
Submitted:
15 June 2023
Posted:
16 June 2023
You are already at the latest version
Abstract
Anabantidae is a large and diverse group of fish cultured both under aquaculture conditions and as hobbyists. These fish share a common structural feature in the form of an additional respiratory organ. Despite the enormous availability of these fish worldwide, little is known to date about their feeding preferences in husbandry and their influence on homeostasis under both industrial and domestic conditions. This study describes, for the first time, the structure of the digestive tracts of two Anabantoidei fishes: African bushfish (Ctenopoma acutirostre) and paradise fish (Macropodus opercularis). The overall structure of the digestive tract and its histological structure were analyzed and compared in both fish species. In conclusion, it was shown that physiological predispositions indicate a predominance of omnivorous fish traits in the M. opercularis, in contrast to the C. acutirostre, which has several morphological traits indicating a greater adaptation to carnivory, particularly ichthyophagy. The results obtained will allow further research to be conducted in the future to optimize the nutrition and feeding of these fish and to develop appropriate dietary recommendations.
Keywords:
labirynth fish
; digestive tract
; fish morphology
; feeding
1. Introduction
Labyrinths (Anabantoidei) are the most famous group of fish in the world, especially guramis (Osphronemidae) and siamense fighting fish (Betta splendens) for their aesthetic qualities [1]. Easy to breed at home, these fish are often chosen not only by hobbyists of labyrinthine fish but also by beginners in the aquarium hobby. The main reason is the intense colouration, small body size, and interesting breeding behaviour unlike other fish groups. Ornamental fish aquaculture is gaining an increasing number of followers every year owing to its high profitability compared with consumer fish aquaculture. It is estimated that the market value of this animal production sector is worth $15-20 trillion/per annum and will continue to grow rapidly in the coming years [2,3]. This branch of aquaculture includes not only the reproduction of aquarium fish but also those that are caught in the wild. Aquarium fish are of great interest to amateurs, but in some cases, to scientists. An excellent example is zebrafish (Danio rerio), medaka (Oryzias latipes) and turquoise killifish (Notobranchius furzeri), which are the subjects of molecular, physiological, and neurodegenerative research (e.g. [4,5,6,7]).
The suborder Anabantoidei includes the following families: Anabantidae (about 30 species), Helostomatidae (1 species), Osphronemidae (120 species) with subfamilies Osphroneminae, Luxiocephalinae, Macropodusinae [8,9]. There is only one species in the Helostomatidae family - Helostoma temmincki. All fish from this suborder are characterized by the presence of a labyrinth, which is an auxiliary respiratory organ transformed from gill arches [10]. Fish from the family Anabantidae are found both in Africa and Asia, while the others are found only in Asia, occupy various ecological niches. A common feature of the environment in which they occur is the low oxygen content in the water [1,11,12,13,14,15].
Ctenopomas are native to Africa Anabantoidei, the bush warbler from the Congo region of central Africa [16]. These fish are animals that swim close to the bottom, slowly, and are more active at night than during the day. They are not only amateur bred but also in production for further distribution around the world. Much better production results are achieved when fed with natural food than with commercial feed, however, the diet does not significantly affect the survival of juvenile stages [15,16]. As predatory fish, Ctenopomas will be more likely to eat natural food than fodder, regardless of its composition. These fish are mainly insectivores, however, they can pray even on small vertebrates [17,18,19].
Macropodusinae is small territorial fishes, bred both as an ornamental species and as a subject of behavioural research. Their dynamic character (agonistic), similar to betta fish, is used in southeast Asia to conduct fish fights. It comes from Taiwan, southern China, northern Vietnam and the Ryukyu Islands [12]. Its production is much simpler compared to Ctenopoma due to the rapid achievement of sexual maturity and sexual dimorphism facilitating breeding.
The structure and organization of the digestive tract in fish differ depending on the type of food ingested by these organisms. In predatory carnivorous fish such as Ctenopoma acutirostre or the paradise fish (Macropodus opercularis), a significant difference can be observed in the length of the digestive tract compared to herbivorous fish such as the kissing gourami (Helostoma temminkii). The digestive tracts of plant- or plankton-eating fish are much shorter than those of carnivorous and omnivorous fishes [20].
Due to the growing interest in aquaristics, both in the countries of origin and for export, it is important to develop optimal conditions for breeding and breeding labyrinths. The development of appropriate and welfare-friendly methods and strategies for feeding juveniles and adults requires basic data on the structure of the digestive tract. In the fish families Anabantidae and Osphronemidae, two species of the most commonly farmed fish of great economic importance were selected to describe their anatomy and compare the structure of their digestive tracts.
2. Materials and Methods
The study material consisted of 15 specimens of Ctenopoma acutirostre and Macropodus opercularis collected in 2017-2019. Fish were euthanized with MS222 and then weighed and measured (Table 1). Whole fish were collected or only their digestive tract if body weight did not allow whole fish fixation. The material was fixed in Bouin's fluid and then subjected to a standard histological procedure. Paraffin-embedded slides were sectioned longitudinally to a thickness of 6um and then subjected to standard haematoxylin and eosin (HE) staining, as well as Alcian blue and Schiff's periodic acid reagent (AB/PAS) staining for visualization of acidic and neutral mucus cells. The microscopical analysis was conducted with a Nikon Eclipse NI-E microscope with a camera Nikon DSFi3 and NIS Elements AR software (Nikon, Tokyo, Japan).
