
Submitted:
20 June 2023
Posted:
21 June 2023
You are already at the latest version
Abstract
Tissue inhibitors of metalloproteinases (TIMPs) are essential players during tumor progression in various types of cancer, including breast cancer. TIMPs regulate the activity of matrix metallo-proteinases (MMPs) at different levels. However, a dual role for TIMPs in breast carcinogenesis, either promoting or inhibiting tumor progression, has been observed. By a comprehensive bioin-formatics analysis using different databases, we report that of the four TIMPs, only TIMP-1 is overexpressed in tumor breast tissue. However, TIMP-2, TIMP-3, and especially TIMP-4 present lower levels in breast tumor tissue than in normal tissue. Interestingly, high levels of TIMP-1 are associated with shorter survival in breast cancer patients. In contrast, low TIMP-2, TIMP-3, and TIMP-4 levels are associated with poor survival. Function enrichment analysis showed that TIMP-3 has the highest interactions with its target proteins and a higher number of processes re-lated to mammary carcinogenesis. We found differential expression of TIMP-1 concerning TIMP-3 and TIMP-4 in breast cancer and normal tissue. TIMP-4 has a significant relationship with cancer staging, including tumor size, lymph node spread, and metastasis. The correlation between immune cell infiltrates and TIMP expression is of great importance as they provide in-formation on breast cancer diagnosis, prognosis, progression, and response to treatment.
Keywords:
breast cancer
; TIMPs
; tumor progression
1. Introduction
Worldwide, breast cancer remains the most common malignancy and the second leading cause of cancer-related deaths in women [1,2]. Breast cancer is a heterogeneous disease with different histopathological and biological features [3]. According to immunohistochemical markers that include the presence or absence of estrogen receptors (ER), progesterone receptors (PR), and human epidermal growth factor receptor 2 (HER2), breast cancer can be classified into molecular subtypes. These subtypes are Luminal A (ER and PR positive, HER2 negative, low Ki67), Luminal B (ER and/or PR positive, HER2 positive or high Ki67), HER2 enriched (ER and PR negative, HER2 positive) and triple-negative (TNBC) (ER, PR, HER2 negative) [4]. Each molecular subtype shows different responses to clinical therapy and requires different treatment strategies [5]. In addition to molecular and histological subtypes, breast cancer is evaluated according to tumor grade and size, lymph node status, and metastasis [6].
Recently, potential biomarkers related to breast cancer development and progression, such as tissue inhibitors of metalloproteinases (TIMPs), have been proposed [7]. TIMPs regulate tissue extracellular matrix turnover, inhibit MMPs, induce apoptosis, inhibit tumor cell migration, invasion, angiogenesis, and mainly inhibit MMPs [8]. TIMPs are 21 to 28 kDa proteins that bind to MMPs and reversibly block their activity. Four TIMPs (TIMP-1, TIMP-2, TIMP-3, TIMP-4) have been described, and they are present in the extracellular matrix (ECM) in soluble form, except TIMP-3, which is bound to the ECM [9,10]. The inhibitory function of TIMPs is related to the presence of a C-terminal and an N-terminal domain, each with three conserved disulfide bonds, where the N-terminal domain folds into itself and binds to the active site of MMPs to inhibit their activity [10,11,12].
Structurally, TIMPs are similar and can selectively inhibit different MMPs; however, they do not do so with the same efficacy [10]. TIMP-1 preferentially inhibits MMP-7, MMP-9, MMP-1 and MMP-3, whereas TIMP-2 more effectively inhibits MMP-2. TIMP-3 can inhibit MMP-2 and MMP-9, and also most ADAMs (disintegrin and metalloproteinase), whereas TIMP-4 inhibits the catalytic activity of MMP-14 and MMP-2 [13]. Therefore, an imbalance between TIMPs and MMPs is an important factor for cancer development by affecting the integrity of the extracellular matrix [14].
