
Submitted:
16 June 2023
Posted:
19 June 2023
You are already at the latest version
Abstract
Foliar fungal diseases in tomatoes include early blight (Alternaria linariae), Septoria leaf spot (Septoria lycopersici), and late blight (Phytophthora infestans) which is oomycetes. These are one of the significant production constraints in tomatoes. We describe the etiology, host range, distribution, symptoms, and disease cycle to understand the biology followed by management practices emphasizing the resistance breeding approach for these diseases. In crop improvement efforts, we provide an analytical review, including conventional and molecular methods for improving this disease resistance. Modern breeding tools, including genomics, genetic transformation, and genome editing, can be used to improve these traits. There is a good possibility of using these tools in the future to improve these traits.
Keywords:
Foliar fungal diseases
; Integrated disease management
; Marker-assisted selection
; Quantitative trait loci
; Resistance breeding
; Solanum lycopersicum
; Tomato
1. Introduction
Tomato is one of the most important vegetable crops in the world. It is produced in more than five million ha area producing more than 186.8 million mt of tomatoes per year globally [1]. It contributes $1.4 billion to the world economy per annum. Among the top 10 tomato-producing countries, China is the number one producer, followed by India and the US (Table 1). Despite such a massive contribution to the global economy, it is affected by several biotic and abiotic factors posing severe threats to its successful production. The biotic problems include bacterial, fungal, oomycetes, viruses, and root-knot nematodes limiting the production of tomatoes. At the time of infection, pathogens adopt various strategies, including delivery of effector molecules or virulence factors into the host plant so that the host plant's defense becomes weak [2,3]. The defense mechanism is also activated on the host plant side, and biochemical activities occur. As a result, if the pathogens cannot grow on the host tissue, the resistance response occurs.
In contrast, disease development occurs if they can establish the relationship and grow successfully [4]. This review will summarize the information on foliar diseases caused by fungi and oomycetes in tomatoes, the biology of its causal agents, host resistance, genetics, and genomics resources available for managing these diseases. These are the major problems of the organic tomato production system in the US [5]. We will expand the discussion to the use of modern cutting-edge tools, including biotechnology and genome editing, to expedite the genetic improvement process.
1. Foliar diseases caused by fungi and oomycetes
2.1. Early blight
Early blight (EB) is one of the devastating foliar fungal diseases of solanaceous crops caused by Alternaria spp. Early blight is a severe disease in tropical, subtropical, and temperate zones or wherever tomatoes and potatoes are grown. This disease impacts several Solanaceaeous crops, including tomato, potato, and eggplant. The taxonomy of Alternaria species causing early blight is confusing with A. solani, A. linariae, A. tomatophila, and A. alternata. Some Taxonomists consider A. linariae to be equivalent to A. tomatophila and A. solani. Although A. solani and A. linariae are indistinguishable morphologically, single nucleotide polymorphisms (SNPs) molecular markers in the second largest subunit of RNA polymerase (RPB2) may differentiate these two Alternaria species [6]. For this review, we consider A. solani and A. linariae (syn A. tomatophila) to be separate species. A. alternata causes a disease commonly referred to as Stem Canker. Alternaria stem canker is caused by A. alternata f. sp. lycopersici and is primarily a problem of coastal-grown tomatoes in California. Symptoms of Alternaria stem canker can appear on the stems, leaves, and fruit of tomato plants. Many commercial tomato varieties have resistance. Collar rot caused by A. linariae may be confused with stem canker. No sexual stage has been reported for this fungus [7,8]. Alternaria linariae is a necrotrophic fungal pathogen capable of causing severe yield loss under conducive environmental conditions. Chaerani and Voorrips (2006) claim that complete defoliation can result from this disease in areas with high rainfall, high humidity, and relatively high temperatures (24°-30°C). When growing conditions are favorable, the disease can cause declines in yield of 40 to 80%. Different countries have experienced yield losses of up to 79% due to EB damage [9]. As tissues approach vegetative maturity, foliar vulnerability to EB pathogens rises, and fungal damage seems to progress from older to younger leaves from the base of the plant to the top. Even though the causative pathogen is widespread worldwide and can decrease productivity, early blight has never caused widespread hunger or other abrupt and substantially adverse effects on humanity. Alternaria linariae have a broad host range although are reported most frequently on tomatoes and potatoes. Other members of the solanaceous crop family such as eggplant, sweet pepper, and solanaceous weeds (such as black nightshade- Solanum pytcanthum, hairy nightshade- Solanum physalifolium) are also infected by this pathogen [8,10].
Early blight is a global problem that affects tomato and potato crops everywhere. All continents are affected by early blight, which is common in tropical, subtropical, and temperate zones. East of the Rocky Mountains in the US, early blight in tomatoes can be problematic, but it is typically not an issue in the less humid mountain or pacific regions [11]. The EB pathogen can affect every part of the plant and cause various symptoms depending on the crop growth stage. Small brown spots on older leaves that quickly expand are the first sign of an infection. The lesions typically have a yellow halo around them. Dark, concentric rings with a "bullseye" look can be visible within these lesions. Typically, the leaf blight phase starts on the lower, older leaves and works its way up the plant. Similar signs on the stem and fruits can also be visible as the disease progresses. On stems, symptoms include collar rot, stem cankers, and sunken, elliptical-shaped brown lesions with a light center and concentric rings (Figure 1).
Early blight pathogens can survive the winter on infected plant debris and soil for several years as a significant inoculum source for the following growing season. It may also spread through contaminated seeds and transplants. New spores are produced during the next crop season when the environment is favorable—high temperature, enough precipitation, or a relative humidity of 90% or more. Lower leaves become infected when they touch contaminated soil or when showers splash conidia onto the leaves. Conidia will germinate, develop a germ tube, and enter the host through direct penetration of the leaf epidermis by the germ tube or entry through stomata at an ideal temperature of (24-30°C) in the presence of free moisture. Even the desiccated germ tubes can re-grow when re-wetted so infection can happen during alternately wet and dry periods. Environmental factors, leave maturity, and cultivar sensitivity all affect how long it takes foliar symptoms to manifest after the first infection [12].
2.2. Late blight
Late blight (LB), caused by an oomycete, Phytophthora infestans (Mont.) de Bary, is one of the most devastating diseases of tomatoes worldwide. Under favorable weather conditions, tomato and potato crops can be destroyed completely within 7 to 10 days if left untreated [12]. It mainly affects leaves, stems, and fruits of tomato plants. Airborne asexual spores carry most infections during the crop cycle [13].Late blight pathogen (P. infestans) is a hemibiotrophic pathogen with a broad host range in the family Solanaceae, including tomato, potato, and pear melon [14]. Infections are also seen in other crops, such as petunia and nightshade, but considerable economic losses are incurred in potato and tomato crops [15].
Phytophthora infestans is found in almost all the places throughout the world where tomatoes and potatoes are grown. This pathogen is a heterothallic oomycete with two mating types: A1 and A2. Individuals of both mating types A1 and A2 were typical only in Mexico, and clonal lineage (US-1) was predominant in other parts of the world [16]. Isolates of the A2 mating type were first reported in Switzerland [17] and afterward in other countries of Northern Europe [18,19] and also in Japan and Korea subsequently. Due to sexual recombination, the pathogen has gained evolutionary advantages which resulted in increased genetic variation leading to rapid evolution [20,21]. Since the late twentieth century, major migrations have altered the global population structure of P. infestans, adding new genotypes of both mating types all over the world resulting in increased genetic diversity of the pathogen [22]. The exotic strains arise due to substantial migration may not respond to conventional disease treatment strategies as they may have "novel" phenotypic traits which may result in considerable damage of the crops [23].
Late blight disease in tomatoes is identified by the initial appearance of small water-soaked brownish lesions starting from the margin of leaves, which may have chlorotic borders (Figure 2). Such lesions expand rapidly, which causes necrosis in entire leaves. During the high humid condition, the pathogen produces sporangiophores and sporangia on the surface of the infected tissue resulting in the whitish color of sporulation on the lower surface of infected leaves, which is characteristic of the disease. As the disease progresses, such lesions on the leaves coalesced, and the whole foliage looks blighted quickly. The disease primarily causes foliar blight, but it causes black discoloration in the stem and shoulders of ripe fruits in a later stage. If fruits get infected, lesions with brownish or purplish discoloration can be observed [12]. Infected tomatoes have shallow, brown, or purple lesions on the surface of the leaves and fruits [24,25].
The disease cycle of Phytophthora infestans consists of the asexual and sexual phases. The asexual cycle is rapid and is a significant factor in disease development and epidemics in the field, while sexual reproduction leads to genetic recombination resulting in new races [26]. The spread and epidemics of late blight in tomato highly depends upon the relative humidity and temperature during different stages of the life cycle of the pathogen. The pathogen requires nearly 100% RH and an optimum temperature of 24°C to cause widespread damage in the field.
2.3. Septoria leaf spot
Septoria leaf spot (SLS) is a foliar disease of tomatoes caused by Septoria lycopersici, a hemibiotrophic fungus, which occurs wherever tomatoes are grown worldwide. It can cause complete defoliation and significant crop losses, especially in the northeastern US and Canada [27]. To date, only one haplotype of this fungus has been reported [28]. The fungus reproduces through conidia formed in pycnidia discharged in the rain or spray irrigation and spread through water droplets.
Every tomato-growing region where susceptible lines are grown has received reports of the disease. It is prevalent worldwide and most severe when prolonged wet, humid weather conditions occur. This pathogen can infect almost all aerial parts of the plant, with rare incidences on fruits [29]. Although this fungus can affect tomatoes at any growth stage, symptoms typically appear on the older, lower leaves and stem when the first sign of fruit set starts under field conditions. Typical symptoms include small, round, or oval water-soaked spots on the undersurface of lower leaves that gradually turn dark in color and develop to become necrotic lesions with a tan or light grey center (Figure 3). In addition to leaves, petioles, stems, and the calyx can also show symptoms [30]. Necrotic patches on older leaves could have a chlorotic ring/yellow halo surrounding them. Numerous circular spots can be seen on infected leaves. The old spots enlarge and frequently merge, giving a blighted appearance. Spotted leaves eventually turn yellow and die too quickly, causing early defoliation. Defoliation typically starts with the oldest leaves and can quickly move up the plant toward new growth [29].
The leaf fragments and other plant waste from infected plants that overwinter in the soil serve as the principal source of inoculum. The pycnidia of the fungus develop spores when it is moist. In addition to human activities, overhead irrigation systems, splashing rain, and insects can transport these spores to healthy tomato leaves. After this initial dissemination, the spores germinate, and the fungus infects tomato plants through stomata or direct penetration of epidermal cells [31]. Spores can begin to grow in as little as 48 hours in the presence of moisture, leaf spots can form within five days, and pycnidia can appear in 7-10 days. Spores are disseminated by rain or irrigation water and can live in the soil for up to three years on infected tissue [32].
2. Management strategies for fungal diseases
Most plant diseases are managed based on five major principles of plant disease management: avoidance, exclusion, eradication, protection, and resistance. Broadly, we can manage the foliar fungal disease of tomatoes by following cultural practices, chemical control, the use of resistant varieties, and integrated pest management strategies.
3.1. Cultural practices
Tomatoes are attacked by numerous pathogens including many foliar fungal diseases such as early blight, and Septoria leaf spot, and oomycetes causing late blight. The primary source of these diseases is water and airborne propagules of the pathogen, which land on the foliage of the plants and initiate the infection. Such sources can be alternate or collateral hosts in the periphery of the primary host, volunteer tomato crops near the field, or diseased plant debris. In many cases, we can deploy various cultural methods in the field to minimize the entry of pathogen propagules in the area. Some practical and popular cultural methods of foliar disease management in tomatoes are as follows.
Field sanitation refers to all the activities that eliminate the amount of inoculum present in the field as crop debris, weed plants, and alternate and collateral hosts, which helps to prevent the further spread of pathogen propagules to other healthy plants. Most of the foliar disease in tomatoes enters the field through crop residues, fruit leftovers after harvesting, and staking poles. Proper removal of such crop residues and periodic disinfestation of the staking poles by using 0.5% bleach helps to minimize the survival and transmission of the pathogen largely.
