
Submitted:
15 June 2023
Posted:
16 June 2023
You are already at the latest version
Abstract
Epidemiological data highlight prostate cancer as a significant global health issue, with high incidence and substantial impact on patients' quality of life. The prevalence of this disease is associated with various factors, including age, heredity, and race. Recent research in prostate cancer genetics has identified several genetic variants that may be associated with an increased risk of developing the disease. However, despite the significance of these findings, genetic markers for prostate cancer are not currently utilized in clinical practice as reliable indicators of the disease. In addition to genetics, epigenetic alterations also play a crucial role in prostate cancer development. Aberrant DNA methylation, changes in chromatin structure, and microRNA (miRNA) expression are major epigenetic events that influence oncogenesis. Existing markers for prostate cancer, such as prostate-specific antigen (PSA), have limitations in terms of sensitivity and specificity. The cost of testing, follow-up procedures, and treatment for false-positive results and overdiagnosis contributes to the overall healthcare expenditure. Improving the effectiveness of prostate cancer diagnosis and prognosis requires either narrowing the risk group by identifying new genetic factors or enhancing the sensitivity and specificity of existing markers. Immunological biomarkers (both circulating and intra-tumoral), including markers of immune response and immune dysfunction, represent a potentially useful area of research for enhancing the diagnosis and prognosis of prostate cancer. Our review emphasizes the need for developing novel immunological biomarkers to improve the diagnosis, prognosis, and management of prostate cancer. We highlight the most recent achievements in the identification of biomarkers provided by circulating monocytes and tumor-associated macrophages (TAMs). We highlight that monocytes-derived and TAMs-derived biomarkers can enable to establish the missing links between genetic predisposition, hormonal metabolism and immune responses in prostate cancer.
Keywords:
cancer genetics
; cancer epigenetic
; tumor microenvironment
; monocytes
; TAMs
; prostate cancer
; cancer biomarkers
1. Epidemiology
The World Health Organization (WHO) estimates that cancer was responsible for 10 million deaths in 2020, accounting for nearly one in six deaths. There were 1,414,259 new cases of prostate cancer (PCa) worldwide and 375,304 deaths from this type of cancer in 2020 according to the Global Cancer Observatory (GCO) (gco.iarc.fr). PCa ranks third in terms of incidence and second in terms of cancers affecting men globally [1]. In terms of mortality, PCa ranks eighth worldwide (Table 1). Overall, PCa statistics are relatively similar across different regions, although it ranks first among cancers affecting men in Latin America and eighth in Asia [2]. This suggests the potential impact of racial, ethnic, socioeconomic factors, and variations in the biology of prostate carcinogenesis, including the genetic predisposition of certain groups to develop biologically aggressive forms of the disease. Even when considering geographically diverse populations with similar access to healthcare, men of African descent have a higher incidence of PCa and worse prognosis. African American men, Caribbean men, and black men in Europe exhibit higher incidence rates than white men, with mortality rates approximately twice as high, indicating a genetic predisposition to PCa development [3,4]. Analyses of PCa biopsies from men in sub-Saharan Africa have revealed a significant proportion of high-grade tumors (Gleason score 8+) However, variations in healthcare access and differences in cancer case registration across countries limit the reliability of conclusions regarding the correlation between race, ethnicity, geography, and PCa aggressiveness based solely on epidemiological data [3]. Furthermore, the significance of racial differences in PCa outcomes diminishes significantly if the male population in the analyzed countries has equal access to healthcare services [4,5,6].
The worldwide incidence and mortality of PCa are strongly correlated with increasing age, with the average age at the time of primary diagnosis being 67 years [7]. Epidemiological studies conducted globally have reported the highest incidence of PCa in individuals aged 75-79 years. In 2022, the incidence rates were recorded as 155 cases per 100,000 for ages 55-59, 510 cases per 100,000 for ages 65-69, and 751 cases per 100,000 for ages 75-79 [2].
Nineteen international studies, spanning from 1935 to 2014 and including countries such as Scandinavia, Caucasus, France, Hungary, Greece, Japan, China, Singapore, and North America, examined the histopathological evidence of PCa in autopsies of 6,024 males aged between 50-59, regardless of the cause of death. These studies found that approximately half of the individuals had histopathological evidence of prostate cancer, although only 3.8% died from PCa [8].
The prognosis for patients newly diagnosed with PCa varies significantly depending on the stage of the disease. If the disease is detected at early stages, men with localized PCa can have a life expectancy of up to 99% for more than 10 years, depending on age, comorbidities, and chosen treatment strategy [9]. International statistics indicate that around 5% of men diagnosed with PCa develop distant metastases, and the overall five-year survival rate for metastatic disease is 30% [9]. Almost all patients with metastatic PCa eventually progress to a castration-resistant form that is resistant to androgen deprivation therapy. These factors contribute significantly to the morbidity and mortality associated with PCa [9].
While it is widely recognized that the immune system plays a crucial role in controlling cancer initiation and progression, there has been insufficient attention given to epidemiological studies examining the link between immune status, subclinical inflammation of infectious or endogenous origin, and the incidence of prostate cancer. It would be of great interest to further investigate how inflammation, leading to subclinical activation of innate immunity, may facilitate the development of prostate cancer.
2. Genetic predisposition for prostate cancer
2.1. Genetic markers of PCa
The vast majority of PCa cases are sporadic and caused by turning off tumor suppressor genes and turning on oncogenes and only up to 5-10% of PCa cases have family history, and are caused by germline mutations [10]. The presence of a first-degree PCa relatives is a risk factor associated with a two- to three-fold PCa compared to men without a family history of PCa [11]. Epidemiological and case-control studies have shown the inheritance of specific mutations in PCa susceptibility genes and reported that patients with these mutations have an increased risk of the disease [9]. Thus BRCA 1/2 germline mutations in men increase the risk of PCa relative to the average population level by 3-8 times. Genetic alterations in BRCA2 in PCa patients are associated with aggressive behavior of the disease [11]. Sequencing of DNA samples in men with a family PCa history reveals a high frequency of DNA repair gene mutations, such as BRCA1/2, HOX, ATM, CHECK2, PALB2 and RAD51D, RNase L (HPC1, lq22), MSR1 (8p), and ELAC2/HPC2 (17p11) (Figure 1) [12]. They are involved in the maintenance of genome stability, particularly in the processes of repair by homologous recombination (HRR), repair of misfolded nucleotides (mismatch repair - MMR), also inter-chain DNA cross-links. BRCA1 and BRCA2 genes predisposing to PCa are also found to be involved in pathways of control of checkpoints for DNA damage and replication fork protection during replication [13].
PCa tumor cells are characterized by genome instability both at chromosomal and gene levels. Genomic gains of chromosomes 7 and 8q and heterozygous losses of 8p, 13q, 16q , and 18 are often met in PCa [14]. Besides, PCa tumors contain a large number of structural gene rearrangements, primarily translocations and deletions, the number of which increases markedly with the disease progression. More than half of primary PCa patients (up to 70%) show formation of the chimeric oncogene TMPRSS2-ERG - a product of fusion of the 5'-untranslated region of androgen-regulated gene TMPRSS2 (chr21q22.2) with ETS family of transcription factors genes, namely ERG (chr21q22.3) that results in aberrant expression of ERG [14,15,16,17]. TMPRSS2:ERG fusion is an early event in prostate carcinogenesis, it is absent in benign prostate hyperplasia (BPH) and in normal prostate tissues and is considered as diagnostic and prognostic marker for PCa [18,19,20]. ERG-positive PCs are associated with a distinct spectrum of hypermethylation accompanied by epigenetically silenced genes, revealing a greater molecular and biological diversity (Cancer Genome Atlas Research Network, 2015). Numerous studies have found a correlation between fusion of the TMPRSS2 and ETS family genes (ERG, ETV1, ETV4, ETV5) and PCa [18]. The fusion of these genes makes it possible for the ETS genes to be activated under by the action of activators of the TMPRSS2 gene, thus triggering the cancer process in prostate cells [17]. ERG fusion genes are regulated by androgen receptor. In primary PCa, ERG binds and involve transcription factors AR, FOXA1, and HOXB13 leading to activating the Wnt and Notch pathways [21], [22]. In ERG-positive PCa cases with PTEN loss PI3K signaling is shown to be activated that results in increased proliferation and invasion. There are no specific prognostic biomarkers for ERG-positive tumors [23]. But it’s important to note that aggressive ERG-negative PCa is associated with SPOP/FOXA1 mutation, CHD1 (5q15-q21) deletion, and SPINK1 expression. Gleason scale, PSA, and other biomarkers are exclusively for ETS-negative patients [24].
Primary PCa germinal and somatic tumors are very diverse genetically. In recent years, due to the rapid development of molecular genetic methods, new approaches to genetic testing were developed. Next Generation Sequencing (NGS) has been introduced into clinical practice and has opened a new approach to genetic testing. It allows to perform multiple genes panel testing of germline and somatic mutations, and to identify rare variants in genes with moderate penetrance [25]. The presence of these mutations leads to an increased risk of PCa phenotype over a lifetime by 35–60%. The Cancer Genome Atlas (TCGA) consortium reported recurrent mutations or copy-number alterations within the following genes - SPOP, FOXA1, IDH1, TP53, PTEN, PIK3CA, BRAF, CTNNB1, HRAS, MED12, ATM, CDKN1B, RB1, NKX3-1, AKT1, ZMYM3, KMT2C, KMT2D, ZNF770, CHD1, BRCA2, CDK12 [14]. Recent investigations revealed metastatic castration-resistant PCa have a similar mutational landscape, but it harbored a higher mutational load and frequency of large-scale structural variants compared to primary PCa [14]. However, the list of driver genes remained almost similar PRAD; AR, TP53, MYC, ZMYM3, PTEN, PTPRD, ZFP36L2, ADAM15, MARCOD2, BRIP1, APC, KMT2C, CCAR2, NKX3-1, C8orf58, and RYBP, except AR gene mutations which were the result of PCa treatment [26]. Somatic alterations affecting AR expression, AR gene amplifications and treatment-associated mutations have been detected to be a major driver of castration resistance [26].
2.2. GWAS in PCa
Genome-wide association studies (GWAS) with the use of multi-ancestry approach are necessary to discover new risk variants for prostate cancer, to refine lead variants in known risk regions and to develop genetic risk scores for PCa for effective stratifying PCa across populations. To date GWAS and precise mapping studies of PCa have been mainly performed in men from European, African, East Asian and Hispanic populations to identify common genetic variants associated with disease risk across populations [27]. Approximately 270 loci harboring hundreds of single nucleotide polymorphisms (SNPs) associated with PCa risk have been revealed [27]. PCa susceptibility loci were found on all chromosomes except chromosomes 15, 16, 21, and 23. PCa risk-associated SNPs were highly enriched in noncoding cis-regulatory genomic regions [28]. Of 191 independent and replicated associations that reached genome-wide significance (i.e., p<10−8), 123 (64%) were reported in populations of European origin, 21 (11%) in populations of Southeast Asia and 45 (24%) in other populations. Only one locus (17q21) reached genome-wide significance in populations of African ancestry [28]. Khan et al. showed little heterogeneity in associated PCa variants across populations [3]. However, many loci found in populations of European or Asian ancestry were not replicated in populations of African ancestry, or the effect size was smaller (or directly opposite) for the race [3]. Several hypotheses have been proposed to explain the complexity of replication and differences in the magnitude of genetic effects depending on race and ethnicity. First, the underlying genetic predisposition, and hence the biology of PCa, can differ fundamentally by racial or ethnic group [29]. This explanation has limitations, as it implies that the biological basis of PCa depends on race or ethnicity. However, this hypothesis cannot be ruled out based on the available data. The most accurate explanation is that risk alleles and the underlying population structure of PCa susceptibility loci differ by ethnicity or race, and that these differences likely affect the ability to detect genetic associations [29]. Polymorphic variants in the 8q24 region are associated not only with PCa, but also with other cancers in different populations [29]. However, the biological mechanism of the influence of SNPs in this region, leading to prostate carcinogenesis, is unclear, since this region does not contain any coding DNA regions. The closest gene to this region is MYC, a proto-oncogene that is disrupted during carcinogenesis [29]. The 8q24 region may influence the expression of MYC gene. The influence of 8q24 on Wnt signaling has also been determined [29]. As a result of the OncoArray project, 63 new loci associated with PCa susceptibility have been identified, 52 of them were identified by imputation of OncoArray genotyping data [30]. A novel 6q27 variant (rs138004030) was found to be significantly associated with early PCa onset [30]. Among the new 63 variants, several candidate genes were revealed; one of them is a missense variant of ATM gene rs1800057 [31]. Although this missense mutation was classified as "benign" in the ClinVar database, ATM has been implicated in the development of PCa and is strongly associated with the aggressive course of the disease. The ATM protein is a key checkpoint kinase that acts as a regulator of a wide range of downstream proteins, including TP53 and BRCA1, the CHEK2 checkpoint kinase, the RAD17 and RAD9 checkpoint proteins, and the NBS1 DNA repair protein [31]. Another missense variant (rs2066827) has been identified in CDKN1B gene (cyclin-dependent kinase 1B inhibitor), which belongs to the Cip/Kip family of cyclin-dependent kinase inhibitors [29]. CDKN1B controls cell cycle progression to the G1 stage, and in vitro studies have shown that CDKN1B levels to be associated with increased tumor size and tumor grade [29]. This particular variant has previously been associated with familial PCa and disease progression. One more candidate gene RASSF3 rs7968403 has been identified. RASSF3 is a plasma membrane GTP-binding protein and it is a member of the RAS signaling pathway that is aberrant in about one-third of cancers [29]. GWAS meta-analysis for Japanese and Chinese populations identified rs12791447/11p15.4 and rs58262369/14q23.2. The rs58262369 polymorphic locus is located in the 3'-untranslated region of ESR2, which encodes for estrogen receptor 2 [32]. Animal studies have shown high expression of ESR2 in normal prostate epithelial cells, and ESR2 knockout mice develop prostatic hyperplasia [32]. TCGA data showed that ESR2 is upregulated in prostate tumors compared to normal prostate tissues, however, this does not correlate with increased mRNA expression [32]. rs12791447 is located in the intron of the PPFIBP2 gene. According to TCGA, the expression level of this gene is significantly lower in tumors compared to normal prostate tissue [32]. To date, almost all available GWAS data have been obtained for the population of Western Europe and the white population of North America (79.8%, [33], therefore, further research is needed with the inclusion of populations from other regions, which will supplement existing information and will identify risk factors specific to different populations [34]. A population-specific approach and the ability of homogeneous populations to detect disease-specific SNPs is important for GWAS studies. At the same time, homogeneous population material provides a resource for checking previous GWAS results performed on mixed populations.
Future GWAS results will provide further support for a contribution of germline variation to ancestry differences in PCa incidence. The clinical benefit of genetic risk scores profiling for targeted screening and early diagnosis also needs to be examined, and larger PCa consortia in men of non-European ancestry will be required to identify additional risk variants, to improve precision of risk estimation and to enhance the predictive ability of the genetic risk scores across populations [33].
Despite the numerous genetic studies of PCa, the results obtained during investigations are contradictory; they are poorly replicated in studies of different populations and are characterized by significant interethnic differences. The wide variability of clinical manifestations of PCa indicates the need to study both general genetic risk factors for the development of the disease and specific factors predisposing to certain clinical and pathogenetic variants of disease course. It has to be taken under consideration, that immune system has strong impact on the creation of permissive conditions both for the initial cell transformation, as well as for the support of primary tumor growth and metastatic spread. Therefore, genetical aspects have to be considered on the immunological landscape, that is affected by lifestyle, exposure to pathogens, and chronic low grade inflammation driven by metabolic and stress factors. In this regard, the identification of risk factors specific for the development of PCa in different populations is relevant. Identification of PCa risk markers will make it possible to predict the risk of PCa development with high accuracy and to develop preventive measures taking into account the individual genetic characteristic of each patient.