Bioinformatic analyses report that TIMP-2 is a potential prognostic factor in several cancer types because it participates in pathways associated with ECM regulators, ECM degradation, and ECM disassembly [15]. Likewise, in another bioinformatics analysis, it was shown that TIMP-2 down-regulates several breast cancer subtypes; moreover, TIMP-2 expression was associated with overall survival with different clinical features; thus, TIMP-2 could serve as a potential target and prognostic biomarker in breast cancer [16]. However, there is little information on the biological role of all TIMPs in breast cancer; here, we elucidate the relationship of TIMPs with breast cancer progression stages, probability of disease-free survival, interaction with target proteins, correlation of TIMPs with MMPs, correlation of immune cell infiltrates with TIMPs, and epigenetic factors regulating TIMPs expression. Here, we report that overexpression of TIMP-1 is associated with longer survival of patients, whereas low levels of TIMP-2, TIMP-3 and TIMP-4 are associated with shorter survival of breast cancer patients. Interestingly, TIMP-3 showed the highest interactions and regulation of cancer-related functions. The correlation between immune cell infiltrates and TIMP expression is of great importance as they provide information on breast cancer diagnosis, prognosis, progression, and response to treatment.
2. Materials and methods
2.1. Expression levels of TIMP-1, TIMP-2, TIMP-3, TIMP-4 in cancer patients
The expression levels of TIMP-1, TIMP-2, TIMP-3, and TIMP-4 in healthy and tumor samples and in the pathological stage of breast cancer patients were queried in the GEPIA database (Gene Expression Profiling Interactive Analysis, http://gepia.cancer-pku.cn/index.html) [17]. For expression analysis, we compared the expression levels of TIMPs in tumors and normal breast tissue; 1085 breast cancer samples and 291 normal tissue samples were included. We entered the "expression analysis" term and "BRCA" as the type of cancer. In pathological stage analysis, each TIMP was entered, BRCA (breast cancer) was added as the cancer type, and the main stage was used for plotting. Data were obtained in log2(TPM+1) for the expression scale.
2.2. Survival analysis of TIMPs expression in cancer patients
Correlation analyses of recurrence-free survival (RFS) of TIMP-1, TIMP-2, TIMP-3, and TIMP-4 expression in cancer patients were queried from the Kaplan-Meier Plotter database (https://kmplot.com/analysis/) [18]. Kaplan-Meier Plotter can assess the correlation between the expression of 30k genes (mRNA, miRNA, protein) and survival in more than 25k samples from 21 tumor types, including breast, ovarian, lung, and gastric cancer. Database sources include GEO (Gene Expression Omnibus), EGA (The European Genome-phenome Archive), and TCGA (The Cancer Genome Atlas) for the discovery and validation of survival biomarkers based on a meta-analysis. Data were queried from expression data by RNA-seq using the symbols TIMP-1, TIMP-2, TIMP-3, and TIMP-4, respectively. Overall survival (OS) analysis of the four breast cancer subtypes was queried from the GENT2 database. Data were selected by subtype and selecting the TIMP to be analyzed.
2.3. TIMP-1, TIMP-2, TIMP-3 and TIMP-4 interactome
To determine the protein-protein interaction (PPI) of TIMPs with target proteins and to explain the cooperative interaction between TIMP-1, TIMP-2, TIMP-3, and TIMP-4, for physical interactions with other proteins and functions was queried in databases such as STRING (https://string-db.org/) and Metascape (https://metascape.org/gp/index.html#/main/step1). Interaction analyses were queried in the BioGRID database (https://thebiogrid.org/) [19] with the UniProt database accession number (https://www.uniprot.org/) for each TIMP. TIMP-1 (https://thebiogrid.org/112932), TIMP-2 (https://thebiogrid.org/112933), TIMP-3 (https://thebiogrid.org/112934) and TIMP-4 (https://thebiogrid.org/112935). The four interactomes were used to generate the global interaction network of the four TIMPs in Cytoscape software [20] that identifies the synergistic interaction of TIMPs with their target proteins.
2.4. Function enrichment analysis of TIMP-1, TIMP-2, TIMP-3 and TIMP-4
Gene lists were obtained from the TIMP-1, TIMP-2, TIMP-3, and TIMP-4 interacting proteins from the interactomes obtained in BioGRID. The user-supplied gene identifiers are first converted into their corresponding Homo sapiens gene IDs using the latest version of the database (last updated on 2022-04-22).
