
Submitted:
15 June 2023
Posted:
16 June 2023
You are already at the latest version
Abstract
The mammalian or mechanistic target of rapamycin (mTOR) integrates multiple intracellular and extra-cellular upstream signals involved in the regulation of anabolic and catabolic processes in cells, and plays a key regulatory role in cell growth and metabolism. Activation of the mTOR signaling pathway has been reported to be associated with a wide range of human diseases. A growing number of in vivo and in vitro studies have demonstrated that the gut microbes and its complex metabolites can regulate host metabolic and immune responses through the mTOR pathway, and result in disorders of host phys-iological functions. In this review, we summarize the regulatory mechanisms of gut microbes and mTOR in different diseases, and discuss the crosstalk between gut microbes and their metabolites and mTOR in the disorders in gastrointestinal tract, liver, heart and other organs. We also discuss the promising appli-cation of multiple potential drugs that can adjust the gut microbiota and mTOR signal pathways. Despite the limited findings between gut microbes and mTOR, elucidating their relationship may provide new clues for the prevention and treatment of various diseases.
Keywords:
mTOR
; gut microbes
; metabolites
; therapy
1. Introduction
The mammalian or mechanistic target of rapamycin (mTOR) is a conserved serine/threonine kinase with two structurally similar but functionally distinct protein complexes named mTOR complex1 (mTORC1) and 2 (mTORC2). mTOR is sensitive to different environmental factors, such as energy and nutrient availability, and is involved in regulating cellular activities such as cell proliferation, cell growth, protein synthesis and autophagy [1,2,3]. Dysregulation of mTOR signaling has been reported in manydiseases such as cancer, diabetes, heart disease, and neurological disorders [2,4,5].
Commensal microorganisms in human body have gained more and more attentions in recent years. The vast majority of commensal microorganisms colonize in the gut and differ between individuals due to genetic background, age, sex, diet, and other factors. It has been found that gut microbiota are involved in the regulation of various diseases such as intestinal diseases, liver diseases, heart diseases, obesity [6], diabetes [7] and Alzheimer's disease [8], and are associated with physiological processes such as immunity and autophagy.
In the present review, we discuss mTOR signaling in detail and review recent evidence that gut microbes and mTOR are involved in multiple diseases and the underlying mechanisms. We also discuss the potential therapeutic drugs currently available to regulate gut microbes and mTOR signaling pathway indifferent disorders.
2. Mechanistic target of rapamycin (mTOR)
2.1. mTORC1 and mTORC2 structure
mTOR is a member of the phosphoinositide 3-kinase(PI3K)-related protein kinase (PIKK) family, which has serine/threonine protein kinase activity [9,10]. In mammals, mTOR combines with different subunits to form two macromolecular complexes, mTOR complex 1 (mTORC1) and 2 (mTORC2). Some subunits exist in both complexes, while others are specific components in only one complex [5].
mTORC1 is composed of mTOR, mammalian lethal with SEC13 protein 8 (mLST8) and regulatory-associated protein of mTOR (RAPTOR) as core components, forming a dimeric structure [11,12,13]. RAPTOR is the defining subunit of mTORC1, which is responsible for recruiting substrates and its subcellular localization [14]. Proteins proline-rich AKT substrate of 40 kDa (PRAS40), DEP domain-containing mTOR-interacting protein (DEPTOR), and the Tti1/Tel2 complex are the remaining component of mTORC1 [1]. Specially, both PRAS40 and DEPTOR are endogenous repressor proteins of mTORC1 [15,16].
The mTORC2-specific combinatorial proteins include rapamycin-insensitive companion of mTOR (RICTOR) [17], protein observed with RICTOR 1/2 (PROTOR 1/2) [18], mammalian stress-activated protein kinase-interaction protein 1 (mSIN1) [19], and some common subunits of mTOR kinase, such as mLST8, DEPTOR, Tel2 and Tti1 with mTORC1 [2]. RICTOR and mSIN1 are the defining subunits of mTORC2. The N-terminal of mSIN1 is embedded in RICTOR and binds to mLST8 to maintain the stability of mTORC2 [20]. Meanwhile, mSIN1 ensures substrate recruitment [21] and plays an important role in plasma membrane localization and inhibition of mTORC2 [22,23].
2.2. Upstream regulation of mTORC1
mTORC1 is more well-characterized of the two complexes. It integrates multiple intracellular and extracellular upstream signals including growth factors, energy inputs, and nutrients. The activation and localization of mTORC1 requires upstream signals of two groups of small G proteins, that is, Ras homolog enriched in the brain (Rheb) and Ras-related GTPases (Rags) [19,20,21]. mTORC1 is recruited from the cytoplasm to the lysosome by Rags and activated by Rheb in the GTP-bound state on the lysosome when the cellular environment contains sufficient cytokines, nutrients, endocrine signals and energy [22].
mTORC1 is regulated by growth factors and other mitogens, and the tuberous sclerosis complexes (TSC)1/2 plays an important role in this process. TSC1/2 acts as a GTPase-activating protein (GAP) on lysosomal Rheb, converting it to an inactive GDP-bound state to inhibit mTORC1 [20,23]. TSC1/2 transmits multiple upstream signals that affect mTORC1. Insulin and insulin-like growth factor 1 (IGF1) phosphorylate TSC2 via the PI3K/AKT pathway, which dissociates TSC from the lysosomal surface and activates mTORC1 [24,25,26]. AKT stimulates phosphorylation of the endogenous repressor of the mTORC1 complex, PRAS40, to promote Rheb-driven mTORC1 activation in a TSC1/2 non-dependent manner [10,27]. Pro-inflammatory cytokines, such as tumor necrosis factor-α (TNF-α), phosphorylate TSC1 via IκB kinase β (IKKβ), leading to TSC1/2 inhibition [28]. Classical Wnt signaling activates mTORC1 by inhibiting glycogen synthase kinase 3 (GSK3), and GSK3 phosphorylates TSC2 by initiating phosphorylation in an AMPK-dependent manner [29]. In addition, TSC is inhibited by regulation of ERK and P90 ribosomal S6 kinase (RSK) [30,31].
mTORC1 is inactive when the environment is in low oxygen and energy,in which the cellular synthetic response is inhibited. When ATP production is poor, AMP-activated protein kinase (AMPK) activation is sensitive to AMP and ADP [32]. AMPK inhibits the mTORC1 complex during energy stress via a twin mechanism: AMPK act directly on mTORC1 to phosphorylate Raptor [33]; it also phosphorylates TSC2 and increase its GAP activity on Rheb, indirectly inhibiting mTORC1 [34]. When oxidative stress is enhanced, thioredoxin 1 (Trx1) in cardiomyocytes interacts directly with mTORC1 to maintain its kinase activity [35]. Hypoxia induces transcriptional regulation of DNA damage response 1 (Redd1) expression to activate TSC1/2 [36,37]. Notably, DNA damage also induces the expression of various p53 target genes, including phosphatase and tensin homolog deleted on chromosome 10,TSC1/2 and AMPK, which deliver signal to mTORC1 [38,39]. However, in cardiomyocytes, DNA-damage-inducible transcript 4-like (DDiT4L) inhibits mTORC1 but activates mTORC2 activity [40]. Additionally, Cellular hypoglycemic conditions inhibit the binding of hexokinase-II (HK-II) to mTORC1 [41]. Other cellular stresses such as amino acid deficiency, hyperosmolarity, and PH stress induce TSC2 recruitment to the lysosome, thereby negatively regulating mTORC1 [42].
mTORC1 can be activated by amino acids, a process that is dependent on the involvement of Rag GTPases. Mammals have four species ranging from Rag-A to Rag-D, while Rag-A or Rag-B form obligate heterodimers with Rag-C or Rag-D, respectively. The active conformation of RAG is related to the loading state of the nucleotide, when in the "on" state, RagA/B binds to GTP and RagC/D binds to GDP. At this point, mTORC1 is recruited into the lysosome by RAG and is then activated by Rheb. In the presence of sufficient nutrients, the Rag in the "on" state interacts with RAPTOR to ensure the activation of mTORC1 [43]. When amino acid is exhausted, Folliculin (FLCN) relocalizes to the lysosome and prevents the exchange of GDP with GTP in Rag-A [44].
mTORC1 perceives amino acid concentrations in lysosomes and envelopes through multiple mechanisms, and these processes require different complexes to deliver amino acid signals to the Rag [45]. Several well-characterized amino acid sensors have been identified. Sestrin2 is a cytoplasmic leucine sensor of the mTORC1 pathway [46]. Under leucine starvation conditions, Sestrin2 binds and inhibits GATOR2; after restoring leucine levels, leucine binds to Sestrin2 and dissociates GATOR2 to activate mTORC1 [47]. Similar to Sestrin2, GTPase-activating protein activity toward Rags-1 (CASTOR1), a cytoplasmic arginine sensor. Arginine binds directly to CASTOR1 and relieves the inhibition of GATOR2 [46]. SLC38A9, a lysosomal arginine sensor, transmits state information of amino acids to mTORC1. SLC38A9 interacts with the Rag GTPase-Ragator complex, which is localized to the lysosome, mediating the transport of leucine-based essential amino acids and activating mTORC1 in an arginine-dependent manner [48,49,50]. S-adenosylmethionine (SAM) sensor upstream of mTORC1(SAMTOR), the upstream sensor of SAM, binds to GATOR1 and negatively regulates mTORC1 when methionine is starved or SAM is at low levels [51]. In addition, vesicular H+-adenosine triphosphate ATPase (v-ATPase) is required for amino acid activation of mTORC1 [52].
2.3. Substrates and functions of mTORC1
Once activated by different inputs, mTORC1 responds by acting on different substrates. It not only promotes anabolic synthesis such as protein, nucleic acid and lipid synthesis, cellular metabolism and energy expenditure, but also inhibits catabolic processes, including autophagy.
Firstly, mTORC1 phosphorylates eukaryotic initiation factor 4E-binding proteins (4E-BPs) and p70 S6 kinase 1 (S6K1) thereby promoting protein synthesis [24]. Phosphorylation of 4E-BP1 releases eukaryotic translation initiation factor 4E (eIF4E) and promotes the formation of its complex, thereby derepressing the translation process. Phosphorylated S6K1 phosphorylates its eponymous target ribosomal protein S6 to participate in controlling the transcriptional process during ribosome genesis [25]. In addition, S6K1 augments protein synthesis through activation of eIF4B and degradation of programmed cell death 4 (PDCD4) [26,27]. Although both are involved in the control of translation, the role of 4E-BP1 is more prominent [3].
Secondly, mTORC1 controls the synthesis of lipids and nucleic acids required for cell proliferation. To a large extent, mTORC1 acts through the transcription factors sterol regulatory element binding protein 1/2 (SREBP1/2), a substrate that controls the expression of multiple lipid genes. Transcription of SREBP1/2 can be regulated by mTORC1 through modulating phosphorylation of lipin 1 or S6K1-dependent manner.mTORC1 inhibition reduces SREBP1/2 expression impairing its function and decreasing lipid synthesis [28,29,30]. Phosphorylation of mTORC1 also increased the expression level of peroxisome proliferator-activated receptor γ (PPAR-γ) to promote adipogenesis [31,32]. In addition, mTORC1 promotes lipid biosynthesis by regulating SR protein kinase 2 (SRPK2) to stabilize lipid biosynthetic enzymes [33].
mTORC1 controls the synthesis of new pyrimidines and purines in different cellular models to enrich the nucleotide pool for nucleic acid synthesis, which is essential to maintain DNA replication and RNA synthesis. De novo synthesis of pyrimidines of mTORC1 is mediated by S6K1 promoting the enzymatic activities of carbamoyl-phosphate synthetase 2, and aspartate transcarbamoylase, dihydroorotase (CAD) [34,35]. mTORC1 stimulates activating transcription factor 4 (ATF4) and mitochondrial tetrahydrofolate cycle enzyme methylenetetrahydrofolate dehydrogenase 2 (MTHFD2) to promote purine synthesis [36].
Moreover, mTORC1 also actively regulates cellular metabolism and ATP production to. mTORC1 increases the expression of glycolytic enzymes by activating the transcription and translation of hypoxia-inducible factor 1α (HIF1α), while the activation of SREBPsenhances the pentose phosphate pathway [29]. It has been shown that mitochondrial biogenesis is regulated by mTORC1 through promoting the binding action of transcription factor yin–yang 1 (YY1) with PPARγ coactivator-1a (PGC1-a) [37]. mTORC1 enhances mitochondrial activity through 4E-BP to increase ATP production [38].