3. Results
- The general structure of the digestive tract
The general structure of the digestive tract was similar in both fish species. The large superior, protrusible mouth was found in C. acutirostre. The following sections of the gastrointestinal tract were specified: headgut from the superior mouth to the oesophagus, oesophagus, stomach, intestine from the pyloric caece opening, sectioned to the anterior, mid- and posterior part (Figure 1). The general morphometrical comparison was showed that intestine length was higher in M. opercularis compared to C. acutirostre. The gut length calculated with standard length was higher too in this species (Table 2).
3.1. Headgut
The opening section of the digestive tract began with the mouth covered by nonkeratinized squamous epithelium with teeth localized in the lips (Figure 2). These teeth had long roots arising in the bone of the maxillary and mandible. In C. acutirostre, jaw teeth were much more developed and prominent compared to M. opercularis (Figure 2A and B). The mouth was lined with thin multi-layered epithelium with numerous acidic mucous cells and singular taste buds (Figure 2C and D). In both species behind the lips were oral valves.
In C. acutirostre, the well-differentiated tongue was characterized by mucous cells and individual taste buds. The latter were found singly throughout the oral cavity, including the interior of the gills and oesophagus. In the M. opercularis, the tongue was merely a protuberance in the oral mucosa with the cartilaginous support of this structure, similar to C. acutirostre (Figure 3). The tongue core in both species was composed of hyaline cartilage with a partially PAS-positive intercellular matrix. Mucosal cells and taste buds lined the mouth cavity and oesophagus, as in C. acutirostre was neutral, mixed, and acidic, unlike in M.opercularis.
The oesophagus also had pharyngeal teeth between the strongly corrugated mucosal divides in this section. The teeth grew from a single dental bone plate. In the following sections, the secretory character of the cells changed to mixed mucous cells (acidic and neutral) and then to exclusively neutral in the stomach (Figure 4).
The stomachs of both C. acutirostre and M. opercularis resembled sacs similar to those of monogastric terrestrial vertebrates. The epithelial cells were covered by neutral mucins-stained magenta in AB/PAS (Figure 4 A and B). M. opercularis stomachs were less folded and complex compared to C. acutirostre. Behind the gastric opening, two short pyloric caeca were observed in both species, in M. opercularis larger than in C. acutirostre (Table 2). Similarly, to follow sections of the digestive tract the epithelium cells were higher compared to the stomach with singular mucosal cells produced acidic and mixed (neutral and acidic) mucins (Figure 4 C-G). In some intestine folds were present singular yellow-brownish macrophages. The pH of mucosal cells had a different pattern in C. acutirostre compared to M. opercularis. In the C. acutirostre the pH of mucosal cells was mostly highly acidic or mixed. In M. opercularis the pH of mucosa cells were acidic too but with more frequently present mixed and neutral cell, in particular the anterior intestine.
4. Discussion
Ornamental fish kept at home or in aquaculture, like well-known commercial fish species, require optimal husbandry conditions. Optimal husbandry conditions require knowledge and a basic understanding of the anatomy, morphology and physiology of the animals kept. For fish, aquatic animals with extremely varied food-acquisition strategies, knowledge of the structure of the digestive tract is particularly important and allows subsequent research into the physiology of this system. Such knowledge can help to avoid feeding and welfare problems, which often occur during fish breeding in home aquaria [3,21].
The studied fish species had similar gastrointestinal structures, though there were significant differences in both of them. The overall structure of the digestive tract of C. acutirostre and M. opercuaris indicated differences in both the ability to obtain food and to swallow and subsequently digest it. Gut length, which contributes to absorption efficiency, concerning total fish length is strongly correlated with diet [22,23,24,25]. In carnivorous fish, the digestive tract with total fish length is shorter compared to omnivorous or herbivorous fish [24,26], which was also confirmed in this study. The comparative analysis conducted in the present study showed that the digestive tracts of C. acutirostre are shorter compared to M. opercularis. These observations indicate a greater adaptation to a carnivorous diet compared to omnivorous M. opercularis. Previous studies on the feeding of C. acutirostre under aquaculture conditions have shown that the best growth rates were achieved with live natural food (silk worms)[15,16]. However, it is important to take into account the significant variation in food preferences depending on the location of C. acutirostre, due to the diverse ecological niches occupied by this species. As an example of such variation, another species Ctenopoma pathereri Gunther known similarly to C. acutirostre as an omnivore, in its habitat in the Oluwa River, Ondo State, Nigeria has greater herbivorous tendencies [27].
Not only the length of the intestines indicated differences in diet, but also the morphological structure of the digestive tract starting with the superior mouth opening [28]. The mouth opening of C. acutirostre was massively built with and deeply indented, indicative of the strong structure and considerable length of the maxillary, premaxillary and dentary bones. This construction pattern also promotes the protrusion of the snout when grasping prey. In contrast to C. acutirostre, in M. opercularis the mouth opening is built less massively, even delicately, and does not show such a deep indentation. In fish living in waters with less light and poor visibility, as is also periodically the case in Anabantoidei, taste buds located on the mouth allow food recognition and search [29]. Their role in low-transparency environments are extremely valuable in enabling not only the identification of food but also the assessment of its tastiness. In predatory fish, the mouth opening is more extensive, equipped with teeth that help to capture and hold prey, further assisting itself by sucking in water along with the prey. These teeth can have a highly variable shape, closely linked to the diet. In carnivorous fish, the teeth are narrow, long and sharp; in omnivorous or predominantly herbivorous fish, they are much lower, broad and blunt. The localised teeth in the two species studied were found both on the lips and in the oesophagus located on the bony plate. Given the mobility and width of the mouth opening of C. acutirostre the teeth present and the central valve help to catch, suck in and impede the retraction of food. The design of the mouth and mouth opening of omnivorous M. opercularis and C. acutirostre clearly indicated that the mouth openings of C. acutirostre were significantly more adapted to capture larger prey. Since in the wild C. acutirostre prey not only on insects, like M. opercularis also does, but also on arachnids and even frogs and other fish, the occurrence of teeth still in the lips indicates their adaptation for hunting live food [30,31].
