
Submitted:
21 June 2023
Posted:
21 June 2023
You are already at the latest version
Abstract
Helicosporous hyphomycetes is a class of filamentous fungi that shows promising application prospects in metabolizing bioactive natural compounds. During a study of helicosporous fungi in China, six new helicosporous taxa were collected and isolated from decaying wood in Guangxi Zhuang Autonomous Region, China. Morphological comparisons with multi-gene phylogenetic analyses revealed that these six taxa belong to Helicosporium (Tubeufiaceae, Tubeufiales), and they can be recognized as three novel species, named Helicosporium liuzhouense, H. multidentatum and H. nanningense. Detailed descriptions and illustrations of the newly discovered taxa and comparisons with similar fungi are provided. In addition, a list and a key to accepted Helicosporium species are provided.
Keywords:
6 new taxa
; ascomycetes
; asexual morph
; filamentous fungi
; phylogeny
; taxonomy
1. Introduction
Based on the type species H. vegetum, Nees [1] established Helicosporium as one of the earliest described genera of helicosporous hyphomycetes. The majority of this group’s species inhabit subtropical to tropical habitats [2,3,4,5,6,7,8,9,10,11]. Typically, they inhabit woody substrates in terrestrial and freshwater environments [3,7,8,9,10,12]. Index Fungorum (accessed on 20 May 2023) [13] currently lists 105 taxa of Helicosporium, of which 75 species have been excluded or transferred to other genera. Most of these taxa were transferred to the genera Helicoma and Neohelicosporium. Currently, there are eighteen accepted species of Helicosporium, twelve of which have molecular data [3,7,8,10,11].
Most species in this genus are reported to have a helicosporous asexual morph. There are three species with reported sexual morph, viz. H. flavum, H. sexuale and H. vegetum [7,8,14]. The sexual morph of Helicosporium is characterized by solitary, yellowish brown, globose to subglobose ascomata, cylindric-clavate, 8-spored bitunicate asci, and hyaline to yellowish brown, fusiform ascospores [7,8,14]. The asexual morph is distinguished by pale yellow to yellow green colonies on the natural woody substratum, erect, setiferous, cylindrical conidiophores with denticulate conidiogenous cells arising laterally from the lower portions of conidiophores resembling tiny tooth-like or bladder-like protrusions, and hyaline to yellow green, pleurogenous helicoid conidia that are smaller than 25 μm diameter with conidial filaments usually not exceeding 4 μm thickness [10,11].
Helicosporium fungi have certain potential in the production of bioactive secondary metabolites. The antimicrobial activity of Helicosporium was first reported by Hardy and Sivasithamparam [15]. The main antimicrobial constituent, 2-methyl resorcinol, was isolated from Helicosporium sp. KCTC 0635BP by Choi et al. [16]. It was reported to have cytotoxicity against mammalian cells, and antimicrobial activity against various types of fungi and bacteria [16].
During a study of helicosporous hyphomycetes in China, six new helicosporous taxa were collected from Guangxi Zhuang Autonomous Region. Three new species, Helicosporium liuzhouense, H. multidentatum and H. nanningense, were identified based on morphological evidence and phylogenetic analyses of combined LSU, ITS, tef1α and rpb2 sequence data. The results of the PHI test support the taxonomic classification of these three newly discovered species. The present study provides descriptive and illustrative morphological information as well.
2. Materials and Methods
2.1. Sample Collection, Specimen Examination, and Isolation
Specimens of decaying wood were randomly sampled from terrestrial and freshwater habitats in Guangxi Zhuang Autonomous Region, China. Freshwater samples were incubated in sterile, moist plastic containers at room temperature for approximately 14 days. After two weeks of collection, fresh specimens were examined and observed using a stereomicroscope (SMZ 745 and SMZ 800N, Tokyo, Nikon, Japan). Morphological characteristics of fresh fungi specimens were recorded with stereomicroscopes (SMZ 745 and SMZ 800N, Nikon, Tokyo, Japan).
Single spore isolation was referred from the method described by Chomnunti et al. [17]. Purified cultures were cultured in a 25 ℃ incubator. The morphological features of colonies, including color, shape, and colony diameter, were recorded regularly.
The dried specimens were deposited in the Herbarium of Cryptogams Kunming Institute of Botany, Academia Sinica (KUN-HKAS), Kunming, China, and the Herbarium of Guizhou Academy of Agriculture Sciences (GZAAS), Guiyang, China. The cultures were deposited in the China General Microbiological Culture Collection Center (CGMCC), Beijing, China, and the Guizhou Culture Collection (GZCC), Guizhou, China.
2.2. DNA Extraction, PCR Amplification, and Sequencing
Using sterile toothpicks, 60-day-old mycelium was scraped from PDA plates and transferred to a 1.5-mL microcentrifuge tube. Using the Ezup fungus genomic DNA extraction kit, DNA was extracted and sequenced following the manufacturer’s instructions. After obtaining the DNA of the fungal strains, EF1-983F/EF1-2218R, FRPB2-5F/FRPB2-7CR, ITS5/ITS4 and LR0R/LR5 were used as primers to amplification [18,19,20]. The amplification reactions of ITS, LSU, tef1α, and rpb2 were carried out according to the methods of Lu et al. [21,22] After PCR amplification, the products were analyzed using 1% agarose gel electrophoresis. The purification and sequencing of PCR products were completed by Beijing Tsingke Biological Engineering Technology and Services Co., Ltd. (Beijing, China).
2.3. Phylogenetic Analyses
BioEdit version 7.0.5.3 was used to inspect the original sequences. The forward and reverse sequences were assembled using SeqMan v. 7.0.0 (DNASTAR, Madison, WI, USA) software and submitted to the GenBank database. Based on recent publications, additional sequences similar to Helicosporium were downloaded from GenBank [7,8,9,10,11,14]. Sequence alignments for each locus were carried using the online multiple alignment program MAFFT version 7, and the alignments were further automatically adjusted using the trimAl tool [23]. The phylogenetic tree was constructed using the methods described by Ma et al. [24], which included Maximum likelihood (ML) and Bayesian Inference (BI).
The phylogenetic trees were edited using FigTree v1.4.0 software. The edit trees and figures layout were edited using Adobe PhotoShop CC 2018 and Adobe Illustrator CC 2021 (Adobe Systems, San Jose, CA, USA) software. Sequences generated in this study were uploaded in GenBank (Table 1).
2.4. Genealogical Concordance Phylogenetic Species Recognition (GCPSR) analysis
Three new species, H. liuzhouense, H. multidentatum and H. nanningense were analyzed using GCPSR with closely related taxa from a combined LSU-ITS-tef1-α-rpb2 gene regions. The pairwise homoplasy index (PHI) test was carried out in SplitsTree4 [25,26]. It indicates that there is no statistically significant evidence for recombination for the selected taxa when the P-value is above 0.05. Both the LogDet transformation and splits decomposition options were used to reveal the relationship among closely related species.
