: a multicenter, single-arm, phase II trial. BMC Med 2022, 20, 42. [Google Scholar] [CrossRef]
- Pacini, L.; Jenks, A.D.; Lima, N.C.; Huang, P.H. Targeting the Fibroblast Growth Factor Receptor (FGFR) Family in Lung Cancer. Cells 2021, 10. [Google Scholar] [CrossRef]
- Krook, M.A.; Reeser, J.W.; Ernst, G.; Barker, H.; Wilberding, M.; Li, G.; Chen, H.Z.; Roychowdhury, S. Fibroblast growth factor receptors in cancer: genetic alterations, diagnostics, therapeutic targets and mechanisms of resistance. Br J Cancer 2021, 124, 880–892. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Radulovich, N.; Ng, C.; Liu, N.; Notsuda, H.; Cabanero, M.; Martins-Filho, S.N.; Raghavan, V.; Li, Q.; Mer, A.S.; et al. Organoid Cultures as Preclinical Models of Non-Small Cell Lung Cancer. Clin Cancer Res 2020, 26, 1162–1174. [Google Scholar] [CrossRef]
- Facchinetti, F.; Hollebecque, A.; Bahleda, R.; Loriot, Y.; Olaussen, K.A.; Massard, C.; Friboulet, L. Facts and New Hopes on Selective FGFR Inhibitors in Solid Tumors. Clin Cancer Res 2020, 26, 764–774. [Google Scholar] [CrossRef]
- Vyse, S.; Huang, P.H. Targeting EGFR exon 20 insertion mutations in non-small cell lung cancer. Signal Transduct Target Ther 2019, 4, 5. [Google Scholar] [CrossRef]
- Yun, J.; Lee, S.H.; Kim, S.Y.; Jeong, S.Y.; Kim, J.H.; Pyo, K.H.; Park, C.W.; Heo, S.G.; Yun, M.R.; Lim, S.; et al. Antitumor Activity of Amivantamab (JNJ-61186372), an EGFR-cMet Bispecific Antibody, in Diverse Models of EGFR Exon 20 Insertion-Driven NSCLC. Cancer Discovery 2020, 15, 15. [Google Scholar] [CrossRef]
- Banda, M.; McKim, K.L.; Myers, M.B.; Inoue, M.; Parsons, B.L. Outgrowth of erlotinib-resistant subpopulations recapitulated in patient-derived lung tumor spheroids and organoids. PLoS One 2020, 15, e0238862. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, L.; Zhu, Y.; Huang, C.; Qin, Y.; Liu, H.; Ren-Heidenreich, L.; Shi, B.; Ren, H.; Chu, X.; et al. Coexistence of EGFR with KRAS, or BRAF, or PIK3CA somatic mutations in lung cancer: a comprehensive mutation profiling from 5125 Chinese cohorts. Br J Cancer 2014, 110, 2812–2820. [Google Scholar] [CrossRef]
- Taverna, J.A.; Hung, C.N.; DeArmond, D.T.; Chen, M.; Lin, C.L.; Osmulski, P.A.; Gaczynska, M.E.; Wang, C.M.; Lucio, N.D.; Chou, C.W.; et al. Single-Cell Proteomic Profiling Identifies Combined AXL and JAK1 Inhibition as a Novel Therapeutic Strategy for Lung Cancer. Cancer Res 2020, 80, 1551–1563. [Google Scholar] [CrossRef] [PubMed]
- Dost, A.F.M.; Moye, A.L.; Vedaie, M.; Tran, L.M.; Fung, E.; Heinze, D.; Villacorta-Martin, C.; Huang, J.; Hekman, R.; Kwan, J.H.; et al. Organoids Model Transcriptional Hallmarks of Oncogenic KRAS Activation in Lung Epithelial Progenitor Cells. Cell Stem Cell 2020, 27, 663–678.e668. [Google Scholar] [CrossRef]
- Vlachogiannis, G.; Hedayat, S.; Vatsiou, A.; Jamin, Y.; Fernandez-Mateos, J.; Khan, K.; Lampis, A.; Eason, K.; Huntingford, I.; Burke, R.; et al. Patient-derived organoids model treatment response of metastatic gastrointestinal cancers. Science 2018, 359, 920–926. [Google Scholar] [CrossRef]