In some fish species [32,33], similarly to terrestrial vertebrates, a tongue is present in the mouth cavity, but its structure and mobility are significantly reduced. The tongue of Anabantoidei fishes was characterized by significantly less mobility and a less complex structure compared to terrestrial vertebrates (lacking such complex undulations, and differentiated taste buds in terms of structure). Despite this, this organ has not only functions related to swallowing and swiping food but also to the identification of taste stimuli through taste buds located also on its surface [33]. Thus far, the fish's tongue has been described in the literature as a thickening at the bottom of the mouth [29,33,34,35], whereas in recent years, increasing literature data indicates the importance of this organ in food intake [36], and its identification and recognition of taste values due to the numerous taste buds present on it [33,34]. The tongues of the Anabantoidei examined were characterised by a convergent overall histological structure, however, the tongue of C. acutirostre was much longer and more freely attached to the floor of the oral cavity, leading to the conclusion that it is probably more mobile compared to the deeply embedded, small tongue of M. opercularis. The motility of the tongue of C. acutirostre, due to its apparent lack of muscular tissue, does not consist of conscious and deliberate movement as is the case in e.g. mammals, instead using it, together with the oral valve, to swallow, restrain, and force further through the swallowed food similar to other ichthyophages. The rigidity of this organ is due to the structure of its core constituted by supporting connective tissue. The staining of the tongue core in HE and AB/PAS staining indicates that it is an osteocartilaginous skeleton, similar to what was observed in another ichthyophage northern pike (Esox lucius)[37]. In contrast, none of the specimens examined revealed the presence of tongue surface undulations with teeth growing from the tongue surface that Levanti et al., described in sea bass (Dicentrarchus labrax), seabream (Sparus aurata) and white seabream (Diplodus sargus sargus) [33].
The food passed through the pharyngeal teeth and entered the stomach. In many fish species, the stomach may not be present at all - its functions are performed by the morphologically transformed anterior intestine. However, in many fish species in which the stomach is absent, partial compensation for this organ is also provided by expanded wide pharyngeal or gizzard teeth, which grind the food more thoroughly than in other fish species, facilitating its further digestion by the transformed initial intestinal region [38,39]. In the case of Andabantoidei representatives, the presence of a stomach is confirmed not only by the present study, as well as those performed on dwarf gourami (Colisa lalia)[40], giant gourami (Osphonemus goramy Lacepede, 1801) [41], or snakeskin gourami (Trichopodus pectoralis)[42] studies. In carnivorous fish, the stomach mucosa is more strongly developed to cope with the digestion of more complex food containing large amounts of protein. Considering this aspect, the stomachs of C. acutirostre were more strongly developed compared to those of M. opercularis. Nonetheless, pyloric caeca were found to be present in both studied species with stomachs secreting the necessary enzymes to partially digest the nutritional matter.
The pyloric appendages are an evolutionary solution to increase the absorptive surface area of the digestive tract. Their structure is very similar to that of the distal intestinal tract. Their number is a species-specific trait, varying significantly from a few appendages in fish such as spotted snakehead (Channa punctata) to several hundred in Atlantic cod (Gadus morhua) [22]. In stomachless fish, they are absent [39,43,44]. Previous studies have not shown a correlation between the number of pyloric caeca and the type of food [22], but their number is observed to be lower in omnivorous and herbivorous species compare to carnivores [45], due to the need to digest chemically and structurally more demanding food. The presence of only two pyloric caeca suggests that the efficiency of digestion in the earlier sections of the digestive tract is probably sufficient to ensure proper absorption of some nutrients already in the first sections of the intestines. In the further sections, no significant differences in the structure of the digestive tract were observed, and the normal layered structure of the intestines, consisting of mucosa, submucosa and musculature, characteristic of all vertebrates, was found. The only difference was a stronger AB-positive staining of the mucosal cells of the anterior and middle intestines of C. acutirostre indicating the more acidic character of the produced mucus.
5. Conclusions
Based on our analyses, we have shown that representatives of the Anabantoidei, belonging to two different families considered omnivorous fish, are characterized by a different structure of the digestive tract indicating that the C. acutirostre has a much better adapted digestive tract for the acquisition and processing of carnivorous food. The absence of differences in the histological structure of the intestines does not imply the lack of differences in their physiology. The results of this study provide a basic knowledge of the morphology and histology of the digestive tracts of these fish, providing a strong foundation for further research into the physiology of the digestive tract and optimizing the feeding of the two species studied. To demonstrate the physiological differences in the two species, it would be necessary to extend the study to analyze the localization and expression of cells that regulate digestive processes, not only under optimal rearing conditions but also during feeding experiments. These studies would provide new information on the nutritional needs of these species allowing the development of a more optimal diet, which could probably benefit the survival rate, especially of fish in the larval and juvenile stages, characterized by the highest mortality.