It is necessary to confirm previously identified markers, to exclude false positive findings is an essential step to evaluate GWAS findings. Thus, replication and validation studies are an effective approach that will enable to disprove random findings and possible random associations. Moreover, in order to bring the genetic aspects in the overall pathophysiological context, it is necessary to analyze how potential genetic predisposition affects PCa risks in the immunocompromised patients. However, such analysis is far away from being trivial, since number of factors can be associated with the compromised immune system. For example, studies on PCa in HIV-infected men are complicated by the significantly enhanced incidence of HIV infection in sexual minority groups while such important parameter as sexual orientation is not routinely included in cancer registers [35]. Additional factors is that sexual minorities can experience poorer health-related quality of life outcomes compared to heterosexual men [35].On another side, the incidence of PCa might be decreased in men from sexual minorities living with HIV due to their shorter life expectations.
2.3. GWAS and PCa aggressiveness
The majority of men diagnosed with PCa demonstrate indolent PCa; therefore, detecting the genetic variants that are able to distinguish aggressive PCa from non-aggressive one is of critical clinical importance for the prevention and treatment of the disease.
A few GWAS devoted to discovery of SNPs for prediction of aggressive PCa have been conducted world-wide [36,37,38,39,40,41,42,43,44]. Risk assessment of PCa is an important tool to distinguish between low-risk from high-risk PCa, preventing PCa overtreatment and helps to choose the optimal and provide tailored treatment strategies for each patient. In 2007, Duggan et al. were the first to conduct a GWAS study for aggressive PCa [36]. They performed genome-wide association scan in 498 men with aggressive PCa and 494 control subjects from Sweden. Among 60,000 SNPs they identified seven that had a similar (positive or negative) and statistically significant (P<.01) association with the risk of aggressive PCa in both studies. Analyzing 1032 PCa patients and 571 control subjects of European descent showed that only rs1571801 has maintained a significant association after the validation stage and was associated with aggressive PCa (one-sided P value = 0.004). (OR: 1.36; 95% CI: 1.13–1.63; p = 1.0 × 10-3). It is located in the DAB2IP gene, which encodes a novel Ras GTPase-activating protein and putative prostate tumor suppressor. Interestingly, the association of this SNP was stronger in patients with a Gleason score higher than 8 [36].
Jielin Sun et al., 2009 found the association of rs9623117*C allele at 22q13 with aggressiveness of PCa. The combined allelic test was highly significant, with P = 5.0 x 10(-7). The odds ratio (OR) of allele C for aggressive PCa was estimated to be 1.18 [95% confidence interval (95% CI), 1.11-1.26]. The risk-associated variant was located within the genomic region of TNRC6B, a gene involved in miRNA-mediated mRNA degradation [37].
One of the studies enrolled 4,829 and 12,205 patients with more and less aggressive disease but did not collected control individuals without prostate cancer. They found that the frequency of the TT genotype of SNP rs4054823 at 17p12 was consistently higher among patients with more aggressive compared with less aggressive disease in each of the seven populations studied, (P= 2.1 x 10(-8)) under a recessive model, exceeding the conservative genome-wide significance level. The difference in frequency was largest between patients with high-grade, non-organ-confined disease compared with those with low-grade, organ-confined disease. This study demonstrates that inherited variants predisposing to aggressive but not indolent PCa exist in the genome, and suggests that the clinical potential of such variants as potential early markers for risk of aggressive PCa should be evaluated. The rs4054823 variant is located in an intergenic region, with the closest gene being HS3ST3A1, which encodes a heparan sulfate biosynthetic enzyme, a protein with no known relation to PCa [38]
The investigation of Liesel M FitzGerald et al., 2011 has analyzed 387,384 autosomal single nucleotide polymorphisms (SNPs), and rs6497287 located on 15q13 chromosome was confirmed to be most strongly associated with more aggressive (P(discovery) = 5.20 × 10(-5), P(validation) = 0.004) than less aggressive disease (P = 0.14) [39]. Besides, rs3774315 on 3q26 was found to be associated with PCa risk; however, the association was not stronger for disease that is more aggressive. Study cohorts included 202 PCa cases with aggressive phenotype and 100 randomly sampled, age-matched prostate-specific antigen screened negative controls. Validation testing in an independent set of 527 cases with more aggressive and 595 cases with less aggressive prostate cancer, and 1,167 age-matched controls confirmed the results obtained [39]. Another study enrolled patients with aggressive forms of PCa and biopsy-proven normal controls ascertained from a PCa screening program to GWAS [40]. They found significant associations between aggressive PCa and five single nucleotide polymorphisms (SNPs) in the 10q26 (rs10788165, rs10749408, and rs10788165, p value for association 1.3 × 10-10 to 3.2 × 10-11 and 15q21 (rs4775302 and rs1994198, p values for association 3.1 × 10-8 to 8.2 × 10-9 regions. Replication study proved the association of combinations of these SNPs in 3439 patients undergoing a prostate biopsy, to be associated with aggressive forms of PCa [40].
One of the most powerful investigations was performed by Amin Al Olama et al [42]. They conducted bioinformatical analysis of previously published GWAS and enrolled datasets of 11085 cases, where c patients had an aggressive disease form and 11463 were healthy individuals. The analysis of 2.6 million SNPs revealed rs11672691 to be significantly associated with aggressive PCa (OR=1.12; 95% CI=1.03–1.21). Validation studies comparing aggressive and indolent PCa cases did not confirm the significance of association of rs11672691 with aggressive PCa risk [45]. The risk allele of rs11672691 (intergenic) was associated with an increased risk for PCa-specific mortality and showed rs11672691 to be associated with both fatal and nonfatal PCa [45]. This intergenic variant is located on 19q13 chromosomal region within a long noncoding RNA gene - PCAT19. According to several investigations this SNP may be also involved in the regulation of HOXA2, PCAT19 and CEACAM21 expression, in PCa cell growth, invasion, metastasis and disease progression [46,47].
Ping Gao J. et al described a statistically significant association of rs11672691 with clinical features of aggressive PCa, such as high tumor stage, high prostate-specific antigen (PSA) levels in 2738 men with disease progression and development of castration-resistant PCa (CRPC) [47]. They analyzed the expression of two previously unknown PCa genes, PCAT19 and CEACAM21, and found that aggressive CRPC is associated with the G allele rs11672691, involved in regulation of PCAT19 and CEACAM21 gene expression and affects the cellular properties of the prostate tumor. It was hypothesized that analysis of the rs11672691 genotypes combined with analysis of PCAT19 and CEACAM21 gene expression levels may predict PCa recurrence and patient survival that may be useful in determining further follow-up tactics [42,43,48,49].
The International Consortium for PCa Genetics (ICPCG) performed GWAS of 2511 (unrelated) familial PCa cases and 1382 unaffected controls, including 1394 PCa aggressive cases and 1096 less-aggressive ones, but none of the SNPs (rs2735839, rs11672691, rs11704416, rs35148638 rs78943174) previously associated with aggressive disease were significant in this investigation [42,43,48,49].
Overall, all germinal mutations which have been highlighted by the studies in different geographical cohorts are found in genes related to the biology of cancers cells themselves, providing transformed cells with the enhanced potential to proliferate or migrate. However, such studies did not focus on the bioinformatics search in the screening data for the mutations in genes that make immune system more permissive for the transformed cells to start uncontrolled proliferation and invasion.
3. MiRNA as prostate cancer biomarker
Despite the significant advances in early diagnosis and treatment, the identification of new markers and design of diagnostic panels of molecular markers providing high accuracy and specificity in discrimination between aggressive forms of PCa and indolent forms, and predicting tumor aggressiveness, is still a relevant challenge. New biomarkers with high precision and specificity are needed for early detection of malignant changes and for population screening to identify predisposition to the development of PC, for disease prognosis and for making therapeutic decisions [49]. Currently the main problem with the diagnostic management of PCa is the poor accuracy of currently available tools. Prostate biopsy performed when the serum prostate-specific antigen (PSA) level is above 2.5-4 ng/mL and/or abnormalities are detected during digital rectal examination confirms the diagnosis of PCa in 24-37% of cases [50].
The discovery of a new class of noncoding RNAs- microRNAs (miRNAs) is a promising new direction in the early diagnosis of PCa [52,53]. Each miRNA can modulate the expression of multiple mRNAs and, furthermore, each mRNA may be targeted by several different miRNAs. Most of the physiological processes are controlled by miRNAs. MiRNAs are involved in the regulation of such cellular functions as the maintenance of stemness, proliferation, differentiation, apoptosis, and metabolism [53,54]. About 60% of all human genes are regulated by miRNAs [55,56,57]. Aberrant expression of miRNAs may have a significant influence on some specific features of cell biology, ultimately leading to various pathological phenomena, including cancer [52]. Epigenetic regulatory mechanisms play a key role in cancer development and epigenetic events are one of the earliest in carcinogenesis [36,37].
MicroRNAs (miRNA) are short noncoding single-stranded RNA molecules, that regulate post-transcriptional gene expression by complementary binding to the 30 untranslated region (3’-UTR) of mRNAs, which, in turn, lead to translational suppression or degradation of the mRNA target. MiRNAs play a crucial role in regulating several biological processes needed to preserving homeostasis and immune balance as they carry out a wide modulatory activity on various molecular signaling pathways. Evidence for their crucial role in prostate carcinogenesis and progression, their stability in biological fluids make microRNAs promising candidates as non-invasive or minimally invasive diagnostic and prognostic biomarkers of PCa.
3.1. MicroRNA biogenesis
In mammals, more than 90% of microRNAs are encoded by nucleotide sequences located in introns. For comparison, this value constitutes 14% in fruit flies and invertebrates [57]. MiRNA biogenesis consists of several stages. At the initial stage, miRNA genes are transcribed by RNA polymerase II, resulting in the formation of precursor miRNAs (pre-miR) that contain from several hundred to several thousand nucleotides. The next stage is catalyzed by ribonuclease III (Drosha) and the RNA- binding protein Pasha (DGCR8), resulting in the cleavage of the miRNA precursor and the formation of the primary miR (pre-miR), which usually consists of about 70 nucleotides [58]. The nuclear transporter factor, exportin-5, subsequently binds to the pre-miR and translocates it into the cytoplasm, where another RNAse, Dicer, interacts with the pre-miR, resulting in the formation of mature RNA duplex consisting of about 22 nucleotides [59]. Mature miRNA in the complex with the Argonaut (Ago2) and TRBP family proteins forms RISC (RNA-induced silencing complex). One of the miRNA duplex strands (passenger strand) is degraded, and the functional strand activates RISC complex and interacts with target mRNA, resulting in the degradation of this target mRNA and inhibiting its translation [60]. Imperfect binding leads to poor contact between the microRNA and target mRNA with subsequent inhibition of translation, while a high degree of complementarity promotes strong binding and proteolytic cleavage of target mRNA with the involvement RISC complex. In the past several years, non-canonical biogenesis pathways of miRNAs such as the synthesis pathway continue to be discovered, but most pathways still require Dicer enzymes [61]. The mirtron pathway is the first non-classical pathway discovered. This pathway does not require the Drosha/Dgcr8 complex to generate pre-miRNA, but still requires the transport of XPO-5 and the cleavage of Dicer enzyme [62].
3.2. The role of miRNA in prostate carcinogenesis
The profiles and functions of miRNAs are altered in different cancer types and their nature varies [54]. miRNA can exert a negative effect by altering the expression level by either loss or increase in the affinity of miRNA sequences within or on its target. The miRNA function is largely determined by the relative accessibility of the target mRNA. For these reasons, individual miRNAs may have different effects in tissues, especially in cancers of different cellular origin [57,64].
Porkka et al. conducted the first study on the expression profiles of 319 miRNAs in PCa with oligonucleotide array hybridization and found differential expression of 51 miRNAs between benign tumors and carcinoma tumors was detected, 37 of them showing down-regulation and 14 up-regulation in carcinoma samples [64]. However, not all of the results of Porkka et al. were confirmed by further research. At present, the number of platforms developed for the study of the miRNA expression profiles is rapidly increasing. For the analysis of tumor-specific microRNAs, microarrays are often used. However, this method was based on previously accumulated data on miRNAs. Bioinformatic analysis of the data identifies a pool of miRNAs that influence PCa carcinogenesis (Figure 2) [65]. However, experimental data have confirmed that not all of the mentioned miRNAs impact the development of malignant prostate tumors. The next-generation sequencing (NGS) technology provided a new approach to identify previously unknown miRNAs. The most appropriate method to confirm the miRNA expression profile is real-time quantitative PCR (RT-qPCR)[66].
3.3. Oncogenic miRNAs in prostate cancer
In malignant tumors of various origin, the expression of majority of miRNAs is reduced. This is consistent with the existing view on the association between miRNA expression and differentiation. However, microRNAs with amplified genes may have the functions of oncogenes. A number of these miRNAs were identified, as well as the well-known fact regarding the overexpression of oncogenic miR-181b in prostate cancer. Hu et al. demonstrated that the inhibition of miR-181b induced apoptosis in the PCa cells [67]. Katia et al. suggested that miR-21 negatively regulated RECK, a matrix metalloproteinase regulator, thereby providing the expression of the invasive properties of PCa cells [68]. In addition, miR-21 can promote the development of the aggressive potential of PCa cells through the regulation of other tumor inhibitors, such as MARCKS protein (Myris- toylated alanine-rich protein kinase c substrate), which is the target of miR-21 [69]. There are other tumor suppressor genes negatively regulated by miR-21, including ANP32A and SMARCA4 genes [70]. Since miR-21 is overexpressed in prostate cancer, targeted inhibition of miR-21 restores apoptosis in the cells [69]. It should be also mentioned that the transfection of miR-21 into PCa cells induces resistance to the antitumor drug docetaxel [71]. Altered expression of miRNA-21 was demonstrated in numerous studies. Recent meta-analysis to assess reliability of miRNA-21 as diagnostic biomarker especially in progressivity of PCa showed miRNA-21 as reliable serum diagnostic biomarker candidate for metastatic progressive PCa. The pooled sensitivity and specificity showed 0.91 (95% CI 0.88-0.94, I2=0%) and 0.89 (95% CI 0.85-0.92, I2=44.8%), respectively. Positive and negative likelihood ratio showed 7.18 (95% CI 4.31-11.96, I2=56%) and 0.11 (95% CI 0.07-0.16, I2=11.8%). SROC were assessed and got Area Under Curve around 97.4% [72]. The expression values of miRNA-21 also correlate with the occurrence of resistant castration of PCa and metastases, therefore miRNA-21 is expected to be a biomarker to estimate the progression of cancer. Seputra et al 2021 examined forty-eight total samples obtained from serum and then performed RT-PCR to obtain expression values of miRNA-21 [72]. The cut-off value of miRNA-21 from the PCa category was 33.595-35.21, PPV = 87.5%, NPV = 66.7% with p value = 0.003, while the value of miRNA-21 in the castrate resistant PCa (CRPC) category was> 35.21, PPV = 80%, NPV = 58.3 with a value of p = 0.04. Taking into account the significant differences in the expression values of miRNA-21 between benign prostatic hypertrophy with CRPC and PCa and CRPC, miRNA-21 cut-off point is potential to differentiate the diagnosis [72].