In addition to its positive effects on anabolism, mTORC1 negatively regulates catabolism, especially autophagy, to promote growth. Autophagy is the process of degradation of intracellular components, which is considered essential for the organism's starvation response as well as for organelle renewal. Unc-51-like autophagy-activating kinase 1 (ULK1) and ATG13 form in complex with 200 kDa FAK family interacting protein (FIP200) and ATG101 to promote the autophagy [39]. mTORC1 phosphorylates and inhibits ULK1 and A TG13 to negatively regulate autophagy [40,41,42]. mTORC1 can also bind and phosphorylate UVRAG thereby enhancing the antagonistic effects of autophagic vesicle maturation in a nutrient-rich environment [43]. When mTORC1 is repressed, nuclear translocation of transcription factor EB (TFEB) and transcription factor E3 (TFE3) are promoted, facilitating the expression of multiple genes associated with autophagy and lysosomes [44,45,46,47]. Recent studies have shown that mTORC1 directly phosphorylates VAMP8 blocking STX17-SNAP29-VAMP8 SNARE complex formation and inhibiting autophagosome-lysosome fusion [48]. mTORC1 may also affect autophagy by regulating effectors such as death-associated protein 1 (DAP1) [49] and WIPI2 [50].
2.4. Upstream regulation of mTORC2
mTORC2 is activated by growth factors such as insulin through PI3K. The PH domain in mSIN1 can combine with phosphatidylinositol 3,4,5-trisphosphate (PIP3), upon PI3K activation, and result in mTORC2 activation [23]. Stimulated by PI3K signaling, the ribosome binds to mTORC2, which is essential for mTORC2 activation [51]. mTORC2 activation also requires the interaction with TSC1/2 independent on Rheb, but the mechanism remains unclear [52].
mTORC2, in addition to being regulated by PI3K/AKT, is also regulated through the mTORC1 negative feedback loop. Degradation of IRS1 induced by mTORC1 and its effector S6K inhibits insulin/PI3K/AKT signaling [53,54,55]. Growth factor receptor-bound protein 10 (Grb10), which negatively regulates insulin/IGF-1 signaling, is activated by mTORC1 and mediates the inhibition of PI3K [56,57]. In addition, SK6 promotes RICTOR and mSIN1 activity, respectively, thereby inhibiting mTORC2 [58,59]. In contrast, AMP-activated protein kinase (AMPK) directly phosphorylates the complex of mTORC2 and RICTOR and promotes AKT signaling, a process that is independent on mTORC1-mediated negative feedback [60].
Compared with mTORC1, the cellular localization of mTORC2 is more diverse and can localize on the plasma membrane, endoplasmic reticulum, mitochondria, and mitochondria-associated ER membranes (MAM) [61]. However, it remains to be unrevealed how mTORC2 is stimulated at these sites.
2.5. Substrates and functions of mTORC2
mTORC2 regulates a variety of important cellular functions,such as cell structure, metabolism, survival, and proliferation, by regulating members of the AGC family including AKT, serum, glucocorticoid-induced protein kinase 1 (SGK1) and protein kinase C-α(PKC-α). mTORC2 can phosphorylate its terminal hydrophobic motif Ser473 to directly activate AKT [62]. Deletion of AKT-Ser473 phosphorylation impaired only forkhead box O1/3a (FoxO1/3a) and did not affect TSC2 and GSK3-β [63,64]. mTORC2 activates SGK1 and its substrate N-myc downstream regulated gene 1 (NDRG1) to promote cell survival in hypoxic conditions, which is similar to the effect of FOXO3a [65,66,67].In addition, mTORC2 mediates cell survival by inhibiting mammalian sterile 20-like kinase (MST1) of the hippo pathway [68,69]. PKC-α is the third AGC kinase activated by mTORC2, which is involved in the regulation of cytoskeleton and structure together with RHO GTPases [17,70].
3. Gut microbiota
Trillions of microorganisms have adapted to inhabit the human intestine and form a complex ecological community, which is an indispensable part of the human body [71]. The microbiota obtained at birth developed synchronously with the development of host, and maintains its stability and diversity after adulthood [72]. Because microorganisms are distributed in different regions with the host, diversity is determined by the local environment including diet [73], high altitude and other extreme weather [74,75].
The gastrointestinal tract (GI) is composed of stomach, small intestine(SI) and large intestine(LI).Specific microbiota reside in the unique microenvironment of each section [72]. Five categories of microbiota reside in the stomach of healthy people, including Firmicutes, Bacteroidetes, Actinobacteria, Fusobacteria and Proteobacteria [76]. Gram-positive bacteria and facultative anaerobic bacteria are mainly found in the SI, such as Lactobacilli, Enterococci and Streptococci, which is benefical to food digestion and nutrition absorption [77]. Firmicutes and Bacteroidetes, mainly settle in the large intestine [78].
In addition to the number, species and composition of the gut microbes, the metabolites of the microbes have attracted more attention these years. Undigested carbohydrates in the SI are fermented by microorganisms in the LI to produce single chain fatty acids (SCFAs), namely acetate, propionate and butyrate.
Gut microbiota contribute to the defens of intestinal pathogens, nutrition and energy absotption from the diet, and maintainace of normal immune function [79]. Disregulation of the normal balance between gut microbiota and host is involved in many disorders, such as obesity [80], aging [81], inflammatory bowel disease [82], neurological diseases [83], and the occurrence and development of tumors [84]. Recent studies have found that intestinal microorganisms are more closely related to mTOR signaling pathway (as shown in Figure 1).
3.1. Gut microbiota, mTOR and Intestinal Diseases
Inflammatory bowel disease (IBD), including ulcerative colitis (UC) and Crohn's disease (CD), is a chronic recurrent intestinal inflammatory disease mediated by immunity. The most common symptoms of these diseases are abdominal pain, diarrhea and weight loss, and the incidence rate is increasing worldwide [85,86,87]. Intestinal inflammation caused by western diet is accompanied by the activation of mTOR, which is contingent on microbial derived pathogen-associated molecular patterns, in intestinal epithelial cells (IEC), such as endotoxin, poly (I: C) and flagellin [88]. It is worth noting that IL-33 can activate the mTOR pathway of intestinal epithelial cells and promote the induction of REG3γ to faciliate the symbiosis of intestinal microflora [89]. Short chain fatty acids (SCFAs), a metabolite of intestinal microorganisms, can promote the production of IL-22 by CD4+ T cells and IEC. In particular, butyrate can accelerate the phosphorylation of STAT3 and mTOR to up regulate the expression of IL-22 to maintain the stability of the intestinal environment [90].In addition, extra-intestinal activation of microbiota-specific CD4+ T cells and concomitant inhibition of mTOR metabolism can remove CD4+ T memory (TM) cells thereby preventing colitis [91].
Chronic inflammation is a characteristic of IBD and the driving force of human colon cancer. Studies have shown that the microbial composition of mouse colon induced by dextran sodium sulfate (DSS) and/or azomethane (AOM) is significantly different from that of the control group. It is worth noting that the abundance of Clostridium and Staphylococcus aureus is increased. Meanwhile, PI3K/AKT/mTOR, fatty acid metabolism and oxidative phosphorylation signals were found to be upregulated in DSS / AOM mice by gene expression profiling [92]. Clostridium butyricum promotes the upregulation of AKT/mTOR and downstream molecule p70 ribosomal protein S6 kinase expression and alters the production of anti-inflammatory cytokines to exert a protective effect on intestinal barrier function [93]. In contrast, Lactobacillus X12 inhibited the growth of colorectal cancer cells by inhibiting mTOR and regulating cell cycle-associated proteins p27 and E1 [94]. In the genesis and development of colon cancer, the imbalance of gut microbiota is accompanied by the release of endotoxin lipopolysaccharide (LPS) [95]. With the increase of LPS level, Cathepsin K (CTSK), a metastasis-related secretory protein, secreted by tumor can interact with macrophage membrane receptor Toll-like receptor 4 (TLR4) to activate mTOR pathway. The TLR4-mTOR-dependent pathway accelerates the M2 polarization of tumor associated macrophages to promote the progression of colorectal cancer [96].
3.2. Gut microbiota, mTOR and Liver Diseases
The gut microbiota and mTOR signaling pathways have been shown to play an important role in liver diseases [97,98,99]. Inoculation of mice treated with high-fat diet with feces from patients with nonalcoholic fatty liver disease (NAFLD) result in aggravated liver injury [100]. Recently, study has shown that NF73-1, an E. coli isolated from the intestines of NASH patients, can enter the liver through TLR2/NLRP3, induce M1 macrophages, and finally promote the development of NAFLD,with the activation of mTOR-S6K1-SREBP-1/PPAR-α [101].In fact, the abundance of microbiota is low and diversity is less in mice with NAFLD. Interestingly, Alisol B 23 acetate (AB23A) could rebalance the gut microbiota, especially reducing the abundance of Firmicutes/Bacteroidaeota and Actinobacteriota/Bacteroidaeota. During this process, AB23A plays the role of probiotics on NAFLD by inhibiting activity of mTOR, TLR4 and NF-κB [102]. Similarly, Lactobacillus rhamnosus, which is considered to be one of the most extensive probiotic, has also been revealed in the role of alcoholic fatty liver. The combination therapy of Lactobacillus rhamnosus culture supernatant and bone marrow mesenchymal stem cells (BMMSCs) can accelerate autophagy and improve alcoholic fatty liver disease by inhibiting PI3K/mTOR pathway [103].In hepatocellular carcinoma cell line Huh-7, Pant et al. found that butyrate, a short chain fatty acid produced by gut microbiota during anaerobic fermentation, inhibited the phosphorylation of AKT and mTOR by inducing reactive oxygen species (ROS), thereby inducing autophagy of liver cancer cells [104]. In general, the disregulation of the gut microbiota and mTOR signal will promote the progression of liver diseases, in which autophagy and immune cells are also involved.
3.3. Gut microbiota, mTOR and Heart disease
The gut microbes and mTOR signaling pathways are both involoved in heart disease. Butyrate, a metabolite of gut microbiota, significantly inhibits PI3K/AKT/mTOR pathway while enhancing ATG5 mediated autophagy in the murine STC-1 enteroendocrine cell line [105]. When mTOR is activated by PI3K and AKT, it can inhibit autophagy by regulating the activity of ULK1 [106]. Urolithin B(UB), a metabolite of gut microbiota, can inhibit autophagy through AKT/mTOR/ULK1 pathway to play a protective role in myocardial ischemia-reperfusion injury in rats. More specifically, the p62/Keap1/Nrf2 signaling pathway protects against oxidative stress and caspase 3-dependent apoptosis [107]. Similarly, this protective effect was also confirmed in the mouse model of myocardial infarction (MI). UB inhibits cardiomyocyte apoptosis by activating AKT/mTOR pathway and simultaneously suppresses NF-κB to reduce the occurrence of arrhythmia after hypoxia [108].
3.4. Gut microbiota, mTOR and Other Diseases
The interaction of gut microbiota and mTOR is also reported in other diseases, in which autophagy, energy metabolism and immunity are involved.
AMP activated protein kinase (AMPK), a key energy receptor, can inhibit the formation of auto phagosomes caused by mTOR [42]. Sodium butyrate can activate the phosphorylation of AMPK in the renal tissue of diabetic mellitus (DM) rats to inhibit mTOR and increase the number of auto phagosomes, thus aggravate the kidney injury of DM rats [109]. In the rat model of diet induced obesity (DIO), Xiexin Tang (XXT) promotes the production of SCFAs by intestinal flora and the expression of AMPK to inhibit the activation of mTOR signal pathway, to adjust the disorder of lipid metabolism and reduce systemic inflammatory response [110]. In addition, both IL-37 and alanylglutamine can increase the expression of AMPK and reduce the expression of mTOR to alleviate chronic inflammation in the mouse models of allergic asthma and atopic dermatitis (AD), respectively. In this process, the diversity and metabolites of intestinal microorganisms are regulated. The difference is that IL-37 increases autophagy related IC3B, decreases autophagy related ubiquitinated protein p62, and the NF-κB and STAT3 signaling pathways may be involoved in the treatment of alanylglutamine [111,112].A recent study showed that environmental low-dose radiation (LDR) compromises the intestinal barrier and increases PA in the organism. This is accompanied by increased expression of PYCR1 in the liver and inhibition of the IRS-1/AKT/mTOR axis, and an accompanying increase in Parabacteria in the gut microbiota, leading to impairment of HFD-induced obesity and insulin resistance [113]. In the mouse models of high-fat diet, simultaneous inhibition of mTORC1 and mTORC2 could aggravate intestinal inflammation and destroy blood glucose homeostasis, while specific inhibition of mTORC1 could alleviate intestinal inflammation and improve glucose tolerance. Interestingly, the chronic inhibition of mTORC2 contributed to the changes of gut microbiota caused by high fat, such as Turicibacter and unclassified Marinilabiliaceae [114]. Long term inhibition of mTOR can prolong the life span of mice and mildly change intestinal metagenes, which are related to immune cells [115]. Moreover, Prevotella Copri obtained from pig intestines can promote chronic inflammatory response and fat deposition through TLR4 and mTOR signaling pathways after its inoculation in mice [116].
