
Submitted:
16 June 2023
Posted:
19 June 2023
You are already at the latest version
Abstract
With the increment of the aging population in recent years, neurodegenerative diseases exert a major global disease burden, essentially as a result of the lack of treatments that can stop the disease progression. Alzheimer’s Disease (AD) is an example of a neurodegenerative disease, that affects millions of people globally, with no effective treatment. Natural compounds have emerged as a viable therapy to fill a huge gap in AD management, and in recent years, mostly fuelled by the Covid-19 pandemic, RNA-based therapeutics have become a hot topic in the treatment of several diseases. Treatments of neurodegenerative diseases face significant limitations due to the complex and interconnected pathways that lead to their hallmarks and, also due to the necessity to cross the blood–brain barrier. Nanotechnology has contributed to surpass this bottleneck in the treatment of AD, by promoting safe and enhanced drug delivery to the brain. In particular, exosome-like nanoparticles, a hybrid delivery system combining exosomes and liposomes' advantageous features, are demonstrating great potential in the treatment of central nervous system diseases.
Keywords:
neurodegenerative diseases
; Alzheimer’s disease
; natural compounds
; RNA therapy
; blood-brain barrier
; exosome-like liposomes
1. Introduction
Neurodegenerative diseases are a diversified group of conditions characterized by progressive degeneration of the function of the central nervous system or peripheral nervous system, and the current therapeutic options do not provide a cure, only slowing down the disease progression. Together, neurodegenerative diseases exert a major burden in global healthcare systems, with dementia being a public health challenge in many developed countries, as aging is a strong risk factor [1].
Dementia is one of the highest global health crises of this century, with Alzheimer’s disease (AD) being the most common form of dementia. In the United States, an estimated 6.7 million individuals aged 65, and older, are living with AD in 2023, and the number is expected to reach 88 million by 2050 [2]. The estimated 2023 cost of caring for those with this disease is $345 billion. Between 2000 and 2019, the number of deaths from AD increased by 145%, while deaths from the number-one cause of death - heart disease - decreased 7.3% [2].
AD is a progressive, irreversible neurodegenerative disease, leading to memory impairment, impacting cognition, and can ultimately affect the behaviour, speech, visuospatial orientation, and the motor system. This disease is characterized by two major pathological hallmarks: progressive accumulation of amyloid beta (Aβ) plaques and neurofibrillary tangles (NFTs). Aβ damages neurons by interfering with neuron communication at synapses and NFTs block the transport of essential molecules for the normal function of neurons. Consequently, these lead to other complications such as oxidative stress, inflammation and brain atrophy due to cell loss [2,3].
Natural compounds or extracts are a viable therapy to fill the huge gap in the treatment of this disease, since they can target several hallmarks. However, for an improved efficacy of these compounds, they can be administered by delivery systems. Also, mostly instigated by the recent COVID-19 pandemic, RNA-based therapies have become a topic of great interest to researchers and pharmaceutical companies. RNA therapies promise to change the current conventional drugs that are not capable to target and treat all type of diseases, and several clinical studies are ongoing for a variety of RNA-based therapeutics against various incurable diseases. RNA therapy presents several advantages such as cost effectiveness, manufacture simplicity and the ability to target previously inaccessible pathways [4,5]. RNA therapies such as microRNAs (miRNAs), small interfering RNAs (siRNAs) and messenger RNAs (mRNAs) seem to be some of the most promising molecules for the treatment of AD [6]. For the effective delivery of RNA, this molecule has to overcome several obstacles. Their hydrophilic, negatively-charged properties make it difficult for the RNA molecule to passively diffuse across the cell’s membrane , and so, it has to undergo endocytosis and escape from the endosome to reach the cytoplasm. Furthermore, this molecule is highly susceptible to ribonucleases degradation and must have enhanced accumulation at targeted tissues [4].
Besides the complex mechanisms that lead to AD, the blood-brain barrier (BBB) is known to be a particular reason for the lack of effective treatments for AD. The BBB is a physiological barrier constituted by blood vessels that vascularize the central nervous system (CNS) and possess unique properties that make a precise control of the molecules allowed to enter the CNS [7].
Nanoparticles have been used to mitigate all of these hindrances in the delivery of RNA molecules into the brain. These drug delivery systems effectively protect RNA from degradation, enable the crossing of biological barriers, and allow a targeted accumulation and release [8]. In the last years, nanoparticles, such as dendrimers, polymeric nanoparticles and gold nanoparticles and carbon quantum dots have shown to be capable of crossing the BBB effectively [8]. Along with these nanoparticles, exosomes and liposomes are delivery systems with promising properties that allow them to cross the BBB. Exosomes are nano-sized extracellular vesicles (EVs) released into surrounding body fluids, by their parental cells, and carry cell-specific cargos of proteins, lipids, and genetic materials. These EVs can be selectively uptaken by neighbouring or distant cells far from their release. On the other hand, while liposomes are synthetic vesicles, comprised of one or several concentrically lipid bilayers surrounding an aqueous lumen, that can be created from cholesterol and natural phospholipids or synthetic surfactants [9,10].
Exosome-like liposomes are a novel concept of nanoparticles that combine the advantages of both the exosomes and liposomes, creating an unique delivery system, with several advantages such as the mimetic constituents of natural exosomes, high biocompatibility, small size, easy production, efficient transport and delivery of therapeutical compounds with low bioavailability (e.g. curcumin and RNA molecules), and the ability to load both hydrophilic and hydrophobic drugs [11].
2. Neurodegenerative Diseases
Neurodegenerative diseases are a heterogeneous group of neurological disorders characterized by a cognitive, psychiatric, and motor deficit, due to neuron loss [12]. In addition to these common features, there is also the fact that the current treatments do not supress the advancement and do not focus on the underlying cause of the diseases. Some of the main reasons for the lack of effective treatment in neurodegenerative diseases are the limitations imposed by the BBB, and also, the complex pathways that lead to the late diagnosis of the diseases [13].
Neurodegenerative diseases are characterised by: (1) protein aggregation; (2) disruptive proteostasis; (3) neuroinflammation; (4) oxidative stress; (5) synaptic failure; and (6) neuronal death (Figure 1). The presence of intracellular and/or extracellular protein aggregation is a key hallmark in a large variety of neurodegenerative disorders. These abnormally deposited proteins are found in brain regions, that when damaged, lead to physical vulnerability [1,14]. Several proteins are associated with neurodegenerative disorders:
Tau protein - microtubule-associated protein - encoded by the microtubule-associated protein tau (MAPT) gene. Tau is substantially expressed in the cytoplasm of neurons and carries an important role mainly in the stabilization and assembly of axonal microtubules and also in a variety of physiological processes, which include axonal transport, signal transmission between neurons, neurogenesis, myelination, motor function, neuronal excitability, glucose metabolism, iron homeostasis, and DNA protection [15,16].
Amyloid-β (Aβ) - derives from the amyloid precursor protein (APP) and this protein aggregates into amyloid plaques with Aβ polypeptides of 40 and 42 amino acids long [17].
Prion - another protein present in neurodegenerative diseases is the prion protein (PrP) encoded by the PRNP gene. In prion diseases, the prion protein misfolds, propagates and aggregates rapidly, being responsible for spreading neurodegeneration between cells, and, consequently, brain regions [14].
α-synuclein – this is a 140 amino acid protein highly expressed in the brain, encoded by the α-synuclein (SCNA) gene, and is abundant in filamentous neuronal and glial inclusions [12].
Disrupted proteostasis is also present in neurodegenerative disorders. In a healthy organism there are protein degradation systems to maintain protein homeostasis, with an important role in the clearance of toxic protein aggregates: the autophagy lysosomal pathway and the ubiquitin-proteosome system. However, these pathways lose activity in elderly individuals, contributing to the accumulation of toxic protein aggregates [14]. Additionally, neuroinflammation is a hallmark in these diseases. Peripheral immune cells don’t pass through the BBB and reach the brain. Actually, glial cells (microglia and astrocyte) substitute peripheral immune cells’ function in the brain. Microglia play a crucial role in defence functions on the brain and these cells are activated in signs of pathogens or injury. In neurotoxic conditions with several aggregated proteins, microglia activation is induced, leading to an unresolved activation. Consequently, microglia interact with astrocytes, which are also essential cells for the maintenance of neuronal health. These activation of glial cells leads to inflammation [14,18]. In neurodegenerative diseases oxidative stress is ubiquitous, and neurons are particularly susceptible to it due to high polyunsaturated fatty acid content in the membranes, high oxygen consumption, and low antioxidant defences, inducing an increased oxidation of proteins and nucleic acids, and lipid peroxidation [1]. Synaptic failure has been described in various neurodegenerative diseases. Synapses are the functional part of the connection between neurons and the key part of the physiological function of neurons. However, when some peripheral factor affects synapses, it leads to reduced synaptic density, supressing brain learning and memory function. Currently, these synaptic changes are the targets of many pharmacological interventions [19]. All these factors result in neuronal cell death and, in some neurodegenerative disorders, can result in brain volume loss [14].
Neurodegenerative diseases include, Parkinson’s disease (PD), amyotrophic lateral sclerosis (ALS), Huntington’s disease (HD), and AD, that will be the focus of this review [1].
The brain of PD’s patients is affected by the presence of intra-neuronal inclusion bodies - Lewy bodies – and by the accumulation of the protein α-synuclein, that spreads from on brain region to another. PD is characterized by movement disorder, motor function impairment of patients, and by other nonmotor symptoms, such as gastrointestinal issues and sleep disturbances [1,20].
ALS is a devastating, progressive disease, and in most of the cases the cause of this condition is unknown. However, oxidative stress and inflammation are significant mechanisms in this neurodegenerative disease, leading to oxidized metabolites. This disease is characterized by a deficit in motor neurons in the spinal cord and in the motor cortex of the cerebrum. Patients witness progressive muscle weakness and atrophy, and respiratory failure, due to the weakening of respiratory muscles, having a life expectancy of 15,8 months post diagnosis [21,22].
HD is an inherited neurodegenerative disorder caused by a mutation in the Huntingtin gene, an abnormal trinucleotide expansion, which gets translated into a mutant protein. The protein leads to the disruption of cellular molecular processes, which can involve both loss- and gain-of-function mechanisms. A hallmark of this disease is the degeneration of the striatum (caudate nucleus and putamen), with specific loss of efferent medium spiny neurons, and brain shrinkage. This disorder is characterized by movement disturbance, cognitive decline, coordination loss, depression, obsessive–compulsive disorder and other psychiatric symptoms. [23,24,25].
2.1. Alzheimer’s Disease
The pathological development of AD is complex and not yet fully understood.[1] At the microscopic level, in the early stage of the disease there are two central hallmarks of AD: amyloid plaques in the extracellular space and neurofibrillary tangles in the intracellular space (Figure 2A) [26].
The presence of amyloid plaques is a consequence of an abnormal accumulation and deposition of amyloid-β, a product of APP processing [27]. APP is a transmembrane precursor protein expressed in numerous human tissues, including the central nervous system, and can be cleaved by different proteases, leading to either the non-amyloidogenic or the amyloidogenic processing of APP [28,29]. APP plays an important role in the generation of Aβ in the brain since APP peptides generated by non-amyloidogenic processing can positively influence the metabolism, while the peptides produced by amyloidogenic processing have negative effects [28,30]. If the APP is cleaved by α-secretase, in the non-amyloidogenic pathway, it prevents the formation of the Aβ and, instead, sAPPα fragments are formed, which are described to be non-cytotoxic [26]. The amyloidogenic pathway includes the combined action of β- and γ- secretases, which generate Aβ peptides with different C-terminal residues [15,27]. Consequently, Aβ is composed of a series of polypeptides of varying lengths [31]. The deposition and accumulation of these Aβ polypeptides leads to the formation of amyloid plaques in the brain, resulting in neuroinflammation and synaptic dysfunction [17,26]. Aβ plaques have a major role in the progression of AD, and can thus be considered a therapeutic target [32]. However, the direct catalytic inhibition of β- and γ-secretases is not the key to AD prevention, as these proteins have an important physiological role [30].
The neurofibrillary tangles, the second major pathological finding in AD, are a consequence of the deposition of abnormally phosphorylated tau proteins and their aggregation in neurons [16]. The microtubule binding protein tau, a cytoskeletal protein produced by alternative splicing of the MAPT gene, has been identified as a key molecule in AD, but also in a series of neurodegenerative diseases referred to as tauopathies, in contrast to Aβ accumulation which is a characteristic exclusive to AD [15,33]. Under pathological conditions, mutations or post-translational modifications, such as phosphorylation, acetylation, ubiquitination, and truncation, of tau can lower its affinity to microtubules [33]. However, the abnormal hyperphosphorylation of this protein is the leading step to tauopathy in AD [34]. The hyperphosphorylated tau around the microtubule can neutralize its positive charges, leading to the dissociation of tau from the microtubule [35,36]. Once hyperphosphorylated, the protein loses its functions in the synthesis and stabilization of microtubules [37]. These tau monomers accumulate into tau oligomers, that act as intermediate structures [38]. These proteins accumulate in neurites and neuronal cell bodies, where it develops into insoluble intracellular aggregates and inclusion bodies, such as NFTs. Nevertheless, tau can also be secreted into the extracellular space either in its naked form or packaged in exosomes [35,36]. Several in vitro and in vivo studies have examined the ability of tau pathology to move from one neuron to the next [39].
Besides the accumulation of these two proteins in the brain, there are other hallmarks in AD, such as synaptic dysfunction, neuroinflammation, oxidative stress and higher activity of acetylcholinesterase (AChE), ultimately leading to neurodegeneration (Figure 2B). Synaptic impairment is responsible for the reduction of presynaptic vesicles release and for the decreasing of glutamatergic receptors [33]. Additionally, microglia cells develop more susceptibility to stimulus and produce inflammatory cytokines and chemokines, leading to cytotoxic and pro-inflammatory events. Consequently, this contributes to the deterioration of the BBB and to the inability to remove neurotoxic molecules, such as Aβ peptides and hyperphosphorylated tau, from the CNS [40]. Damages in the mitochondrial structure, integrity, and biogenesis lead to excessive reactive oxygen species (ROS) production, causing damage to the cellular structure. Oxidative damage is superior in the AD brain compared to the healthy brain of elderly individuals, which indicates that AD patients have a cellular imbalance between the production of reactive oxygen nitrogen species and the antioxidant defence systems, damaging proteins, lipids and nucleic acids and contributing to the progression of AD [17,41]. The equilibrium of distinct neurotransmitters systems, such as acetylcholine (Ach), is crucial for a healthy brain function [42]. In AD an alteration in the cholinergic system is often present, since there is a loss of cholinergic neurons that leads to an extensive decline of ACh, and consequently to a deficit of cholinergic transmission at pre-synaptic level. This ACh decrease is due to a higher activity of AChE, the enzyme responsible for its degradation [43]. All of the aforementioned microscopic hallmarks in AD alter the brain at a macroscopic level. These cause a brain shrinkage with cortical thinning and atrophy, that are highly common in AD, leading to decreased brain weight in most patients affected [3,27].
