
Submitted:
21 June 2023
Posted:
21 June 2023
You are already at the latest version
Abstract
Mass mortality events and anthropogenic impacts were increasingly documented affecting Paramuricea clavata (Risso, 1826) during the last decades. These impacts enhance the settling of epibiontic organisms on the injured colonies. The epibiosis was studied by photographic sampling carried out on the granitic outcrops of the Tavolara Channel within the Tavolara-Punta Coda Cavallo Marine Protected Area (NE Sardinia) between 2017-2023 at 35-55 m. The number of colonies and percent surface involved in epibiosis, the specific richness of the epibiontic community and the temporal evolution of the phenomenon were investigated. Almost all the investigated gorgonians (93%) showed parts involved in epibiosis, with a high percentage of covering (one-third of the surface). Out of the 37 epibiontic species recorded, the most recurrent described an ecological succession dominated by hydrozoans, sponges, bryozoans, serpulids and the parasitic soft coral Alcyomiun coralloides. Nevertheless, single colonies studied through time revealed the unpredictability of the colonization process. The peculiar habitat of a high naturalistic value of the granitic outcrops hosting the P. clavata forests demonstrates a widespread suffering condition, supported both by environmental and anthropogenic sources of stress. Such considerations make it necessary to review the current zonation of the area, where the actual vulnerability and usability evaluations are based on obsolete information.
Keywords:
Gorgonians
; benthic assemblages
; fishing impact
; global change
; conservation
; Mediterranean Sea
1. Introduction
The purple gorgonian Paramuricea clavata (Risso, 1826) is an endemic Mediterranean species [1], considered “vulnerable” in the Red List of the International Union for Conservation of Nature [2] because of its large size, branched shape and limited flexibility, that render it highly susceptible to a wide range of natural and human impacts [3,4,5,6]. A pool of evidence regarding the reduction of some Mediterranean populations in the last three decades has been addressed to a mix of consequences due to the increase of positive thermal anomalies with summer diseases, development of filamentous algae, mucilage and invasive invertebrate epibionts [7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22].
Moreover, it has been recognized that P. clavata forests are highly sensitive to direct or indirect anthropogenic impacts (boat anchoring, recreational and commercial fishing and SCUBA diving). In 1997, Bavestrello et al. [4] described the multiple damages inflicted on these gorgonians by fishing activity: the lost lines in contact with the colonies scrape the coenenchyme and these abrasions create the suitable conditions for the settlement of numerous epibiontic organisms that weigh down the branches until the total or partial breaking of the colony under the action of the currents. This idea has been shared by several Authors who have employed the number of colonies involved in epibiosis as an index of the stress of the population due to fishing [2,5,6,23,24]. In fact, Linares et al. [25] stated that, in the absence of extraordinary events, the extent of colony injury and proportion of affected gorgonian colonies remain low over time and therefore, a substantial increase in these parameters may serve as a good indicator of recent disturbances.
The Tavolara - Punta Coda Cavallo Marine Protected Area (40.8830 N, 9.7020 E, Sardinia, Italy) hosts a large population of P. clavata widely studied in recent years. Sea fan forests settle both on Tavolara Island’s limestones and Tavolara Channel’s granitic outcrops [26,27]. During the last decades, several surveys were conducted to evaluate the health status of these forests after diseases/mortality events occurred since the 2000s [14,16,21,28]. Unfortunately, this pool of data regards shallow sites (up to 35 m depth) under the medium-high protection regime (A-B Zones) and does not include gorgonians settled on the deeper granitic shoals at a depth ranging from 35 to 55 m [27], characterized by low protection level (C Zone) and prone to intense fishing activity [29,30].
The aim of this paper is to obtain data about the level of epibiosis of the sea fans present in this charismatic MPA, also in relation to the different regimes of protection. Moreover, a particular focus was provided on the diversity, structure, and development of the epibiontic community. In fact, despite the wide use of epibiosis as an index of injuries, few data are available about the description of the community.
2. Materials and Methods
The study area involved granitic outcrops emerging from a detrital plain located in the middle of the Tavolara Channel (Zone C, partial protection), between the Tavolara and Molara Islands, in the depth range 35-55m (Figure 1).
During SCUBA dives performed from 2017 to 2023, 10 granitic sites (out of 57 where Paramuricea clavata is present), were selected on the base of the gorgonian abundance. For each shoal, on average, 40 photos were obtained applying the multi-zoom photographic approach [31], to evaluate the occurrence of damages and epibionts on gorgonians. Images were taken using a Sony A6000 camera (24 megapixels, two Inon Z330 flashes, colour temperature 5000 K) with Sony 16–50 lens (focal length 19 mm), Nauticam WW1 wet wide lens (130° rectilinear field angle), and a Sea & Sea MDX-A6000 underwater case with a flat porthole.
Photos were post-processed on Adobe Photoshop©, while formal analysis was performed on ImageJ Software [32]. Out of 410 photographs, 582 colonies of P. clavata with a suitable perspective were examined to evaluate the presence and the diversity of the epibiontic assemblage, the area of the gorgonian involved in the epibiosis and the presence of entangled abandoned lines. We have considered epibionted only colonies with more than 5% of covered surface.
Epibiontic organisms were identified and classified at the lower taxonomic level; when specific or generic identification was not possible, other taxonomic and morphological units (OTUs) were adopted. Only taxa with a recurrence higher than 10% employed the formal analysis.
Moreover, thanks to the wide photographic archive of one of us (1986-2023) (ET), we had the opportunity to analyze images of the same colonies taken across different spans of time. These data allowed to reconstruct the temporal evolution of the epibiosis process.
3. Results
3.1. Paramuricea clavata of the Tavolara Channel granitic shoals
The granitic outcrops emerging from the detritic bottom of the Tavolara Channel are characterized by a modest development of the basal coralline algal layer that only occasionally forms the typical coralligenous bioherms. The density of the Paramuricea clavata varied from 5 to 25 colonies m-2. Moreover, the macrobenthic community is mainly represented by other gorgonians (Eunicella cavolini (Koch, 1887), E. singularis (Esper, 1791), E. verrucosa (Pallas, 1776), erected sponges (Axinella polypoides Schmidt, 1862, Spongia (Spongia) officinalis Linnaeus, 1759, Sarcotragus foetidus Schmidt, 1862) and branched bryozoans (Figure 2a,b).
During the period of observations, serious damages to colonies resulting from heat waves, with necrosis and loss of coenenchyme documented in 2008 and 2018 (Figure 2c,d); other less intense episodes were recorded in 2015, 1017, 2019 and 2020. The P. clavata spawning (Figure 2e) has been always recorded between 2009 and 2022, with the exceptions of 2010, 2018 and 2019. Recruits, as small unbranched colonies composed of less than ten polyps (Figure 2f), were observed in 2017, 2018, and 2020.
3.2. Epibiosis
Of 582 studied colonies of P. clavata, 541 presented some degree of epibiosis (93.0%) and only 41 (7.0%) were healthy (Figure 3a–c); 455 (78.2%) showed epibiosis on more than 5% of the colony’s surface, 15 (2.6%) were recorded completely dead in standing position and 31 (4.6%) specimens were found entangled in lost fishing gears (Figure 3d) and 58 were uprooted (Figure 3e). On average, the epibiosis affected 27.3 ± 1.2% of the colony surface.
From the diversity point of view, the epibiontic assemblages on the gorgonians of the Tavolara MPA accounted for 37 species (or OTUs), of which, 5 were vagile (Table 1).
The most diversified group was sponges with 11 species (or OTUs), followed by bryozoans and ascidians, respectively with 7 and 5 species (or OTUs). About 13% of the observed colonies showed only one epibiontic species (OTUs), about 45% hosted 2-3 species, while 23 % was found with 4-5 species (OTUs). The highest value of species (OTUs) per colony was 16, recorded on one colony. The most common species/OTUs (Figure 4) resulted in the ubiquitarians hydrozoans recorded in 91.9% of the epibionted colonies, 4 sponges (Dysidea fragilis (Montagu, 1814) (43.8%), Crella (Crella) elegans Schmidt, 1862) (10.4%), D. perfistulata Pulitzer-Finali & Pronzato, 1980 (10.2%), Pleraplysilla spinifera (Schulze, 1879) (9.9%)), 3 bryozoans (Adeonella calveti Canu & Bassler, 1930 (52.3%), Turbicellepora avicularis (Hincks, 1860) (26.9%), Flustra sp. (9.6%)), the parasitic anthozoan Alcyonium coralloides (Pallas, 1766) (22.9%), the serpulids Filograna/Salmacina spp. (33.3%) (Figure 5).