- Liu, L.; Yu, L.; Li, Z.; Li, W.; Huang, W. Patient-derived organoid (PDO) platforms to facilitate clinical decision making. Journal of translational medicine 2021, 19, 40. [Google Scholar] [CrossRef] [PubMed]
- Putker, M.; Millen, R.; Overmeer, R.; Driehuis, E.; Zandvliet, M.M.J.M.; Clevers, H.; Boj, S.F.; Li, Q.-X. Medium-Throughput Drug- and Radiotherapy Screening Assay using Patient-Derived Organoids. Journal of visualized experiments : JoVE. [CrossRef]
- Dzobo, K. Taking a Full Snapshot of Cancer Biology: Deciphering the Tumor Microenvironment for Effective Cancer Therapy in the Oncology Clinic. Omics 2020, 24, 175–179. [Google Scholar] [CrossRef]
- Mazzocchi, A.; Devarasetty, M.; Herberg, S.; Petty, W.J.; Marini, F.; Miller, L.; Kucera, G.; Dukes, D.K.; Ruiz, J.; Skardal, A.; et al. Pleural Effusion Aspirate for use in 3D Lung Cancer Modeling and Chemotherapy Screening. ACS Biomater Sci Eng 2019, 5, 1937–1943. [Google Scholar] [CrossRef] [PubMed]
- Mazzocchi, A.; Dominijanni, A.; Soker, S. Pleural Effusion Aspirate for Use in 3D Lung Cancer Modeling and Chemotherapy Screening. Methods Mol Biol 2022, 2394, 471–483. [Google Scholar] [CrossRef]
- Palechor-Ceron, N.; Krawczyk, E.; Dakic, A.; Simic, V.; Yuan, H.; Blancato, J.; Wang, W.; Hubbard, F.; Zheng, Y.L.; Dan, H.; et al. Conditional Reprogramming for Patient-Derived Cancer Models and Next-Generation Living Biobanks. Cells 2019, 8, 27. [Google Scholar] [CrossRef]
- Dijkstra, K.K.; Monkhorst, K.; Schipper, L.J.; Hartemink, K.J.; Smit, E.F.; Kaing, S.; de Groot, R.; Wolkers, M.C.; Clevers, H.; Cuppen, E.; et al. Challenges in Establishing Pure Lung Cancer Organoids Limit Their Utility for Personalized Medicine. Cell Rep 2020, 31, 107588. [Google Scholar] [CrossRef]
- Xu, H.; Jiao, Y.; Qin, S.; Zhao, W.; Chu, Q.; Wu, K. Organoid technology in disease modelling, drug development, personalized treatment and regeneration medicine. Exp Hematol Oncol 2018, 7, 30. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, S.N.; Ingber, D.E. Microfluidic organs-on-chips. Nat Biotechnol 2014, 32, 760–772. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.J.; Chieh, J.A.; Chen, Y.C.; Lee, K.Y.; Chan, Y.F.; Ho, S.C.; Sun, W.l.; Wang, Y.S.; Huang, W.C.; Chang, W.C.; et al. Lung Cancer On Chip for Testing Immunotherapy. In Proceedings of the 2021 21st International Conference on Solid-State Sensors, Actuators and Microsystems (Transducers), 20-24 June 2021; 2021; pp. 1032–1035. [Google Scholar]
- Jiang, X.; Ren, L.; Tebon, P.; Wang, C.; Zhou, X.; Qu, M.; Zhu, J.; Ling, H.; Zhang, S.; Xue, Y.; et al. Cancer-on-a-Chip for Modeling Immune Checkpoint Inhibitor and Tumor Interactions. Small 2021, 17, e2004282. [Google Scholar] [CrossRef]
- Hassell, B.A.; Goyal, G.; Lee, E.; Sontheimer-Phelps, A.; Levy, O.; Chen, C.S.; Ingber, D.E. Human Organ Chip Models Recapitulate Orthotopic Lung Cancer Growth, Therapeutic Responses, and Tumor Dormancy In Vitro. Cell Rep 2017, 21, 508–516. [Google Scholar] [CrossRef]