Currently, the therapeutical options for these patients are still very limited [44,45]. The main classes of drugs available to treat AD are acetylcholinesterase inhibitors, which include donepezil, rivastigmine and galantamine. Although, the equilibrium of different neurotransmitters, such as acetylcholine, play a key role in normal brain function, the cholinergic system is not the only system affected in this pathology. Thus, the cholinesterase inhibitors only treat symptoms, and do not prevent disease progression. Treatments directed to other hallmarks, such as amyloid-β accumulation and tau hyperphosphorylation failed to provide effects [42,46]. Therefore, an innovative approach that can efficiently treat and control each hallmark of this multifactorial disease is required for an effective AD treatment [47].
3. Natural Compounds for AD Treatment
Natural drugs are gaining an increased interest by both for the scientific academia and the pharmaceutical industries, for the therapy of several diseases. In the last years, due to their multiple beneficial properties, more than 100 natural products have been proposed as a promising approach for AD therapy [48]. Many molecules, including lignans, flavonoids, tannins, polyphenols, triterpenes, sterols, and alkaloids, can act by different pathways, since they have anti-inflammatory, anti-amyloidogenic, anticholinesterase, anti-inflammatory properties, and can reduce oxidative stress [49]. It is reasonable to speculate that the progression of AD could be slowed down, or even prevented by natural products working on multiple pathological targets [50]. Table 1 presents an array of natural compounds or extracts that were researched upon for AD, in the last fifteen years and found on PubMed and Web of Science.
3.1. Overcoming Limitations of Natural Compounds with Delivery Systems
Besides the multiple beneficial properties of natural products, these display several limitations such as low hydrophilicity, rapid metabolism and degradation, low bioavailability, reduced targeting, susceptibility to physiological media, and poor permeability through lipid bilayers. Consequently, in vivo, natural drugs require a high dose administration beyond a safe range, to result in an effective and safe bioavailability [48,79]. Nanotechnology represents a new method to overcome these challenges. Delivery systems for natural compounds lead to enhancement of pharmacological activity, by improving stability of drugs in vivo, bioavailability, controlled release, increasing accumulation of active ingredients in target sites, promoting the solubility of insoluble drugs and reducing the required doses to produce therapeutic effects [79]. Nanotechnology offers multiple advantages in the delivery of natural products, since by this method these drugs can exert their therapeutic effect in the treatment of AD (Figure 3) [80].
4. RNAs as a Promising Tool in the Treatment of AD
The small molecule- and protein-based therapies that interact with a particular biologic molecule to obtain a pharmacological response to control a disease have been successfully dealing with many diseases over the past years [81]. However, this conventional pharmacotherapy has several limitations. Protein-based medications target mainly proteins to inhibit its activity and only ∼1.5% of the human genome encodes proteins. Consequently, the range of disease targets of this drugs are limited and unable to meet the required demands [5,82]. Most protein-based drugs are too large to enter their target cells and therefore are only effective when their target molecule is extracellular, demonstrating difficult tissue penetration [82,83].
The study of RNA therapeutics started decades ago, leading to a long scientific journey. However, just recently this field of research has developed dramatically, as a result to the response to COVID-19 pandemic, that revealed how RNA-based therapeutics could lead to a new era of different and accessible new technology in combatting a wide range of diseases [40,84]. Besides the inherent instability of RNA, these molecules possess particular features and versatility, having multiple advantages over protein and DNA-based drugs. These features include inducement of protein coding, binding specificity to target molecules and inhibition of protein translation. Additionally, RNA has the capacity to recognize a wide range of ligands, targeting almost any genetic component within the cell, that are out of reach for the most established drug models, since protein drugs essentially act on genome-derived proteins to exert pharmacological effects [81,84].
RNA can be modified in the base, backbone and sugar, increasing target affinity and preventing nuclease digestion [85]. These modifications make them completely different from cellular RNAs transcribed from the genome [81]. Thus, unlike DNA-based therapeutics which must cross the cytoplasmic and nuclear membrane and, can integrate the host genome and cause a mutation, RNA therapeutics have no risk of chromosomal integration, exhibiting a safer profile [5,86]. Another important advantage of RNA therapy is its long-lasting effects when using siRNAs as a drug, benefiting patients who cannot receive frequent treatments [87]. The fast production of RNA-based therapies is also a distinct advantage, considering the increased knowledge that will provide a faster and easier design of RNA molecules, when compared to the process to produce novel protein-based drugs, which takes years. The vaccine’s rapid production and the successful reduction of the severity of the disease in infected people in the COVID-19 pandemic is an evidence of how quickly this type of therapies can be developed and implemented [86]. These RNA features, together with emerging innovative technologies in diverse scientific areas, such as nanomedicine and bioinformatics have motivated researchers to address RNA through different angles, which makes it a very promising tool for therapeutic applications [84,88].
A long scientific journey has led to prominent technological advances in the RNA field, and several new types of RNA molecules have been discovered, leading to the development of several studies designed to expecting the implement more effective treatments for diseases that still remain without any cure [40]. RNA-based therapeutics can be classified into five different categories: (1) messenger RNAs that encode for proteins; (2) small interfering RNAs that are double-stranded and primarily cause translational repression of their target protein; (3) microRNAs that are small RNAs that can either inhibit protein synthesis when they bind to an mRNA target (miRNA mimics) or free up mRNA by binding to the miRNA that represses the translation of that particular mRNA (miRNA inhibitors); (4) antisense oligonucleotides that are small (~15–25 nucleotides) single-stranded RNAs that can either promote or repress their targets expression; and, (5) aptamers that are short single-stranded nucleic acids that form secondary and tertiary structures, that inhibit several types of target molecules, including proteins [83,84,89].
The use of nucleic acid therapy has a limitless potential to treat not only neurological diseases, but also a considerable range of disorders, since preclinical studies in cellular and animal models proved that mRNAs and short RNAs can be a new class of medicine [88]. Considering the unmet needs of effective treatments for AD patients, it is crucial to evaluate diverse therapeutic targets and strategies to cure this disease [44]. Over the last years, Aβ and tau proteins had been the principal targets for researchers. Although being well documented hallmarks of AD, treatments involving the regulation of these proteins , unfortunately, are still an unsuccessful strategy [45,90]. In this review we are going to focus on RNA therapies such as siRNA, miRNA and mRNA for the treatment of AD (Figure 4) and describe some examples in Table 2.
miRNA
miRNAs are small non-coding single-stranded RNAs with about ~22 nucleotides of length [91]. Endogenous miRNAs are essential in cell development and plays a key role in post-transcriptional gene regulation, since they regulate the expression of multiple mRNAs both by promoting mRNA degradation and by blocking translation of multiple target mRNAs, inhibiting protein synthesis [84]. They can regulate mRNA translation by binding to the 3’untranslated region, allowing the reduction in the amount of target protein, instead of only inhibiting its activity [40,92]. In addition to gene expression regulation, miRNA can also act as signalling molecules for intercellular communication, revealing that it can be packaged into exosomes to exert this function [93]. The miRNA therapeutic strategy could be categorized into two types: miRNAs mimics and miRNAs inhibitors [5,82]. miRNA mimics are synthetic RNA molecules that are designed to act as endogenous miRNA to silence genes and can be applied when increased levels of mRNA are prevalent. In contrast, miRNAs inhibitors are synthetic ssRNA molecules and can interrupt the miRNA function via sequence-specific binding to mature miRNA, without causing gene silencing. This option is interesting, when protein synthesis restoration is needed [82]. These particular features allow miRNA to target multiple sites of various molecular deregulated cascades in disease conditions, similar to what occurs in AD [40,91]. miRNAs molecules have crucial functions in the nervous system, such as neuronal differentiation, neurite outgrowth and synaptic plasticity and are responsive to neuropathological processes, including oxidative stress, neuroinflammation and protein aggregation. This proves that miRNAs are key molecules in AD and dysfunction of miRNAs in this neurological disease is being recognized [45,91].
siRNA
siRNAs are short synthetic double-stranded RNA oligonucleotides (20-25 nt) that take advantage of the RNA interference pathway to silence gene expression, by targeting their complementary mRNA [83,84]. siRNAs offer promising therapeutics for brain disease treatment by directly blocking causative gene expression with high targeting specificity, requiring low effective dosages, and benefiting form a relatively simple drug development process [94]. Several siRNA-based therapeutics were already approved by FDA for other diseases, supporting their potential use for AD therapeutics [40].
mRNA
mRNA is a type of single-stranded RNA involved in protein synthesis. The role of mRNA is to carry protein information from the DNA in a cell’s nucleus to the cell’s cytoplasm. Compared to other RNA therapies, mRNA can provide advantages such as (1) safety, (2) effectiveness, particularly in slowly-dividing or non-dividing cells such as neural cells and, (3) better control in protein expression [40].

Table 2.
miRNA, siRNA and mRNA therapeutic applications in Alzheimer’s disease.
| Role in AD | Reference | |
|---|---|---|
| Types of miRNA | ||
| miR-101 | Significantly reduced the expression of a reporter under control of APP 3’-UTR in HeLa cells. | [95] |
| miR-106b | Overexpression of miR-106b inhibited Aβ1-42-induced tau phosphorylation at Tyr18 in SH-SY5Y cells stably expressing tau. | [96] |
| miR-137 | miR-137 inhibited increased expression levels of p-tau induced by Aβ1-42 in SH-SY5Y and inhibited the hyperphosphorylation of tau protein in a transgenic mouse model of AD. | [97] |
| miR-219 | In a Drosophila model that produces human tau, reduction of miR-219 exacerbated tau toxicity, while overexpression of miR-219 partially annulled toxic effects. | [98] |
| miR-17 | miR-17 inhibits elevated miR-17 in adult AD (5xFAD) mice microglia improves Aβ degradation. | [99] |
| miR-20b-5p | Treatment with miR-20b-5p reduced APP mRNA and protein levels in cultured human neuronal cells. | [100] |
| miR-29c | Over-expression of miR-29c in SH-SY5Y, HEK-293T cell lines and miR-29c in transgenic mice downregulated BACE1 protein levels. | [101] |
| miR-298 | miR-298 is a repressor of APP, BACE1, and the two primary forms of Aβ (Aβ40 and Aβ42) in a primary human cell culture model. Thus, miR-298 significantly reduced levels of ~55 and 50 kDa forms of the tau protein without significant alterations of total tau or other forms. | [102] |
| miR-485-5p | miR-485-5p overexpression facilitated the learning and memory capabilities of APP/PS1 mice and promoted pericyte viability and prohibited pericyte apoptosis in this model. | [103] |
| miR-9-5p | miR-9-5p overexpression inhibited Aβ25-35-induced mitochondrial dysfunction, cell apoptosis, and oxidative stress by regulating GSK-3β expression in HT22 cells. | [104] |
| miR-132 | miR-132 inhibited hippocampal iNOS expression and oxidative stress by inhibiting MAPK1 expression to improve the cognitive function of rats with AD. | [105] |
| miR-153 | Using miR-153 transgenic mouse model, was verified that miR-153 downregulated the expression of APP and APLP2 protein in vivo. | [106], p. 2 |
| Targeted gene silencing by siRNA | ||
| Tau | siRNA against MAPT can effectively suppress tau expression in vitro and in vivo without a specific delivery agent. | [107] |
| BACE1 | Polymeric siRNA nanomedicine targeting BACE1 in APP/PS1 transgenic AD mouse model can efficiently penetrate the BBB via glycemia-controlled glucose transporter-1–mediated transport, ensuring that siRNAs decrease BACE1 expression. | [94] |
| Presenilin1 (PS1) | Down regulation of PS1 and Aβ42 in IMR32 cells transfected with siRNA against PS1 was verified. | [108] |
| APP | Infusion of siRNAs that down-regulated mouse APP protein levels into the ventricular system for 2 weeks down-regulated APP mRNA in mouse brain. | [109] |
| Proteins encoded by mRNA | ||
| mRNA encoding neprilysin | Neprilysin plays a major role in the clearance of Aβ in the brain. New mRNA therapeutic strategy utilizing mRNA encoding the mouse neprilysin protein has shown to decrease Aβ deposition and prevent pathogenic changes in the brain. | [110] |
4.1. Overcoming Limitations of RNA Therapies with Delivery Systems
In spite of being hot research topic in the present days, the development of novel RNA therapeutics has proven to be highly challenging in the past two decades. Some of the major disadvantages that stop RNA from being a clinical success are its instability for in vivo application, since the human body has several intrinsic defence systems to protect the cells against exogenous molecules, such as ribonucleases (RNases). RNA’s low targeted tissue accumulation, decreases its therapeutic efficacy, requiring high therapeutic RNA doses that can induce toxicity [111]. Early degradation of naked RNAs is another hurdle for their therapeutic efficiency, since they tend to be promptly eliminated from the body, via renal or hepatic clearance, shortly after the systemic administration [82,111]. For reference, the half-life of naked siRNA is about 15 minutes, and the half-life of naked mRNA can vary between 2 and 25 minutes, depending on the presence of 5’ capping, the length of the 3’ poly-A tail and on the RNA secondary structure.[112] Finally, being a large and negatively charged molecule, RNA is difficult to deliver into the cellular cytoplasm, where it exerts its action. Even if cellular internalization of RNA occurs, there is also the risk that RNA can’t escape the endosomal pathway, with only ~1–2% of siRNAs uptaken by the cells escaping the endosome [81,82].
These obstacles have been considerably surpassed, thanks to the recent advancements in research areas such as RNA biology and nanotechnology, that allowed the development of new materials and technologies for the delivery of RNA molecules [81,84]. These new advances transformed RNA technology into the novel therapeutic tool, since RNA can now be safely transported and delivered to the target, thanks to delivery systems [82] (Figure 5).