These species produced different growth patterns on the gorgonian branches (Figure 5). The observed hydrozoan generally did not make large colonies but a carpet of small creeping colonies involving the entire naked skeleton. A. calveti, T. avicularis and Filograna/Salmacina spp. were able to produce subspherical large colonies that develop starting from a small portion of the free skeleton. Finally, sponges and A. coralloides grew covering entire branches of the gorgonians.
Single epibiontic species adopted a different pattern of colonization in relation to the richness of the assemblage. Hydrozoans were always recorded as pioneers and occurred in the epibiontic community, independently of its richness. The group of bushy species (bryozoans and serpulids) followed hydrozoans and their occurrence quickly increased according to the number of species present in the community. A similar trend was shown by a pair of sponges (D. fragilis and C. elegans) that grew covering a wide portion of the gorgonian’s branches. Finally, other species such as D. perfistulata, P. spinifera and A. coralloides needed more than 4 species or OTUs of the assemblage for their settling (Figure 6).
During the colonization process, new species were added to the community that became more and more complex without substitution (Figure 7a): in fact, the average percentage of the gorgonian surface involved in the epibiosis colonies increased according to the diversity of the epibiontic community (Figure 7b).
3.3. Observations on single gorgonians through time
Observing single colonies through time (months, seasons and years), it was possible to describe the development of the epibiosis on P. clavata, which showed great variability in mode and time even if some general patterns can be described.
When the lesions of the coenenchyme originate from thermal stress the fate of the epibiosis depends on the extension of the denudated scleraxis. In fact, when the involved surface was not too extended, the coenenchyme was able to re-grow on the naked skeleton, even if already covered by the hydrozoan carpet. Two colonies photographed in 2020 showed a complete recovery in two years (Figure 8a–d). When the lesions resulted too wide, the recovery was not possible and the epibiosis developed very quickly involving the entire colony (Figure 8e–f).
In some rare cases, the epibiontic community did not increase its complexity and branches remained covered by hydrozoans for several years. A colony in 2018 showed several naked portions of the skeleton partially covered by hydrozoans (18.9% of the colony surface); two years after (2020), the epibionted surface decreased to 9.1% thanks to a re-growth of coenenchyme on the involved branches. After four years (2023), the colony surface showed again a modest increase of epibiosis (19.3%) (Figure 8g–j).
Quick colonization was also observed when the epibiosis was due to the scraping of entangled lines. A colony photographed in March 2023 entangled by a lost line already colonized by hydrozoans and A. calveti showed a very fast development of the epibiontic community: in 47 days (May 2023) it included also D. fragilis, Terpios fugax Duchassaing & Michelotti, 1864, serpulids, Beania magellanica (Busk, 1852) with a covered surface shifting from 15.7% to 24.4% (Figure 9a,b).
Another colony recorded entangled in April 2018 by a line showed an epibiontic community composed by A. calveti, T. avicularis and Filograna/Salmacina spp. In the following December, the epibiotic community progressively developed in size without increasing the number of species or OTUs. In particular, in this period Filograna/Salmacina spp. appeared drastically reduced. The comparison with an image obtained four years after (October 2022) showed the settlement of the sponge D. fragilis and a shift in the involved surface from 22.73% to 50.74% (Figure 9c–e).
A colony during March 2018 was recorded with some nude branches, partially covered by hydroids and A. calveti; at the end of the year (November/December 2018), naked parts were covered by a massive development of Filograna/Salmacina spp. (Figure 9f–h). In some cases, when the scraping of the lost line is blocked by the growth of the carbonatic epibionts, the colonies are able, after several years, to recover damaged portions (Figure 9i,j). The observation of one dead extirpated colony (line still in situ) showed a long persistence of the gorgonin skeleton (five years) that became heavily colonized by sponges and encrusting coralline algae (Figure 9k–m).
4. Discussion
The first impressive evidence emerging from the study of epibionts on the Paramuricea clavata forests in Tavolara MPA was the high percentage of involved colonies: more than 70% of the studied specimens showed, on average, one-third of the surface covered by epibionts. For comparison, a study conducted in the Medes Islands (Catalan Sea) indicated that between 10% and 33% of the colonies of unprotected populations were partially colonized by epibionts, whereas only from 4% to 10% of the populations in the marine protected area were affected [33]. Inside the Portofino MPA (Ligurian Sea), 51% of colonies showed epibionts [19]; along the Eastern Adriatic coasts, less than 10% of the observed colonies presented denuded branches or epibiosis [34]. Very probably, the present data indicate that the Tavolara MPA is the Mediterranean area with the highest recorded values.
It was already stated that the epibiosis origins from the loss of coenenchyme integrity that leaves free portions of the gorgonin skeleton on which organisms can settle. Gorgonian diseases and fishing impact are considered the main causes of this phenomenon involving P. clavata [3,10,25,35]. In the area of Tavolara MPA, diseases/mass mortalities occurred at least since 2001 [28] and other episodes followed in 2008 [16], 2018 [14] and 2019-20 [21]. Recent mortality events resulted increasingly aggressive, as previous anomalies did not hit populations at 35−40 m depth [25,36,37]. These phenomena had a wide impact on the populations: for example, during the 2008 event, about 100% of the colonies living between 20 and 30 m depth were damaged, and this value decreased to 40-60% under 35m depth [16]. Currently, 15 years later in the area then considered, no colony of P. clavata can be found at depths of less than 29 m.
Regarding the fishing impact, demersal activities represent one of the major causes of gorgonian mortality, determining entanglements, partial breakages, and detachment [19,24,38]. The persistent abrasion of the lost lines may cause wounds and infections and lead to extended epibiosis and/or necrosis [4,5,39,40,41,42]. Artisanal fishing activity is particularly intense within the Tavolara Channel, an area under low protection regime, as demonstrated by the presence of lost fishing gears in 81 of 82 visited sites [30] and the high percentage of impacted erect organisms and marine litter [27,29].
Previous evaluation of the environmental quality of the Tavolara MPA [43] assigned low values to the Tavolara Channel because of the absence of a biogenic secondary substrate, limited by the lithological nature of the bedrock [26]. Considering the data presented here, the same priority habitat value of typical coralligenous reefs should be assigned to this area.
Although it is not easy to discriminate the kind of causative event by the observation of the epibiont colonization, some hints can be obtained. Generally, the diseases due to heat waves are restricted in time determining acute stress, while damages related to entangled lines work over a long span of time, producing chronic stress. In the first case, the fate of epibiosis depends on the percent of involved gorgonian’ surface [44]: when the affected portions are small, the coenenchyme is able to re-grow on the naked skeleton, plausibly quickly, despite the presence of pioneers epibionts (hydroids) [25,44,45,46,47]. Fast tissue regeneration in about 1 month was observed after the mass mortality event in the Portofino MPA in 2008 in case of minor injuries [48]. Studies on soft corals [49] demonstrated that they can act recovery mechanisms to quickly heal minor and momentary injuries and prevent the settlement of pioneer epibiont species such as hydroids on lesioned surfaces [50]. Recolonization of necrotic tissue might be faster and less expensive in terms of energy consumption than the production of new colony branches or tissue, and therefore influenced mainly by competition for space with epibionts. If the lesions involve a large portion of the denudated skeleton, the recovery is not sufficient to contrast the settling and development of calcified organisms that make permanent the epibiosis.
On the other hand, the continuous stress produced by the friction of lines, also if applied on a small coenenchyme surface, completely prevents the tissue recovery, favoring the development of complex epibiotic assemblages characterized by several calcified organisms (bryozoans, serpulids), sponges and Alcyonium coralloides. In fact, a chronic injury allows a permanent colonization phase of a diversified epibiont community [4,51,52].