For function enrichment analysis, process and pathway enrichment analysis was performed with the following ontology sources in the Metascape database (https://metascape.org/gp/index.html#/main/step1) using pathways obtained from KEGG, GO biological processes, Reactome gene sets, canonical pathways, cell type signatures, CORUM, TRRUST, DisGeNET, PaGenBase, transcription factor targets, WikiPathways, PANTHER Pathway, and COVID.
Metascape analyzes protein-protein interaction enrichment. Protein-protein interaction enrichment analysis has been performed for each given gene list with the following databases: STRING, BioGrid, OmniPath, and InWeb_IM. Only physical interactions in STRING (physical score > 0.132) and BioGrid are used. The resulting network contains the subset of proteins that form physical interactions with at least one list member. If the network includes 3 and 500 proteins, the molecular complex detection algorithm 10 (MCODE) was applied to identify densely connected network components.
We also used the GeneMANIA database (http://genemania.org/) [21] to analyze the functions and interaction of TIMPs with other target proteins. GeneMANIA generates hypotheses about gene functions, analyzes gene lists, and determines the priority of genes according to their functions. The analysis was performed with the functional list of differentially expressed TIMPs family genes and related molecules.
2.5. Correlation analysis between TIMP-1, TIMP-2, TIMP-3, and TIMP-4 with MMPs
To perform the correlation between the expression of TIMP-1, TIMP-2, TIMP-3, and TIMP-4 with MMPs, the GEPIA database was used [17]. Gene A corresponds to TIMP-1, TIMP-2, TIMP-3, and TIMP-4, while gene B corresponds to the MMPs that we observed have physical interaction with each of the TIMPs. The correlation was obtained from breast cancer and normal tissue samples. Expression data are obtained in log2(TPM) for the expression scale.
2.6. Immune infiltration status of TIMPs
The TIMER2.0 database [22] (http://timer.comp-genomics.org/) was used to systematically evaluate the infiltration of different immune cells and their clinical effects. This analysis evaluated the correlation between the expression of TIMPs in breast cancer and the infiltration of immune cells: CD4+ T cells, CD8+ T cells, B cells, macrophages, neutrophils, and dendritic cells in breast cancer samples.
2.7. Statistical data
The expression levels of TIMPs in normal and tumor tissue and survival analysis were considered statistically significant (p< 0.05). The results between survival correlation were besides being considered significant with p< 0.05, and the Hazard ratio (HR) was also considered. In the functional enrichment analysis, all genes in the genome were used as enrichment background. Terms with a p-value < 0.01, a minimum count of 3, and an enrichment factor > 1.5 (enrichment factor is the ratio of observed counts to expected counts by chance) are collected and grouped according to their membership similarities. The p-values were calculated based on the cumulative hypergeometric distribution 2, and the values were calculated using the Benjamini-Hochberg procedure for accounting for multiple testing. For GO function analysis, they were considered using the range of -log10(p). Correlation (R) was calculated using Spearman’s coefficients; a non-logarithmic scale was used for calculation and a logarithmic scale axis for visualization. p < 0.05 (*), p < 0.01 (**), p < 0.001 (***) was considered statistically significant.
3. Results
3.1. Expression levels of TIMPs and correlation with progression stages of breast cancer patients
TIMPs possess several biological functions, such as regulating MMP activation, angiogenesis, cell growth, apoptosis, and metastasis [8,23]. For this reason, it is important to determine the expression levels of TIMPs in tumors and normal breast tissue and their relationship with cancer staging.
