
Submitted:
13 June 2023
Posted:
16 June 2023
You are already at the latest version
Abstract
Parkinson’s-disease (PD) is an incurable age-related neurodegenerative disease and its global prevalence of disability and death has increased exponentially. Although motor symptoms are the characteristic manifestations of PD, the clinical spectrum also contains a wide variety of non-motor symptoms, which are the main cause of disability and determinants of the decrease in a patient's quality of life. Noteworthy in this regard is the stress on the cardiac system that is often observed in the course of PD, however its effects have not yet been adequately researched. Here, an untargeted metabolomics approach was used to assess changes in cardiac metabolism in the 6-hydroxydopamine model of PD. Beta-sitosterol, campesterol, cholesterol, monoacylglycerol, α-tocopherol, stearic acid, beta-glycerophosphoric acid, o-phosphoethanolamine, myo-inositol-1-phosphate, alanine, valine and allothreonine, are the metabolites that significantly discriminate Parkinsonian rats from sham counterparts. Upon analysis of the metabolic pathways with the aim of uncovering the main biological pathways involved in concentration patterns of cardiac metabolites, biosynthesis of both phosphatidylethanolamine and phosphatidylcholine, glucose-alanine cycle, the glutathione metabolism and plasmalogen synthesis most adequately differentiated sham and Parkinsonian rats. Our results reveal that both lipid and energy metabolism are particularly involved in changes in cardiac metabolism in PD. These results provide insight into cardiac metabolic signatures in PD and indicate potential targets for further investigation.
Keywords:
Parkinson’s disease
; cardiac metabolism
; heart
; 6-hydroxydopamine
; metabolites
1. Introduction
The dramatic increase in the world's elderly population brings with it an increased risk of developing neurodegenerative diseases [1]. Parkinson’s disease (PD) is a common age-related neurodegenerative disease identified as the fastest growing disease among neurological disorders in terms of disability and death [2] Motor impairment is the most well-known symptom of PD, but the disease is also known to cause a plethora of non-motor manifestations that impose a significant burden on patients, with an emphasis on cardiovascular abnormalities, that are prevalent in individuals with PD and associated increase in morbidity and mortality [3]. Urgent questions are raised about the cardiovascular implications of PD with the aim of better understanding the impact of the disease on heart health and proposing strategies to improve patient outcomes. Nevertheless, the heart is still given too little attention in PD research. Our study is dedicated to examining the metabolic changes associated with disease within the heart itself. Cardiac tissue metabolomics provides a comprehensive view of the metabolic status by identifying the disease-specific metabolites and their associated metabolic pathways, thereby elucidating the biochemical mechanisms underlying the disease. Here, an untargeted metabolomics approach was used to comprehensively understand the cardiac metabolic changes in the 6-OHDA rat model of PD, a widely used neurotoxic model that mimics PD-like motor and non-motor aspects, and provide valuable insights into disease-associated with metabolic changes in Parkinsonian animals.
2. Results
2.1. Tyrosine hydroxylase immunohistochemistry
Tyrosine hydroxylase (TH) is a rate limiting enzyme in the biosynthesis of dopamine. Here, TH-positive neurons and fibers were evaluated in the SNpc and striatum, respectively (Figure 1). Significant differences were observed between 6-OHDA and sham groups. There was a 42% reduction in TH-positive striatal fibers in the 6-OHDA group compared to the sham group (Figure 2A). The 6-OHDA group also showed a 55% reduction in the TH-immunoreactive neurons in the SNpc (Figure 2B), indicating significant dopaminergic neuronal death.
3.2. Untargeted metabolomics
Metabolomic analysis performed on the cardiac tissue of animals (n=8 per group) subjected to an orthogonal partial least squares test (OPLS) in the heart revealed that there was a robust separation of metabolites between the sham and 6-OHDA groups (Figure 3A-B). The metabolites were plotted hierarchically according to their variable importance (VIP) score (figure 3B-C) and compared to assess significant differences in their concentrations (False Discovery Rate (FDR) adjusted to p-value < 0.05).
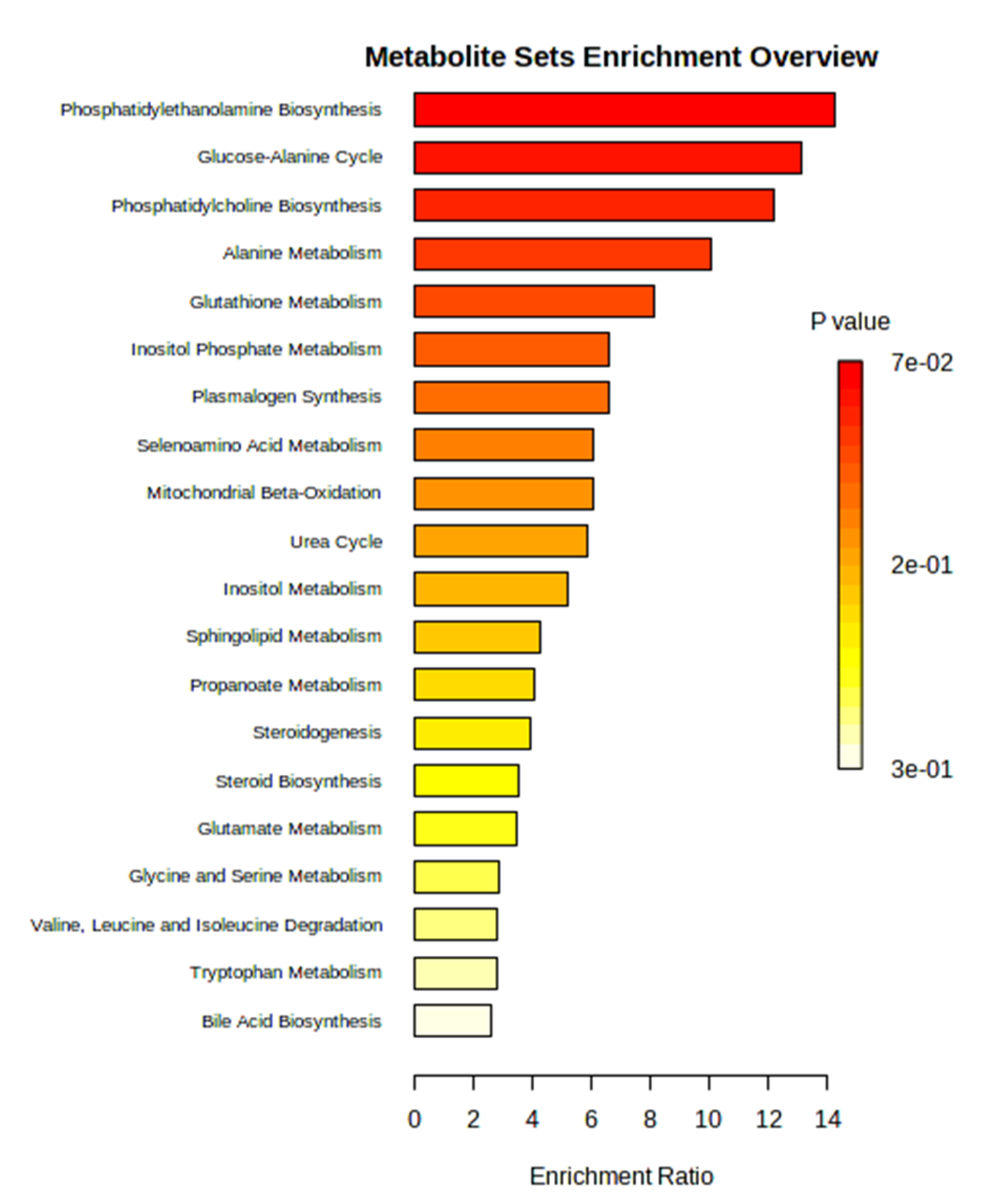
Levels of selected cardiac metabolites were compared to assess differences between the two groups. Lower amounts of the metabolites alanine, allothreonine and valine were identified in the 6-OHDA group compared to the sham group, while higher levels of alpha-tocopherol, beta-glycerophosphoric acid, beta-sitosterol, campesterol, stearic acid, organic phosphoethanolamine, myo- inositol -1-phosphate, monoacylglycerol and cholesterol were found in the 6-OHDA group when compared to the sham group (Figure 4). Metaboanalyst 5.0 software was used to identify the major metabolic pathways associated with the metabolites of interest (Figure 5).