Although several authors have recorded the epibiosis process involving P. clavata, less attention was dedicated to the diversity of epibiontic community: during this study, the 37 species (OTUs) found demonstrated a flourishing situation. The recorded assemblage is similar to that previously observed in the Ligurian Sea [4]. No species are strictly related to gorgonians, although the sponge Pleraplysilla spinifera and the bivalve Pteria hirundo (Linneaus, 1758) were frequently recorded as acrophilic on P. clavata [53,54,55,56]. The soft coral A. coralloides, a typical parasite of many gorgonians, grows by fouling (‘runner’ type) the apical axis of colonies [57,58]. A. coralloides exploit the stressed conditions of its host due to reproductive window and oligotrophy of water column during summertime, synchronizing its reproduction cycle with those of P. clavata [59]. Indeed, the emission of sexual products by A. coralloides has been documented in the Tavolara MPA area within the middle of May (Figure 2g) with the settling of recruits a couple of weeks after (ET, unpublished) simultaneously with the spawning of P. clavata.
A study conducted in Medes MPA, suggested that the epibiosis of this species was caused by the intense diving activity and not directly by fishing impact; this is not the situation of the Tavolara Channel, where diving activity is negligible and sporadically limited to 2 sites, while, on the contrary, fishing is widely performed [30]. Another example could be the appearance of the opportunistic tube-dwelling polychaetes Filograna/Salmacina spp. particularly after summertime [60,61].
Lacking the possibility to observe a temporal pattern of colonization, we have related the presence of each epibiont with the total diversity of the epibiontic community. Our observations clearly indicate that when the assemblage was made by only one species this was generally represented by pioneer hydrozoan turf (72%) and more rarely by bryozoans (9%) and sponges (4%). At this stage, the parasitic anthozoan A. coralloides was never found. When the assemblage increased in complexity, the species already present continued to be recorded with the progressive addition of other taxa following this order: hydrozoans, bryozoans, serpulids, sponges, and A. coralloides. Considering the gorgonians with more than 7 epibiontic species/OTUs, this anthozoan was present in more than 90% of them. In general, in a hypothetical model of community development, new species are added without substitution of the previous ones. The general scenario seems to put in evidence that the epibiontic species cooperate to maintain their occupation on the branches instead of competing. Probably, the only observed competition is between the epibiontic community and the coenenchyme of the gorgonian. A similar pattern of cooperation was already observed in sponge populations living in extremely competitive assemblages [62].
Another problem regards the age estimation of epibiontic assemblage. Some data are available in literature for the first phases after mass mortality. Perez et al. (2000) [9] recorded micro-organisms colonization after a few days while hydroids, followed by bryozoans and serpulids were observed some weeks after. This evolution was confirmed by other Authors [4,13,16,63]. Our data, obtained on single colonies photographed in different years indicate that the epibiontic assemblage persists for several years. According to Wahl [64], epibiont species commonly grow fast. This is certainly true for pioneer species (filamentous green and ⁄ or red algae and ⁄ or hydrozoans) that colonize wide stripped portions of the colonies in a few weeks [4,13,16]. Enrichetti et al. [38] studied the evolution of epibiosis on lost nylon lines: after 4 years, the diversity of the recorded community was very similar to those found on P. clavata, but only after a couple of years large colonies of calcified bryozoans were found. These data suggested a possibility to obtain a rough age of epibionts of the gorgonians as indicated by the temporal sequence of images of the single colonies. Very likely, the epibionts living on the gorgonians may persist for a very long time cooperating to maintain the substrate occupation.
These results highlight how the development of epibiosis appears with a partially stochastic phenomenon. The understanding of the dynamic of the processes requires continuous monitoring techniques, on a consistent number of colonies. The possibility of obtaining images over long periods of time requires a high effort in terms of resources but on the other hand, can allow the acquisition of fundamental information for the conservation of habitats so important and fragile.
Author Contributions
Conceptualization, G.B., M.C. and E.T.; methodology, M.C. and E.T.; validation, M.C., E.T.; formal analysis, M.C.; I.A.; investigation, E.T., P.P; resources, E.T., P.P.; data curation, E.T., I.A.; writing—original draft preparation, M.C., E.T., G.B..; writing—review and editing, M.C., E.T., I.A., and G.B.; visualization, M.C. and E.T.; supervision, G.B.; project administration, G.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received external fund by the National Biodiversity Future Center.
Data Availability Statement
The data collected during and/or analysed for the present study are available from the corresponding author upon request.
Acknowledgments
The authors would like to thank the Tavolara MPA management for the permission to use the GIS environment and for the possibility to independently develop the underwater surveys. The authors would like to thank the “Slow dive” team for its support during the diving activities.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Coelho, M.A. G. , Pearson, G.A., Boavida, J.R.H., Paulo, D., Aurelle, D., Arnaud-Haond, S., Gómez-Gras, D., Bensoussan, N., López-Sendino, P., Cerrano, C., Kipson, S., Bakran-Petricioli, T., Ferretti, E., Linares, C., Garrabou, J., Serrão, E.A., Ledoux, J.-B. (2023). Not out of the Mediterranean: Atlantic populations of the gorgonian Paramuricea clavata are a separate sister species under further lineage diversification. Ecology and Evolution 2023, 13, e9740. [Google Scholar] [CrossRef] [PubMed]
- Otero, M.M., Numa, C., Bo, M., Orejas, C., Garrabou, J., Cerrano, C., KružicÅL, P., Antoniadou, C., Aguilar, R., Kipson, S., Linares, C., Terr.n-Sigler, A., Brossard, J., Kersting, D., Casado-Amez.a, P., Garc.a, S., Goffredo, S., Ocana, O., Caroselli, E., Maldonado, M., Bavestrello, G., Cattaneo-Vietti, R. and .zalp, B. (2017). Overview of the Conservation Status of Mediterranean Anthozoans; IUCN: Málaga, Spain, 2017, pp. 1–73. Available online: http://hdl.handle.net/10508/11253ISBN 978-2-8317-1845-3. [CrossRef]
- Harmelin, J.G. , Marinopoulos, J. Population structure and partial mortality of the gorgonian Paramuricea clavata (Risso) in the north-western Mediterranean (France, Port-Cros Island). Marine life 1994, 4, 5–13. [Google Scholar]
- Bavestrello, G., Cerrano, C., Zanzi, D., Cattaneo-Vietti, R. Damage by fishing activities to the Gorgonian coral Paramuricea clavata in the Ligurian Sea. Aquatic Conservation: Marine and Freshwater Ecosystems 1997, 7, 253-262.