- Ingber, D.E. From tensegrity to human organs-on-chips: implications for mechanobiology and mechanotherapeutics. Biochem J 2023, 480, 243–257. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Karim, M.; Hasan, M.M.; Hooper, J.; Wahab, R.; Roy, S.; Al-Hilal, T.A. Cancer-on-a-Chip: Models for Studying Metastasis. Cancers (Basel) 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, S.; Pinho, D.; Vila, A. Development of organoid-on-a-chip platform for preclinical drug screening. In Proceedings of the 5th World Congress on Recent Advances in Nanotechnology, RAN 2020; 2022; pp. 117-111–117-112. [Google Scholar]
- Charelli, L.E.; Ferreira, J.P.D.; Naveira-Cotta, C.P.; Balbino, T.A. Engineering mechanobiology through organoids-on-chip: A strategy to boost therapeutics. Journal of tissue engineering and regenerative medicine 2021, 15, 883–899. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.W.; Lee, S.Y.; Lee, D.W. A Cancer Spheroid Array Chip for Selecting Effective Drug. Micromachines (Basel) 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Konar, D.; Devarasetty, M.; Yildiz, D.V.; Atala, A.; Murphy, S.V. Lung-On-A-Chip Technologies for Disease Modeling and Drug Development. Biomed 2016, 7, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, S.; Palmberg, L. Air-Liquid Interface: Relevant In Vitro Models for Investigating Air Pollutant-Induced Pulmonary Toxicity. Toxicol Sci 2018, 164, 21–30. [Google Scholar] [CrossRef]
- Dvorak, A.; Tilley, A.E.; Shaykhiev, R.; Wang, R.; Crystal, R.G. Do airway epithelium air-liquid cultures represent the in vivo airway epithelium transcriptome? Am J Respir Cell Mol Biol 2011, 44, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Pezzulo, A.A.; Starner, T.D.; Scheetz, T.E.; Traver, G.L.; Tilley, A.E.; Harvey, B.G.; Crystal, R.G.; McCray, P.B., Jr.; Zabner, J. The air-liquid interface and use of primary cell cultures are important to recapitulate the transcriptional profile of in vivo airway epithelia. Am J Physiol Lung Cell Mol Physiol 2011, 300, L25–31. [Google Scholar] [CrossRef]
- Fulcher, M.L.; Randell, S.H. Human nasal and tracheo-bronchial respiratory epithelial cell culture. Methods Mol Biol 2013, 945, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Mathis, C.; Poussin, C.; Weisensee, D.; Gebel, S.; Hengstermann, A.; Sewer, A.; Belcastro, V.; Xiang, Y.; Ansari, S.; Wagner, S.; et al. Human bronchial epithelial cells exposed in vitro to cigarette smoke at the air-liquid interface resemble bronchial epithelium from human smokers. Am J Physiol Lung Cell Mol Physiol 2013, 304, L489–503. [Google Scholar] [CrossRef]
- Sachs, N.; Papaspyropoulos, A.; Zomer-van Ommen, D.D.; Heo, I.; Bottinger, L.; Klay, D.; Weeber, F.; Huelsz-Prince, G.; Iakobachvili, N.; Amatngalim, G.D.; et al. Long-term expanding human airway organoids for disease modeling. Embo J 2019, 38. [Google Scholar] [CrossRef] [PubMed]
- Barbosa Rabago, D.; Blakely, C.M.; Haderk, F.; Bivona, T.G. Profiling Sensitivity to Targeted Therapies in EGFR-Mutant NSCLC Patient-Derived Organoids. Journal of visualized experiments : JoVE 2021. [Google Scholar] [CrossRef]
Figure 1.
The illustration of applications of patient-derived lung cancer organoids for precision medicine.
Figure 1.
The illustration of applications of patient-derived lung cancer organoids for precision medicine.