Author Contributions
Conceptualization D.A.U.; methodology D.A.U.; formal analysis, D.A.U., M.K., W.W., A.S.; investigation, D.A.U., R.F., J.B.; resources D.A.U., M.K.; data curation D.A.U.; writing—original draft preparation D.A.U. M.K., W.W.; writing—review and editing, D.A.U.; visualization, D.A.U., R.F., J.B. A.S., supervision, J.Ś.; project administration D.A.U., W.W.; funding acquisition, D.A.U. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Science Center (Poland), grant number 2021/05/X/NZ4/00761
Institutional Review Board Statement
Ethical review and approval were waived for this study, as no experimental, stress-inflicting procedures were performed on the studied fish. Fish were euthanized according to the guidelines of the 2nd Warsaw Local Ethics Committee for Animal Experimentation (at Warsaw University of Life Sciences – SGGW), which follow Directive 2010/63/EU of the European Union, on the protection of animals used for scientific purposes.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Yu, T.; Guo, Y. Early Normal Development of the Paradise Fish Macropodus opercularis. Russ. J. Dev. Biol. 2018 494 2018, 49, 240–244. [Google Scholar] [CrossRef]
- Evers, H.G.; Pinnegar, J.K.; Taylor, M.I. Where are they all from? – sources and sustainability in the ornamental freshwater fish trade. J. Fish Biol. 2019, 94, 909–916. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.; Alexander, M.E.; Snellgrove, D.; Smith, P.; Bramhall, S.; Carey, P.; Henriquez, F.L.; McLellan, I.; Sloman, K.A. How should we monitor welfare in the ornamental fish trade? Rev. Aquac. 2022, 14, 770–790. [Google Scholar] [CrossRef]
- Kowalska, A.; Kamaszewski, M.; Czarnowska-Kujawska, M.; Podlasz, P.; Kowalski, R.K. Dietary ARA Improves COX Activity in Broodstock and Offspring Survival Fitness of a Model Organism (Medaka Oryzias latipes). Anim. 2020, Vol. 10, Page 2174 2020, 10, 2174. [Google Scholar] [CrossRef] [PubMed]
- Wiweger, M.; Majewski, L.; Adamek-Urbanska, D.; Wasilewska, I.; Kuznicki, J. npc2-Deficient Zebrafish Reproduce Neurological and Inflammatory Symptoms of Niemann-Pick Type C Disease. Front. Cell. Neurosci. 2021, 15. [Google Scholar] [CrossRef]
- Reeves, G.A.; Singh, P.P.; Brunet, A. Chromatin Accessibility Profiling and Data Analysis using ATAC-seq in Nothobranchius furzeri. 2023. [Google Scholar] [CrossRef]
- Žák, J.; Anil, A.N.; Dyková, I. Dissolved oxygen saturation is crucial for gas bladder inflation in turquoise killifish (Nothobranchius furzeri). Environ. Biol. Fishes 2023, 106, 673–683. [Google Scholar] [CrossRef]
- Rüber, L. Labyrinth fishes (Anabantoidei). Timetree Life 2009, 344–347. [Google Scholar]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World; John Wiley & Sons, Inc: Hoboken, NJ, USA, 2016; Vol. 27, ISBN 9781119174844. [Google Scholar]
- Adamek-Urbańska, D.; Błażewicz, E.; Sobień, M.; Kasprzak, R.; Kamaszewski, M. Histological study of suprabranchial chamber membranes in anabantoidei and clariidae fishes. Animals 2021, 11, 1158. [Google Scholar] [CrossRef]
- Dadgar Sh., H. M.. Z.D.; A.R., G. Optimum dietary protein requirement of Paradise fish, Macropodus opercularis based on growth and reproduction performances. J. Surv. Fish. Sci. 2020, 7, 91–104. [Google Scholar] [CrossRef]
- Kitagawa, T.; Hosoya, K. Origin of the Ryukyuan paradise fish Macropodus opercularis inferred from literature survey. Biogeography 2016, 18, 11–16. [Google Scholar] [CrossRef]
- Miklosi, A.; Csanyi, V.; Gerlai, R. Antipredator behavior in paradise fish (Macropodus opercularis) larvae: The role of genetic factors and paternal influence. Behav. Genet. 1997, 27, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Dutta, H.M. The suspensorium of Ctenopoma acutirostre: A comparative functional analysis with Anabas testudineus. J. Morphol. 1975, 146, 457–477. [Google Scholar] [CrossRef] [PubMed]
- Prasetya, O.E.S.; Muarif, M.; Mumpuni, F.S. PENGARUH PEMBERIAN PAKAN CACING SUTERA (Tubifex sp.) DAN Daphnia sp. TERHADAP PERTUMBUHAN DAN TINGKAT KELANGSUNGAN HIDUP LARVA IKAN LELE SANGKURIANG (Clarias gariepinus). J. MINA SAINS 2020, 6, 8. [Google Scholar] [CrossRef]
- Santanumurti, M.B.; Nugroho, A.K.; Santoso, L.; Hudaidah, S. Study of Different Protein Content of Feeding of Local Raw Materials on Gourami Fish (Osphronemus goramy Lac.) Aquaculture Performance. IOP Conf. Ser. Earth Environ. Sci. 2022, 1036, 012119. [Google Scholar] [CrossRef]
- Hickley, P.; Bailey, R.G. Food and feeding relationships of fish in the Sudd swamps (River Nile, southern Sudan). J. Fish Biol. 1987, 30, 147–159. [Google Scholar] [CrossRef]
- Louca, V.; Lucas, M.C.; Green, C.; Majambere, S.; Fillinger, U.; Lindsay, S.W. Role of fish as predators of mosquito larvae on the floodplain of the Gambia River. J. Med. Entomol. 2009, 46, 546–556. [Google Scholar] [CrossRef] [PubMed]
- G.B. Kassi; M.G. Zeré; S.Y. Stanislas; V. N’douba Régime alimentaire de Ctenopoma petherici (Perciformes, Anabantidae) dans la rivière Agnéby et dans le lac de barrage hydroélectrique d’Ayamé 2 (Côte d’Ivoire). https://popups.uliege.be/2295-8010 2018. [CrossRef]
- Kilarski, W. Anatomia ryb; 2012; ISBN 9788309010807.