Several studies identified miR-18a-5p as a tumor promoter in prostate cancer. The results of Liang et al 2021 suggest that miR-18a-5p might function as a tumor-promoting factor in PCa and might contribute to its proliferation [73]. They found miR-18a-5p expression to be upregulated in human PCa tissues while SLC40A1 was down-regulated. Cell proliferation assay demonstrated that miR-18a-5p promoted PCa cell proliferation. They also found out that SLC40A1 was downregulated by miR-18a-5p in PCa cell lines and restoration of SLC40A1 reversed the effects of miR-18a-5p in PCa cells [74]. Besides, diagnostic significance of miR-18a was shown among Egyptian patients. MiR-18a was a powerful discriminator of PCa patients from healthy controls as it had the highest AUC (0.966; 95% CI, 0.937-1.000), while miR-221 provided better differentiation of metastatic from localized PC (sensitivity was 92.9% at 100% specificity), and when miR-18a and miR-221were combined for differentiating patients with PC, that would increase the sensitivity to 96.4% at a specificity of 100% (AUC, 0.997; 95% CI, 0.988-1.0) (p < .000) [74].
It is known that the miR-106b-25 cluster is located in intron 13 of the MCM7 gene (Minichromosome maintenance protein 7) [75]. The co-expression of MCM7 and miR-106b-25 mediates prostatic intraepithelial neoplasia in transgenic mice) [75]. Moreover, PTEN expression is negatively regulated by miR-106b-25 [75]. It is interesting that miR-106b-25 regulates the ZBTB4 (zinc finger and BTB domain-containing 4) at the post-transcriptional level. ZBTB4 functions as a tumor suppressor gene and is involved in the inhibition of specific target gene expression by competitive binding with the promoter regions [75]. The miR-106b-25 cluster also negatively regulates caspase-7 [76]. There are conflicting data on the regulation miR-125b via androgen signaling. It is suggested that androgen receptor (AR) inhibits miR-125b to initiate the expression of various mRNA transcripts [77]. In contrast to these data, it was reported in one of the studies that androgen signaling stimulated miR-125b expression, while targeted inhibition of miR-125b led to a decrease in androgen-independent growth [78]. Moreover, miR-125b negatively regulates various proapoptotic genes, including p53, Puma, and Bak1 [79]. It was also demonstrated that microRNA-4534 (miR-4534) is overexpressed in PCa. MiR-4534 was found to be hypermethylated in normal tissues and cell lines compared to PCa tissues/cells. MiR-4534 exerts its oncogenic effects partly by downregulating the tumor suppressor PTEN gene. Knockdown of miR-4534 impaired cell proliferation, migration/invasion and induced G0/G1 cell cycle arrest and apoptosis in PCa. Hannah Nip et al., 2016 revealed that overexpression of miR-4534 induced pro-cancerous characteristics in this non-cancer cell line. Statistical analyses revealed that miR-4534 has potential to independently distinguish malignant from normal tissues and positively correlated with poor overall and PSA recurrence free survival [80]. Information on oncogenic miRNA is summarized in Table 2.
3.4. Tumor suppressor miRNAs in prostate cancer
MiR-15a and miR-16 function as a tumor suppressor by participating in the regulation of the expression of such oncogenes as BCL2, MCL1, CCND1, and WNT3A [75,76]. It was reported that the injection of a new class of miRNA inhibitors, named antagomirs, in normal mouse prostate with the purpose of miR-15a and miR-16 silencing resulted in considerable hyperplasia [77,78,79]and an increase in survival, proliferation, invasion, as well as increased cancer disease severity in the immunodeficient NOD-SCID mice [80]. There is evidence suggesting low expression levels of these microRNAs in various malignancies, including chronic lymphocytic leukemia, pituitary adenoma, and prostate carcinoma. In humans, miR-15a and miR-16 are located in the 13q14.3 region of chromosome 13, which has been reported in the studies to undergo deletions not only in PCa but also in other cancers such as chronic lymphocytic leukemia, pituitary adenoma, mantle cell lymphoma, and prostate cancer. [81]. Zidan et al. showed that the expression of miR-15a and miR-16-1 was considerably decreased in 80% of the examined PCa samples as compared to normal tissues [81].
It is also known that expression of miR-224, miR-16, miR-31, miR-125b, miR-143, miR-145, miR-149, miR-181b, miR-184, miR-205, miR-221, and miR-222 is decreased in PCa [82]. miR-143 and miR-145 play an important role in the biology of PCa and act as tumor suppressors by inhibiting cell growth and activation of apoptosis [83]. MiR-205 is a tumor suppressor, the transfection of miR-205 in PCa cells induces apoptosis. Note that miR-205 is directly involved in stimulating the expression of the IL24 and IL32 tumor suppressor genes [87][84]. This miRNA also participates in down- regulation of Bcl-2. Enhanced apoptosis was observed in PCa cells with restored miR-205 (by means of treatment with cisplatin and doxorubicin) [84]. MiR-574-3p controls Bcl-xL and substantially induces apoptosis in the PCa cells. The expression of miR-574-3p in PCa is decreased[85]
The Polycomb group gene Bmi-1 is overexpressed in PCa stem cells; however, as was demonstrated previously, there was a decrease in Bmi-1 expression in cells treated with NVP-LDE-225 (erismodegib) [86]. The inhibitory effect of erismodegib on Bmi-1 expression is realized through up-regulation of miR-128. The introduction of antagomirs in the cells to knock down miR-128 revealed that erismodegib-induced apoptosis was significantly disrupted [86].
The potential applications of exosomal miRNAs in the treatment and diagnosis of PCa have gained increasing interest. Many studies highlight the potential for EVs to be collected from the blood as marker-containing capsules for both the diagnosis of PC and for making a determination of the patient’s stage and prognosis. Thus, exosomal miRNAs miR-107 and miR-574-3p, were successfully quantified in the urine of men with prostate cancer. Recently, miR-375 and miR-141 were identified as the best markers for high risk PCa [88]. In addition, it was found that miR-141, miR-298, miR-346, and miR-375 were upregulated in serum from prostate patients compared with controls [89,90]. The role of the miR-141 in PCa is further supported by a study that found that it could be a valuable diagnostic tool for patients with this disease [91]. Moreover, exosomal miR-141 could differentiate patients with localized PCa from those with metastatic PCa with higher AUC (0.869) in comparison with AUC of the PSA (0.775). Also, high diagnostic capabilities had the miR-196a-5p and the miR-501-3p with the AUC 0.73 and 0.69, respectively [92]. Perspective non-invasive biomarkers was found by Li et al. in urine samples collected from PCa patients, namely they shown remarkable upregulation of exosomal miR-451a and exosomal miR-486-3p/5p as compared to the healthy controls [93]. In addition, as a predictive biomarker for early detection and castration-resistance PCa was serve exosomal miR-423-3p [94]. It was also demonstrated, that the miR-125a-5p/miR-141-5p ratio was significantly higher in PCa patients compared to HCs. According to the data presented, the miR-125a-5p/miR-141-5p ratio appeared to perform better as an early PCa biomarker than either of the miRNAs taken individually [95]. Fredsoe et al. identiffied different miRs by qPCR in cell-free urine samples from patients with benign prostatic hyperplasia and from those with clinically localized PCa. Furthermore, they developed a new diagnostic model of three miRs (miR-222-3p*miR-24-3p/miR-30c-5p) which distinguished benign prostatic hyperplasia from PCa [78].
Yao et al. found that hsa-miR-182 was the single most upregulated miRNA in PC tissue . Hirata etal. provided evidence that hsa-miR-182 promotes PC by targeting RECK, FOXF2, and MTSS1—tumor suppressor transcripts [96]. Also, it was shown that miR-146a is upregulated in PC cell lines and tissues, oncoMir potential was demonstrated [97]. MiR-15b-5p and miR-106b-5p were confirmed to be significantly upregulated in tissue of aggressive PCa when their level was associated with disease aggressiveness [98]. Furthermore, prognostic score combining the level of miR-15b-5p and miR-106b-5p with serum PSA level discriminated indolent PCa from an aggressive form with even higher analytical parameters [98]. Also, as a result of the comparison of tissue and peripheral blood mononuclear cells samples, it was idenntified that down regulated hsa-miR-494-3p, hsa-miR-3128, hsa-miR-8084 were common miRNAs [96]. Three (HIF1A, NHS,INSL4) targets identified for hsa-miR-494-3p, two (HIF1A, AVRP1A) for hsa-miR-3128 and three (AVRP1A, NHS, INSL4) for hsa-miR-8084. These results suggested that hsa-miR-494-3p, hsa-miR-3128, hsa-miR-8084 and their targets may play a crucial role in therapeutic and early diagnostic strategies for PCa [96].
4. Pathogenesis and staging
PCa is the result of a complicated synergistic action of accumulated genetic changes aimed at boosting cell proliferation compared to cell death. Detection and recognition of these events is essential for disease control in the earliest stages of transformation, for developmental progression to an invasive tumor, for prognosis and determining points of therapeutic intervention. There is epidemiological evidence that indicates a potential association between symptomatic prostatitis and the risk of developing prostate cancer. [100,101,102]. Data from 746,176 patients over 50 years of age diagnosed with PCa from 2010 to 2013 were analysed, with follow-up to 2019 using the Korean National Health Insurance Service patient database [102]. Control groups were matched for age, disease (diabetes, hypertension), and Charlson comorbidity index. The incidence of PCa was statistically significantly higher in the prostatitis group (1.8% vs. 0.6%, p<0.001). The HR for patients with PCa was significantly higher in patients with prostatitis (HR 2.99; 95% CI 2.89-3.09, p<0.001), with HR for PCa being significantly higher in acute prostatitis than in chronic prostatitis (3.82; 95% CI 3.58-4.08; p<0.001; HR 2.77; 95% CI 2.67-2.87, p<0.001)[102].
The exact mechanisms underlying the association between prostatitis and PCa risk are not fully understood. However, several factors have been proposed, where inflammation plays a prominent role. Prostatitis is characterized by inflammation of the prostate gland. Chronic inflammation can lead to DNA damage, and alterations in cell signaling pathways [103]. Inflammatory processes in the prostate can result in the release of various cytokines and growth factors. These molecules can promote cell proliferation, angiogenesis (formation of new blood vessels), and tissue remodeling, which are all processes involved in tumor development and progression [104]. The immune response triggered by prostatitis may involve the recruitment of immune cells, such as macrophages and T-cells, into the prostate gland. Although these immune cells are intended to fight infection, their prolonged presence may contribute to chronic inflammation and create conditions favorable for evading immune surveillance [104]. It is important to note that while the association between prostatitis and PCa risk has been observed in epidemiological studies, not all individuals with prostatitis will develop prostate cancer. Further research is needed to better understand the complex mechanisms underlying these correlations and to identify potential therapeutic targets for intervention.
As mentioned above, uncontrolled activation of signaling pathways (which can be caused by inflammation), activated growth factors, in particular signaling through the lipid kinase group phosphoinositide 3-kinase (PI3K), contributes to carcinogenesis. [105]. As part of the activation mechanism, the catalytic subunits of PI3K, directly bind to small GTPases, which appear to be members of the RAS subgroup. The role of PTEN in this pathway is to catalyze the opposite reaction by metabolizing PI3K [105]. Eventually the triggering of signaling pathways leads to genomic instability, increased cell proliferation, cell invasion and migration (Figure 3).
In the majority of patients localized PCa is multifocal, due to the fact that cancer most often occurs in the peripheral zone of the prostate, the surrounding pseudocapsule is involved in 80% of clinically detected cases of cancer [107,108]. In the following stages, the cancer cells invade the seminal vesicles, paraprostatic tissue, which is regarded by the international TNM classification as T3 or in the urinary bladder, levator muscles, external sphincter and/or anterior abdominal wall - T4 stage [108]. The T1 stage of PCa is typically assigned when the tumor is confined to the prostate gland and is clinically non-palpable and non-visualized. This means that the tumor was detected either during a prostate biopsy performed due to elevated PSA levels or incidentally during a transurethral resection. In other cases where the tumor does not extend beyond the prostate, it is classified as stage T2 [108]. Lymphatic metastasis involves the hypogastric, obturator, external iliac, presacral, common iliac, or retroperitoneal lymph nodes. Depending on the spread of the tumor to regional or distant lymph nodes, the diagnosis is N1 or N2, respectively. When PCa spreads through the hematogenous route, it most often involves the bones of the axial skeleton, and less often the lungs, liver, and other soft tissues, which is assessed as stage M1 [109,110].
Risk assessment of PCa at diagnosis and after treatment is based on the grading system, PSA level, tumor-node-metastasis classification (TNM), and/or previous treatment history in order to predict potential fatal outcomes of PCa and justify treatment decisions. Some patients with intermediate-risk PCa and every high-risk PCa patient should undergo additional imaging studies [87,88,89].
Signs of chronic inflammation are a common finding in malignant prostate tissues. By histological examination, inflammatory infiltrates are defined mainly by CD3+ T lymphocytes, CD19 or CD20 B lymphocytes (10-15%) and macrophages (15%) [110]. It is likely that the damage associated with the inflammatory response and subsequent chronic tissue healing can lead to proliferative inflammatory atrophy (PIA).
Currently, there exists inadequate evidence to definitively establish a direct causal connection between prostate inflammation and prostate cancer. Ongoing investigations aim to explore a potential association between these two conditions; however, additional dependable and precise evidence is necessary to firmly establish this association. For instance, it has been suggested that the absence of glutathione S-transferase P1 (GSTP1) might be accountable for the progression from prostatic inflammation to high-grade intraepithelial neoplasia (HGPIN) and prostate cancer (PCa) in individuals with a genetic predisposition [110]. Conducting further research will enable us to gain a better understanding of these mechanisms and their impact on the development of cancer.
Chronic inflammation preceding PCa onset or cooperating with early stages of PCa development can be an essential source for new biomarkers that can be used in combination with PCs or other relevant cancer cell-derived factors. Such biomarkers can be released by immune cells in TME, or can be identified in circulating immune cells, in particular in monocytes. We have previously demonstrated that monocytes provide clinically valuable information for patients with breast and colorectal cancer [1,2,3]. In breast cancer CD163+ CD14lowCD16+ and CD163+ CD14+CD16+ monocytes were indicative for the presence of the malignancy, and CD14lowCD16+HLA-DR+monocytes were predictive for the good response to neoadjuvant chemotherapy [111]. In colorectal cancer (CRC), the monocyte biomarkers showed distinct correlations with metastatic processes [112]. Increased levels of CCR2+ monocytes in rectal cancer was indicative for the absence of both lymphatic and hematogenous metastasis. In contrast, in patients with colon cancer CD163+ monocytes positively correlated with lymphatic [112]. In this study we also made full transcriptome profiling in circulating monocytes by NGS, and identified PFKFB3, activator of glycolysis that is currently an attractive candidate for several solid cancers [112]. PFKFB3+ monocyte-derived macrophages massively infiltrated tumor in colon, and Nanostring spatial profiling identified correlation of PFKFB3 with tumor-promoting properties of TAMs in colon but not in rectal cancer. Monocytes-expressed PFKFB3 was indicative for tumor relapse specifically in colon but not rectal cancer [112]. Monocytes are not only indicators of the systemic immune status, but can be also programmed by systemic changes caused by all types of anti-cancer therapy: surgery, radiotherapy, chemotherapy and immunotherapy [113]. In cardio-metabolic disorders, monocytes are crucial biomarkers predicting progression of vascular complications related to the pathological lipid metabolism, and key role of monocyte-derived foamy macrophages is established [114,115,116].Considering recently identified by us and other foamy macrophages in PC, the potential of circulating monocytes as predictors of therapy efficiency and biomarkers that can be used for the personification of therapeutic schemas is only emerging but has solid theoretic background. For PCa the clinical value of monocytes programming and subpopulations urgently needs to be identified.