RNA delivery is key for the treatment of diseases such as neurological disorders, as it gives RNA the ability to target diseases that cannot be treated with other conventional drug groups, by encapsulating these molecules in delivery vectors. These RNA vectors have the capacity to effectively protect this molecule from biodegradation, increase bioavailability, solubility and permeation, surpass biologic barriers and promote a targeted delivery and release. Hence, RNA nanoencapsulation potentiate the treatment of diseases, by silencing genes or expressing therapeutic proteins [113,114].
5. Nanoparticles and the BBB
Delivery systems can overcome that limit RNA becoming a successful therapy, particularly for facilitating the crossing of the blood-brain barrier, which is the main culprit for the shortage of new and effective treatments for AD [115].
In this review we will explain the BBB anatomic composition and characteristics, as well as the BBB pathways into the CNS. Subsequently, we will discuss exosomes, liposomes and exosome-like liposomes as possible tools for RNA transport across the BBB. From an anatomic point of view, the BBB is composed by different cell types such as endothelial cells, pericytes and astrocytes (Figure 6A)[116]. In between the endothelial cells there are tight junctions , which are surrounded by a thin basal membrane and astrocytes vascular feet . These highly restrictive tight junctions, , are a key BBB feature, since they are responsible for the barrier properties and limit the transfer of almost all drugs [116,117]. The pericytes cover 20% of the outer surface of endothelial cells and are responsible for the regulation of the blood flow in the brain capillary through contraction and relaxation. The astrocytes are glial cells connect the brain capillary and neurons and also maintain BBB functions, by providing nutrients to neurons and protecting the brain from oxidative stress and metal toxicity [117]. Additionally, the basal membrane provides structural support around the pericytes and endothelial cells [118].
This specialized barrier acts like an interface with the capacity to regulate the entry of plasma components, red blood cells and leukocytes into the CNS, and ensures the export of potentially neurotoxic molecules, from the brain to the blood [119,120] This barrier strictly controls the molecule movements between the blood and the brain, regulating the homeostasis of the nervous system [119]. Moreover, more than 98% of all small molecule drugs and approximately 100% of biologic drugs are incapable to cross the BBB. Additionally, water-soluble molecules in the blood are prevented from entering the CNS, while lipid-soluble molecules are reduced by enzymes or efflux pumps [115].
The BBB possess several permanently active transport mechanisms to ensure the transport of nutrients into the CNS while excluding blood-borne molecules that could be detrimental [121]. On one hand, these BBB properties are proof of its vital role in maintaining the specialized microenvironment of the brain tissue. On the other hand, these features makes CNS access one of the most difficult of the body, limiting the development of novel effective drugs to treat AD [119]. Even though the BBB is a strict barrier for the circulation of molecules between the blood and the CNS, there are a few pathways that allow the delivery of essential molecules that maintain the brain homeostasis (Figure 5B). These include transcellular pathway, paracellular pathway, efflux pumps, carrier mediated transcytosis, receptor-mediated transcytosis, and adsorptive mediated transcytosis [114].
In a healthy BBB, transcellular diffusion (Figure 6B1) consists in the diffusion of solute particles through the endothelial cells. Particles transported through this route are small lipophilic molecules that penetrate through the cells. On the other hand, paracellular transport (Figure 5B2) is restricted by the tight junctions between the endothelial cells, allowing only hydrophilic molecules to pass, with a molecular weight <500 Da.
Efflux pumps (Figure 6B3) are a set of proteins, responsible for limiting the accumulation of various potentially toxic molecules, and eventually, for expelling these molecules from the brain. These proteins are a limiting factor for the delivery of bioactive compounds to the brain.
The carrier mediated transcytosis (Figure 6B4) consists in an active transport with the support of carrier proteins such as glucose transporter isoform (GLUT-1), large amino acid transporter (LAT), allowing the entrance to glucose or amino acids. The transport of these molecules occurs when they bind to the protein at the blood side of the BBB, and a subsequent conformational change allows their transport to the brain [114,123].
Receptor-mediated transcytosis (Figure 6B5) is a specialized transport system by which endogenous molecules can cross the BBB, through receptors present on the cell surface. This type of transport relies on the following mechanisms: endocytosis, intracellular vesicular trafficking and exocytosis. Active components bind to their specific receptors, on the luminal side of the endothelial cells, and an intracellular vesicle is formed through membrane invagination. The formed vesicles cross the cell to release the ligand at the basolateral side via exocytosis. The most common receptors involved in this process are the transferrin receptor (TfR), the insulin and insulin like growth factor receptors, the low-density lipoprotein receptor (LDLR), the low-density lipoprotein -receptor-related protein 1 and 2 (LRP1 and LRP2), the scavenger receptor class B type I (SR-B1), the leptin receptor, the albumin receptor, and the lactoferrin receptor. Receptor-mediated transcytosis is one of the one of the most promising pathways for nanoparticle drug delivery through the BBB. [114,122].
Adsorptive-mediated transcytosis (Figure 6B6) is another important BBB-crossing pathway, without the involvement of specific plasma-membrane receptors. The basic mechanism is responsible for the transport of charged particles, by taking advantage of the electrostatic interactions between the positively charged drug carriers and the negatively charged luminal membrane of the brain endothelial cells. This transport has lower affinity but higher capacity compared to receptor mediated transcytosis [114,122,123].
There is a crucial need for an ideal and safe approach to effectively carry pharmaceutical agents in a target-specific and sustained-release manner into the CNS, without disrupting the BBB [122,123]. A promising approach is taking advantage of the receptor-mediated transcytosis for drug delivery to the brain with the help of ligand functionalized nanoparticles. Nanoparticles are gaining popularity as drug carriers for the treatment of neurological disorders, , due to their small size and unique physical properties [124]. By virtue of their biochemical composition, lipidic nanoparticles provide biomimicking and bio-degradable platforms. As a result of their lipophilicity and size, nanosized particles such as exosomes and liposomes are promising drug delivery carriers to increase penetration of the BBB [113,125].
5.1. Exosomes
Exosomes are a subset of EVs with a diameter ranging from 30 to100 nm, and their composition includes lipids, proteins and nucleic acids [126,127]. Their lipid content incudes sphingomyelin, phosphatidylserine, cholesterol, and ceramide, or derivatives. Exosomes also carry non-specific proteins (e.g., cytoplasmic enzymes, cytosolic proteins, heat shock proteins, and transferring proteins) and specific proteins that differ from one exosome to another, depending on their origin. The genetic material of these vesicles includes microRNAs, mRNAs, long non-coding RNAs, and DNA fragments [126]. Exosomes are also enriched in late endosome components such as CD63, CD9 and CD81, since they originate from the endocytic compartment of the producer cell, in a process that generates multi-vesicular endosomes (MVEs), that subsequently fuse with the plasma membrane to release exosomes into the extracellular space.
Exosomes yield information that can reflect the phenotype of the parental cell, since these vesicles carry distinct RNA and protein cargoes, that allow the identification of their parental cells, as well as cell-specific or tissue specific factors that can be used to determine their site of origin [128,129]. Also, when exosomes are secreted, undesirable proteins and other molecules are discarded, making these vesicles a compartment of cellular debris for subsequent disposal [130].
Originally, exosomes were mainly described as a via for elimination of excessive and unnecessary molecules from the cells. However, in the last decade, it has been shown that they have other key functions in in both physiological and pathological processes. In regard of the physiological roles, exosomes play an important role in intercellular communication, since they are able to deliver a number of bioactive cargos to near or distant target cells [126]. Moreover, exosomes display a role in tissue homeostasis and have anti-inflammatory functions. An example of their function is the neuronal communication via secretion of exosomes, that can contribute to a range of neurobiological functions, including synaptic plasticity., In relation to the pathological functions, they control the expansion and progression of diseases, such as cancer and neurodegenerative diseases [130]. Exosomes are involved in the complex mechanisms of secretion, spread, and degradation of the Aβ and tau proteins and are, specially, involved in tau propagation between neuronal cells [131].
The unique properties of exosomes include their small size, durability, stability, potential cell selectivity, low immunogenicity, and ideal biocompatibility. These properties make them good candidates to be a therapeutic delivery system. Compared to traditional therapeutic drugs, exosomes have higher potential to pass through the BBB, which helps the drugs they might carry to reach the CNS. Also, since exosomes can be isolated from all body fluids, they can be candidates for analysis as part of a non-invasive liquid biopsy [129,130,131].
5.2. Liposomes
In the last two decades, lipid-based nanoparticles (LNPs), especially liposomes, have been attractive nanometric delivery systems, for being the most well-studied nonviral platforms for the delivery of RNA molecules, and achieving significant clinical success [113,132] highlighted by the highly effective mRNA COVID-19 vaccines [132,133].
There are several types of LNPs such as liposomes, niosomes, transfersomes, nanoemulsions, solid lipid nanoparticles (SLNs), lipid nanocapsules (LNCs), nanostructured lipid carriers (NLCs), lipid-based micelles, core–shell lipid nanoparticles (CLNs), and hybrid lipid-polymeric nanoparticles [132,134]. Solid lipid nanoparticles (SNLPs) are nanoparticles , that combines all the advantageous characteristics of delivery systems such as polymeric nanoparticles and liposomes.[135], [136] LNPs range in size from 40 nm to 1000 nm and are colloidal lipophilic systems constituted by of four main components; a pH-sensitive cationic lipid, a helper lipid, cholesterol, and a PEG-lipid. The cationic lipid is a synthetic lipid, constituted by a hydrophilic head with a protonable tertiary amino group (pKa 6–6.7), and a long hydrophobic tail. Cholesterol is incorporated into the LNPs formulation with the goal of increasing their flexibility, whilst the helper phospholipid assists the process of endosomal escape and contribute to the stability of LNPs. Finally, The insertion of a short-chain PEG-lipid derivative (normally of 14 carbon atoms) is essential to maximize the ex vivo stability, and control the particle size before administration [137].
Due to their resemblance to biological and natural components, these nano systems show tremendous promise as carriers for therapeutic applications. The main advantages of LNPs over other nanoparticles are their low toxicity and biocompatibility, biodegradability, safety, high mechanical and chemical versatility, and the capacity to protect the active ingredient from degradation processes induced by external factors. Along with these features, these lipid-based nanoparticles can incorporate and delivery of both hydrophobic and hydrophilic molecules, and most of their preparation methods can be easily scaled up. Additionally, because of their lipophilicity, LNPs possess the ability to overcome difficult physiological barriers, such as the BBB, even without surface modification [124,134].
Among all the nano-based drug delivery systems, liposomes are the most biocompatible and least toxic, since they are composed by phospholipids and cholesterol, the main components of cell membranes. Liposomes are an extremely versatile nanocarrier platform that have the capacity of loading multiple drugs, provide protection from degradation, have controlled and targeted drug release, and enhance drug endocytosis into cells [11]. Additionally, these nanoparticles are able to incorporate both hydrophilic and hydrophobic therapeutic agents. The hydrophilic compounds may either be entrapped into the aqueous core of the liposomes, or be located at the interface between the lipid bilayer and the external water phase, while the hydrophobic compounds are generally entrapped in the hydrophobic core of their lipid bilayers. The of positively charged lipids in the liposome’s constitution, allow the electrostatic interaction with negatively charged nucleic acids, such as DNA and RNA, in gene delivery applications [138].
There is evidence that lipid-based nanoplatforms will play a key role in the development of RNA neuro-therapies, with liposomes being one of the main lipidic platforms for RNA-delivery to the CNS [113]. In order to enhance drug delivery into the CNS, the liposome surface can be modified by the inclusion of biologically active ligands, such as peptides, polysaccharides, antibodies, or aptamers, which specifically bind to receptors expressed on the surface of the brain endothelial cells, facilitating their binding and transport across the BBB. The addition of polyethylene glycol (PEG) offers superficial protection for the liposomes, by avoiding binding with plasma proteins, and hence, preventing their opsonization and subsequent clearance. PEGylation of liposomes prolongs their circulation time in the body and, also has a crucial role in brain drug delivery, allowing liposomes to cross the BBB [138].
5.3. Exosome-Like Liposomes as a Novel Strategy
Scientists have explored various nanomaterials for targeted delivery through the BBB with significant efficacy, such as dendrimers, polymeric nanoparticles, gold nanoparticles and exosome-like liposomes [8,139].
- Dendrimers: Dendrimers are highly branched, characterized by defined molecular weights and specific encapsulation properties. This type of delivery system is composed by a symmetrical polymeric macromolecules with large number of reactive surface groups, with three distinctive architectural components, an interior core, an interior layer consisting of repeating units radially attached to the inner core, and functional end groups on the outside layer. Because of these unique features, dendrimers can cross impaired BBB, target astrocytes and microglia after systemic administration in animal models [139].
- Polymeric nanoparticles: Polymeric nanoparticles can be produced from synthetic or natural polymers. However, to be applied in brain drug delivery, these nanoparticles need to be biodegradable and biocompatible. PBCA, PLA and PLGA nanoparticles are nanoparticles able to cross the BBB. These nanocarriers possess controlled drug release, targeting efficiency, and they can avoid phagocytosis by the reticuloendothelial system, thus improving the concentration of drugs in the brain [140].
- Gold nanoparticles: Nanoparticles (mostly < 10 nm in size) composed of a gold core and with covalently or non-covalently attached surface ligands. Multiple in vivo studies on rodents have shown that low amounts of this delivery system were able to cross the BBB. However, the greater amount of the administered dose was found in the liver and in the blood.[8] Additionally, Sela et al. proved that gold nanoparticles could penetrate the BBB of rat without the use of external field or surface modification and were found to be distributed uniformly in both hypothalamus and hippocampus indicating there is no selective binding in these regions of brain [141].
- Carbon quantum dots: This delivery system retains a polymeric core structure and various functional groups on the surface, facilitating their conjugation with drug molecules for specific delivery. Carrier with several efficient features for BBB crossing such as excellent biocompatibility and low toxicity due to the lack of metal elements, small size and possess photoluminescence which can be utilized to track the penetration of CDs through the BBB [117].