3. Phylogenetic Results
The partial LSU-ITS-tef1α-rpb2 nucleotide sequences were used to determine the phylogenetic position of our newly isolated taxa. The concatenated sequence matrix consisted of LSU (1–842 bp), ITS (843–1398 bp), tef1α (1399–2310 bp), and rpb2 (2311–3337 bp), totaling 3337 characters for 30 taxa and two outgroups, Acanthostigma chiangmaiensis (MFLUCC 10–0125) and A. perpusillum (UAMH 7237). The concatenated LSU, ITS, tef1α, and rpb2 datasets were analyzed using ML and BI methods, yielding similar tree topologies.
The resulting multigene phylogenetic tree confirms that our newly isolated Helicosporium liuzhouense, H. multidentatum and H. nanningense have a distant clade from other taxa within the genus Helicosporium (Figure 1).
4. Genealogical Concordance Phylogenetic Species Recognition (GCPSR) analysis
Application of the PHI test to the concatenated tree-locus sequences of LSU-ITS-tef1α-rpb2 revealed the recombination level within phylogenetically related species. No significant recombination events were observed between our species Helicosporium liuzhouense, H. multidentatum and H. nanningense and closely related species in Helicosporium (Figure 2). The test results show Φw = 1 for the combined sequence data, Φw = 1 for LSU dataset, Φw = 0.80 for ITS dataset, Φw = 0.09 for tef1α and Φw = 0.93 for rpb2 data.
5. Taxonomy
Helicosporium liuzhouense X.J. Xiao, J. Ma & Y.Z. Lu, sp. nov., Figure 3.
Index Fungorum number: IF900461
Etymology: The epithet “liuzhouense” named after the city from which the holotype was found.
Holotype: HKAS 125865
Saprobic on decaying wood in a freshwater stream. Sexual morph: Undetermined. Asexual morph: Hyphomycetous, helicosporous. Colonies on the substratum superficial, effuse, gregarious, bright lime green. Mycelium partly immersed, partly superficial, composed of branched, septate, hyphae, pale brown to brown hyphae. Conidiophores macronematous, mononematous, erect, flexuous or straight, unbranched, apical sterile, septate, cylindrical, (102) 110–180 (213) × 4–5 μm (= 145 ×5 μm, n = 25), brown to dark brown, thick-walled. Conidiogenous cells holoblastic, monoblastic to polyblastic, discrete, determinate, arising laterally from the lower portion of the conidiophores as tiny tooth-like protrusions, hyaline to pale brown, smooth-walled. Conidia solitary, pleurogenous, helicoid, rounded at tip, 13–15 μm diam., and conidial filament 1–2 μm wide ( = 14 × 1.5 μm, n = 25), 90–105 μm long, tightly coiled 2–3 times, becoming loosely coiled in water, indistinctly multi-septate, guttulate, hyaline to pale green, smooth-walled.
Culture characteristics: Conidia germinated on water agar and produced germ tubes within 8 h. The colonies grew on PDA, had a circular shape with a flat surface and undulate edge. They reached a size of 46 mm in 6 weeks at 25 ℃, and exhibited a pale brown center with brown edges on PDA.
Material examined: CHINA, Guangxi Zhuang Autonomous Region, Liuzhou City, Luzhai County, on decaying wood in a freshwater stream, 4 May 2021, Xing-Juan Xiao & Jian Ma, LZ3 (HKAS 125865, holotype; GZAAS 22–2014, isotype), ex-type living culture CGMCC, GZCC 22–2014. Ibid., LZ3-2 (HKAS 125870, paratype), living culture GZCC 23–0586.
Notes: Phylogenetically, Helicosporium liuzhouense is strongly supported as a sister species to H. multidentatum and H. nanningense (97% ML/ 1.00 PP). Morphologically, Helicosporium liuzhouense is distinguished from H. multidentatum by having conidiogenous cells with tiny tooth-like protrusions, while H. multidentatum has conidiogenous cells with integrated multi-dentate protrusions. Additionally, H. liuzhouense is characterized by shorter conidia (90–105 µm vs. 105–128 µm) and a larger conidial diameter (13–15 µm vs. 12–13 µm) compared to H. multidentatum. H. liuzhouense can be differentiated from H. nanningense by having shorter conidiophores (90–115 µm vs. 100–215 µm) and conidial filaments (82–92 µm vs. 90–105 µm). The PHI test provides strong evidence showing that they are separate species (Figure 2). Although H. liuzhouense and H. viridisporum share similar conidiophores, conidiogenous cells and conidial features[11], the phylogenetic analyses indicating that they are distinct species.
Helicosporium multidentatum X.J. Xiao, J. Ma & Y.Z. Lu, sp. nov., Figure 4.
Index Fungorum number: IF900460
Etymology: The epithet “multidentatum” referring to the multi-dentate integration protrusions conidiogenous cells.
Holotype: HKAS 125856
Saprobic on decaying wood in a terrestrial habitat. Sexual morph: Undetermined. Asexual morph: Hyphomycetous, helicosporous. Colonies on the substratum superficial, effuse, gregarious, bright lime green. Mycelium partly immersed, partly superficial, composed of branched, septate, pale brown to brown hyphae. Conidiophores macronematous, mononematous, erect, flexuous or straight, unbranched, apical sterile, septate, cylindrical, 130–200 × 4–6 μm ( = 165 × 5 μm, n = 20), brown to dark brown, thick-walled. Conidiogenous cells holoblastic, monoblastic to polyblastic, discrete, determinate, arising laterally from the lower portion of the conidiophores as integrated multi-dentate protrusions, hyaline to pale brown, smooth-walled. Conidia solitary, pleurogenous, helicoid, rounded at tip, 12–13 μm diam., and conidial filament 1.5–3 μm wide ( = 12.5× 2.5 μm, n = 30), 105–130 μm long, tightly coiled 31/4–33/4 times, becoming loosely coiled in water, indistinctly multi-septate, guttulate, hyaline to pale green, smooth-walled.
Culture characteristics: Conidia germinated on water agar and produced germ tubes within 8 h. The colonies grew on PDA, had a circular shape with a flat surface and undulate edge. They reached a size of 35 mm in 5 weeks at 25 ℃, and exhibited a pale brown center with brown edges on PDA.
Material examined: CHINA, Guangxi Zhuang Autonomous Region, Guilin City, Qixingyan Scenic spot, on decaying wood, 4 May 2021, Xing-Juan Xiao & Jian Ma, QXY8 (HKAS 125856, holotype; GZAAS 22–2013, isotype), ex-type culture CGMCC, GZCC 22–2013. Ibid., QXY8-2 (HKAS 125855, paratype), living culture GZCC 23–0585.