3. Discussion
In this study, we showed that 6-OHDA-induced PD-like pathology resulted in altered cardiac metabolite expression in rats. By profiling the metabolites in cardiac tissue, beta-sitosterol, campesterol, monoacylglycerol, beta-glycerophosphoric acid, o-phosphoethanolamine, myo-inositol-1-phosphate, alanine, valine, allothreonine, α-tocopherol, stearic acid and cholesterol, were identified as those metabolites that significantly discriminated Parkinsonian rats and sham counterparts. Energy metabolism is central to cardiac function and the dynamic ability of this organ to switch metabolic substrates to meet its energy need is well established. The heart has more lipid fluxes than other body organs, possibly with the exception of the liver [4]. Most abundantly expressed in cardiac tissue, lipoprotein lipase is a crucial molecule in lipid metabolism that catalyses the hydrolysis of triacylglycerols (or triglycerides), resulting in the production of free fatty acids and monoacylglycerol (MG) [5]. Decreased lipid metabolism in the heart can lead to triacylglycerol accumulation, leading to abnormal lipid signaling [6], overproduction of reactive oxygen species and mitochondrial dysfunction. Therefore it is critical to maintain the heart´s metabolic flexibility [7]. In this study, increased cardiac MG levels (MG; 0:0/18:1(9Z)/0:0) were found in 6-OHDA-lesioned rats compared to sham therapy. The MG pathway of fat absorption is a key process to resynthesize triglycerides. A study using metabolomics analysis found significantly elevated serum MG levels in PD patients compared to healthy controls, suggesting higher lipid metabolism associated with the disease [8]. The importance of the pathogenic substrates of PD are still unclear but a pivotal role in lipid metabolism is currently emerging [9]. In a recent study of lipidome analysis of the catecholaminergic neuronal cell line SH-SY5Y treated with 6-OHDA, a model of PD, increased levels of unsaturated lipids have been found in accordance with prior research using biological samples of PD patients [10].
Results from metabolomics-based studies in PD have shown that metabolic pathways associated with lipid metabolism are among those most affected by the disease [11]. It is worth noting that triglycerides, sterols and phopholipids are lipids of nutritional importance. Since lipid metabolism is affected by dietary components, it should be noted that all rats of the present work were fed a solid formulation diet for laboratory rats consisting of corn, wheat, nuts, sunflower seeds, soybean, vegetable oils, enriched with minerals, and vitamins, suggesting that the higher cardiac MG levels found in the Parkinsonian rats may be a reflection of changes in lipid metabolism in PD. In the same line, elevated levels of phytosterol metabolite (campesterol and sitosterol) were found in the hearts of Parkinsonian rats in our study. Phytosterols or plant sterols, not synthetized in mammalian cells, are structurally similar to cholesterol and obtained exclusively from dietary sources, being found in nuts, seeds and vegetal oils [12]. Campesterol and sitosterol are the most abundant dietary phytosterols in humans. Despite the recognized effect of phytosterol supplementation on lowering both LDL cholesterol and circulating triglycerides, an growing body of data suggests that higher serum concentrations of these natural plant compounds are causally associated with increased risk of cardiovascular diseases [13,14,15].
There’s a long body of research supporting the atherogenic effect of phytosterols [15]. Phytosterolemia or sitosterolemia is a rare genetic condition that causes coronary heart disease, aortic valve disease and premature death in untreated humans [16]. In line with human findings, results from studies in animal models of phytosterolemia pointed to cardiovascular outcomes in mice including cardiac fibrosis, impaired cardiac function, arrhythmias and sudden cardiac death [13]. Findings from studies on the impact of phytosterols against inflammation are still inconsistent [17,18]. On the other hand, phytosterols robustly impact cell membrane dynamics by regulating lipid rafts, fluidity, permeability and curvature [19]. Research indicates that β-sitosterol is involved in the inhibition of muscle atrophy and, in conjunction with its anticancer properties, modulates several cell signaling pathways such as apoptosis, proliferation, survival and angiogenesis [19,20,21]. The cardiac accumulation of phytosterols in Parkinsonian rats likely affects several cellular signaling processes that can impair heart function. In the coming years, it will be decisive to clarify how such changes in cardiac substrate availability lead to processes that in the end may culminate in cardiac dysfunction.
In this reasoning, our results showed that PD-like pathology changed the concentrations of cholesterol, an important fat-like substance, that was found to be increased in the hearts of 6-OHDA rats. Cholesterol plays a central role in a range of cellular processes and impairments, and its biosynthesis has been widely described in PD [22]. A study based on bilateral brain injection of 6-OHDA showed no significant differences in plasma cholesterol levels between Parkinsonian rats and control counterparts, while a work using 6-OHDA unilateral lesion found reduced concentrations of cholesterol ester in the cerebrospinal fluid from neurotoxin-lesioned rats [23,24]. Although available data from studies clearly indicate that higher serum cholesterol concentrations can increase the risk of heart disease, there is growing evidence that elevated serum levels of cholesterol are associated with reduced prevalence of PD and are even beneficial for decelerating clinical progression [25,26]. Moreover, our study identified increased amounts of stearic acid, a saturated long-chain fatty acid, in cardiac tissue of 6-OHDA rats. Shah and colleagues [27] investigating the plasma metabolic profile in the unilateral model of 6-OHDA also detected increased levels of this metabolite in the Parkinsonian rats. We also found increased levels of α-tocopherol (vitamin E), a potent lipophilic antioxidant in 6-OHDA-treated animals, probably pointing to a counter-regulatory mechanism against damage provoked by free radicals in the heart [28]. Other studies described that pretreatment with α-tocopherol provided a neuroprotective role in the 6-OHDA model [29,30]. The metabolomics analysis of our study revealed a substantial increase in phosphoethanolamine in the cardiac tissues of Parkinsonian rats. Organic phosphoethanolamine is a substance naturally synthesized by living cells that is deeply involved in phospholipid turnover and in the lipid signaling pathways. Kataoka and colleagues assessed the developmental patterns of phosphoethanolamine expression in the heart, brain, and kidney of mice from 5 to 110 days old and showed that phosphoethanolamine levels decreased as a function of age up to 24-day-old, then remained constant, suggesting that tissue concentrations of phosphoethanolamine are regulated in adulthood [31]. A study in which the heart of patients during cardioplegic cardiac arrest was examined using microdialysis for in-vivo monitoring of extracellular phosphoethanolamine levels to assess degradation of membrane phospholipids associated to myocardial pathophysiology, revealed a significant 25-fold increase in phosphoethanolamine levels in just 20 minutes after cardiac arrest [32].
It has been also shown that the rate of membrane phospholipid synthesis is impacted in PD, and findings from a human postmortem study showed an increase in the activity of key phospholipid biosynthesis enzymes, such as phosphoethanolaminecytidylyltransferase, in the substantia nigra of PD patients [33]. For this reason, increased phospholipid turnover was reported in a previous study using the unilateral 6-OHDA rat model of PD, which showed increased uptake of radiolabeled arachidonic acid in ipsilateral basal ganglia circuit structures compared to those from the contralateral hemisphere of Parkinsonian animals, reflecting increased phospholipase A2 activity [34]. Beta-Glycerophosphoric acid, also known as β-glycerophosphate, is another metabolite that significantly discriminates Parkinsonian rats and sham counterparts and was found elevated in the heart of 6-OHDA animals. In addition to changing crucial parameters of mitochondrial respiration in vascular smooth muscle cells, β-glycerophosphate induced mitochondrial-dependent oxidative stress and also contributed to calcification by modifying cellular bioenergetics [35]. A recent study showed that obesity-related metabolomic signatures are correlated with clinical and inflammatory marks of asthmatic patients and that beta-glycerophosphoric acid was among the most significant differential metabolites identified in the serum of obese asthmatic individuals compared to lean asthmatics, with increased expression in the obese group [36]. Here, we also encountered that significantly changed cardiac metabolites were related to amino acid metabolites (allothreonine, alanine and valine), which were found to be markedly decreased in Parkinsonian rats compared with sham animals. A nonessential amino acid in humans, allothreonine, beyond serving as a substrate for the enzyme serine hydroxymethyltransferase1, which catalyzes the reversible and simultaneous interconversion of serine and glycine, also exerts pivotal roles in fat and fatty acid metabolism [37]. A study using metabolomics to assess the metabolic profile of amino acids in atrial fibrillation, the predominant type of cardiac arrhythmia, found that plasma allothreonine was among the significantly altered metabolites between patients and control, and had diagnostic implications for atrial fibrillation [38]. Plasma metabolomics was also used by Shoaib and colleagues to compare metabolic changes between human patients and a rat model of cardiac arrest, and similar patterns of change were found, with allothreonine among the analyzed metabolites [39]. Allothreonin stood out as a significantly altered amino acid in a study examining the effect of a 6-month exercise intervention on serum metabolites in men with insomnia [40]. Myo-Inositol 1-phosphate is a crucial intermediate in the biosynthesis of inositol in mammals. The rate limiting step in inositol synthesis is the conversion of glucose 6-phosphate to myo-inositol-1-phosphate, which is the first step in the generation of all inositol-containing elements, encompassing phospholipids. Myo-inositol 1-phosphate is crucial for the biosynthesis of myo-inositol, a constituent of animal cells and a key growth-promoting factor that has central roles in a wide range of metabolic and regulatory biochemical processes, including lipid, glucose and insulin metabolism, osmoregulation, and many others [41,42,43].