- Kasprzak, R.; Grzeszkiewicz, A.B.; Górecka, A. Performance of Co-Housed Neon Tetras ( Paracheirodon innesi) and Glowlight Rasboras ( Trigonostigma hengeli) Fed Commercial Flakes and Lyophilized Natural Food. Anim. an open access J. from MDPI 2021, 11. [Google Scholar] [CrossRef]
- Ray, A.K.; Ringø, E. The Gastrointestinal Tract of Fish. Aquac. Nutr. Gut Heal. Probiotics Prebiotics 2014, 1–13. [Google Scholar] [CrossRef]
- Karachle, P.K.; Stergiou, K.I. Intestine morphometrics: a compilation and analysis of bibliographic data. Acta Ichthyol. Piscat. 2010, 40, 45–54. [Google Scholar] [CrossRef]
- German, D.P.; Horn, M.H. Gut length and mass in herbivorous and carnivorous prickleback fishes (Teleostei: Stichaeidae): ontogenetic, dietary, and phylogenetic effects. Mar. Biol. 2006, 148, 1123–1134. [Google Scholar] [CrossRef]
- Cho, J.H.; Park, J.W.; Ryu, Y.W.; Kim, K.W.; Hur, S.W. Morphology, Histology, and Histochemistry of the Digestive Tract of the Marbled Flounder Pseudopleuronectes yokohamae. Anim. 2023, Vol. 13, Page 936 2023, 13, 936. [Google Scholar] [CrossRef] [PubMed]
- Karachle, P.K.; Stergiou, K.I. Gut length for several marine fish: relationships with body length and trophic implications. Mar. Biodivers. Rec. 2010, 3, e106. [Google Scholar] [CrossRef]
- Omoniyi, I.T.; Agbon, A.O.; Akegbejo-Samsons, Y. The Food Habits of Ctenopoma pethereci, Gunther (Pisces: Anabantidae) in River Oluwa, Ondo State, Nigeria. West African J. Appl. Ecol. 2012, 19. [Google Scholar] [CrossRef]
- Skelton, P.H. The taxonomic identity of dwarf or blackspot Ctenospoma (Pisces, Anabantidae) in Southern Africa. Cybium (Paris) 1988, 12, 73–89. [Google Scholar]
- Buddington, R.K.; Kuz’mina, V. Digestive System. Lab. Fish 2000, 379–384. [Google Scholar] [CrossRef]
- Meunier, F.J.; Brito, P.M.; Leal, M.-E.C. Morphological and histological data on the structure of the lingual toothplate of Arapaima gigas (Osteoglossidae; Teleostei). Cybium 2013, 37, 263–271. [Google Scholar]
- El Bakary, N.E.S.R. Morphological study of the asymmetrical buccal cavity of the flatfish common solea (Solea solea) and its relation to the type of feeding. Asian Pac. J. Trop. Biomed. 2014, 4, 13. [Google Scholar] [CrossRef]
- MEYER-ROCHOW, V.B. Fish tongues—surface fine structures and ecological considerations. Zool. J. Linn. Soc. 1981, 71, 413–426. [Google Scholar] [CrossRef]
- Levanti, M.; Germanà, A.; Montalbano, G.; Guerrera, M.C.; Cavallaro, M.; Abbate, F. The Tongue Dorsal Surface in Fish: A Comparison Among Three Farmed Species. Anat. Histol. Embryol. 2017, 46, 103–109. [Google Scholar] [CrossRef]
- Abbate, F.; Guerrera, M.C.; Montalbano, G.; Ciriaco, E.; Germanà, A. Morphology of the tongue dorsal surface of gilthead seabream (Sparus aurata). Microsc. Res. Tech. 2012, 75, 1666–1671. [Google Scholar] [CrossRef] [PubMed]
- Abbate, F.; Guerrera, M.C.; Montalbano, G.; De Carlos, F.; Suárez, A.Á.; Ciriaco, E.; Germanà, A. Morphology of the european sea bass (dicentrarchus labrax) tongue. Microsc. Res. Tech. 2012, 75, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Camp, A.L.; Brainerd, E.L. Role of axial muscles in powering mouth expansion during suction feeding in largemouth bass (Micropterus salmoides). J. Exp. Biol. 2014, 217, 1333–1345. [Google Scholar] [CrossRef] [PubMed]
- Sadeghinezhad, J.; Rahmati-Holasoo, H.; Fayyaz, S.; Zargar, A. Morphological study of the northern pike (Esox lucius) tongue. Anat. Sci. Int. 2015, 90, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Palasai, A.; Lampang, P.N.; Gerald Plumley, F.; Kettratad, J.; Senarat, S.; Kangwanrangsan, N.; Jiraungkoorskul, W. Feeding apparatus, digestive system structure, and gut contents of Priapium fish, Neostethus lankesteri Regan 1916. Songklanakarin J. Sci. Technol 44, 602–608.
