
Submitted:
07 July 2023
Posted:
10 July 2023
You are already at the latest version
Abstract
Tilapia tilapinevirus, or tilapia lake virus (TiLV), is a virus that has caused disease outbreaks in many countries worldwide since 2009. We recently developed two simple but effective TiLV vaccines based on heat-killed virus (HKV) and formalin-killed virus (FKV). In the present study, we evaluated the immunological response and efficacy of MontanideTM ISA 763A VG oil-based vaccines (O-HKV and O-FKV) in Nile tilapia (Oreochromis niloticus) and compared these responses in fish vaccinated with water-based formulations (W-HKV and W-FKV). Juvenile Nile tilapia were given primary and booster vaccinations with the vaccine formulations. The relative fold change in expression of five immunoglobulin genes (IgM, IgT, IgD, CD4 and CD8) was assessed before and after booster vaccination, and a challenge with TiLV was performed to evaluate the efficacy of the vaccines. The results showed that the water-based vaccines induced higher levels of Ig transcripts and conferred significantly higher levels of protection compared to the oil-based formulations, with relative percent survival (RPS) values ranging from 54.6% to 59.7% for W-HKV and W-FKV vaccinated fish, and 32% and 0% for the O-HKV and O-FKV vaccinated fish, respectively. These findings suggest that water-based vaccines are more effective at protecting tilapia against TiLV infection, and MontanideTM ISA 763A VG may not be a suitable adjuvant for formulating whole-inactivated TiLV vaccines.
Keywords:
tilapia lake virus
; TiLV
; Nile tilapia
; vaccine
; adjuvants
; MontanideTM ISA 763A VG
1. Introduction
Tilapia (Oreochromis spp.) is one of the most common fish species farmed worldwide (over 140 countries) because of their many attributes, including their high protein content, their large size and rapid growth, and their ability to adapt to different geographical locations [1]. The increasing demand for protein from aquaculture has seen an intensification of tilapia farming systems [2]. This has led to increased disease outbreaks caused by bacteria, viruses, parasites, and fungi. Massive die-offs due to disease have resulted in substantial economic losses to tilapia producers and related sectors [3].
Tilapia lake virus (TiLV) or Tilapinevirus tilapiae is an emerging RNA virus affecting both wild and farmed tilapia. The virus has now been reported in 17 countries, with cumulative mortalities due to TiLV infection ranging from 20 to 90% [4]. The initial signs of infection include loss of appetite, erratic swimming, darkening skin, and scale protrusion, followed by more serious symptoms such as abdominal extension, ocular alteration, pale gills, necrosis of internal organs and fin loss [5,6]. The virus has been shown to be transmitted both horizontally and vertically [7,8]. Therefore, initiatives to maintain a TiLV-free status for tilapia broodstock and their progeny are important for controlling TiLV disease in tilapia aquaculture.
Vaccination is one of the effective solutions for fish health management to mitigate against disease outbreaks by triggering a specific immune response against the pathogen [3,9]. Some injectable vaccines have been reported to be efficacious against TiLV infections in tilapia. The first vaccines developed against TiLV were based on attenuated viral strains, which underwent 17 and 20 passages through cell culture, called strains P17 and P20. These vaccines resulted in relative percentage survival (RPS) values of over 50% [10]. Development of DNA and recombinant vaccines based on segment 8, administered with adjuvant M402, resulted in higher survival levels in vaccinated fish compared to unvaccinated fish [11]. Another study by the same research group demonstrated that inactivated whole virus vaccines, combined with adjuvant Montanide IMS 1312VG, were relatively effective in protecting tilapia against TiLV, with RPS values ranging from 32.1% - 85.7%, depending on the virus concentration in the vaccine [12]. Meanwhile, water-based, heat-killed, and formalin-killed vaccines resulted in RPS values of 71.3% and 79.6%, respectively [13]. A subsequent study with these vaccines found that broodstock immunization with water-based inactivated TiLV vaccines produced protective antibodies that were transferred to offspring, indicating that broodstock immunization could potentially prevent vertical transmission of TiLV [14]. Recently, a nano chitosan-based, inactivated whole virus vaccine developed for immersion delivery to small fish, resulted in a relatively high RPS value of 68.17% compared to 25.01% for the group receiving the non-encapsulated inactivated vaccine [15].
Several studies have demonstrated that combining potential antigens with adjuvants can improve co-stimulatory signals, prolong antigen release, and activate lymphocyte proliferation. However, the effectiveness of the resulting vaccines can vary depending on the type of antigen used [16,17,18]. Water in oil (w/o) emulsion is widely used to formulate vaccines for aquatic animals. The antigen is deposited in the oil phase, which can enhance antigen delivery to antigen-presenting cells, prolong specific immune responses, and attract more effective lymphocytes to the injection site, leading to stronger localized immune responses [19,20,21]. Various w/o adjuvanted vaccines have been shown to be effective formulations for fish and are often administered via intraperitoneal (IP) injection. MontanideTM ISA 763A VG is one such adjuvant that has been found to be a non-mineral metabolizable oil-based adjuvant. It has been reported to strongly stimulate humoral immunity and be a highly effective adjuvant for bacterin vaccines in a variety of fish species, such as Atlantic salmon (Salmo salar), rainbow trout (Oncorhynchus mykiss), turbot (Scophthalmus maximus L.), tilapia (O. niloticus), etc. [20,21,22,23]. Recently, MontanideTM ISA 763A VG was used as an adjuvant for a recombinant TiLV vaccine expressed from TiLV segments 9 and 10, showing relatively good levels of protection with an RPS value of 55.56% [24]. Although MontanideTM ISA 763A VG is a potential adjuvant and TiLV water-based inactivated vaccines have demonstrated high efficacy, there are currently no reports of its use in TiLV-inactivated vaccinations. Thus, the objective of this study was to investigate whether this adjuvant can enhance immunological responses and the efficacy of TiLV-inactivated vaccines.