The MetaboAnalyst platform was used to identify the relevant metabolic pathways to interpret our results in a biologically meaningful scenario from the concentration patterns of the cardiac metabolites that discriminated sham and Parkinsonian rats. The biosynthesis of both phosphatidylethanolamine (PE) and phosphatidylcholine (PC) have been identified as relevant metabolic pathways in the present study. Crucial for a great variety of cellular processes, the glycerophospholipids PC and PE are the two most abundant phospholipids in eukaryotic cells and in addition to their central roles in the structure and function of membranes, they are essential regulators of lipids and cell energy metabolism [44,45]. The cytidinediphosphate (CDP)- ethanolamine and CDP-choline are branches of the Kennedy pathways employed by mammalian cells for PE and PC biosynthesis in the endoplasmic reticulum, representing major biosynthetic routes in the formation of these phospholipids [46,47]. Alterations in tissue expression of PC and PE have been involved in metabolic disorders including atherosclerosis [45]. Comprising roughly 20% of membrane phospholipids, plasmalogens are a particular subclass of glycerophospholipids containing a sn-1 vinyl ether linkage plus an ester bond at sn-1 and sn-2 positions of the glycerol backbone, playing a diversity of cellular functions [52]. Like other lipid mediators, plasmalogens are vital molecular messengers involved in inflammatory responses and defects in their synthesis are implicated in metabolic disorders and neurodegenerative diseases [48].
The metabolism of phospholipids is closely linked with that of other lipids within the cell. The glucose-alanine cycle, otherwise known as the alanine (a glucogenic amino acid) cycle or Cahill cycle, links carbohydrate metabolism to amino acid metabolism and mediates the degradation of muscle protein in order to deliver extra glucose to produce more ATP to sustain muscle contraction, regulating energy metabolism [49,50]. Found in all mammalian tissues, glutathione is a rich antioxidant synthesized through two sequential reactions in the cytosol through the action of ATP-dependent enzymes: first, γ-glutamylcysteine is formed from L-glutamate and cysteine, catalyzed by glutamate–cysteine ligase; the second reaction produces glutathione by adding glycine to the C-terminal of γ-glutamylcysteine, a step catalyzed by glutathione synthase [51]. Glutathione is involved in biological processes such as redox reactions, cell homeostasis and is a modulator of cell proliferation, immune response, apoptosis, and others [52]. Furthermore, glutathione system dysfunction has been implicated in brain disorders, including PD [53]. Inositol phosphates are an acclaimed group of small signaling molecules ubiquitously found in eukaryote cells, which exhibit diverse biological activities, with emphasis on energy homeostasis and anti-inflammatory actions [54]. Therefore, many studies have been focused on investigating the potential activity of these molecules for the treatment of chronic inflammation and cardiovascular diseases [55,56]. The biological and pharmacological properties of inositol phosphates rely on the specific phosphate group content in the inositol ring, required for the precise propagation of cellular information. Moreover, the phosphorylated derivates of inositol phosphates are the inositol pyrophosphates, a class of high-energy molecules with a range of metabolic and signaling functions [54,57].
In conclusion, our study found that both lipid and energy metabolism are deeply implicated in cardiac metabolic changes in PD-like pathologies. Our results provide new insights into cardiac metabolism in experimental PD and indicate potential targets for further investigation. As complex multicellular organisms, mammalian metabolism involves interconnected organ systems. In this study, we focused on the metabolic changes associated with PD in a specific tissue, the heart. Future studies should adopt a more integrated approach focused on exploring inter-tissue communication. Therefore, the next steps to gain new insights into PD pathogenesis should involve applying a network-based approach to uncover PD related multi-tissue crosstalk. Future advancements in omics and mutli-omics research complemented by the systematic study of biological species, including samples from PD patients, will advance our understanding and lead to a greater recognition of the importance of the regulatory components for this disabling disease.
4. Materials and Methods
4.1. Animals
The rats were provided by the Center for Development of Experimental Models for Medicine and Biology (CEDEME)/Universidade Federal de São Paulo and were kept in the bioterium of the Neuroscience laboratory, housed in groups of 4 per cage with sawdust as bedding. The cages were maintained under a 12h light-dark cycle (light: 7:00–19:00), temperature at 21 ± 2 °C with food and water ad libitum. Sixteen adult male Wistar rats weighing between 230-300g and 8 weeks of age were used. Animals were distributed in two experimental groups: sham and 6-OHDA. All experiments followed the guidelines of the Ethics Committee of the Federal University of São Paulo to avoid unnecessary animal suffering. The research has the authorization number 7848020920 of the ethics committee on the use of animals (CEUA-Unifesp).
4.2. Study Design
The rats were randomly divided into two groups. In the 6-OHDA group, the animals received bilateral injections of 6-OHDA in the striatum while the sham group received saline solution (vehicle). After 40 days, the brains and hearts were removed for immunohistochemistry and metabolomics, respectively.
4.3. 6-OHDA Lesion
Fifteen minutes before stereotactic surgery, the animals were anesthetized intraperitoneally with a single injection of ketamine (100 mg/kg) and xylazine (10 mg/kg) and after complete anesthesia, they were fixed in a stereotaxic device (EFF 331-Insight™, RibeirãoPreto, São Paulo, Brazil). A 10 µL Hamilton syringe attached to the stereotaxic rod was used to inject 1 µL of 6-OHDA solution (Sigma©, Saint Louis, Missouri, USA) (12 µg/µL concentration in 0.3% ascorbic acid) or saline solution in four different coordinates referent to the bregma: (1) latero-lateral: −2.7 mm, anteroposterior: bregma, dorsoventral: −4.5 mm; (2) latero-lateral: −3.2 mm, anteroposterior: +0.5 mm, dorsoventral: −4.5 mm; (3) latero-lateral: +2.7 mm, anteroposterior: bregma, dorsoventral: -4.5 mm; and (4) latero-lateral: +3.2 mm, anteroposterior: +0.5 mm, dorsoventral: -4.5 mm, according to the rat brain atlas by Paxinos and Watson [58]. Bilateral administration of the neurotoxin 6-OHDA, which is structurally similar to catecholamines, to the dorsal striatum causes death of dopaminergic neurons in the substantia nigra pars compacta (SNc), mimicking Parkinson's pathology [59,60].
4.4. Immunohistochemistry
The rats were anesthetized with lidocaine (10 mg/kg i.p.) and sodium thiopental (80 mg/kg i.p.). After complete anesthesia, the rats were decapitated, their hearts were removed and quickly frozen with liquid nitrogen and then stored in a freezer at -80 °C until use. The rat brains were removed and placed in 4% paraformaldehyde solution for 24 hours for fixation and then transferred to phosphate buffered saline (PBS) (0.01 M pH 7.4). Finally, the brains were immersed in a 30% sucrose cryoprotective solution for 2 days, allowing a 40 μΜ coronal cut with a cryostat (Microm HM 505E). Brain tissue slices were washed for 5 minutes in 0.01 M PBS (pH 7.4), treated with 0.1% hydrogen peroxide to remove endogenous peroxides and, after washing, incubated in 10% albumin solution and 0.3% Triton X-100 for 2 hours. After this period, the slices were incubated overnight with primary antibody (1:500 tyrosine hydroxylase (TH), diluted in 0.01 M PBS (pH 7.4) and 2% albumin. The brain slices were treated with biotinylated secondary antibody (anti-rabbit 1:200-Abcam®) diluted in 0.01 M PBS and 2% albumin per 2 hours. The tissue was then washed and incubated with avidinbiotin-peroxidase complex (ABC Elite; Vector Labs, Burlingame, CA, USA). The sections were stained with 3,3'-Diaminobenzidine tetrachloride (DAB) dissolved in TRIS-HCl 0.05 M (PH 7.6) and activated by 0.3% hydrogen peroxide. The images were analyzed using the free ImageJ program to indicate cell nuclei and fibers in the striatum and substantia nigra region.