- Bo, M. , Bava, S., Canese, S., Angiolillo, M., Cattaneo-Vietti, R., Bavestrello, G. Fishing impact on deep Mediterranean rocky habitats as revealed by ROV investigation. Biol. Conserv. 2014, 171, 167–176. [Google Scholar] [CrossRef]
- Angiolillo, M.; di Lorenzo, B.; Farcomeni, A.; Bo, M.; Bavestrello, G.; Santangelo, G.; Cau, A.; Mastascusa, V.; Cau, A.; Sacco, F.; et al. Distribution and assessment of marine debris in the deep Tyrrhenian Sea (NW Mediterranean Sea, Italy). Mar. Pollut. Bull. 2015, 92, 149–159. [Google Scholar] [CrossRef]
- Bavestrello, G. , Bertone, S., Cattaneo-Vietti, R., Cerrano, C., Gaino, E., Zanzi, D. Mass mortality of Paramuricea clavata (Anthozoa, Cnidaria) on Portofino Promontory cliffs, Ligurian Sea, Mediterranean Sea. Mar. Life 1994, 4, 15–19. [Google Scholar]
- Cerrano, C. , Bavestrello, G., Bianchi, C. N., Cattaneo-Vietti, R. Bava, S. Morganti, C. Morri, C., Picco, P. Sara, G., Schiaparelli, S., Siccardi, A., Sponga, F. A catastrophic mass-mortality episode of gorgonians and other organisms in the Ligurian Sea (NW Mediterranean), summer. Ecol. Letters 2000, 3, 284–293. [Google Scholar] [CrossRef]
- Perez, T. , Garrabou, J., Sartoretto, S., Harmelin, J.G., Francour, P., Vacelet, J. Massive mortality of marine invertebrates: an unprecedented event in northwestern Mediterranean. Comptes rendus de l'Academie des sciences. Serie III, Sciences de la vie 2000, 323, 853–865. [Google Scholar] [CrossRef]
- Coma, R. , Pola, E., Ribes, M., Zabala, M. Long-term assessment of temperate octocoral mortality patterns, protected vs. unprotected areas. Ecological Applications 2004, 14, 1466–1478. [Google Scholar] [CrossRef]
- Coma R, Ribes M, Serrano E, Jiménez E, Salat J, Pascual J (2009) Global warming-enhanced stratification and mass mortality events in the Mediterranean. Proc Natl Acad Sci USA 106:6176–6181. [CrossRef]
- Schiaparelli, S. , Castellano, M., Povero, P., Sartoni, G., Cattaneo-Vietti, R. A benthic mucilage event in North-Western Mediterranean Sea and its possible relationships with the summer 2003 European heatwave: short term effects on littoral rocky assemblages. Mar. Ecol. 2007, 28, 341–353. [Google Scholar] [CrossRef]
- Garrabou, J. , Coma, R., Bensoussan, N., Bally, M., Chevaldonne, P. et al. Mass mortality in Northwestern Mediterranean rocky benthic communities: effects of the 2003 heat wave. Global Change Biology 2009, 15, 1090–1103. [Google Scholar] [CrossRef]
- Garrabou, J. , Gómez-Gras, D., Ledoux, J. B., Linares, C., Bensoussan, N., López-Sendino, P., et al. Collaborative database to track mass mortality events in the Mediterranean Sea. Front. Mar. Sci. 2019, 707. [Google Scholar] [CrossRef]
- Garrabou, J. , Gómez-Gras, D., Medrano, A., Cerrano, C., Ponti, M., Schlegel, R., et al. Marine heatwaves drive recurrent mass mortalities in the Mediterranean Sea. Global Change Biology 2022, 28, 5708–5725. [Google Scholar] [CrossRef]
- Huete-Stauffer, C. , Vielmini, I., Palma, M., Navone, A., Panzalis, P., Vezzulli, L., Misic, C., Cerrano, C. Paramuricea clavata (Anthozoa, Octocorallia) loss in the Marine Protected Area of Tavolara (Sardinia, Italy) due to a mass mortality event. Mar. Ecol. 2011, 32, 107–116. [Google Scholar] [CrossRef]
- Teixido, N. , Casas, E., Cebrian, E., Linares, C., Garrabou, J. Impacts on coralligenous outcrop biodiversity of a dramatic coastal storm. PloS one 2013, 8, e53742. [Google Scholar] [CrossRef] [PubMed]
- Turicchia, E. , Abbiati, M., Sweet, M., Ponti, M. Mass mortality hits gorgonian forests at Montecristo Island. Diseases of Aquatic Organisms 2018, 131, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Betti, F. , Bavestrello, G., Bo, M., Ravanetti, G., Enrichetti, F., Coppari, M., Cappanera, V., Venturini, S., Cattaneo-Vietti, R. Evidences of fishing impact on the coastal gorgonian forests inside the Portofino MPA (NW Mediterranean Sea). Oc. Coast. Manag. 2020, 187, 105105. [Google Scholar] [CrossRef]
- Casoli, E. , Ventura, D., Mancini, G., Cardone, S., Farina, F., Donnini, L., Pace, D.S., Shaul, R., Belluscio, A., Ardizzone, G. Rehabilitation of Mediterranean animal forests using gorgonians from fisheries by-catch. Restor. Ecol. 2022, 30, e13465. [Google Scholar] [CrossRef]
- Chimienti, G. , De Padova, D., Adamo, M., Mossa, M., Bottalico, A., Lisco, A., Ungaro, N., Mastrototaro, F. Effects of global warming on Mediterranean coral forests. Sci. Rep. 2021, 11, 20703. [Google Scholar] [CrossRef]
- Chimienti, G. , Maiorca, M., Digenis, M., Poursanidis, D. Conservation status of upper-mesophotic octocoral habitats at Sporades Archipelago (Aegean Sea). Mar. Pollut. Bull. 2023, 190, 114868. [Google Scholar] [CrossRef]
- Angiolillo, M. , Fortibuoni, T. Impacts of marine litter on Mediterranean reef systems: from shallow to deep waters. Front. Mar. Sci. 2020, 7, 581966. [Google Scholar] [CrossRef]
- Sini, M. , Kipson, S., Linares, C., Koutsoubas, D., Garrabou, J. The yellow gorgonian Eunicella cavolini: demography and disturbance levels across the Mediterranean Sea. PloS one 2015, 10, e0126253. [Google Scholar] [CrossRef]
- Linares, C. , Coma, R., Diaz, D., Zabala, M., Hereu, B., Dantart, L. Immediate and delayed effects of a mass mortality event on gorgonian population dynamics and benthic community structure in the NW Mediterranean Sea. Mar. Ecol. Prog. Ser. 2005, 305, 127–137. [Google Scholar] [CrossRef]
- Canessa, M. , Bavestrello, G., Bo, M., Trainito, E., Panzalis, P., Navone, A., Caragnano, A., Betti, F., Cattaneo-Vietti, R. Coralligenous assemblages differ between limestone and granite: a case study at the Tavolara-Punta Coda Cavallo Marine Protected Area (NE Sardinia, Mediterranean Sea). Reg. Stud. Mar. Sci. 2020, 35, 101159. [Google Scholar] [CrossRef]
- Canessa, M. , Bavestrello, G., Trainito, E., Bianchi, C. N., Morri, C., Navone, A., Cattaneo-Vietti, R. (2021). A large and erected sponge assemblage on granite outcrops in a Mediterranean Marine Protected Area (NE Sardinia). Reg. Stud. Mar. Sci. 2021, 44, 101734. [Google Scholar] [CrossRef]
- Calvisi, G. , Trainito, E., Pais, M., Franci, G., Schiaparelli, S. Prima segnalazione di un episodio di mortalità di Gorgonacei lungo la costa dell’isola di Tavolara (Sardegna settentrionale). Biol. Mar. Mediterr. 2003, 10, 506–508. [Google Scholar]
- Canessa, M. , Bavestrello, G. Valutazione sperimentale dello stato di conservazione del coralligeno dell'AMP Tavolara - Punta Coda Cavallo nell’ambito della Strategia Marina. Relazione finale. Genova, 28 Novembre 2018.
- Amedeo, I. , Canessa, M., Panzalis, P., Trainito, E. The need for change in a changing world: fishing impact within the Tavolara-Punta Coda Cavallo MPA. In 4th Mediterranean Symposium on the Conservation of the Coralligenous and other calcareous bio-concretions Genoa, 2022, (p. 11).
- Pititto, F.; Trainito, E.; Macic, V.; Rais, C.; Torchia, G. The resolution in benthic carthography: A detailed mapping technique and a multiscale GIS approach with applications to coralligenous assemblages. In Proceedings of the Conference: 2° Mediterranean Symposium on the Conservation of Coralligenous & Other Calcareous Bio-Concretions, Portorož, Slovenia, 29–30 October 2014. [Google Scholar] [CrossRef]
- Rasband, W.S. ImageJ: Image processing and analysis in Java. Astrophysics Source Code Library, 2012, record ascl:1206.013. Bibcode:2012ascl.soft06013R.
- Tsounis, G.; Martinez, L.; Bramanti, L.; Viladrich, N.; Gili, J.M. , Martinez, Á.; Rossi, S. Anthropogenic effects on reproductive effort and allocation of energy reserves in the Mediterranean octocoral Paramuricea clavata. Mar. Ecol. Prog. Ser. 2012, 449, 161–172. [Google Scholar] [CrossRef]
- Kipson, S. , Linares, C., Čižmek, H., Cebrián, E., Ballesteros, E., Bakran-Petricioli, T., Garrabou, J. Population structure and conservation status of the red gorgonian Paramuricea clavata (Risso, 1826) in the Eastern Adriatic Sea. Mar. Ecol. 2015, 36, 982–993. [Google Scholar] [CrossRef]
- Arnoux A., J.G. Harmelin, J.L. Monod, L.A. Romana, H. Zibrowius - Altérations des peuplements benthiques de roches profondes en Méditerranée nord-occidentale : quelques aspects biologiques et molysmologiques. Comptes rendus de l'Académie des sciences. Série 3, Sciences de la vie. 1992, Vol 314, Num 5, pp 219-225. Available online: http://pascal-francis.inist.fr/vibad/index.php?action=getRecordDetail&idt=5153892.