Besides the fact that these nanoparticles are promising in overcoming the BBB, exosome-like liposomes are also delivery systems with unique properties to achieve this goal. Exosomes and liposomes are promising nanocarriers with unique properties. However, these delivery systems have some limitations. For instance, liposomal targeting efficiency is limited, and they can induce immunogenicity,[11] whilst efficient and reliable isolation and purification is needed for the clinical application of exosomes. It is also crucial to identify appropriate strategies to increase loading capacity and specificity to use exosomes as a nanocarrier [10,142]. When comparing the advantages and disadvantages of exosomes and liposomes, it becomes evident that the two systems are complimentary, since the advantages of the one system mitigate the disadvantages of the other, and vice-versa (figure 7). So, the development of a system that incorporates the desirable features of the two carriers into one hybrid delivery system, led to an innovative carrier for drug delivery applications, the exosome-like liposome [11,143]. These novel exosome-like liposomes are formulated with a lipid composition that mimics that of exosomes, that impart them with some of the desirable characteristics of exosomes, such as enhanced passive targeting, biocompatibility and RES-evasion, and the ability to cross biological barriers, whilst allowing much higher encapsulation efficiencies and larger scale production in good manufacturing practice, that the use exosomes themselves does not allow.
Thus, particle size plays a major role in drug delivery efficacy, including in the therapeutic effect achieved. The endothelial cells’ slit width from 200 to 500 nm in width. Consequently, for the long-term circulation of these nanoparticles, they need to have a size that does not exceed 200 nm, leading to better brain drug delivery across the BBB. . Also, the fact that exosome-like liposomes are coated with this suitable hydrophilic polymer, leads to advantages such as escaping phagocytosis, avoiding opsonization, and further increasing the blood circulation time, since the hydrophobic nature of the nanoparticle containing the drug is shielded. These unique characteristics drive researchers to believe that this type of carrier can have a key impact in BBB-crossing, helping in the therapy of neurodegenerative diseases such as AD [123].
Additionally, exosome-like liposomes were highly efficient encapsulating curcumin, with encapsulation efficiencies ranging between 85 and 94% and were effective in delivering curcumin into neuronal cells to promote a superior neuroprotective effect, after oxidative insult, compared to free curcumin, at the same concentrations. Thus, these delivery systems have shown to be highly biocompatible, without significantly affecting cell viability or causing hemolysis. Finally, the exosome-like liposomes have shown to be internalized by zebrafish embryos and accumulate in lipid rich zones, such as the brain and yolk sac, also in stages where the BBB is already formed [144].
6. Conclusion
AD is a neurodegenerative disease that as a tremendous impact in people’s quality of life, all around an aging world. This burden is further impacted by the lack of therapies, due to the complex mechanisms that lead to the disease onset, and, also due to limitations that the BBB imposes on the entrance, and maintenance, of therapeutic molecules in the CNS.
However, the emergence of RNA as an especially versatile tool for the treatment of several diseases, has open multiple new possibilities for the development of effective treatments for AD.
The accelerated growth of RNA therapies requires the development of efficient delivery systems to transport a selectively deliver the RNA molecules into their target cells or tissues , due to the low bioavailability of these molecules.
Nanoparticles have provided new and safer avenues for the delivery of natural compounds and RNA. In this field, the novel exosome-like liposomes are positioning themselves as a prime solution for the vehiculation of therapeutical compounds to the CNS, due to the combination of the favourable characteristics of exosomes, with those of liposomes, which entitle scalable production in good manufacturing practices, that is currently unfeasible for the exosomes, themselves.
Patents
The authors are co-inventors in a patent application (EXOSOME-MIMETIC LIPOSOME COMPOSITION AND USE, Instituto Nacional da Propriedade Industrial PPP nº 116560 P, 3/7/2020).
Author Contributions
Conceptualization, J.R., I.L. and A.C.G.; investigation, formal analysis, J.R.; writing—original draft preparation, J.R.; writing—review and editing, J.R., I.L. and A.C.G.; supervision, I.L. and A.C.G.; funding acquisition, A.C.G. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the strategic programme UID/BIA/04050/2020 funded by National funds through the Fundação para a Ciência e Tecnologia I.P. Ivo Lopes is a recipient of a scholarship from the Direção Regional da Ciência e Tecnologia, Governo Regional dos Açores (M3.1.a/F/128/2015).
Data Availability Statement
Data will be made available upon request.
Acknowledgments
We acknowledge Mario Fernandes for support in revisions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bloomingdale P, Karelina T, Ramakrishnan V, et al. Hallmarks of neurodegenerative disease: A systems pharmacology perspective. CPT Pharmacomet Syst Pharmacol. 2022;11(11):1399-1429. [CrossRef]
- 2023 Alzheimer’s disease facts and figures. Alzheimers Dement. Published online March 14, 2023:alz.13016. [CrossRef]
- DeTure MA, Dickson DW. The neuropathological diagnosis of Alzheimer’s disease. Mol Neurodegener. 2019;14(1):32. [CrossRef]
- Zhu Y, Zhu L, Wang X, Jin H. RNA-based therapeutics: an overview and prospectus. Cell Death Dis. 2022;13(7):644. [CrossRef]
- Damase TR, Sukhovershin R, Boada C, Taraballi F, Pettigrew RI, Cooke JP. The Limitless Future of RNA Therapeutics. Front Bioeng Biotechnol. 2021;9:628137. [CrossRef]
- Luo M, Lee LKC, Peng B, Choi CHJ, Tong WY, Voelcker NH. Delivering the Promise of Gene Therapy with Nanomedicines in Treating Central Nervous System Diseases. Adv Sci. 2022;9(26):2201740. [CrossRef]
- Daneman R, Prat A. The Blood–Brain Barrier. Cold Spring Harb Perspect Biol. 2015;7(1):a020412. [CrossRef]
- Lombardo SM, Schneider M, Türeli AE, Günday Türeli N. Key for crossing the BBB with nanoparticles: the rational design. Beilstein J Nanotechnol. 2020;11:866-883. [CrossRef]
- Akbarzadeh A, Rezaei-Sadabady R, Davaran S, et al. Liposome: classification, preparation, and applications. Nanoscale Res Lett. 2013;8(1):102. [CrossRef]
- Zhang Y, Liu Y, Liu H, Tang WH. Exosomes: biogenesis, biologic function and clinical potential. Cell Biosci. 2019;9(1):19. [CrossRef]
- Antimisiaris S, Mourtas S, Marazioti A. Exosomes and Exosome-Inspired Vesicles for Targeted Drug Delivery. Pharmaceutics. 2018;10(4):218. [CrossRef]
- Forrest SL, Kovacs GG. Current Concepts of Mixed Pathologies in Neurodegenerative Diseases. Can J Neurol Sci J Can Sci Neurol. Published online March 31, 2022:1-17. [CrossRef]
- Masoudi Asil S, Ahlawat J, Guillama Barroso G, Narayan M. Nanomaterial based drug delivery systems for the treatment of neurodegenerative diseases. Biomater Sci. 2020;8(15):4109-4128. [CrossRef]
- Wilson DM, Cookson MR, Van Den Bosch L, Zetterberg H, Holtzman DM, Dewachter I. Hallmarks of neurodegenerative diseases. Cell. 2023;186(4):693-714. [CrossRef]
- Roda A, Serra-Mir G, Montoliu-Gaya L, Tiessler L, Villegas S. Amyloid-beta peptide and tau protein crosstalk in Alzheimer’s disease. Neural Regen Res. 2022;17(8):1666. [CrossRef]
- Liang SY, Wang ZT, Tan L, Yu JT. Tau Toxicity in Neurodegeneration. Mol Neurobiol. 2022;59(6):3617-3634. [CrossRef]
- Yu H, Wu J. Amyloid-β: A double agent in Alzheimer’s disease? Biomed Pharmacother. 2021;139:111575. [CrossRef]
- Xiao Y, Wang SK, Zhang Y, et al. Role of extracellular vesicles in neurodegenerative diseases. Prog Neurobiol. 2021;201:102022. [CrossRef]
- Wei Z, Wei M, Yang X, Xu Y, Gao S, Ren K. Synaptic Secretion and Beyond: Targeting Synapse and Neurotransmitters to Treat Neurodegenerative Diseases. Parmar M, ed. Oxid Med Cell Longev. 2022;2022:1-22. [CrossRef]
- Van Den Berge N, Ulusoy A. Animal models of brain-first and body-first Parkinson’s disease. Neurobiol Dis. 2022;163:105599. [CrossRef]
- Sun X, Song J, Huang H, Chen H, Qian K. Modeling hallmark pathology using motor neurons derived from the family and sporadic amyotrophic lateral sclerosis patient-specific iPS cells. Stem Cell Res Ther. 2018;9(1):315. [CrossRef]
- Xiong L, McCoy M, Komuro H, et al. Inflammation-dependent oxidative stress metabolites as a hallmark of amyotrophic lateral sclerosis. Free Radic Biol Med. 2022;178:125-133. [CrossRef]
- Mehler MF, Petronglo JR, Arteaga-Bracho EE, et al. Loss-of-Huntingtin in Medial and Lateral Ganglionic Lineages Differentially Disrupts Regional Interneuron and Projection Neuron Subtypes and Promotes Huntington’s Disease-Associated Behavioral, Cellular, and Pathological Hallmarks. J Neurosci. 2019;39(10):1892-1909. [CrossRef]
- Jimenez-Sanchez M, Licitra F, Underwood BR, Rubinsztein DC. Huntington’s Disease: Mechanisms of Pathogenesis and Therapeutic Strategies. Cold Spring Harb Perspect Med. 2017;7(7):a024240. [CrossRef]
- Machiela E, Southwell AL. Biological Aging and the Cellular Pathogenesis of Huntington’s Disease. J Huntingt Dis. 2020;9(2):115-128. [CrossRef]
- Gallego Villarejo L, Bachmann L, Marks D, Brachthäuser M, Geidies A, Müller T. Role of Intracellular Amyloid β as Pathway Modulator, Biomarker, and Therapy Target. Int J Mol Sci. 2022;23(9):4656. [CrossRef]
- Chen XQ, Mobley WC. Alzheimer Disease Pathogenesis: Insights From Molecular and Cellular Biology Studies of Oligomeric Aβ and Tau Species. Front Neurosci. 2019;13:659. [CrossRef]
- Guo Y, Wang Q, Chen S, Xu C. Functions of amyloid precursor protein in metabolic diseases. Metabolism. 2021;115:154454. [CrossRef]
- Liu X, Liu Y, Ji S. Secretases Related to Amyloid Precursor Protein Processing. Membranes. 2021;11(12):983. [CrossRef]
- Zhang T, Chen D, Lee TH. Phosphorylation Signaling in APP Processing in Alzheimer’s Disease. Int J Mol Sci. 2019;21(1):209. [CrossRef]
- Lane CA, Hardy J, Schott JM. Alzheimer’s disease. Eur J Neurol. 2018;25(1):59-70. [CrossRef]
- Beera AM, Seethamraju SM, Nori LP. Alzheimer’s Disease: Perspective on Therapeutic Options and Recent Hallmarks in Clinical Research. Int J Pharm Res Allied Sci. 2021;10(4):110-120. https://doi.org/10.51847/ViC6sAGCyq. [CrossRef]
- Wu M, Zhang M, Yin X, et al. The role of pathological tau in synaptic dysfunction in Alzheimer’s diseases. Transl Neurodegener. 2021;10(1):45. [CrossRef]
- Chu D, Liu F. Pathological Changes of Tau Related to Alzheimer’s Disease. ACS Chem Neurosci. 2019;10(2):931-944. [CrossRef]
- Zhang H, Cao Y, Ma L, Wei Y, Li H. Possible Mechanisms of Tau Spread and Toxicity in Alzheimer’s Disease. Front Cell Dev Biol. 2021;9:707268. https://doi.org/10.3389/fcell.2021.707268. [CrossRef]
- Front Cell Neurosci, Fleeman RM, Proctor EA. Astrocytic Propagation of Tau in the Context of Alzheimer’s Disease. Front Cell Neurosci. 2021;15:645233. [CrossRef]
- Silva MVF, Loures C de MG, Alves LCV, de Souza LC, Borges KBG, Carvalho M das G. Alzheimer’s disease: risk factors and potentially protective measures. J Biomed Sci. 2019;26(1):33. [CrossRef]
- Rawat P, Sehar U, Bisht J, Selman A, Culberson J, Reddy PH. Phosphorylated Tau in Alzheimer’s Disease and Other Tauopathies. Int J Mol Sci. 2022;23(21):12841. [CrossRef]
- DeVos SL, Corjuc BT, Oakley DH, et al. Synaptic Tau Seeding Precedes Tau Pathology in Human Alzheimer’s Disease Brain. Front Neurosci. 2018;12:267. [CrossRef]
- Riscado M, Baptista B, Sousa F. New RNA-Based Breakthroughs in Alzheimer’s Disease Diagnosis and Therapeutics. Published online 2021:27.