5. Methods for diagnosing prostate cancer.
5.1. Currently used diagnostic approaches
Clinical guidelines from various countries and leading cancer societies recommend similar algorithms for the diagnosis of PCa [108,109,117]:
- digital rectal examination (DRE)
- and/or transrectal ultrasound (TRUS)
- serum PSA (prostate-specific antigen): total PSA and the ratio of free PSA to total PSA.
- biopsy confirmation.
Table 3 summarizes the information about the sensitivity (the ability of the test to correctly identify patients with the disease) and specificity (the test's ability to correctly identify patients without the disease) of the PCa tests available for clinical appellation.
Tumor biopsy testing for somatic gene mutations (e.g., BRCA1, BRCA2, ATM, PALB2, FANCA, RAD51D, and CHEK2), MSI, or dMMR should be considered as additional diagnostic options for patients with regional or distant metastases [108]. Table 3 summarizes data on the sensitivity and specificity of each type of diagnostic method, and their advantages and disadvantages. Each type of test has limitations and can miss a case of prostate cancer; only the combination of several tests provides the most complete picture. The maximal sensitivity of a single text is not exceeding 77%, and the maximal specificity of a single test is below 50%. Therefore, it is necessary to improve the test systems or their combination to prove precise clinical decision.
5.2. Necessity of early screening
Early PCa can be asymptomatic, so screening is necessary. The gold standard for screening now are PSA testing and Digital Rectal Examination [109,110]. Since the introduction of the PSA testing and subsequent biopsies, the incidence of PCa in the USA doubled, beginning in the late 1980s [113,114]. The UK interview study used EPIC-26 questionnaire to analyze 3 523 patients 18-42 months after they were diagnosed for prostate cancer. This study revealed that quality of life is higher in patients diagnosed with PSA-screened PCa compared to patients diagnosed due to symptoms of PCa [120].PCa was detected through PSA testing in 31.3% of cases, while it presented with symptoms in 59.7% of cases. In a multivariate analysis, men with symptoms reported more severe problems with urinary incontinence (Adjusted mean ratio (AMR): 0.96, 95% CI: 0.96-0.97), bladder irritation (AMR: 0.95, 95% CI: 0.95-0.96), bowel function (see AMR: 0.97, 95% CI: 0.97-0.98), sexual function (AMR: 0.90, 95% CI: 0.88-0.92), and vital/hormonal function (AMR: 0.96, 95% CI: 0.96-0.96 compared to men whose PCa was found via the PSA test. These differences were consistent among respondents of different ages, stages, Gleason scales, and treatment types [120].
Early detection of PCa improves overall patient survival in low-risk patients, but has no effect on the survival of patients with metastatic cancer [115,116,117]. Since the management strategy for low-risk patients is an active monitoring, careful patient selection and accuracy in risk assessment require new more precise and more specific biomarkers, since randomized controlled trials have shown that PSA screening has no effect on overall mortality or prostate cancer–caused mortality [123]. A meta-analysis of five randomized controlled trials involving 721,718 men found that screening probably leads to a small reduction in specific mortality over 10 years, but has no effect on overall mortality [124]. It has no effect on all-cause mortality (IRR 0.99, 95% CI 0.98-1.01) and may have no effect on prostate-specific mortality (IRR 0.96, 0.85-1.08;), or a small effect on prostate-specific mortality (IRR 0.79, 0.69-0.91). Using Der Simonian and Laird’s inverse of variance random effects modeling , it was estimated that for every 1,000 men screened, about 0,1% will be hospitalized because of biopsy complications, 0,3% will have reduced quality of life because of bladder control problems, and 2,5% will have erectile dysfunction [124]. In addition to the side effects caused by screening procedures, it is necessary to consider the patient's psychological discomfort due to false positive PSA tests.
There is also evidence of the benefits of measuring baseline PSA levels in men in middle age to determine the subsequent risk of PCa [125]. Analysing 13 years of data from a cohort of 10,968 men between the ages of 55 and 60 included in the PCa screening group showed baseline prostate-specific antigen levels were associated with any future prostate cancer. The risk was significantly lower among men with baseline prostate-specific antigen levels less than 2.00 ng/mL [126].
From the standpoint of reducing mortality, overtreatment, which refers to unnecessary or excessive medical interventions, offers little or no benefit. Many years after treatment with radical prostatectomy or radiation therapy for high-risk prostate cancer, a significant proportion of men still have significant deterioration in one or many functional areas: sexual, urinary, and intestinal function [7,121,122,123]. Modeling of the lifelong consequences of annual PSA screening at the age of 55-69 years compared with the absence of screening estimated the loss of 23% of the years of life obtained during screening, mainly due to deterioration in the quality of life due to long-term side effects from treatment [125].
The study covered a cohort of 182,160 men aged 55 to 69 from eight European countries calculated the number of men that have to be invited for screening to prevent one death from PCa [129]. They used the Wald method to calculate confidence intervals for the difference in the risk of death.The result of this calculation demonstrated that prevention of one death prom PC requires screening of PSA levels for 9 years in 1947 men, for 13 years in 742 men, and for 16 years in 570 men [129].
5.3. Biopsy examination and how TAMs can help
Prostate biopsy is a medical procedure used to diagnose prostate cancer. It involves the removal of a small tissue sample from the prostate gland, which is then examined under a microscope for the presence of cancer cells. To assess the degree of differentiation prognostically, pathologists evaluate PCa biopsies. When examining biopsy samples, an assessment is made of the shape, size of cells, nuclear changes, and the presence of anomalies in cellular structure [130]. Additionally, the degree of differentiation of cancer cells is evaluated. Furthermore, the quantity and distribution of cancer cells in the sample, as well as the presence of invasion into surrounding tissues, are assessed. This helps to determine the stage of PCa and its potential for spread or metastasis [130].
The histological evaluation of PCa samples also includes a description of specific pathological features such as the presence of glandular structures, cribriform patterns, perineural invasion, and others. To unify the examination of samples, Donald Gleason developed a scale for evaluating the architectural details of malignant glands under at low and medium zoom [131]. According to the Gleason scale, microscopic estimation of histopathological features is used to classify cancerous tissue from low-differentiated (highest grade) to highly differentiated (lowest grade). In 2014, the classification system was reorganized into groups 1-5 according to the International Society of Urological Pathology (ISUP) classification [132]. The Gleason classification of acinar adenocarcinoma of the prostate is one of the earliest and most successful applications of evidence-based medicine in routine clinical practice. The original Gleason system has shown excellent correlation with clinical outcomes [128,129].
The field of PCa diagnosis and treatment has witnessed significant advancements in recent years, with the development of several diagnostic tests based on new biomarkers. These tests aim to enhance the specificity and sensitivity of prostate-specific antigen (PSA) screening, while also improving treatment efficiency. In light of these advancements, it becomes increasingly crucial to consider the incorporation of newly identified biomarkers to further enhance diagnostic accuracy and optimize patient management.
Among the emerging biomarkers, the prostate health index (PHI), TMPRSS2-ERG fusion gene, 4K test, and PCA3 test have shown promising potential in PCa diagnosis [135]. The 4K test, for instance, utilizes a panel comprising total PSA (tPSA), free PSA (fPSA), intact PSA (iPSA), and human kallikrein 2 (hK2). By analyzing these biomarkers, it becomes possible to differentiate between various causes of elevated PSA values, offering valuable insights into the presence and characteristics of PCa [136]. Unbound PSA is known as free PSA (fPSA), which is about 5%-35% of total PSA. The investigation of different forms of PSA, such as free PSA (fPSA) and intact PSA (iPSA), has provided valuable information for the identification and stratification of prostate cancer. While fPSA represents the unbound fraction of PSA, accounting for approximately 5% to 35% of total PSA, iPSA demonstrates a higher ratio in men with cancer compared to those without tumors [117]. Furthermore, human kallikrein 2 (hK2), a closely related protein to PSA, has exhibited structural and functional similarities, offering additional insights into PCa detection[138]. These biomarkers will help to identify high-grade PCa in men who have not previously been tested for elevated PSA. The PHI (prostate health index) defined by the formula [([-2]proPSA / free PSA) × √total PSA] was developed for prognostic purposes [117,119]. The PHI, compared to other PSA related tests, has been shown to better differentiate PCa from benign prostatic hyperplasia ,and thus to prevent unnecessary prostate biopsy [133,135]. The PHI test allows to detect the progression of PCa with active monitoring of the condition. Additional non-invasive biomarkers used for diagnostic purposes are TMPRSS2-ERG Fusion and PCA3 Progensa PCa Antigen 3 based on urine [135]. One of the markers of castrate resistance in PCa is the level of TMPRSS2-ERG. Serine protease 2 (TMPRSS2), as well as the ERG gene, can be found in 50% of patients with prostate cancer, overexpression of the PCA3 or DD3 gene (specific non-coding mRNA) is observed in more than 95% of cases of primary PCa [141]. However, the assessment of the above markers in comparison with the prostate specific antigen has a lower sensitivity. The accuracy of PCa biomarkers is critical in determining the further tactics - the transition to invasive methods of examination [141].
As can be seen from the above, the accuracy of PCa biomarkers is critical in determining the further tactics - the transition to invasive methods of examination. Deciding the necessary of screening, it is needed to take into account and individually discuss the strategy with transgender patients taking or not taking hormone therapy, since the risk of PCa for them has not been determined [137,138]. However, the biopsy examination in the current clinical practice does not consider evaluation of any immunological parameter in tumor microenvironment. Tumor-associated macrophages massively infiltrate solid tumors, and in several cancers, including prostate cancer, bad prognosis correlates with increased amounts of TAMs, identified in majority of studies by highly specific macrophage marker CD68, that belongs to scavenger receptor family [144].
However, there are also cancer types or specific intertumoral compartments, where levels of CD68+ TAMs in parenchima in breast cancer have a negative correlations with lymphatic metastasis, and in colorectal cancer total CD68 levels positively correlate with overall survival and good prognosis [144,145,146]. Therefore number of studies are focusing o the specific TAM biomarkers, there scavenger/endocytic receptors for example CD206, stabilin-1, MARCO and others) demonstrated specific correlations with stages, metastasis and even therapy responses for lung, breast, colorectal, ovarian, prostate and other types of cancer [144]. However, scavenger receptors (SRs) are limited in the demonstrating of specific TAM functional polarization to high redundancy of SRs’ ligand repertoire and functions [147]. More specific biomarkers with clear distinct functions are needed to distinguish between tumor-promoting and anti-tumor TAMs. Chitinase-like proteins, potent regulators of angiogenesis and immune cell recruitment can offer here much more precise biomarker value [148,149]. Metabolic regulators, as we identified for colorectal cancer, is also of a great potential [112]. In case of prostate cancer, we have observed even on the morphological level 2 major phenotypes of TAMs: regular in size and shape, and large foamy-types TAMs [150]. Identification of the molecular profile and functions of these 2 major phenotypes of TAMs is needed to decipher their biomarker potential.
The limited integration of these new biomarkers in routine clinical practice can be attributed to several factors. Further validation and standardization across different populations and settings, cost-effectiveness, and accessibility challenges, as well as interpretational complexities, hinder their widespread use. The reliance on PSA as the primary screening tool and the need for a shift in established practices also contribute to their limited utilization. Continued research, standardization efforts, and evidence-based guidelines are crucial for facilitating their integration into PCa care.
5.4. State-of-the art imaging for staging and metastasis detection
A multicenter, paired-cohort, confirmatory study was conducted to test the diagnostic accuracy of MP-MRI and TRU-biopsy [159]. A total of 740 men with prostate-specific antigen concentrations up to 15 ng/mL, without prior biopsy, who underwent MRI followed by TRU biopsy were included in the study. The performance and reporting of each test was blinded to the results of the other tests. Clinically significant cancer was defined as a Gleason score ≥4 + 3. For clinically significant cancer, MP-MRI was more sensitive (93%, 95% CI 88-96%) than TRUSI biopsy (48%, 42-55%; p<0-0001) and less specific (41%, 36-46% for MP-MRI versus 96%, 94-98% for TRUS biopsy; p<0-0001). 44 (5-9%) of 740 patients reported serious adverse events, including 8 cases of sepsis [159].
Modern imaging techniques (CT, MRI, PET) can accurately determine the presence of metastases and the stage of development of PCa. MRI with high accuracy can reduce the number of unnecessary biopsies compare with standard TRUS. PI-RADS was introduced by the European Society of Urogenital Radiology to standardize the results of MRI. Key elements of assessment include prostate volume measurement, lesion mapping, lesion measurement, and lesion assessment [160].
However, the high cost and low availability of these methods is an obstacle to accurate diagnosis (Table 2)
5.5. Raman spectroscopy
One of the advanced optical diagnostic methods is the Raman spectroscopic analysis, the use of which is acceptable in the study of tissues and / or biofluids for in vitro and in vivo diagnostics. Raman imaging provides real-time results, contributing a molecular portrait of tissue in malignant diseases such as prostate cancer. The basic principle of Raman spectroscopy is associated with the interaction of light with the molecules of substances contained in the analyzed samples [161]. Raman spectroscopy has been positively evaluated as a diagnostic method for differential analysis of malignant and non-malignant tumors of the prostate tissue. Aubertin et al. conducted a study of 32 fresh prostatectomy tissue samples with Raman spectroscopy to detect and analyze the severity of PCa [162]: from samples of fresh prostate tissue, sections were obtained, from each of which 20 to 50 scattering spectra were obtained, the total number of which was 947 spectra. Using Raman spectroscopy, it was found that the sensitivity in identifying the prostate among non-prostatic tissue was 82%, the specificity was 83%. During the analysis, it was also noted that the method made it possible to distinguish benign prostate tissue from malignant tissue with a sensitivity of 87% and a specificity of 86% [162]. Pinto et al incorporated Raman spectroscopy for in vivo (n = 4) and ex vivo (based on 599 spectra from 20 prostatectomy specimens) into the operational process of robotic radical prostatectomy to differentiate between cancerous and benign prostate tissue [163]. A study by Medipalli et al. compared the Raman spectra of plasma from 43 patients diagnosed with PCa and 33 healthy volunteers. As a result, it was found that in the plasma of patients with prostate cancer, during the interpretation of Raman spectra, an increase in the bands associated with nucleic acids was revealed [164]. In interpreting the results, it is worth noting that the increase in nucleic acid concentration may be associated with abnormally increased gene expression, which correlates with increased release of nucleic acids due to cell death. In patients diagnosed with prostate cancer, compared with healthy people, absorption bands corresponding to lipids are more common. This may be due to the fact that the risks of tumor progression and treatment resistance are associated with high levels of lipids and cholesterol in the tumor [165,166].