2. Materials and Methods
2.1. Fish
Fingerling Nile tilapia weighing 7.3±1.2 g were kindly provided by Kasetsart University, Thailand. The fish were acclimatized for seven days in 100 liter tanks supplied with fresh water at a density of 45 fish per tank. The water temperature ranged from 25oC to 28oC. Water parameters (pH, NO2, NH3) were checked every two days using standard water quality kits (Sera, Germany), and water was changed twice weekly. Five fish, randomly selected, were confirmed TiLV-free by real-time PCR [25]. Approval for the use of animals in this study was granted by Kasetsart University Institutional Animal Care and Use Committee (ACKU62- FIS- 008).
2.2. Virus culture
TiLV propagation was performed as described in a previous study [14]. Briefly, TiLV-KT strain was propagated on E11 cells in Leibovitz’s L15 medium (Sigma, Saint Louis, MO, USA) containing 5% fetal bovine serum for 5-7 days or until a cytopathic effect (CPE) of around 80%-90% was obtained. The viral supernatant was clarified by centrifugation at 4500 g for 10 min to remove cell debris. The virus was quantified as a 50% tissue culture infectious dose (TCID50 ml-1) [26]. The cell culture supernatant containing the virus was used for vaccine preparation.
2.3. Vaccine preparation
Water-based heat-killed and formalin-killed vaccines were prepared as described in previous studies [13]. Briefly, the clarified viral supernatant was inactivated by heating at 60oC for 2.5 h or incubating with 0.006% formaldehyde solution at 25oC for 24 h to generate W-HKV and W-FKV, respectively. Virus-free cell-culture supernatant was used as a control (W-C) for water-based vaccine treatment. Virus inactivation was confirmed by incubating the inactivated viral solutions on E11 cells and observing for 7 days for the absence of a CPE. Oil-based O-HKV and O-FKV were generated by mixing the water-based vaccines with adjuvant Motanide ISA 763A VG (Seppic, France) at a ratio of 3:7 using a high shear homogenizer (IKA T25 digital Ultra Turrax). Virus-free cell-culture supernatant mixed with adjuvant was used as control (O-C) for oil-based vaccine treatment. Vaccines were stored at 4oC until used.
2.4. Immunization, sampling, and challenge test
The study consisted of 6 groups, with 45 fish per group. Fish in treatment groups 1, 2 and 3 were IP injected with 100 µl of W-C, W-HKV and W-FKV, while fish in treatment groups 4, 5 and 6 were injected IP with 100 µl of O-C, O-HKV and O-FKV, respectively (Table 1). Booster vaccinations were performed in the same manner 2 weeks post-primary vaccination (wppv). Before sampling, fish were anaesthetized with clove oil at 100 ppm. Three fish were randomly selected from every treatment group. Blood was sampled for measurement of serum IgM antibody levels, and spleen and kidney were sampled for analysis of immune gene expression at 7, 14 and 21 days post-primary vaccination (dppv). Blood was collected from the caudal vein and centrifuged at 5000g at 4oC for 15 min. Fish sera were collected and stored at -20 °C until analysis. Tissues were placed immediately into 250 µl RNA later solution (Sigma) and stored at -20oC until RNA extraction. Virus stock (TCID50 is 107.25 ml-1) was diluted 100X for the challenge test. The remained fish from 6 treatments (see Table 1) were injected IP with 100 μl of the diluted viral solution. The cumulative mortality was recorded for the next 15 days. The RPS value was calculated using the following formula:
2.5. Immune gene expression by RT-qPCR
The relative fold change of immunoglobulin genes IgM, IgT, IgD, CD4 and CD8 was investigated. RNA samples were extracted from head kidney and spleen using Trizol (Invitrogen) following the protocol from the producer. DNA contamination in RNA samples was removed using DNAseI (Ambicon, USA) according to the manufacturer’s procedure. RNA samples were re-purified using phenol-chloroform (5:1, pH 4.7) and their qualities were determined by Nanodrop (Thermo Scientific, USA). iScriptTM Reverse Transcription Supermix (Bio- Rad) was used to synthesize cDNA with 100 ng RNA template. The protocol steps included incubation at 25°C for 5 min for priming, followed by 46°C for 20 min for reverse transcription and then 95°C for 1 min for inactivation of the reverse transcriptase. Immunoglobulin gene IgM, IgT and IgD expression were analyzed using quantitative real-time PCR (RT-qPCR) using the primers listed in Table 2 and iTaq Universal SYBR Supermix (Bio-rad, USA). The 20 µl reaction consisted of 10.0 µl 2X Supermix, 1.0 µl forward and reverse primers (10 µM each), 1.0 µl cDNA and 7.0 µl distilled water. The protocol for qPCR comprised of one step for initial activation at 95oC for 2 mins, followed by 40 cycles of denaturation at 95oC for 30s, annealing at the optimal temperature of each primer pair for 30s (Table 2) and extension at 72oC for 30s. Melt curves were analyzed from 65oC to 95oC during the heating process. EF-1α gene was used as a housekeeping gene for normalization in gene expression of vaccinated and control fish using the 2-ΔΔCt method [27].