Crop Rotation is another crucial cultural method. We are continuously cultivating the same type of crops on the same piece of land, resulting in a higher population of pathogens in the soil. So, crop rotation with unrelated crops is a very effective method to manage soil borne diseases [33].
Crop Management Practices to minimize foliar diseases is advisable. Tomatoes are grown in low fertilized fields or use fertilizers in the improper ratio of NPK and other nutrients resulting in higher crop susceptibility. Such conditions may promote blight, particularly excess or lack of nitrogen. Those fields that are well supplied with potassium may have the opposite effect. Several common foliar diseases in the soil are spread to the plant by splashing water from irrigation and rain. Mulching around plants will help by reducing water-splashed soil and the pathogens that are spread with the particles [34]. Similarly, avoiding sprinkler irrigation and adopting drip irrigation helps to minimize the leaf wetness period in the field, which is correlated with germination and penetration of fungal spores in the leaf tissue.
3.2. Chemical control
Various agrochemicals have been used to manage different foliar fungal diseases in the tomato field. Such chemicals are used as prophylactic as well as curative measures. In general, the chemicals are sprayed before the symptoms appear in the plant. They are continued until the crop harvest, which demands a considerable amount of monetary expenditure from the growers. Most used prophylactic compounds are copper-based fungicides; however, they have undesirable toxicity issues in soilborne microorganisms [35,36]. Synthetic broad-spectrum chemicals like Azoxystrobin, chlorothalonil, difenoconazole, metalaxyl, mancozeb, fluazinam, pyrimethanil, strobilurin, fenamidone, trifloxystrobin, cyprodinil, and chlorothalonil are sprayed in the field either singly or combined for controlling various fungal diseases and oomycetes such as early blight, Septoria leaf spot, and late blight. However, due to the continuous use of such chemicals in the field, the pathogens have developed resistance against chemicals, so it is advisable to rotate different fungicides in the field for resistance management.
3.3. Nanotechnology in plant protection
The application of nanotechnology to agriculture is a relatively new development. Nanomaterials are becoming increasingly popular in crop production as cutting-edge antimicrobial agents, control agent delivery tools, disease detection, and Nano fertilizers to improve plant health. This is particularly true considering that Nanomaterials have unique qualities such as the high surface area to volume ratio ability to exchange ions and chelation, enhanced reaction, unique configuration, huge ion adsorption ratio, dissolve in water, and are less harmful to mammalian cells than conventional antibiotics [37,38]. Additionally, they require simpler preparation and superior chemical stability. Because of their abundance and low toxicity, many nanoparticles are often used as antimicrobial compounds [39]. Only Au, Cu, and Zn-based Nanomaterials are drawing attention, even though several Nanomaterials are efficient against various microorganisms, including bacteria, fungi, and viruses [40]. While certain nanomaterials may strengthen plants' natural defenses against pathogens, others act directly as antibacterial agents. Most nanotechnology studies for controlling plant diseases have used nanoparticles made from metalloids, metallic oxides, nonmetals, and carbon nanomaterials [40,41]. There are numerous examples of the use of nanomaterials for the control of plant pathogens or use as antimicrobial agents, including the use of Ag colloidal against rose powdery mildew [42] and silica (Si) and silver (Ag) nanoparticles mixed with water-soluble polymer against Podosphaera xanthi [43].
Alternaria linariae in tomatoes has been successfully treated using synthetic mesoporous silica nanoparticles [44]. Quick detection and diagnosis of plant diseases have been achieved by using nanomaterials' capacity to conjugate with nucleic acids, proteins, and other biomolecules. For fast disease diagnosis, nanoparticle-based kits and sensors have recently been developed. Quantum dots can potentially be employed in diagnostics since they are programmable fluorescent nanocrystals. For example, [45] created a particular quantum dot-based Nano sensor to detect Candidatus, Phytoplasma aurantifolia in lime. Similarly, Brusca (2003) also created a chip-based hybridization method that uses AgNPs to diagnose Phytophthora species. Further, Fukamachi et al. (2019) reported using poly lactic-co-glycolic acid (PLGA) nanoparticles to encapsulate cyazofamid and develop a precise pesticide delivery system to control the Phytophthora infestans effectively.
As we know, numerous plant micronutrients are crucial for growth and protection against plant diseases. Therefore, essential micronutrients must be available for plants to grow and develop properly. Nanoparticles can use to improve foliar availability and proper micronutrient delivery. Ali et al. (2015) synthesized Ag-based nanoparticles in combination with plant extract, sprinkled them in plants, and saw a decline in the incidence and progression of Phytophthora parasitica and P. capsici disease because of enhanced defense mechanisms in plants.
The use of foliar and soil-applied agrochemicals, adopting resistant cultivars, crop rotation, and other management techniques are examples of current control tactics. Since current methods fail to provide complete crop protection, newer, less expensive, and less harmful technologies like nanomaterials should be investigated more.
Nanotechnology deals with materials smaller than 100 nm, which can be used at the molecular level. These molecules may be Ag-based, C-based, Ce-based, Cu-based, Mg-based, Si-based, or Ti-based, as reported by Bella et al. (2012). This technology has several applications and is being explored in agriculture. Cu-based nanoparticles were effective against early blight pathogens [50]. Some of the applications have already been developed and evaluated. For example, Silver Oxide nanoparticle was developed and used to manage multiple bacterial and fungal diseases in tomatoes, including early blight. It was not only effective in suppressing development but also reducing the fungal spore count and increasing the chlorophyll content in plants [51]. In another study, mesoporous silica nanoparticles were significantly better than metalaxyl (a control or recommended fungicide) in controlling early blight in tomatoes [44] and improving the overall growth-related traits, including plant height, fresh weight, and dry weight of tomatoes.
3.4. Breeding and use of resistant cultivars
Breeding for resistance to the early blight was initiated in the early 1940s. Resistance identified from Solanum habrachaites, and S. pimpinellifolium is still useful as a source of resistance at present. Several public and private tomato-breeding programs have released multiple EB-resistant breeding lines and hybrids using those sources of resistance. Moderate foliar resistance derived from Campbell 1943 (C.1943) has been advanced into greatly improved horticultural backgrounds, and resistant breeding lines were released NC EBR-2, NC EBR-3, and NC EBR-4 in the 1980s [52,53]. These lines are extensively used to incorporate EB resistance throughout the world. The C.1943 resistance source also confers a high level of resistance to the collar rot (stem lesion) phase of early blight, which is an occasional problem in western NC. In a field study in 1986, it was shown that some of the lines identified as resistant to stem lesions in the greenhouse were also resistant to the foliar blight phase of early blight [54]. Solanum hirsutum PI 126,445 was used as a source of early blight resistance in developing the breeding line NC EBR-1 [52]. Combining resistance from C.1943 and PI 126,445 sources resulted in the development and release of the breeding lines NC EBR-3 and NC EBR-4 and their F1 hybrid combination as 'Mountain Supreme' [53]. The early blight-resistant hybrid 'Plum Dandy' and its parents, NC EBR-5 and NC EBR-6, were released in 1996 [55]. The plum tomato hybrid 'Plum Crimson' and its parental lines, NC EBR-7 and NC EBR-8 were released in 2002. 'Plum Regal' and breeding line NC25P are resistant to late blight (Ph-3 gene) and moderately resistant to early blight [56]. Another hybrid, ' Mountain Magic', and breeding line NC 2CELBR are resistant to both early and late blight [57]. Using the same source, we released more late blight-resistant hybrids, including 'Mountain R', 'Mountain Bebe', and 'Mountain Crown' [58,59,60].
Genetic resistance to LB in tomatoes has been of interest for many years. Three significant resistance genes have been identified in the red-fruited tomato wild species S. pimpinellifolium, including Ph-1, Ph-2, and Ph-3, mapped to tomato chromosomes 7, 10, and 9, respectively. Ph-1 is a single dominant gene providing resistance to race T-0, but new races of the pathogen rapidly overcame it. Ph-1 was mapped to the distal end of chromosome 7 using morphological markers [61]. However, no molecular marker associated with this resistance gene has been reported. Currently, P. infestans race T-1 predominates, rendering the resistance conferred by the Ph-1 gene ineffective. The resistance conditioned by Ph-2, a single incomplete-dominant gene mapped to the lower end of the long arm of tomato chromosome 10 [62], provides partial resistance to several isolates of race T-1 [61,63]. Ph-2 slows but does not stop the disease progress [62]. Furthermore, Ph-2 often fails in the presence of more aggressive isolates [10,64]. Ph-2 has been mapped to an 8.4 cm interval on the long arm of chromosome 10 between RFLP markers CP105 and TG233 [62]. A much stronger resistance gene, Ph-3, was discovered in S. pimpinellifolium accessions L3707 and L3708 (a.k.a. LA 1269 or PI365957) at the Asian Vegetable Research and Development Center (AVRDC) in Taiwan [10]. Currently, this gene is much more useful than Ph-1 and Ph-2 and confers incomplete dominant resistance to a wide range of P. infestans isolates of tomato, including those that overcome Ph-1 and Ph-2 [65]. Ph-3 has been mapped to the long arm of chromosome 9 near RFLP marker TG591a [65]. However, a combination of Ph-2 and Ph-3 confers strong resistance to such isolates. Recently, several tomato-breeding programs have been held around the world, including the North Carolina State University, Pennsylvania State University, Cornell University, and AVRDC. The World Vegetable Center have succeeded in transferring LB resistance genes to fresh-market and/or processing tomato breeding lines or hybrid cultivars using a combination of phenotypic screening and MAS. For example, most recently, several fresh-market tomato breeding lines (e.g., NC1 CELBR (Ph-2 + Ph-3) and NC2 CELBR (Ph-2 + Ph-3) and hybrid cultivars Plum Regal (Ph-3), Mountain Magic (Ph-2 + Ph-3), Mountain Merit (Ph-2 + Ph-3) and Mountain Rouge, have been released by the North Carolina State University Tomato Breeding Program, USA [56,57,66,67]. Also, more breeding lines and cultivars are in the pipeline from these and other tomato breeding programs. However, the availability of more useful PCR-based markers for Ph-2 and Ph-3 will make the selection and breeding for LB resistance in tomato more expedient. The objective of the present study was to map the genes and QTL associated with late blight resistance in a tomato population derived from intra-specific crosses.
Multiple foliar fungal and oomucetes disease resistant hybrids were developed from Cornell University (CU) in collaboration with various seed companies. These hybrids not only combine disease resistance but also fruit quality and are being marketed by seed companies in various parts of the countries. For instance Defiant, Iron lady, and Plum Perfect were developed in collaboration with NCSU to combine the LB resistance whereas Stellar has similar disease resistance and earlier matirity. Brandywise and Summer Sweetheart combine the disease resistance and fruit quality, particularly flavor [68,69]. Using this as a background information, they used three EB-resistant lines CU151011-146, CU151011-170, and CU151095-146 as parents from the CU and two lines (OH08-7663 and OH7536) from the Ohio State University (OSU) to develop mapping populations for EB-resistance [68]. They identified three QTL from chromosomes 1, 5 and 9. The QTL-EB9 was contributed by CU151095-146 whereas the QTL-EB5 was contributed by OH08-7663. Resistance in CU151095-146 was derived from C.1943 whereas that in OH08-7663 was derived from the HI7998 [68,70].
Research on early blight and late blight resistance has been advanced at Penn State University Tomato Breeding Program. It has released multiple breeding lines and hybrids including the award-winning grape hybrid ‘Valentine’. The genetic analysis and reporting at molecular breeding has been useful not only for PSU but also for the entire tomato breeding community. For instance, the heritability estimates reported was 65 to 71% whereas the correlation between earliness and EB resistance was r=-0.46 when they used the population derived from NC84173 (S) x NC39E (R) [71]. In a backcross population, developed between NC84173 (S) x PI126445 (R) to estimate the heritability for early blight resistance, the heritability was close to 70%. They also reported a weak negative correlation (r=-0.26) between maturity and EB resistance [72]. Using this population, they identified ten QTL from various chromosomes explaining a total of 56.4% of phenotypic variance [73]. However, they have reported seven QTL from the chromosomes 3, 4, 5, 6, 8, 10 and 11 in a population derived from the same parents (NC84173 (S) x PI126445 (R) but a different generation [74]. A summary of these information is reported by [75]. A more recent molecular mapping research is summarized in Table 2.