Notes: Morphologically, Helicosporium multidentatum is similar to H. hainanense and H. vesicarium in having brown to dark brown, unbranched and septate conidiophores with integrated multi-dentate protrusions arising laterally from its lower portion, and hyaline to pale green or yellowish, pleurogenous, helicoid conidia [10,11]. However, H. multidentatum differs from H. hainanense in having longer conidial filaments (105–130 μm vs. 55–60 μm), and from H. vesicarium in having smaller conidial diameter (12–13 µm vs. 13–18 µm) [10,11]. Phylogenetically, H. multidentatum forms a sister clade to H. liuzhouense with strong support and is distant from H. hainanense and H. vesicarium (Figure 1).
Helicosporium nanningense X.J. Xiao, J. Ma & Y.Z. Lu, sp. nov., Figure 5.
Index Fungorum number: IF900556
Etymology: The epithet “nanningense” named after the city from which the holotype was found.
Holotype: HKAS 128858
Saprobic on decaying wood in a terrestrial habitat. Sexual morph: Undetermined. Asexual morph: Hyphomycetous, helicosporous. Colonies on the substratum superficial, effuse, gregarious, bright green. Mycelium partly immersed, partly superficial, branched, septate, pale brown to brown hyphae. Conidiophores macronematous, mononematous, flexuous or straight, unbranched, apical sterile, septate, cylindrical, 90–115 × 4–5 μm ( = 102 × 5 μm, n = 20), brown to dark brown, thick-walled. Conidiogenous cells holoblastic, monoblastic to polyblastic, discrete, determinate, arising laterally from the lower portion of the conidiophores as tiny tooth-like protrusions, hyaline to pale brown, smooth-walled. Conidia solitary, pleurogenous, helicoid, rounded at tip, 11–14 μm diam., and conidial filament 1.5–2.0 μm wide ( = 13 × 1.8 μm, n = 30), 82–92 μm long, tightly coiled 2–31/2 times, becoming loosely coiled in water, indistinctly multi-septate, guttulate, hyaline to pale green, smooth-walled.
Culture characteristics: Conidia germinated on water agar and produced germ tubes within 12 h. The colonies grew on PDA, had a circular shape with a flat surface and undulate edge. They reached a size of 42 mm in 5 weeks at 25 ℃, and exhibited a brown center with nigger-brown edges on PDA.
Material examined: CHINA, Guangxi Medicinal Botanical Garden, Nanning City, on decaying wood in a terrestrial habitat, 4 May 2021, Xing-Juan Xiao & Jian Ma, GXZWY3.2 (HKAS 128858, holotype; GZAAS 23–0591, isotype), ex-type living culture CGMCC, GZCC 23–0587. Ibid., GXZWY3.5 (GZAAS 23–0592, paratype), living culture GZCC 23–0588.
Notes: Phylogenetically, Helicosporium nanningense shares a sister relationship to H. multidentatum with high statistical support (94% ML/ 1.00 PP). Morphologically, H. nanningense differs from H. multidentatum in having different conidiogenous cells (tiny tooth-like protrusions vs. integrated multi-dentate protrusions). Additionally, H. nanningense differs from H. multidentatum in having shorter conidiophores (90–115 µm vs. 130–200 µm) and larger conidia (105–130 µm vs. 82–92 µm). Moreover, H. nanningense is similar to H. viridisporum in conidiophores, conidiogenous cells and conidial features [11], but the phylogenetic analysis result supports they are distinct species.
6. Discussion
Helicosporous fungi are filamentous fungi whose conidia curve through at least 180 degrees in one plane or three-dimensional space as they grow longer [2,3,9,10,11,27,28,29,30] Due to their ability to produce active secondary metabolites with distinct structures, these fungi have attracted scientists’ interest and become a popular study area. Numerous novel helicosporous taxa have recently been discovered in subtropical to tropical terrestrial and freshwater habitats [9,10,11,12,21,22,30,31,32]. In addition, a growing number of active secondary metabolites have been isolated from helicosporous fungi [33,34,35,36].
Helicomyces [37], Helicosporium [1] and Helicoma [38] are the three earliest described helicosporous genera. Based on morphological characteristics, researchers including Linder [2], Moore [39] and Goos [3,27,28,29] carried out systematic classification studies on these three genera. Tsui et al. [49] conducted a phylogenetic analysis of helicosporous fungi. They discovered that the species of Helicomyces, Helicosporium, and Helicoma did not cluster within their respective genus-level taxonomy units but instead interbred and dispersed within the family Tubeufiaceae. Kuo and Goh [41] also reported the chaotic phylogenetic relationships between these three genera. Lu et al. [10] reevaluated these three genera, redefined their generic concepts based on morphological and phylogenetic evidence, and provided recommendations for classifying and identifying helicosporous fungi. However, some taxa within these genera still require additional morphological and molecular data to resolve their taxonomic issues. For example, Boonmee et al. [7] combined Helicosporium sp. NBRC 9014 (as Tubeufia cerea NBRC 9014 in Tsui et al. [40] with H. vegetum based on phylogenetic analyses. However, Lu et al. [10] disagreed with this treatment as Helicosporium sp. NBRC 9014 did not cluster with other H. vegetum strains in the multi-gene phylogenetic tree. The taxonomic status of this strain remains unresolved due to insufficient morphological information [10].
Lu et al. [11] highlighted the challenge of taxonomic studies on helicosporous fungi due to their similar morphological characteristics. In this study, three new helicosporous fungi, namely Helicosporium liuzhouense, H. multidentatum and H. nanningense, were identified using morphological and phylogenetic analyses, with supporting evidence from the PHI test. Helicosporium liuzhouense and H. nanningense share similarities with H. sexuale and H. viridisporum in terms of conidiophores, conidiogenous cells and conidial features, respectively, while H. multidentatum is comparable to H. hainanense and H. vesicarium. Notably, they have distinct phylogenetic positions (Figure 1). These findings reinforce the significance of molecular data in precisely distinguishing helicosporous hyphomycetes.
A checklist of accepted Helicosporium species is provided in this study (Table 2). Nine species are found in freshwater habitats and 11 species in terrestrial habitats, including the newly described species in this study. H. sexuale exists in both freshwater and terrestrial habitats. Among them, 18 species are reported only in their helicosporous asexual morph, while three species, viz. H. flavum, H. sexuale and H. vegetum have asexual-sexual links that have been confirmed by molecular data. The taxonomic status of 15 species has been determined through phylogenetic analyses, while six species do not have any molecular data and require further research to determine their phylogenetic relationships [42,43,44,45]. A key to the species accepted in Helicosporium is provided as well.