2.6. Measurement of IgM antibody levels
Ninety-six-well ELISA plates were coated by 0.01% poly-L- lysine solution (Sigma, Saint Louis, MO, USA) for 1 h in coating buffer (15 mM Na2CO3; 35 mM NaHCO3, pH 9.6). The plates were then rinsed three times with low salt wash buffer (LSWB, 2 mM Tris; 38 mM NaCl; 0.005% Tween 20, pH 7.3) before incubating them with 100 µL of inactivated TiLV supernatant (TCID50 is 107.25 ml-1) overnight at 4oC. The next day, 50 μL glutaraldehyde 0.05% (EMS, USA) was added to the wells. After 20 mins the contents of the wells were removed, the plates were washed as described above and 200 μL of 1% bovine serum albumin (BSA, Sigma, Saint Louis, MO, USA) in 1x phosphate-buffered saline (PBS) (137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4 and 1.8 mM KH2PO4, pH 7.4) was added to each well f for 2 h at 28oC to block non-specific binding sites. Fish sera were diluted 1:512 in 1x PBS and 100 µL of the diluted fish sera was added to the plate, then incubated overnight at 4oC. On the third day, the plates were washed 5 times with high salt wash buffer (HSWB, 2 mM Tris; 50 mM NaCl; 0.01% Tween 20, pH 7.7). Then, 100 µL of anti-tilapia IgM monoclonal antibody [28] (diluted 1:200 in 1× PBS + 1% BSA) was added to each well and incubated for 2 h at 28oC, and the plate was again washed 5 times with HSWB. Goat anti-mouse antibody conjugated with horseradish peroxidase (diluted 1:3000 in LSWB + 1% BSA) was added to the late for 1 h at 28°C. Finally, the plates were washed 5 times with HSWB, and 100 µL of 3,3′,5,5′-Tetramethylbenzidine (TMB) substrate solution was added to each well. The reaction was allowed to develop in the dark for 5 mins before adding 50 μL of stop solution (2M H2SO4). The absorbance of the reaction was measured at 450 nm using the microplate reader (SpectraMax ID3, San Jose, CA, USA).
2.7. Statistical analysis
The IgM antibody levels and gene expression data were compared using one way ANOVA and LSD post-hoc test. Kaplan-Meier survival analysis was performed to illustrate the cumulative mortality. Graphpad prism 6 (Graphpad software, USA) was used to generate the graphs.
3. Results
3.1. Vaccine efficacy
After the challenge with TiLV, mortalities started to appear in the oil-based groups before the water-based groups. The first mortality was seen in O-C at 4-day post-challenge (dpc), followed by the O-FHV and O-HKV groups with moribund or dead fish found at 6- and 7-dpc, respectively (Figure 1). Meanwhile, the first mortality in the W-C group was recorded 7-dpc, whereas mortalities in the W-HKV and W-FKV groups were first observed at 8-9 dpc (Figure 1). The RPS values were 54.6% and 59.7% for W-HKV and W-FKV treatments, compared to 32.0% and 0% for O-HKV and O-FKV (p < 0.01) treatments, respectively (Table 1).
3.2. Immunoglobulin gene expression
The mRNA transcript levels of five immunoglobulin genes (IgM, IgD, IgT, CD4 and CD8) in head kidney and spleen were compared between vaccinated and control groups (Table 3A-B). For the head-kidney from the oil-based treatment groups, significant up-regulation of IgT, IgD transcripts (almost 3-fold) and CD4 (around 8-fold) were observed in the O-HKV group at 7-dppv (p<0.05), followed by a significant increase in IgD mRNA levels after the booster vaccination (21-dppv) compared to the control group O-C (p<0.01). In the O-FKV group, mild upregulation of IgD was observed at 7 and 21-dppv (p<0.05) compared to the O-C treatment group. For the water-based treatments groups, all immunoglobulin transcript levels were up-regulated in the head kidney of fish vaccinated with W-FKV with significant increases in IgM and IgD at 7- dppv in the head-kidney (with 6- and 3-fold changes, respectively) (p<0.05). (Table 3A).
Regarding the expression of immune genes in the spleen, for the oil-based treatments, CD4 transcript levels were found to be significantly up-regulated in the O-HKV group (almost 3 times, p<0.01) and O-FKV group (around 4 times, p<0.05) at 7- and 14-dppv, respectively. For the water-based treatment groups, a considerable rise in IgM transcripts was observed (p < 0.05) at 14-dppv in fish vaccinated with W-HKV (almost a 7-fold change) and W-FKV (around 4 times). In contrast, an increase in IgT and IgD mRNA levels were found in fish vaccinated with W-FKV (a 6-time increase) (p < 0.05) compared to water-based control (W-C). Only W-HKV treatment could generate a significant increase in CD8 transcript level after booster (p<0.05) (Table 3B).
3.3. IgM antibody levels
Specific levels of TiLV antibodies at 0 (pre-vaccination), 7, 14 and 21 dppv are presented as optical density (OD) values in Figure 2. Prior to vaccination, the average OD at 450 nm value of fish was 0.148 ± 0.091. For the oil-based treatment groups, no significant differences were seen in OD values between the O-C, O-HKV and O-FKV vaccinated fish, with values of 0.358 ± 0.070, 0.313 ± 0.047 and 0.304 ± 0.041, respectively, obtained at 7 dppv. The OD values raised slightly for the O-HKV group prior to the booster at 14 dppv, with readings of 0.403 ± 0.060 compared to the control group (p < 0.05). One-week post booster vaccination, a rapid increase of OD reading of almost 0.5 was seen in all three groups, but no significant differences were observed between the groups.
For the water-based treatment groups, specific TiLV antibody levels in the FKV group rose rapidly at 7 dppv (0.266), around double the value obtained for the HKV and control groups. The levels continued to increase at 14 dppv before the booster vaccination in both the W-HKV and W-FKV groups, with values of 0.248 ± 0.066 and 0.312 ± 0.241 compared to 0.159 ± 0.149 for the W-C group. No significant increases were found in vaccinated groups compared to the control one week post-booster vaccination.
4. Discussion
Emulsification of antigens in oil-based adjuvants is commonly used in vaccine formulations for fish. These types of formulations have been shown to be effective in protecting farmed fish against certain bacterial diseases such as Edwardsiella ictaluri, Edwardsiella tarda, Aeromonas salmonicida, Vibrio harveyi, and Streptococcus agalactiae, for example [20,21,23,29]. However, the oil adjuvant MontanideTM ISA 763A VG might not be suitable for formulation with antigen derived from inactivated virus, such as TiLV in the current study, as this vaccine resulted in very low or no protection with RPS values of 32% and 0%, respectively, compared to the corresponding water-based vaccines, W-HKV and W-FKV (54.6 and 59.7%). This outcome may have resulted from lower concentrations of viral particles in the oil-based vaccines after formulating them with the oil adjuvant or the slower release of antigens due to the oil coating covering the virus. Veenstra et al (2021) showed that different adjuvants can elicit different degrees of responsiveness [20], suggesting that the efficacy of other adjuvants should be investigated in the TiLV vaccine to establish if they produce a more efficacious vaccine formulation. Further research should also focus on whether different antigen concentrations formulated with adjuvant affect vaccine efficacy and whether the duration of immunity post-vaccination can impact the challenge results.