Septoria leaf spot resistance breeding was also initiated in the 1940s [76,77,78]. Some of the progress made at that time is still useful to advance the SLS breeding forward. However, little progress has been made toward developing SLS-resistant breeding lines and hybrids. This disease was not in priority in most of the tomato breeding programs. With climate change, the average temperature is rising and getting warmer every year. Average temperature and humidity are higher, creating a more conducive environment for the SLS. Considering the economic importance of the problem in tomato production, we have initiated the SLS breeding program. While several advanced breeding lines have been developed or are in the pipeline, we have not released them yet.
3.5. Integrated Disease Management (IDM) for the management of foliar fungal diseases of tomato
Integrated disease management in any cropping system uses different methods to prevent and manage various diseases. It relies on closely monitoring the crop health status and deciding on applying action to manage the crop health. IDM is known as a method of disease management in which various disease management methods including biological, chemical, cultural, and physical are integrated for managing the diseases in crops. It is a collection of good agricultural practices which helps in the profitable production of crops with the use of the maximum possible sustainable approach to crop health management.
Integrated disease management in tomato production system starts early from a selection of appropriate varieties resistant or tolerant against various diseases such as early blight, late blight, and Septoria leaf spots. Cropping patterns such as a proper irrigation system that minimizes the leaf wetness period, appropriate fertilization program for optimum growth of the crop, optimum planting density, timely crop canopy management activities, regular and effective disease monitoring activities during the growing season, and finally, proper harvesting and post-harvest handling activities all fall under the package of Integrated disease management system [79]. This helps achieve maximum yield with better quality and having the least footprints in the environment.
Integration of various disease management strategies including using African marigolds as a trap crop, root dip treatment of seedlings by Imidacloprid, application of neem and Pongamia cake in soil, a spray of biopesticides like Pongamia soap were found effective in the management of both foliar diseases in tomatoes production systems [80]. Similarly, plant growth-promoting bacteria are also used as efficient and effective alternatives to various chemicals for managing diseases in tomatoes. Different microbes are also used as soil and plant inoculants in various crops like tomatoes which have shown good results in disease inhibition and growth promotion in crops [81]. Utilization of host plant resistance is one of the economically viable, technically feasible, environmentally safe, and socially acceptable management strategies for tomato foliar fungal disease IDM programs [82] (Bombarely et al., 2011).
3. Crop improvement efforts through molecular and conventional methods
4.1. Early blight
It is reported that multiple genes are involved in tomatoes conferring resistance to early blight (EB) [75,83,84,85]. Top emphasis was placed on improving early blight resistance in tomatoes since the beginning of the systematic tomato breeding program. There are reports on screening for early blight resistance as early as the 1940s. Those reports and genetic analyses are still valid. For instance, the source of resistance identified in Solanum pimpinellium C1943 from Rutger University is the primary source of early blight resistance, which is still valid in the tomato breeding program. Genomic regions and QTL conferring resistance to the EB have been identified. Those QTLs have been reported to explain as high as 25% of the phenotypic variance. Some of the QTL analysis research in connection with breeding effort is presented in section 3.4. A comprehensive review of the availability of QTL has been reported by Beattie et al. (2007) and Adhikari et al. (2017). Additional novel QTLs associated with EB resistance resulting from new research is presented in Table 2.
Contrary to the reporting of QTL conferring resistance to the EB, some of the studies have reported the presence of major gene(s) to conrol the EB resistance [87]. However, there are no follow-up related to these studies. Because of lack of the identificatoin of the major gene, it has impeded the overall breeding progress for the EB resistance in tomatoes. Limited progress has been made in EB resistance by releasing fresh-market resistant lines from the NC State University tomato breeding program. Those lines include NC EBR1 through NCEBR8, NC 1CELBR, and NC 2CELBR [52,53,55,88]. Hybrids were also developed using these lines, growing widely in the US and worldwide. Those hybrids include Mountain Supreme, Mountain Magic, and Mountain Merit [53,57,66], among others. These breeding lines and hybrids are used in the US and worldwide to improve early blight resistance. The World Vegetable Center has expanded its effort to improve the early blight resistance in toamto vareities suitable for Asian and African countries [89].

Table 2.
Quantitative trait loci (QTL) associated with early blight (EB) and late blight (LB) resistance in tomatoes reported in various studies in the past ten years.
Table 2.
Quantitative trait loci (QTL) associated with early blight (EB) and late blight (LB) resistance in tomatoes reported in various studies in the past ten years.
| Trait | Population | QTL | Chr | Position | LOD Score | Additive effect | R2-value (%) | Reference |
|---|---|---|---|---|---|---|---|---|
| Early blight | NC 1CELBR x Fla. 7775 | qEBR-2 | 2 | 16.6-20.0 | 4.2 | -1.42 | 3.8 | [90] |
| qEBR-8 | 8 | 32.4-51.3 | 4.2 | -1.44 | 12.1 | |||
| qEBR-11 | 11 | 44.1-50.9 | 4.0 | -1.44 | 11.7 | |||
| Early blight | - | EB-1.2 | 1 | 85.0 | 5.7 | 87.9 | 4.9 | [70] |
| - | EB-5 | 5 | 64.4 | 10.4 | -126.4 | 11.0 | ||
| - | EB-9 | 9 | 66.9 | 24.0 | 12.0 | 26.4 | ||
| Early blight | NC EBR1 x LA2093 | cLEC73K6b-CT205 | 2 | 1.1-12.2 | 3 | -184.9 | 8 | [91] |
| cTOF19J9-TG-463 | 2 | 46.9-64.9 | 3.4 | -197.4 | 8 | |||
| EB5.1 cLEY-18H8-Ctoc20j21 | 5 | 69.4-81.5 | 5.6 | 283.2 | 18 | |||
| EB6.1 TG274-TG590 | 6 | 14.8-17.3 | 4.6 | -182.2 | 16 | |||
| TG274-cLEN10H12 | 6 | 14.8-29.0 | 3.7 | -224.2 | 10 | |||
| EB9.1 TG348-cTOE10J18 | 9 | 52.8-54.6 | 5.1 | 205.6 | 14 | |||
| TG343-cLED4N20 | 9 | 60.8-69.8 | 3 | 179.7 | 7 | |||
| Late blight | Fla. 8059 X PI 270441 | 02g30527779 | 2 | 11.7 | 0.97 | -0.12 | 3 | [92] |
| 02g30827526 | 2 | 13.7 | 1.95 | -0.17 | 6 | |||
| 2 | 14.7 | 1.87 | -0.17 | 6 | ||||
| 09g66536514 | 9 | 114.9 | 9.37 | -0.76 | 39 | |||
| 09g66864250 | 9 | 116.4 | 10.14 | -0.78 | 42 | |||
| 09g67494653 | 9 | 119.3 | 9.54 | -0.76 | 40 | |||
| Late blight | NC 1CELBR x Fla 7775 | solcap_snp_sl_65677 | 6 | 0 | 2.52 | 0.03 | 2 | [93] |
| solcap_snp_sl_65677 | 6 | 0.01 | 2.52 | 0.03 | 2 | |||
| solcap_snp_sl_11588 | 8 | 0.27 | 2.01 | -0.15 | 8 | |||
| solcap_snp_sl_22830 | 9 | 0.32 | 9.18 | -0.33 | 81 | |||
| CL016855-0847 | 9 | 0.67 | 41.99 | -1.69 | 66 | |||
| solcap_snp_sl_69978 | 9 | 0.67 | 42.44 | -1.72 | 67 | |||
| solcap_snp_sl_8807 | 10 | 0.64 | 4 | -0.44 | 2 | |||
| solcap_snp_sl_1490 | 12 | 0.01 | 3.1 | -0.37 | 2 |
Recently, Akhtar et al. (2019) evaluated an extensive list of tomatoes consisting of 401 genotypes with diverse genetic backgrounds in Pakistan for early bight resistance using a scoring scale of 0 to 5, where 0 = no disease at all and 5=100% disease on the plant. The list of genotypes consisted of genotypes from NC-released breeding lines, which were found susceptible, indicating that the pathogen isolate in Pakistan may be different from NC or the resistance introduced in those lines may already be broken down or may not be effective in Pakistan. However, they found only one resistant line- '21396', and 56 with mild resistance.
Singh et al. (2017a) investigated the inheritance pattern of EB resistance in tomatoes. They used multiple sources of resistance from wild relatives, including S. habrachaites. They found a ratio for a single dominant gene in the F2 generation in only one cross (EC520061). In contrast, in the rest of the crosses, it was 1:2:1. Regardless of the ratio, the level of resistance was reasonably good. Monogenic inheritance was also reported derived from PI 134417. However, most of the inheritance pattern was quantitative.
A recent review on molecular breeding for EB resistance in tomatoes is provided by Chaerani (2020) in a conference paper. The QTL mapping and identification of genes have not been reported in the case of EB. This is the first step towards developing molecular markers associated with any trait. Although EB QTL mapping has been reported, there is a lack of information on the major QTL or a gene conferring resistance to the EB, which is the basis of the development of molecular markers. Because of this lack of information, marker-assisted selection (MAS) has not been employed in EB resistance.
Oliveira et al. (2016) have reviewed the mechanism of plant resistance induction to various diseases. They reported using salicylic acid (SA), Jasmonic acid (JA), reactive oxygen species (ROS), callose deposition, and synthesis of defense enzymes, among other compounds. Tripathi et al. (2019) also conducted a detailed review of the involvement of SA in plant systemic acquired resistance (SAR), and host resistance mechanism at the molecular level. It has been shown that SAR was induced in tomatoes by the application of Paenibacillus lentimorbus B-30488 to suppress early blight and was found to reduce the disease development by 45.3% [99].
4.2. Late blight
There are three major genes Ph-1, Ph-2, and Ph-3, conferring resistance to the late blight of tomatoes. Ph-1 is a dominant gene, whereas Ph-2 and Ph-3 are co-dominant genes. These genes were identified in Solanum pimpinellifolium, which is cross-compatible with S. lycopersicum. Late blight resistance conferred by these genes is race-specific. While the gene Ph-1 is no more effective, a combination of Ph-2 and Ph-3 is still effective in conferring resistance to resistance to the existing race (Figure 4). By combining these two genes, several breeding lines and hybrids of tomatoes have been released from NC State University tomato breeding programs worldwide, including The World Vegetable Center in Taiwan. Those breeding lines and hybrids are NC 1CELBR, NE 2CLBR, NC 161L, NC 25P, NC 8Grape, Mountain Merit, Mountain Rouge, Plum regal, Mountain Crown, and Mountain Bebe [52,53,56,57,59,66,67,88]. Several wild accessions are resistant to the EB, including LA2157, GI 1556, LA3111, PI126445, PI390513, and PI134417, among others, as reported by Belkhadir et al. (2004) in her review paper. The Ph-3 gene was fine-mapped on the long-arm of the chromosome 9 [101]. This was very useful for the development of molecualr markers and eventually cloning of this gene, which encodes a CC-NBS-LRR protein [102].
Molecular markers associated with Ph-2 and Ph-3 are reported in tomatoes. These markers are CAPS type requiring restriction enzyme digestion before resolving the fragment sizes. It makes the genotyping process not only time-consuming but also costly. While the markers are universal, we are trying to develop SCAR-type markers associated with Ph-2 and Ph-3, which do not involve restriction digestion. Since the Ph-1 gene does not have any use at present, there is little use in developing new molecular markers associated with it [103,104].