4.5. Metabolomics
For the metabolomic analyses, the hearts stored in the ultra-low freezer (-80ºC) were removed and homogenized in a suitable solution for disrupting lipid molecules (TissueLyser; Qiagen, Germantown, MD). For gas chromatography analysis (GC-MS), aliquots of 100 uL samples from both groups were shaken with pure methanol in a 1:3 ratio in such a way that deproteinization occurred and centrifuged at high speed. 100uL of the supernatant was placed in GC-MS vials with glass pellets for the derivatization process. In the methoximation step, the solvent was evaporated at 30°C in a SpeedVac and the O-methoxyamine hydrochloride (15mg/mL) in pyridine was placed in the flasks, vortexed and incubated for 16 hours (dark and at room temperature). After this time, silylation with 10uL of BSTFA (1% TMCS (v/v)) started and the samples were incubated at 70°C for 1 hour. Finally, 100uL of heptane (with 20 ppm of internal standard pentadecanoic acid) were added to each analysis vial. Blanks were processed together to correct the baseline chromatograms [61]. Analyzes were performed on a quadrupole-type GCMS-QP2020NX system (Shimadzu Co., Kyoto, Japan), with 1uL of the sample loaded onto a DB5-MS column (30m x 0.25 mm, 0.25 µm, Restek) and injected in splitless mode with helium gas flow (20 mL/min). Carrier gas was conducted at a constant flow of 1.36 ml/min. The initial column temperature was 100°C followed by increasing the temperature at 15°C/min until it reached 300°C. The temperature was then left at 300°C for five minutes before cooling. The injector (280°C), transfer line (200°C) and quadrupole source filament (150°C) temperatures remained unchanged. The system operated in full scan mode (40-650 m/z and 3 spectra/s) with the EI set to 70eV. LabSolutions software (GCMS version 4.5, Shimadzu Co., Japan), allows real-time control of each analyte analyzed to label the metabolites in the Scan. The detected metabolites were processed to create a unified matrix with the different charge states, adducts and groups of the same analytes from all samples using GCMS Solution (v.3.30), NIST 17 MASS (v.1.00.1) and GCMS software Smart Metabolite (v.3.01), all developed by Shimadzu Co. After identification of molecules by NIST [14] and Smart Metabolite libraries, samples were exported to Excel (Microsoft Office©) software for statistical treatment using the free online tool available metaboanalyst 5.0. Public databases available on the internet, such as the Human Metabolome Database (hmdb.ca/) were used (www.metlin.scripps.edu, genome.jp/kegg/, www.lipidmaps .org or www.hmdb.ca).
4.6. Statistics
Since the Shapiro-Wilk test showed that the immunohistochemistry was not normally distributed. The study groups were compared simultaneously with the non-parametric Kruskal-Wallis test. Bonferroni's post hoc test determined whether the groups were significantly different from each other. The metabolomic data were normalized, the parametric ANOVA test was used to simultaneously compare the two groups. Tukey's post hoc test determined whether there were significant differences between the groups. The level of statistical significance was set at 0.05. Statistical tests were performed using MATLAB R2017a, GraphPad Prism 5.0 and Metaboanalyst 5.0
Author Contributions
Conceptualization, Carla Alessandra Scorza; Data curation, Valeria de Cassia Gonçalves; Formal analysis, Victor Silva da Fonsêca, Valeria de Cassia Gonçalves, Mario Augusto Izidoro and Fernando Luiz Affonso Fonseca; Funding acquisition, Valeria de Cassia Gonçalves and Carla Alessandra Scorza; Investigation, Valeria de Cassia Gonçalves; Methodology, Victor Silva da Fonsêca and Valeria de Cassia Gonçalves; Supervision, Carla Alessandra Scorza; Writing – original draft, Victor Silva da Fonsêca, Valeria de Cassia Gonçalves, Mario Augusto Izidoro, Antônio-Carlos G. de Almeida, Fernando Luiz Affonso Fonseca, Fulvio Alexandre Scorza, Josef Finsterer and Carla Alessandra Scorza; Writing – review & editing, Valeria de Cassia Gonçalves, Mario Augusto Izidoro, Antônio-Carlos G. de Almeida, Fernando Luiz Affonso Fonseca, Fulvio Alexandre Scorza, Josef Finsterer and Carla Alessandra Scorza.
Funding
The present study was financially supported by the São Paulo Research Foundation (FAPESP) through grant 2020/12791-6. This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Brazil (CAPES), Finance Code 001; and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq; Process 304268/2020-8). VCG. and VSF received CAPES scholarship.
Acknowledgments
The authors would like to thank Shimadzu do Brasil, who has kindly lent us the GC-MS-QP2020NX.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hou, Y.; Dan, X.; Babbar, M.; Wei, Y.; Hasselbalch, S.G.; Croteau, D.L.; Bohr, V.A. Ageing as a risk factor for neurodegenerative disease. Nat. Rev. Neurol. 2019, 15, 565–581. [Google Scholar] [CrossRef] [PubMed]
- Ou, Z. , Pan, J., Tang, S., Duan, D., Yu, D., Nong, H., Wang, Z. 2021. Global trends in the incidence, prevalence, and years lived with disability of Parkinson's disease in 204 countries/territories from 1990 to 2019. Frontiers in public health 9: 776847.
- Değirmenci, H. , Bakirci, E.M., Hamur, H. 2020. Cardiac Effects of Parkinson’s Disease. Open Journal of Parkinson's Disease and Treatment 3(1): 006-007.
- Park, T.-S.; Yamashita, H.; Blaner, W.S.; Goldberg, I.J. Lipids in the heart: a source of fuel and a source of toxins. Curr. Opin. Infect. Dis. 2007, 18, 277–282. [Google Scholar] [CrossRef]
- Kumari, A.; Kristensen, K.K.; Ploug, M.; Winther, A.L.-M. The Importance of Lipoprotein Lipase Regulation in Atherosclerosis. Biomedicines 2021, 9, 782. [Google Scholar] [CrossRef]
- Goldberg, I.J.; Trent, C.M.; Schulze, P.C. Lipid Metabolism and Toxicity in the Heart. Cell Metab. 2012, 15, 805–812. [Google Scholar] [CrossRef]
- Pascual, F.; Coleman, R.A. Fuel availability and fate in cardiac metabolism: A tale of two substrates. Biochim. et Biophys. Acta (BBA) - Mol. Cell Biol. Lipids 2016, 1861, 1425–1433. [Google Scholar] [CrossRef]
- Toczylowska, B.; Zieminska, E.; Michałowska, M.; Chalimoniuk, M.; Fiszer, U. Changes in the metabolic profiles of the serum and putamen in Parkinson’s disease patients – In vitro and in vivo NMR spectroscopy studies. Brain Res. 2020, 1748, 147118. [Google Scholar] [CrossRef]
- Fanning, S.; Selkoe, D.; Dettmer, U. Parkinson’s disease: proteinopathy or lipidopathy? npj Park. Dis. 2020, 6, 1–9. [Google Scholar] [CrossRef]
- Xicoy, H.; Brouwers, J.F.; Kalnytska, O.; Wieringa, B.; Martens, G.J.M. Lipid Analysis of the 6-Hydroxydopamine-Treated SH-SY5Y Cell Model for Parkinson’s Disease. Mol. Neurobiol. 2019, 57, 848–859. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Le, W. Recent advances and perspectives of metabolomics-based investigations in Parkinson’s disease. Mol. Neurodegener. 2019, 14, 1–12. [Google Scholar] [CrossRef]
- Poli, A. , Marangoni, F., Corsini, A., Manzato, E., Marrocco, W., Martini, D., Medea, G., Visioli, F. 2021. Phytosterols, Cholesterol Control, and Cardiovascular Disease. Nutrients 13(8), 2810.