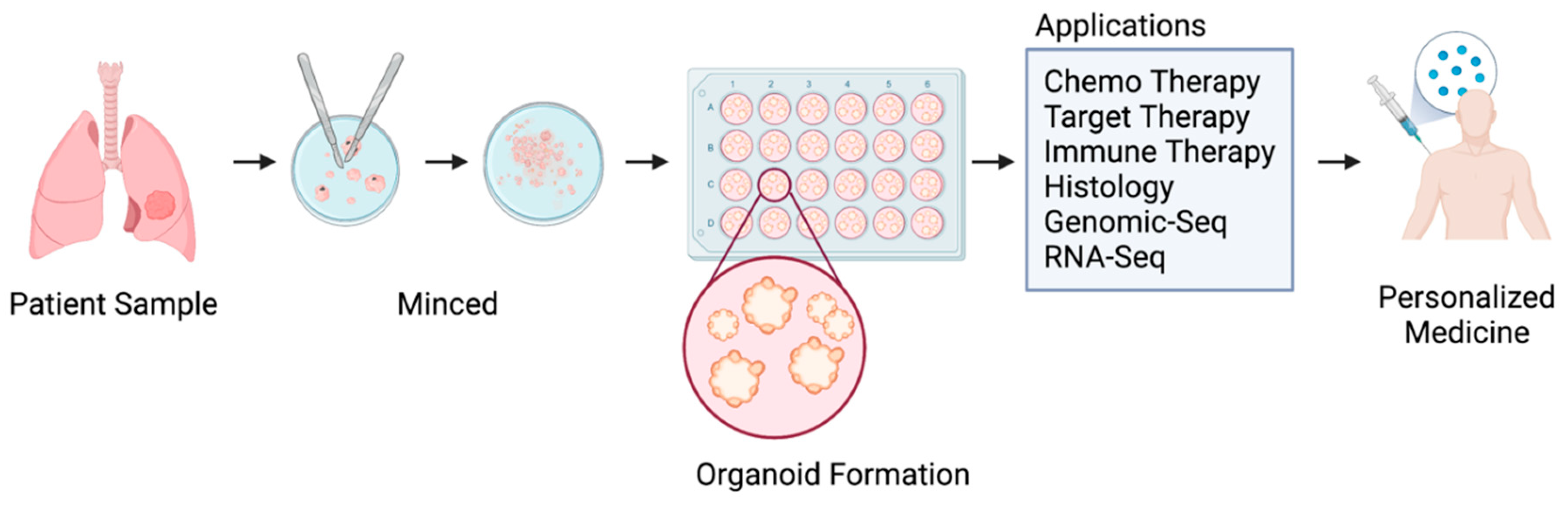

Table 1.
Literature search strategy in Database(s): Ovid MEDLINE(R) ALL from 1946 to March 23, 2023.
Table 1.
Literature search strategy in Database(s): Ovid MEDLINE(R) ALL from 1946 to March 23, 2023.
| # | Searches | Results |
|---|---|---|
| 1 | (((patient-derived adj3 organoid*) or patient derived) adj3 organoid*).ti,ab. | 1187 |
| 2 | exp Organoids/de [Drug Effects] | 1031 |
| 3 | (((human-derived adj3 organoid*) or human derived) adj3 organoid*).ti,ab. | 35 |
| 4 | 1 or 3 | 1218 |
| 5 | 2 and 4 | 96 |
| 6 | drug screening.mp. or exp Drug Evaluation, Preclinical/ | 297911 |
| 7 | 4 and 6 | 269 |
| 8 | 5 or 7 | 312 |
Table 2.
Summary of organoids as an enhanced model for personalized drug screening.