This study found a correlation between the level of immunoglobulin mRNA expression and the effectiveness of vaccines. The W-HKV and W-FKV were found to stimulate the expression of three classes of IgM, IgD and IgT at 7 to 14 dppv. On the other hand, the O-HKV and O-FKV triggered slight up-regulation of IgT and IgD. The highest expression level of IgM mRNA (6 to 7-fold increase) was observed with the water-based vaccines, suggesting that IgM may be more important than the other immunoglobulins in protecting fish from TiLV infection. This assumption is supported by the higher protection offered by the water-based TiLV vaccines compared to the oil-based TiLV vaccines. Although levels of secreted IgM were not performed in this study, our previous studies showed that the same TiLV water-based vaccines formulations were effective in eliciting an IgM antibody response in both the serum and the mucus and provided protection when vaccinated fish were challenged with TiLV [13,14]. The level of IgM and neutralizing antibodies in fish serum after vaccination with a TiLV vaccine were also reported by Zeng et al., (2021). The upregulation of CD4 transcript levels found in the oil-based and water-base vaccine groups may reflect differentiation of CD4+ naïve cell into the helper T cell (Th1 and Th2 cells), which can trigger B cells differentiate into plasma cells to produce TiLV-specific antibodies [29,30]. Only the W-HKV group triggered a significant increase in CD8 levels, suggesting that this vaccine may stimulate differentiation of CD8+ naïve cells into the cytotoxic T-cells [30,[30,31].
When it comes to the TiLV- specific antibody levels, the IgM levels were found to significantly increase at 14-dppv before booster for both water-based and oil-based treatments compared to the control, which may indicate that specific IgM level may reach the highest level at 2-week post primary immunization. This was in agreement with our previous study where we investigated only water-based vaccines using intraperitoneal injection to administer the vaccine to juvenile fish [13]. The results from both studies were similar with the kinetics of IgM antibody secretion from red tilapia infected with TiLV [[33]. Co-relation of between IgM mRNA levels with antibody secretion was found at 14 dppv in head-kidney for O-HKV treatment group and spleen of the W-HKV group. However, it is important to note that the dynamics of the antibody level response observed in this study differ from those reported by Mai et al. (2021). Specifically, the antibody levels did not show a significant increase after administration of the booster dose. This may due to the antigen was neutralized by the remaining TiLV-specific antibodies before booster, suggesting that the timing of the booster vaccination is important to elicit an increased antibody response. That may explain for the low protection for both oil-based and water-based vaccines in this study.
In summary, these findings suggest several key considerations for developing TiLV vaccines. Firstly, water-based vaccines showed high potential for the development of injectable autogenous vaccines against TiLV. Secondly, the MontanideTM ISA 763A VG oil adjuvant may not be suitable for formulating TiLV inactivated vaccines. Lastly, the booster dose given at 3 weeks post-primary vaccination (as reported by Mai et al., 2021) is more effective in terms of immunological responses and efficacy than a booster given at 2-weeks post-primary vaccination in this study. This knowledge can be used to improve the design of future vaccination strategies against TiLV and possibly other fish pathogens.
Data Availability Statement
The data that support the findings of this study are available on request.
Acknowledgments
This study was financially supported by the GCRF Networks in Vaccines Research and Development, co-funded by the MRC and BBSRC and supported by the International Veterinary Vaccinology Network (IVVN). The authors thank for the partial support from the Royal Golden Jubilee Ph.D. program for ASEAN students (RGJ-ASEAN) and Thailand Science Research and Innovation (TSRI) Fund (grant number CU_FRB65_food (25)189_31_08). Much appreciation is given to Asst. Prof. Dr. Pattanapon Kayansamruaj (deceased) for his significant contribution during the project.
Conflicts of Interest
The authors declare no conflict of interest.
References
- FAO. In Brief to The State of World Fisheries and Aquaculture 2022. 2022. Available online: https://www.fao.org/3/cc0461en/cc0461en.pdf.
- Prabu, E.; Rajagopalsamy, C.; Ahilan, B.; Jeevagan, I.; Renuhadevi, M. Tilapia—An excellent candidate species for world aquaculture: A review. Annu. Res. Rev. Biol. 2019, 31, 1–14. [Google Scholar] [CrossRef]
- Kayansamruaj, P.; Areechon, N.; Unajak, S. Development of fish vaccine in Southeast Asia: A challenge for the sustainability of SE Asia aquaculture. Fish Shellfish. Immunol. 2020, 103, 73–87. [Google Scholar] [CrossRef]
- Tang, K.F.J.; Bondad-Reantaso, M.G.; Surachetpong, W.; Dong, H.T.; Fejzic, N.; Wang, Q.; Wajsbrot, N.; Hao, B. FAO Fisheries and Aquaculture Circular NFIM / C1220 ( En ) TILAPIA LAKE VIRUS Fisheries and. vol. 1220. 2021.