- Ge, H.; Liu, G.; Yamawaki, T.M.; Tao, C.; Alexander, S.T.; Ly, K.; Fordstrom, P.; Shkumatov, A.A.; Li, C.-M.; Rajamani, S.; et al. Phytosterol accumulation results in ventricular arrhythmia, impaired cardiac function and death in mice. Sci. Rep. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Makhmudova, U.; Schulze, P.C.; Lütjohann, D.; Weingärtner, O. Phytosterols and Cardiovascular Disease. Curr. Atheroscler. Rep. 2021, 23, 1–8. [Google Scholar] [CrossRef]
- Scholz, M. , Horn, K., Pott, J., Gross, A., Kleber, M.E., Delgado, G.E., Mishra, P.P., Kirsten, H., Gieger, C., Müller-Nurasyid, M., Tönjes, A., Kovacs, P., Lehtimäki, T., Raitakari, O., Kähönen, M., Gylling, H., Baber, R., Isermann, B., Stumvoll, M., Loeffler, M., März, W., Meitinger, T., Peters, A., Thiery, J., Teupser, D., Ceglarek, U. 2022. Genome-wide meta-analysis of phytosterols reveals five novel loci and a detrimental effect on coronary atherosclerosis. Nature communications 13(1), 143.
- Sudhop, T.; Gottwald, B.M.; von Bergmann, K. Serum plant sterols as a potential risk factor for coronary heart disease. Metabolism 2002, 51, 1519–1521. [Google Scholar] [CrossRef] [PubMed]
- A Othman, R.; Moghadasian, M.H. Beyond cholesterol-lowering effects of plant sterols: clinical and experimental evidence of anti-inflammatory properties. Nutr. Rev. 2011, 69, 371–382. [Google Scholar] [CrossRef]
- Stanasila, L.; Marques-Vidal, P. Serum Phytosterols Are Not Associated with Inflammatory Markers in Two Cross-Sectional, Swiss Population-Based Studies (The CoLaus|PsyCoLaus Study). Nutrients 2022, 14, 2500. [Google Scholar] [CrossRef] [PubMed]
- Fakih, O.; Sanver, D.; Kane, D.; Thorne, J.L. Exploring the biophysical properties of phytosterols in the plasma membrane for novel cancer prevention strategies. Biochimie 2018, 153, 150–161. [Google Scholar] [CrossRef]
- Hah, Y.-S.; Lee, W.K.; Lee, S.; Kim, E.J.; Lee, J.H.; Lee, S.-J.; Ji, Y.H.; Kim, S.G.; Lee, H.-H.; Hong, S.Y.; et al. β-Sitosterol Attenuates Dexamethasone-Induced Muscle Atrophy via Regulating FoxO1-Dependent Signaling in C2C12 Cell and Mice Model. Nutrients 2022, 14, 2894. [Google Scholar] [CrossRef]
- Khan, Z.; Nath, N.; Rauf, A.; Bin Emran, T.; Mitra, S.; Islam, F.; Chandran, D.; Barua, J.; Khandaker, M.U.; Idris, A.M.; et al. Multifunctional roles and pharmacological potential of β-sitosterol: Emerging evidence toward clinical applications. Chem. Interactions 2022, 365, 110117. [Google Scholar] [CrossRef]
- Xicoy, H.; Wieringa, B.; Martens, G.J.M. The Role of Lipids in Parkinson’s Disease. Cells 2019, 8, 27. [Google Scholar] [CrossRef]
- Gomes, F.A.; Flores, R.A.; Bruxel, M.A.; da Silva, F.N.; Moreira, E.L.G.; Zoccal, D.B.; Prediger, R.D.; Rafacho, A. Glucose Homeostasis Is Not Affected in a Murine Model of Parkinson’s Disease Induced by 6-OHDA. Front. Neurosci. 2019, 12, 1020. [Google Scholar] [CrossRef]
- Qiu, J.; Peng, G.; Tang, Y.; Li, S.; Liu, Z.; Zheng, J.; Wang, Y.; Liu, H.; Wei, L.; Su, Y.; et al. Lipid profiles in the cerebrospinal fluid of rats with 6-hydroxydopamine-induced lesions as a model of Parkinson’s disease. Front. Aging Neurosci. 2023, 14, 1077738. [Google Scholar] [CrossRef]
- Fu, X.; Wang, Y.; He, X.; Li, H.; Liu, H.; Zhang, X. A systematic review and meta-analysis of serum cholesterol and triglyceride levels in patients with Parkinson’s disease. Lipids Heal. Dis. 2020, 19, 1–10. [Google Scholar] [CrossRef]
- Hurh, K.; Park, M.; Jang, S.-I.; Park, E.-C.; Jang, S.-Y. Association between serum lipid levels over time and risk of Parkinson’s disease. Sci. Rep. 2022, 12, 1–9. [Google Scholar] [CrossRef]
- Shah, A.; Han, P.; Wong, M.-Y.; Chang, R.C.-C.; Legido-Quigley, C. Palmitate and Stearate are Increased in the Plasma in a 6-OHDA Model of Parkinson’s Disease. Metabolites 2019, 9, 31. [Google Scholar] [CrossRef]
- van der Loo, B.; Labugger, R.; Aebischer, C.P.; Skepper, J.N.; Bachschmid, M.; Spitzer, V.; Kilo, J.; Altwegg, L.; Ullrich, V.; Lüscher, T.F.; et al. Cardiovascular aging is associated with vitamin E increase. Circulation 2002, 105, 1635–1638. [Google Scholar] [CrossRef]
- Heim, C.; Kolasiewicz, W.; Kurz, T.; Sontag, K.H. Behavioral alterations after unilateral 6-hydroxydopamine lesions of the striatum. Effect of alpha-tocopherol. Pol. J. Pharmacol. 2002, 53. [Google Scholar]
- Roghani, M.; Behzadi, G. Neuroprotective effect of vitamin E on the early model of Parkinson’s disease in rat: behavioral and histochemical evidence. Brain Res. 2001, 892, 211–217. [Google Scholar] [CrossRef]
- Kataoka, H. , Maeda, M., Makita, M. 1991. O-phosphoethanolamine content in mouse tissues during development. Agricultural and biological chemistry 55(1): 289-290.
- Kennergren, C.; Mantovani, V.; Lönnroth, P.; Nyström, B.; Berglin, E.; Hamberger, A. Extracellular Amino Acids as Markers of Myocardial Ischemia during Cardioplegic Heart Arrest. Cardiology 1999, 91, 31–40. [Google Scholar] [CrossRef]
- Ross, B.M. , Mamalias, N., Moszczynska, A., Rajput, A.H., Kish, S.J. 2001. Elevated activity of phospholipid biosynthetic enzymes in substantia nigra of patients with Parkinson's disease. Neuroscience 102(4), 899–904.
- Hayakawa, T. , Chang, M.C., Bell, J.M., Seeman, R., Rapoport, S.I., Appel, N.M. 1998. Fatty acid incorporation depicts brain activity in a rat model of Parkinson's disease. Brain research 807(1-2), 177–181.