| Cancer type | Organoid Model | PDX Model (n) | Compounds Tested | Technology/ Unique culture supplement | Limitations | Ref |
|---|---|---|---|---|---|---|
| Adenocarcinoma | Primary lung tumor spheroids (n=14) were cultured in 3D-Matrigel culture methods | N/A | Erlotinib |
|
The number of different drug and dose combinations that can be investigated at one time is limited. | [78] |
| Adenocarcinoma (NSCLC patients with tumors stage I–III, EGFR L858R, EGFR Ex20 ins, KRAS G12C) |
1) Fresh tumor samples harvested for organoid culture. 2) Primary tumor samples and PDOs were analyzed via whole-exome sequencing and IHC. (n=7) |
N/A | 26 antineoplastic drugs tested (gefitinib, osimertinib, afatinib) | Small cohort sized used and will require further large-scale analyses to validate the findings. | [50] | |
| Small cell lung cancer – refractory (tumors stage I–III, EGFR L858R, EGFR Ex20 ins, KRAS G12C) | PDOs (n=4) were developed from human SCLC PDX samples to test if TS inhibition could be a viable strategy for SCLC treatment. | PDXs were generated from SCLC tumor biopsy samples. |
|
|
[59] | |
| 1)Adenocarcinoma 2)Squamous cell carcinoma 3)Adenosquamous carcinoma 4)Large cell carcinoma 5)Small cell lung cancer |
Surgically resected lung cancer tissues from 36 patients were embedded in Matrigel and submerged in MBM to create PDOs (n=80). |
|
|
Limitation of cancer organoid models is the lack of a cancer microenvironment. | [16] | |
| NSCLC | PDOs derived from NSCLC were cultured in vitro (n=10) | N/A |
|
[52] | ||
| Adenocarcinoma | Developed PDOs from human lung adenocarcinoma biopsy samples (n=12) | N/A |
|
Small sample size limits the power to detect molecular markers of drug response. | [53] | |
| Small cell lung cancer (n=1) | Tumor organoids were generated from primary lung cancer cells from patients with SCLC. | N/A |
|
|
Small sample sized used and will require further large-scale analyses to validate the findings. | [58] |
| Adenocarcinoma Metastatic lung cancer |
Developed PDOs from pleural effusion of patients with lung adenocarcinoma (n=2) | N/A | Cisplatin + pemetrexed; carboplatin + pemetrexed; crizotinib |
|
Small sample size. Will require further large-scale analyses to validate the findings. | [86] |
| Non-small cell lung cancer | PDO (n=1) | N/A | S-40 (oral potent tubulin destabilizing agent) | No detail description of organoid culture | Small sample size | [60] |
| 1)Squamous cell carcinoma 2)Adenosquamous carcinoma |
PDOs developed from human tumor lung cancer surgical specimens, (n=3) | N/A | 86 antineoplastic drugs tested, including molecular targeted drugs, immune checkpoint inhibitors, and cytotoxic chemotherapy |
|
Few and limited PDO examples were selected for each type drug screen. | [17] |
| 1)Lung adenocarcinoma 2)Squamous cell |
PDOs developed from 19 surgically resected lungadenocarcinomas (LUAD) and 15 lung squamous cell carcinomas (LUSC), |
|
|
[74] | ||
| Lung adenocarcinoma | Three primary lung cancer organoid models:
|
N/A |
|
Organoids culture condition previously described [74] | [63] | |
| 1) Squamous cell carcinoma 2) Adenocarcinoma 3) small cell lung cancer |
PDOs (n=21) generated from 16 adenocarcinoma and 4 squamous cell carcinoma, two small cell lung cancer organoids | On-chip drug responses of the PDX derived organoids (n=3) were consistent with the in vivo PDX results |
|
|
Well-designed pilot study would improve sensitivity and specificity of the assay. | [64] |
| Lung adenocarcinoma. | Organoids (n=84) were established from patients with advanced lung adenocarcinoma. |
|
Organoids culture medium previously described [106] | This was a retrospective study which cannot provide real-time information for clinical decision making. |
[61] | |
| Lung adenocarcinoma | PDOs (n=5) were developed from fresh lung tumors obtained from treatment naïve patients. | N/A |
|
Single cell CyTOF analysis of tumor tissues | Not for high throughput screening with a higher cost | [80] |
| Adenocarcinoma HER2 mutant |
PDO (n=1) was developed from human lung tumor specimens, | Human lung tumor fragments were subcutaneously implanted in mice for generating PDX |
|
One PDO and small size of clinical trial patient cohort | [49] | |
| NSCLC | PDOs (n=2) were developed from malignant pleural effusions of patients with NSCLC with Exon20ins mutations. | Human lung tissue specimens were implanted subcutaneously in mice | Amivantamab | Organoids culture medium previously described [106] | [77] | |
| EGFR-Mutant NSCLC | Patient surgical resection or tumor biopsy specimens (n=3). | N/A | Osimertinib | Organoids culture medium previously described [106] |
|
[107] |
Table 3.