- Eyngor, M.; Zamostiano, R.; Tsofack, J.E.K.; Berkowitz, A.; Bercovier, H.; Tinman, S.; Lev, M.; Hurvitz, A.; Galeotti, M.; Bacharach, E.; et al. Identification of a Novel RNA Virus Lethal to Tilapia. J. Clin. Microbiol. 2014, 52, 4137–4146. [Google Scholar] [CrossRef]
- Ferguson, H.W.; Kabuusu, R.; Beltran, S.; Reyes, E.; A Lince, J.; del Pozo, J. Syncytial hepatitis of farmed tilapia, Oreochromis niloticus (L.): a case report. J. Fish Dis. 2013, 37, 583–589. [Google Scholar] [CrossRef]
- Dong, H.T.; Senapin, S.; Gangnonngiw, W.; Nguyen, V.V.; Rodkhum, C.; Debnath, P.P.; Delamare-Deboutteville, J.; Mohan, C.V. Experimental infection reveals transmission of tilapia lake virus (TiLV) from tilapia broodstock to their reproductive organs and fertilized eggs. Aquaculture 2020, 515, 734541. [Google Scholar] [CrossRef]
- Yamkasem, J.; Tattiyapong, P.; Kamlangdee, A.; Surachetpong, W. Evidence of potential vertical transmission of tilapia lake virus. J. Fish Dis. 2019, 42, 1293–1300. [Google Scholar] [CrossRef]
- Secombes, C.J.; Belmonte, R. Overview of the Fish Adaptive Immune System. Birkhauser Adv Infect Dis 2016, 35–52. [Google Scholar] [CrossRef]
- Bacharach, E.; Eldar, A. (2016). Tilapia lake virus vaccines. US Patent Application Publication no.US2016/0354458A1. Available online: https://patents.google.com/paten t/US201 60354 458A1/en.
- Zeng, W.; Wang, Y.; Chen, X.; Wang, Q.; Bergmann, S.M.; Yang, Y.; Wang, Y.; Li, B.; Lv, Y.; Li, H.; et al. Potency and efficacy of VP20-based vaccine against tilapia lake virus using different prime-boost vaccination regimens in tilapia. Aquaculture 2021, 539, 736654. [Google Scholar] [CrossRef]
- Zeng, W.; Wang, Y.; Hu, H.; Wang, Q.; Bergmann, S.; Wang, Y.; Li, B.; Lv, Y.; Li, H.; Yin, J.; et al. Cell Culture-Derived Tilapia Lake Virus-Inactivated Vaccine Containing Montanide Adjuvant Provides High Protection against Viral Challenge for Tilapia. Vaccines 2021, 9, 86. [Google Scholar] [CrossRef]
- Mai, T.T.; Kayansamruaj, P.; Taengphu, S.; Senapin, S.; Costa, J.Z.; Del-Pozo, J.; Thompson, K.D.; Rodkhum, C.; Dong, H.T. Efficacy of heat-killed and formalin-killed vaccines against Tilapia tilapinevirus in juvenile Nile tilapia ( Oreochromis niloticus ). J. Fish Dis. 2021, 44, 2097–2109. [Google Scholar] [CrossRef]
- Mai, T.T.; Kayansamruaj, P.; Soontara, C.; Kerddee, P.; Nguyen, D.-H.; Senapin, S.; Costa, J.Z.; Del-Pozo, J.; Thompson, K.D.; Rodkhum, C.; et al. Immunization of Nile Tilapia (Oreochromis niloticus) Broodstock with Tilapia Lake Virus (TiLV) Inactivated Vaccines Elicits Protective Antibody and Passive Maternal Antibody Transfer. Vaccines 2022, 10, 167. [Google Scholar] [CrossRef] [PubMed]
- Tattiyapong, P.; Kitiyodom, S.; Yata, T.; Jantharadej, K.; Adamek, M.; Surachetpong, W. Chitosan nanoparticle immersion vaccine offers protection against tilapia lake virus in laboratory and field studies. Fish Shellfish. Immunol. 2022, 131, 972–979. [Google Scholar] [CrossRef] [PubMed]
- Bonam, S.R.; Partidos, C.D.; Halmuthur, S.K.M.; Muller, S. An Overview of Novel Adjuvants Designed for Improving Vaccine Efficacy. Trends Pharmacol. Sci. 2017, 38, 771–793. [Google Scholar] [CrossRef] [PubMed]
- Tafalla, C.; Bøgwald, J.; Dalmo, R.A. Adjuvants and immunostimulants in fish vaccines: Current knowledge and future perspectives. Fish Shellfish. Immunol. 2013, 35, 1740–1750. [Google Scholar] [CrossRef]
- Jazayeri, S.D.; Lim, H.X.; Shameli, K.; Yeap, S.K.; Poh, C.L. Nano and Microparticles as Potential Oral Vaccine Carriers and Adjuvants Against Infectious Diseases. Front. Pharmacol. 2021, 12, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Hoare, R.; Jung, S.-J.; Ngo, T.; Bartie, K.; Bailey, J.; Thompson, K.; Adams, A. Efficacy and safety of a non-mineral oil adjuvanted injectable vaccine for the protection of Atlantic salmon (Salmo salar L.) against Flavobacterium psychrophilum. Fish Shellfish. Immunol. 2019, 85, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Veenstra, K.A.; Wang, T.; Russell, K.S.; Tubbs, L.; Ben Arous, J.; Secombes, C.J. Montanide™ ISA 763A VG and ISA 761 VG induce different immune pathway responses in rainbow trout (Oncorhynchus mykiss) when used as adjuvant for an Aeromonas salmonicida bacterin. Fish Shellfish. Immunol. 2021, 114, 171–183. [Google Scholar] [CrossRef]
- Arous, J.B. Vaccine oil adjuvants for the development of aquaculture. Veterinary Science Today 2016;4:62-65.