While Ph-2 and Ph-3 are single genes conferring resistance, QTL conferring resistance to late blight has also been reported from wild relatives [105,106]. Three QTL have been reported from chromosomes 4, 5, and 11 in the near-isogenic lines derived from a backcross population developed from Lycopersicum esculentum (currently Solanum lycopersicum) x L. hirsitum (currently, S. habrachaites). QTL on chromosome 4 mapped in an interval of 6.9 cM between TG192 and CT19,4 whereas that on chromosome 5 mapped near TG23, and that on chromosome 11 mapped to an interval of 15.1 cM between TG194 and TG400. The source of resistance traces back to LA2099 [105]. These QTLs conferring resistance to LB were mapped and fine-mapped from chromosome 5 [107] and also from chromosome 11 separately later [108]. Near-isogenic lines were used to create sub-NIL recombinant inbred populations one for each target chromosome region for fine-mapping. From the single QTL of each chromosome, they found two and six QTL, respectively. Most of the growth chambers QTL were found to be co-located with field QTL [109]. Five QTLs derived from LA1777 (S. habrachaites) were identified and associated with late blight resistance in terms of lesion size and were located on chromosomes 4 (two QTL), 7, 8, and 12 [110]. QTL associated with late blight resistance has also been reported from the same chromosomes derived from LA2099 (S. habrachaites) before [106], which has shown a high level of resistance to LB [105,108,111]. Quantitative resistance to LB has also been reported from LA716 (S. penelli) [112]. To make the resistance durable, [110] have suggested the pyramiding of resistance genes and/or QTL from multiple species.
In another study, it was reported that there were 15 QTL detected in a backcross population derived from Lycopersicum esculentum (currently, S. lycopersicum) x L. hirsutum (currently, S. habrachaites) located on chromosomes 1, 2, 3, 4, 5 and 11 [106]. Yet another QTL was found on chromosome 6 derived from S. penelli explaining about 25% of the total phenotypic variations [112]. Three minor QTLs were found in chromosomes 6, 8, and 12 [93]. These QTLs are also reported in Table 2. The World Vegetable Center has advanced the late blight resistance breeding at molecular level and developed multiple lines of tomato that are suitable for Asian production system [113,114]. These lines can also be used in other parts of the world to introgress the resistant gene.
Four QTL located on chromosomes 1, 10 and 11 were reported in a population derived from Fla. 8059 x PI 270,441 [115], and, additional QTL associated with LB resistance were located on chromosomes 2, 3, 10, and 11 in a population derived from Fla. 8059 x PI 163,245 [116]. These QTLs were identified based on selective genotyping approach.
4.3. Septoria leaf spot
Genetic control of Septoria leaf spot resistance is not well understood. It has a narrow pathogen diversity. Based on our research at NC State University, there may be a quantitative mode of resistance for SLS at an early stage. However, a study conducted at Cornell University revealed that two genes may be involved in conferring resistance to the SLS, which is not published yet. Since the genetic control is unclear, the SLS-resistant breeding lines and hybrids have not yet been developed. One of the most vital resources for disease resistance germplasm is found in wild tomato species. Of the 13 wild species of tomato (Solanum L. section Lycopersicon [Mill.]) [117], SLS resistance has been discovered in varying degrees in S. pimpinellifolium, S. chilense, S. habrochaites, S. peruvianum and S. pennelli [104,118], with the best source of resistance found in S. habrochaites and S. peruvianum [118]. Unfortunately, crossing between cultivated tomato and wild species is very difficult. When it can be achieved, the resulting SLS resistance in progeny are often linked with traits unfit for commercial production [75]. A study exploring the SLS resistance of various Solanum accessions found that the most susceptible accessions tested were cultivated tomatoes (S. lycopersicum and S. lycopersicum var. cerasiforme). Only one cultivated tomato accession was found to be moderately resistant: S. lycopersicum var. cerasiforme (CNPH-0633), and it crosses readily with other cultivated varieties [119]. Blum et al. (2002) offer a detailed list of partially SLS-resistant accessions to be explored in potential breeding work.
SLS resistance in tomatoes is thought to be primarily qualitative, with several QTLs supplementing the resistance of a single gene. In 1945, SLS resistance was reported to be significantly affected by a single dominant resistance gene called Se. A chromosome position for Se has not yet been determined [76]. Joshi et al. (2015) reported that Se's dominance was incomplete, although it was suggested that this could have resulted from using a parent that was not completely susceptible to SLS. Cornell University has been trying to determine if homozygous plants for the Se gene are more resistant than heterozygous plants [120].
In a study conducted in Brazil, they evaluated a collection of 124 accessions for the SLS under greenhouse conditions. They found that ten accessions were highly resistant (HR), whereas 33 were classified as resistant (R) and the rest of the accessions were susceptible [121]. The greenhouse observations were verified by field experiments by using only the HR and R sub-set of accessions. Five new sources with high levels of resistance were found in S. peruvianum accessions including PI-306811, CNPH-1036, LA-1910, LA-1984 and LA-2744 [121].
No major gene or QTL associated with the SLS resistance in tomatoes is reported. Because of the lack of this information, molecular breeding has yet to be realized for SLS. We are going to map the QTL associated with the SLS in our program now. We are optimistic that we will be able to identify major QTL and eventually develop molecular markers associated with this important disease.
Only a few tomato cultivars have been released, claiming SLS resistance. Some cultivars with good SLS resistance are 'Iron Lady,' 'Stellar,' 'Summer Sweetheart,' and 'Plum Perfect. The cultivars mentioned above have some degree of tolerance/resistance to several other diseases, such as late blight and early blight [68,69,122].
4. Potential to improve using modern tools
5.1. Genomic resources
Tomato is one of the well-studied crop plants for its genomics. Its whole genome sequence was published in 2012 [123]. The total estimated number of genes in tomatoes are around 34,075. These genes have been annotated for their gene ontology and gene function. This information is summarized and presented in various resources, including online databases such as www.solgenomics.net, https://solgenomics.net/organism/Solanum_lycopersicum/genome, http://ted.bti.cornell.edu/, http://www.kazusa.or.jp/tomato/, http://www.g2p-sol.eu/, http://www.kazusa.or.jp/jsol/microtom/, https://tomatoma.nbrp.jp/, and review papers [124]. Gene expression data with various treatment combinations have been generated, including biotic and abiotic stress treatments [125,126,127,128]. QTL mapping populations, QTL and genes associated with various traits, and eventually molecular markers are also available. Various portals report this information, including genes, gene expression, micro RNAs, different marker systems, and quantitative trait loci (QTL) analysis. Molecular markers, including Amplified fragment length polymorphism (AFLP), Cleaved amplified polymorphic sequence (CAPS), Restriction fragment length polymorphism (RFLP), sequence characterized amplified region (SCAR), single nucleotide polymorphism (SNP), and simple sequence region (SSR). Among different reporting portal systems, the most informative portal is Sol Genomics Network (www.solgenomics.net). It has so much information on various aspects, including biotic and abiotic stress conditions. The Tomato Genetics Resource Database (TGRD), consisting of similar information, was developed before. However, it is no longer active now [129]. However, major genes and molecular markers associated with EB and SLS are not available for marker-assisted selection. With the availability of next-generation sequencing facilities for lower costs, most programs are inclined to use SNPs now [130,131,132]. Now gene expression analysis data has been generated for various pathogens in tomatoes by inoculating the resistant and susceptible tomato genotypes and collecting tissue samples at different time points [133,134,135]. Performing RNA-Seq analysis and gene ontology analysis, we can get a list of up-regulated and down-regulated genes and their association with various proteins. RNA-seq analysis provides information on differential gene expression and the extraction of SNP molecular markers, which can eventually be used for QTL mapping, as shown by Shi and Panthee, (2020). Casa et al. (2008) revealed that early infection occurs in potatoes among three species, including potato, tobacco, and tomato when inoculated with Phytophthora infestans. They also found that gene expression pattern was in the same trend, i.e., defense-related genes, such as reactive oxygen species, were early expressed. There were some common genes among the three species. This type of research concerning EB, LB, and SLS in tomatoes are yet to be investigated in detail.
An updated tomato reference genome was published recently Wang et al. (2020). This study has compared the S. lycopersicum Heinz1706 and S. pimpinellifolium LA2093. Even more, comprehensive reference genomes are being reported by Pan-genome analysis so that it covers all available genes. More SNP molecular markers are being developed by using cultivated and wild relatives, which will be helpful in mapping and gene identification, and cloning [138,139].
5.2. Genetic transformation
Genetic transformation has been an important tool for crop improvement, including crop yield, quality, and disease resistance [140]. Tomato was one of the first crop plants to transform and improve fruit quality. Since then, several traits have been improved. In this series, rolB gene from Agrobacterium rhizogenes was introduced into Agrobacterium tumefaciens, and eventually, tomato Rio Grande was transformed. When phenotypic traits, including early blight resistance, were monitored, the disease incidence ranged from 4.17 to 25% whereas disease severity ranged from 1.75 to 16.75%, significantly less than control values [141]. This indicated that early blight could be managed using genetic transformation. Khan et al. (2011) transformed tomatoes by fusing miR4026 and nucleotide-binding leucine-rich repeat revealed that miR4026 negatively regulates the resistance to early blight caused by Alternaria solani in tomatoes. This is important information for developing early blight resistance by genetic transformation or genome editing by targeting miR4026. Tomato transformation protocols for various purposes are available [143,144].
5.3. Genome editing
Genome editing is the technique by which DNA mutation in insertion and deletion (InDels) or base substitutions are introduced to create an organism with a new or modified product [124,145,146]. There are different types of genome-editing methods, as described below.
Zinc-finger nuclease-based genome-editing approach utilizes the synthetic restriction enzymes' Zinc-finger nucleases, which have DNA-binding domains that specifically bind three base pairs at the target sites. Three base-pair specificities can be designed to target a specific amino acid and modify the amino acid of interest, ultimately leading to changes in the protein of interest [147]. It has broader applications in crop improvement.
TALEN-based genome-editing includes the role of tandem repeats for the specificity of the proteins domain, based on which the development of chimeric genome-editing tool known as transcription factor-like effector nucleases (TALEN) was developed. The mechanism of TALEN-based genome editing is basically via the disruption of the effector-binding element of the S-gene promoter, which eventually impairs the comparable molecular interactions between the effector and the target S-gene [147].
Oligonucleotide-directed mutagenesis (ODM) is another tool of genome editing in which 20 to 100 base pairs long sequences are identical to the target sequence except in a single nucleotide where the intended point mutation is required. The oligonucleotide-directed host, DNA repair system introduces the mutation that disrupts the target gene's function. This process introduces the desired single nucleotide mutation into the target genome resulting in the expression of a novel trait or function following subsequent regenerations either by tissue culture or classical plant breeding [148,149].
CRISPR/Cas-based genome editing has been the most popular and adopted to manipulate the genome of many crop plants to achieve various breeding objectives recently. The components for genome editing using the CRISPR/Cas-based method are DNA endo-nuclease Cas9 protein and a customizable single-stranded guide-RNA. Once the target DNA sequence is hybridized with the complementary sgRNA, high-fidelity Cas9 triggers dsDNA breaks [147,150].
The concept of genome editing of the susceptible gene (S gene) has emerged and is practiced in some of the diseases in tomatoes. A summary of all those disease resistance achieved by genome editing is presented by Barka and Lee (2022). They discuss early blight and the Septoria leaf spot of tomato. In a separate study, tomato plants were transformed with Agrobacterium rhizogenese with the rolB gene. They assessed the phenotype for early blight and insect resistance, although the mode of action of rolB is mainly unknown.
5. Future prospects
Foliar fungal diseases, early blight, and septoria leaf spots, late blight (caused by oomycetes) are still the primary threat to tomato production worldwide. There are novel resources available to manage these diseases, including nanotechnology, breeding, genetics, genomics, genome editing, and biotechnology. While some genetic resources have already been utilized to manage some of the above diseases, such as early blight and late blight, other resources are yet to be used to manage these diseases better. For MAS to employ effectively, it should be mapped precisely in the genome of the tomato. Early blight and Septoria leaf spot resistance should be mapped precisely and reliable molecular markers should be developed to employ the MAS for these diseases. Genomic resources developed from gene expression analysis are beneficial for gene identification. Gene expression data on EB and SLS is still lacking. Future experiments in this direction will help generate the expression data and fill the gene identification gap. Depending upon their mode of action, these genes can eventually be used in genetic transformation or genome editing. Progress is being made in various programs in this direction. Genome editing can be exploited either on host resistance or to understand the pathogen's virulence. These resources are likely to be used for the integrated management of foliar fungal diseases as required.
Acknowledgement
We want to thank Dr. Tika Adhikari for reviewing this manuscript. The second and third authors (AP and RP) are supported by USDA NIFA Organic Transition Program.