- Plascencia-Villa G, Perry G. Neuropathologic Changes Provide Insights into Key Mechanisms of Alzheimer Disease and Related Dementia. Am J Pathol. 2022;192(10):1340-1346. https://doi.org/10.1016/j.ajpath.2022.07.002. [CrossRef]
- Marucci G, Buccioni M, Ben DD, Lambertucci C, Volpini R, Amenta F. Efficacy of acetylcholinesterase inhibitors in Alzheimer’s disease. Neuropharmacology. 2021;190:108352. https://doi.org/10.1016/j.neuropharm.2020.108352. [CrossRef]
- Vecchio I, Sorrentino L, Paoletti A, Marra R, Arbitrio M. The State of The Art on Acetylcholinesterase Inhibitors in the Treatment of Alzheimer’s Disease. J Cent Nerv Syst Dis. 2021;13:117957352110291. https://doi.org/10.1177/11795735211029113. [CrossRef]
- Bennett CF, Kordasiewicz HB, Cleveland DW. Antisense Drugs Make Sense for Neurological Diseases. Annu Rev Pharmacol Toxicol. 2021;61(1):831-852. [CrossRef]
- Angelucci F, Cechova K, Valis M, Kuca K, Zhang B, Hort J. MicroRNAs in Alzheimer’s Disease: Diagnostic Markers or Therapeutic Agents? Front Pharmacol. 2019;10:665. https://doi.org/10.3389/fphar.2019.00665. [CrossRef]
- Noori T, Dehpour AR, Sureda A, Sobarzo-Sanchez E, Shirooie S. Role of natural products for the treatment of Alzheimer’s disease. Eur J Pharmacol. 2021;898:173974. [CrossRef]
- Lee CY, Ryu IS, Ryu JH, Cho HJ. miRNAs as Therapeutic Tools in Alzheimer’s Disease. Int J Mol Sci. 2021;22(23):13012. [CrossRef]
- Ramalho MJ, Andrade S, Loureiro JA, do Carmo Pereira M. Nanotechnology to improve the Alzheimer’s disease therapy with natural compounds. Drug Deliv Transl Res. 2020;10(2):380-402. [CrossRef]
- Alhazmi HA, Albratty M. An update on the novel and approved drugs for Alzheimer disease. Saudi Pharm J. 2022;30(12):1755-1764. [CrossRef]
- Chen X, Drew J, Berney W, Lei W. Neuroprotective Natural Products for Alzheimer’s Disease. Cells. 2021;10(6):1309. [CrossRef]
- Said MM, Rabo MMA. Neuroprotective effects of eugenol against aluminiuminduced toxicity in the rat brain. Arch Ind Hyg Toxicol. 2017;68(1):27-37. [CrossRef]
- Casares N, Alfaro M, Cuadrado-Tejedor M, et al. Improvement of cognitive function in wild-type and Alzheimer´s disease mouse models by the immunomodulatory properties of menthol inhalation or by depletion of T regulatory cells. Front Immunol. 2023;14:1130044. [CrossRef]
- Campos HM, da Costa M, da Silva Moreira LK, et al. Protective effects of chrysin against the neurotoxicity induced by aluminium: In vitro and in vivo studies. Toxicology. 2022;465:153033. [CrossRef]
- Hase T, Shishido S, Yamamoto S, et al. Rosmarinic acid suppresses Alzheimer’s disease development by reducing amyloid β aggregation by increasing monoamine secretion. Sci Rep. 2019;9(1):8711. [CrossRef]
- Guan X, Xu J, Liu J, Wu J, Chen L. Ginkgo biloba preparation prevents and treats senile dementia by inhibiting neuro-inflammatory responses. Trop J Pharm Res. 2019;17(10):1961. [CrossRef]
- Islam F, Nafady MH, Islam MdR, et al. Resveratrol and neuroprotection: an insight into prospective therapeutic approaches against Alzheimer’s disease from bench to bedside. Mol Neurobiol. 2022;59(7):4384-4404. [CrossRef]
- 57. Villegas C, Perez R, Petiz LL, Glaser T, Ulrich H, Paz C. Ginkgolides and Huperzine A for complementary treatment of Alzheimer’s disease. IUBMB Life. 2022;74(8):763-779. [CrossRef]
- Dubey T, Chinnathambi S. Brahmi (Bacopa monnieri): An ayurvedic herb against the Alzheimer’s disease. Arch Biochem Biophys. 2019;676:108153. [CrossRef]
- Snow AD, Castillo GM, Nguyen BP, et al. The Amazon rain forest plant Uncaria tomentosa (cat’s claw) and its specific proanthocyanidin constituents are potent inhibitors and reducers of both brain plaques and tangles. Sci Rep. 2019;9(1):561. [CrossRef]
- Huang M, Jiang X, Liang Y, Liu Q, Chen S, Guo Y. Berberine improves cognitive impairment by promoting autophagic clearance and inhibiting production of β-amyloid in APP/tau/PS1 mouse model of Alzheimer’s disease. Exp Gerontol. 2017;91:25-33. [CrossRef]
- Mani RJ, Mittal K, Katare DP. Protective Effects of Quercetin in Zebrafish Model of Alzheimer’s Disease.
- Leiteritz A, Dilberger B, Wenzel U, Fitzenberger E. Betaine reduces β-amyloid-induced paralysis through activation of cystathionine-β-synthase in an Alzheimer model of Caenorhabditis elegans. Genes Nutr. 2018;13(1):21. [CrossRef]
- Finley JW, Gao S. A Perspective on Crocus sativus L. (Saffron) Constituent Crocin: A Potent Water-Soluble Antioxidant and Potential Therapy for Alzheimer’s Disease. J Agric Food Chem. 2017;65(5):1005-1020. [CrossRef]
- Singh M, Ramassamy C. In vitro screening of neuroprotective activity of Indian medicinal plant Withania somnifera. J Nutr Sci. 2017;6:e54. [CrossRef]
- Kim JK, Bae H, Kim MJ, et al. Inhibitory Effect of Poncirus trifoliate on Acetylcholinesterase and Attenuating Activity against Trimethyltin-Induced Learning and Memory Impairment. Biosci Biotechnol Biochem. 2009;73(5):1105-1112. [CrossRef]
- Bihaqi SW, Sharma M, Singh AP, Tiwari M. Neuroprotective role of Convolvulus pluricaulis on aluminium induced neurotoxicity in rat brain. J Ethnopharmacol. 2009;124(3):409-415. [CrossRef]
- Azimi A, Ghaffari SM, Riazi GH, Arab SS, Tavakol MM, Pooyan S. α-Cyperone of Cyperus rotundus is an effective candidate for reduction of inflammation by destabilization of microtubule fibers in brain. J Ethnopharmacol. 2016;194:219-227. [CrossRef]
- Rivera DS, Lindsay C, Codocedo JF, et al. Andrographolide recovers cognitive impairment in a natural model of Alzheimer’s disease (Octodon degus). Neurobiol Aging. 2016;46:204-220. [CrossRef]
- Balez R, Steiner N, Engel M, et al. Neuroprotective effects of apigenin against inflammation, neuronal excitability and apoptosis in an induced pluripotent stem cell model of Alzheimer’s disease. Sci Rep. 2016;6(1):31450. [CrossRef]
- Shi J, Li Y, Zhang Y, et al. Baicalein Ameliorates Aβ-Induced Memory Deficits and Neuronal Atrophy via Inhibition of PDE2 and PDE4. Front Pharmacol. 2021;12:794458. [CrossRef]
- Celik Topkara K, Kilinc E, Cetinkaya A, Saylan A, Demir S. Therapeutic effects of carvacrol on beta-amyloid-induced impairments in in vitro and in vivo models of Alzheimer’s disease. Eur J Neurosci. 2022;56(9):5714-5726. [CrossRef]
- Li L, Li W, Jung SW, Lee YW, Kim YH. Protective Effects of Decursin and Decursinol Angelate against Amyloid β-Protein-Induced Oxidative Stress in the PC12 Cell Line: The Role of Nrf2 and Antioxidant Enzymes. Biosci Biotechnol Biochem. 2011;75(3):434-442. [CrossRef]
- Duan X, Li Y, Xu F, Ding H. Study on the neuroprotective effects of Genistein on Alzheimer’s disease. Brain Behav. 2021;11(5). https://doi.org/10.1002/brb3.2100. [CrossRef]
- Huang DS, Yu YC, Wu CH, Lin JY. Protective Effects of Wogonin against Alzheimer’s Disease by Inhibition of Amyloidogenic Pathway. Evid Based Complement Alternat Med. 2017;2017:1-13. [CrossRef]
- Xu P xin, Wang S wei, Yu X lin, et al. Rutin improves spatial memory in Alzheimer’s disease transgenic mice by reducing Aβ oligomer level and attenuating oxidative stress and neuroinflammation. Behav Brain Res. 2014;264:173-180. [CrossRef]
- Daily JW, Kang S, Park S. Protection against Alzheimer’s disease by luteolin: Role of brain glucose regulation, anti-inflammatory activity, and the gut microbiota-liver-brain axis. BioFactors. 2021;47(2):218-231. https://doi.org/10.1002/biof.1703. [CrossRef]
- Sabogal-Guáqueta AM, Osorio E, Cardona-Gómez GP. Linalool reverses neuropathological and behavioral impairments in old triple transgenic Alzheimer’s mice. Neuropharmacology. 2016;102:111-120. [CrossRef]
- Asiatic acid nullified aluminium toxicity in in vitro model of Alzheimer’s disease. Accessed May 30, 2023. https://www.imrpress.com/journal/FBE/10/2/10.2741/E823.
- Li Z, Zhao T, Li J, et al. Nanomedicine Based on Natural Products: Improving Clinical Application Potential. Wang R, ed. J Nanomater. 2022;2022:1-11. [CrossRef]
- Woon CK, Hui WK, Abas R, Haron MH, Das S, Lin TS. Natural Product-based Nanomedicine: Recent Advances and Issues for theTreatment of Alzheimer’s Disease. Curr Neuropharmacol. 2022;20(8):1498-1518. [CrossRef]
- Yu AM, Jian C, Yu AH, Tu MJ. RNA therapy: Are we using the right molecules? Pharmacol Ther. 2019;196:91-104. [CrossRef]
- Shin H, Park SJ, Yim Y, et al. Recent Advances in RNA Therapeutics and RNA Delivery Systems Based on Nanoparticles. Adv Ther. 2018;1(7):1800065. [CrossRef]
- Zogg H, Singh R, Ro S. Current Advances in RNA Therapeutics for Human Diseases. Int J Mol Sci. 2022;23(5):2736. [CrossRef]
- Mollocana-Lara EC, Ni M, Agathos SN, Gonzales-Zubiate FA. The infinite possibilities of RNA therapeutics. J Ind Microbiol Biotechnol. 2021;48(9-10):kuab063. [CrossRef]
- DeLong R. Ushering in a new era of RNA-based therapies. Commun Biol. 2021;4(1):577, s42003-021-02150-w. [CrossRef]
- Kim YK. RNA therapy: rich history, various applications and unlimited future prospects. Exp Mol Med. Published online April 19, 2022. [CrossRef]
- Kim YK. RNA Therapy: Current Status and Future Potential. Chonnam Med J. 2020;56(2):87. [CrossRef]
- Lee MJ, Lee I, Wang K. Recent Advances in RNA Therapy and Its Carriers to Treat the Single-Gene Neurological Disorders. Biomedicines. 2022;10(1):158. [CrossRef]
- Anthony K. RNA-based therapeutics for neurological diseases. RNA Biol. 2022;19(1):176-190. [CrossRef]
- Jurcău MC, Andronie-Cioara FL, Jurcău A, et al. The Link between Oxidative Stress, Mitochondrial Dysfunction and Neuroinflammation in the Pathophysiology of Alzheimer’s Disease: Therapeutic Implications and Future Perspectives. Antioxidants. 2022;11(11):2167. [CrossRef]
- Walgrave H, Zhou L, De Strooper B, Salta E. The promise of microRNA-based therapies in Alzheimer’s disease: challenges and perspectives. Mol Neurodegener. 2021;16(1):76. [CrossRef]
- Kreth S, Hübner M, Hinske LC. MicroRNAs as Clinical Biomarkers and Therapeutic Tools in Perioperative Medicine: Anesth Analg. 2018;126(2):670-681. h. [CrossRef]
- Martier R, Konstantinova P. Gene Therapy for Neurodegenerative Diseases: Slowing Down the Ticking Clock. Front Neurosci. 2020;14:580179. [CrossRef]
- Zhou Y, Zhu F, Liu Y, et al. Blood-brain barrier–penetrating siRNA nanomedicine for Alzheimer’s disease therapy. Sci Adv. 2020;6(41):eabc7031. [CrossRef]
- Long JM, Lahiri DK. MicroRNA-101 downregulates Alzheimer’s amyloid-β precursor protein levels in human cell cultures and is differentially expressed. Biochem Biophys Res Commun. 2011;404(4):889-895. [CrossRef]
- Liu W, Zhao J, Lu G. miR-106b inhibits tau phosphorylation at Tyr18 by targeting Fyn in a model of Alzheimer’s disease. Biochem Biophys Res Commun. 2016;478(2):852-857. [CrossRef]
- Jiang Y, Xu B, Chen J, et al. Micro-RNA-137 Inhibits Tau Hyperphosphorylation in Alzheimer’s Disease and Targets the CACNA1C Gene in Transgenic Mice and Human Neuroblastoma SH-SY5Y Cells. Med Sci Monit. 2018;24:5635-5644. [CrossRef]
- Santa-Maria I, Alaniz ME, Renwick N, et al. Dysregulation of microRNA-219 promotes neurodegeneration through post-transcriptional regulation of tau. J Clin Invest. 2015;125(2):681-686. [CrossRef]
- Estfanous S, Daily KP, Eltobgy M, et al. Elevated Expression of MiR-17 in Microglia of Alzheimer’s Disease Patients Abrogates Autophagy-Mediated Amyloid-β Degradation. Front Immunol. 2021;12:705581. [CrossRef]
- Wang R, Chopra N, Nho K, et al. Human microRNA (miR-20b-5p) modulates Alzheimer’s disease pathways and neuronal function, and a specific polymorphism close to the MIR20B gene influences Alzheimer’s biomarkers. Mol Psychiatry. 2022;27(2):1256-1273. [CrossRef]
- Zong Y, Wang H, Dong W, et al. miR-29c regulates BACE1 protein expression. Brain Res. 2011;1395:108-115. [CrossRef]
- Chopra N, Wang R, Maloney B, et al. MicroRNA-298 reduces levels of human amyloid-β precursor protein (APP), β-site APP-converting enzyme 1 (BACE1) and specific tau protein moieties. Mol Psychiatry. 2021;26(10):5636-5657. [CrossRef]
- He C, Su C, Zhang W, Wan Q. miR-485-5p alleviates Alzheimer’s disease progression by targeting PACS1. Transl Neurosci. 2021;12(1):335-345. [CrossRef]
- Liu J, Zuo X, Han J, et al. MiR-9-5p inhibits mitochondrial damage and oxidative stress in AD cell models by targeting GSK-3β. Biosci Biotechnol Biochem. 2020;84(11):2273-2280. [CrossRef]
- Deng Y, Zhang J, Sun X, et al. miR-132 improves the cognitive function of rats with Alzheimer’s disease by inhibiting the MAPK1 signal pathway. Exp Ther Med. 2020;20(6):159. [CrossRef]
- 106. Liang C, Zhu H, Xu Y, et al. MicroRNA-153 negatively regulates the expression of amyloid precursor protein and amyloid precursor-like protein 2. Brain Res. 2012;1455:103-113. [CrossRef]
- Xu H, Rosler T, Carlsson T, et al. Tau Silencing by siRNA in the P301S Mouse Model of Tauopathy. Curr Gene Ther. 2014;14(5):343-351. [CrossRef]
- Kandimalla RJ, Wani WY, Bk B, Gill KD. siRNA against presenilin 1 (PS1) down regulates amyloid b42 production in IMR-32 cells. Published online 2012.