- Bramanti, L. , Magagnini, G., De Maio, L., Santangelo, G. Recruitment, early survival and growth of the Mediterranean Red Coral Corallium rubrum 1758, a four-year study. J. Exp. Mar. Biol. Ecol 2005, 314, 69–78. [Google Scholar] [CrossRef]
- Cupido, R. , Cocito, S., Sgorbini, S., Bordone, A., Santangelo, G. Response of a gorgonian (Paramuricea clavata) population to mortality events: recovery or loss. In Aquatic conservation marine and freshwater ecosystems; John Wiley & Sons, Ltd., 2008; Volume 18, pp. 984–992. ISSN 1052-7613. [Google Scholar]
- Enrichetti, F. , Bavestrello, G., Betti, F., Rindi, F., Tregrosso, A., Bo, M. Fate of lost fishing gears: Experimental evidence of biofouling colonization patterns from the northwestern Mediterranean Sea. Environ. Pollut. 2021, 268, 115746. [Google Scholar] [CrossRef]
- Asoh, K. , Yoshikawa, T., Kosaki, R., Marschall, E.A. Damage to cauliflower coral by monofilament fishing lines in Hawaii. Conserv. Biol. 2004, 18, 1645–1650. [Google Scholar] [CrossRef]
- Yoshikawa, T. , Asoh, K. Entanglement of monofilament fishing lines and coral death. Biol. Conserv. 2004, 117, 557–560. [Google Scholar] [CrossRef]
- Chiappone, M. , Dienes, H., Swanson, D.W., Miller, S.L. Impacts of lost fishing gear on coral reef sessile invertebrates in the Florida Keys National Marine Sanctuary. Biol. Conserv. 2005, 121, 221–230. [Google Scholar] [CrossRef]
- Hinz, H. Impact of bottom fishing on animal forests: science, conservation, and fisheries management. Marine Animal Forests: The Ecology of Benthic Biodiversity Hotspots; Rossi, S., Ed, 2017, 1041-1059.
- Rovere, A.; Ferraris, F.; Parravicini, V.; Navone, A.; Morri, C.; Bianchi, C.N. Characterization and evaluation of a marine protected area: ‘Tavolara–Punta Coda Cavallo’ (Sardinia, NW Mediterranean). J. Maps 2013, 9, 279–288. [Google Scholar] [CrossRef]
- Cupido, R. , Cocito, S., Barsanti, M., Sgorbini, S., Peirano, A., Santangelo, G. Unexpected long-term population dynamics in a canopy-forming gorgonian coral following mass mortality. Mar. Ecol. Prog. Ser. 2009, 394, 195–200. [Google Scholar] [CrossRef]
- Lindsay, S.M. Frequency of injury and the ecology of regeneration in marine benthic invertebrates. Integr. Comparat. Biol. 2010, 50, 479–493. [Google Scholar] [CrossRef]
- Wahle, C.M. Regeneration of injuries among Jamaican gorgonians: the roles of colony physiology and environment. Biol. Bull. 1983, 165, 778–790. [Google Scholar] [CrossRef]
- Castanaro, J. , Lasker, H.R. Colony growth responses of the Caribbean octocoral, Pseudopterogorgia elisabethae, to harvesting. Invertebrate Biology 2003, 122, 299–307. [Google Scholar] [CrossRef]
- Cerrano, C. , Bavestrello, G. Medium-term effects of die-off of rocky benthos in the Ligurian Sea. What can we learn from gorgonians? Chem. Ecol. 2008, 24(S1), 73–82. [Google Scholar] [CrossRef]
- Sánchez, J.A. , Lasker, H.R. Do multi–branched colonial organisms exceed normal growth after partial mortality? Proceedings of the Royal Society of London. Series B: Biological Sciences 2004, 271 (suppl. 3), S117–S120. [Google Scholar] [CrossRef]
- Loya, Y. Skeletal regeneration in a Red Sea scleractinian coral population. Nature 1976, 261, 490–491. [Google Scholar] [CrossRef]
- Riegl, B. , Riegl, A. Studies on coral community structure and damage as a basis for zoning marine reserves. Biol. Conserv. 1996, 77, 269–277. [Google Scholar] [CrossRef]
- Cerrano, C.; Arillo, A.; Azzini, F.; Calcinai, B.; Castellano, L.; Muti, C.; Valisano, L.; Zega, G.; Bavestrello, G. Gorgonian population recovery after a mass mortality event. Aquat. Conserv. 2005, 15, 147–157. [Google Scholar] [CrossRef]
- Ros, J.D., Romero, J., Ballesteros, E, Gili, J.M. Diving in blue water. The benthos. Western Mediterranean. R. Margalef, ed., Pergamon Press, Oxford, 1985, pp: 233-295.
- Hrs-Brenko, M. , Legac, M. Inter-and intra-species relationships of sessile bivalves on the eastern coast of the Adriatic Sea. Natura Croatica 2006, 15, 203. [Google Scholar]
- Basile, G. , Cerrano, C., Radjasa, O., Povero, P., Zocchi, E. ADP-ribosyl cyclase and abscisic acid are involved in the seasonal growth and in post-traumatic tissue regeneration of Mediterranean sponges. J. Exp. Mar. Biol. Ecol. 2009, 381, 10–17. [Google Scholar] [CrossRef]
- Chimienti, G. Vulnerable forests of the pink sea fan Eunicella verrucosa in the Mediterranean Sea. Diversity 2000, 12, 176. [Google Scholar] [CrossRef]
- McFadden, C.S. Genetic and taxonomic relationships among Northeastern Atlantic and Mediterranean populations of the soft coral Alcyonium coralloides. Mar. Biol. 1999, 133, 171–184. [Google Scholar] [CrossRef]
- McFadden, C.S. , Donahue, R., Hadland, B.K., Weston, R. A molecular phylogenetic analysis of reproductive trait evolution in the soft coral genus Alcyonium. Evolution 2001, 55, 54–67. [Google Scholar] [CrossRef]
- Quintanilla, E. , Gili, J. M., López-González, P. J., Tsounis, G., Madurell, T., Fiorillo, I., Rossi, S. Sexual reproductive cycle of the epibiotic soft coral Alcyonium coralloides (Octocorallia, Alcyonacea). Aquat. Biol. 2013, 18, 113–124. [Google Scholar] [CrossRef]
- Ten Hove, H.A. Different causes of mass occurrence in serpulids. Biology and systematics of colonial organisms 1979, 11, 281–298. [Google Scholar]
- Ten Hove, H.A., Van den Hurk, P. A review of Recent and fossil serpulid'reefs'; actuopalaeontology and'Upper Malm'serpulid limestones in NW Germany. Geologie en Mijnbouw 1993, (1). Netherlands Journal of Geosciences. Kluwer/Cambridge University Press: Den Haag, Cambridge. ISSN 0016-7746.
- Sarà, M. Competition and cooperation in sponge populations. In Symposium of the Zoological Society of London 1970, Vol. 25, pp. 273-284.
- Denoual, D. Mortalité massive de gorgones rouges (Paramuricea clavata) en méditerranée durant l'été 1999 (Doctoral dissertation). Ecole Nationale Vétérinaire de Toulouse - ENVT, 2004, 86 p.
- Wahl, M. Marine epibiosis. I. Fouling and antifouling: some basic aspects. Mar. Ecol. Prog. Ser. /, 1989; 175–189. Available online: https://www.jstor.org/stable/24842178.
Figure 1.
The investigated area within the Tavolara – Punta Coda Cavallo MPA. Red spots granitic shoals with the presence of Paramuricea clavata; yellow spots, shoals considered in this study.
Figure 1.
The investigated area within the Tavolara – Punta Coda Cavallo MPA. Red spots granitic shoals with the presence of Paramuricea clavata; yellow spots, shoals considered in this study.