Raman spectroscopy is a minimally invasive analysis technique that can be used to analyze samples of urine, blood, saliva, and other body fluids. Del Mistro et al. conducted an experimental surface-enhanced Raman scattering (SERS) analysis of urine samples from 9 patients diagnosed with PCa and 9 healthy volunteers. Using machine learning algorithms, the obtained spectra were divided into two groups (PCa or healthy people), resulting in a sensitivity of the analysis of 100%, a specificity of 89% [167]. Similarly, Ma et al. used SERS to obtain urine spectra of 75 patients, of which 12 patients with recurrent PCa and 63 patients with non-recurrent PCa [168]. While comparing Raman bands in patients without recurrence and with recurrent prostate cancer, it was found that the latter have enhanced Raman bands associated with lipids, proteins, amino acids and DNA [168]. Review by Chen et al. demonstrates the most complete information about Raman scattering methods based on urine [169] . Serum measurement of prostate specific antigen (PSA) protein is the most common method for identifying men at risk of developing prostate cancer, but the PSA test has certain limitations due to type I errors [153]. False elevated PSA can be observed with prostatitis, benign prostatic hyperplasia, urinary tract infections [154]. In men, PSA levels in the 4–10 ng/mL range represent a significant diagnostic challenge, Chen et al. investigated the use of serum SERS as a screening method to differentiate between PCa and BPH in patients with above-specified PSA levels [170].
Raman imaging techniques allow specific identification of individually pure substances, often without the need for contrast agents. This represents great promise for in vivo applications in disease diagnosis, treatment outcome monitoring, disease prognosis, and disease staging. By developing special nanoparticles in the SERS process, it is possible to achieve high sensitivity and specificity for diagnostic purposes in various biological fluids.
6. Conclusions
In conclusion, the current limitations of clinical and pathological parameters in accurately differentiating prostate cancer types present a significant obstacle in preventing unnecessary treatments. The application of modern molecular genetic techniques has provided an opportunity to gather comprehensive data encompassing genomic, transcriptomic, epigenomic, proteomic, and metabolomic profiles from various sources such as biopsies, prostatectomies, and individual cells. The integration of these data into clinical practice is of utmost importance [7]. However, each tumor develops in the close contact with the immune system, both systemic and local, where the immune system can be 1) pre-programmed to be more cancer permissive or restrictive; 2) programmed by growing tumor; 3) programmed by cancer therapy. Such programming can be be on epigenetic, transcriptional and metabolic levels, and each level can offer clinically applicable [111,171]. Thus, the immune biomarkers, in particular biomarkers provided by innate immunity(circulating monocytes and TAMs), is essential to significantly enhance the precision of the diagnostics and to achieve maximal therapeutic efficacy.
By harnessing this wealth of information, we can gain a better understanding of the disease's variability and tumor progression, thereby identifying valuable biomarkers for effectively managing newly diagnosed cases and tailoring treatment strategies to individual patients. This approach holds great promise in optimizing prostate cancer care and ensuring that patients receive the most appropriate and personalized treatments.
Funding
The study was funded by the state grant of the Ministry of Science and Higher Education of the Russian Federation “Genetic and epigenetic editing of tumor cells and the microenvironment to block metastasis” No. 075-15-2021-1073 and BSMU Strategic Academic Leadership Program PRIORITY-2030
Conflicts of Interest
The authors declare no conflict of interest
Abbreviations
21-kDa protein
BAD Bcl-2 agonist of cell death
Bcl-2 - family of regulator proteins that regulate cell death (apoptosis), by either inhibiting (anti-apoptotic) or inducing (pro-apoptotic) apoptosis
Bcl-xL It is a member of the Bcl-2 family of proteins, and acts as an anti-apoptotic protein.
Bmi-1 Polycomb complex protein BMI-1
cAMP responsive element binding protein (CREB)
CASP9 - caspase 9
CBP CREB-binding protein
CDKN1B cyclin dependent kinase inhibitor 1B
CDK2 - cyclin dependent kinase 2
ERK extracellular signal-regulated kinase
FKHR - a subfamily of "forkhead" proteins that regulate transcription.
FOXA1 s a transcription factor (TF) of the Forkhead box (FOX) protein family.
GFR - growth factor receptors
GF - growth factor
Grb2 Growth Factor Receptor bound 2
GSK-3 Glycogen synthase kinase 3
GSTP1 - glutathione S-transferase pi 1
HIV - human immunodeficiency virus
HCs-96
HOX subset of homeobox genes, are a group of related genes that specify regions of the body plan of an embryo along the head-tail axis
HRas proto-oncogene Harvey-RAS
IL1R2 - interleukin 1 receptor type 2
IKK IκB kinase
IκB [I-kappa-B], a protein complex
MSI microsatellite instability
MDM2 proto-oncogene encodes a nuclear-localized E3 ubiquitin ligase.
MEK Mitogen-activated protein kinase - MAPK/ERK (MAPK/ERK kinase или MEK MAPK mitogen-activated protein kinase ERK extracellular signal-regulated kinase
MMP Matrix metalloproteinases
mTOR mammalian target of rapamycin
NF-kB Nuclear Factor Kappa B
NKX3-1 NK3 homeobox 1
NBS1 - is a protein which in humans is encoded by the NBN gene
p21 wildtype activating factor-1/cyclin-dependent kinase inhibitory protein-1 or WAF1/CIP1
P27 - tumor suppressor, inhibit all types of cyclin-dependent kinase (cyclin dependent kinase CDK)
P53 protein regulates the repair of cellular DNA.
PDK1 - Phosphoinositide-dependent kinase-1
PI3K phosphatidylinositol-3-kinase
PIP3 - phosphatidylinositol-3,4,5-triphosphate
pkb/akt - protein kinase B
PLAU plasminogen activator, urokinase
PLAT (tissue plasminogen activator)
PSA prostate specific antigen
RNase L (HPC1) interferon (IFN)-induced ribonuclease which, upon activation, destroys all RNA within the cell (both cellular and viral)
Rb - Retinoblastoma protein
SRD5a2 - Steroid 5-α-reductase type 2
SPINT1 - serine peptidase inhibitor, Kunitz type 1
TCF/LEF The TCF/LEF family (T cell factor/lymphoid enhancer factor family)
TMPRSS2 - Transmembrane serine protease 2
TMPRSS2-ERG - fusion of transmembrane serine protease 2 (TMPRSS2) genes with ETS transcription factor
TF Tissue factor
TRBP transactivation response element RNA-binding protein
References
- F. Bray, J. Ferlay, I. Soerjomataram, R. L. Siegel, L. A. Torre, и A. Jemal, «Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries», CA. Cancer J. Clin., 68, pp. 394–424, . 2018. [CrossRef]
- T. I. A. for R. on Cancer (IARC), «Global Cancer Observatory». https://gco.iarc.fr/ (accessed 9 march 2023 ).
- T. R. Rebbeck, «Prostate Cancer Genetics: Variation by Race, Ethnicity, and Geography», Semin. Radiat. Oncol., 27, 1, pp. 3–10, 2017. [CrossRef]
- A. J. Vickers, A. Elfiky, V. L. Freeman, и M. Roach, «Race, Biology, Disparities, and Prostate Cancer», Eur. Urol., 81, 5, pp. 463–465, 2022. [CrossRef]
- T. M. Ma et all, «Comparison of Response to Definitive Radiotherapy for Localized Prostate Cancer in Black and White Men: A Meta-analysis», JAMA Netw. Open, 4, 12, с. e2139769, 2021. [CrossRef]
- M. Koscuiszka et all, «Impact of race on survival in patients with clinically nonmetastatic prostate cancer who deferred primary treatment: Race and Survival in PCa Patients», Cancer, 118, 12, pp. 3145–3152, 2012. [CrossRef]
- G. Wang, D. Zhao, D. J. Spring, и R. A. DePinho, «Genetics and biology of prostate cancer», Genes Dev., 32, 17–18, pp. 1105–1140, 2018. [CrossRef]
- J. L. Jahn, E. L. Giovannucci, и M. J. Stampfer, «The high prevalence of undiagnosed prostate cancer at autopsy: implications for epidemiology and treatment of prostate cancer in the Prostate-specific Antigen-era: High prostate cancer prevalence: Research implications in the PSA-ERA», Int. J. Cancer, 137, 12, pp. 2795–2802, 2015. [CrossRef]
- S. Wasim, S.-Y. Lee, и J. Kim, «Complexities of Prostate Cancer», Int. J. Mol. Sci., 23, 22, с. 14257, 2022. [CrossRef]
- B. Turanli et all, «Drug Repositioning for Effective Prostate Cancer Treatment», Front. Physiol., 9, с. 500, 2018. [CrossRef]
- H. Han et all, «Characteristics of BRCA2 Mutated Prostate Cancer at Presentation», Int. J. Mol. Sci., 23, 21, с. 13426, 2022. [CrossRef]
- M. D. Bardis et all, «Applications of Artificial Intelligence to Prostate Multiparametric MRI (mpMRI): Current and Emerging Trends», Cancers, 12, 5, с. 1204, 2020. [CrossRef]
- J. M. Santos-Pereira и A. Aguilera, «R loops: new modulators of genome dynamics and function», Nat. Rev. Genet., 16, 10, pp. 583–597, 2015. [CrossRef]
- A. Abeshouse et all, «The Molecular Taxonomy of Primary Prostate Cancer», Cell, 163, 4, pp. 1011–1025, 2015. [CrossRef]
- C. Kumar-Sinha, S. Kalyana-Sundaram, и A. M. Chinnaiyan, «Landscape of gene fusions in epithelial cancers: seq and ye shall find», Genome Med., 7, 1, с. 129, 2015. [CrossRef]
- L. F. van Dessel et all, «The genomic landscape of metastatic castration-resistant prostate cancers reveals multiple distinct genotypes with potential clinical impact», Nat. Commun., 10, 1, с. 5251, 2019. [CrossRef]
- D. Tong, «Unravelling the molecular mechanisms of prostate cancer evolution from genotype to phenotype», Crit. Rev. Oncol. Hematol., 163, с. 103370, 2021. [CrossRef]
- M. Adamaki и V. Zoumpourlis, «Prostate Cancer Biomarkers: From diagnosis to prognosis and precision-guided therapeutics», Pharmacol. Ther., 228, с. 107932, 2021. [CrossRef]
- D.-P. Kong et all, «Prevalence and clinical application of TMPRSS2-ERG fusion in Asian prostate cancer patients: a large-sample study in Chinese people and a systematic review», Asian J. Androl., 22, 2, с. 200, 2020. [CrossRef]
- J. St. John, K. Powell, и M. K. C. -LaComb, «TMPRSS2-ERG Fusion Gene Expression in Prostate Tumor Cells and Its Clinical and Biological Significance in Prostate Cancer Progression», J. Cancer Sci. Ther., 04, 04, 2012. [CrossRef]
- U. Testa, G. Castelli, и E. Pelosi, «Cellular and Molecular Mechanisms Underlying Prostate Cancer Development: Therapeutic Implications», Medicines, 6, 3, с. 82, 2019. [CrossRef]
- S. Stelloo et all, «Integrative epigenetic taxonomy of primary prostate cancer», Nat. Commun., 9, 1, с. 4900, 2018. [CrossRef]
- I. Dudka et all, «Comprehensive metabolomics analysis of prostate cancer tissue in relation to tumor aggressiveness and TMPRSS2-ERG fusion status», BMC Cancer, 20, 1, с. 437, 2020. [CrossRef]
- H. Koide et all, «Comparison of ERG and SPINK1 expression among incidental and metastatic prostate cancer in Japanese men», The Prostate, 79, 1, pp. 3–8, 2019. [CrossRef]
- R. D. Webster, J. L. Ross, и B. K. Arun, «The changing landscape of hereditary cancer genetic testing: Hereditary Cancer Genetic Testing», Cancer, 124, 4, pp. 664–666, 2018. [CrossRef]
- E. Dathathri et all, «Liquid Biopsy Based Circulating Biomarkers in Metastatic Prostate Cancer», Front. Oncol., 12, с. 863472, 2022. [CrossRef]
- P. Tian, M. Zhong, и G.-H. Wei, «Mechanistic insights into genetic susceptibility to prostate cancer», Cancer Lett., 522, pp. 155–163, 2021. [CrossRef]
- The COGS–Cancer Research UK GWAS–ELLIPSE (part of GAME-ON) Initiative et all, «Identification of 23 new prostate cancer susceptibility loci using the iCOGS custom genotyping array», Nat. Genet., 45, 4, pp. 385–391, 2013. [CrossRef]
- S. Benafif, Z. Kote-Jarai, и R. A. Eeles, «A Review of Prostate Cancer Genome-Wide Association Studies (GWAS)», Cancer Epidemiol. Biomarkers Prev., 27, 8, pp. 845–857, 2018. [CrossRef]
- The Profile Study et all, «Association analyses of more than 140,000 men identify 63 new prostate cancer susceptibility loci», Nat. Genet., 50, 7, pp. 928–936, 2018. [CrossRef]
- The UKGPCS Collaborators et all, «Frequent germline deleterious mutations in DNA repair genes in familial prostate cancer cases are associated with advanced disease», Br. J. Cancer, 110, 6, pp. 1663–1672, 2014. [CrossRef]
- M. Wang et all, «Large-scale association analysis in Asians identifies new susceptibility loci for prostate cancer», Nat. Commun., 6, 1, с. 8469, 2015. [CrossRef]
- D. V. Conti et all, «Trans-ancestry genome-wide association meta-analysis of prostate cancer identifies new susceptibility loci and informs genetic risk prediction», Nat. Genet., 53, 1, pp. 65–75, 2021. [CrossRef]
- C. Sipeky, T. L. J. Tammela, A. Auvinen, и J. Schleutker, «Novel prostate cancer susceptibility gene SP6 predisposes patients to aggressive disease», Prostate Cancer Prostatic Dis., 24, 4, pp. 1158–1166, 2021. [CrossRef]
- C. Amarasekera, V. Wong, E. Yura, A. Manjunath, E. Schaeffer, и S. Kundu, «Prostate cancer in sexual minorities and the influence of HIV status», Nat. Rev. Urol., 16, 7, pp. 404–421, 2019. [CrossRef]
- D. Duggan et all, «Two Genome-wide Association Studies of Aggressive Prostate Cancer Implicate Putative Prostate Tumor Suppressor Gene DAB2IP», JNCI J. Natl. Cancer Inst., 99, 24, pp. 1836–1844, 2007. [CrossRef]
- J. Sun et all, «Sequence Variants at 22q13 Are Associated with Prostate Cancer Risk», Cancer Res., 69, 1, pp. 10–15, 2009. [CrossRef]