Key to species of Helicosporium
| 1. Ascomata present | 2 |
| 1. Ascomata lacking | 4 |
| 2. Ascomata bright yellow to brown yellow, without setae | H. vegetum |
| 2. Ascomata bright yellow to brown yellow, with sparsely setae | 3 |
| 3. Ascomata globose to subglobose, 185–235 µm high, 116–214 µm diam. | H. sexuale |
| 3. Ascomata globose to subglobose, 320–350 µm high, 220–260 µm diam. | H. flavum |
| 4. Fresh colonies on decaying woody substrate are bright green | 5 |
| 4. Fresh colonies on decaying woody substrate are hyaline, gray, yellow, yellow green, or dark brown | 7 |
| 5. Conidiophores unbranched, with multi-dentate protrusions conidiogenous cells | H. multidentatum |
| 5. Conidiophores unbranched, with tiny tooth-like protrusions conidiogenous cells | 6 |
| 6. Conidiophores < 120 μm long | H. nanningense |
| 6. Conidiophores > 120 μm long | H. liuzhouense or H. viridisporum |
| 7. Fresh colonies on decaying woody substrate are yellow green | 8 |
| 7. Fresh colonies on decaying woody substrate are hyaline, gray, yellow, or dark brown | 14 |
| 8. Conidiophores with bladder-like protrusions conidiogenous cells | 9 |
| 8. Conidiophores with tiny tooth-like protrusions conidiogenous cells | 10 |
| 9. Conidiophores < 120 μm long | H. vesicarium |
| 9. Conidiophores > 120 μm long | H. hainanense |
| 10. Conidia are acropleurogenous | H. thailandense |
| 10. Conidia are pleurogenous and apical sterile in conidiophores | 11 |
| 11. Conidiophores < 180 μm long | 12 |
| 11. Conidiophores > 180 μm long | 13 |
| 12. Conidia 10–14 µm diam., 70–90 µm long | H. aquaticum |
| 12. Conidia 13–21 µm diam., 100–130 µm long | H. setiferum |
| 13. Conidiophores 184–257 µm long | H. flavidum |
| 13. Conidiophores 250–425 µm long | H. viridiflavum |
| 14. Fresh colonies on decaying woody substrate are yellow | 15 |
| 14. Fresh colonies on decaying woody substrate are hyaline, gray, or dark brown | 16 |
| 15. Conidiophores with bladder-like protrusions conidiogenous cells | H. flavisporum |
| 15. Conidiophores with tiny tooth-like protrusions conidiogenous cells | H. luteosporum |
| 16. Fresh colonies on decaying woody substrate are hyaline, branched conidiophores 200–300 µm long | H. albidum |
| 16. Fresh colonies on decaying woody substrate are gray, or dark brown | 17 |
| 17. Fresh colonies on decaying woody substrate are gray, conidiophores 350–400 µm long, with bladder-like protrusions conidiogenous cells | H. neesii |
| 17. Fresh colonies on decaying woody substrate are dark brown | 18 |
| 18. Conidiophores are unbranched with tooth-like protrusions conidiogenous cells | H. murinum |
| 18. Conidiophores are rarely branched | 19 |
| 19. Tooth- or bladder-like protrusions conidiogenous cells, conidia 6–9 µm diam. | H. decumbens |
| 19. Tooth-like protrusions conidiogenous cells, conidia 11.4–19 µm diam. | H. melghatianum |
Author Contributions
The specimens in this study were collected by X.-J.X., J.M. and Y.-Z.L. Morphological data were collected by X.-J.X. and J.M. Molecular data and phylogenetic analyses were performed by X.-J.X. and J.M. Writing-original were completed by X.-J.X., J.M. Review and editing were done by X.-J.X., J.M., L.-J.Z., N.-G.L., Y.-P.X., Z.-L.L. and Y.-Z.L.
Funding
This work was funded by Guizhou Provincial Key Technology R&D Program (grant No. Qian Ke He Zhi Cheng [2021] Generally 200), Guizhou Province high-level talent innovation and entrepreneurship merit funding project (No. 202104), Youth Science and Technology Talent Development Project from Guizhou Provincial Department of Education (QJHKYZ [2021]263) and the academic emerging project of the Guizhou Institute of Technology (GZLGXM-15).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All sequences generated in this study were submitted to GenBank database.
Acknowledgments
We would like to thank Dr. Shaun Pennycook (Manaaki Whenua Landcare Research, New Zealand) for advising on fungal nomenclature.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nees Von Esenbeck, C.D.G. System der Pilze und Schwämme. Würtzburg, 1817; 1–334. [Google Scholar]
- Linder, D.H. A monograph of the helicosporous fungi imperfecti. Ann. Mo. Bot. Gard. 1929, 16, 227–388. [Google Scholar] [CrossRef]
- Goos, R.D. On the anamorph genera Helicosporium and Drepanospora. Mycologia 1989, 81, 356–374. [Google Scholar] [CrossRef]
- Ho, W.H.; Hyde, K.D.; Hodgkiss, I.J. Seasonality and sequential occurrence of fungi on wood submerged in Tai Po Kau Forest Stream, Hong Kong. Fungal Divers. 2002, 10, 21–43. [Google Scholar]
- Pinruan, U.; Hyde, K.D.; Lumyong, S.; McKenzie, E.H.C.; Jones, E.B.G. Occurrence of fungi on tissues of the peat swamp palm Licuala longicalycata. Fungal Divers. 2007, 25, 157–173. [Google Scholar]
- Zhao, G.Z.; Liu, X.Z.; Wu, W.P. Helicosporous hyphomycetes from China. Fungal Divers. 2007, 26, 313–524. [Google Scholar]
- Boonmee, S.; Rossman, A.Y.; Liu, J.K.; Li, W.J.; Dai, D.Q.; Bhat, J.D.; Jones, E.B.G.; McKenzie, E.H.C.; Xu, J.C.; Hyde, K.D. Tubeufiales, ord. nov., integrating sexual and asexual generic names. Fungal Divers. 2014, 68, 239–298. [Google Scholar] [CrossRef]
- Boonmee, S.; Wanasinghe, D.N.; Calabon, M.S.; Huanraluek, N.; Chandrasiri, S.K.U.; Jones, G.E.B.; Rossi, W.; Leonardi, M.; Singh, S.K.; Rana, S.; et al. Fungal diversity notes 1387–1511: taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2021, 111, 1–335. [Google Scholar] [CrossRef]
- Lu, Y.Z.; Boonmee, S.; Bhat, D.J.; Hyde, K.D.; Kang, J.C. Helicosporium luteosporum sp. nov. and Acanthohelicospora aurea (Tubeufiaceae, Tubeufiales) from terrestrial habitats. Phytotaxa 2017, 319, 241–253. [Google Scholar] [CrossRef]
- Lu, Y.Z.; Liu, J.K.; Hyde, K.D.; Jeewon, R.; Kang, J.C.; Fan, C.; Boonmee, S.; Bhat, D.J.; Luo, Z.L.; Lin, C.G.; Eungwanichayapant, P.D. A taxonomic reassessment of Tubeufiales based on multi-locus phylogeny and morphology. Fungal Divers. 2018, 92, 131–344. [Google Scholar] [CrossRef]
- Lu, Y.Z.; Ma, J.; Xiao, X.J.; Zhang, L.J.; Xiao, Y.P.; Kang, J.C. Four new species and three new records of helicosporous hyphomycetes from China and their multi-gene phylogenies. Front. Microbiol. 2022, 13, 1053849. [Google Scholar] [CrossRef]
- Dong, W.; Wang, B.; Hyde, K.D.; McKenzie, E.H.; Raja, H.A.; Tanaka, K.; Abdel-Wahab, M.A.; Abdel-Aziz, F.A.; Doilom, M.; Phookamsak, R.; et al. Freshwater Dothideomycetes. Fungal Divers. 2020, 105, 319–575. [Google Scholar] [CrossRef]
- Index Fungorum. Available online: https://www.indexfungorum.org/Names/Names.asp (accessed on 20 May 2023).