3.5. Gut microbes and mTOR in the analysis of big data
The human intestinal microbes are so numerous and diverse that they have been called the "second genome". The microbial census began with 16s rRNA gene sequencing, and PCR is applied to amplify 16s rRNA and then two to three times sanger sequencing is utilized to complete the gene sequencing. In order to make the sequencing more accurate, second-generation sequencing and third-generation technologies were developed to make the study of microbiota genomics more efficient and rapid [117].
In order to understand the impact of gut microbes on human health, researchers analyzed 124 human-derived feces by macro genome sequencing to create a genomic catalog of the human gut microbiome, covering most of the prevalent gut microbial genes in humans [118]. More studies are delving into the role of microbiota in different health and disease conditions through transcriptomic and metabolomics. Analysis of microorganisms in feces by 16S rRNA sequencing revealed a relative decrease in bacterial abundance in mice with colorectal cancer [119]. In addition, analysis of colonic tissues by whole transcriptome profiling revealed that enhanced PI3K-AKT-mTOR signaling in colorectal cancer mice was accompanied by a significant increase in the expression level of phosphorylated S6 ribosomal protein (a downstream target of the mTOR pathway) [92]. In 129 stool samples from NAFLD patients consuming high carbohydrate (HC), alterations in intestinal microbial species were found, with a significant increase in the abundance of Enterobacteriaceae and a significant decrease in the abundance of Ruminococcaceae compared to 75 normal samples. Meanwhile, analysis of 90 liver transcripts revealed that the expression of SREBF2 and mTOR increased with the enhancement of NAFLD activity [120].
4. Application of multiple drugs affecing gut microbes and mTOR in the treatment of different diseases
In recent years, further studies on, the direct or indirect interaction between gut microbiota and mTOR in various diseases have been revealed. The significance of a variety of physiological activities in the body, including metabolic response, autophagy, immune response and so on, is also shown above. Many kinds of drugs have demanstrated promising evidence in preclinical studies in different diseases as shown in Table 1. More effective treatment methods need further evaluation one effectiveness and safety in clinical trials.
4.1. Treatment of Intestinal diseases
Mu Xia Li et al. revealed the mechanism of Huangqin decoction (HQD) in the treatment of gastrointestinal diseases such as UC. As a traditional Chinese medicine therapy, HQD could improve the clinical performance of DSS induced UC model, inhibit the inflammatory response in vivo and rebalance the gut microbiota. HQD treatment activated PI3K/AKT/mTOR signaling by up regulating amino acid metabolism, andimproved the barrier function of intestinal epithelial [121,122]. Dandan Wang et al. found that a polysaccharide was isolated from Panax ginseng (GP) reduced the intestinal injury of DSS induced colitis in rats. GP treatment increased microbial community diversity, improved the compositions of gut microbiota,reduced the phosphorylation level of mTOR and activated autophagy to inhibit inflammation [122]. In addition, SCFAs and metformin (MTF) can regulate intestinal immunity to prevent colitis, and have potential therapeutic applications [90,123]. Theabrownin (TB) inhibited the development of CRC by decreasing Bacteroidceae and Bacteroides associated with CRC and increasing the production of SCFAs, thereby inhibiting cell proliferation through suppression of PI3K/AKT/mTOR phosphorylation [5].
4.2. Treatment of Liver Diseases
In the HFD-induced rat model of NAFLD, fecal levels of Firmicutes and Bacteroidetes and short chain fatty acids returned to normal in the treatment with L. reuteri + MTZ alone or in combination with MTF More precisely, combined therapy prevented steatosis and the progression of liver injury by inducing autophagy by p-AKT/mTOR/LC-3Ⅱ pathways in the liver [124]. Fan Xia et al. found that AB23A not only reduced the abundance of Firmicutes/Bacteroidaeota and Actinobacteriota /Bacteroidaeota, but also decreased the activities of mTOR and TLR4 to prevent the progress of NAFLD [102]. Besides, the combined LGG-s and BMMSC treatment also inhibited PI3K/mTOR signal to accelerate autophagy, which has the potential to alleviate alcoholic steatohepatitis [125].Interestingly,in the HFD-induced metabolic syndrome, Zhenzhen Deng et al. found that low molecular weight fucoidan fraction LF2 and MTF have similar effects on gut microbiota, increasing the proportion of Verrucomicrobia and enriching the abundance of Akkermansia muciniphila. LF2 promoted the phosphorylation of PI3K and AKT in a dose-dependent manner, but reversed the over activation of mTOR, thereby improving lipid metabolism [126].
According to the report, the gut microbiota can regulate the immune response of hepatocellular carcinoma (HCC), thus readjust the gut microbiota could be a potential option for HCC treatment [127]. Butyrate, considered as a potential candidate drug for the treatment of liver cancer, could inhibit the phosphorylation of AKT and mTOR through reactive oxygen species, resulting in the up regulation of autophagy proteins beclin 1, ATG 5, LC3-Ⅱ, thereby promoting the formation of autophagy bodies [104]. Curcumin can significantly sensitize hepatoma cells to 5-FU cytotoxicity and increased the apoptosis rate through synergistic effects. The gut microbiota facilitates oral utilization of curcumin in vivo and enhances chemo-sensitivity of hepatocellular carcinoma cells to 5-FU by blocking PI3K/AKT/mTOR signaling pathway in vitro [128].
4.3. Treatment of Other Diseases
Probiotics regulate PI3K/AKT/mTOR signaling pathway, which is beneficial to coordinate immune response. Probiotics fermentation technology (PFT) activated PI3K/AKT signal transduction pathway but inhibited glycogen synthase kinase-3β (GSK-3β) and mTOR signal, its potential role in the treatment of Alzheimer's disease was parallel to that of pioglitazone [129]. Additionally, there are reports that Aronia melanocarpa polysaccharide (AMP) activated PI3K/AKT/mTOR signaling pathway and its downstream apoptotic protein family, inhibitd brain cell apoptosis and enriched intestinal beneficial bacteria to delay aging, which had similar function to MogrosideV and its metabolite 11-oxo-mogrol [130,131]. In contrast to the mechanism of action of AMP, Xiexin Tang (XXT) ameliorates obesity by promoting the activity of key enzymes for SCFAs synthesis and inhibiting AMPK while activating mTOR signaling [110].
Ophiopogonin D (OPD) could increase the abundance of Bacteroidetes, reduce the relative abundance of Firmiuts, inhibit the phosphorylation of mTOR and the expression of SREBP1 and SCD1 to alleviate fat metabolism, and result in to prevention of atherosclerosis and metabolic syndrome [132]. The mechanism that β-hydroxyβ-methylbutyrate(HMB) functions through Bacteroidetes-acetic acid-AMPKα axis to reduce the lipid metabolism of Bama Xiang mini-pigs was somewhat similar [133]. By the way, Flammulina velutipes polysaccharide (FVP) affected the abundance of gut microbiota, especially Bacteroidetes phylum and Muribaculaceae family, and up regulated mTOR signal pathway in cardiac tissue [134]. However, the specific mechanism remains to be determined.
Oral administration of bruceae Frutus oil (Bo) under the influence of gut microbiota inhibited breast cancer. At the same time, Bo changed the dominant strains of gut microbiota and promoted mTOR activity, leading to the inhibition of autophagy [135]. In contrast, 20(s)-ginsenoside Rh2 (grh2) played an anti-tumor role by inhibiting PI3K/AKT/mTOR signal [136]. Both Engineered resistant-starch (ERS) diet and ketogenic diet (KD) reduced mTOR phosphorylation and regulated microorganisms [137,138]. Diet may serve as a synergistic approach to improve the treatment of diseases.
The direct interaction between mTOR and intestinal microorganisms provides potential ideas for treatment. Firstly, microencapsulated rapamycin (eRapa), the best pharmacological mTOR inhibitor studied in the study of life span and health extension, had strong immune effects and can gently change intestinal metagenes, which was worthy of further study [139]. Then, resveratrol, a specific inhibitor of mTOR complex 1, alleviated the changes of intestinal microflora in diet induced obese mice [114]. Furthermore, The microflora metabolite SCFAs activated mTOR and STAT3 of IEC to produce antimicrobial peptides to balance the intestinal environment [140].
Overall, in view of the limitations of current treatment, more drugs can only be used as a potential choice for disease treatment, which has broad clinical application prospects in the future.
5. Conclusions and Future Perspective
mTOR signal pathway plays a significant role in various physiological processes in the cell. The gut microbiota and its complex metabolites regulate a wide range of host functions, including autophagy, fatty acid metabolism, oxidative phosphorylation, and immune response. Dysregulation of the gut microbiota is associated with various disorders, such as cancers, diabetes, and inflammatory diseases, in which the mTOR signal pathway is also involoved.. In the present review, we introduced mTOR signal pathway, the correlation between gut microbes and mTOR, their function the mechanisms in multiple diseases (the crosstalk between the two is shown in Figure 1), and the possible treatments utilizing microbiota and mTOR inhibitors in these diseases.
Currently, there are still unsolved problems remain to be elucidated regarding the study of the gut microbiota and mTOR. Most existing studies suggest a phenomenal association between gut microbes and mTOR, with disease development causing composition changes in the gut microbiota or regulation of its metabolites, accompanied by inhibition or activation of mTOR.
The translation of basic mTOR and microbiota studies to the clinical setting remains challenging. Most studies focus on the evidence in animal models, few is known about mTOR in clinical trials. More data needs to be verified in clinical conditions. As inhibition of mTOR does not always lead to protective effects, it may also be controversial. Furthermore, due to the complexity of the organism, mTOR activity is increased in some pathological conditions and decreased by others, making it difficult to estimate the exact effects of therapeutic intervention. More in-depth systematic studies are in great need to elucidate how mTOR is associated with gut microbes, to produce more conclusive and valid results, and to further explore new drug candidates for the treatment of relevant diseases.
Author Contributions
Writing—original draft preparation, Y.G. and T.T.; writing—review and editing, Y.G. and T.T.; supervision and funding acquisition, T.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Natural Science Foundation of Beijing (grant number 5222019).