- J. Xu et all, «Inherited genetic variant predisposes to aggressive but not indolent prostate cancer», Proc. Natl. Acad. Sci., 107, 5, pp. 2136–2140, 2010. [CrossRef]
- L. M. FitzGerald et all, «Genome-wide Association Study Identifies a Genetic Variant Associated with Risk for More Aggressive Prostate Cancer», Cancer Epidemiol. Biomarkers Prev., 20, 6, pp. 1196–1203, 2011. [CrossRef]
- R. K. Nam et all, «New variants at 10q26 and 15q21 are associated with aggressive prostate cancer in a genome-wide association study from a prostate biopsy screening cohort», Cancer Biol. Ther., 12, 11, pp. 997–1004, 2011. [CrossRef]
- F. R. Schumacher et all, «Genome-wide association study identifies new prostate cancer susceptibility loci», Hum. Mol. Genet., 20, 19, pp. 3867–3875, 2011. [CrossRef]
- A. Amin Al Olama et all, «A meta-analysis of genome-wide association studies to identify prostate cancer susceptibility loci associated with aggressive and non-aggressive disease», Hum. Mol. Genet., 22, 2, pp. 408–415, 2013. [CrossRef]
- S. I. Berndt et all, «Two susceptibility loci identified for prostate cancer aggressiveness», Nat. Commun., 6, 1, с. 6889, 2015. [CrossRef]
- C. C. Teerlink et all, «Genome-wide association of familial prostate cancer cases identifies evidence for a rare segregating haplotype at 8q24.21», Hum. Genet., 135, 8, pp. 923–938, 2016. [CrossRef]
- I. M. Shui et all, «Prostate cancer (PCa) risk variants and risk of fatal PCa in the National Cancer Institute Breast and Prostate Cancer Cohort Consortium», Eur. Urol., 65, 6, pp. 1069–1075, 2014. [CrossRef]
- J.-H. Xia и G.-H. Wei, «Oncogenic regulatory circuits driven by 19q13 rs11672691 underlies prostate cancer aggressiveness», Mol. Cell. Oncol., 5, 6, с. e1516451, 2018. [CrossRef]
- P. Gao et all, «Biology and Clinical Implications of the 19q13 Aggressive Prostate Cancer Susceptibility Locus», Cell, 174, 3, pp. 576-589.e18, 2018. [CrossRef]
- B. T. Helfand et all, «Associations of prostate cancer risk variants with disease aggressiveness: results of the NCI-SPORE Genetics Working Group analysis of 18,343 cases», Hum. Genet., 134, 4, pp. 439–450, 2015. [CrossRef]
- D. Vanacore et all, «Micrornas in prostate cancer: an overview», Oncotarget, 8, 30, pp. 50240–50251, 2017. [CrossRef]
- G. Cochetti, G. Poli, G. Guelfi, A. Boni, M. G. Egidi, и E. Mearini, «Different levels of serum microRNAs in prostate cancer and benign prostatic hyperplasia: evaluation of potential diagnostic and prognostic role», OncoTargets Ther., Volume 9, pp. 7545–7553, 2016. [CrossRef]
- H. S. Zhou, T. Zhao, X. M. Rao, и A. L. Beaudet, «Production of helper-dependent adenovirus vector relies on helper virus structure and complementing», J. Gene Med., 4, 5, pp. 498–509, 2002. [CrossRef]
- K. Kasomva et all, «Roles of microRNA in prostate cancer cell metabolism», Int. J. Biochem. Cell Biol., 102, pp. 109–116, 2018. [CrossRef]
- N. Sharma и M. M. Baruah, «The microRNA signatures: aberrantly expressed miRNAs in prostate cancer», Clin. Transl. Oncol., 21, 2, pp. 126–144, 2019. [CrossRef]
- M. Hill и N. Tran, «miRNA interplay: mechanisms and consequences in cancer», Dis. Model. Mech., 14, 4, с. dmm047662, 2021. [CrossRef]
- S. Khan, H. Ayub, T. Khan, и F. Wahid, «MicroRNA biogenesis, gene silencing mechanisms and role in breast, ovarian and prostate cancer», Biochimie, 167, pp. 12–24, 2019. [CrossRef]
- C. Massillo, G. N. Dalton, P. L. Farré, P. De Luca, и A. De Siervi, «Implications of microRNA dysregulation in the development of prostate cancer», Reproduction, 154, 4, pp. R81–R97, 2017. [CrossRef]
- D. Trümbach и N. Prakash, «The conserved miR-8/miR-200 microRNA family and their role in invertebrate and vertebrate neurogenesis», Cell Tissue Res., 359, 1, pp. 161–177, 2015. [CrossRef]
- M. Majidinia et all, «MicroRNAs, DNA damage response and ageing», Biogerontology, 21, 3, pp. 275–291, 2020. [CrossRef]
- Rawat et all, «MicroRNA in Pancreatic Cancer: From Biology to Therapeutic Potential», Genes, 10, 10, с. 752, 2019. [CrossRef]
- T. Takahashi et all, «LGP2 virus sensor regulates gene expression network mediated by TRBP-bound microRNAs», Nucleic Acids Res., 46, 17, pp. 9134–9147, 2018. [CrossRef]
- Y.-K. Kim, B. Kim, и V. N. Kim, «Re-evaluation of the roles of DROSHA , Exportin 5 , and DICER in microRNA biogenesis», Proc. Natl. Acad. Sci., 113, 13, 2016. [CrossRef]
- C. Stavast и S. Erkeland, «The Non-Canonical Aspects of MicroRNAs: Many Roads to Gene Regulation», Cells, 8, 11, с. 1465, 2019. [CrossRef]
- A. Khan, E. Ahmed, N. Elareer, K. Junejo, M. Steinhoff, и S. Uddin, «Role of miRNA-Regulated Cancer Stem Cells in the Pathogenesis of Human Malignancies», Cells, 8, 8, с. 840, 2019. [CrossRef]
- K. P. Porkka, M. J. Pfeiffer, K. K. Waltering, R. L. Vessella, T. L. J. Tammela, и T. Visakorpi, «MicroRNA Expression Profiling in Prostate Cancer», Cancer Res., 67, 13, pp. 6130–6135, 2007. [CrossRef]
- «KEGG PATHWAY: MicroRNAs in cancer - Reference pathway». https://www.kegg.jp/pathway/map=map05206&keyword=prostate%20cancer (accessed 13 march 2023 ).
- W. Wittling и M. Pflüger, «Neuroendocrine hemisphere asymmetries: Salivary cortisol secretion during lateralized viewing of emotion-related and neutral films», Brain Cogn., 14, 2, pp. 243–265, 1990. [CrossRef]
- A. R. Rhodes et all, «The malignant potential of small congenital nevocellular nevi», J. Am. Acad. Dermatol., 6, 2, pp. 230–241, 1982. [CrossRef]
- K. R. M. Leite et all, «Controlling RECK miR21 Promotes Tumor Cell Invasion and Is Related to Biochemical Recurrence in Prostate Cancer», J. Cancer, 6, 3, pp. 292–301, 2015. [CrossRef]
- T. Li, D. Li, J. Sha, P. Sun, и Y. Huang, «MicroRNA-21 directly targets MARCKS and promotes apoptosis resistance and invasion in prostate cancer cells», Biochem. Biophys. Res. Commun., 383, 3, pp. 280–285, 2009. [CrossRef]
- K. Schramedei et all, «MicroRNA-21 targets tumor suppressor genes ANP32A and SMARCA4», Oncogene, 30, 26, pp. 2975–2985, 2011. [CrossRef]
- M. Rizzo, «Mechanisms of docetaxel resistance in prostate cancer: The key role played by miRNAs», Biochim. Biophys. Acta BBA - Rev. Cancer, 1875, 1, с. 188481, 2021. [CrossRef]
- K. Seputra, B. Purnomo, H. Susianti, H. Kalim, и A. Purnomo, «miRNA-21 as Reliable Serum Diagnostic Biomarker Candidate for Metastatic Progressive Prostate Cancer: Meta-analysis Approach», Med. Arch., 75, 5, с. 347, 2021. [CrossRef]
- K. Seputra, B. Purnomo, H. Susianti, H. Kalim, и A. Purnomo, «miRNA-21 Serum Evaluation in BPH, Hormone Sensitive Prostate Cancer, and Castrate Resistant Prostate Cancer: Attempt for Diagnostic Biomarker Evaluation», Acta Inform. Medica, 29, 4, с. 266, 2021. [CrossRef]
- N. H. Ibrahim, M. S. Abdellateif, S. H. Kassem, M. A. Abd El Salam, и M. M. El Gammal, «Diagnostic significance of miR-21, miR-141, miR-18a and miR-221 as novel biomarkers in prostate cancer among Egyptian patients», Andrologia, 51, 10, 2019. [CrossRef]
- D. Mehlich, F. Garbicz, и P. K. Włodarski, «The emerging roles of the polycistronic miR-106b∼25 cluster in cancer – A comprehensive review», Biomed. Pharmacother., 107, pp. 1183–1195, 2018. [CrossRef]
- R. S. Hudson et all, «MicroRNA-106b-25 cluster expression is associated with early disease recurrence and targets caspase-7 and focal adhesion in human prostate cancer», Oncogene, 32, 35, pp. 4139–4147, 2013. [CrossRef]
- S. Rezaei, M. Mahjoubin Tehran, A. Sahebkar, A. Jalili, и S. H. Aghaee-Bakhtiari, «Androgen receptor-related micro RNAs in prostate cancer and their role in antiandrogen drug resistance», J. Cell. Physiol., 235, 4, pp. 3222–3234, 2020. [CrossRef]
- J. Fredsøe et all, «Diagnostic and Prognostic MicroRNA Biomarkers for Prostate Cancer in Cell-free Urine», Eur. Urol. Focus, 4, 6, pp. 825–833, 2018. [CrossRef]
- A. A. Svoronos, S. G. Campbell, и D. M. Engelman, «MicroRNA function can be reversed by altering target gene expression levels», iScience, 24, 10, с. 103208, 2021. [CrossRef]
- H. Nip et all, «Oncogenic microRNA-4534 regulates PTEN pathway in prostate cancer», Oncotarget, 7, 42, pp. 68371–68384, 2016. [CrossRef]
- H. E. Zidan, R. S. Abdul-Maksoud, W. S. H. Elsayed, и E. A. M. Desoky, «Diagnostic and prognostic value of serum miR-15a and miR-16-1 expression among egyptian patients with prostate cancer: SERUM MIR-15A AND MIR-16-1 IN PROSTATE CANCER», IUBMB Life, 70, 5, pp. 437–444, 2018. [CrossRef]
- G. Deng et all, «Targeting androgen receptor (AR) with antiandrogen Enzalutamide increases prostate cancer cell invasion yet decreases bladder cancer cell invasion via differentially altering the AR/circRNA-ARC1/miR-125b-2-3p or miR-4736/PPARγ/MMP-9 signals», Cell Death Differ., 28, 7, pp. 2145–2159, 2021. [CrossRef]
- H. Liang, L. Studach, R. L. Hullinger, J. Xie, и O. M. Andrisani, «Down-regulation of RE-1 silencing transcription factor (REST) in advanced prostate cancer by hypoxia-induced miR-106b~25», Exp. Cell Res., 320, 2, pp. 188–199, 2014. [CrossRef]
- S. J. Oh-Hohenhorst и T. Lange, «Role of Metastasis-Related microRNAs in Prostate Cancer Progression and Treatment», Cancers, 13, 17, с. 4492, 2021. [CrossRef]
- X. Yang et all, «Down-Regulation of mir-221 and mir-222 Restrain Prostate Cancer Cell Proliferation and Migration That Is Partly Mediated by Activation of SIRT1», PLoS ONE, 9, 6, с. e98833, 2014. [CrossRef]
- Z.-H. Zuo et all, «Oncogenic Activity of miR-650 in Prostate Cancer Is Mediated by Suppression of CSR1 Expression», Am. J. Pathol., 185, 7, pp. 1991–1999, 2015. [CrossRef]
- R. J. Bryant et all, «Changes in circulating microRNA levels associated with prostate cancer», Br. J. Cancer, 106, 4, pp. 768–774, 2012. [CrossRef]
- J. C. Brase et all, «Circulating miRNAs are correlated with tumor progression in prostate cancer», Int. J. Cancer, 128, 3, pp. 608–616, 2011. [CrossRef]
- L. A. Selth et all, «Discovery of circulating microRNAs associated with human prostate cancer using a mouse model of disease», Int. J. Cancer, 131, 3, pp. 652–661, 2012. [CrossRef]
- P. Porzycki, E. Ciszkowicz, M. Semik, и M. Tyrka, «Combination of three miRNA (miR-141, miR-21, and miR-375) as potential diagnostic tool for prostate cancer recognition», Int. Urol. Nephrol., 50, 9, pp. 1619–1626, 2018. [CrossRef]
- Z. Li et all, «Exosomal microRNA-141 is upregulated in the serum of prostate cancer patients», OncoTargets Ther., 9, pp. 139–148, 2016. [CrossRef]
- M. Rodríguez et all, «Identification of non-invasive miRNAs biomarkers for prostate cancer by deep sequencing analysis of urinary exosomes», Mol. Cancer, 16, 1, с. 156, 2017. [CrossRef]
- Z. Li, L.-X. Li, Y.-J. Diao, J. Wang, Y. Ye, и X.-K. Hao, «Identification of Urinary Exosomal miRNAs for the Non-Invasive Diagnosis of Prostate Cancer», Cancer Manag. Res., 13, pp. 25–35, 2021. [CrossRef]
- T. Guo et all, «The Identification of Plasma Exosomal miR-423-3p as a Potential Predictive Biomarker for Prostate Cancer Castration-Resistance Development by Plasma Exosomal miRNA Sequencing», Front. Cell Dev. Biol., 8, с. 602493, 2020. [CrossRef]
- C. Alix-Panabières, «Circulating Tumor Cells: Finding Rare Events for a Huge Knowledge of Cancer Dissemination», Cells, 9, 3, с. 661, 2020. [CrossRef]
- J. Chisholm, S. Haas-Neill, P. Margetts, и K. Al-Nedawi, «Characterization of proteins, mRNAs, and miRNAs of circulating extracellular vesicles from prostate cancer patients compared to healthy subjects», Front. Oncol., 12, с. 895555, 2022. [CrossRef]
- M. Worthington et all, «A Study to Investigate the Role of Noncoding RNA miR146 Alpha as a Potential Biomarker in Prostate Cancer», J. Anal. Oncol., 11, pp. 21–23, 2022. [CrossRef]
- J. Juracek et all, «A tissue miRNA expression pattern is associated with disease aggressiveness of localized prostate cancer», The Prostate, 83, 4, pp. 340–351, 2023. [CrossRef]
- L. K. Dennis, C. F. Lynch, и J. C. Torner, «Epidemiologic association between prostatitis and prostate cancer», Urology, 60, 1, pp. 78–83, 2002. [CrossRef]
- G. S. Palapattu et all, «Prostate carcinogenesis and inflammation: emerging insights», Carcinogenesis, 26, 7, pp. 1170–1181, 2005. [CrossRef]
- R. O. Roberts, E. J. Bergstralh, S. E. Bass, M. M. Lieber, и S. J. Jacobsen, «Prostatitis as a Risk Factor for Prostate Cancer»:, Epidemiology, 15, 1, pp. 93–99, 2004. [CrossRef]
- G. Jung, J. K. Kim, H. Kim, J. Lee, и S. K. Hong, «The association between prostatitis and risk of prostate cancer: a National Health Insurance Database study», World J. Urol., 40, 11, pp. 2781–2787, 2022. [CrossRef]
- M. Murata, «Inflammation and cancer», Environ. Health Prev. Med., 23, с. 50, 2018. [CrossRef]
- «Chronic prostatitis/chronic pelvic pain syndrome and prostate cancer: study of immune cells and cytokines - Liu - 2020 - Fundamental & Clinical Pharmacology - Wiley Online Library». https://onlinelibrary.wiley.com/doi/10.1111/fcp.12517 (accessed 19 march 2023 ).
- H. M. Wise, M. A. Hermida, и N. R. Leslie, «Prostate cancer, PI3K, PTEN and prognosis», Clin. Sci., 131, 3, pp. 197–210, 2017. [CrossRef]
- W. G. Nelson, A. M. De Marzo, и W. B. Isaacs, «Prostate Cancer», N. Engl. J. Med., 349, 4, pp. 366–381, 2003. [CrossRef]
- V. T. DeVita, T. S. Lawrence, и S. A. Rosenberg, Ред., DeVita, Hellman, and Rosenberg’s cancer: principles & practice of oncology, 11th edition. Philadelphia: Wolters Kluwer, 2019.