- Cao, T.T.; Tsai, M.-A.; Yang, C.-D.; Wang, P.-C.; Kuo, T.-Y.; Chen, H.-C.G.; Chen, S.-C. Vaccine efficacy of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) from Edwardsiella ictaluri against E. tarda in tilapia. J. Gen. Appl. Microbiol. 2014, 60, 241–250. [Google Scholar] [CrossRef]
- Xu, W.; Jiao, C.; Bao, P.; Liu, Q.; Wang, P.; Zhang, R.; Liu, X.; Zhang, Y. Efficacy of Montanide™ ISA 763 A VG as aquatic adjuvant administrated with an inactivated Vibrio harveyi vaccine in turbot (Scophthalmus maximus L.). Fish Shellfish. Immunol. 2018, 84, 56–61. [Google Scholar] [CrossRef]
- Chamtim, P.; Suwan, E.; Dong, H.T.; Sirisuay, S.; Areechon, N.; Wangkahart, E.; Hirono, I.; Mavichak, R.; Unajak, S. Combining segments 9 and 10 in DNA and recombinant protein vaccines conferred superior protection against tilapia lake virus in hybrid red tilapia (oreochromis sp.) compared to single segment vaccines. Front. Immunol. 2022, 13, 935480. [Google Scholar] [CrossRef]
- Taengphu, S.; Kayansamruaj, P.; Kawato, Y.; Delamare-Deboutteville, J.; Mohan, C.V.; Dong, H.T.; Senapin, S. Concentration and quantification of Tilapia tilapinevirus from water using a simple iron flocculation coupled with probe-based RT-qPCR. PeerJ 2022, 10, e13157. [Google Scholar] [CrossRef]
- Reed, L. .; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Soonthonsrima, T.; Wangman, P.; Chaivisuthangkura, P.; Pengsuk, C.; Sithigorngul, P.; Longyant, S. Generation of mouse monoclonal antibodies specific to tilapia immunoglobulin using fish immunoglobulin/BSA complex for monitoring of the immune response in Nile tilapia Oreochromis niloticus. Aquac. Res. 2018, 50, 277–283. [Google Scholar] [CrossRef]
- Wangkahart, E.; Thongsrisuk, A.; Vialle, R.; Pholchamat, S.; Sunthamala, P.; Phudkliang, J.; Srisapoome, P.; Wang, T.; Secombes, C.J. Comparative study of the effects of Montanide™ ISA 763A VG and ISA 763B VG adjuvants on the immune response against Streptococcus agalactiae in Nile tilapia (Oreochromis niloticus). Fish Shellfish. Immunol. 2023, 134, 108563. [Google Scholar] [CrossRef] [PubMed]
- Smith, N.C.; Rise, M.L.; Christian, S.L. A Comparison of the Innate and Adaptive Immune Systems in Cartilaginous Fish, Ray-Finned Fish, and Lobe-Finned Fish. Front. Immunol. 2019, 10, 2292. [Google Scholar] [CrossRef] [PubMed]
- Secombes, C.; Wang, T. The innate and adaptive immune system of fish. Woodhead Publishing Limited; 2012. [CrossRef]
- Somamoto, T.; Nakanishi, T.; Okamoto, N. Role of Specific Cell-Mediated Cytotoxicity in Protecting Fish from Viral Infections. Virology 2002, 297, 120–127. [Google Scholar] [CrossRef]
- Tattiyapong, P.; Dechavichitlead, W.; Waltzek, T.B.; Surachetpong, W. Tilapia develop protective immunity including a humoral response following exposure to tilapia lake virus. Fish Shellfish. Immunol. 2020, 106, 666–674. [Google Scholar] [CrossRef]
- Velázquez, J.; Acosta, J.; Lugo, J.M.; Reyes, E.; Herrera, F.; González, O.; Morales, A.; Carpio, Y.; Estrada, M.P. Discovery of immunoglobulin T in Nile tilapia (Oreochromis niloticus): A potential molecular marker to understand mucosal immunity in this species. Dev. Comp. Immunol. 2018, 88, 124–136. [Google Scholar] [CrossRef]
- Wang, B.; Wang, P.; Wu, Z.-H.; Lu, Y.-S.; Wang, Z.-L.; Jian, J.-C. Molecular Cloning and Expression Analysis of IgD in Nile Tilapia (Oreochromis niloticus) in Response to Streptococcus agalactiae Stimulus. Int. J. Mol. Sci. 2016, 17, 348. [Google Scholar] [CrossRef]
Figure 1.
Percent survival of oil-based vaccine and water-based-vaccine groups compared to the control during 15-day post-challenge with TiLV (strain TiLV-KT). Level of significant difference were indicated by * and ** indicating levels of significance of p<0.05 and p<0.01, respectively. Significant differences were found between O-FKV and W-HKV (p < 0.01), between O-FKV and W-FKV (p < 0.01), and between W-FKV and O-C (p< 0.05).
Figure 1.
Percent survival of oil-based vaccine and water-based-vaccine groups compared to the control during 15-day post-challenge with TiLV (strain TiLV-KT). Level of significant difference were indicated by * and ** indicating levels of significance of p<0.05 and p<0.01, respectively. Significant differences were found between O-FKV and W-HKV (p < 0.01), between O-FKV and W-FKV (p < 0.01), and between W-FKV and O-C (p< 0.05).

Figure 2.
Specific TiLV- IgM antibody levels in fish sera (diluted 1:512) of treatment groups at day 0 (pre-vaccination), 7, 14 and 21-dppv in the oil-based (A) and water-based (B) groups expressed as an absorbance value at 450 nm . Data were present as mean ± SEM (n=3). O: oil-based treatment, W: water-based treatment, C: control group (unvaccinated group), HKV: heat-killed vaccine, FKV: formalin-killed vaccine. * significant level p < 0.05.
Figure 2.
Specific TiLV- IgM antibody levels in fish sera (diluted 1:512) of treatment groups at day 0 (pre-vaccination), 7, 14 and 21-dppv in the oil-based (A) and water-based (B) groups expressed as an absorbance value at 450 nm . Data were present as mean ± SEM (n=3). O: oil-based treatment, W: water-based treatment, C: control group (unvaccinated group), HKV: heat-killed vaccine, FKV: formalin-killed vaccine. * significant level p < 0.05.