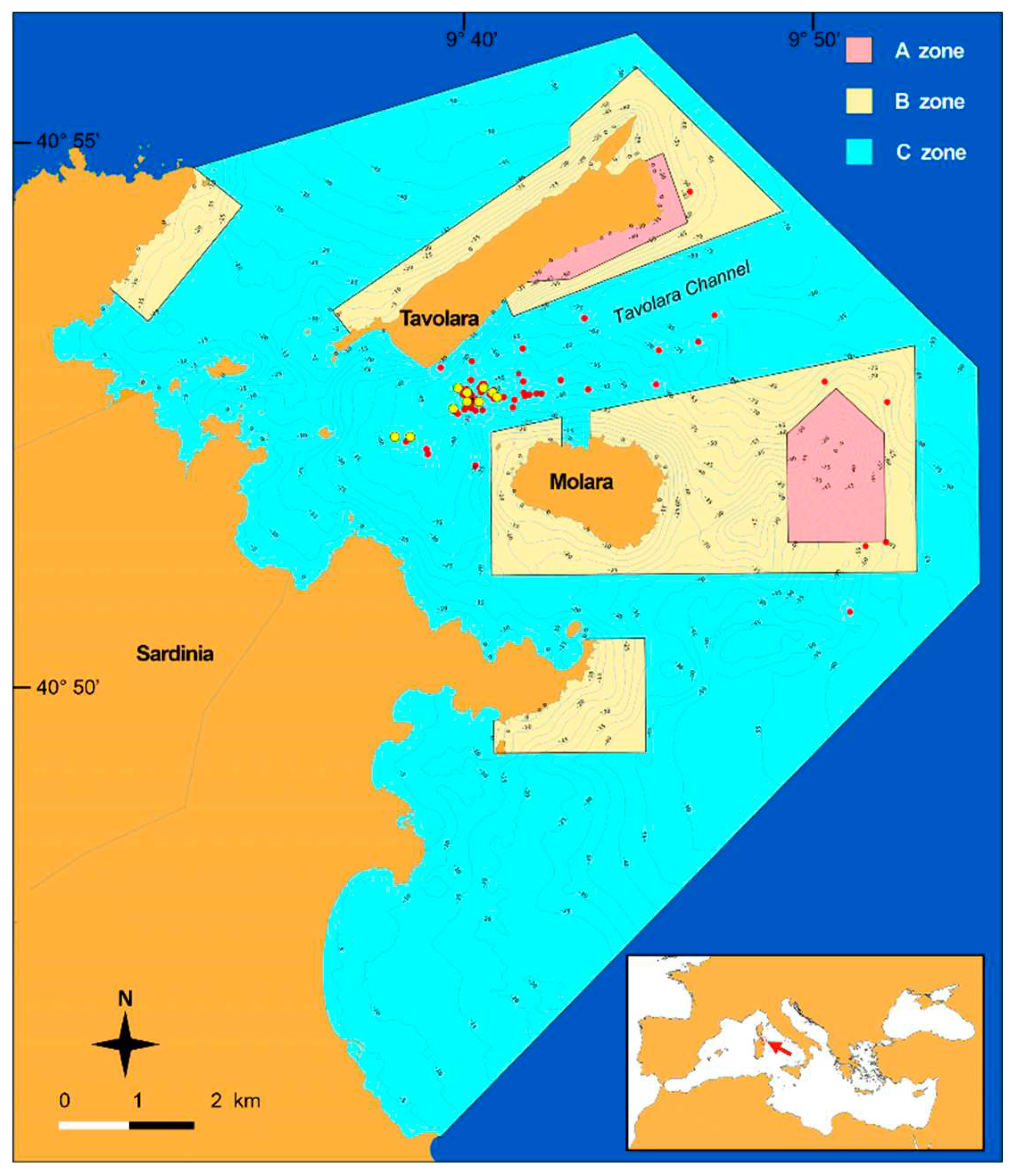
Figure 2.
(a-b) The benthic community settled on the granitic shoals of the Tavolara Channel mainly characterized by large sponges (Sarcotragus foetidus, Axinella spp.), gorgonians (Eunicella verrucosa, P. clavata). (c-d) Diseased colonies showed a massive coenenchyme loss during the mass mortality events as a consequence of summer heat weaves. (e) Spawning and (f) recruits of P. clavata; (g) spawning of the parasitic soft coral Alcyonium coralloides.
Figure 2.
(a-b) The benthic community settled on the granitic shoals of the Tavolara Channel mainly characterized by large sponges (Sarcotragus foetidus, Axinella spp.), gorgonians (Eunicella verrucosa, P. clavata). (c-d) Diseased colonies showed a massive coenenchyme loss during the mass mortality events as a consequence of summer heat weaves. (e) Spawning and (f) recruits of P. clavata; (g) spawning of the parasitic soft coral Alcyonium coralloides.

Figure 3.
(a-c) Paramuricea clavata forests showing a high level of epibiosis due to hydrozoan carpet (Hy), the false black coral Savaglia savalia (Ss), Filograna/Salmacina spp. (FS) and the calcified branched bryozoan Adeonella calveti (Aca). (d) Colonies recorded entangled in lost fishing gears and (e) totally uprooted.
Figure 3.
(a-c) Paramuricea clavata forests showing a high level of epibiosis due to hydrozoan carpet (Hy), the false black coral Savaglia savalia (Ss), Filograna/Salmacina spp. (FS) and the calcified branched bryozoan Adeonella calveti (Aca). (d) Colonies recorded entangled in lost fishing gears and (e) totally uprooted.

Figure 4.
percent recurrence of the recorded species/OTUs with values greater than 10%, calculated on the whole photograpic dataset.
Figure 4.
percent recurrence of the recorded species/OTUs with values greater than 10%, calculated on the whole photograpic dataset.
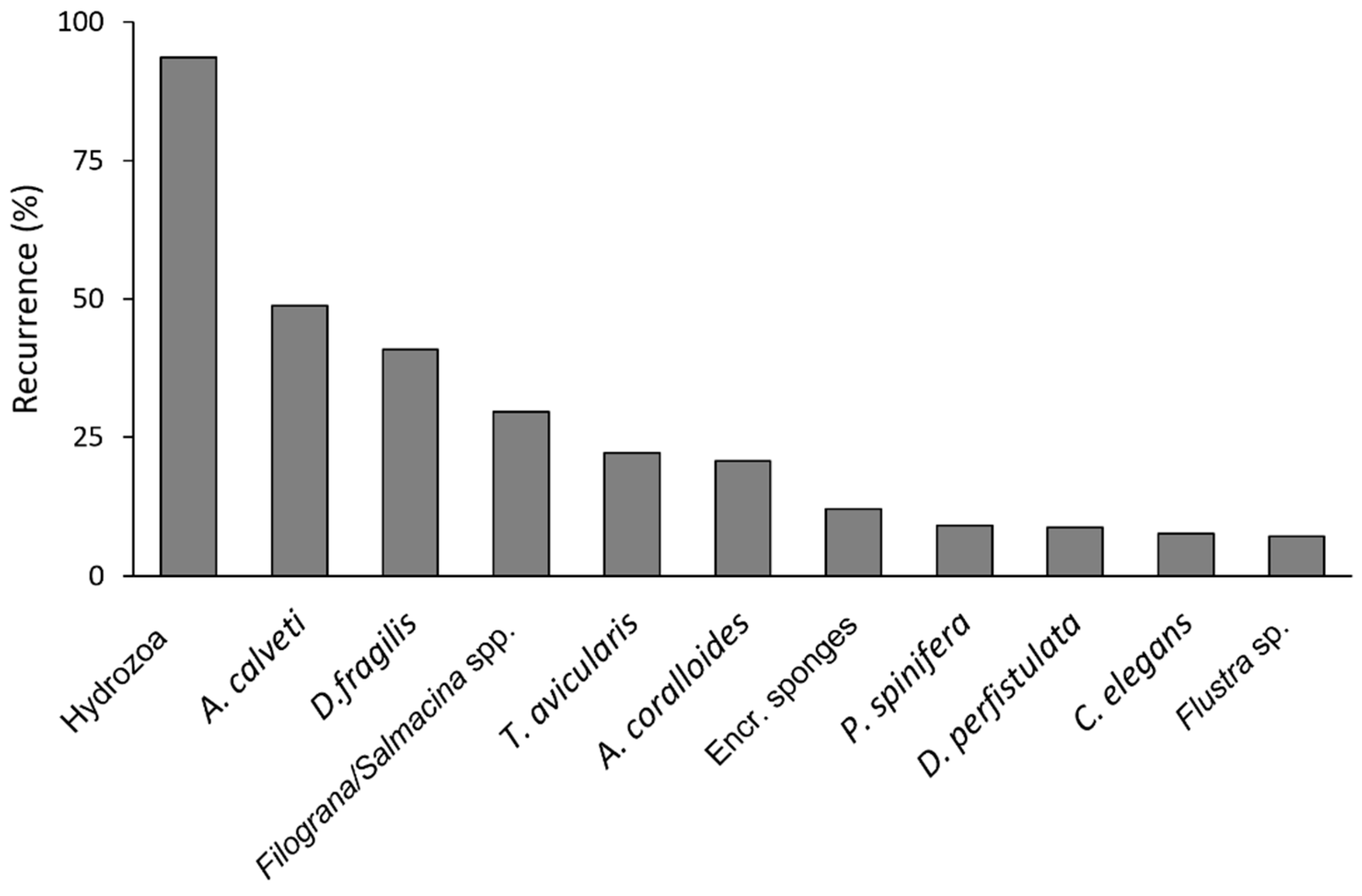
Figure 5.