References
- FAOSTAT Crop and Livestock Products Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 2 June 2023).
- Abramovitch, R.B.; Kim, Y.J.; Chen, S.; Dickman, M.B.; Martin, G.B. Pseudomonas Type III Effector AvrPtoB Induces Plant Disease Susceptibility by Inhibition of Host Programmed Cell Death. EMBO J 2003, 22, 60–69. [CrossRef]
- Sobczak, M.; Avrova, A.; Jupowicz, J.; Phillips, M.S.; Ernst, K.; Kumar, A. Characterization of Susceptibility and Resistance Responses to Potato Cyst Nematode (Globodera Spp.) Infection of Tomato Lines in the Absence and Presence of the Broad-Spectrum Nematode Resistance Hero Gene. Mol Plant Microbe Interact 2005, 18, 158–168. [CrossRef]
- Zhang, Y.; Lubberstedt, T.; Xu, M. The Genetic and Molecular Basis of Plant Resistance to Pathogens. Journal of Genetics and Genomics 2013, 40, 23–35. [CrossRef]
- Hoagland, L.; Navazio, J.; Zystro, J.; Kaplan, I.; Vargas, J.G.; Gibson, K. Key Traits and Promising Germplasm for an Organic Participatory Tomato Breeding Program in the U.S. Midwest. HortScience 2015, 50, 1301–1308. [CrossRef]
- Adhikari, T.B.; Ingram, T.; Halterman, D.; Louws, F.J. Gene Genealogies Reveal High Nucleotide Diversity and Admixture Haplotypes within Three Alternaria Species Associated with Tomato and Potato. Phytopathology 2020, 110, 1449–1464. [CrossRef]
- Chaerani, R.; Voorrips, R.E. Tomato Early Blight (Alternaria Solani): The Pathogen, Genetics, and Breeding for Resistance. Journal of General Plant Pathology 2006, 72, 335–347. [CrossRef]
- Beattie, A.D.; Scoles, G.J.; Rossnagel, B.G. Identification of Molecular Markers Linked to a Pyrenophora Teres Avirulence Gene. Phytopathology 2007, 97, 842–849. [CrossRef]
- Sherf, A.F.; MacNab, A.A. Vegetable Diseases and Their Control; J. Wiley, 1986; ISBN 978-0-471-05860-1.
- Black, L.L.; Wang, T.C.; Hanson, P.M.; Chen, J.T. Late Blight Resistance in Four Wild Tomato Accessions: Effectiveness in Diverse Locations and Inheritance of Resistance Available online: https://scholar.google.com/scholar?hl=en&as_sdt=0%2C34&q=Late+blight+resistance+in+four+wild+tomato+accessions%3A+effectiveness+in+diverse+locations+and+inheritance+of+resistance.+&btnG= (accessed on 4 June 2023).
- Kemmitt, G. Early Blight of Potato and Tomato. The Plant Health Instructor 2002. [CrossRef]
- Abbasi, P.A.; Cuppels, D.A.; Lazarovits, G. Effect of Foliar Applications of Neem Oil and Fish Emulsion on Bacterial Spot and Yield of Tomatoes and Peppers. Canadian Journal of Plant Pathology 2003, 25, 41–48. [CrossRef]
- Bock, C.H.; Gottwald, T.R.; Parker, P.E.; Ferrandino, F.; Welham, S.; Van Den Bosch, F.; Parnell, S. Some Consequences of Using the Horsfall-Barratt Scale for Hypothesis Testing. Phytopathology 2010, 100, 1030–1041. [CrossRef]
- Biswas, M.K.; Chai, L.; Amar, M.H.; Zhang, X.; Deng, X. xin Comparative Analysis of Genetic Diversity in Citrus Germplasm Collection Using AFLP, SSAP, SAMPL and SSR Markers. Sci Hortic 2011, 129, 798–803. [CrossRef]
- Blanco, A.; Gadaleta, A. and; Simeone, R. Variation for Yield and Quality Components in Durum Wheat Backcross Inbred Lines Derived from Ssp. Dicoccoides. BODENKULTUR (diebodenkultur.boku.ac.at) 2004, 54, 163–170.
- Goodwin, S.B.; Cohen, B.A.; Fry, W.E. Panglobal Distribution of a Single Clonal Lineage of the Irish Potato Famine Fungus. Proceedings of the National Academy of Sciences 1994, 91, 11591–11595. [CrossRef]
- Hohl, H.R.; Iselin, K. Strains of Phytophthora Infestans from Switzerland with A2 Mating Type Behaviour. Transactions of the British Mycological Society 1984, 83, 529–530.
- Fry, W.; Goodwin, S.; Dyer, A.; Matuszak, J.; Drenth, A.; Tooley, P.; Sujkowski, L.; Koh, Y.; Cohen, B.; Spielman, L.; et al. Historical and Recent Migrations of Phytophthora-Infestans - Chronology, Pathways, and Implications. Plant Dis 1993, 77, 653–661. [CrossRef]
- Fry, W.E.; McGrath, M.T.; Seaman, A.; Zitter, T.A.; McLeod, A.; Danies, G.; Small, I.M.; Myers, K.; Everts, K.; Gevens, A.J.; et al. The 2009 Late Blight Pandemic in the Eastern United States – Causes and Results. Plant Dis 2013, 97, 296–306. [CrossRef]
- Fry, W.E. Population Genomics of Phytophthora Infestans. - Google Scholar. Phytopathology 2015, 105.
- Fry, W.E.; Birch, P.R.J.; Judelson, H.S.; Grünwald, N.J.; Danies, G.; Everts, K.L.; Gevens, A.J.; Gugino, B.K.; Johnson, D.A.; Johnson, S.B.; et al. Five Reasons to Consider Phytophthora Infestans a Reemerging Pathogen. Phytopathology 2015, 105, 966–981. [CrossRef]
- Tian, Y.E.; Yin, J.L.; Sun, J.P.; Ma, Y.F.; Wang, Q.H.; Quan, J.L.; Shan, W.X. Population Genetic Analysis of Phytophthora Infestans in Northwestern China. Plant Pathol 2016, 65, 17–25. [CrossRef]
- Fry, W.E. Phytophthora Infestans: The Itinerant Invader; “Late Blight”: The Persistent Disease. Phytoparasitica 2020, 48, 87–94. [CrossRef]
- Schumann, G.L.; D’Arcy, C.J. Late Blight of Potato and Tomato. The Plant Health Instructor 2000. [CrossRef]
- Saville, A.C.; Martin, M.D.; Ristaino, J.B. Historic Late Blight Outbreaks Caused by a Widespread Dominant Lineage of Phytophthora Infestans (Mont.) de Bary. PLoS One 2016, 11, e0168381. [CrossRef]
- Majeed, A.; Muhammad, Z.; Ullah, Z.; Ullah, R.; Ahmad, H. Late Blight of Potato (Phytophthora Infestans) I: Fungicides Application and Associated Challenges. Turkish Journal of Agriculture - Food Science and Technology 2017, 5, 261–266. [CrossRef]
- Botella-Pavía, P.; Rodríguez-Concepción, M. Carotenoid Biotechnology in Plants for Nutritionally Improved Foods. Physiol Plant 2006, 126, 369–381. [CrossRef]
- da Costa, C.A.; Lourenço, V.; Santiago, M.F.; Veloso, J.S.; Reis, A. Molecular Phylogenetic, Morphological, and Pathogenic Analyses Reveal a Single Clonal Population of Septoria Lycopersici with a Narrower Host Range in Brazil. Plant Pathol 2022, 71, 621–633. [CrossRef]
- Broggini, G.A.L.; Galli, P.; Parravicini, G.; Gianfranceschi, L.; Gessler, C.; Patocchi, A. HcrVf Paralogs Are Present on Linkage Groups 1 and 6 of Malus. Genome 2009, 52, 129–138. [CrossRef]
- Blauth ’, S.L.; Ste4ens, J.C.; Churchill ’, G.A.; Mutschler, M.A.; Ste, J.C.; Ens ’, !; Churchill, G.A.; Hall, E. Identification of QTLs Controlling Acylsugar Fatty Acid Composition in an Intraspecific Population of Lycopersicon Pennellii (Corr.) D’Arcy. Theoretical Applied Genetics 1999, 99, 373–381.
- Broman, K.W.; Wu, H.; Sen, Ś.; Churchill, G.A. R/Qtl: QTL Mapping in Experimental Crosses. Bioinformatics 2003, 19, 889–890. [CrossRef]
- Hardy, O.J.; Vekemans, X. Spagedi: A Versatile Computer Program to Analyse Spatial Genetic Structure at the Individual or Population Levels. Mol Ecol Notes 2002, 2, 618–620. [CrossRef]
- Bohs, L.; Olmstead, R.G. Phylogenetic Relationships in Solanum (Solanaceae) Based on NdhF Sequences. Syst Bot 1997, 22, 5–17. [CrossRef]
- Brommonschenkel, S.H.; Frary, A.; Frary, A.; Tanksley, S.D. The Broad-Spectrum Tospovirus Resistance Gene Sw-5 of Tomato Is a Homolog of the Root-Knot Nematode Resistance Gene Mi. Mol Plant Microbe Interact 2000, 13, 1130–1138. [CrossRef]
- Broome, C.R.; Terrel, E.E.; Reveal, J.L. Proposal to Conserve Lycopersicon Esculentum Milleras the Scientific Name of the Tomato.; 1983.
- Brumfield, R.T.; Beerli, P.; Nickerson, D.A.; Edwards, S. V. The Utility of Single Nucleotide Polymorphisms in Inferences of Population History. Trends Ecol Evol 2003, 18, 249–256. [CrossRef]
- Broun, P.; Tanksley, S.D. Characterization and Genetic Mapping of Simple Repeat Sequences in the Tomato Genome. Molecular and General Genetics 1996, 250, 39–49. [CrossRef]
- Pandey, A.; Devkota, A.; Yadegari, Z.; Dumenyo, K.; Taheri, A. Antibacterial Properties of Citric Acid/β-Alanine Carbon Dots against Gram-Negative Bacteria. Nanomaterials 2021, 11. [CrossRef]
- Charles Brummer, E.; Casler, M.D. Improving Selection in Forage, Turf, and Biomass Crops Using Molecular Markers. Molecular Breeding of Forage and Turf 2009, 193–210. [CrossRef]
- Brummer, E.C.; Graef, G.L.; Orf, J.; Wilcox, J.R.; Shoemaker, R.C. Mapping QTL for Seed Protein and Oil Content in Eight Soybean Populations. Crop Sci 1997, 37, 370–378. [CrossRef]
- Yadav, A.; Yadav, K. Nanoparticle-Based Plant Disease Management: Tools for Sustainable Agriculture. Nanotechnology in the Life Sciences 2018, 29–61. [CrossRef]
- Kim, J.S.; Kuk, E.; Yu, K.N.; Kim, J.H.; Park, S.J.; Lee, H.J.; Kim, S.H.; Park, Y.K.; Park, Y.H.; Hwang, C.Y.; et al. Antimicrobial Effects of Silver Nanoparticles. Nanomedicine 2007, 3, 95–101. [CrossRef]
- Bonnema, G.; Hontelez, J.; Verkerk, R.; Zhang, Y.Q.; Van Daelen, R.; Van Kammen, A.; Zabel, P. An Improved Method of Partially Digesting Plant Megabase DNA Suitable for YAC Cloning: Application to the Construction of a 5.5 Genome Equivalent YAC Library of Tomato. The Plant Journal 1996, 9, 125–133. [CrossRef]
- Derbalah, A.; Shenashen, M.; Hamza, A.; Mohamed, A.; El Safty, S. Antifungal Activity of Fabricated Mesoporous Silica Nanoparticles against Early Blight of Tomato. Egyptian Journal of Basic and Applied Sciences 2018, 5, 145–150. [CrossRef]
- Safarpour, H.; Safarnejad, M.R.; Tabatabaei, M.; Mohsenifar, A.; Rad, F.; Basirat, M.; Shahryari, F.; Hasanzadeh, F. Development of a Quantum Dots FRET-Based Biosensor for Efficient Detection of Polymyxa Betae. Canadian Journal of Plant Pathology 2012, 34, 507–515. [CrossRef]
- Brusca, J. Inheritance of Tomato Late Blight Resistance from’Richter’s Wild Tomato’and Evaluation of Late Blight Resistance Gene Combinations in Adapted Fresh Market Tomato. 2003.