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| 4E-BPs | 4E-binding proteins |
| AB23A | Alisol B 23-acetate |
| AD | Alzheimer’s disease |
| AMP | Aronia melanocarpa polysaccharide |
| AMPK | AMP-activated protein kinase |
| AMPK | AMP-activated protein kinase |
| AOM | Azomethane |
| ASD | Autism spectrum disorder |
| ATF4 | Activating transcription factor 4 |
| BMMSCs | Bone marrow mesenchymal stem cells |
| BMMSCs | Bone marrow mesenchymal stem cells |
| BO | Bruceae fructus oil |
| CASTOR1 | GTPase-activating protein activity toward Rags-1 |
| CD | Crohn's disease |
| CTSK | Cathepsin K |
| DAP1 | Death-associated protein 1 |
| DDiT4L | DNA-damage-inducible transcript 4-like |
| DEPTOR | DEP domain-containing mTOR-interacting protein |
| DSS | Dextran sodium sulfate |
| eIF4E | Eukaryotic translation initiation factor 4E |
| ERS | Engineered Resistant-Starch |
| FLCN | Folliculin |
| FoxO1/3a | Forkhead box O1/3a |
| FVP | Flammulina velutipes polysaccharide |
| GAP | GTPase-activating protein |
| GI | Gastrointestinal tract |
| Grb10 | Growth factor receptor-bound protein 10 |
| GSK3 | Glycogen synthase kinase 3 |
| HCC | Hepatocellular Carcinoma |
| HK-II | Hexokinase-II |
| HMB | β-hydroxy-β-methylbutyrate |
| HQD | Huangqin decoction |
| IBD | Inflammatory bowel disease |
| IEC | Intestinal epithelial cells |
| IGF1 | Insulin and insulin-like growth factor 1 |
| IKKβ | IκB kinase β |
| L. reuteri | Lactobacillus reuteri DSM 17938 |
| LGG-s | Lactobacillus rhamnosus culture supernatant |
| LI | Large intestine |
| LPS | Lipopolysaccharide |
| METS | Metabolic syndrome |
| mLST8 | Mammalian lethal with SEC13 protein 8 |
| MogV | MogrosideV |
| mSIN1 | Mammalian stress-activated protein kinase interacting protein 1 |
| MST1 | Mammalian sterile 20-like kinase |
| MTF | Metformin |
| MTHFD2 | Methylenetetrahydrofolate dehydrogenase 2 |
| mTOR | The mammalian or mechanistic target of rapamycin |
| mTORC | mTOR complex |
| NAFLD | Nonalcoholic fatty liver disease |
| NDRG1 | N-myc downstream regulated gene 1 |
| NKB | Natural killer B cell |
| OPD | Ophiopogonin D |
| P. ginseng | Polysaccharides from Panax ginseng C. A. Meyer |
| PC | Pancreatic cancer |
| PDCD4 | Programmed cell death 4 |
| PFT | Probiotics fermentation technology |
| PGC-1α | Proliferator-activated receptor gamma coactivator 1-alpha |
| PI3K | Phosphoinositide 3-kinase |
| PIKK | Phosphoinositide 3-kinase related protein kinase |
| PKC-α | Protein kinase C-α |
| PPAR-γ | Peroxisome proliferator-activated receptor γ |
| PRAS40 | Proline-rich substrate of 40 kDa |
| PROTOR1/2 | Protein observed with RICTOR 1/2 |
| Rags | Ras-related GTPases |
| RAPTOR | Regulatory-associated protein of mTOR |
| Redd1 | Regulation of DNA damage response 1 |
| Rheb | Ras homolog enriched in the brain |
| ROS | Reactive oxygen species |
| RSK | Ribosomal S6 kinase |
| S6K1 | S6 kinase 1 |
| SAM | S-adenosylmethionine |
| SCFAs | Short chain fatty acids |
| SGK1 | Serum and glucocorticoid-induced protein kinase 1 |
| SI | Small intestine |
| SREBP1/2 | Sterol regulatory element binding protein 1/2 |
| SRPK2 | SR protein kinase 2 |
| STZ | Streptozotocin |
| T-ALL | T-cell acute lymphoblastic leukemia |
| TB | Theabrownin |
| TFH | Follicular helper T cell |
| TLR4 | Toll-like receptor 4 |
| TNBC | Triple-negative breast cancer |
| TNF-α | Tumor necrosis factor-α |
| Trx1 | Thioredoxin 1 |
| TSC | Tuberous sclerosis complexes |
| UC | Ulcerative colitis |
| UCP-2 | Uncoupling protein-2 |
| ULK1 | Unc-51-like autophagy-activating kinase 1 |
| XXT | Xiexin Tang |
References
- Laplante, M.; Sabatini, D. M. , mTOR signaling in growth control and disease. Cell 2012, 149, 274–293. [Google Scholar] [CrossRef]
- Saxton, R. A.; Sabatini, D. M. , mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef] [PubMed]
- Liu, G. Y.; Sabatini, D. M. , mTOR at the nexus of nutrition, growth, ageing and disease. Nat Rev Mol Cell Biol 2020, 21, 183–203. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Li, X.; Zhang, J. , mTOR Signaling in Cancer and mTOR Inhibitors in Solid Tumor Targeting Therapy. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Sciarretta, S.; Forte, M.; Frati, G.; Sadoshima, J. , The complex network of mTOR signalling in the heart. Cardiovasc Res 2022, 118, 424–439. [Google Scholar] [CrossRef]
- Ruth, E. Ley, P. J. T., Samuel Klein, Jeffrey I. Gordon, Human gut microbes associated with obesity. NATURE 2006, 444. [Google Scholar]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; Peng, Y.; Zhang, D.; Jie, Z.; Wu, W.; Qin, Y.; Xue, W.; Li, J.; Han, L.; Lu, D.; Wu, P.; Dai, Y.; Sun, X.; Li, Z.; Tang, A.; Zhong, S.; Li, X.; Chen, W.; Xu, R.; Wang, M.; Feng, Q.; Gong, M.; Yu, J.; Zhang, Y.; Zhang, M.; Hansen, T.; Sanchez, G.; Raes, J.; Falony, G.; Okuda, S.; Almeida, M.; LeChatelier, E.; Renault, P.; Pons, N.; Batto, J. M.; Zhang, Z.; Chen, H.; Yang, R.; Zheng, W.; Li, S.; Yang, H.; Wang, J.; Ehrlich, S. D.; Nielsen, R.; Pedersen, O.; Kristiansen, K.; Wang, J. , A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef]
- Hu, X.; Wang, T.; Jin, F. , Alzheimer's disease and gut microbiota. Sci China Life Sci 2016, 59, 1006–1023. [Google Scholar] [CrossRef]
- Stephen, B. Helliwell, P. W., Jeannette Kunz, Maja Deuter-Reinhard, Ruben Henriquez, and Michael N. Hall, TOR1 and TOR2 Are Structurally and Functionally Similar but not Identical Phosphatidylinositol Kinase Homologues in Yeast. Molecular biology of the cell 1994, 5, 105–118. [Google Scholar]
- Schreiber, C. T. K. a. S. L. , PIK-Related Kinases: DNA Repair, Recombination, and Cell Cycle Checkpoints. Science 1995, 270, 50–50. [Google Scholar]
- Loewith, R.; Jacinto, E.; Wullschleger, S.; Lorberg, A.; Crespo, J. L.; Bonenfant, D.; Oppliger, W.; Jenoe, P.; Hall, M. N. , Two TOR Complexes, Only One of which Is Rapamycin Sensitive, Have Distinct Roles in Cell Growth Control. Mol Cell 2002, 10, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Rudge, D. G.; Koos, J. D.; Vaidialingam, B.; Yang, H. J.; Pavletich, N. P. , mTOR kinase structure, mechanism and regulation. Nature 2013, 497, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Baretic, D.; Berndt, A.; Ohashi, Y.; Johnson, C. M.; Williams, R. L. , Tor forms a dimer through an N-terminal helical solenoid with a complex topology. Nat Commun 2016, 7, 11016. [Google Scholar] [CrossRef] [PubMed]
- Chao, L. H.; Avruch, J. , Cryo-EM insight into the structure of MTOR complex 1 and its interactions with Rheb and substrates. F1000Res 2019, 8. [Google Scholar] [CrossRef]
- Peterson, T. R.; Laplante, M.; Thoreen, C. C.; Sancak, Y.; Kang, S. A.; Kuehl, W. M.; Gray, N. S.; Sabatini, D. M. , DEPTOR is an mTOR inhibitor frequently overexpressed in multiple myeloma cells and required for their survival. Cell 2009, 137, 873–886. [Google Scholar] [CrossRef]
- Sancak, Y.; Thoreen, C. C.; Peterson, T. R.; Lindquist, R. A.; Kang, S. A.; Spooner, E.; Carr, S. A.; Sabatini, D. M. , PRAS40 is an insulin-regulated inhibitor of the mTORC1 protein kinase. Mol Cell 2007, 25, 903–915. [Google Scholar] [CrossRef]
- Sarbassov, D. D.; Ali, S. M.; Kim, D. H.; Guertin, D. A.; Latek, R. R.; Erdjument-Bromage, H.; Tempst, P.; Sabatini, D. M. , Rictor, a novel binding partner of mTOR, defines a rapamycin-insensitive and raptor-independent pathway that regulates the cytoskeleton. Curr Biol 2004, 14, 1296–1302. [Google Scholar] [CrossRef]
- Pearce, L. R.; Sommer, E. M.; Sakamoto, K.; Wullschleger, S.; Alessi, D. R. , Protor-1 is required for efficient mTORC2-mediated activation of SGK1 in the kidney. Biochem J 2011, 436, 169–179. [Google Scholar] [CrossRef]
- Frias, M. A.; Thoreen, C. C.; Jaffe, J. D.; Schroder, W.; Sculley, T.; Carr, S. A.; Sabatini, D. M. , mSin1 is necessary for Akt/PKB phosphorylation, and its isoforms define three distinct mTORC2s. Curr Biol 2006, 16, 1865–1870. [Google Scholar] [CrossRef]
- Scaiola, A.; Mangia, F.; Imseng, S.; Boehringer, D.; Berneiser, K.; Shimobayashi, M.; Stuttfeld, E.; Hall, M. N.; Ban, N.; Maier, T. , The 3.2-Å resolution structure of human mTORC2. Sci Adv 2020, 6, 1–10. [Google Scholar] [CrossRef]
- Tatebe, H.; Murayama, S.; Yonekura, T.; Hatano, T.; Richter, D.; Furuya, T.; Kataoka, S.; Furuita, K.; Kojima, C.; Shiozaki, K. , Substrate specificity of TOR complex 2 is determined by a ubiquitin-fold domain of the Sin1 subunit. Elife 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Berchtold, D.; Walther, T. C. , TORC2 plasma membrane localization is essential for cell viability and restricted to a distinct domain. Mol Biol Cell 2009, 20, 1565–1575. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Gan, W.; Chin, Y. R.; Ogura, K.; Guo, J.; Zhang, J.; Wang, B.; Blenis, J.; Cantley, L. C.; Toker, A.; Su, B.; Wei, W. , PtdIns(3,4,5)P3-Dependent Activation of the mTORC2 Kinase Complex. Cancer Discov 2015, 5, 1194–1209. [Google Scholar] [CrossRef] [PubMed]
- Ma, X. M.; Blenis, J. , Molecular mechanisms of mTOR-mediated translational control. Nat Rev Mol Cell Biol 2009, 10, 307–318. [Google Scholar] [CrossRef]
- Chauvin, C.; Koka, V.; Nouschi, A.; Mieulet, V.; Hoareau-Aveilla, C.; Dreazen, A.; Cagnard, N.; Carpentier, W.; Kiss, T.; Meyuhas, O.; Pende, M. , Ribosomal protein S6 kinase activity controls the ribosome biogenesis transcriptional program. Oncogene 2014, 33, 474–483. [Google Scholar] [CrossRef]
- Holz, M. K.; Ballif, B. A.; Gygi, S. P.; Blenis, J. , mTOR and S6K1 mediate assembly of the translation preinitiation complex through dynamic protein interchange and ordered phosphorylation events. Cell 2005, 123, 569–580. [Google Scholar] [CrossRef]
- Dorrello, N. V.; Peschiaroli, A.; Guardavaccaro, D.; Colburn, N. H.; Sherman, N. E.; Pagano, M. , S6K1- and betaTRCP-mediated degradation of PDCD4 promotes protein translation and cell growth. Science 2006, 314, 467–471. [Google Scholar] [CrossRef]
- Porstmann, T.; Santos, C. R.; Griffiths, B.; Cully, M.; Wu, M.; Leevers, S.; Griffiths, J. R.; Chung, Y. L.; Schulze, A. , SREBP activity is regulated by mTORC1 and contributes to Akt-dependent cell growth. Cell Metab 2008, 8, 224–236. [Google Scholar] [CrossRef]
- Duvel, K.; Yecies, J. L.; Menon, S.; Raman, P.; Lipovsky, A. I.; Souza, A. L.; Triantafellow, E.; Ma, Q.; Gorski, R.; Cleaver, S.; Vander Heiden, M. G.; MacKeigan, J. P.; Finan, P. M.; Clish, C. B.; Murphy, L. O.; Manning, B. D. , Activation of a metabolic gene regulatory network downstream of mTOR complex 1. Mol Cell 2010, 39, 171–183. [Google Scholar] [CrossRef]