- Alesutan, I.; Moritz, F.; Haider, T.; Shouxuan, S.; Gollmann-Tepeköylü, C.; Holfeld, J.; Pieske, B.; Lang, F.; Eckardt, K.-U.; Heinzmann, S.S.; et al. Impact of β-glycerophosphate on the bioenergetic profile of vascular smooth muscle cells. J. Mol. Med. 2020, 98, 985–997. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zheng, J.; Zhang, H.P.; Zhang, X.; Wang, L.; Wood, L.; Wang, G. Obesity-Associated Metabolic Signatures Correlate to Clinical and Inflammatory Profiles of Asthma: A Pilot Study. Allergy, Asthma Immunol. Res. 2018, 10, 628–647. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Yin, C.; Tian, Z.; Wang, T.; Sun, W.; Xiang, Q.; Guo, G. Metabolomic Analysis of Plasma From Patients With Acute Mountain Sickness Using Chromatography–Mass Spectrometry. Medicine 2015, 94, e1988–e1988. [Google Scholar] [CrossRef] [PubMed]
- She, J.; Guo, M.; Li, H.; Liu, J.; Liang, X.; Liu, P.; Zhou, B.; Liu, S.; Deng, Y.; Lou, B.; et al. Targeting amino acids metabolic profile to identify novel metabolic characteristics in atrial fibrillation. Clin. Sci. 2018, 132, 2135–2146. [Google Scholar] [CrossRef]
- Shoaib, M.; Choudhary, R.C.; Choi, J.; Kim, N.; Hayashida, K.; Yagi, T.; Yin, T.; Nishikimi, M.; Stevens, J.F.; Becker, L.B.; et al. Plasma metabolomics supports the use of long-duration cardiac arrest rodent model to study human disease by demonstrating similar metabolic alterations. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, X.; Le, S.; Ojanen, X.; Tan, X.; Wiklund, P.; Cheng, S. Effects of exercise and dietary interventions on serum metabolites in men with insomnia symptoms: A 6-month randomized controlled trial. Sports Med. Heal. Sci. 2020, 2, 95–101. [Google Scholar] [CrossRef]
- Fisher, S.K.; Novak, J.E.; Agranoff, B.W. Inositol and higher inositol phosphates in neural tissues: homeostasis, metabolism and functional significance. J. Neurochem. 2002, 82, 736–754. [Google Scholar] [CrossRef]
- Chhetri, D.R. Myo-Inositol and Its Derivatives: Their Emerging Role in the Treatment of Human Diseases. Front. Pharmacol. 2019, 10, 1172. [Google Scholar] [CrossRef]
- Gonzalez-Uarquin, F.; Rodehutscord, M.; Huber, K. Myo-inositol: its metabolism and potential implications for poultry nutrition—a review. Poult. Sci. 2019, 99, 893–905. [Google Scholar] [CrossRef]
- Calzada, E.; Onguka, O.; Claypool, S.M. Phosphatidylethanolamine Metabolism in Health and Disease. 2015, 321, 29–88. [CrossRef]
- van der Veen, J.N.; Lingrell, S.; da Silva, R.P.; Jacobs, R.L.; Vance, D.E. The Concentration of Phosphatidylethanolamine in Mitochondria Can Modulate ATP Production and Glucose Metabolism in Mice. Diabetes 2014, 63, 2620–2630. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.; Witt, S.N. Ethanolamine and Phosphatidylethanolamine: Partners in Health and Disease. Oxidative Med. Cell. Longev. 2017, 2017, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Tavasoli, M.; Lahire, S.; Reid, T.; Brodovsky, M.; McMaster, C.R. Genetic diseases of the Kennedy pathways for membrane synthesis. J. Biol. Chem. 2020, 295, 17877–17886. [Google Scholar] [CrossRef] [PubMed]
- Bozelli, J.C.; Azher, S.; Epand, R.M. Plasmalogens and Chronic Inflammatory Diseases. Front. Physiol. 2021, 12. [Google Scholar] [CrossRef]
- Sarabhai, T.; Roden, M. Hungry for your alanine: when liver depends on muscle proteolysis. J. Clin. Investig. 2019, 129, 4563–4566. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Kumaran, S.S.; Goyal, V.; Bose, S.; Jain, S.; Dwivedi, S.N.; Srivastava, A.K.; Jagannathan, N.R. Metabolomic analysis of serum using proton NMR in 6-OHDA experimental PD model and patients with PD. Neurochem. Int. 2020, 134, 104670. [Google Scholar] [CrossRef]
- Bachhawat, A.K.; Yadav, S. The glutathione cycle: Glutathione metabolism beyond the γ-glutamyl cycle. IUBMB Life 2018, 70, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Lushchak, V.I. Glutathione Homeostasis and Functions: Potential Targets for Medical Interventions. J. Amino Acids 2012, 2012, 736837. [Google Scholar] [CrossRef]
- Bjørklund, G.; Peana, M.; Maes, M.; Dadar, M.; Severin, B. The glutathione system in Parkinson’s disease and its progression. Neurosci. Biobehav. Rev. 2020, 120, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Chatree, S.; Thongmaen, N.; Tantivejkul, K.; Sitticharoon, C.; Vucenik, I. Role of Inositols and Inositol Phosphates in Energy Metabolism. Molecules 2020, 25, 5079. [Google Scholar] [CrossRef]
- Schantl, A.E.; Verhulst, A.; Neven, E.; Behets, G.J.; D’haese, P.C.; Maillard, M.; Mordasini, D.; Phan, O.; Burnier, M.; Spaggiari, D.; et al. Inhibition of vascular calcification by inositol phosphates derivatized with ethylene glycol oligomers. Nat. Commun. 2020, 11, 1–17. [Google Scholar] [CrossRef]
- Weinberg, S.E.; Sun, L.Y.; Yang, A.L.; Liao, J.; Yang, G.Y. Overview of Inositol and Inositol Phosphates on Chemoprevention of Colitis-Induced Carcinogenesis. Molecules 2020, 26, 31. [Google Scholar] [CrossRef]
- Wundenberg, T.; Mayr, G.W. Synthesis and biological actions of diphosphoinositol phosphates (inositol pyrophosphates), regulators of cell homeostasis. Biol. Chem. 2012, 393, 979–998. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Elsevier Academic Press: Amsterdam, The Netherlands; London, UK, 2007; ISBN 9780123919496. [Google Scholar]
- Shimohama, S. , Sawada, H., Kitamura, Y., Taniguchi, T. 2003. Disease model: Parkinson's disease. Trends in molecular medicine 9(8), 360–365.
- Perlbarg, V.; Lambert, J.; Butler, B.; Felfli, M.; Valabrègue, R.; Privat, A.-L.; Lehéricy, S.; Petiet, A. Alterations of the nigrostriatal pathway in a 6-OHDA rat model of Parkinson’s disease evaluated with multimodal MRI. PLOS ONE 2018, 13, e0202597. [Google Scholar] [CrossRef]
- Mastrangelo, A.; Ferrarini, A.; Rey-Stolle, F.; García, A.; Barbas, C. From sample treatment to biomarker discovery: A tutorial for untargeted metabolomics based on GC-(EI)-Q-MS. Anal. Chim. Acta 2015, 900, 21–35. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Illustrative images of tyrosine hydroxylase (TH) immunoreactivity TH-positive neurons (A,B) and fibers (C,D) in the SNpc and striatum, respectively. Sham group (A,C) and 6-OHDA (B,D). Note the evident reduction of TH-immunostaining in brain slices from the 6-OHDA group compared to the sham group.
Figure 1.
Illustrative images of tyrosine hydroxylase (TH) immunoreactivity TH-positive neurons (A,B) and fibers (C,D) in the SNpc and striatum, respectively. Sham group (A,C) and 6-OHDA (B,D). Note the evident reduction of TH-immunostaining in brain slices from the 6-OHDA group compared to the sham group.
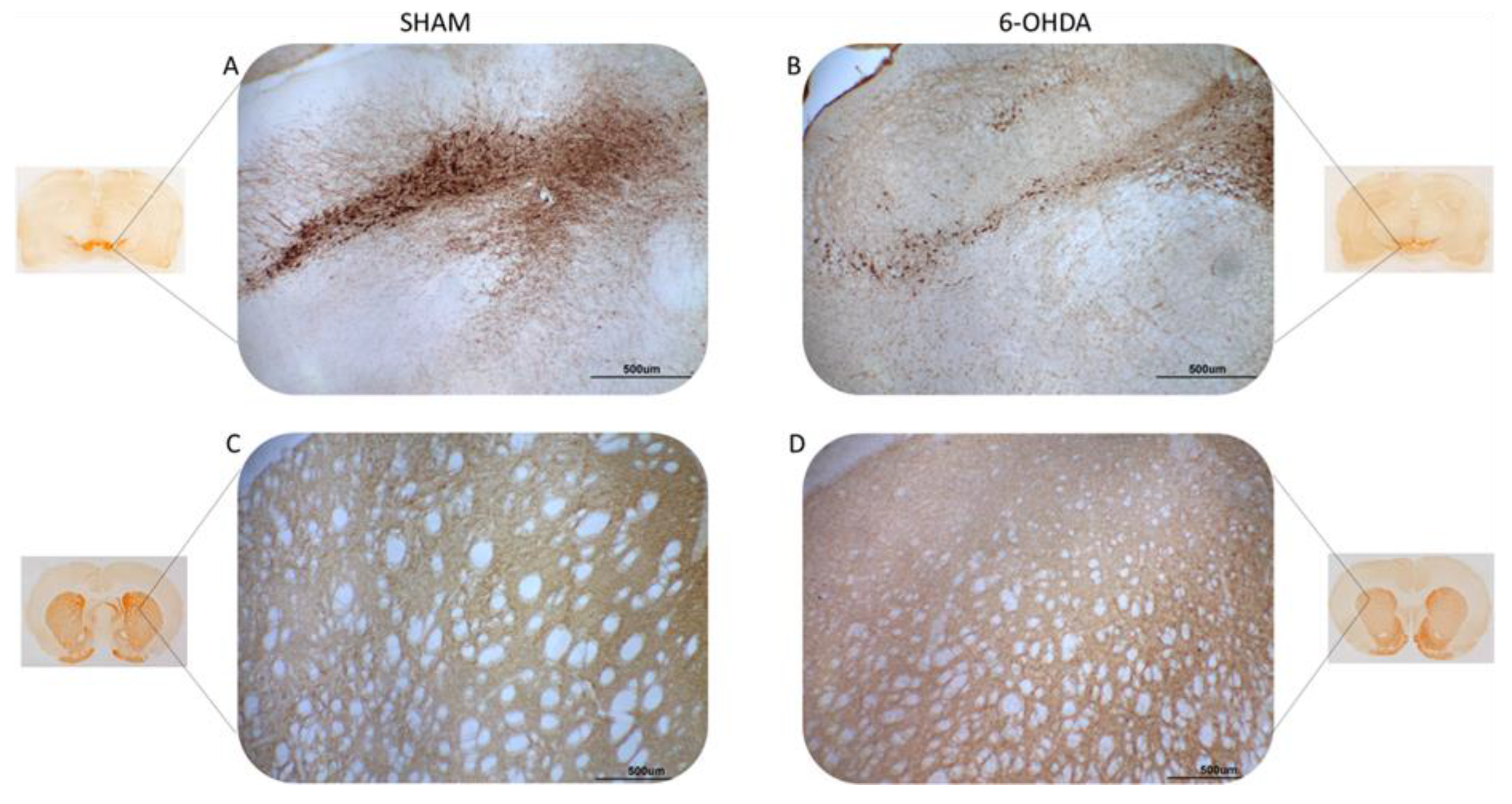
Figure 2.