- Fukamachi, K.; Konishi, Y.; Nomura, T. Disease Control of Phytophthora Infestans Using Cyazofamid Encapsulated in Poly Lactic-Co-Glycolic Acid (PLGA) Nanoparticles. Colloids Surf A Physicochem Eng Asp 2019, 577, 315–322. [CrossRef]
- Ali, M.; Kim, B.; Belfield, K.D.; Norman, D.; Brennan, M.; Ali, G.S. Inhibition of Phytophthora Parasitica and P. Capsici by Silver Nanoparticles Synthesized Using Aqueous Extract of Artemisia Absinthium. Phytopathology 2015, 105, 1183–1190. [CrossRef]
- Bella, P.; Ialacci, G.; Licciardello, G.; Rosa, R.; Catara, V. CHARACTERIZATION OF ATYPICAL CLAVIBACTER MICHIGANENSIS Subsp. MICHIGANENSIS POPULATIONS IN GREENHOUSE TOMATOES IN ITALY. Journal of Plant Pathology 2012. [CrossRef]
- Kanhed, P.; Birla, S.; Gaikwad, S.; Gade, A.; Seabra, A.B.; Rubilar, O.; Duran, N.; Rai, M. In Vitro Antifungal Efficacy of Copper Nanoparticles against Selected Crop Pathogenic Fungi. Mater Lett 2014, 115, 13–17. [CrossRef]
- Kumari, M.; Pandey, S.; Bhattacharya, A.; Mishra, A.; Nautiyal, C.S. Protective Role of Biosynthesized Silver Nanoparticles against Early Blight Disease in Solanum Lycopersicum. Plant Physiology and Biochemistry 2017, 121, 216–225. [CrossRef]
- Gardner, R.G. NC EBR-1 and NC EBR-2 Early Blight Resistant Tomato Breeding Lines. HortScience 1988, 23, 779–781. [CrossRef]
- Gardner, R.G.; Shoemaker, P.B. “Mountain Supreme” Early Blight-Resistant Hybrid Tomato and Its Parents, NC EBR-3 and NC EBR-4. HortScience 1999.
- Gardner, R.G. Greenhouse Disease Screen Facilitates Breeding Resistance to Tomato Early Blight. HortScience 1990, 25, 222–223. [CrossRef]
- Gardner, R.G. “Plum Dandy”, a Hybrid Tomato, and Its Parents, NC EBR-5 and NC EBR-6. HortScience 2000, 35, 962–963.
- Gardner, R.G.; Panthee, D.R. ‘Plum Regal’ Fresh-Market Plum Tomato Hybrid and Its Parents, NC 25P and NC 30P. HortScience 2010, 45, 824–825. [CrossRef]
- Gardner, R.G.; Panthee, D.R. ‘Mountain Magic’: An Early Blight and Late Blight-Resistant Specialty Type F1 Hybrid Tomato. HortScience 2012, 47, 299–300. [CrossRef]
- Panthee, D.R. ‘Mountain Regina’: Multiple Disease Resistant Fresh-Market Hybrid Tomato and Its Parents, NC 1LF and NC 2LF. HortScience 2021, 56, 736–738. [CrossRef]
- Panthee, D.R.; Gardner, R.G. ‘Mountain Bebe’: Hybrid Grape Tomato and Its Parents NC 7 Grape and NC 8 Grape. HortScience 2022, 57, 444–446. [CrossRef]
- Panthee, D.R. ‘Mountain Crown’: Late Blight and Tomato Mosaic Virus-Resistant Plum Hybrid Tomato and Its Parent, NC 1 Plum. HortScience 2020, 55, 2056–2057. [CrossRef]
- Peirce, L.C. Linkage Tests with Ph Conditioning Resistance to Race 0, Phytophthora Infestans. . Report of the Tomato Genetics Cooperative 1971, 21, 30.
- Moreau, P.; Thoquet, P.; Olivier, J.; Laterrot, H.; Grimsley, N. Genetic Mapping of Ph-2, a Single Locus Controlling Partial Resistance to Phytophthora Infestans in Tomato. Molecular Plant Microbe Interaction 1998, 11, 259–269. [CrossRef]
- Gallegly, M.E. Resistance to the Late Blight Fungus in Tomato. Proceedings of Plant Science Seminar, Camden, New Jersey 1960. 1960.
- Goodwin, S.B.; Schneider, R.E.; Fry, W.E. Use of Cellulose-Acetate Electrophoresis for Rapid Identification of Allozyme Genotypes of Phytophthora Infestans. Plant Dis 1995, 79, 1181–1185. [CrossRef]
- Chunwongse, J.; Chunwongse, C.; Black, L.; Hanson, P. Molecular Mapping of the Ph-3 Gene for Late Blight Resistance in Tomato. J Hortic Sci Biotechnol 2002, 77, 281–286. [CrossRef]
- Panthee, D.R.; Gardner, R.G. ‘Mountain Merit’: A Late Blight-Resistant Large-Fruited Tomato Hybrid. HortScience 2010, 45, 1547–1548. [CrossRef]
- Panthee, D.R.; Gardner, R.G. ‘Mountain Rouge’: A Pink-Fruited, Heirloom-Type Hybrid Tomato and Its Parent Line NC 161L. HortScience 2014, 49, 1463–1464. [CrossRef]
- Anderson, T.; Dejong, D.; Glos, M.; Bojanowski, J.B.; Mutschler, M. Mapping Campbell 1943 Stem Early Blight Resistance and Adding an Additional Source of Foliar Early Blight Resistance to Cornell Fungal Resistant Tomato Line. Ithaca, NY: Cornell University 2019.
- Mutschler, M.A.; McGrath, M. VEGEdge : Cornell Cooperative Extension. 2019.
- Anderson, T.A.; Zitter, S.M.; De Jong, D.M.; Francis, D.M.; Mutschler, M.A. Cryptic Introgressions Contribute to Transgressive Segregation for Early Blight Resistance in Tomato. Theor Appl Genet 2021, 134, 2561–2575. [CrossRef]
- Foolad, M.R.; Subbiah, P.; Ghangas, G.S. Parent-Offspring Correlation Estimate of Heritability for Early Blight Resistance in Tomato, Lycopersicon Esculentum Mill. Euphytica 2002, 126, 291–297. [CrossRef]
- Foolad, M.R.; Lin, G.Y. Heritability of Early Blight Resistance in a Lycopersicon Esculentum×Lycopersicon Hirsutum Cross Estimated by Correlation between Parent and Progeny. Plant Breeding 2001, 120, 173–177. [CrossRef]
- Foolad, M.R.; Zhang, L.P.; Khan, A.A.; Niño-Liu, D.; Lin, G.Y. Identification of QTLs for Early Blight (Alternaria Solani) Resistance in Tomato Using Backcross Populations of a Lycopersicon Esculentum × L. Hirsutum Cross. Theoretical and Applied Genetics 2002, 104, 945–958. [CrossRef]
- Zhang, L.P.; Lin, G.Y.; Niño-Liu, D.; Foolad, M.R. Mapping QTLs Conferring Early Blight (Alternaria Solani) Resistance in a Lycopersicon Esculentum x L. Hirsutum Cross by Selective Genotyping. Molecular Breeding 2003, 12, 3–19. [CrossRef]
- Foolad, M.R.; Merk, H.L.; Ashrafi, H. Genetics, Genomics and Breeding of Late Blight and Early Blight Resistance in Tomato. Critical Reviews in Plant Sciences 2008, 27, 75–107. [CrossRef]
- Andrus, C.F.; Reynard, G.B. Resistance to Septoria Leaf Spot and Its Inheritance in Tomatoes. Phytopathology 1945, 35, 16–24.
- Locke, S.B. Resistance to Early Blight and Septoria Leaf Spot in the Genus Lycopersicon. Phytopathology 1949, 39, 829–836.
- Joshi, B. Molecular Tagging of Resistance Genes to Septoria Leaf Spot and Late Blight in Tomato (Solanum Lycopersicum L.). 2011.
- Paret, M.L.; Dufault, N.; Momol, T.; Marois, J.; Olson, S. Integrated Disease Management for Vegetable Crops in Florida. EDIS 2012, 2012. [CrossRef]
- Bubeck, D.M.; Carlone, M.R.; Fox, R.L.; Hoffbeck, M.D.; Segebart, R.L.; Stucker, D.S. Breeding Progress Measured in Eight Elite Inbred Families. Maydica 2006, 51, 141–149.
- Singh, V.K.; Singh, A.K.; Kumar, A. Disease Management of Tomato through PGPB: Current Trends and Future Perspective. 3 Biotech 2017, 7, 1–10. [CrossRef]
- Bombarely, A.; Menda, N.; Tecle, I.Y.; Buels, R.M.; Strickler, S.; Fischer-York, T.; Pujar, A.; Leto, J.; Gosselin, J.; Mueller, L.A. The Sol Genomics Network (Solgenomics.Net): Growing Tomatoes Using Perl. Nucleic Acids Res 2011, 39, D1149–D1155. [CrossRef]
- Nash, A.F.; Gardner, R.G. Tomato Early Blight Resistance in a Breeding Line Derived from Lycopersicon Hirsutum PI 126445. Plant Dis 1988.