- Brahmanage, R.S.; Lu, Y.Z.; Bhat, D.J.; Wanasinghe, D.N.; Yan, J.Y.; Hyde, K.D.; Boonmee, S. Phylogenetic investigations on freshwater fungi in Tubeufiaceae (Tubeufiales) reveals the new genus Dictyospora and new species Chlamydotubeufia aquatica and Helicosporium flavum. Mycosphere 2017, 8, 917–933. [Google Scholar] [CrossRef]
- Hardy, G.E.S.J.; Sivasithamparam, K. Antagonism of fungi and actinomycetes isolated from composted Eucalyptus bark to Phytophthora drechsleri in a steamed and non-steamed composted Eucalyptus bark-amended container medium. Soil Biol. Biochem. 1995, 27, 243–246. [Google Scholar] [CrossRef]
- Choi, H.J.; Lee, S.M.; Kim, S.H.; Kim, D.W.; Choi, Y.W.; Joo, W.H. A novel Helicosporium isolate and its antimicrobial and cytotoxic pigment. J. Microbiol. and Biotechn. 2012, 22, 1214–1217. [Google Scholar] [CrossRef]
- Chomnunti, P.; Hongsanan, S.; Hudson, B.A.; Tian, Q.; Peršoh, D.; Dhami, M.K.; Alias, A.S.; Xu, J.C.; Liu, X.Z.; Stadler, M.; Hyde, K.D. The sooty moulds. Fungal Divers. 2014, 66, 1–36. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97(1), 84–98. [Google Scholar] [CrossRef]
- Lu, Y.Z.; Boonmee, S.; Dai, D.Q.; Liu, J.K.; Hyde, K.D.; Bhat, D.J.; Ariyawansa, H.; Kang, J.C. Four new species of Tubeufia (Tubeufiaceae, Tubeufiales) from Thailand. Mycol. Prog. 2017, 16, 403–417. [Google Scholar] [CrossRef]
- Lu, Y.Z.; Boonmee, S.; Liu, J.K.; Hyde, K.D.; McKenzie, E.H.C.; Eungwanichayapant, P.D.; Kang, J.C. Multi-gene phylogenetic analyses reveals Neohelicosporium gen. nov. and five new species of helicosporous hyphomycetes from aquatic habitats. Mycol. Prog. 2018, 17, 631–646. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Ma, J.; Zhang, J.Y.; Xiao, X.J.; Xiao, Y.P.; Tang, X.; Boonmee, S.; Kang, J.C.; Lu, Y.Z. Multi-gene phylogenetic analyses revealed five new species and two new records of Distoseptisporales from China. J. Fungi 2022, 8. [Google Scholar] [CrossRef]
- Bruen, T.C.; Philippe, H.; Bryant, D. A simple and robust statistical test for detecting the presence of recombination. Genetics 2006, 172, 2665–2681. [Google Scholar] [CrossRef]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Goos, R.D. Some helicosporous fungi from Hawaii. Mycologia 1980, 72, 595–610. [Google Scholar] [CrossRef]
- Goos, R.D. A review of the anamorph genus Helicomyces. Mycologia 1985, 77, 606–618. [Google Scholar] [CrossRef]
- Goos, R.D. A review of the anamorph genus Helicoma. Mycologia 1986, 78, 744–761. [Google Scholar] [CrossRef]
- Luo, Z.L.; Bhat, D.J.; Jeewon, R.; Boonmee, S.; Bao, D.F.; Zhao, Y.C.; Chai, H.M.; Su, H.Y.; Su, X.J.; Hyde, K.D. Molecular phylogeny and morphological characterization of asexual fungi (Tubeufiaceae) from freshwater habitats in Yunnan, China. Cryptogamie Mycol. 2017, 38, 27–53. [Google Scholar] [CrossRef]
- Liu, J.K.; Lu, Y.Z.; Cheewangkoon, R.; To-Anun, C. Phylogeny and morphology of Helicotubeufia gen. nov, with three new species in Tubeufiaceae from aquatic habitats. Mycosphere 2018, 9, 495–509. [Google Scholar] [CrossRef]
- Tian, X.; Karunarathna, S.C.; Xu, R.; Lu, Y.; Suwannarach, N.; Mapook, A.; Bao, D.; Xu, J.; Tibpromma, S. Three new species, two new records and four new collections of Tubeufiaceae from Thailand and China. J. Fungi 2022, 8. [Google Scholar] [CrossRef]
- Zeng, X.; Qian, S.; Lu, Y.; Li, Y.; Chen, L.; Qian, Y.; He, Z.; Kang, J. A novel Nitrogen-containing Glyceride from fungal saprobe Tubeufia rubra reverses MDR of tumor cell lines to Doxorubicin. Rec. Nat. Prod. 2022, 16, 622–632. [Google Scholar] [CrossRef]
- Zhang, L.J.; Yang, M.F.; Ma, J.; Xiao, X.J.; Ma, X.Y.; Zheng, D.G.; Han, M.Y.; Xia, M.L.; Jayawardena, R.S.; Mapook, A.; et al. Neogrisphenol A, a potential ovarian cancer inhibitor from a new record fungus Neohelicosporium griseum. Metabolites 2023, 13, 435. [Google Scholar] [CrossRef]
- Zheng, W.; Han, L.; He, Z.J.; Kang, J.C. A new alkaloid derivative from the saprophytic fungus Neohelicomyces hyalosporus PF11-1. Nat. Prod. Res. 2023, 1–5. [Google Scholar] [CrossRef]
- Qian, S.Y.; Zeng, X.B.; Qian, Y.X.; Lu, Y.Z.; He, Z.J.; Kang, J.C. A saprophytic fungus Tubeufia rubra Produces novel rubracin D and E reversing multidrug resistance in cancer cells. J. Fungi 2023, 9, 309. [Google Scholar] [CrossRef]
- Link, H.F. Observationes in ordines plantarum naturales. Dissertatio I. Mag Ges Naturf Fr. Berl. 1809, 3, 3–42. [Google Scholar]
- Corda, A.K.J. Icones fungorum hucusque cognitorum, Vol 1. Prague: J.G. Calve, 1837, 1–32.