The main epibionts recorded on Paramuricea clavata: (a) hydrozoan turf (Hy), (b) large masses of the serpulids Filograna/Salmacina spp. (FS), (c) the sponge Crella elegans (Ce), (d-h) complex epibiontic communities composed by the sponges Dysidea fragilis (Df), D. perfistulata (Dp) and Pleraplysilla spinifera (Ps), the parasitic soft coral Alcyonium coralloides (Aco), the serpulids Filograna/Salmacina spp., the bryozoans Adeonella calveti (Aca), Flustra sp. ((Fl) and Turbicellepora avicularis (Ta) and the ascidian Aplidium undulatum (Au).
Figure 5.
The main epibionts recorded on Paramuricea clavata: (a) hydrozoan turf (Hy), (b) large masses of the serpulids Filograna/Salmacina spp. (FS), (c) the sponge Crella elegans (Ce), (d-h) complex epibiontic communities composed by the sponges Dysidea fragilis (Df), D. perfistulata (Dp) and Pleraplysilla spinifera (Ps), the parasitic soft coral Alcyonium coralloides (Aco), the serpulids Filograna/Salmacina spp., the bryozoans Adeonella calveti (Aca), Flustra sp. ((Fl) and Turbicellepora avicularis (Ta) and the ascidian Aplidium undulatum (Au).

Figure 6.
Percent recurrence of the main species/OTUs showing different colonization strategies in relation to the diversity of the assemblage. (a) Hydrozoans are the main pioneer species and remain dominant at all the colonization phases; (b-i) branched bryozoans (Adeonella calveti, Flustra sp. and Turbicellepora avicularis), the serpulids Filograna/Salmacina spp. and the sponges Dysidea fragilis and Crella elegans settled immediately after hydrozoans and quickly increased according to the community richness. (j-l) D. perfistulata, Pleraplysilla spinifera and Alcyonium coralloides need a certain degree of diversity of the assemblage for their settling (> 4 species/OTUs); after that their growth is continuous.
Figure 6.
Percent recurrence of the main species/OTUs showing different colonization strategies in relation to the diversity of the assemblage. (a) Hydrozoans are the main pioneer species and remain dominant at all the colonization phases; (b-i) branched bryozoans (Adeonella calveti, Flustra sp. and Turbicellepora avicularis), the serpulids Filograna/Salmacina spp. and the sponges Dysidea fragilis and Crella elegans settled immediately after hydrozoans and quickly increased according to the community richness. (j-l) D. perfistulata, Pleraplysilla spinifera and Alcyonium coralloides need a certain degree of diversity of the assemblage for their settling (> 4 species/OTUs); after that their growth is continuous.
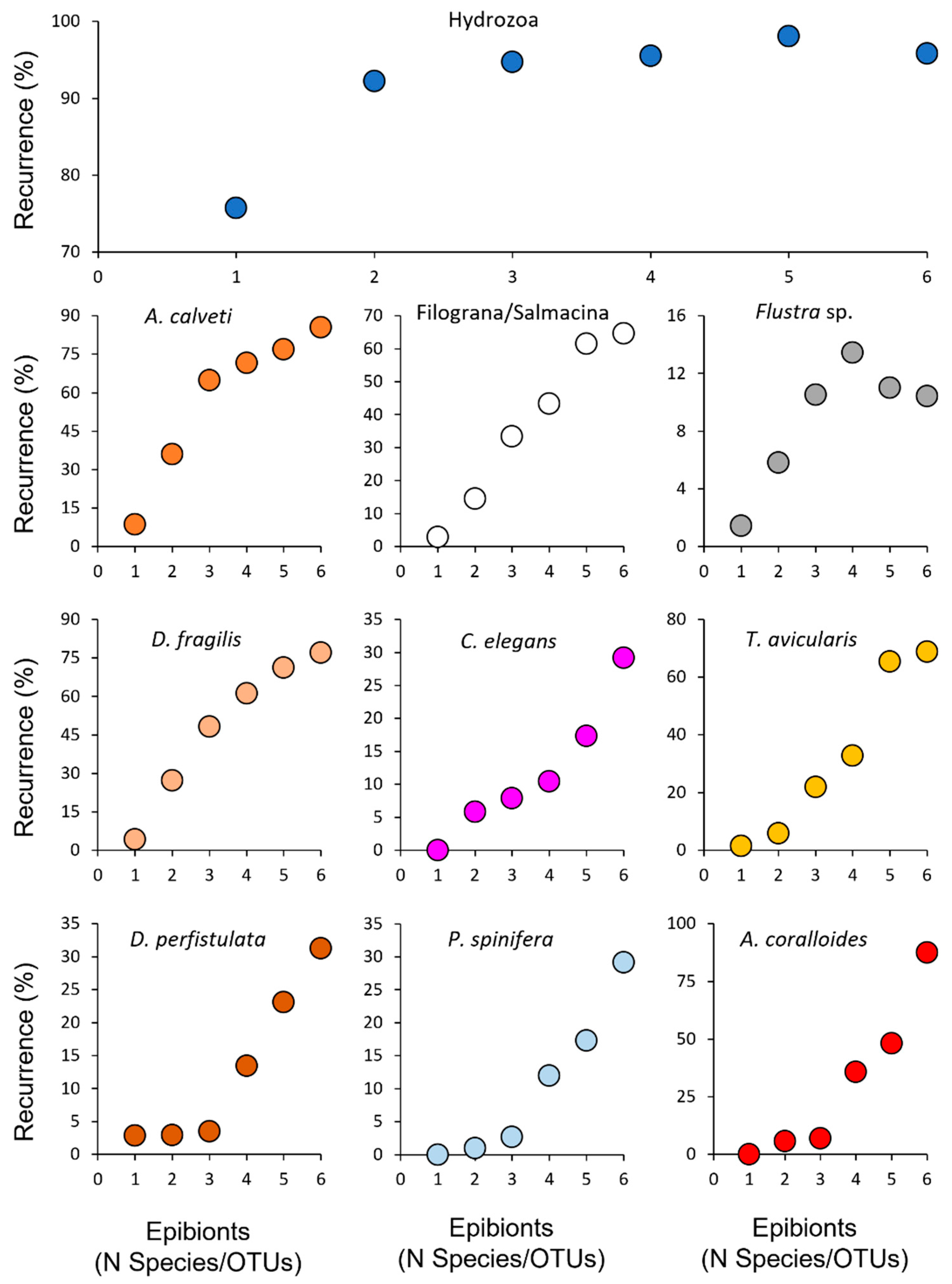
Figure 7.
(a) Percent recurrence of the main species/OTUs according to the diversity of the epibiontic community. (b) Average percentage of the gorgonian surface involved in the epibiosis, showing a progressive increase according to the diversity of the epibiontic community.
Figure 7.
(a) Percent recurrence of the main species/OTUs according to the diversity of the epibiontic community. (b) Average percentage of the gorgonian surface involved in the epibiosis, showing a progressive increase according to the diversity of the epibiontic community.