- J. L. Mohler et all, «Prostate Cancer, Version 2.2019, NCCN Clinical Practice Guidelines in Oncology», J. Natl. Compr. Canc. Netw., 17, 5, pp. 479–505, 2019. [CrossRef]
- C. Parker et all, «Prostate cancer: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up», Ann. Oncol., 31, 9, pp. 1119–1134, 2020. [CrossRef]
- C. De Nunzio et all, «The Controversial Relationship Between Benign Prostatic Hyperplasia and Prostate Cancer: The Role of Inflammation», Eur. Urol., 60, 1, pp. 106–117, 2011. [CrossRef]
- M. Patysheva et all, «Effect of Early-Stage Human Breast Carcinoma on Monocyte Programming», Front. Oncol., 11, с. 800235, 2022. [CrossRef]
- I. Larionova et all, «PFKFB3 overexpression in monocytes of patients with colon but not rectal cancer programs pro-tumor macrophages and is indicative for higher risk of tumor relapse», Front. Immunol., 13, с. 1080501, 2023. [CrossRef]
- M. Patysheva et all, «Monocyte programming by cancer therapy», Front. Immunol., 13, с. 994319, 2022. [CrossRef]
- J. Kzhyshkowska, C. Neyen, и S. Gordon, «Role of macrophage scavenger receptors in atherosclerosis», Immunobiology, 217, 5, pp. 492–502, 2012. [CrossRef]
- S. Mosig et all, «Different functions of monocyte subsets in familial hypercholesterolemia: potential function of CD14 + CD16 + monocytes in detoxification of oxidized LDL», FASEB J., 23, 3, pp. 866–874, 2009. [CrossRef]
- L. Willemsen и M. P. Winther, «Macrophage subsets in atherosclerosis as defined by single-cell technologies», J. Pathol., 250, 5, pp. 705–714, 2020. [CrossRef]
- Д. А. Нoсoв et all, «Практические рекoмендации пo лечению рака предстательнoй железы», Malig. Tumours, 12, 3s2-1, pp. 607–626, 2022. [CrossRef]
- H. G. Welch и P. C. Albertsen, «Reconsidering Prostate Cancer Mortality — The Future of PSA Screening», N. Engl. J. Med., 382, 16, pp. 1557–1563, 2020. [CrossRef]
- P. Rawla, «Epidemiology of Prostate Cancer», World J. Oncol., 10, 2, pp. 63–89, 2019. [CrossRef]
- D. W. Donnelly et all, «Quality of life among symptomatic compared to PSA-detected prostate cancer survivors - results from a UK wide patient-reported outcomes study», BMC Cancer, 19, 1, с. 947, 2019. [CrossRef]
- P. C. Sotomayor et all, «Active Surveillance in Prostate Cancer: Current and Potentially Emerging Biomarkers for Patient Selection Criteria», Urol. Int., 106, 12, pp. 1201–1213, 2022. [CrossRef]
- J. Andersson et all, «Digital Rectal Examination in Stockholm3 Biomarker-based Prostate Cancer Screening», Eur. Urol. Open Sci., 44, pp. 69–75, 2022. [CrossRef]
- M. Bilal, A. Javaid, F. Amjad, T. A. Youssif, и S. Afzal, «An overview of prostate cancer (PCa) diagnosis: Potential role of miRNAs», Transl. Oncol., 26, с. 101542, 2022. [CrossRef]
- D. Ilic et all, «Prostate cancer screening with prostate-specific antigen (PSA) test: a systematic review and meta-analysis», BMJ, с. k3519, 2018. [CrossRef]
- S. V. Carlsson и A. J. Vickers, «Screening for Prostate Cancer», Med. Clin. North Am., 104, 6, pp. 1051–1062, 2020. [CrossRef]
- F. H. Schröder et all, «Screening and Prostate-Cancer Mortality in a Randomized European Study», N. Engl. J. Med., 360, 13, pp. 1320–1328, 2009. [CrossRef]
- F. H. Schröder et all, «Prostate-Cancer Mortality at 11 Years of Follow-up», N. Engl. J. Med., 366, 11, pp. 981–990, 2012. [CrossRef]
- G. L. Andriole et all, «Mortality Results from a Randomized Prostate-Cancer Screening Trial», N. Engl. J. Med., 360, 13, pp. 1310–1319, 2009. [CrossRef]
- «A 16-yr Follow-up of the European Randomized study of Screening for Prostate Cancer». [CrossRef]
- M. Noureldin et all, «Review article: MRI-targeted biopsies for prostate cancer diagnosis and management», World J. Urol., 39, 1, pp. 57–63, 2021. [CrossRef]
- T. B. J. Murray, «The Pathogenesis of Prostate Cancer», в Prostate Cancer, Urology Department, Frimley Park Hospital, Portsmouth Rd, Frimley, Camberley GU16 7UJ, UK, S. R. Bott, и K. Lim Ng, Ред., Exon Publications, 2021, pp. 29–42. [CrossRef]
- G. J. L. H. Van Leenders et all, «The 2019 International Society of Urological Pathology (ISUP) Consensus Conference on Grading of Prostatic Carcinoma», Am. J. Surg. Pathol., 44, 8, pp. e87–e99, 2020. [CrossRef]
- G. Sauter et all, «Clinical Utility of Quantitative Gleason Grading in Prostate Biopsies and Prostatectomy Specimens», Eur. Urol., 69, 4, pp. 592–598, 2016. [CrossRef]
- J. I. Epstein et all, «The 2019 Genitourinary Pathology Society (GUPS) White Paper on Contemporary Grading of Prostate Cancer», Arch. Pathol. Lab. Med., 145, 4, pp. 461–493, 2021. [CrossRef]
- A. V. Alford, J. M. Brito, K. K. Yadav, S. S. Yadav, A. K. Tewari, и J. Renzulli, «The Use of Biomarkers in Prostate Cancer Screening and Treatment», Rev. Urol., 19, 4, pp. 221–234, 2017. [CrossRef]
- Y. Naya и K. Okihara, «Role of Complexed PSA in the Early Detection of Prostate Cancer», J. Natl. Compr. Canc. Netw., 2, 3, pp. 209–212, 2004. [CrossRef]
- A. christensson et all, «Serum Prostate Specific Antigen Complexed to α 1-Antichymotrypsin as an Indicator of Prostate Cancer», J. Urol., 150, 1, pp. 100–105, 1993. [CrossRef]
- M. F. Darson et all, «Human glandular kallikrein 2 (hK2) expression in prostatic intraepithelial neoplasia and adenocarcinoma: A novel prostate cancer marker», Urology, 49, 6, pp. 857–862, 1997. [CrossRef]
- Y.-H. Fan et all, «Prostate Health Index outperforms other PSA derivatives in predicting a positive biopsy in men with tPSA <10 ng/mL: Largest prospective cohort in Taiwan», J. Chin. Med. Assoc., 82, 10, pp. 772–777, 2019. [CrossRef]
- P. Nurmikko, K. Pettersson, T. Piironen, J. Hugosson, и H. Lilja, «Discrimination of Prostate Cancer from Benign Disease by Plasma Measurement of Intact, Free Prostate-specific Antigen Lacking an Internal Cleavage Site at Lys145-Lys146», Clin. Chem., 47, 8, pp. 1415–1423, 2001. [CrossRef]
- «Modern biomarkers in prostate cancer diagnosis», Cent. Eur. J. Urol., 73, 2020. [CrossRef]
- N. A. Deebel, J. P. Morin, R. Autorino, R. Vince, B. Grob, и L. J. Hampton, «Prostate Cancer in Transgender Women: Incidence, Etiopathogenesis, and Management Challenges», Urology, 110, pp. 166–171, 2017. [CrossRef]
- M. D. Ingham, R. J. Lee, D. MacDermed, и A. F. Olumi, «Prostate cancer in transgender women», Urol. Oncol. Semin. Orig. Investig., 36, 12, pp. 518–525, 2018. [CrossRef]
- I. Larionova et all, «Tumor-Associated Macrophages in Human Breast, Colorectal, Lung, Ovarian and Prostate Cancers», Front. Oncol., 10, с. 566511, 2020. [CrossRef]
- I. Mitrofanova et all, «Tumor-associated macrophages in human breast cancer parenchyma negatively correlate with lymphatic metastasis after neoadjuvant chemotherapy», Immunobiology, 222, 1, pp. 101–109, 2017. [CrossRef]
- T. Cavalleri et all, «Tumor-associated macrophages and risk of recurrence in stage III colorectal cancer», J. Pathol. Clin. Res., 8, 4, pp. 307–312, 2022. [CrossRef]
- E. Kazakova, P. Iamshchikov, I. Larionova, и J. Kzhyshkowska, «Macrophage scavenger receptors: Tumor support and tumor inhibition», Front. Oncol., 12, с. 1096897, 2023. [CrossRef]
- J. Kzhyshkowska, S. Yin, T. Liu, V. Riabov, и I. Mitrofanova, «Role of chitinase-like proteins in cancer», Biol. Chem., 397, 3, pp. 231–247, 2016. [CrossRef]
- I. Larionova, E. Kazakova, T. Gerashchenko, и J. Kzhyshkowska, «New Angiogenic Regulators Produced by TAMs: Perspective for Targeting Tumor Angiogenesis», Cancers, 13, 13, с. 3253, 2021. [CrossRef]
- K. V. Danilko et all, «Morphological heterogeneity of intratumoral macrophages in prostate tumors», Sib. J. Oncol., 21, 6, pp. 81–90, 2022. [CrossRef]
- J.-L. Descotes, «Diagnosis of prostate cancer», Asian J. Urol., 6, 2, pp. 129–136, 2019. [CrossRef]
- L. Naji et all, «Digital Rectal Examination for Prostate Cancer Screening in Primary Care: A Systematic Review and Meta-Analysis», Ann. Fam. Med., 16, 2, pp. 149–154, 2018. [CrossRef]
- A. M. D. Wolf et all, «American Cancer Society Guideline for the Early Detection of Prostate Cancer: Update 2010», CA. Cancer J. Clin., 60, 2, pp. 70–98, 2010. [CrossRef]
- K. A. O. Tikkinen et all, «Prostate cancer screening with prostate-specific antigen (PSA) test: a clinical practice guideline», BMJ, с. k3581, 2018. [CrossRef]
- M. Shiva, C. Wei, H. Molana, и G. Nabi, «Cost-Effectiveness of Prostate Cancer Detection in Biopsy-Naïve Men: Ultrasound Shear Wave Elastography vs. Multiparametric Diagnostic Magnetic Resonance Imaging», Healthcare, 10, 2, с. 254, 2022. [CrossRef]
- O. Derin, L. Fonseca, R. Sanchez-Salas, и M. J. Roberts, «Infectious complications of prostate biopsy: winning battles but not war», World J. Urol., 38, 11, pp. 2743–2753, 2020. [CrossRef]
- F.-J. H. Drost et all, «Prostate MRI, with or without MRI-targeted biopsy, and systematic biopsy for detecting prostate cancer», Cochrane Database Syst. Rev., 2019, 4, 2019. [CrossRef]
- J. S. Quon, B. Moosavi, M. Khanna, T. A. Flood, C. S. Lim, и N. Schieda, «False positive and false negative diagnoses of prostate cancer at multi-parametric prostate MRI in active surveillance», Insights Imaging, 6, 4, pp. 449–463, 2015. [CrossRef]
- H. U. Ahmed et all, «Diagnostic accuracy of multi-parametric MRI and TRUS biopsy in prostate cancer (PROMIS): a paired validating confirmatory study», The Lancet, 389, 10071, pp. 815–822, 2017. [CrossRef]
- A. O’Shea и M. Harisinghani, «PI-RADS: multiparametric MRI in prostate cancer», Magn. Reson. Mater. Phys. Biol. Med., 35, 4, pp. 523–532, 2022. [CrossRef]
- V. V. Rafalsky, A. Yu. Zyubin, E. M. Moiseeva, и I. G. Samusev, «Prospects for Raman spectroscopy in cardiology», Cardiovasc. Ther. Prev., 19, 1, pp. 70–77, 2020. [CrossRef]
- K. Aubertin et all, «Mesoscopic characterization of prostate cancer using Raman spectroscopy: potential for diagnostics and therapeutics», BJU Int., 122, 2, pp. 326–336, 2018. [CrossRef]
- M. Pinto et all, «Integration of a Raman spectroscopy system to a robotic-assisted surgical system for real-time tissue characterization during radical prostatectomy procedures», J. Biomed. Opt., 24, 02, с. 1, 2019. [CrossRef]
- D. K. R. Medipally et all, «Vibrational spectroscopy of liquid biopsies for prostate cancer diagnosis», Ther. Adv. Med. Oncol., 12, с. 175883592091849, 2020. [CrossRef]
- R. Patel et all, «Sprouty2 loss-induced IL 6 drives castration-resistant prostate cancer through scavenger receptor B1», EMBO Mol. Med., 10, 4, 2018. [CrossRef]
- S. Yue et all, «Cholesteryl Ester Accumulation Induced by PTEN Loss and PI3K/AKT Activation Underlies Human Prostate Cancer Aggressiveness», Cell Metab., 19, 3, pp. 393–406, 2014. [CrossRef]
- G. Del Mistro et all, «Surface-enhanced Raman spectroscopy of urine for prostate cancer detection: a preliminary study», Anal. Bioanal. Chem., 407, 12, pp. 3271–3275, 2015. [CrossRef]
- Y. Ma, J. Chi, Z. Zheng, A. Attygalle, I. Y. Kim, и H. Du, «Therapeutic prognosis of prostate cancer using surface-enhanced Raman scattering of patient urine and multivariate statistical analysis», J. Biophotonics, 14, 1, 2021. [CrossRef]
- S. Chen et all, «Raman Spectroscopy Reveals Abnormal Changes in the Urine Composition of Prostate Cancer: An Application of an Intelligent Diagnostic Model with a Deep Learning Algorithm», Adv. Intell. Syst., 3, 4, с. 2000090, 2021. [CrossRef]
- N. Chen et all, «Surface-enhanced Raman spectroscopy of serum accurately detects prostate cancer in patients with prostate-specific antigen levels of 4–10 ng/mL», Int. J. Nanomedicine, Volume 12, pp. 5399–5407, 2017. [CrossRef]
- I. Larionova, E. Kazakova, M. Patysheva, и J. Kzhyshkowska, «Transcriptional, Epigenetic and Metabolic Programming of Tumor-Associated Macrophages», Cancers, 12, 6, с. 1411, 2020. [CrossRef]
Figure 1.
The location of mutations in DNA repair genes detected by sequencing DNA samples from men with a family history of prostate cancer.
Figure 1.
The location of mutations in DNA repair genes detected by sequencing DNA samples from men with a family history of prostate cancer.