- Le, H.T.M.D.; Shao, X.; Krogdahl, Å.; Kortner, T.M.; Lein, I.; Kousoulaki, K.; Lie, K.K.; Sæle, Ø. Intestinal function of the stomachless fish, ballan wrasse (Labrus bergylta). Front. Mar. Sci. 2019, 6, 438019. [Google Scholar] [CrossRef]
- Y, H. INFLUENCES OF GUARDING TERRITORY ON REPRODUCTIVE ACTIVITY IN THE MALE DWARF GOURAMI, COLISA LALIA. Indian J. Sci. Technol. 2011, 4, 113–113. [Google Scholar] [CrossRef]
- Nugraha, A.A.; Yustiati, A.; Bangkit, I.; Andriani, Y. Growth performance and survival rate of giant gourami fingerlings (Osphronemus goramy Lacepede, 1801) with potassium diformate addition. World Sci. News 2020, 143. [Google Scholar]
- Boonanuntanasarn, S.; Jangprai, A.; Yoshizaki, G. Characterization of proopiomelanocortin in the snakeskin gourami (Trichopodus pectoralis) and its expression in relation to food intake. Domest. Anim. Endocrinol. 2015, 50, 1–13. [Google Scholar] [CrossRef]
- Tan, X.; Hoang, L.; Du, S.J. Characterization of muscle-regulatory genes, Myf5 and Myogenin, from striped bass and promoter analysis of muscle-specific expression. Mar. Biotechnol. 2002, 4, 537–545. [Google Scholar] [CrossRef]
- Moreira, E.; Novillo, · Manuel; Eastman, J.T.; Barrera-Oro, E. Degree of herbivory and intestinal morphology in nine notothenioid fishes from the western Antarctic Peninsula. 2020, 43, 535–544. [CrossRef]
- Rust, M.B. Nutritional Physiology. Fish Nutr. 2003, 367–452. [Google Scholar] [CrossRef]
Figure 1.
Macroscopic view of the digestive tract of C. acutirostre (A) and M. opercularis (B) OS – oesophagus ST – stomach PC - pyloric caeca AI - anterior intestine MI - mid-intestine PI - posterior intestine R – rectum.
Figure 1.
Macroscopic view of the digestive tract of C. acutirostre (A) and M. opercularis (B) OS – oesophagus ST – stomach PC - pyloric caeca AI - anterior intestine MI - mid-intestine PI - posterior intestine R – rectum.

Figure 2.
Transverse sections of mouth in (A, C) the African bushfish and (B, D) paradise fish. In cross-section through the lower lips, the weakly staining PAS-positive bone from which the teeth emerge is evident. In African bush-fish (A), this is a uniform block of tissue, unlike in the paradise fish (B). In HE staining, taste buds located both on the external and internal parts of the mouth cavity were also visible (magnifications in red frame); A, B - AB/PAS staining; C, D – HE staining.
Figure 2.
Transverse sections of mouth in (A, C) the African bushfish and (B, D) paradise fish. In cross-section through the lower lips, the weakly staining PAS-positive bone from which the teeth emerge is evident. In African bush-fish (A), this is a uniform block of tissue, unlike in the paradise fish (B). In HE staining, taste buds located both on the external and internal parts of the mouth cavity were also visible (magnifications in red frame); A, B - AB/PAS staining; C, D – HE staining.
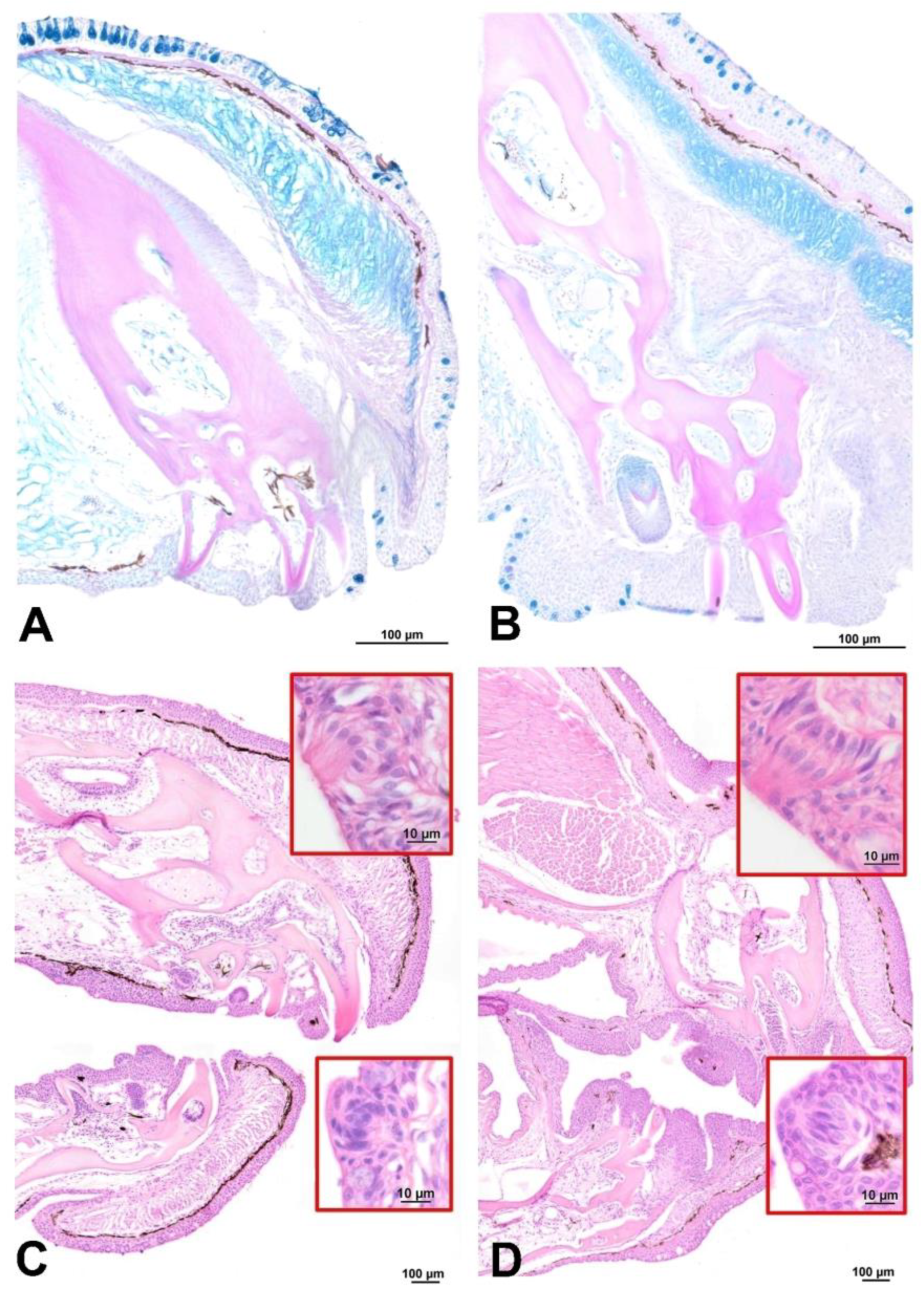
Figure 3.