- Moore, R.T. Index to the Helicosporae. Mycologia 1955, 47, 90–103. [Google Scholar] [CrossRef]
- Tsui, C.K.; Sivichai, S.; Berbee, M.L. Molecular systematics of Helicoma, Helicomyces and Helicosporium and their teleomorphs inferred from rDNA sequences. Mycologia 2006, 98, 94–104. [Google Scholar] [CrossRef]
- Kuo, C.H.; Goh, T.K. Two new species of helicosporous hyphomycetes from Taiwan. Mycol. Prog. 2018, 17, 557–569. [Google Scholar] [CrossRef]
- Dharkar, N.; Subhedar, A.; Hande, D.; Shahezad, M.A. Two new fungal species from Vidarbha, India. J. Mycol. Plant Pathol. 2010, 40, 235–237. [Google Scholar]
- Grove, W.B. New or noteworthy fungi. III. J. Bot. Br. Foreign 1886, 24, 197–207. [Google Scholar]
- Hsieh, S.Y.; Goh, T.K.; Kuo, C.H. New species and records of Helicosporium sensu lato from Taiwan, with a reflection on current generic circumscription. Mycol. Prog. 2021, 20, 169–190. [Google Scholar] [CrossRef]
- Moore, R.T. Index to the Helicosporae: addenda. Mycologia 1957, 49, 580–587. [Google Scholar] [CrossRef]
Figure 1.
The phylogenetic tree generated by maximum likelihood (ML) analysis based on a combined LSU, ITS, tef1α, and rpb2 sequence data. ML and Bayesian posterior probabilities (PP) near the nodes are indicated as ML/PP. The Acanthostigma chiangmaiensis (MFLUCC 10–0125) and A. perpusillum (UAMH 7237) were used as outgroup taxa. Ex-type strains represented in bold. Newly generated sequences are represented in red.
Figure 1.
The phylogenetic tree generated by maximum likelihood (ML) analysis based on a combined LSU, ITS, tef1α, and rpb2 sequence data. ML and Bayesian posterior probabilities (PP) near the nodes are indicated as ML/PP. The Acanthostigma chiangmaiensis (MFLUCC 10–0125) and A. perpusillum (UAMH 7237) were used as outgroup taxa. Ex-type strains represented in bold. Newly generated sequences are represented in red.
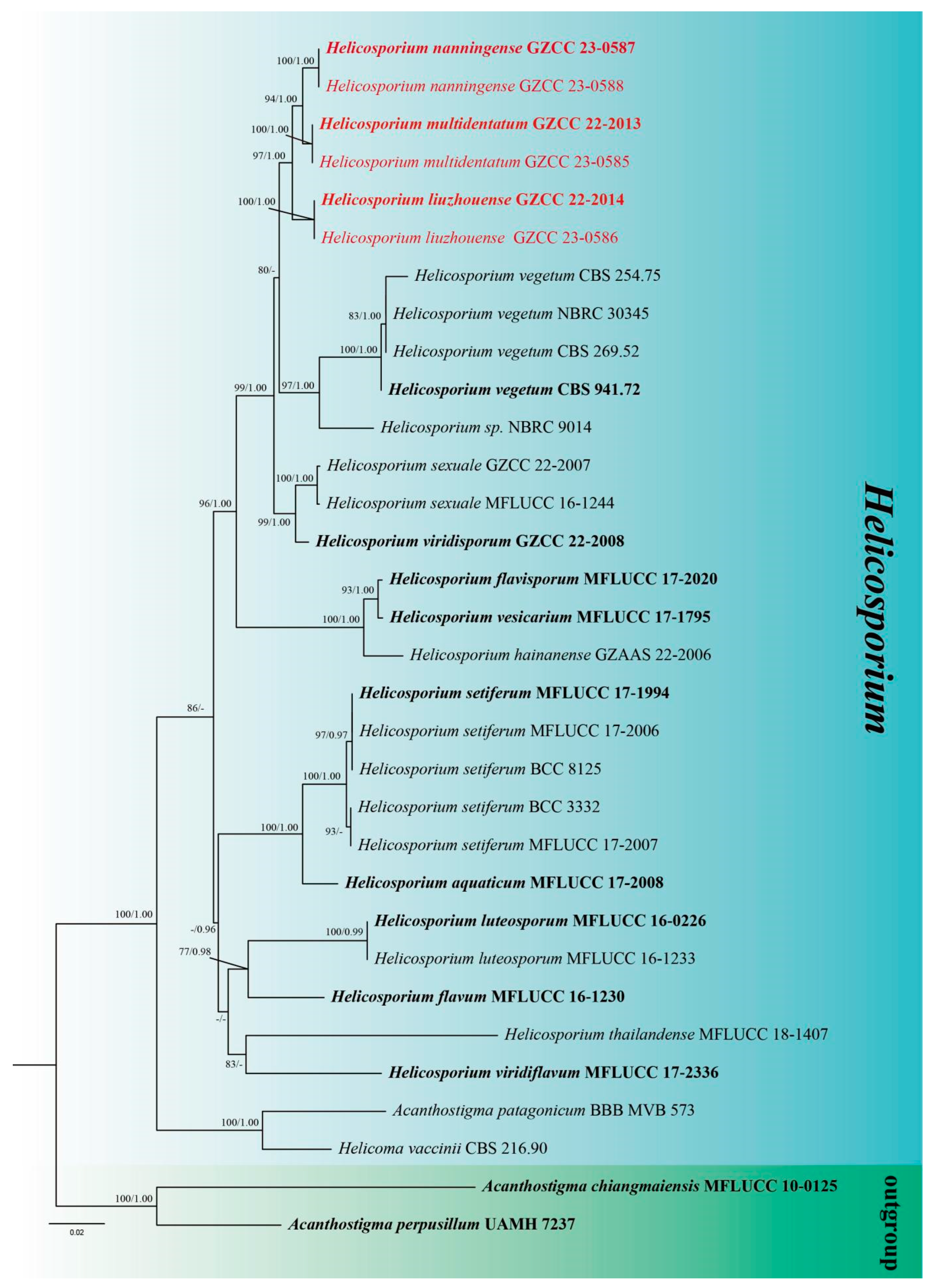
Figure 2.