- Peterson, T. R.; Sengupta, S. S.; Harris, T. E.; Carmack, A. E.; Kang, S. A.; Balderas, E.; Guertin, D. A.; Madden, K. L.; Carpenter, A. E.; Finck, B. N.; Sabatini, D. M. , mTOR complex 1 regulates lipin 1 localization to control the SREBP pathway. Cell 2011, 146, 408–420. [Google Scholar] [CrossRef]
- Chen, J. E. K. a. J. , Regulation of Peroxisome Proliferator–Activated Receptor-γ Activity by Mammalian Target of Rapamycin and Amino Acids in Adipogenesis. Diabetes 2004, 53, 2748–2756. [Google Scholar]
- Zhang, H. H.; Huang, J.; Duvel, K.; Boback, B.; Wu, S.; Squillace, R. M.; Wu, C. L.; Manning, B. D. , Insulin stimulates adipogenesis through the Akt-TSC2-mTORC1 pathway. PLoS One 2009, 4, e6189. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; Zheng, Y.; Cho, S.; Jang, C.; England, C.; Dempsey, J. M.; Yu, Y.; Liu, X.; He, L.; Cavaliere, P. M.; Chavez, A.; Zhang, E.; Isik, M.; Couvillon, A.; Dephoure, N. E.; Blackwell, T. K.; Yu, J. J.; Rabinowitz, J. D.; Cantley, L. C.; Blenis, J. , Post-transcriptional Regulation of De Novo Lipogenesis by mTORC1-S6K1-SRPK2 Signaling. Cell 2017, 171, 1545–1558. [Google Scholar] [CrossRef] [PubMed]
- Aaron, M. Robitaille; Stefan Christen; Mitsugu Shimobayashi; Marion Cornu; Luca L. Fava; Suzette Moes; Cristina Prescianotto-Baschong; Uwe Sauer; Paul Jenoe; Hall, M. N., Quantitative Phosphoproteomics Reveal mTORC1 Activates de Novo Pyrimidine Synthesis. Science 2013, 339, 1320–1323. [Google Scholar]
- Ben-Sahra, I.; Howell, J. J.; Asara, J. M.; Manning, B. D. , Stimulation of de novo pyrimidine synthesis by growth signaling through mTOR and S6K1. Science 2013, 339, 1323–1328. [Google Scholar] [CrossRef]
- Ben-Sahra, I.; Hoxhaj, G.; Ricoult, S. J. H.; Asara, J. M.; Manning, B. D. , mTORC1 induces purine synthesis through control of the mitochondrial tetrahydrofolate cycle. Science 2016, 351, 728–733. [Google Scholar] [CrossRef]
- Cunningham, J. T.; Rodgers, J. T.; Arlow, D. H.; Vazquez, F.; Mootha, V. K.; Puigserver, P. , mTOR controls mitochondrial oxidative function through a YY1-PGC-1alpha transcriptional complex. Nature 2007, 450, 736–740. [Google Scholar] [CrossRef]
- Zid, B. M.; Rogers, A. N.; Katewa, S. D.; Vargas, M. A.; Kolipinski, M. C.; Lu, T. A.; Benzer, S.; Kapahi, P. , 4E-BP extends lifespan upon dietary restriction by enhancing mitochondrial activity in Drosophila. Cell 2009, 139, 149–160. [Google Scholar] [CrossRef]
- Dikic, I.; Elazar, Z. , Mechanism and medical implications of mammalian autophagy. Nat Rev Mol Cell Biol 2018, 19, 349–364. [Google Scholar] [CrossRef]
- Ganley, I. G.; Lam du, H.; Wang, J.; Ding, X.; Chen, S.; Jiang, X. , ULK1.ATG13.FIP200 complex mediates mTOR signaling and is essential for autophagy. J Biol Chem 2009, 284, 12297–12305. [Google Scholar] [CrossRef]
- Hosokawa, N.; Hara, T.; Kaizuka, T.; Kishi, C.; Takamura, A.; Miura, Y.; Iemura, S.; Natsume, T.; Takehana, K.; Yamada, N.; Guan, J. L.; Oshiro, N.; Mizushima, N. , Nutrient-dependent mTORC1 association with the ULK1-Atg13-FIP200 complex required for autophagy. Mol Biol Cell 2009, 20, 1981–1991. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kundu, M.; Viollet, B.; Guan, K. L. , AMPK and mTOR regulate autophagy through direct phosphorylation of Ulk1. Nat Cell Biol 2011, 13, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y. M.; Jung, C. H.; Seo, M.; Kim, E. K.; Park, J. M.; Bae, S. S.; Kim, D. H. , mTORC1 phosphorylates UVRAG to negatively regulate autophagosome and endosome maturation. Mol Cell 2015, 57, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Martina, J. A.; Chen, Y.; Gucek, M.; Puertollano, R. , MTORC1 functions as a transcriptional regulator of autophagy by preventing nuclear transport of TFEB. Autophagy 2012, 8, 903–914. [Google Scholar] [CrossRef] [PubMed]
- Roczniak-Ferguson, A.; Petit, C. S.; Froehlich, F.; Qian, S.; Ky, J.; Angarola, B.; Walther, T. C.; Ferguson, S. M. , The transcription factor TFEB links mTORC1 signaling to transcriptional control of lysosome homeostasis. Sci Signal 2012, 5, ra42. [Google Scholar] [CrossRef] [PubMed]
- Settembre, C.; Zoncu, R.; Medina, D. L.; Vetrini, F.; Erdin, S.; Erdin, S.; Huynh, T.; Ferron, M.; Karsenty, G.; Vellard, M. C.; Facchinetti, V.; Sabatini, D. M.; Ballabio, A. , A lysosome-to-nucleus signalling mechanism senses and regulates the lysosome via mTOR and TFEB. EMBO J 2012, 31, 1095–1108. [Google Scholar] [CrossRef]
- Martina, J. A.; Diab, H. I.; Lishu, L.; Jeong, A. L.; Patange, S.; Raben, N.; Puertollano, R. , The nutrient-responsive transcription factor TFE3 promotes autophagy, lysosomal biogenesis, and clearance of cellular debris. Sci Signal 2014, 7, ra9. [Google Scholar] [CrossRef]
- Ouyang, Q.; Liu, R. , MTOR-mediates hepatic lipid metabolism through an autophagic SNARE complex. Autophagy 2022, 18, 1467–1469. [Google Scholar] [CrossRef]
- Koren, I.; Reem, E.; Kimchi, A. , DAP1, a novel substrate of mTOR, negatively regulates autophagy. Curr Biol 2010, 20, 1093–1098. [Google Scholar] [CrossRef]
- Wan, W.; You, Z.; Zhou, L.; Xu, Y.; Peng, C.; Zhou, T.; Yi, C.; Shi, Y.; Liu, W. , mTORC1-Regulated and HUWE1-Mediated WIPI2 Degradation Controls Autophagy Flux. Mol Cell 2018, 72, 303–315. [Google Scholar] [CrossRef]
- Zinzalla, V.; Stracka, D.; Oppliger, W.; Hall, M. N. , Activation of mTORC2 by association with the ribosome. Cell 2011, 144, 757–768. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Dibble, C. C.; Matsuzaki, M.; Manning, B. D. , The TSC1-TSC2 complex is required for proper activation of mTOR complex 2. Mol Cell Biol 2008, 28, 4104–4115. [Google Scholar] [CrossRef] [PubMed]
- Shah, O. J.; Wang, Z.; Hunter, T. , Inappropriate activation of the TSC/Rheb/mTOR/S6K cassette induces IRS1/2 depletion, insulin resistance, and cell survival deficiencies. Curr Biol 2004, 14, 1650–1656. [Google Scholar] [CrossRef] [PubMed]
- Sung Hee Um, F. F. M. W.; Fre De Ric Picard, M. J. M. S.; Peter, R. Allegrini, S. C. K. J., Absence of S6K1 protects against age- and diet-induced obesity while enhancing insulin sensitivity. Nature 2004, 431, 200–205. [Google Scholar]
- Yoneyama, Y.; Inamitsu, T.; Chida, K.; Iemura, S. I.; Natsume, T.; Maeda, T.; Hakuno, F.; Takahashi, S. I. , Serine Phosphorylation by mTORC1 Promotes IRS-1 Degradation through SCFbeta-TRCP E3 Ubiquitin Ligase. iScience 2018, 5, 1–18. [Google Scholar] [CrossRef]
- Hsu, P. P.; Kang, S. A.; Rameseder, J.; Zhang, Y.; Ottina, K. A.; Lim, D.; Peterson, T. R.; Choi, Y.; Gray, N. S.; Yaffe, M. B.; Marto, J. A.; Sabatini, D. M. , The mTOR-regulated phosphoproteome reveals a mechanism of mTORC1-mediated inhibition of growth factor signaling. Science 2011, 332, 1317–1322. [Google Scholar] [CrossRef]
- Yu, Y.; Yoon, S. O.; Poulogiannis, G.; Yang, Q.; Ma, X. M.; Villen, J.; Kubica, N.; Hoffman, G. R.; Cantley, L. C.; Gygi, S. P.; Blenis, J. , Phosphoproteomic analysis identifies Grb10 as an mTORC1 substrate that negatively regulates insulin signaling. Science 2011, 332, 1322–1326. [Google Scholar] [CrossRef]
- Julien, L. A.; Carriere, A.; Moreau, J.; Roux, P. P. , mTORC1-activated S6K1 phosphorylates Rictor on threonine 1135 and regulates mTORC2 signaling. Mol Cell Biol 2010, 30, 908–921. [Google Scholar] [CrossRef]
- Liu, P.; Gan, W.; Inuzuka, H.; Lazorchak, A. S.; Gao, D.; Arojo, O.; Liu, D.; Wan, L.; Zhai, B.; Yu, Y.; Yuan, M.; Kim, B. M.; Shaik, S.; Menon, S.; Gygi, S. P.; Lee, T. H.; Asara, J. M.; Manning, B. D.; Blenis, J.; Su, B.; Wei, W. , Sin1 phosphorylation impairs mTORC2 complex integrity and inhibits downstream Akt signalling to suppress tumorigenesis. Nat Cell Biol 2013, 15, 1340–1350. [Google Scholar] [CrossRef]
- Kazyken, D.; Magnuson, B.; Bodur, C.; Acosta-Jaquez, H. A.; Zhang, D.; Tong, X.; Barnes, T. M.; Steinl, G. K.; Patterson, N. E.; Altheim, C. H.; Sharma, N.; Inoki, K.; Cartee, G. D.; Bridges, D.; Yin, L.; Riddle, S. M.; Fingar, D. C. , AMPK directly activates mTORC2 to promote cell survival during acute energetic stress. Sci Signal 2019, 12. [Google Scholar] [CrossRef]
- Betz, C.; Hall, M. N. , Where is mTOR and what is it doing there? J Cell Biol 2013, 203, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Sarbassov, D. D.; Guertin, D. A.; Ali, S. M.; Sabatini, D. M. , Phosphorylation and regulation of Akt/PKB by the rictor-mTOR complex. FEBRUARY 2005, 307, 1098–1101. [Google Scholar] [CrossRef] [PubMed]
- Jacinto, E.; Facchinetti, V.; Liu, D.; Soto, N.; Wei, S.; Jung, S. Y.; Huang, Q.; Qin, J.; Su, B. , SIN1/MIP1 maintains rictor-mTOR complex integrity and regulates Akt phosphorylation and substrate specificity. Cell 2006, 127, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Guertin, D. A.; Stevens, D. M.; Thoreen, C. C.; Burds, A. A.; Kalaany, N. Y.; Moffat, J.; Brown, M.; Fitzgerald, K. J.; Sabatini, D. M. , Ablation in mice of the mTORC components raptor, rictor, or mLST8 reveals that mTORC2 is required for signaling to Akt-FOXO and PKCalpha, but not S6K1. Dev Cell 2006, 11, 859–871. [Google Scholar] [CrossRef]
- Bakker, W. J.; Harris, I. S.; Mak, T. W. , FOXO3a is activated in response to hypoxic stress and inhibits HIF1-induced apoptosis via regulation of CITED2. Mol Cell 2007, 28, 941–953. [Google Scholar] [CrossRef]
- Garcia-Martinez, J. M.; Alessi, D. R. , mTOR complex 2 (mTORC2) controls hydrophobic motif phosphorylation and activation of serum- and glucocorticoid-induced protein kinase 1 (SGK1). Biochem J 2008, 416, 375–385. [Google Scholar] [CrossRef]
- Weiler, M.; Blaes, J.; Pusch, S.; Sahm, F.; Czabanka, M.; Luger, S.; Bunse, L.; Solecki, G.; Eichwald, V.; Jugold, M.; Hodecker, S.; Osswald, M.; Meisner, C.; Hielscher, T.; Rubmann, P.; Pfenning, P. N.; Ronellenfitsch, M.; Kempf, T.; Schnolzer, M.; Abdollahi, A.; Lang, F.; Bendszus, M.; von Deimling, A.; Winkler, F.; Weller, M.; Vajkoczy, P.; Platten, M.; Wick, W. , mTOR target NDRG1 confers MGMT-dependent resistance to alkylating chemotherapy. Proc Natl Acad Sci U S A 2014, 111, 409–414. [Google Scholar] [CrossRef]
- Sciarretta, S.; Zhai, P.; Maejima, Y.; Del Re, D. P.; Nagarajan, N.; Yee, D.; Liu, T.; Magnuson, M. A.; Volpe, M.; Frati, G.; Li, H.; Sadoshima, J. , mTORC2 regulates cardiac response to stress by inhibiting MST1. Cell Rep 2015, 11, 125–136. [Google Scholar] [CrossRef]
- Ma, S.; Meng, Z.; Chen, R.; Guan, K. L. , The Hippo Pathway: Biology and Pathophysiology. Annu Rev Biochem 2019, 88, 577–604. [Google Scholar] [CrossRef]
- Jacinto, E.; Loewith, R.; Schmidt, A.; Lin, S.; Ruegg, M. A.; Hall, A.; Hall, M. N. , Mammalian TOR complex 2 controls the actin cytoskeleton and is rapamycin insensitive. Nat Cell Biol 2004, 6, 1122–1128. [Google Scholar] [CrossRef]