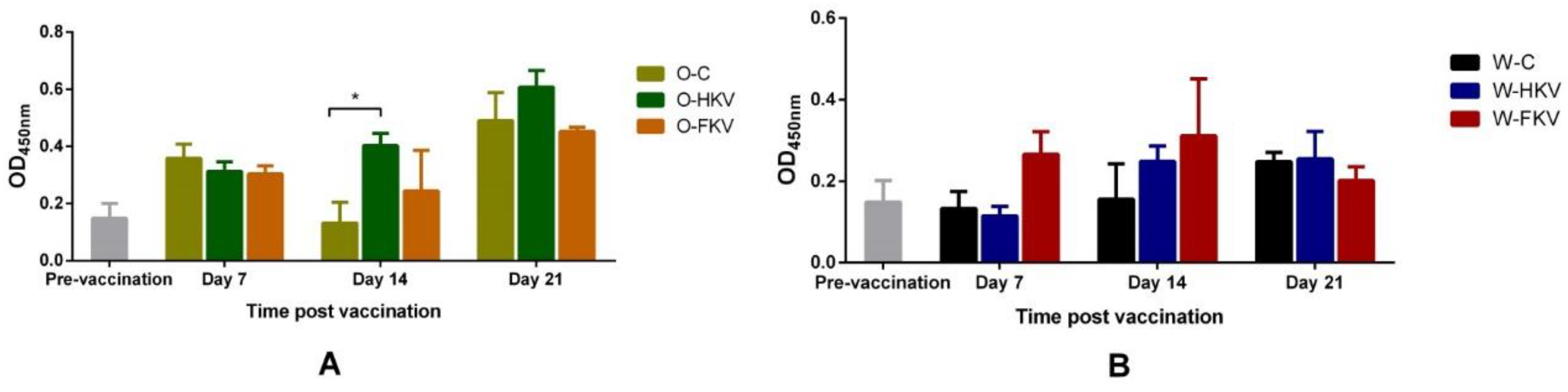

Table 1.
Details of experiment groups and challenge results.
| Treatment | Abbreviation | Number of fish challenged | Number of total dead fish at 15 dpc | RPS (%) 15 dpc |
|---|---|---|---|---|
| Water based (W) | ||||
| Control (Cell culture supernatant, L15) | W-C | 31 | 5 | |
| Heat-killed vaccine | W-HKV | 27 | 2 | 54.6 |
| Formalin-killed vaccine | W-FKV | 31 | 2 | 59.7 |
| Oil-based (O) (MontanideTM ISA 763A VG) | ||||
| Control (L15 + Oil adjuvant) | O-C | 32 | 8 | |
| Heat-killed vaccine | O-HKV | 28 | 5 | 32.0 |
| Formalin-killed vaccine | O-FKV | 24 | 9 | 0 |
dpc, day post challenge; RPS: relative percentage survival.
Table 2.
Detail of primers used for gene expression.
| Gene | Oligo sequences | Annealing temperature (oC) | Product size (bp) | Reference |
|---|---|---|---|---|
| EF-1α | F-5’-CTACAGCCAGGCTCGTTTCG-3’ R-5’-CTTGTCACTGGTCTCCAGCA-3’ |
56 | 139 | [34] |
| IgM | F-5’-GGATGACGAGGAAGCAGACT-3’ R-5’-CATCATCCCTTTGCCACTGG-3’ |
53 | 122 | [34] |
| IgT | F-5’-TGACCAGAAATGGCGAAGTCTG-3’ R-5’-GTTATAGTCACATTCTTTAGAATTACC-3’ |
53 | 163 | [34] |
| IgD | F-5′- AACACCACCCTGTCCCTGAAT- 3′ R-5’-GGGTGAAAACCACATTCCAAC- 3’ |
61 | 127 | [35] |
| CD4 | F- 5′- GCTCCAGTGTGACGTGAAA- 3′ R- 5′- TACAGGTTTGAGTTGAGCTG- 3′ |
61 | 106 | [13] |
| CD8 | F- 5′- GCTGGTAGCTCTGGCCTTT- 3′ R- 5′- TGTGATGGTGTGGGCATCTC- 3′ |
49.5 | 91 | [13] |
Table 3A.
Fold change for IgM, IgT, IgD, CD4 and CD8 gene expression in head kidney for the oil-based and water-based vaccine groups at 7, 14, and 21 days post-immunization (↑: significant up-regulation of gene expression at a significant level of p<0.05; ↑↑: significant up-regulation of gene expression at a significant level of e p< 0.01). Data were present as mean ± SEM (n=3).
Table 3A.
Fold change for IgM, IgT, IgD, CD4 and CD8 gene expression in head kidney for the oil-based and water-based vaccine groups at 7, 14, and 21 days post-immunization (↑: significant up-regulation of gene expression at a significant level of p<0.05; ↑↑: significant up-regulation of gene expression at a significant level of e p< 0.01). Data were present as mean ± SEM (n=3).
| Time point | Treatment groups | Head kidney | ||||
|---|---|---|---|---|---|---|
| IgM | IgT | IgD | CD4 | CD8 | ||
| D7 | O-C | 1.07 ± 0.46 | 1.02 ± 0.20 | 1.07 ± 0.44 | 1.09 ± 0.56 | 1.08 ± 0.51 |
| O-HKV | 1.75 ± 0.78 | 3.14 ± 0.73↑ | 2.93 ± 1.24↑ | 8.53 ± 4.97↑ | 0.79 ± 0.11 | |
| O-FKV | 1.53 ± 0.47 | 1.61 ± 0.99 | 1.89 ± 0.11↑ | 2.22 ± 0.46 | 0.68 ± 0.46 | |
| W-C | 1.55 ± 1.77 | 1.48 ± 1.61 | 1.07 ± 0.44 | 1.05 ± 0.40 | 1.03 ± 0.31 | |
| W-HKV | 0.96 ± 0.74 | 0.55 ± 0.32 | 1.19 ± 0.86 | 0.33 ± 0.05 | 0.20 ± 0.01 | |
| W-FKV | 6.00 ± 2.81↑ | 1.43 ± 0.85 | 3.80 ± 1.81↑ | 3.51 ± 2.55 | 2.26 ± 2.22 | |
| D14 | O-C | 1.58 ± 1.21 | 1.09 ± 0.48 | 1.01 ± 0.17 | 1.20 ± 0.73 | 1.40 ± 1.42 |
| O-HKV | 4.94 ± 4.11 | 0.77 ± 0.45 | 1.46 ± 0.92 | 2.89 ± 3.02 | 0.95 ± 1.00 | |
| O-FKV | 1.84 ± 2.25 | 0.37 ± 0.36 | 1.08 ±1.14 | 0.91 ± 0.42 | 1.40 ±1.03 | |
| W-C | 1.44 ± 1.05 | 0.96 ± 0.80 | 1.53 ± 1.70 | 1.02 ± 0.28 | 1.47 ± 1.59 | |
| W-HKV | 1.81 ± 0.64 | 1.09 ± 0.53 | 1.35 ± 0.45 | 2.25 ± 0.80 | 2.04 ± 1.37 | |
| W-FKV | 7.61 ± 8.34 | 2.93 ± 2.19 | 3.46 ± 2.27 | 5.65 ± 2.31↑ | 1.40 ± 1.15 | |
| D21 | O-C | 1.16 ± 0.80 | 1.02 ± 0.22 | 1.04 ± 0.33 | 1.80 ± 0.45 | 1.02 ± 0.23 |
| O-HKV | 0.91 ± 0.42 | 1.44 ± 0.40 | 3.22 ± 0.44↑↑ | 2.33 ± 0.71 | 2.31 ± 2.30 | |
| O-FKV | 1.91 ± 0.26 | 0.90 ± 0.13 | 2.09 ± 0.45↑ | 0.96 ± 0.43 | 1.00 ± 0.75 | |
| W-C | 1.36 ±1.30 | 1.29 ± 0.93 | 1.17 ± 0.82 | 1.22 ± 0.73 | 2.28 ± 3.21 | |
| W-HKV | 1.25 ± 0.94 | 1.20 ± 1.30 | 2.01 ±1.45 | 0.82 ± 0.50 | 5.29 ± 2.81 | |
| W-FKV | 0.83 ±0.66 | 0.66 ± 0.53 | 0.93 ± 0.61 | 0.46 ± 0.43 | 1.48 ± 1.66 | |
Table 3B.