Results of the PHI test of Helicosporium liuzhouense, H. multidentatum and H. nanningense with closely related species (combined LSU-ITS-tef1α-rpb2) using both LogDet transformation and splits decomposition. PHI test results (Φw) < 0.05 indicate significant recombination within the dataset. The new taxon is in red bold type. Our new species are indicated in red, type strains are marked with “T”.
Figure 2.
Results of the PHI test of Helicosporium liuzhouense, H. multidentatum and H. nanningense with closely related species (combined LSU-ITS-tef1α-rpb2) using both LogDet transformation and splits decomposition. PHI test results (Φw) < 0.05 indicate significant recombination within the dataset. The new taxon is in red bold type. Our new species are indicated in red, type strains are marked with “T”.
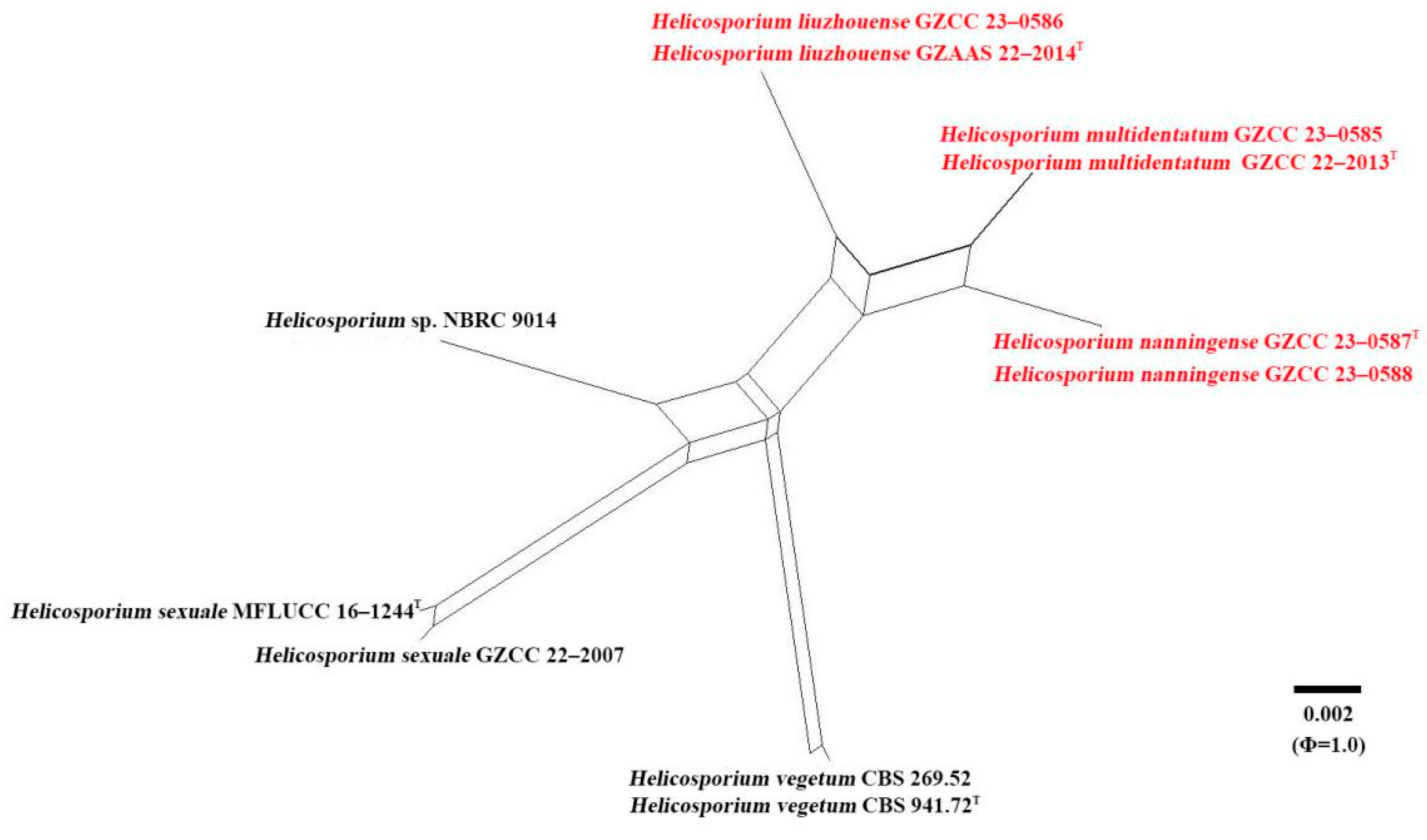
Figure 3.
Helicosporium liuzhouense (HKAS 125865, holotype). (a,b) Colonies on natural substrate. (c–f) Conidiophores. (g–i) Conidiogenous cells. (j) Germinating conidium. (k–p) Conidium. (q–r) Colonies on PDA from above and below. Scale bars: (c–f) 50 μm, (j) 20 μm, (g–i,k–p) 10 μm.
Figure 3.
Helicosporium liuzhouense (HKAS 125865, holotype). (a,b) Colonies on natural substrate. (c–f) Conidiophores. (g–i) Conidiogenous cells. (j) Germinating conidium. (k–p) Conidium. (q–r) Colonies on PDA from above and below. Scale bars: (c–f) 50 μm, (j) 20 μm, (g–i,k–p) 10 μm.

Figure 4.
Helicosporium multidentatum (HKAS 125856, holotype). (a,b) Colonies on natural substrate. (c–f) Conidiophores. (g–i) Conidiogenous cells. (j) Germinating conidium. (k–p) Conidium. (q–r) Colonies on PDA from above and below. Scale bars: (c–f) 50 μm, (j–k, n–p) 20 μm, (g–i,l–m) 10 μm.
Figure 4.
Helicosporium multidentatum (HKAS 125856, holotype). (a,b) Colonies on natural substrate. (c–f) Conidiophores. (g–i) Conidiogenous cells. (j) Germinating conidium. (k–p) Conidium. (q–r) Colonies on PDA from above and below. Scale bars: (c–f) 50 μm, (j–k, n–p) 20 μm, (g–i,l–m) 10 μm.

Figure 5.