- Lozupone, C. A.; Stombaugh, J. I.; Gordon, J. I.; Jansson, J. K.; Knight, R. , Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Adak, A.; Khan, M. R. , An insight into gut microbiota and its functionalities. Cell Mol Life Sci 2019, 76, 473–493. [Google Scholar] [CrossRef]
- Rampelli, S.; Schnorr, S. L.; Consolandi, C.; Turroni, S.; Severgnini, M.; Peano, C.; Brigidi, P.; Crittenden, A. N.; Henry, A. G.; Candela, M. , Metagenome Sequencing of the Hadza Hunter-Gatherer Gut Microbiota. Curr Biol 2015, 25, 1682–1693. [Google Scholar] [CrossRef] [PubMed]
- Adak, A.; Maity, C.; Ghosh, K.; Mondal, K. C. , Alteration of predominant gastrointestinal flora and oxidative damage of large intestine under simulated hypobaric hypoxia. Z Gastroenterol 2014, 52, 180–186. [Google Scholar] [CrossRef]
- Adak, A.; Maity, C.; Ghosh, K.; Pati, B. R.; Mondal, K. C. , Dynamics of predominant microbiota in the human gastrointestinal tract and change in luminal enzymes and immunoglobulin profile during high-altitude adaptation. Folia Microbiol (Praha) 2013, 58, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Nardone, G.; Compare, D. , The human gastric microbiota: Is it time to rethink the pathogenesis of stomach diseases? United European Gastroenterol J 2015, 3, 255–260. [Google Scholar] [CrossRef] [PubMed]
- El Aidy, S.; van den Bogert, B.; Kleerebezem, M. , The small intestine microbiota, nutritional modulation and relevance for health. Curr Opin Biotechnol 2015, 32, 14–20. [Google Scholar] [CrossRef]
- Hollister, E. B.; Gao, C.; Versalovic, J. , Compositional and functional features of the gastrointestinal microbiome and their effects on human health. Gastroenterology 2014, 146, 1449–1458. [Google Scholar] [CrossRef]
- Belkaid, Y.; Hand, T. W. , Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef]
- Gerard, P. , Gut microbiota and obesity. Cell Mol Life Sci 2016, 73, 147–162. [Google Scholar] [CrossRef]
- Ling, Z.; Liu, X.; Cheng, Y.; Yan, X.; Wu, S. , Gut microbiota and aging. Crit Rev Food Sci Nutr 2022, 62, 3509–3534. [Google Scholar] [CrossRef]
- Ni, J.; Wu, G. D.; Albenberg, L.; Tomov, V. T. , Gut microbiota and IBD: causation or correlation? Nat Rev Gastroenterol Hepatol 2017, 14, 573–584. [Google Scholar] [CrossRef]
- Jameson, K. G.; Olson, C. A.; Kazmi, S. A.; Hsiao, E. Y. , Toward Understanding Microbiome-Neuronal Signaling. Mol Cell 2020, 78, 577–583. [Google Scholar] [CrossRef]
- Xu, J. Y.; Liu, M. T.; Tao, T.; Zhu, X.; Fei, F. Q. , The role of gut microbiota in tumorigenesis and treatment. Biomed Pharmacother 2021, 138, 111444. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Albenberg, L.; Compher, C.; Baldassano, R.; Piccoli, D.; Lewis, J. D.; Wu, G. D. , Diet in the pathogenesis and treatment of inflammatory bowel diseases. Gastroenterology 2015, 148, 1087–1106. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, K.; Kanai, T. , The gut microbiota and inflammatory bowel disease. Semin Immunopathol 2015, 37, 47–55. [Google Scholar] [CrossRef]
- Lloyd-Price, J.; Arze, C.; Ananthakrishnan, A. N.; Schirmer, M.; Avila-Pacheco, J.; Poon, T. W.; Andrews, E.; Ajami, N. J.; Bonham, K. S.; Brislawn, C. J.; Casero, D.; Courtney, H.; Gonzalez, A.; Graeber, T. G.; Hall, A. B.; Lake, K.; Landers, C. J.; Mallick, H.; Plichta, D. R.; Prasad, M.; Rahnavard, G.; Sauk, J.; Shungin, D.; Vazquez-Baeza, Y.; White, R. A., 3rd; Investigators, I.; Braun, J.; Denson, L. A.; Jansson, J. K.; Knight, R.; Kugathasan, S.; McGovern, D. P. B.; Petrosino, J. F.; Stappenbeck, T. S.; Winter, H. S.; Clish, C. B.; Franzosa, E. A.; Vlamakis, H.; Xavier, R. J.; Huttenhower, C. , Multi-omics of the gut microbial ecosystem in inflammatory bowel diseases. Nature 2019, 569, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Zhao, Y.; Shi, L.; Li, W.; Chen, K.; Li, M.; Chen, X.; Zhang, H.; Li, T.; Matsuzawa-Ishimoto, Y.; Yao, X.; Shao, D.; Ke, Z.; Li, J.; Chen, Y.; Zhang, X.; Cui, J.; Cui, S.; Leng, Q.; Cadwell, K.; Li, X.; Wei, H.; Zhang, H.; Li, H.; Xiao, H. , Gut epithelial TSC1/mTOR controls RIPK3-dependent necroptosis in intestinal inflammation and cancer. J Clin Invest 2020, 130, 2111–2128. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Huang, X.; Zhao, Y.; Chen, F.; Sun, M.; Yang, W.; Chen, L.; Yao, S.; Peniche, A.; Dann, S. M.; Sun, J.; Golovko, G.; Fofanov, Y.; Miao, Y.; Liu, Z.; Chen, D.; Cong, Y. , Interleukin-33 Promotes REG3gamma Expression in Intestinal Epithelial Cells and Regulates Gut Microbiota. Cell Mol Gastroenterol Hepatol 2019, 8, 21–36. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Yu, T.; Huang, X.; Bilotta, A. J.; Xu, L.; Lu, Y.; Sun, J.; Pan, F.; Zhou, J.; Zhang, W.; Yao, S.; Maynard, C. L.; Singh, N.; Dann, S. M.; Liu, Z.; Cong, Y. , Intestinal microbiota-derived short-chain fatty acids regulation of immune cell IL-22 production and gut immunity. Nat Commun 2020, 11, 4457. [Google Scholar] [CrossRef]
- Qing Zhao; W., L.; Duck1 Fengyuan Huang; Katie L.Alexander; L., C.; Maynard; J., P.; Mannon; Elson, C. O., CD4+T cell activation and concomitant mTOR metabolic inhibition can ablate microbiota-specific memory cells and prevent colitis. SCIENCE IMMUNOLOGY.
- Sakai, K.; De Velasco, M. A.; Kura, Y.; Nishio, K. , Transcriptome Profiling and Metagenomic Analysis Help to Elucidate Interactions in an Inflammation-Associated Cancer Mouse Model. Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Xie, W.; Wan, X.; Deng, T. , Clostridium butyricum protects intestinal barrier function via upregulation of tight junction proteins and activation of the Akt/mTOR signaling pathway in a mouse model of dextran sodium sulfate-induced colitis. Exp Ther Med 2020, 20, 10. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Shan, Y.-J.; He, C.-X.; Ren, M.-H.; Tian, P.-J.; Song, W. , Effects of L. paracasei subp. paracasei X12 on cell cycle of colon cancer HT-29 cells and regulation of mTOR signalling pathway. Journal of Functional Foods 2016, 21, 431–439. [Google Scholar] [CrossRef]
- Jorgensen, S. F.; Troseid, M.; Kummen, M.; Anmarkrud, J. A.; Michelsen, A. E.; Osnes, L. T.; Holm, K.; Hoivik, M. L.; Rashidi, A.; Dahl, C. P.; Vesterhus, M.; Halvorsen, B.; Mollnes, T. E.; Berge, R. K.; Moum, B.; Lundin, K. E.; Fevang, B.; Ueland, T.; Karlsen, T. H.; Aukrust, P.; Hov, J. R. , Altered gut microbiota profile in common variable immunodeficiency associates with levels of lipopolysaccharide and markers of systemic immune activation. Mucosal Immunol 2016, 9, 1455–1465. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhou, R.; Wang, H.; Li, W.; Pan, M.; Yao, X.; Zhan, W.; Yang, S.; Xu, L.; Ding, Y.; Zhao, L. , Gut microbiota-stimulated cathepsin K secretion mediates TLR4-dependent M2 macrophage polarization and promotes tumor metastasis in colorectal cancer. Cell Death Differ 2019, 26, 2447–2463. [Google Scholar] [CrossRef]
- Cassard, A. M.; Ciocan, D. , Microbiota, a key player in alcoholic liver disease. Clin Mol Hepatol 2018, 24, 100–107. [Google Scholar] [CrossRef]
- Miura, K.; Ohnishi, H. , Role of gut microbiota and Toll-like receptors in nonalcoholic fatty liver disease. World J Gastroenterol 2014, 20, 7381–7391. [Google Scholar] [CrossRef]
- Ferrin, G.; Guerrero, M.; Amado, V.; Rodriguez-Peralvarez, M.; De la Mata, M. , Activation of mTOR Signaling Pathway in Hepatocellular Carcinoma. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef]
- Chiu, C. C.; Ching, Y. H.; Li, Y. P.; Liu, J. Y.; Huang, Y. T.; Huang, Y. W.; Yang, S. S.; Huang, W. C.; Chuang, H. L. , Nonalcoholic Fatty Liver Disease Is Exacerbated in High-Fat Diet-Fed Gnotobiotic Mice by Colonization with the Gut Microbiota from Patients with Nonalcoholic Steatohepatitis. Nutrients 2017, 9. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, W.; Xu, J.; Wu, N.; Wang, Y.; Lin, T.; Liu, Y.; Liu, Y., E. coli NF73-1 Isolated From NASH Patients Aggravates NAFLD in Mice by Translocating Into the Liver and Stimulating M1 Polarization. Front Cell Infect Microbiol 2020, 10, 535940. [Google Scholar] [CrossRef]
- Xia, F.; Xiang, S.; Chen, Z.; Song, L.; Li, Y.; Liao, Z.; Ge, B.; Zhou, B. , The probiotic effects of AB23A on high-fat-diet-induced non-alcoholic fatty liver disease in mice may be associated with suppressing the serum levels of lipopolysaccharides and branched-chain amino acids. Arch Biochem Biophys 2021, 714, 109080. [Google Scholar] [CrossRef] [PubMed]
- Chao Cai1*, D.-Z. C. , Li-Chao Ge1, Wen-Kai Chen1, Sha-Sha Ye1, Wei-Wei Ye1,3, Ying Tao1, Rui Wang1, Ji Li1, Zhuo Lin1, Xiao-Dong Wang1, Lan-Man Xu1,4,5, Yong-Ping Chen1, Synergistic effects of Lactobacillus rhamnosus culture supernatant and bone marrow mesenchymal stem cells on the development of alcoholic steatohepatitis in mice. Am J Transl Res 2019. [Google Scholar]
- Pant, K.; Saraya, A.; Venugopal, S. K. , Oxidative stress plays a key role in butyrate-mediated autophagy via Akt/mTOR pathway in hepatoma cells. Chem Biol Interact 2017, 273, 99–106. [Google Scholar] [CrossRef]
- Qiao, C.-M.; Sun, M.-F.; Jia, X.-B.; Shi, Y.; Zhang, B.-P.; Zhou, Z.-L.; Zhao, L.-P.; Cui, C.; Shen, Y.-Q. , Sodium butyrate causes α-synuclein degradation by an Atg5-dependent and PI3K/Akt/mTOR-related autophagy pathway. Experimental Cell Research 2020, 387. [Google Scholar] [CrossRef]
- Al-Bari, M. A. A.; Xu, P. , Molecular regulation of autophagy machinery by mTOR-dependent and -independent pathways. Ann N Y Acad Sci 2020, 1467, 3–20. [Google Scholar] [CrossRef]
- Zheng, D.; Liu, Z.; Zhou, Y.; Hou, N.; Yan, W.; Qin, Y.; Ye, Q.; Cheng, X.; Xiao, Q.; Bao, Y.; Luo, J.; Wu, X. , Urolithin B, a gut microbiota metabolite, protects against myocardial ischemia/reperfusion injury via p62/Keap1/Nrf2 signaling pathway. Pharmacol Res 2020, 153, 104655. [Google Scholar] [CrossRef]
- Huang, X.; Gao, H.; Jiang, X.; Zheng, Z. , Urolithin B, a Gut Microbiota Metabolite, Reduced Susceptibility to Myocardial Arrhythmic Predisposition after Hypoxia. Dis Markers 2022, 2022, 6517266. [Google Scholar] [CrossRef] [PubMed]
- Cai, K.; Ma, Y.; Cai, F.; Huang, X.; Xiao, L.; Zhong, C.; Ren, P.; Luo, Q.; Chen, J.; Han, F. , Changes of gut microbiota in diabetic nephropathy and its effect on the progression of kidney injury. Endocrine 2022, 76, 294–303. [Google Scholar] [CrossRef]
- Xiao, S.; Zhang, Z.; Chen, M.; Zou, J.; Jiang, S.; Qian, D.; Duan, J. , Xiexin Tang ameliorates dyslipidemia in high-fat diet-induced obese rats via elevating gut microbiota-derived short chain fatty acids production and adjusting energy metabolism. J Ethnopharmacol 2019, 241, 112032. [Google Scholar] [CrossRef]