Statistical results of the Kruskall-Wallis and post hoc Bonferroni for tyrosine hydroxylase (TH) immunohistochemistry analysis. (A) Optical density of the TH-positive fibers in the striatum (chi-square = 8.27, p = 0.004); median and confidence interval were as follows: Sham (550040 ± 86764); 6-OHDA (299536 ± 49874). (B) TH-positive neuronal nuclei in the substantianigra pars compacta (chi-square = 11.76, p = 0.0006); median and confidence interval were as follows: Sham (501 ± 71); 6-OHDA (212 ± 29). The * represents significant difference with p-value < 0.05.
Figure 2.
Statistical results of the Kruskall-Wallis and post hoc Bonferroni for tyrosine hydroxylase (TH) immunohistochemistry analysis. (A) Optical density of the TH-positive fibers in the striatum (chi-square = 8.27, p = 0.004); median and confidence interval were as follows: Sham (550040 ± 86764); 6-OHDA (299536 ± 49874). (B) TH-positive neuronal nuclei in the substantianigra pars compacta (chi-square = 11.76, p = 0.0006); median and confidence interval were as follows: Sham (501 ± 71); 6-OHDA (212 ± 29). The * represents significant difference with p-value < 0.05.
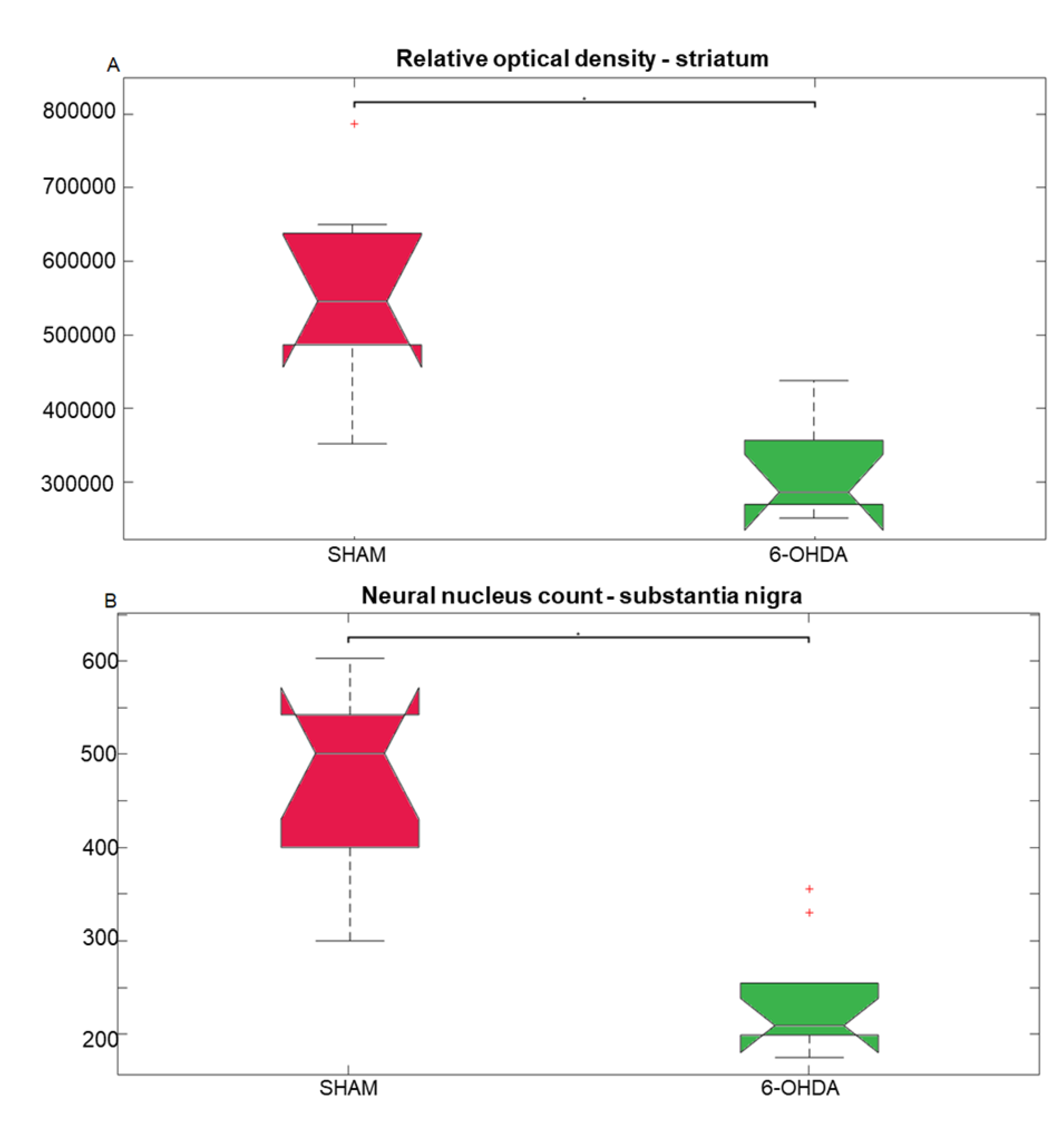
Figure 3.
Multivariate Statistical Analysis of Metabolites between 6-OHDA and Sham groups. (A) PCA analysis to identify the most important variables (PC1, main component one; PC2, main component two) between the two groups and ensure data reliability. (B) PLS plot showing the separation of groups according to their different characteristics in relation to the metabolites (C,D) Main metabolites identified in the cardiac tissue of animals, presented in Feature importance and VIP score.
Figure 3.
Multivariate Statistical Analysis of Metabolites between 6-OHDA and Sham groups. (A) PCA analysis to identify the most important variables (PC1, main component one; PC2, main component two) between the two groups and ensure data reliability. (B) PLS plot showing the separation of groups according to their different characteristics in relation to the metabolites (C,D) Main metabolites identified in the cardiac tissue of animals, presented in Feature importance and VIP score.