Helicosporium nanningense (HKAS 128858, holotype). (a,b) Colonies on natural substrate. (c–g) Conidiophores. (h–j) Conidiogenous cells. (k–q) Conidium. (r,s) Colonies on PDA from above and below. Scale bars: (c–g) 20 μm, (h–q) 10 μm.
Figure 5.
Helicosporium nanningense (HKAS 128858, holotype). (a,b) Colonies on natural substrate. (c–g) Conidiophores. (h–j) Conidiogenous cells. (k–q) Conidium. (r,s) Colonies on PDA from above and below. Scale bars: (c–g) 20 μm, (h–q) 10 μm.


Table 1.
The taxa used in this study, along with their corresponding GenBank accession numbers for DNA sequences, are presented in the following table.
Table 1.
The taxa used in this study, along with their corresponding GenBank accession numbers for DNA sequences, are presented in the following table.
| Taxon | Strain | GenBank Accessions | |||
|---|---|---|---|---|---|
| ITS | LSU | tef1α | rpb2 | ||
| Acanthostigma chiangmaiensis | MFLUCC 10-0125T | JN865209 | JN865197 | KF301560 | - |
| Acanthostigma patagonicum | BBB MVB 573 | JN127358 | JN127359 | - | - |
| Acanthostigma perpusillum | UAMH 7237T | AY916492 | AY856892 | - | - |
| Helicoma vaccinii | CBS 216.90 | AY916486 | AY856879 | - | - |
| Helicosporium aquaticum | MFLUCC 17-2008T | MH558733 | MH558859 | MH550924 | MH551049 |
| Helicosporium flavisporum | MFLUCC 17-2020T | MH558734 | MH558860 | MH550925 | MH551050 |
| Helicosporium flavum | MFLUCC 16-1230T | KY873626 | KY873621 | KY873285 | - |
| Helicosporium hainanense | GZCC 22-2006T | OP508730 | OP508770 | OP698081 | OP698070 |
| Helicosporium liuzhouense | GZCC 22-2014T | OQ981394 | OQ981402 | OQ980476 | OQ980474 |
| Helicosporium liuzhouense | GZCC 23-0586 | OR066416 | OR066423 | OR058862 | OR058855 |
| Helicosporium luteosporum | MFLUCC 16-0226T | KY321324 | KY321327 | KY792601 | - |
| Helicosporium luteosporum | MFLUCC 16-1233 | - | KY873624 | - | - |
| Helicosporium multidentatum | GZCC 22-2013T | OQ981395 | OQ981403 | OQ980477 | OQ980475 |
| Helicosporium multidentatum | GZCC 23-0585 | OR066417 | OR066424 | OR058863 | OR058856 |
| Helicosporium nanningense | GZCC 22-2175T | OR066418 | OR066425 | OR058864 | OR058857 |
| Helicosporium nanningense | GZCC 23-0588 | OR066419 | OR066426 | OR058865 | OR058858 |
| Helicosporium setiferum | BCC 3332 | AY916490 | AY856907 | - | - |
| Helicosporium setiferum | BCC 8125 | AY916491 | - | - | - |
| Helicosporium setiferum | MFLUCC 17-1994T | MH558735 | MH558861 | MH550926 | MH551051 |
| Helicosporium setiferum | MFLUCC 17-2006 | MH558736 | MH558862 | MH550927 | MH551052 |
| Helicosporium setiferum | MFLUCC 17-2007 | MH558737 | MH558863 | MH550928 | MH551053 |
| Helicosporium sexuale | GZCC 22-2007 | OP508731 | OP508771 | OP698082 | OP698071 |
| Helicosporium sexuale | MFLUCC 16-1244T | MZ538503 | MZ538537 | MZ567082 | MZ567111 |
| Helicosporium sp. | NBRC 9014 | AY916489 | AY856903 | - | - |
| Helicosporium thailandense | MFLUCC 18-1407T | MT627698 | MN913718 | MT954371 | - |
| Helicosporium vegetum | CBS 254.75 | - | DQ470982 | DQ471105 | - |
| Helicosporium vegetum | CBS 269.52 | AY916487 | AY856893 | - | - |
| Helicosporium vegetum | CBS 941.72T | AY916488 | AY856883 | - | - |
| Helicosporium vegetum | NBRC 30345 | - | AY856896 | - | - |
| Helicosporium vesicarium | MFLUCC 17-1795T | MH558739 | MH558864 | MH550930 | MH551055 |
| Helicosporium viridiflavum | MFLUCC 17-2336T | MH558738 | - | MH550929 | MH551054 |
| Helicosporium viridisporum | GZCC 22-2008T | OP508736 | OP508776 | OP698087 | OP698076 |
Note: “T” represents the ex-type strain. Newly generated sequences are represented in bold. “-” indicates that no sequence data are available in GenBank.
Table 2.
Checklist of accepted Helicosporium species.
| Species | Habitats | Distribution | Molecular data | References |
|---|---|---|---|---|
| H. albidum | Terrestrial | Belgium, Britain (Birminghan) | No | Grove 1886 |
| H. aquaticum | Freshwater | Thailand | Yes | Lu et al. 2018b |
| H. decumbens | Terrestrial | Austria, Brazil | No | Linder 1929 |
| flavidum | Freshwater | China | No | Hsieh 2021 |
| H. flavisporum | Freshwater | Thailand | Yes | Lu et al. 2018b |
| H. flavum | Freshwater | Thailand | Yes | Brahamanage et al. (2017) |
| H. hainanense | Terrestrial | China | Yes | Lu et al. 2022 |
| H. liuzhouense | Freshwater | China | Yes | This study |
| H. luteosporum | Terrestrial | Thailand | Yes | Lu et al. 2017a |
| H. melghatianum | Terrestrial | India | No | Dharkar et al. 2010 |
| H. murinum | Terrestrial | Argentina, Austria, Brazil, Canada, China, Cuba, Malaysia, USA | No | Linder 1929; Goos 1989; Zhao et al. 2007 |
| H. multidentatum | Terrestrial | China | Yes | This study |
| H. nanningense | Terrestrial | China | Yes | This study |
| H. neesii | Terrestrial | USA | No | Moore 1957 |
| H. setiferum | Freshwater | Thailand | Yes | Lu et al. 2018b |
| H. sexuale | Freshwater, Terrestrial | China, Thailand | Yes | Boonmee et al. 2021; Lu et al. 2022 |
| H. thailandense | Freshwater | Thailand | Yes | Dong et al. 2020 |
| H. vegetum | Terrestrial | Worldwide | Yes | Boonmee et al. 2014 |
| H. vesicarium | Freshwater | Thailand | Yes | Lu et al. 2018b |
| H. viridiflavum | Terrestrial | Thailand | Yes | Lu et al. 2018b |
| viridisporum | Freshwater | China | Yes | Lu et al. 2022 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