- Hou, T.; Sun, X.; Zhu, J.; Hon, K. L.; Jiang, P.; Chu, I. M.; Tsang, M. S.; Lam, C. W.; Zeng, H.; Wong, C. K. , IL-37 Ameliorating Allergic Inflammation in Atopic Dermatitis Through Regulating Microbiota and AMPK-mTOR Signaling Pathway-Modulated Autophagy Mechanism. Front Immunol 2020, 11, 752. [Google Scholar] [CrossRef]
- Liu, S. K.; Ma, L. B.; Yuan, Y.; Ji, X. Y.; Sun, W. J.; Duan, J. X.; Zeng, Q. P.; Wasti, B.; Xiao, B.; Zheng, J. F.; Chen, P.; Xiang, X. D. , Alanylglutamine Relieved Asthma Symptoms by Regulating Gut Microbiota and the Derived Metabolites in Mice. Oxid Med Cell Longev 2020, 2020, 7101407. [Google Scholar] [CrossRef] [PubMed]
- Ju, Z.; Guo, P.; Xiang, J.; Lei, R.; Ren, G.; Zhou, M.; Yang, X.; Zhou, P.; Huang, R. , Low-dose radiation exaggerates HFD-induced metabolic dysfunction by gut microbiota through PA-PYCR1 axis. Communications Biology 2022, 5. [Google Scholar] [CrossRef] [PubMed]
- Jung, M. J.; Lee, J.; Shin, N. R.; Kim, M. S.; Hyun, D. W.; Yun, J. H.; Kim, P. S.; Whon, T. W.; Bae, J. W. , Chronic Repression of mTOR Complex 2 Induces Changes in the Gut Microbiota of Diet-induced Obese Mice. Sci Rep 2016, 6, 30887. [Google Scholar] [CrossRef] [PubMed]
- Hurez, V.; Dao, V.; Liu, A.; Pandeswara, S.; Gelfond, J.; Sun, L.; Bergman, M.; Orihuela, C. J.; Galvan, V.; Padrón, Á.; Drerup, J.; Liu, Y.; Hasty, P.; Sharp, Z. D.; Curiel, T. J. , Chronic mTOR inhibition in mice with rapamycin alters T, B, myeloid, and innate lymphoid cells and gut flora and prolongs life of immune-deficient mice. Aging Cell 2015, 14, 945–956. [Google Scholar] [CrossRef]
- Chen, C.; Fang, S.; Wei, H.; He, M.; Fu, H.; Xiong, X.; Zhou, Y.; Wu, J.; Gao, J.; Yang, H.; Huang, L. , Prevotella copri increases fat accumulation in pigs fed with formula diets. Microbiome 2021, 9, 175. [Google Scholar] [CrossRef]
- Weinstock, G. M. , Genomic approaches to studying the human microbiota. Nature 2012, 489, 250–256. [Google Scholar] [CrossRef]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K. S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; Mende, D. R.; Li, J.; Xu, J.; Li, S.; Li, D.; Cao, J.; Wang, B.; Liang, H.; Zheng, H.; Xie, Y.; Tap, J.; Lepage, P.; Bertalan, M.; Batto, J. M.; Hansen, T.; Le Paslier, D.; Linneberg, A.; Nielsen, H. B.; Pelletier, E.; Renault, P.; Sicheritz-Ponten, T.; Turner, K.; Zhu, H.; Yu, C.; Li, S.; Jian, M.; Zhou, Y.; Li, Y.; Zhang, X.; Li, S.; Qin, N.; Yang, H.; Wang, J.; Brunak, S.; Dore, J.; Guarner, F.; Kristiansen, K.; Pedersen, O.; Parkhill, J.; Weissenbach, J.; Meta, H. I. T. C.; Bork, P.; Ehrlich, S. D.; Wang, J. , A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef]
- Hong, Y.; Chen, B.; Zhai, X.; Qian, Q.; Gui, R.; Jiang, C. , Integrated analysis of the gut microbiome and metabolome in a mouse model of inflammation-induced colorectal tumors. Front Microbiol 2022, 13, 1082835. [Google Scholar] [CrossRef]
- Kang, H.; You, H. J.; Lee, G.; Lee, S. H.; Yoo, T.; Choi, M.; Joo, S. K.; Park, J. H.; Chang, M. S.; Lee, D. H.; Kim, W.; Ko, G.; Innovative Target Exploration of, N. c. , Interaction effect between NAFLD severity and high carbohydrate diet on gut microbiome alteration and hepatic de novo lipogenesis. Gut Microbes 2022, 14, 2078612. [Google Scholar] [CrossRef]
- Li, M. X.; Li, M. Y.; Lei, J. X.; Wu, Y. Z.; Li, Z. H.; Chen, L. M.; Zhou, C. L.; Su, J. Y.; Huang, G. X.; Huang, X. Q.; Zheng, X. B. , Huangqin decoction ameliorates DSS-induced ulcerative colitis: Role of gut microbiota and amino acid metabolism, mTOR pathway and intestinal epithelial barrier. Phytomedicine 2022, 100, 154052. [Google Scholar] [CrossRef]
- Wang, D.; Shao, S.; Zhang, Y.; Zhao, D.; Wang, M. , Insight Into Polysaccharides From Panax ginseng C. A. Meyer in Improving Intestinal Inflammation: Modulating Intestinal Microbiota and Autophagy. Front Immunol 2021, 12, 683911. [Google Scholar] [CrossRef] [PubMed]
- Qing Zhao; Lennard W. Duck; Fengyuan Huang; Katie L. Alexander; Craig L. Maynard; Peter J. Mannon; Elson, C. O., CD4+ T cell activation and concomitant mTOR metabolic inhibition can ablate microbiota-specific memory cells and prevent colitis. SCIENCE IMMUNOLOGY 2020, 5.
- Seif El-Din, S. H.; Salem, M. B.; El-Lakkany, N. M.; Hammam, O. A.; Nasr, S. M.; Okasha, H.; Ahmed, L. A.; Saleh, S.; Botros, S. S. , Early intervention with probiotics and metformin alleviates liver injury in NAFLD rats via targeting gut microbiota dysbiosis and p-AKT/mTOR/LC-3II pathways. Hum Exp Toxicol 2021, 40, 1496–1509. [Google Scholar] [CrossRef] [PubMed]
- Chao Cai; Da-Zhi Chen; Ge,, L. -C.; Chen,, W.-K.; Ye,, S.-S.; Ye,, W.-W.; Tao,, Y.; Rui Wang; Ji Li; Zhuo Lin; Xiao-Dong Wang; Xu, L.-M.; Chen, Y.-P., Synergistic effects of Lactobacillus rhamnosus culture supernatant and bone marrow mesenchymal stem cells on the development of alcoholic steatohepatitis in mice. Am J Transl Res 2019, 11, 5703–5715. [Google Scholar]
- Deng, Z.; Wu, N.; Wang, J.; Geng, L.; Yue, Y.; Wang, F.; Zhang, Q. , Low molecular weight fucoidan fraction LF2 improves metabolic syndrome via up-regulating PI3K-AKT-mTOR axis and increasing the abundance of Akkermansia muciniphila in the gut microbiota. Int J Biol Macromol 2021, (Pt A), 789–798. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, M.; Ericsson, A. C. , The Potential Gut Microbiota-Mediated Treatment Options for Liver Cancer. Front Oncol 2020, 10, 524205. [Google Scholar] [CrossRef]
- Jin, M.; Kong, L.; Han, Y.; Zhang, S. , Gut microbiota enhances the chemosensitivity of hepatocellular carcinoma to 5-fluorouracil in vivo by increasing curcumin bioavailability. Phytother Res 2021, 35, 5823–5837. [Google Scholar] [CrossRef]
- El Sayed, N. S.; Kandil, E. A.; Ghoneum, M. H. , Enhancement of Insulin/PI3K/Akt Signaling Pathway and Modulation of Gut Microbiome by Probiotics Fermentation Technology, a Kefir Grain Product, in Sporadic Alzheimer's Disease Model in Mice. Front Pharmacol 2021, 12, 666502. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, X.; Zheng, Y.; Liu, W.; Ding, C. , Aronia melanocarpa polysaccharide ameliorates inflammation and aging in mice by modulating the AMPK/SIRT1/NF-kappaB signaling pathway and gut microbiota. Sci Rep 2021, 11, 20558. [Google Scholar] [CrossRef]
- Ju, P.; Ding, W.; Chen, J.; Cheng, Y.; Yang, B.; Huang, L.; Zhou, Q.; Zhu, C.; Li, X.; Wang, M.; Chen, J. , The protective effects of Mogroside V and its metabolite 11-oxo-mogrol of intestinal microbiota against MK801-induced neuronal damages. Psychopharmacology (Berl) 2020, 237, 1011–1026. [Google Scholar] [CrossRef]
- Zhang, Y. X.; Qu, S. S.; Zhang, L. H.; Gu, Y. Y.; Chen, Y. H.; Huang, Z. Y.; Liu, M. H.; Zou, W.; Jiang, J.; Chen, J. Q.; Wang, Y. J.; Zhou, F. H. , The Role of Ophiopogonin D in Atherosclerosis: Impact on Lipid Metabolism and Gut Microbiota. Am J Chin Med 2021, 49, 1449–1471. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Zheng, C.; Song, B.; Guo, Q.; Zhong, Y.; Zhang, S.; Zhang, L.; Duan, G.; Li, F.; Duan, Y. , HMB Improves Lipid Metabolism of Bama Xiang Mini-Pigs via Modulating the Bacteroidetes-Acetic Acid-AMPKalpha Axis. Front Microbiol 2021, 12, 736997. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Liao, X.; Wang, X.; Lao, S.; Liao, W. , The biological regulatory activities of Flammulina velutipes polysaccharide in mice intestinal microbiota, immune repertoire and heart transcriptome. Int J Biol Macromol 2021, 185, 582–591. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Chen, X.; Xiao, Y.; Li, D.; Li, M.; Li, H.; Huang, J.; Lai, Z.; Su, Z.; Xie, Y.; Zhu, D.; Chen, Q.; Lu, H.; He, J.; Xia, C. , Bruceae Fructus Oil Inhibits Triple-Negative Breast Cancer by Restraining Autophagy: Dependence on the Gut Microbiota-Mediated Amino Acid Regulation. Front Pharmacol 2021, 12, 727082. [Google Scholar] [CrossRef]
- Xia, T.; Zhang, B.; Li, Y.; Fang, B.; Zhu, X.; Xu, B.; Zhang, J.; Wang, M.; Fang, J. , New insight into 20(S)-ginsenoside Rh2 against T-cell acute lymphoblastic leukemia associated with the gut microbiota and the immune system. Eur J Med Chem 2020, 203, 112582. [Google Scholar] [CrossRef] [PubMed]
- Panebianco, C.; Adamberg, K.; Adamberg, S.; Saracino, C.; Jaagura, M.; Kolk, K.; Di Chio, A. G.; Graziano, P.; Vilu, R.; Pazienza, V. , Engineered Resistant-Starch (ERS) Diet Shapes Colon Microbiota Profile in Parallel with the Retardation of Tumor Growth in In Vitro and In Vivo Pancreatic Cancer Models. Nutrients 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Liang, J.; Fu, N.; Han, Y.; Qin, J. , A Ketogenic Diet and the Treatment of Autism Spectrum Disorder. Front Pediatr 2021, 9, 650624. [Google Scholar] [CrossRef] [PubMed]
- Hurez, V.; Dao, V.; Liu, A.; Pandeswara, S.; Gelfond, J.; Sun, L.; Bergman, M.; Orihuela, C. J.; Galvan, V.; Padron, A.; Drerup, J.; Liu, Y.; Hasty, P.; Sharp, Z. D.; Curiel, T. J. , Chronic mTOR inhibition in mice with rapamycin alters T, B, myeloid, and innate lymphoid cells and gut flora and prolongs life of immune-deficient mice. Aging Cell 2015, 14, 945–956. [Google Scholar] [CrossRef]
- Zhao, Y.; Chen, F.; Wu, W.; Sun, M.; Bilotta, A. J.; Yao, S.; Xiao, Y.; Huang, X.; Eaves-Pyles, T. D.; Golovko, G.; Fofanov, Y.; D'Souza, W.; Zhao, Q.; Liu, Z.; Cong, Y. , GPR43 mediates microbiota metabolite SCFA regulation of antimicrobial peptide expression in intestinal epithelial cells via activation of mTOR and STAT3. Mucosal Immunol 2018, 11, 752–762. [Google Scholar] [CrossRef]
- Leung, H. K. M.; Lo, E. K. K.; El-Nezami, H. , Theabrownin Alleviates Colorectal Tumorigenesis in Murine AOM/DSS Model via PI3K/Akt/mTOR Pathway Suppression and Gut Microbiota Modulation. Antioxidants (Basel) 2022, 11. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of crosstalk between gut microbes and mTOR signaling pathway in different diseases. Gut microbes directly or indirectly stimulate mTOR through changes in their own abundance or metabolites (mainly SCFAs), integrating intra- and extracellular signals, which in turn influence disease development. A detailed description of these interactions is provided in the main text. Arrows represent activation and horizontal lines indicate inhibition. The different coloured lines represent different regulatory mechanisms. Abbreviations: AMPK: AMP-activated protein kinase, ERK1/2: Extracellular signal–regulated kinases 1/2, ROS: Reactive oxygen species, SCFAs: Short chain fatty acids, IL33: Interleukin-33, IL-22: Interleukin-22, CTSK: Cathepsin K, LPS: lipopolysaccharide, TAMs: Tumor-associated macrophages.