- Senechal Y, Kelly PH, Cryan JF, Natt F, Dev KK. Amyloid precursor protein knockdown by siRNA impairs spontaneous alternation in adult mice: In vivo knockdown of APP by RNAi. J Neurochem. 2007;102(6):1928-1940. https://doi.org/10.1111/j.1471-4159.2007.04672.x. [CrossRef]
- Lin CY, Perche F, Ikegami M, Uchida S, Kataoka K, Itaka K. Messenger RNA-based therapeutics for brain diseases: An animal study for augmenting clearance of beta-amyloid by intracerebral administration of neprilysin mRNA loaded in polyplex nanomicelles. J Controlled Release. 2016;235:268-275. [CrossRef]
- 111. Lim SA, Cox A, Tung M, Chung EJ. Clinical progress of nanomedicine-based RNA therapies. Bioact Mater. 2022;12:203-213. [CrossRef]
- Gorshkov A, Purvinsh L, Brodskaia A, Vasin A. Exosomes as Natural Nanocarriers for RNA-Based Therapy and Prophylaxis. Nanomaterials. 2022;12(3):524. [CrossRef]
- Tsakiri M, Zivko C, Demetzos C, Mahairaki V. Lipid-based nanoparticles and RNA as innovative neuro-therapeutics. Front Pharmacol. 2022;13:900610. [CrossRef]
- Fernandes F, Dias-Teixeira M, Delerue-Matos C, Grosso C. Critical Review of Lipid-Based Nanoparticles as Carriers of Neuroprotective Drugs and Extracts. Nanomaterials. 2021;11(3):563. [CrossRef]
- Pardridge WM. Treatment of Alzheimer’s Disease and Blood–Brain Barrier Drug Delivery. Pharmaceuticals. 2020;13(11):394. [CrossRef]
- Wohlfart S, Gelperina S, Kreuter J. Transport of drugs across the blood–brain barrier by nanoparticles. J Controlled Release. 2012;161(2):264-273. [CrossRef]
- Zhou Y, Peng Z, Seven ES, Leblanc RM. Crossing the blood-brain barrier with nanoparticles. J Controlled Release. 2018;270:290-303. https://doi.org/10.1016/j.jconrel.2017.12.015. [CrossRef]
- Bors L, Erdő F. Overcoming the Blood–Brain Barrier. Challenges and Tricks for CNS Drug Delivery. Sci Pharm. 2019;87(1):6. [CrossRef]
- Wong K, Riaz M, Xie Y, et al. Review of Current Strategies for Delivering Alzheimer’s Disease Drugs across the Blood-Brain Barrier. Int J Mol Sci. 2019;20(2):381. [CrossRef]
- Zenaro E, Piacentino G, Constantin G. The blood-brain barrier in Alzheimer’s disease. Neurobiol Dis. 2017;107:41-56. [CrossRef]
- Sharma C, Woo H, Kim SR. Addressing Blood–Brain Barrier Impairment in Alzheimer’s Disease. Biomedicines. 2022;10(4):742. [CrossRef]
- Juhairiyah F, de Lange ECM. Understanding Drug Delivery to the Brain Using Liposome-Based Strategies: Studies that Provide Mechanistic Insights Are Essential. AAPS J. 2021;23(6):114. [CrossRef]
- Satapathy MK, Yen TL, Jan JS, et al. Solid Lipid Nanoparticles (SLNs): An Advanced Drug Delivery System Targeting Brain through BBB. Pharmaceutics. 2021;13(8):1183. [CrossRef]
- Musielak E, Feliczak-Guzik A, Nowak I. Synthesis and Potential Applications of Lipid Nanoparticles in Medicine. Materials. 2022;15(2):682. [CrossRef]
- Heidarzadeh M, Gürsoy-Özdemir Y, Kaya M, et al. Exosomal delivery of therapeutic modulators through the blood–brain barrier; promise and pitfalls. Cell Biosci. 2021;11(1):142. [CrossRef]
- Negahdaripour M, Vakili B, Nezafat N. Exosome-based vaccines and their position in next generation vaccines. Int Immunopharmacol. 2022;113:109265. [CrossRef]
- Jafari D, Malih S, Eini M, et al. Improvement, scaling-up, and downstream analysis of exosome production. Crit Rev Biotechnol. 2020;40(8):1098-1112. [CrossRef]
- Hu Q, Su H, Li J, et al. Clinical applications of exosome membrane proteins. Precis Clin Med. 2020;3(1):54-66. https://doi.org/10.1093/pcmedi/pbaa007. [CrossRef]
- Ludwig N, Whiteside TL, Reichert TE. Challenges in Exosome Isolation and Analysis in Health and Disease. Int J Mol Sci. 2019;20(19):4684. [CrossRef]
- H. Rashed M, Bayraktar E, K. Helal G, et al. Exosomes: From Garbage Bins to Promising Therapeutic Targets. Int J Mol Sci. 2017;18(3):538. [CrossRef]
- Zhang N, He F, Li T, et al. Role of Exosomes in Brain Diseases. Front Cell Neurosci. 2021;15:743353. [CrossRef]
- Xu L, Wang X, Liu Y, Yang G, Falconer RJ, Zhao CX. Lipid Nanoparticles for Drug Delivery. Adv NanoBiomed Res. 2022;2(2):2100109. [CrossRef]
- Feng R, Patil S, Zhao X, Miao Z, Qian A. RNA Therapeutics - Research and Clinical Advancements. Front Mol Biosci. 2021;8:710738. [CrossRef]
- Burdușel AC, Andronescu E. Lipid Nanoparticles and Liposomes for Bone Diseases Treatment. Biomedicines. 2022;10(12):3158. [CrossRef]
- Duan Y, Dhar A, Patel C, et al. A brief review on solid lipid nanoparticles: part and parcel of contemporary drug delivery systems. RSC Adv. 2020;10(45):26777-26791. [CrossRef]
- Tenchov R, Bird R, Curtze AE, Zhou Q. Lipid Nanoparticles─From Liposomes to mRNA Vaccine Delivery, a Landscape of Research Diversity and Advancement. ACS Nano. 2021;15(11):16982-17015. [CrossRef]
- Abdellatif AAH, Younis MA, Alsowinea AF, et al. Lipid nanoparticles technology in vaccines: Shaping the future of prophylactic medicine. Colloids Surf B Biointerfaces. 2023;222:113111. [CrossRef]
- Vieira D, Gamarra L. Getting into the brain: liposome-based strategies for effective drug delivery across the blood–brain barrier. Int J Nanomedicine. 2016;Volume 11:5381-5414. [CrossRef]
- Gauro R, Nandave M, Jain VK, Jain K. Advances in dendrimer-mediated targeted drug delivery to the brain. J Nanoparticle Res. 2021;23(3):76. [CrossRef]
- Teleanu D, Chircov C, Grumezescu A, Volceanov A, Teleanu R. Blood-Brain Delivery Methods Using Nanotechnology. Pharmaceutics. 2018;10(4):269. [CrossRef]
- Sela H, Cohen H, Elia P, Zach R, Karpas Z, Zeiri Y. Spontaneous penetration of gold nanoparticles through the blood brain barrier (BBB). J Nanobiotechnology. 2015;13(1):71. [CrossRef]
- Li X, Corbett AL, Taatizadeh E, et al. Challenges and opportunities in exosome research—Perspectives from biology, engineering, and cancer therapy. APL Bioeng. 2019;3(1):011503. [CrossRef]
- Schiffelers R, Kooijmans S, Vader, van Dommelen, Van Solinge. Exosome mimetics: a novel class of drug delivery systems. Int J Nanomedicine. Published online March 2012:1525. [CrossRef]
- Fernandes M, Lopes I, Magalhães L, et al. Novel concept of exosome-like liposomes for the treatment of Alzheimer’s disease. J Controlled Release. 2021;336:130-143. [CrossRef]
Figure 1.
Hallmarks of neurodegenerative diseases. The scheme illustrates the six hallmarks of these disorders: protein aggregation, disruptive proteostasis, neuroinflammation, oxidative stress, synaptic failure, and neuronal death.
Figure 1.
Hallmarks of neurodegenerative diseases. The scheme illustrates the six hallmarks of these disorders: protein aggregation, disruptive proteostasis, neuroinflammation, oxidative stress, synaptic failure, and neuronal death.
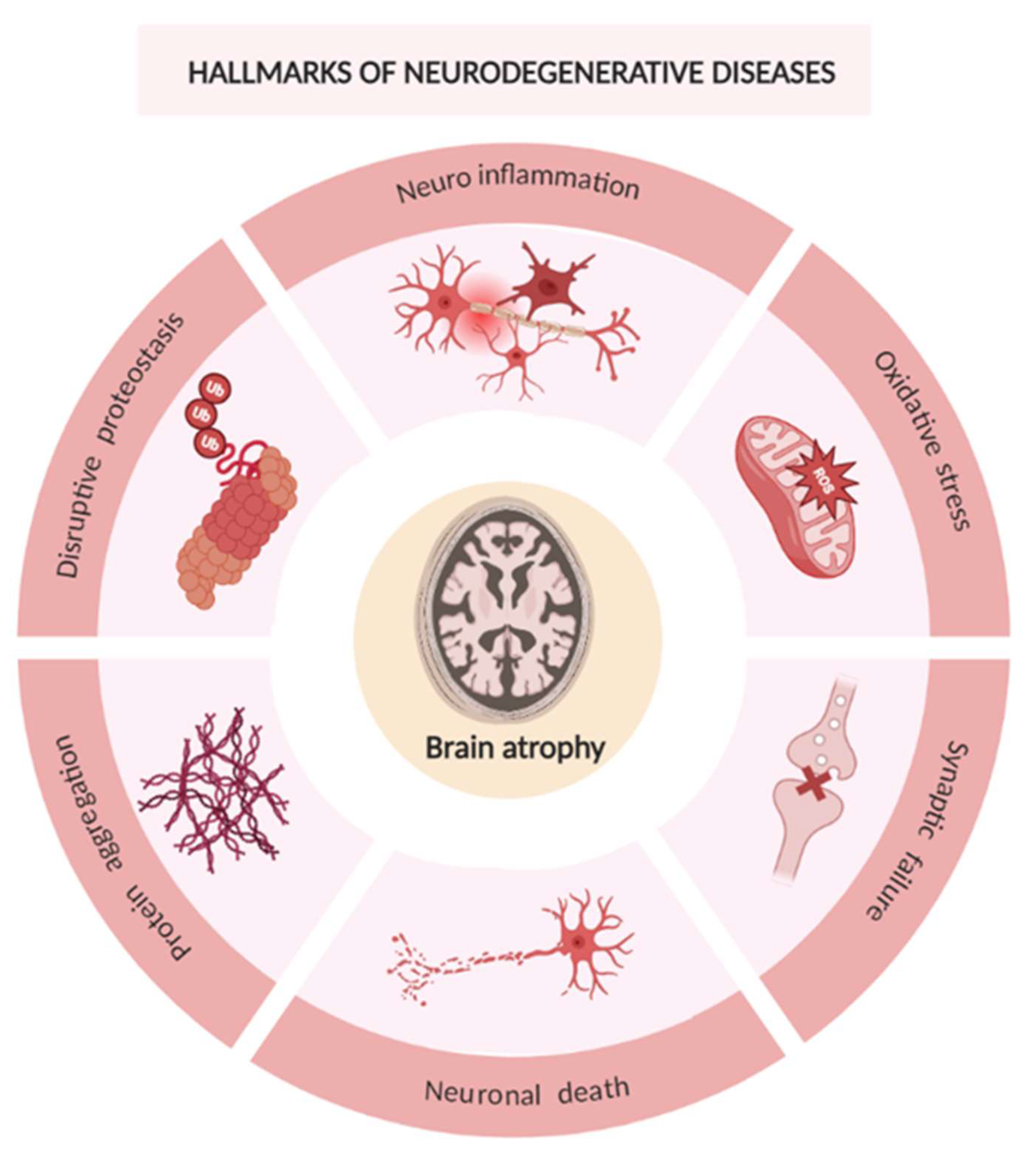
Figure 2.
Alzheimer’s disease hallmarks. (A) pathway that leads to the formation of amyloid plaques and the mechanism responsible for the formation of neurofibrillary tangles. (B) Accumulation of amyloid plaques and neurofibrillary tangles provoke disorders such as synaptic dysfunction, neuroinflammation, oxidative stress, and higher activity of acetylcholinesterase. These result in neurodegeneration.
Figure 2.
Alzheimer’s disease hallmarks. (A) pathway that leads to the formation of amyloid plaques and the mechanism responsible for the formation of neurofibrillary tangles. (B) Accumulation of amyloid plaques and neurofibrillary tangles provoke disorders such as synaptic dysfunction, neuroinflammation, oxidative stress, and higher activity of acetylcholinesterase. These result in neurodegeneration.
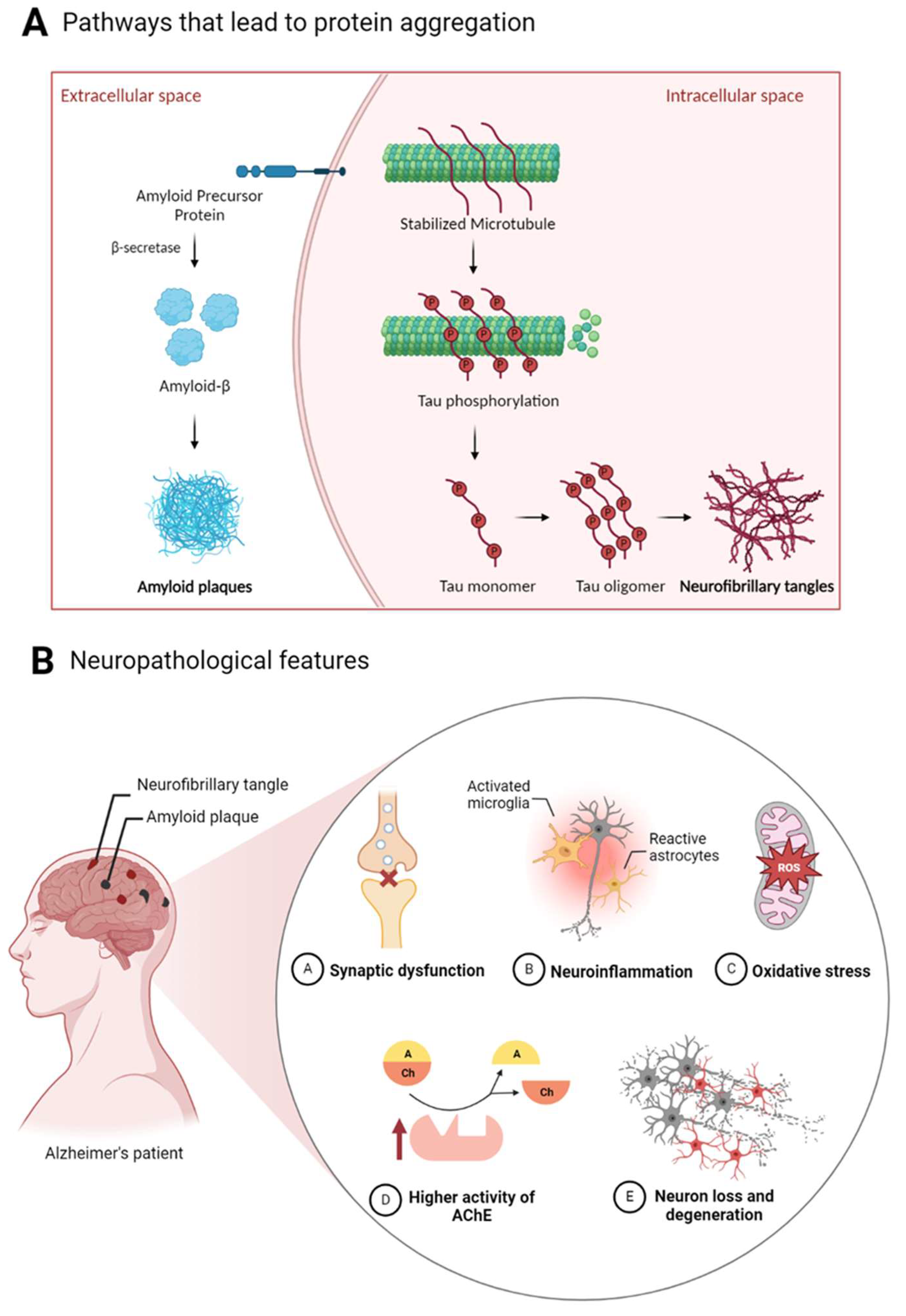
Figure 3.