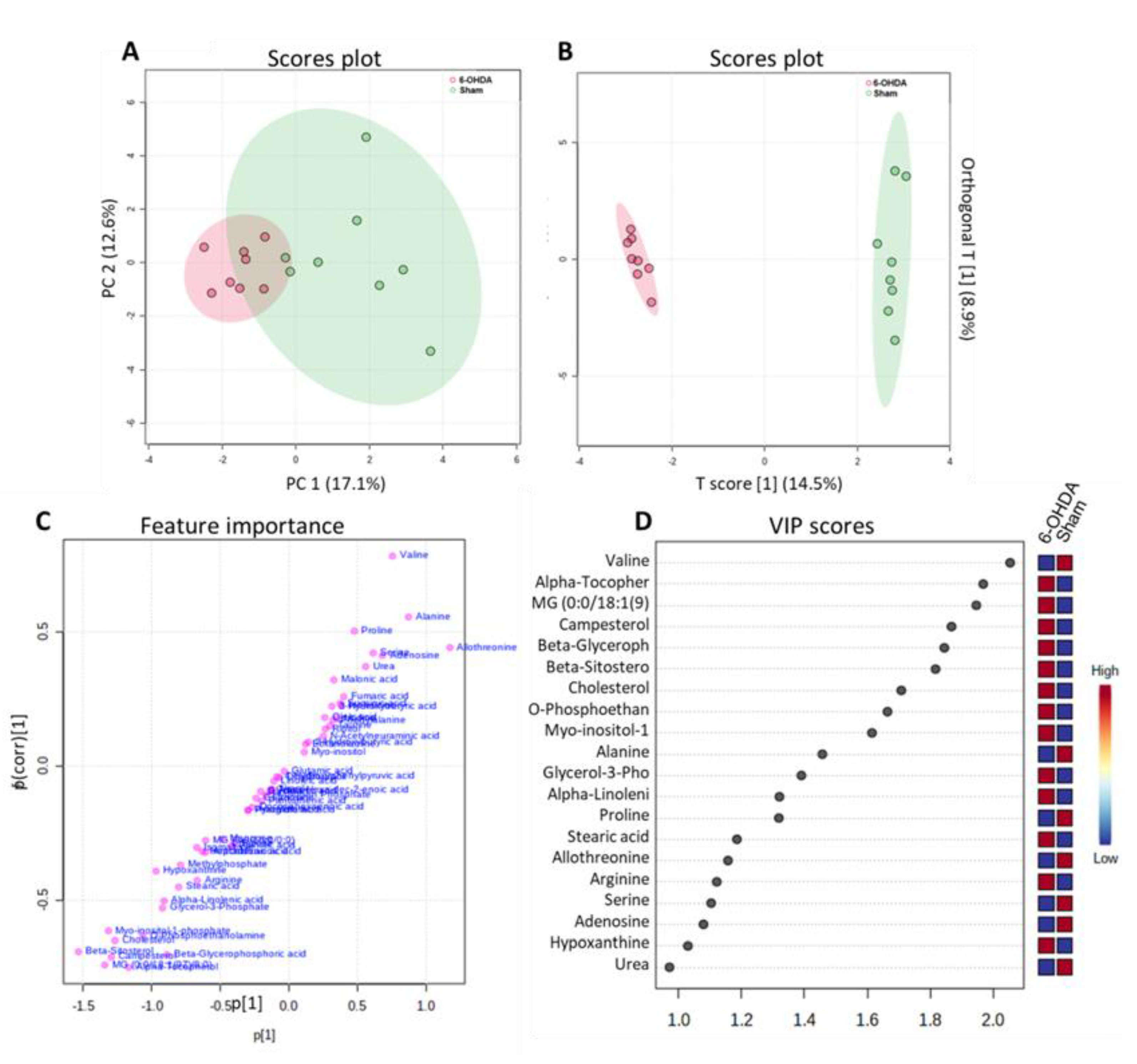
Figure 4.
Statistical analysis of metabolites with higher amounts in the comparison between the two groups. The (p-value, FDR) for the metabolites are: Beta-Glycerophosphoric acid (0.04662, 0.035601); Alpha-Tocopherol (0.001088, 0.020886); Allothreonine (0.004662, 0.035601); Alanine (0.006993, 0.035601); Beta-Sitosterol (0.01865, 0.020886); Campesterol (0.001865, 0.020886); Stearic acid (0.010412, 0.048588); O-Phosphoethanolamine (0.006993, 0.035601); Myo-inositol-1-phosphate (0.006993, 0.035601); MG (0:0/18:1(9Z)/0:0) (0.001088, 0.020886); Cholesterol (0.006993, 0.035601); Valine (0.001865, 0.020886). The * represents significant difference with p-value < 0.05.
Figure 4.
Statistical analysis of metabolites with higher amounts in the comparison between the two groups. The (p-value, FDR) for the metabolites are: Beta-Glycerophosphoric acid (0.04662, 0.035601); Alpha-Tocopherol (0.001088, 0.020886); Allothreonine (0.004662, 0.035601); Alanine (0.006993, 0.035601); Beta-Sitosterol (0.01865, 0.020886); Campesterol (0.001865, 0.020886); Stearic acid (0.010412, 0.048588); O-Phosphoethanolamine (0.006993, 0.035601); Myo-inositol-1-phosphate (0.006993, 0.035601); MG (0:0/18:1(9Z)/0:0) (0.001088, 0.020886); Cholesterol (0.006993, 0.035601); Valine (0.001865, 0.020886). The * represents significant difference with p-value < 0.05.
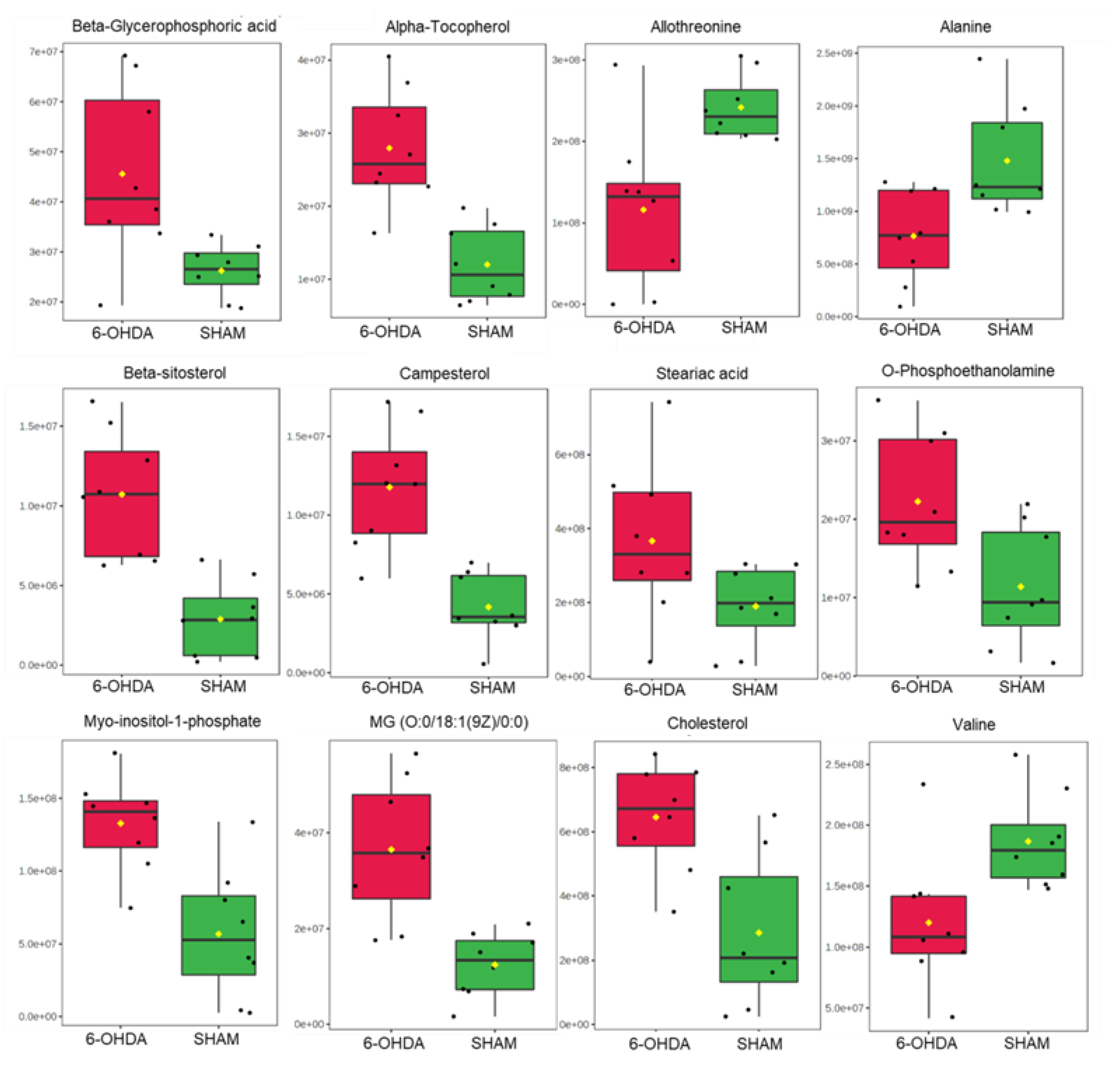
Figure 5.
Analysis of metabolite enrichment between 6-OHDA and Sham groups. The relevant metabolic pathways to the interpretation of the concentration patterns of the metabolites of interest are illustrated.
Figure 5.
Analysis of metabolite enrichment between 6-OHDA and Sham groups. The relevant metabolic pathways to the interpretation of the concentration patterns of the metabolites of interest are illustrated.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