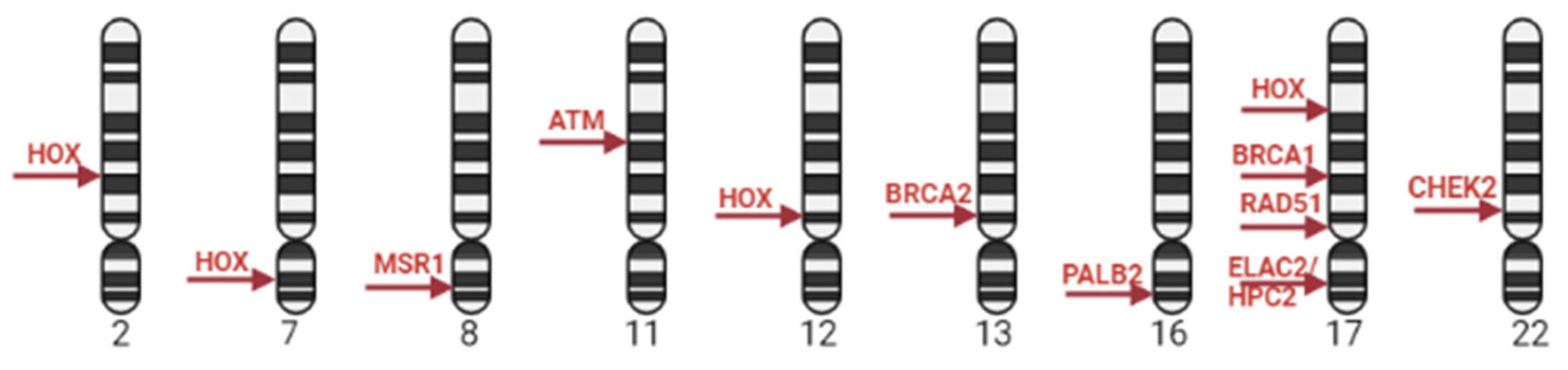
Figure 2.
The up-regulated and down-regulated miRNA, and their role in the pathogenesis of prostate cancer.
Figure 2.
The up-regulated and down-regulated miRNA, and their role in the pathogenesis of prostate cancer.
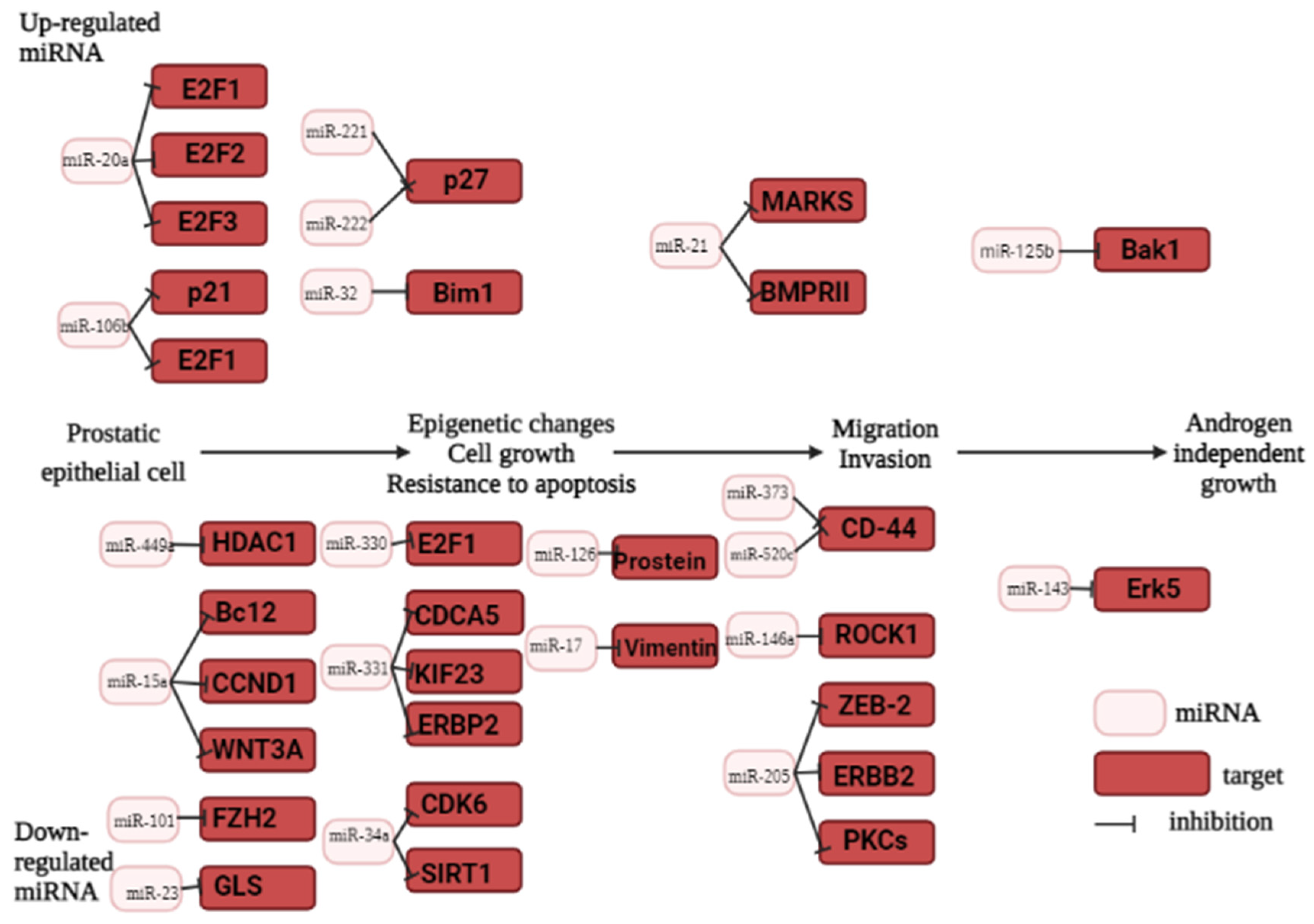
Figure 3.
Prostate cancer pathways [65]. This is a schematic presentation of prostate cancer pathogenesis, driving factors, and connections between genes, signalig pathways. Key events in carcionogenesi include DNA hypermethylation, triggering the MARK signaling pathway, which leads to cell proliferation and tumor growth. Open question are marked in red.
Figure 3.
Prostate cancer pathways [65]. This is a schematic presentation of prostate cancer pathogenesis, driving factors, and connections between genes, signalig pathways. Key events in carcionogenesi include DNA hypermethylation, triggering the MARK signaling pathway, which leads to cell proliferation and tumor growth. Open question are marked in red.
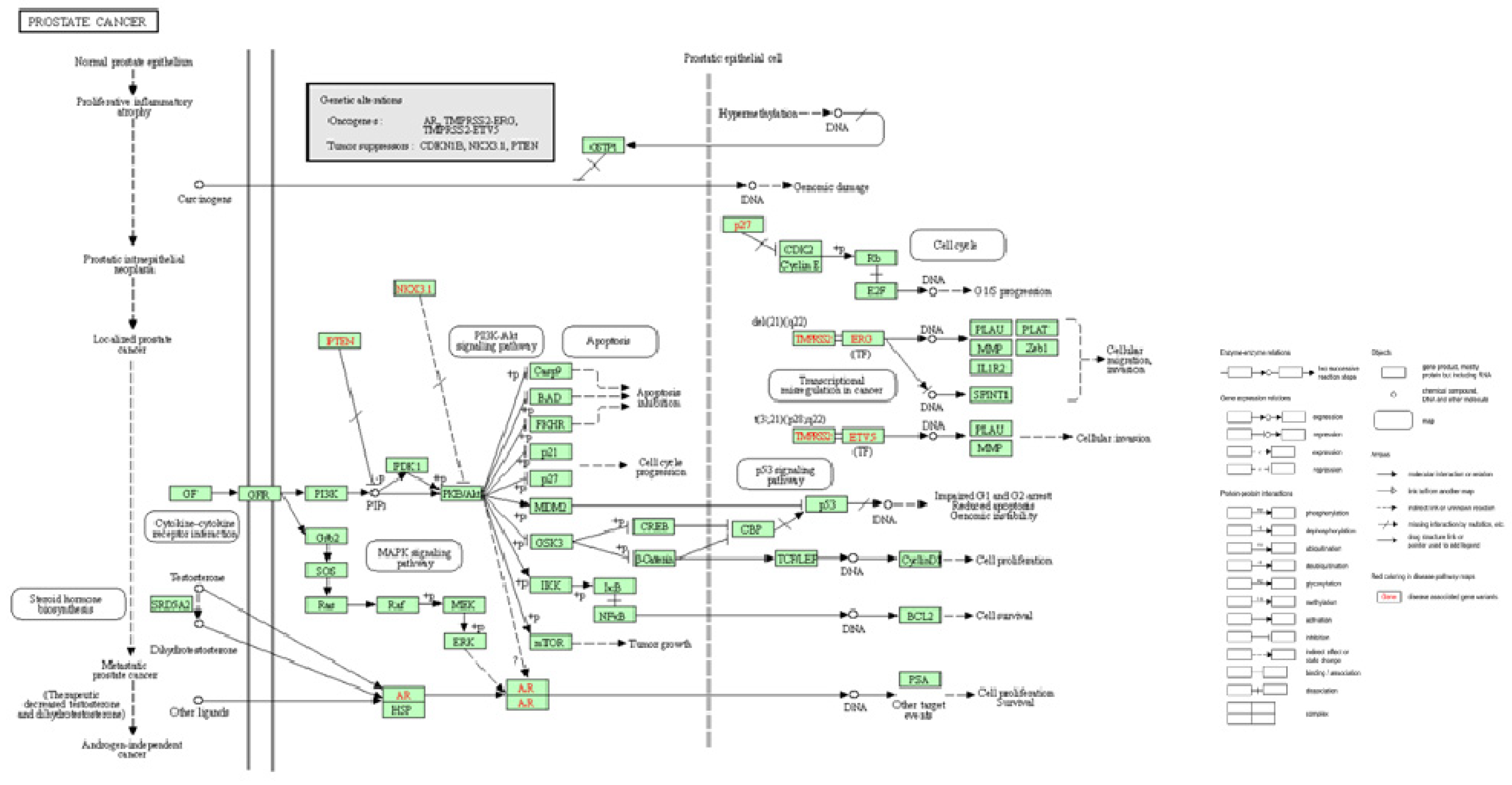

Table 1.
The epidemiology of prostate cancer in 2020 according to the Global Cancer Observatory (GCO) [2].
Table 1.
The epidemiology of prostate cancer in 2020 according to the Global Cancer Observatory (GCO) [2].
| Region | Incidence | Deaths | 5-year prevalence (all ages) | |||||
| number | % of all sites | Rank | number | % of all sites | Rank | number | Per 100 000 | |
| World | 1 414 259 | 7.3% | 3 | 375 304 | 3,8% | 8 | 4 956 901 | 126,13 |
| Europe | 473 344 | 10.8% | 3 | 108 088 | 5.5% | 5 | 1 873 814 | 518.11 |
| Northern America | 239 574 | 9.4% | 3 | 37 192 | 5.3% | 5 | 929 921 | - |
| Latin America and the Caribbean | 214 522 | 14.6% | 1 | 57 415 | 8.0% | 3 | 709 119 | 220.48 |
| Asia | 371 225 | 3.9% | 8 | 120 593 | 2,1% | 14 | 1 176 781 | 49.59 |
| Africa | 93 173 | 8,4% | 3 | 47 249 | 6.6% | 4 | 178 197 | 26.60 |
| Oceania | 22 421 | 8.8% | 2 | 4 767 | 6,9 | 4 | 89 069 | 416.92 |
| Russia Federation | 46 454 | 7,9% | 3 | 14 434 | 4,6 | 7 | 169 221 | 250.18 |
Table 2.
Oncogenic miRNA in prostate cancer.
| miRNA | Function | Experimental models (Cell lines, animal models) | Patient cohort, size, age, and geographic location, groups of comparison | Reference |
| miR-18a | Increasing cancer progression | - | 160 patients, average age 56.8 ± 12 including stage I, II, and IV presented the National Cancer Institute Cairo compared to 50 normal control healthy male individuals |
[74] |
| miR-21 | Accelerating tumor invasion and inducing castration resistance | - | 170 patient older 45 years from Zagazig University Hospitals, Egypt compared to 70 healthy men | [81] |
| miR-32 | Inhibition of apoptosis and increased proliferation | transgenic mir-32 mice | - | [30] |
| miR-106/miR-25 | Increasing cancer progression | LNCaP cells PC-3 cells |
- |
[31] |
| miR-125b |
Increase in cell proliferation and suppression of apoptosis | The human PCa cell lines: C4-2 CWR22Rv1 BCa cell lines: T24, TCC-SUP, UMUC3, TCC-5637, and 293T |
- | [82] |
| miR-141 | Development of castration resistance | LNCaP cells PC-3 cells |
- | [82,83,84] |
| miR-221/miR-222 | Increased cell proliferation, invasion, cell survival | LNCaP, PC3 | - | [85] |
| miR-375 |
Diagnostics | LNCaP, PC3 | - | [84] |
| miR-650 | Reduced expression of the cellular stress response gene 1 (CSR1). | PC3 | 216 patients from 45 through 79 years from Pittsburgh, USA compared with 77 healthy men | [86] |
| miR-4534 | Downregulating the tumor suppressor PTEN gene | LNCaP, PC3 | [80] |
LNCaP cells are androgen-sensitive human prostate adenocarcinoma cells derived from the left supraclavicular lymph node metastasis from a 50-year-old caucasian male in 1977. PC-3 is a cell line initiated from a bone metastasis of a grade IV prostatic adenocarcinoma from a 62-year-old male. C4-2 is a cell line with epithelial-like morphology that was isolated from a human prostate cancer LNCaP cell subcutaneous xenograft tumor of castrated mouse CWR22Rv1 human prostate carcinoma epithelial cell line derived from a xenograft BC- bladder cancer. T24 a cell line established from a human urinary bladder cancer patien TCC-SUP is a cell line isolated from the urinary bladder of a female with grade IV transitional cell carcinoma. UMUC3 is an epithelial-like cell that was isolated from the urinary bladder male of a patient and can be used in cancer research TCC-5637 is a cell line exhibiting epithelial morphology that was isolated from the urinary bladder of a 68-year-old, White male patient with grade II carcinoma 293T is an immortalized cell line derived from the embryonic human kidney that is transfected with sheared human adenovirus type 5DNA.
Table 3.
Diagnostic procedures for prostate cancer according to the international clinical guidelines [108,109].
| Diagnostic method | Principle | Sensitivity (0-1) |
Specificity (0-1) | False-negative cases (%) | False positive cases (%) | Benefits | Limitations | Reference |
| Digital Rectal Examination (DRE) | Palpation of the lower part of the rectum, pelvis and lower abdomen | 0,51 | 0,59 | - | - | availability and affordability non-invasive |
low sensitivity lack specificity More than 60% are identified as asymptomatic |
[118,139,140] |
| Prostate Specific Antigen (PSA) | Venous blood sampling for prostate-specific antigen, a glycoprotein expressed in both cancerous and normal columnar prostate epithelial cells. | 0,21-0,5 | 0,91 | 10-15% | - | availability and affordability | lack specificity predictive accuracy of 8% to 10% |
[118,141,142] |
| Transrectal Ultrasound Scan (TRUS) | Ultrasound examination of the prostate with insertion of the sensor into the rectum. | - | - | 11,34-29,31 | 4,61-6,11% | availability and affordability non-invasive |
lack specificity | [123] |
| Transrectal biopsy (TRB) | Tissue sampling with a thin needle that is inserted through the rectum into the prostate. | 0,53 | 1 | 11-46% | - | Availability affordability |
most lesion are small and sometime located in regions that are not identifiable Complications of prostate biopsy (eg, infection, pain, bleeding, urinary obstruction) |
[118,139,143,144,145,146] |
| MRI -guided biopsy | MRI-guided sampling of prostate tumor tissue | 0.77 | 1 | 6% | 4,2% | accuracy | Complications of prostate biopsy (eg, infection, pain, bleeding, urinary obstruction); expensive |
[143,144,145] |
| MRI | Creation of detailed volumetric images of areas using a magnetic tomography | 0.67 | 0.92 | 2,7% | 44,1% | non-invasive |
expensive |
[143,145] |
“-“ no data available
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