Potential problems of natural products in vivo and advantages of natural products delivered by nanoparticles.
Figure 3.
Potential problems of natural products in vivo and advantages of natural products delivered by nanoparticles.
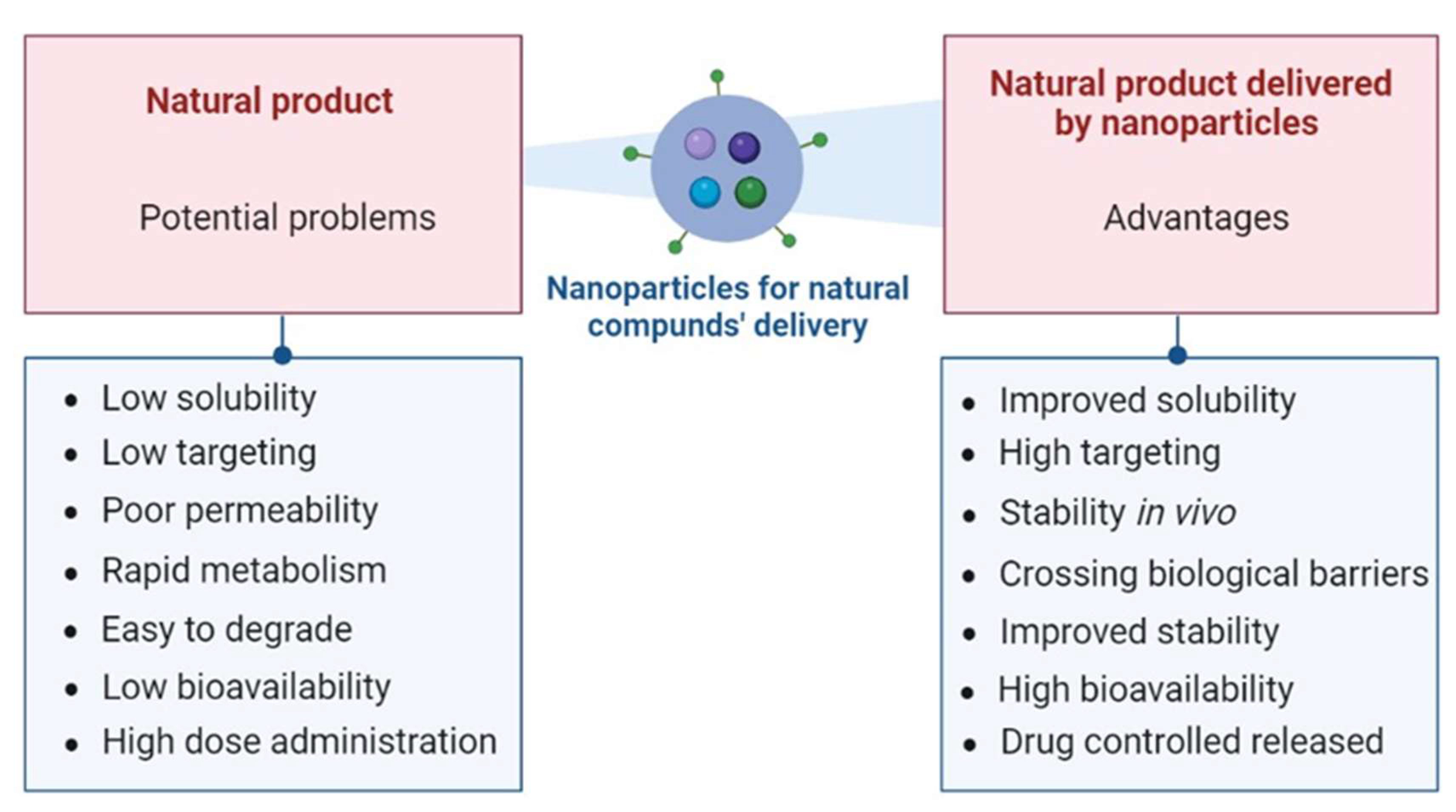
Figure 4.
Possible RNA-based therapeutic strategies for AD. (A) mRNA leads to protein expression and (B) siRNA, miRNA and antisense oligonucleotide (ASO) inhibit protein expression.
Figure 4.
Possible RNA-based therapeutic strategies for AD. (A) mRNA leads to protein expression and (B) siRNA, miRNA and antisense oligonucleotide (ASO) inhibit protein expression.
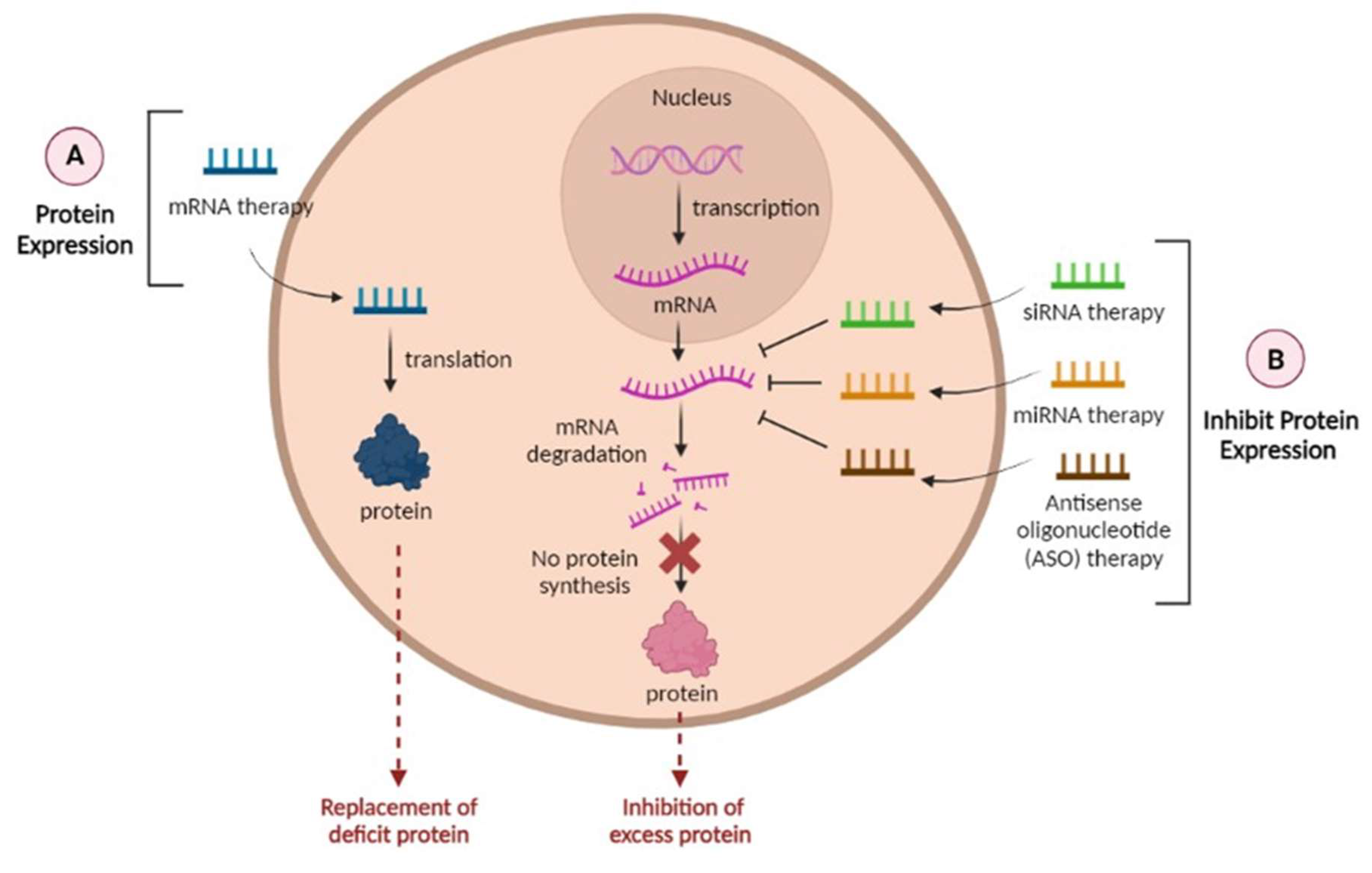
Figure 5.
How RNA delivery systems can overcome the physiological obstacles faced by naked RNAs. (a) RNA can be delivered across the BBB; (b) protection of RNAs from degradation by ribonucleases (RNAses); (c) enhanced cellular internalization by controlled or specific pathways; (d) intracellular endosomal degradation can be avoided by endosomal escape; (e) improved RNA-mediated bioactivities; (f) enhanced accumulation of RNA in target tissue.
Figure 5.
How RNA delivery systems can overcome the physiological obstacles faced by naked RNAs. (a) RNA can be delivered across the BBB; (b) protection of RNAs from degradation by ribonucleases (RNAses); (c) enhanced cellular internalization by controlled or specific pathways; (d) intracellular endosomal degradation can be avoided by endosomal escape; (e) improved RNA-mediated bioactivities; (f) enhanced accumulation of RNA in target tissue.
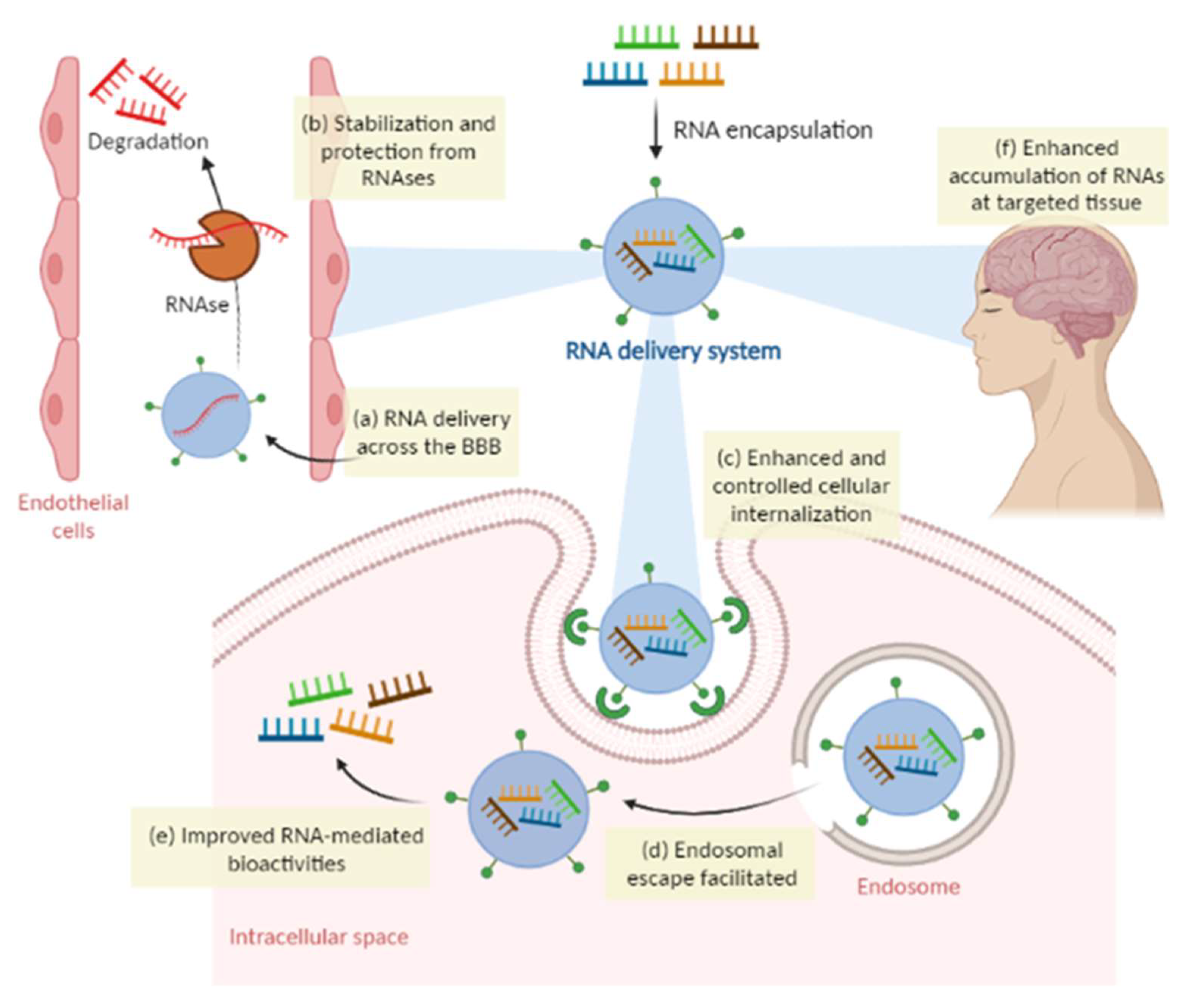
Figure 6.