- Nash, A.F.; Gardner, R.G. Heritability of Tomato Early Blight Resistance Derived from Lycopersicon Hirsutum P.I. 126445. Journal of the American Society for Horticultural Science 1988, 113, 264–268. [CrossRef]
- Chaerani, R.; Smulders, M.J.M.; Van Der Linden, C.G.; Vosman, B.; Stam, P.; Voorrips, R.E. QTL Identification for Early Blight Resistance (Alternaria Solani) in a Solanum Lycopersicum × S. Arcanum Cross. Theoretical and Applied Genetics 2007, 114, 439–450. [CrossRef]
- Adhikari, P.; Oh, Y.; Panthee, D.R. Current Status of Early Blight Resistance in Tomato: An Update. Int J Mol Sci 2017, 18. [CrossRef]
- Rao, E.S.; Munshi, A.D.; Sinha, P.; Rajkumar Genetics of Rate Limiting Disease Reaction to Alternaria Solani in Tomato. Euphytica 2008, 159, 123–134. [CrossRef]
- Gardner, R.G.; Panthee, D.R. NC 1 CELBR and NC 2 CELBR: Early Blight and Late Blight-Resistant Fresh Market Tomato Breeding Lines. HortScience 2010, 45, 975–976. [CrossRef]
- Bihon, W.; Ognakossan, K.E.; Tignegre, J.B.; Hanson, P.; Ndiaye, K.; Srinivasan, R. Evaluation of Different Tomato (Solanum Lycopersicum L.) Entries and Varieties for Performance and Adaptation in Mali, West Africa. Horticulturae 2022, Vol. 8, Page 579 2022, 8, 579. [CrossRef]
- Adhikari, T.B.; Siddique, M.I.; Louws, F.J.; Sim, S.-C.; Panthee, D.R. Molecular Mapping of Quantitative Trait Loci for Resistance to Early Blight in Tomatoes. Front Plant Sci 2023, 14, 1684. [CrossRef]
- Beavis, W.D. QTL Analyses: Power, Precision, and Accuracy. Molecular Dissection of Complex Traits 1998, 145–162. [CrossRef]
- Carvalho, L.J.C.B.; Schaal, B.A. Assessing Genetic Diversity in the Cassava (Manihot Esculenta Crantz) Germplasm Collection in Brazil Using PCR-Based Markers. Euphytica 2001, 120, 133–142. [CrossRef]
- Panthee, D.R.; Piotrowski, A.; Ibrahem, R. Mapping Quantitative Trait Loci (QTL) for Resistance to Late Blight in Tomato. International Journal of Molecular Sciences 2017, Vol. 18, Page 1589 2017, 18, 1589. [CrossRef]
- Akhtar, K.P.; Ullah, N.; Saleem, M.Y.; Iqbal, Q.; Asghar, M.; Khan, A.R. Evaluation of Tomato Genotypes for Early Blight Disease Resistance Caused by Alternaria Solani in Pakistan. Journal of Plant Pathology 2019, 101, 1159–1170. [CrossRef]
- Singh, A.K.; Rai, N.; Singh, R.K.; Saha, S.; Rai, R.K.; Singh, R.P. Genetics of Resistance to Early Blight Disease in Crosses of Wild Derivatives of Tomato. Sci Hortic 2017, 219, 70–78. [CrossRef]
- IOP Conf Ser Earth Environ Sci 2020, 482. [CrossRef]
- Oliveira, M.D.M.; Varanda, C.M.R.; Félix, M.R.F. Induced Resistance during the Interaction Pathogen x Plant and the Use of Resistance Inducers. Phytochem Lett 2016, 15, 152–158. [CrossRef]
- Tripathi, D.; Raikhy, G.; Kumar, D. Chemical Elicitors of Systemic Acquired Resistance—Salicylic Acid and Its Functional Analogs. Curr Plant Biol 2019, 17, 48–59. [CrossRef]
- Camejo, D.; Martí, M.D.C.; Nicolás, E.; Alarcón, J.J.; Jiménez, A.; Sevilla, F. Response of Superoxide Dismutase Isoenzymes in Tomato Plants (Lycopersicon Esculentum) during Thermo-Acclimation of the Photosynthetic Apparatus. Physiol Plant 2007, 131, 367–377. [CrossRef]
- Belkhadir, Y.; Nimchuk, Z.; Hubert, D.A.; Mackey, D.; Dangl, J.L. Arabidopsis RIN4 Negatively Regulates Disease Resistance Mediated by RPS2 and RPM1 Downstream or Independent of the NDR1 Signal Modulator and Is Not Required for the Virulence Functions of Bacterial Type III Effectors AvrRpt2 or AvrRpm1. Plant Cell 2004, 16, 2822–2835. [CrossRef]
- Zhang, C.; Liu, L.; Zheng, Z.; Sun, Y.; Zhou, L.; Yang, Y.; Cheng, F.; Zhang, Z.; Wang, X.; Huang, S.; et al. Fine Mapping of the Ph-3 Gene Conferring Resistance to Late Blight (Phytophthora Infestans) in Tomato. Theor Appl Genet 2013, 126, 2643–2653. [CrossRef]
- Zhang, C.; Liu, L.; Wang, X.; Vossen, J.; Li, G.; Li, T.; Zheng, Z.; Gao, J.; Guo, Y.; Visser, R.G.F.; et al. The Ph-3 Gene from Solanum Pimpinellifolium Encodes CC-NBS-LRR Protein Conferring Resistance to Phytophthora Infestans. Theor Appl Genet 2014, 127, 1353. [CrossRef]
- Robbins, M.D.; Masud, M.A.T.; Panthee, D.R.; Gardner, R.G.; Francis, D.M.; Stevens, M.R. Marker-Assisted Selection for Coupling Phase Resistance to Tomato Spotted Wilt Virus and Phytophthora Infestans (Late Blight) in Tomato. HortScience 2010, 45, 1424–1428. [CrossRef]
- Foolad, M.R.; Panthee, D.R. Marker-Assisted Selection in Tomato Breeding. CRC Crit Rev Plant Sci 2012, 31, 93–123. [CrossRef]
- Brouwer, D.J.; St. Clair, D.A. Fine Mapping of Three Quantitative Trait Loci for Late Blight Resistance in Tomato Using near Isogenic Lines (NILs) and Sub-NILs. Theoretical and Applied Genetics 2004, 108, 628–638. [CrossRef]
- Brouwer, D.; Jones, E.; Genome, D.C.-; 2004, undefined QTL Analysis of Quantitative Resistance to Phytophthora Infestans (Late Blight) in Tomato and Comparisons with Potato. Genome 2004, 47, 475–492. [CrossRef]
- Haggard, J.E.; Johnson, E.B.; St. Clair, D.A. Linkage Relationships among Multiple QTL for Horticultural Traits and Late Blight (P. Infestans) Resistance on Chromosome 5 Introgressed from Wild Tomato Solanum Habrochaites. G3: Genes, Genomes, Genetics 2013, 3, 2131–2146. [CrossRef]
- Haggard, J.E.; Johnson, E.B.; St. Clair, D.A. Multiple QTL for Horticultural Traits and Quantitative Resistance to Phytophthora Infestans Linked on Solanum Habrochaites Chromosome 11. G3: Genes, Genomes, Genetics 2015, 5, 219–233. [CrossRef]
- Johnson, E.B.; Erron Haggard, J.; St.Clair, D.A. Fractionation, Stability, and Isolate-Specificity of QTL for Resistance to Phytophthora Infestans in Cultivated Tomato (Solanum Lycopersicum). G3: Genes, Genomes, Genetics 2012, 2, 1145–1159. [CrossRef]
- Li, J.; Liu, L.; Bai, Y.; Finkers, R.; Wang, F.; Du, Y.; Yang, Y.; Xie, B.; Visser, R.G.F.; van Heusden, A.W. Identification and Mapping of Quantitative Resistance to Late Blight (Phytophthora Infestans) in Solanum Habrochaites LA1777. Euphytica 2011, 179, 427–438. [CrossRef]
- Haggard, J.E.; St.Clair, D.A. Combining Ability for Phytophthora Infestans Quantitative Resistance from Wild Tomato. Crop Sci 2015, 55, 240–254. [CrossRef]
- Smart, C.D.; Tanksley, S.D.; Mayton, H.; Fry, W.E. Resistance to Phytophthora Infestans in Lycopersicon Pennellii. Plant Dis 2007, 91, 1045–1049. [CrossRef]
- Hanson, P.; Lu, S.F.; Wang, J.F.; Chen, W.; Kenyon, L.; Tan, C.W.; Tee, K.L.; Wang, Y.Y.; Hsu, Y.C.; Schafleitner, R.; et al. Conventional and Molecular Marker-Assisted Selection and Pyramiding of Genes for Multiple Disease Resistance in Tomato. Sci Hortic 2016, 201, 346–354. [CrossRef]
- Wang, Y.Y.; Chen, C.H.; Hoffmann, A.; Hsu, Y.C.; Lu, S.F.; Wang, J.F.; Hanson, P. Evaluation of the Ph-3 Gene-Specific Marker Developed for Marker-Assisted Selection of Late Blight-Resistant Tomato. Plant Breeding 2016, 135, 636–642. [CrossRef]
- Sullenberger, M.T.; Jia, M.; Gao, S.; Ashrafi, H.; Foolad, M.R. Identification of Late Blight Resistance Quantitative Trait Loci in Solanum Pimpinellifolium Accession PI 270441. Plant Genome 2022, 15, e20251. [CrossRef]
- Ohlson, E.W.; Ashrafi, H.; Foolad, M.R. Identification and Mapping of Late Blight Resistance Quantitative Trait Loci in Tomato Accession PI 163245. Plant Genome 2018, 11, 180007. [CrossRef]
- Kimura, S.; Sinha, N. Tomato (Solanum Lycopersicum): A Model Fruit-Bearing Crop. Cold Spring Harb Protoc 2008, 2008, pdb.emo105. [CrossRef]
- Joshi, B.K.; Louws, F.J.; Yencho, G.C.; Sosinski, B.R.; Arellano, C.; Panthee, D.R. Molecular Markers for Septoria Leaf Spot (Septoria Lycopersicii Speg.) Resistance in Tomato (Solanum Lycopersicum L.). Nepal Journal of Biotechnology 2015, 3, 40–47. [CrossRef]
- Blum, E.; Liu, K.; Mazourek, M.; Yoo, E.Y.; Jahn, M.; Paran, I. Molecular Mapping of the C Locus for Presence of Pungency in Capsicum. Genome 2002, 45, 702–705. [CrossRef]
- Zitter, T.A.; Mutschler-Chu, M.A. Choosing LB, EB and SLS Resistant Tomato Varieties for 2014 What Tomato Growers Need to Know About Foliar Disease Resistance Issues? Cornell University: Cooperative Extension 2013.
- Satelis, J.F.; Boiteux, L.S.; Reis, A. Resistance to Septoria Lycopersici in Solanum (Section Lycopersicon) Species and in Progenies of S. Lycopersicum × S. Peruvianum. Sci Agric 2010, 67, 334–341. [CrossRef]
- Camejo, D.; Nicolás, E.; Torres, W.; Alarcón, J.J. Differential Heat-Induced Changes in the CO2 Assimilation Rate and Electron Transport in Tomato (Lycopersicon Esculentum Mill.). The Journal of Horticultural Sciences and Biotechnology 2010, 85, 137–143. [CrossRef]
- Sato, S.; Tabata, S.; Hirakawa, H.; Asamizu, E.; Shirasawa, K.; Isobe, S.; Kaneko, T.; Nakamura, Y.; Shibata, D.; Aoki, K.; et al. The Tomato Genome Sequence Provides Insights into Fleshy Fruit Evolution. Nature 2012 485:7400 2012, 485, 635–641. [CrossRef]
- Rothan, C.; Diouf, I.; Causse, M. Trait Discovery and Editing in Tomato. The Plant Journal 2019, 97, 73–90. [CrossRef]
- Alwala, S.; Suman, A.; Arro, J.A.; Veremis, J.C.; Kimbeng, C.A. Target Region Amplification Polymorphism (TRAP) for Assessing Genetic Diversity in Sugarcane Germplasm Collections. Crop Sci 2006, 46, 448–455. [CrossRef]