Fold change for IgM, IgT, IgD, CD4 and CD8 gene expression in the spleen of the oil-based and water-based vaccine groups at 7, 14, and 21 days post-immunization (↑: significant up-regulation of gene expression at a significant level of p<0.05; ↑↑: significant up-regulation of gene expression at a significant level of e p< 0.01). Data were present as mean ± SEM (n=3).
Table 3B.
Fold change for IgM, IgT, IgD, CD4 and CD8 gene expression in the spleen of the oil-based and water-based vaccine groups at 7, 14, and 21 days post-immunization (↑: significant up-regulation of gene expression at a significant level of p<0.05; ↑↑: significant up-regulation of gene expression at a significant level of e p< 0.01). Data were present as mean ± SEM (n=3).
| Time point | Treatment groupsgroups | Spleen | ||||
|---|---|---|---|---|---|---|
| IgM | IgT | IgD | CD4 | CD8 | ||
| D7 | O-C | 1.51 ± 1.60 | 1.20 ± 0.75 | 1.05 ± 0.38 | 1.13 ± 0.72 | 1.08 ± 0.52 |
| O-HKV | 1.14 ± 0.45 | 0.90 ± 0.49 | 1.00 ± 0.78 | 2.80 ± 1.62↑↑ | 0.58 ± 0.73 | |
| O-FKV | 1.51 ± 0.94 | 0.93 ± 0.75 | 2.04 ± 1.34 | 1.93 ± 0.82 | 0.75 ± 0.49 | |
| W-C | 1.72 ± 1.97 | 1.79 ± 2.09 | 1.24 ± 0.86 | 1.44 ± 1.33 | 1.18 ± 0.71 | |
| W-HKV | 1.04 ±1.02 | 1.09 ± 0.96 | 1.23 ± 0.86 | 1.01 ± 0.69 | 0.50 ± 0.27 | |
| W-FKV | 0.31 ± 0.12 | 0.38 ± 0.22 | 0.42 ± 0.16 | 0.51 ± 0.21 | 0.19 ± 0.16 | |
| D14 | O-C | 1.19 ± 0.70 | 1.67 ± 1.72 | 1.20 ± 0.89 | 1.25 ± 1.01 | 2.60 ±2.90 |
| O-HKV | 1.09 ± 0.80 | 1.00 ± 0.67 | 1.22 ± 1.04 | 2.87 ± 1.96 | 1.50 ± 1.63 | |
| O-FKV | 1.81 ± 0.87 | 1.11 ± 0.32 | 2.71 ± 1.71 | 4.29 ± 1.00↑ | 6.75 ± 5.60 | |
| W-C | 1.43 ± 1.19 | 2.31 ± 2.80 | 1.66 ± 1.88 | 1.50 ± 1.11 | 1.06 ± 0.39 | |
| W-HKV | 6.91 ± 3.32↑ | 6.07 ± 4.82 | 4.12 ± 1.40 | 3.12 ± 1.03 | 6.14 ± 4.74 | |
| W-FKV | 4.30 ± 3.22 | 5.89 ± 0.41↑ | 5.69 ± 1.15↑ | 2.31 ± 0.51 | 3.44 ± 2.36 | |
| D21 | O-C | 1.02 ± 0.25 | 1.21 ± 0.94 | 1.02 ± 0.25 | 1.10 ± 0.51 | 1.08 ± 0.53 |
| O-HKV | 0.70 ± 0.24 | 1.15 ± 0.32 | 2.12 ± 0.56↑ | 0.83 ± 0.38 | 0.92 ± 0.75 | |
| O-FKV | 1.60 ± 2.38 | 0.43 ± 0.52 | 2.20 ± 3.05 | 0.61 ± 0.87 | 0.98 ± 1.36 | |
| W-C | 1.10 ± 0.55 | 1.70 ± 1.82 | 1.11 ± 0.65 | 1.83 ± 1.65 | 1.03 ± 0.29 | |
| W-HKV | 1.82 ± 1.79 | 2.76 ± 2.03 | 2.23 ± 2.55 | 1.58 ± 1.37 | 2.82 ± 1.55↑ | |
| W-FKV | 0.37 ± 0.10 | 0.79 ± 0.59 | 0.57 ± 0.19 | 0.33 ± 0.24 | 0.46 ± 0.38 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