In African bush-fish (A, C in black frame) the tongue structure was visible macroscopically (A green arrow) with the oral valve in the upper mouth. The histological structure of the tongue showed that the whole structure is based on two fragments of hyaline cartilage fith with the front growing cartilage full of magenta-positive droplets (magnification in red frame). The tongue of paradise fish was structured with based hyaline cartilage with singular areas of ossification (magnification in blue frame).
Figure 3.
In African bush-fish (A, C in black frame) the tongue structure was visible macroscopically (A green arrow) with the oral valve in the upper mouth. The histological structure of the tongue showed that the whole structure is based on two fragments of hyaline cartilage fith with the front growing cartilage full of magenta-positive droplets (magnification in red frame). The tongue of paradise fish was structured with based hyaline cartilage with singular areas of ossification (magnification in blue frame).
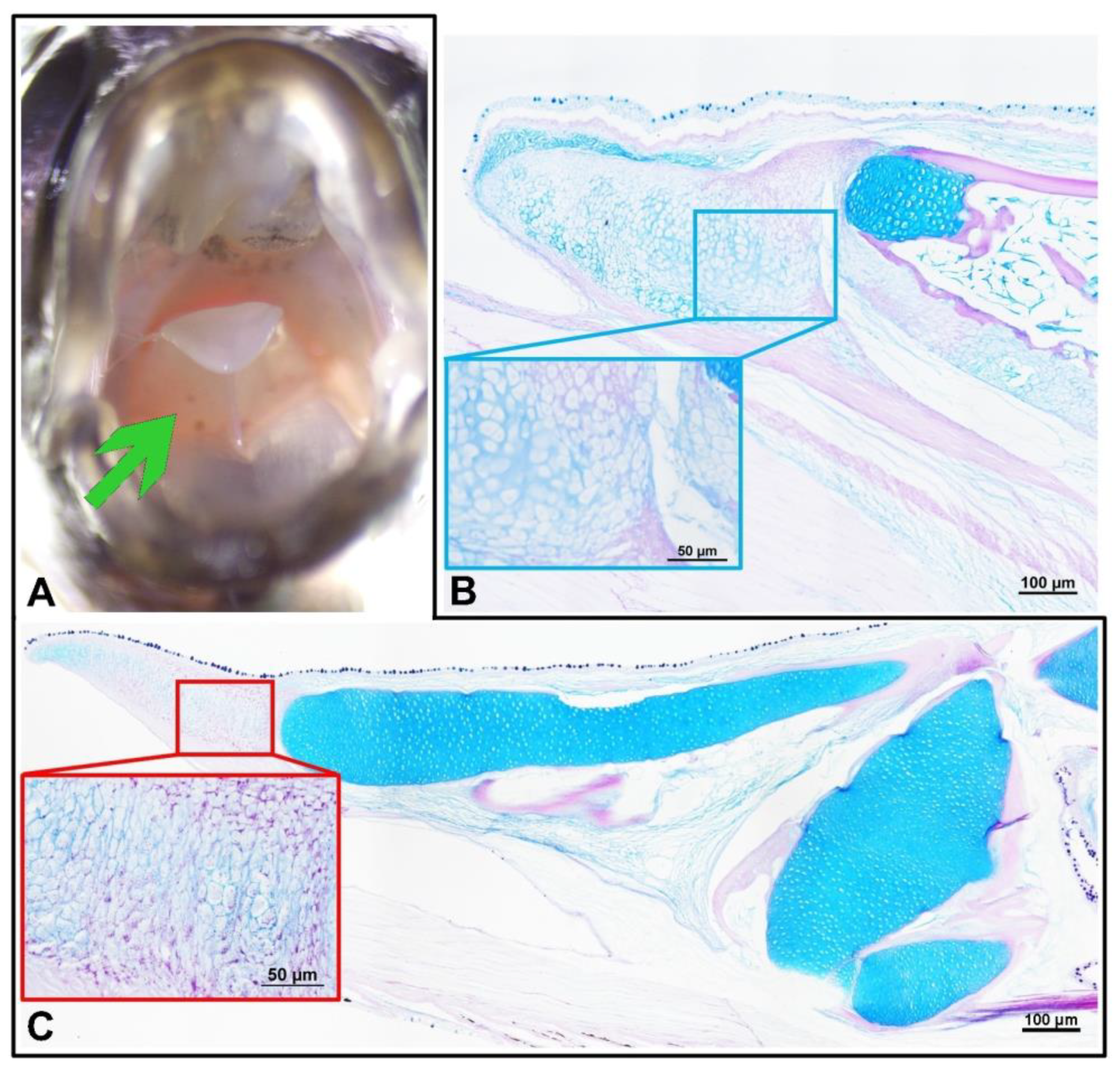
Figure 4.
Histological structure of digestive tract sections: stomach, anterior, mid-, posterior intestine and pyloric caeca. In the stomach mucosa, the neutral mucin was stained in a magenta colour, clearly visible in the apical part of the cells. In the following sections, the mucosal cells were stained in the blue colour; AB/PAS staining.