Figure 1.
Schematic representation of crosstalk between gut microbes and mTOR signaling pathway in different diseases. Gut microbes directly or indirectly stimulate mTOR through changes in their own abundance or metabolites (mainly SCFAs), integrating intra- and extracellular signals, which in turn influence disease development. A detailed description of these interactions is provided in the main text. Arrows represent activation and horizontal lines indicate inhibition. The different coloured lines represent different regulatory mechanisms. Abbreviations: AMPK: AMP-activated protein kinase, ERK1/2: Extracellular signal–regulated kinases 1/2, ROS: Reactive oxygen species, SCFAs: Short chain fatty acids, IL33: Interleukin-33, IL-22: Interleukin-22, CTSK: Cathepsin K, LPS: lipopolysaccharide, TAMs: Tumor-associated macrophages.
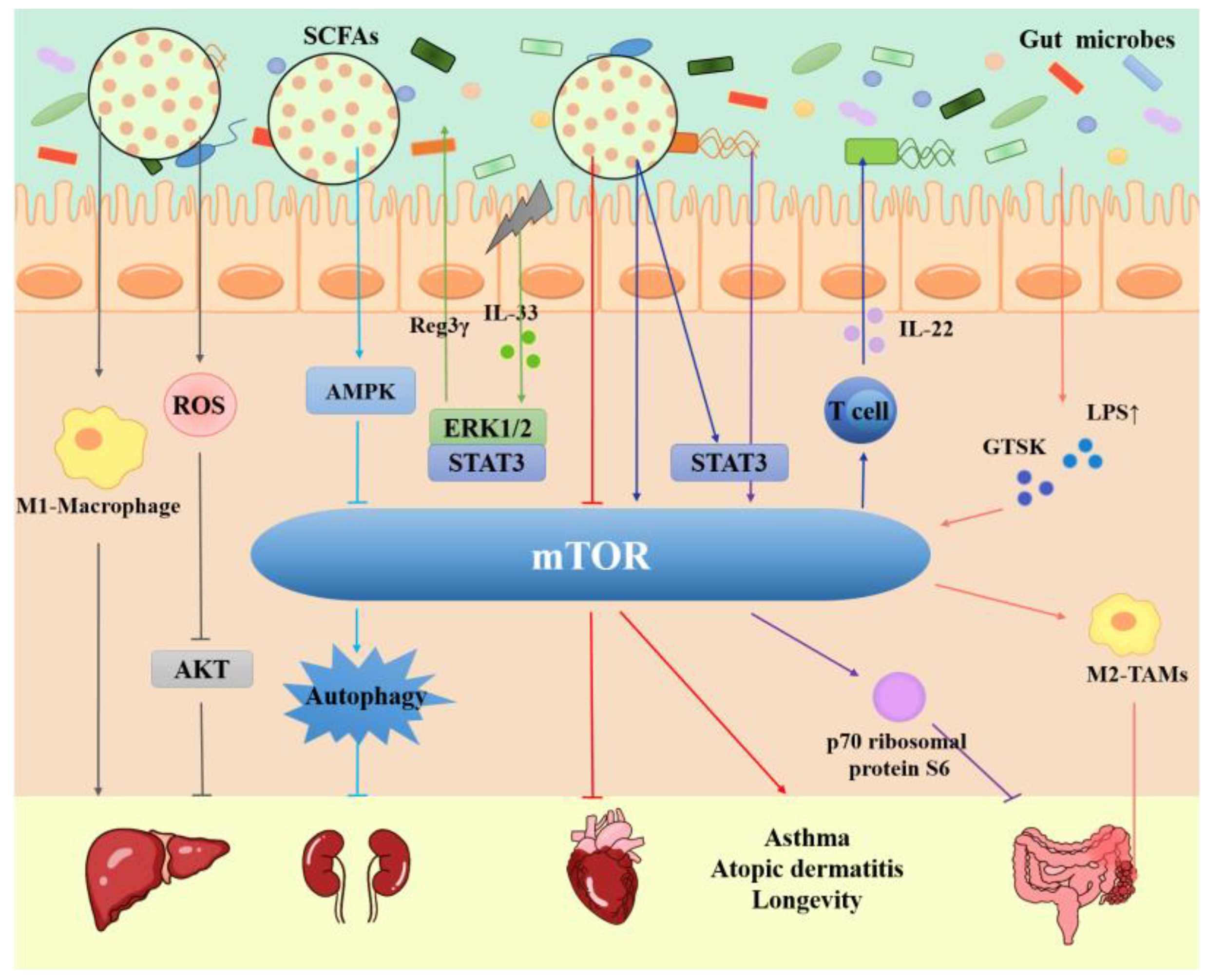

Table 1.
List of drugsaffecting microbiota and mTOR signal pathway in different disorders.
| Name | Disease | Pathway affected | Changes of gut microbiota | Cell response |
|
HQD [121] |
UC | ↑PI3K/AKT/mTOR |
↑Firmicutes, Bacteroidetes, |
↑amino acid metabolism, p-S6 and p-4EBP1 ↓Apoptosis |
|
P.ginseng [122] |
IBD | ↓mTOR, TLR4, NF-kB |
↓Gram-negative bacteria | ↑Autophagy ↓p62 |
|
SCFAs [90] |
CD and UC | ↑mTOR, STAT3 |
against enteric infection of Citrobacter rodentium |
↑HIF1α, AhR, IL-22 ↓Gpr41, HDAC |
|
TB [141] |
CRC | ↓PI3K/AKT/mTOR |
↓Bacteroidceae and Bacteroides ↑Prevotellaceae and Alloprevotella |
↑cyclin D1 protein, cleaved caspase 3 |
|
SCFAs [140] |
—— | ↑mTOR, STAT3 |
—— | ↑AMP, RegIIIγ, β-defensins |
| L. reuteri + MTZ [124] | NAFLD |
↓mTOR, AKT |
↑Akkermansia muciniphila, Firmicutes, butyrate |
↑Autophagy, LC-3II ↓LPS, NF-kB, TNF-α |
|
AB23A [102] |
NAFLD |
↓mTOR, TLR4, NF-kB |
↓Firmicutes/Bacteroidaeota, Actinobacteriota/Bacteroidaeota | ↑ZO-1, occludin |
|
LGG-s and BMMSC [125] |
Alcoholic liver disease | ↓PI3K/mTOR, PI3K/NF-kB |
—— | ↑Autophagy ↓NKB cells, TFH cells |
|
LF2 [126] |
METS |
↓PI3K/AKT/mTOR |
↑Verrucomicrobia, Akkermansia muciniphila |
↓SREBP-1c, PPARγ |
|
Butyrate [104] |
HCC | ↓mTOR, AKT |
—— | ↑ROS, Autophagy: beclin 1, ATG 5, LC3-II |
|
Curcumin [128] |
HCC | ↓PI3K/AKT/mTOR |
↑family Helicobacteraceaeorder, order Campylobacterales, and genus Helicobacter and Campylobacteria |
↑apoptosis |
|
PFT [129] |
AD | ↑PI3K/AKT ↓mTOR, GSK-3β |
—— | ↓oxidative stress, inflammation |
|
AMP [130] |
Brain aging | ↑PI3K/AKT/mTOR ↓AMPK/SIRT1/NF-κB |
↑Bacteroides ↓Firmicutes |
↓apoptosis, NLRP3 |
| MogV and 11-oxo-mogrol [131] | neuronal damages | ↑AKT/mTOR | —— | ↑neurite outgrowth ↓apoptosis, [Ca2+]i release |
|
OPD [132] |
atherosclerosis | ↓mTOR/SREBP1/SCD1 | ↑Bacteroidetes, Faecalibaculum ↓Firmicutes, Ileibacterium |
↑insulin resistance ↓lipid metabolism |
|
HMB [133] |
Obesity | ↑AMPKα, Sirt1, and FoxO1 ↓mTOR |
↑Bacteroidetes, acetic acid | ↓lipid metabolism |
|
XXT [110] |
Obesity | ↑AMPK ↓mTOR |
↑key synthetic enzymes of SCFAs | ↑energy expenditure:PGC-1α, UCP-2 ↓energy intake |
|
FVP [134] |
Heart | ↑mTOR, etc ↓AMPK, PI3K-Akt, etc |
↑Bacteroidetes, Muribaculaceae | ↑Immunity |
|
BO [135] |
TNBC | ↑mTOR | ↑Candidatus Melainabacteria bacterium MEL.A1, Ndongobacter massiliensis, Prevotella ruminicola | ↓Autophagy Regulate amino acid metabolism |
|
GRh2 [136] |
T-ALL | ↓PI3K/AKT/mTOR |
↑Bacteroidetes, Verrucomicrobia ↓Firmicutes, Proteobacteria |
↑Immunity, tight junction proteins, antimicrobial peptides, IgA |
|
ERS Diet [137] |
PC | ↓mTOR, ERK1/2 | ↑diversity of microbiota ↑Formate, Lactate ↓Propionate |
↓Proliferation |
|
Resveratrol [114] |
Obesity and Diabetes |
↓mTOR |
↓Lactococcus, Clostridium XI, Oscillibacter, and Hydrogenoanaerobacterium | ↑insulin resistance |
|
ERapa [115] |
Longevity | ↓mTOR | Alteration of gut metagenomes | Regulate T, B, myeloid, and innate lymphoid cells |
1 Arrows indicate up-regulation or down-regulation.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