Summary of organoids culture medium.
| Ref | Matrigel | Base medium | Bovine serum albumin | N2 | B27 | EGF (ng/mL) |
bFGF (ng/mL) |
FGF-10 (ng/mL) |
FGF-4 (ng/mL) |
Y-27632 (ROCK inhibitor) |
GlutaMax (L-glutamine alternative) |
HEPES (mM) |
R-spondin 1 (ng/mL) |
Noggin (ng/mL) |
Nicotinaminde(mM) | Prostaglandin E2 | SB202190 | N-acetylcysteine | A83-01 | Note |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| [78] | Yes | DMEM/F12 | 25% | Yes | 10 μM | Yes | Stem Pro hESC Supplement, 0.1mM 2-mercaptoethanol | |||||||||||||
| [50] | Yes | DMEM/F12 | Yes | Yes | 50 | 10 μM | ||||||||||||||
| [59] | N/A | HyStem-HP hydrogel kits | ||||||||||||||||||
| [16] | Yes | DMEM/F12 | Yes | Yes | 50 | 20 | 10 μM | |||||||||||||
| [52] | Yes | Advanced DMEM/F12 | 0.01% | Yes | Yes | 50 | 1 | 20 | 250 | 100 | 10 | 1μM | 10 μM | 1 mM | 500 nM | 2mM L-glutamine, 100 ng/ml Wnt3a, 10 nM gastrin 1 | ||||
| [53] | Yes | Advanced DMEM/F12 | Yes | Yes | 20 | 10 mM | Yes | 500 | 100 | 10 | 10 mM | 1.25 mM | 500 nM | 25 ng/mL FGF-7 | ||||||
| [58] | Yes | DMEM/F12 | Yes | Yes | 50 | 20 | 10 μM | |||||||||||||
| [86] | hydrogel cultures | RPMI 1640 | 5% fetal bovine serum | |||||||||||||||||
| [60] | No detailed description | |||||||||||||||||||
| [17] | FBIM001 medium [54] | |||||||||||||||||||
| [74] | Yes | RPMI 1640 | Yes | Yes | 50 | 100 | 100 | 10 μM | Yes | 10 | 100 | 500 nM | 250 nM CHIR 99021, 100 nM SAG | |||||||
| [63] | Yes | RPMI 1640 | Yes | Yes | 50 | 100 | 100 | 10 μM | Yes | 10 | 100 | 500 nM | 250 nM CHIR 99021, 100 nM SAG | |||||||
| [64] | Yes | DMEM/F12 | Yes | Yes | 50 | 10 μM | Yes | 10 | 5 | 3 μM | 1 mM | 5 μM | 10 μM Forskolin, 3 nM Dexamethasone | |||||||
| [61] | Yes | Advanced DMEM/F12 | Yes | 100 | 5 μM | Yes | 10 | 500 | 100 | 5 | 500 nM | 1.25 mM | 500 nM | 25 ng/mL FGF-7 | ||||||
| [80] | Yes | Advanced DMEM/F12 | Yes | Yes | 50 | 10 | 10 | 10 μM | 500 | 100 | 4 | 1μM | 5 μM | 500 nM | 20 ng/ml HGF | |||||
| [49] | Yes | Advanced DMEM/F12 | Yes | 50 | 10 | Yes | 10 | Yes* | Yes* | 10 | 10 μM | 1.25 mM | 500 nM | R-spondin and Noggin form condition medium, 1 ng/ml FGF2, Dihydrotestosterone (DHT) | ||||||
| [77] | Yes | Advanced DMEM/F12 | Yes | 100 | 5 μM | Yes | 10 | 500 | 100 | 5 | 500 nM | 1.25 mM | 500 nM | 25 ng/mL FGF-7 | ||||||
| [107] | Yes | Advanced DMEM/F12 | Yes | 100 | 5 μM | Yes | 10 | 500 | 100 | 5 | 500 nM | 1.25 mM | 500 nM | 25 ng/mL FGF-7 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