Figure 4.
Histological structure of digestive tract sections: stomach, anterior, mid-, posterior intestine and pyloric caeca. In the stomach mucosa, the neutral mucin was stained in a magenta colour, clearly visible in the apical part of the cells. In the following sections, the mucosal cells were stained in the blue colour; AB/PAS staining.
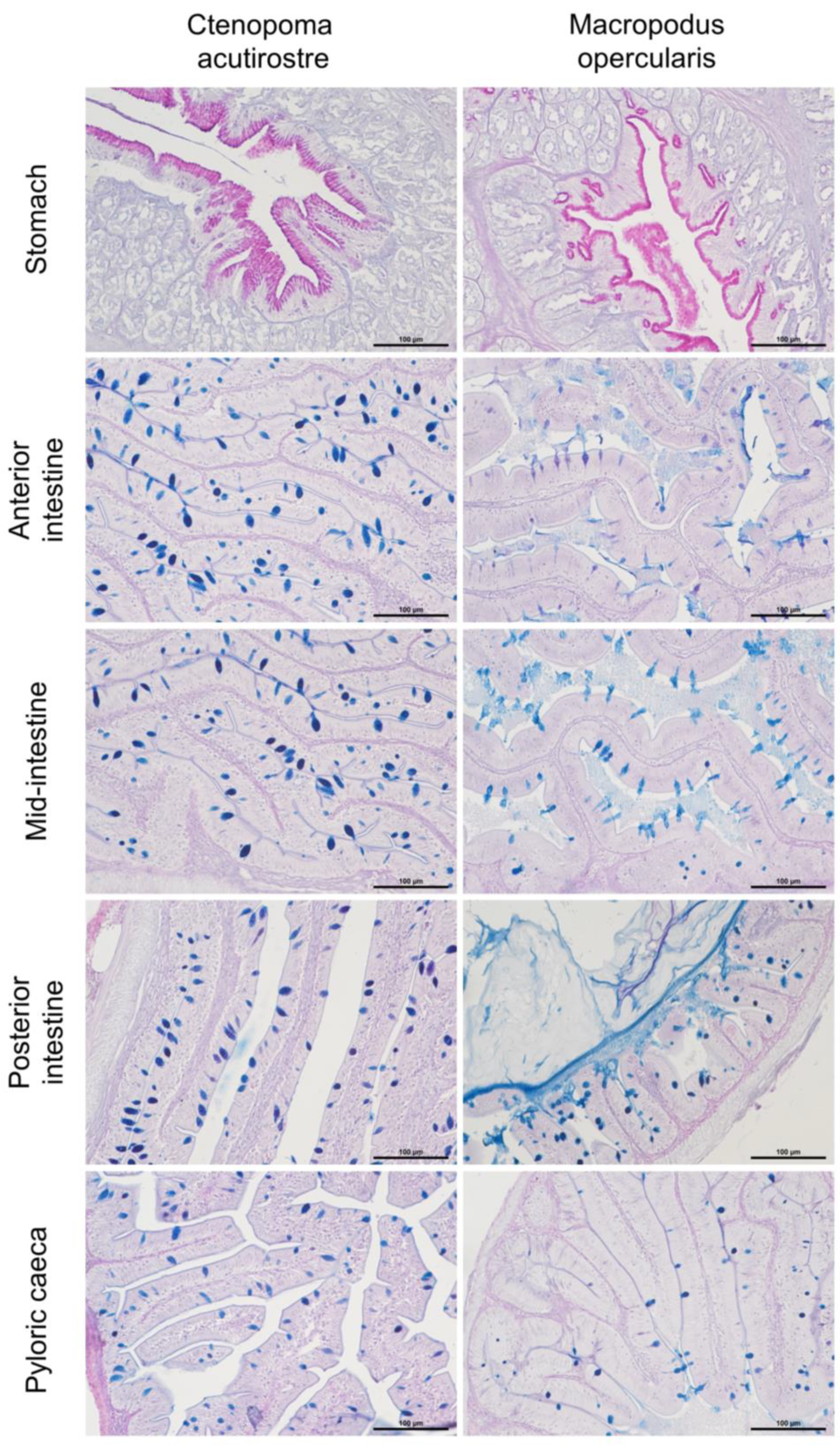

Table 1.
Morphometrical data of investigated fish [n=15].
| Total length [cm] | Standard length [cm] | Fork length [cm] | Body weight [g] | |
|---|---|---|---|---|
| Ctenopoma acutirostre | 5,39 ± 0.23 | 4,58 ± 0.24 | 0,82 ± 0.09 | 2,88 ± 0.56 |
| Macropodus opercularis | 5.47 ± 0.49 | 3,68 ± 0.18 | 1,79 ± 0.37 | 1,30 ± 0.49 |
Table 2.
General morphometrics of C. acutirostre and M. opercularis (SL standard length, IL intestine length).
Table 2.
General morphometrics of C. acutirostre and M. opercularis (SL standard length, IL intestine length).
| Length [cm] | Ctenopoma acutirostre | Macropodus opercularis |
|---|---|---|
| SL [cm] | 4.58 ± 0.24 | 3.68 ± 0.18 |
| IL [cm] | 3.38 ± 0.47 | 4.14 ± 0.70 |
| Pyloric caeca length [cm] | 0.25 ± 0.13 | 0.41 ± 0.09 |
| IL/SL | 0.74 ± 0.9 | 1.13 ± 0.14 |
| Range of IL/SL | 0.59-0.83 | 0.93-1.31 |
| % of body length | 59-83 | 93-131 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