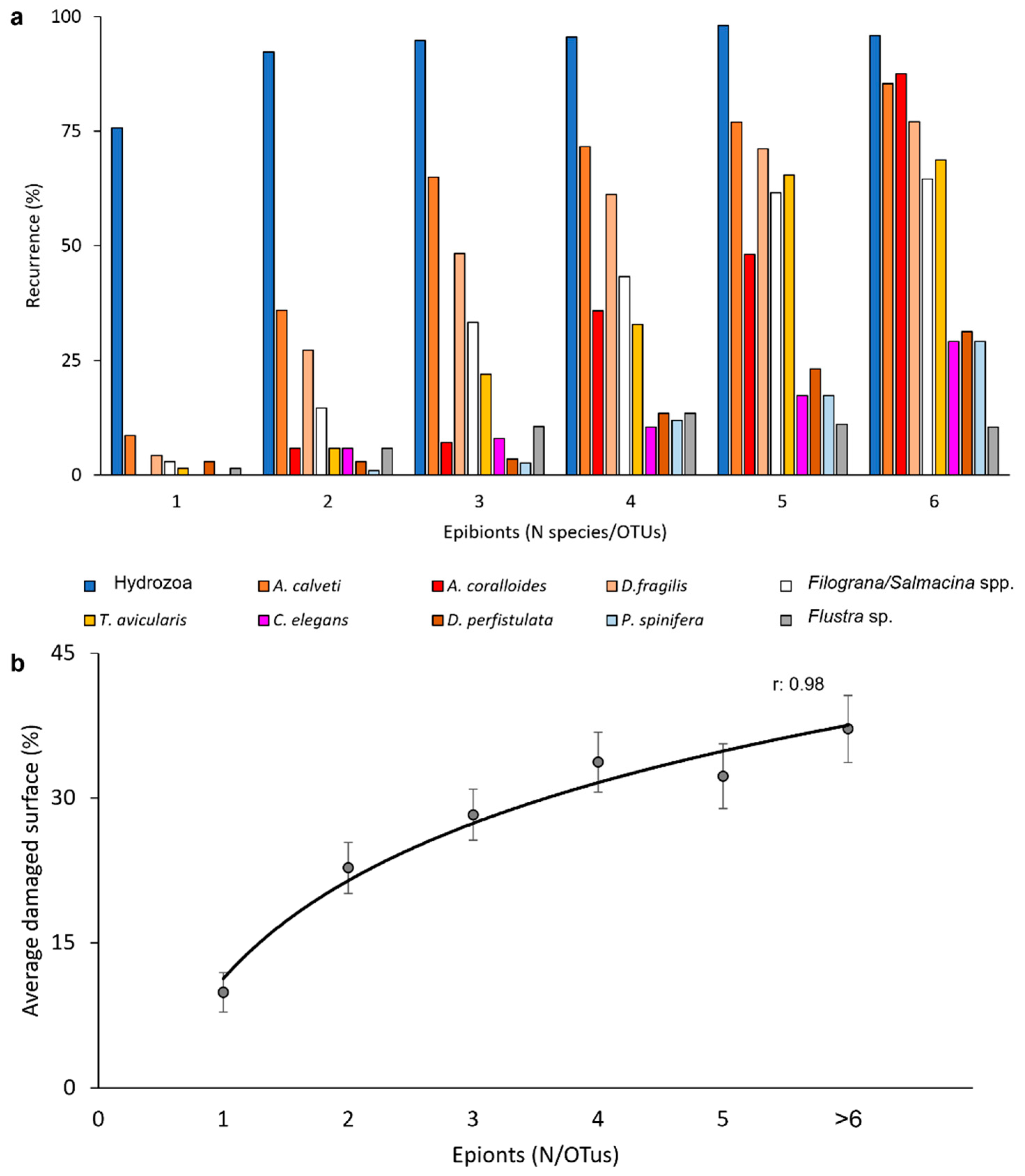
Figure 8.
Examples of different evolutions of the coenenchyme lesions and consequent epibiosis due to thermal stress. (a-d) Two cases of damages on small portions of branches were recovered after two years. (e-f) Extensive lesions were not recovered by the gorgonian and the epibiosis developed very quickly involving the entire colony. (g-j) A colony involved in the thermal disease of 2018 showed minor damage severity changes across four years.
Figure 8.
Examples of different evolutions of the coenenchyme lesions and consequent epibiosis due to thermal stress. (a-d) Two cases of damages on small portions of branches were recovered after two years. (e-f) Extensive lesions were not recovered by the gorgonian and the epibiosis developed very quickly involving the entire colony. (g-j) A colony involved in the thermal disease of 2018 showed minor damage severity changes across four years.

Figure 9.
Development of epibiosis linked to lost fishing lines entanglement. (a-b) Quick colonization (47 days) by sponges and bryozoans. (c-e) Increasing in complexity of the epibiontic community on a gorgonian entangled in a line (arrows) during 54 months; note that the sponge Dysidea fragilis appears later. (f-h) The settlement of Filograna/Salamacina spp. covered almost the entire naked parts (arrows) of a colony after the summertime 2018. (i-j) The settling of calcified organisms stops the movement of the entangled lines so preventing further damage. (k-m) A dead colony totally covered by sponges and encrusting coralline algae after five years.
Figure 9.
Development of epibiosis linked to lost fishing lines entanglement. (a-b) Quick colonization (47 days) by sponges and bryozoans. (c-e) Increasing in complexity of the epibiontic community on a gorgonian entangled in a line (arrows) during 54 months; note that the sponge Dysidea fragilis appears later. (f-h) The settlement of Filograna/Salamacina spp. covered almost the entire naked parts (arrows) of a colony after the summertime 2018. (i-j) The settling of calcified organisms stops the movement of the entangled lines so preventing further damage. (k-m) A dead colony totally covered by sponges and encrusting coralline algae after five years.


Table 1.
Species/OTUs found on Paramuricea clavata and their percent recurrence, calculated on the whole photographic dataset. (*) vagile species and taxa with a recurrence < of 10% were not taken into account for the formal analysis.
Table 1.
Species/OTUs found on Paramuricea clavata and their percent recurrence, calculated on the whole photographic dataset. (*) vagile species and taxa with a recurrence < of 10% were not taken into account for the formal analysis.
| Species/OTUs | Recurrence (%) |
Species/OTUs | Recurrence (%) |
|---|---|---|---|
| Algae | Anellids | ||
| Crustose Coralline Algae | 2.5 | Filograna/Salmacina spp. | 33.3 |
| Flabellia petiolata | 0.2 | Serpulids | 5.6 |
| Valonia sp. | 0.2 | Bryozoans | |
| Sponges | Adeonella calveti | 52.3 | |
| Anchinoe tenacior | 0.2 | Beania magellanica | 0.9 |
| Crella elegans | 10.4 | Cellaria salicornioides | 0.5 |
| Dysidea avara | 0.2 | Flustra sp. | 9.6 |
| Dysidea fragilis | 43.8 | Pentapora fascialis | 1.6 |
| Dysidea perfistulata | 10.2 | Turbicellepora avicularis | 26.9 |
| Haliclona mediterranea | 1.4 | Encr. Bryozoans n.i. | 1.4 |
| Ircinia variabilis | 0.2 | Crustaceans | |
| Oscarella lobularis | 2.5 | Periclimenes scriptum* | 0.5 |
| Pleraplysilla spinifera | 9.9 | Echinoderms | |
| Terpios fugax | 2.8 | Antedon mediterranea* | 0.2 |
| Encr. Sponges | 14.4 | Astrospartus mediterraneus* | 0.2 |
| Cnidarians | Chordata | ||
| Hydrozoa | 91.9 | Aplidium undulatum | 7.9 |
| Alcyonium coralloides | 22.9 | Clavelina dellavallei | 1.6 |
| Epizoanthus sp. | 0.2 | Clavelina oblonga | 0.9 |
| Molluscs | Didemnids | 0.9 | |
| Pteria hirundo | 0.7 | Pycnoclavella communis | 0.7 |
| Calliostoma conulus* | 0.2 | ||
| Flabellina ischitana* | 0.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