(A) Structural representation of the BBB anatomy and possible pathways of entrance into the central nervous system (B).
Figure 6.
(A) Structural representation of the BBB anatomy and possible pathways of entrance into the central nervous system (B).
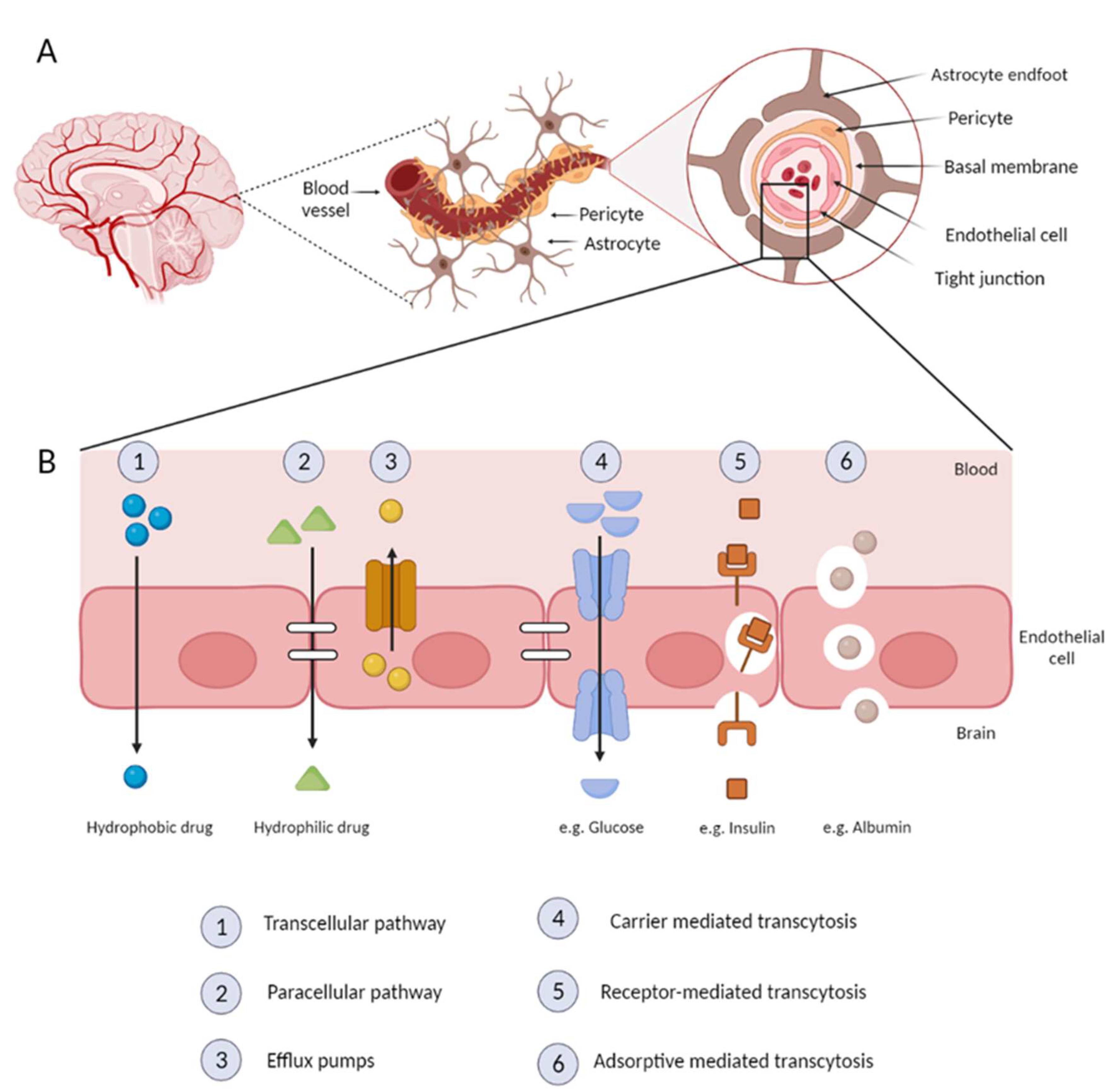
Figure 7.
Exosome-like liposomes combine advantages of liposomes and exosomes, having the capacity to be loaded with different types of molecules due to their constitution.
Figure 7.
Exosome-like liposomes combine advantages of liposomes and exosomes, having the capacity to be loaded with different types of molecules due to their constitution.
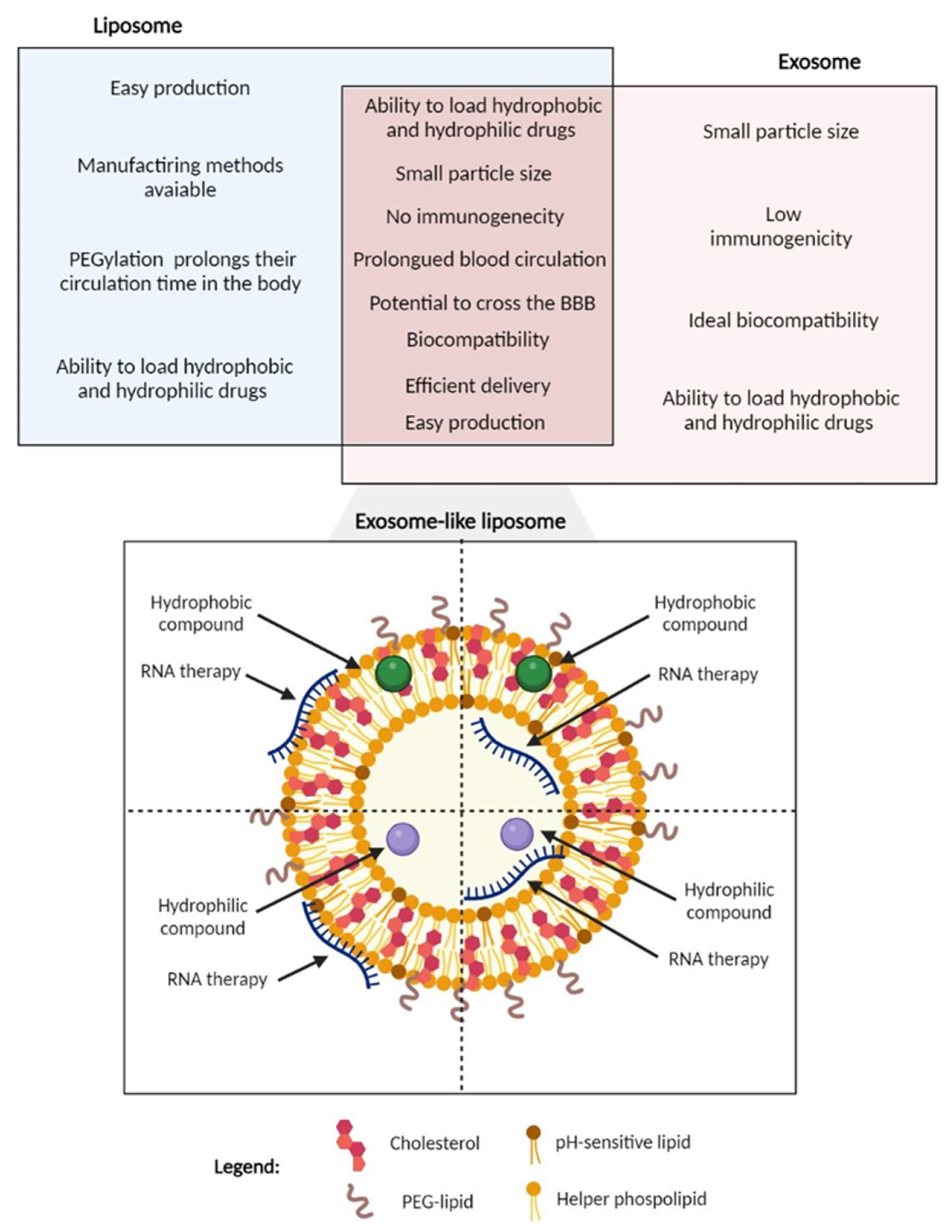
Table 1.
Natural compounds or extracts that can be used for the AD therapy.
| Natural Compound | Role in AD | Reference |
|---|---|---|
| Eugenol | Rats were fed aluminium, a neurotoxic metal that leads to oxidative brain injury and enhanced lipid peroxidation, disruption of neurotrophic, cholinergic, and serotonergic functions, and induce apoptosis with ultimate neuronal and astrocyte damages. A neuroprotective role of eugenol against the aluminium effects, was verified through its antioxidant, antiapoptotic potential and its neurotrophic properties. | [51] |
| Menthol | Menthol inhalation by mice (1 week per month, for 6 months) prevented cognitive impairment in the APP/PS1 mouse model of Alzheimer’s. | [52] |
| Chrysin | Chrysin showed the ability to act as a membrane shield against early oxidative events mediated by O2˙- and other ROS that contribute to neuronal death triggered by AlCl3 exposure, showing chrysin neuroprotective action. | [53] |
| Rosmarinic acid | Suppresses amyloid β accumulation in mice. | [54] |
| Ginkgo biloba | Ginkgo biloba improves microcirculation, inhibits the expression of inflammatory factors, and reduces inflammatory damage to neurons, thereby improving the spatial exploration memory of dementia model rats. | [55] |
| Resveratrol | Multiple studies demonstrated that resveratrol has a neuroprotective, anti-inflammatory, and antioxidant characteristics and the ability to minimize Aβ peptide aggregation and toxicity in the hippocampus of Alzheimer’s patients, stimulating neurogenesis and inhibiting hippocampal degeneration. Furthermore, resveratrol’s antioxidant effect promotes neuronal development by activating the silent information regulator-1 (SIRT1), which can protect against the detrimental effects of oxidative stress. | [56] |
| Huperzine A | Huperzine A is natural, potent, highly specific reversible inhibitor of acetylcholinesterase, with the ability to cross the BBB. | [57] |
| Brahmi | The neuroprotective properties of Brahmi include the reduction of ROS and neuroinflammation, the inhibition of the aggregation of Aβ and the improvement of cognitive and learning behaviour. | [58] |
| Uncaria tomentosa | Inhibits plaques and tangles formation. | [59] |
| Berberine | Berberine has antioxidant activity and promotes AChE and monoamine oxidase inhibition. Berberine has been shown to improve memory, lower Aβ and APP concentration, and diminish Aβ plaque accumulation. | [60] |
| Quercetin | Behavioural and biochemical tests confirm that quercetin promotes the reduction in oxidative stress and increased cognition in zebrafish AD models induced with aluminium chloride. | [61] |
| Betaine | Betaine has shown to decrease homocysteine levels and Aβ toxicity in Caenorhabditis elegans AD model. | [62] |
| Curcumin | Curcumin is known to be a potent antioxidant, anti-inflammatory and anti-amyloidogenic compound, that plays a beneficial role in treating AD through several mechanisms. Curcumin can promote a significant reduction Aβ oligomers and fibril formation. | [46] |
| Crocin | Crocin, the main constituent of Crocus sativus L., has a multifunctional role in protecting brain cells, modulating aggregation of Aβ and tau proteins, attenuating cognitive and memory impairments, and improving oxidative stress. | [63] |
| Withania somnifera | Withania somnifera (WS) extract can protect against Aβ peptide- and acrolein-induced toxicity. Treatment with WS extract significantly protected against Aβ peptide and acrolein, in various cell survival assays with the human neuroblastoma cell line SK-N-SH , significantly reduced the generation of reactive oxygen species and demonstrated to be a potent inhibitor of acetylcholinesterase activity. | [64] |
| Poncirus trifoliate | The extract of Poncirus trifoliate is a naturally occurring AChE inhibitor. It showed a 47.31% inhibitory effect on the activity of acetylcholine. | [65] |
| Convolvulus pluricaulis | Convolvulus pluricaulis prevented aluminium induced neurotoxicity in rat cerebral cortex. | [66] |
| α-Cyperone | α-Cyperone binds and interacts with tubulin, being capable of destabilizing microtubule polymerization. The effect of this interaction could result in reduction of inflammation. | [67] |
| Andrographolide | Andrographolide has beneficial effects in the recovery of spatial memory and learning performance, recovery of synaptic basal transmission, partial or complete protection of certain synaptic proteins and shows a specific neuroprotective effect, that includes the reduction of phosphorylated tau and amyloid beta aggregate maturation, in aged degus. | [68] |
| Apigenin | Apigenin has been shown to have anti-inflammatory and neuroprotective properties in a number of cell and animal models. This compound is also able to protect human induced pluripotent stem cell-derived AD neurons via multiple pathways, by reducing the frequency of spontaneous Ca2+ signals and significantly reducing caspase-3/7 mediated apoptosis. | [69] |
| Baicalein | Baicalein has antioxidant and anti-inflammatory effects. | [70] |
| Carvacrol | Carvacrol possesses anti-acetylcholinesterase, antioxidant, and neuroprotective properties. This compound alleviated Aβ-induced deficits by reducing cellular neurotoxicity and oxidative stress in the SH-SY5Y cell line, and by reducing oxidative stress and memory impairment in a rat model of AD. | [71] |
| Decursin/ Decursinol angelate | Decursin and decursinol angelate increase cellular resistance to Aβ-induced oxidative injury in PC12 cells. | [72] |
| Genistein | In vivo studies have shown that genistein improves brain function, antagonizes the toxicity of Aβ and has neuroprotective effects. | [73] |
| Wogonin | Wogonin has various neuroprotective and neurotrophic activities, such as inducing neurite outgrowth. | [74] |
| Rutin | Rutin is an antioxidant, anti-inflammatory, and has the capacity of reducing Aβ oligomer activities. | [75] |
| Luteolin | Luteolin has the capacity to cross the BBB and can inhibit β- and γ-secretase to decrease Aβ. It can also reduce neuroinflammation, and attenuate the phosphorylation of tau and the formation of tangles. | [76] |
| Linalool | A linalool-treated mice model of AD showed improved learning and spatial memory. This compound reverses the histopathological hallmarks of AD and restores cognitive and emotional functions via an anti-inflammatory effect. | [77] |
| Asiatic acid | Pre-treatment with Asiatic Acid enhanced cell viability, attenuated rotenone-induced ROS, mitochondrial membrane dysfunction and apoptosis regulating AKT/GSK-3β signalling pathway, after aluminium maltolate neurotoxicity induction in SH-SY5Y neuroblastoma cells. | [78] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