- Albuquerque, P.; Caridade, C.M.R.; Rodrigues, A.S.; Marcal, A.R.S.; Cruz, J.; Cruz, L.; Santos, C.L.; Mendes, M. V.; Tavares, F. Evolutionary and Experimental Assessment of Novel Markers for Detection of Xanthomonas Euvesicatoria in Plant Samples. PLoS One 2012, 7, e37836. [CrossRef]
- Liu, S.; Yeh, C.T.; Tang, H.M.; Nettleton, D.; Schnable, P.S. Gene Mapping via Bulked Segregant RNA-Seq (BSR-Seq). PLoS One 2012, 7, e36406. [CrossRef]
- Chen, Y.; Lun, A.T.L.; Smyth, G.K. Differential Expression Analysis of Complex RNA-Seq Experiments Using EdgeR. Statistical Analysis of Next Generation Sequencing Data 2014, 51–74. [CrossRef]
- Suresh, B.V.; Roy, R.; Sahu, K.; Misra, G.; Chattopadhyay, D. Tomato Genomic Resources Database: An Integrated Repository of Useful Tomato Genomic Information for Basic and Applied Research. PLoS One 2014, 9, e86387. [CrossRef]
- Yano, K.; Aoki, K.; Shibata, D. Genomic Databases for Tomato. Plant Biotechnology 2007, 24, 17–25. [CrossRef]
- Matsukura, C.; Aoki, K.; Fukuda, N.; Mizoguchi, T.; Asamizu, E.; Saito, T.; Shibata, D.; Ezura, H. Comprehensive Resources for Tomato Functional Genomics Based on the Miniature Model Tomato Micro-Tom. Curr Genomics 2008, 9, 436–443. [CrossRef]
- Barone, A.; Matteo, A.; Carputo, D.; Frusciante, L. High-Throughput Genomics Enhances Tomato Breeding Efficiency. Curr Genomics 2009, 10, 1. [CrossRef]
- Campbell, J.K.; Rogers, R.B.; Lila, M.A.; Erdman, J.W. Biosynthesis of 14C-Phytoene from Tomato Cell Suspension Cultures (Lycopersicon Esculentum) for Utilization in Prostate Cancer Cell Culture Studies. J Agric Food Chem 2006, 54, 747–755. [CrossRef]
- Shi, R.; Panthee, D.R. Transcriptome-Based Analysis of Tomato Genotypes Resistant to Bacterial Spot (Xanthomonas Perforans) Race T4. International Journal of Molecular Sciences 2020, Vol. 21, Page 4070 2020, 21, 4070. [CrossRef]
- Kim, M.; Nguyen, T.T.P.; Ahn, J.H.; Kim, G.J.; Sim, S.C. Genome-Wide Association Study Identifies QTL for Eight Fruit Traits in Cultivated Tomato (Solanum Lycopersicum L.). Hortic Res 2021, 8, 203. [CrossRef]
- Casa, A.M.; Pressoir, G.; Brown, P.J.; Mitchell, S.E.; Rooney, W.L.; Tuinstra, M.R.; Franks, C.D.; Kresovich, S. Community Resources and Strategies for Association Mapping in Sorghum. Crop Sci 2008, 48, 30–40. [CrossRef]
- Wang, X.; Gao, L.; Jiao, C.; Stravoravdis, S.; Hosmani, P.S.; Saha, S.; Zhang, J.; Mainiero, S.; Strickler, S.R.; Catala, C.; et al. Genome of Solanum Pimpinellifolium Provides Insights into Structural Variants during Tomato Breeding. Nature Communications 2020 11:1 2020, 11, 1–11. [CrossRef]
- Gao, L.; Gonda, I.; Sun, H.; Ma, Q.; Bao, K.; Tieman, D.M.; Burzynski-Chang, E.A.; Fish, T.L.; Stromberg, K.A.; Sacks, G.L.; et al. The Tomato Pan-Genome Uncovers New Genes and a Rare Allele Regulating Fruit Flavor. Nature Genetics 2019, 51, 1044–1051. [CrossRef]
- Gonda, I.; Ashrafi, H.; Lyon, D.A.; Strickler, S.R.; Hulse-Kemp, A.M.; Ma, Q.; Sun, H.; Stoffel, K.; Powell, A.F.; Futrell, S.; et al. Sequencing-Based Bin Map Construction of a Tomato Mapping Population, Facilitating High-Resolution Quantitative Trait Loci Detection. Plant Genome 2019, 12, 180010. [CrossRef]
- Chetty, V.J.; Ceballos, N.; Garcia, D.; Narváez-Vásquez, J.; Lopez, W.; Orozco-Cárdenas, M.L. Evaluation of Four Agrobacterium Tumefaciens Strains for the Genetic Transformation of Tomato (Solanum Lycopersicum L.) Cultivar Micro-Tom. Plant Cell Rep 2013, 32, 239–247. [CrossRef]
- Arshad, W.; Haq, I.U.; Waheed, M.T.; Mysore, K.S.; Mirza, B. Agrobacterium-Mediated Transformation of Tomato with RolB Gene Results in Enhancement of Fruit Quality and Foliar Resistance against Fungal Pathogens. PLoS One 2014, 9, e96979. [CrossRef]
- Khan, R.S.; Nakamura, I.; Mii, M. Development of Disease-Resistant Marker-Free Tomato by R/RS Site-Specific Recombination. Plant Cell Rep 2011, 30, 1041–1053. [CrossRef]
- Catanzariti, A.M.; Lim, G.T.T.; Jones, D.A. The Tomato I-3 Gene: A Novel Gene for Resistance to Fusarium Wilt Disease. New Phytologist 2015, 207, 106–118. [CrossRef]
- Kaplanoglu, E.; Kolotilin, I.; Menassa, R.; Donly, C. Plastid Transformation of Micro-Tom Tomato with a Hemipteran Double-Stranded RNA Results in RNA Interference in Multiple Insect Species. Int J Mol Sci 2022, 23, 3918. [CrossRef]
- Čermák, T.; Gasparini, K.; Kevei, Z.; Zsögön, A. Genome Editing to Achieve the Crop Ideotype in Tomato. Methods in Molecular Biology 2021, 2264, 219–244. [CrossRef]
- Nagamine, A.; Takayama, M.; Ezura, H. Genetic Improvement of Tomato Using Gene Editing Technologies. J Hortic Sci Biotechnol 2022, 98, 1–9. [CrossRef]
- Barka, G.D.; Lee, J. Advances in S Gene Targeted Genome-Editing and Its Applicability to Disease Resistance Breeding in Selected Solanaceae Crop Plants. Bioengineered 2022, 13, 14646–14666. [CrossRef]
- Bhargava, A.; Shukla, S.; Ohri, D. Evaluation of Foliage Yield and Leaf Quality Traits in Chenopodium Spp. in Multiyear Trials. Euphytica 2007, 153, 199–213. [CrossRef]
- Hong, Y.; Meng, J.; He, X.; Zhang, Y.; Liu, Y.; Zhang, C.; Qi, H.; Luan, Y. Editing Mir482b and Mir482c Simultaneously by Crispr/Cas9 Enhanced Tomato Resistance to Phytophthora Infestans. Phytopathology 2021, 111, 1008–1016. [CrossRef]
- Tiwari, J.K.; Singh, A.K.; Behera, T.K. CRISPR/Cas Genome Editing in Tomato Improvement: Advances and Applications. Front Plant Sci 2023, 14. [CrossRef]
Figure 1.
Early blight symptoms shown by infected plants: Concentric ring symptoms are seen on leaves on the left side (A), whereas stem girdling and spots on the right side (B) are caused by Alternaria linariae infection. (Photo by Anju Pandey).
Figure 1.
Early blight symptoms shown by infected plants: Concentric ring symptoms are seen on leaves on the left side (A), whereas stem girdling and spots on the right side (B) are caused by Alternaria linariae infection. (Photo by Anju Pandey).

Figure 2.
Late blight symptoms on leaf, stem, and fruits. Symptoms progress rapidly and can cover the whole plant within seven to ten days, showing a brown discoloration of infected tomatoes over the entire plant. The picture on the right is a resistant hybrid released as ‘Mountain Rouge’ (A) whereas the center and right are susceptible genotypes showing symptoms on the whole plant (B), and the fruits (C). (Photo by Dilip Panthee).
Figure 2.
Late blight symptoms on leaf, stem, and fruits. Symptoms progress rapidly and can cover the whole plant within seven to ten days, showing a brown discoloration of infected tomatoes over the entire plant. The picture on the right is a resistant hybrid released as ‘Mountain Rouge’ (A) whereas the center and right are susceptible genotypes showing symptoms on the whole plant (B), and the fruits (C). (Photo by Dilip Panthee).

Figure 3.
Symptoms on leaf and stem caused by Septoria leaf spot in tomato. A B, and C show symptoms on the leaf and stem. D. Black color pycnidia formed on spots developed by Septoria on leaves. (Photo by Anju Pandey).
Figure 3.
Symptoms on leaf and stem caused by Septoria leaf spot in tomato. A B, and C show symptoms on the leaf and stem. D. Black color pycnidia formed on spots developed by Septoria on leaves. (Photo by Anju Pandey).

Figure 4.
Tomato genotypes with various combinations of genes conferring resistance to late blight in tomatoes. In the figure, NC 2CELBR is homozygous for Ph2 and Ph3, Mountain Merit is heterozygous for Ph-2 and Ph-3, Plum Regal is heterozygous for Ph3, and NC 30P does not have any resistance gene. Phenotypic data were collected in the summer of 2013 when late blight infestation was extremely high at Mountain Research Station, Waynesville, NC.
Figure 4.
Tomato genotypes with various combinations of genes conferring resistance to late blight in tomatoes. In the figure, NC 2CELBR is homozygous for Ph2 and Ph3, Mountain Merit is heterozygous for Ph-2 and Ph-3, Plum Regal is heterozygous for Ph3, and NC 30P does not have any resistance gene. Phenotypic data were collected in the summer of 2013 when late blight infestation was extremely high at Mountain Research Station, Waynesville, NC.
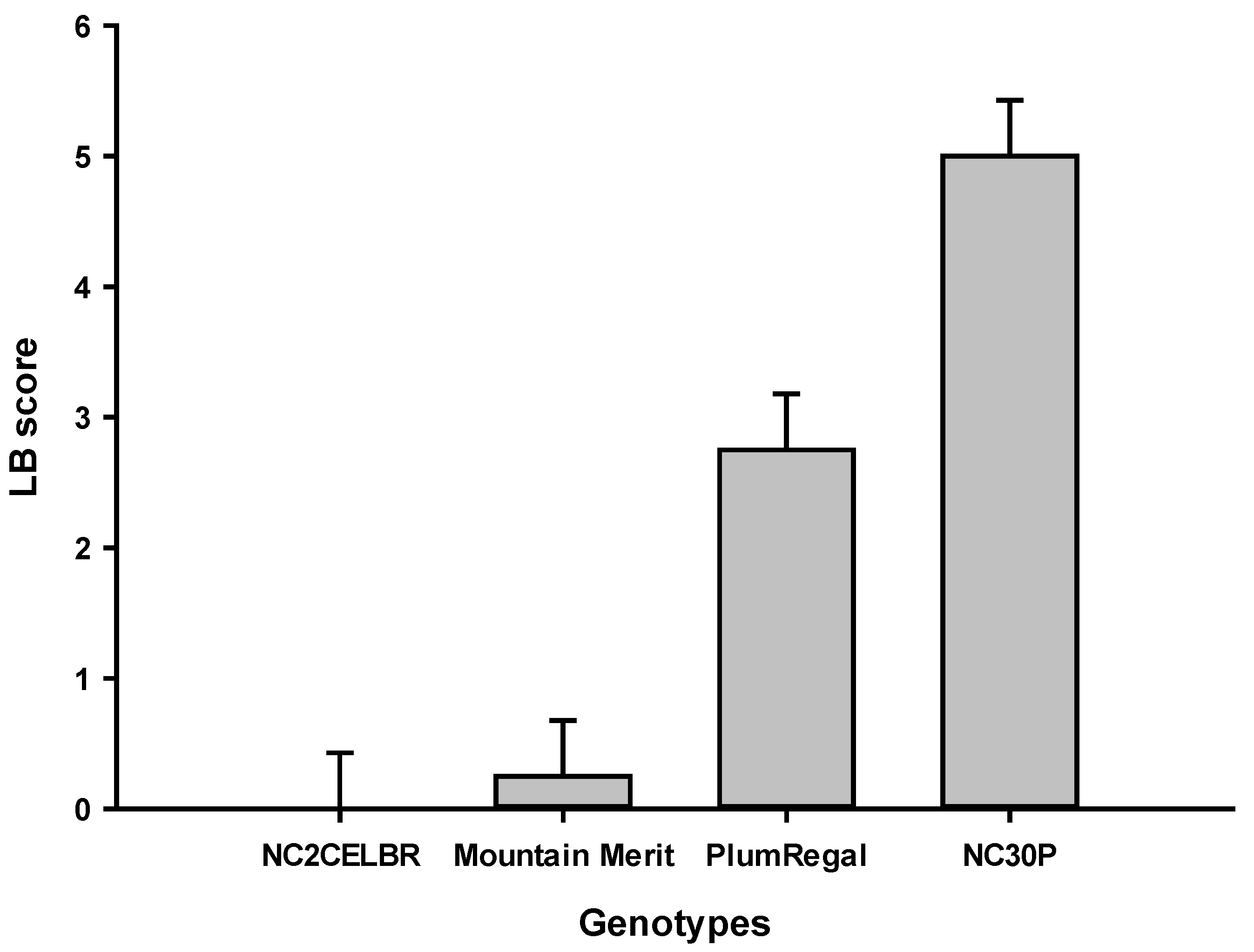
Table 1.
Total area (000 ha) under production, total production (million mt), yield (ton/ha), and percentage of the total production of tomato at the global level in 2022 [1].
Table 1.
Total area (000 ha) under production, total production (million mt), yield (ton/ha), and percentage of the total production of tomato at the global level in 2022 [1].
| Country | Production (million tons) | Area ('000 ha) | Yield (ton/ha) | World production (%) |
|---|---|---|---|---|
| China | 64.9 | 1111.5 | 58.4 | 34.7 |
| India | 20.6 | 812.0 | 25.3 | 11.0 |
| Turkey | 13.2 | 181.9 | 72.6 | 7.1 |
| USA | 12.2 | 110.4 | 110.7 | 6.5 |
| Egypt | 6.7 | 170.9 | 39.4 | 3.6 |
| Italy | 6.2 | 99.8 | 62.6 | 3.3 |
| Iran | 5.8 | 129.1 | 44.8 | 3.1 |
| Spain | 4.3 | 55.5 | 77.8 | 2.3 |
| Mexico | 4.1 | 84.9 | 48.7 | 2.2 |
| Brazil | 3.8 | 52.0 | 72.2 | 2.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