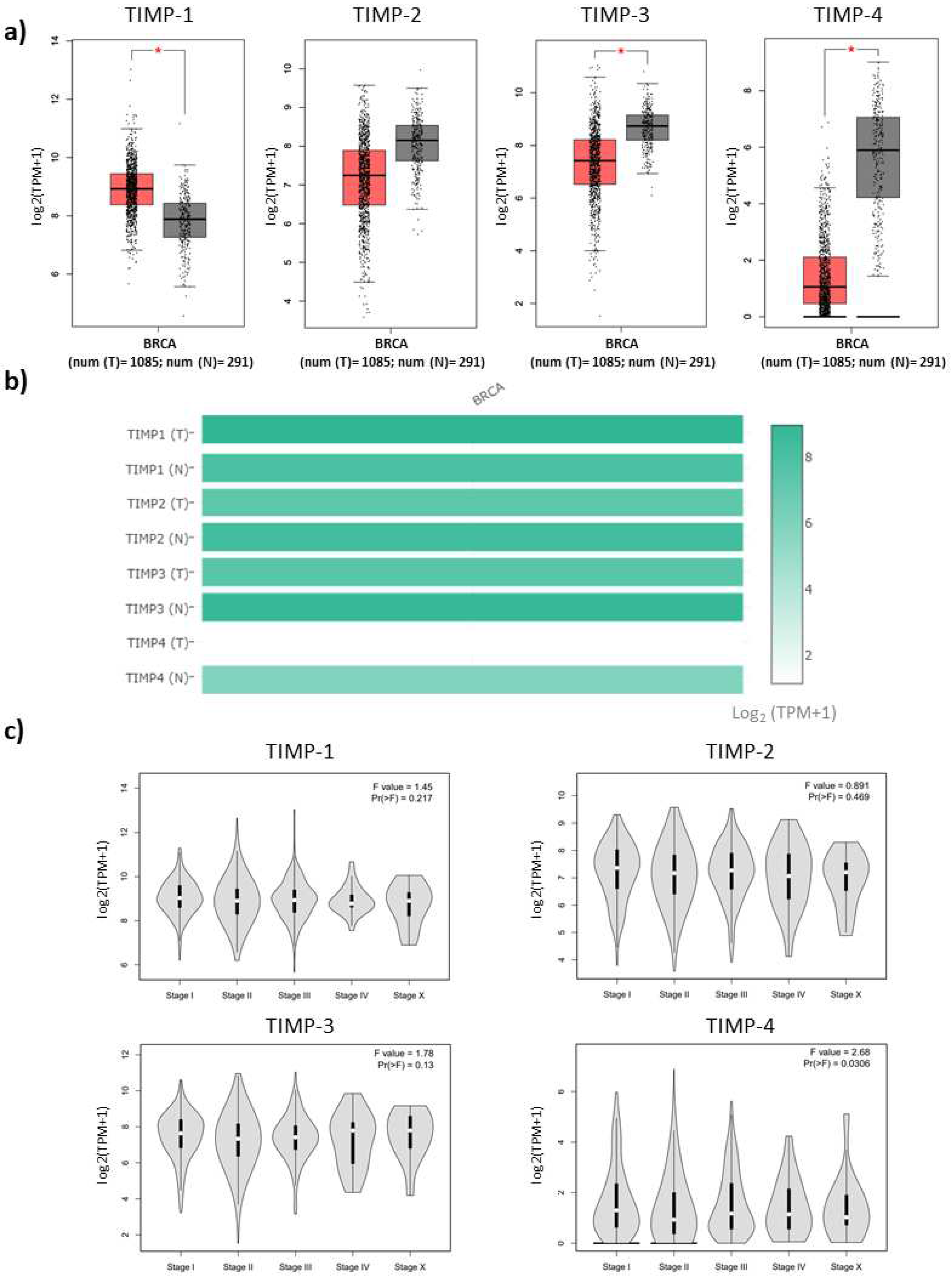
In the GEPIA database (http://gepia.cancer-pku.cn/index.html), we compared the expression levels of TIMPs in tumors and normal breast tissue; 1085 breast cancer samples and 291 normal tissue samples were included. We observed that TIMP-1 expression is 8.9 of log2(TPM+1) in tumors compared to 7.9 of log2(TPM+1) in normal tissues. TIMP-2 expression levels were lower by one log2(TPM+1) compared to normal tissue, TIMP-3 expression levels are 1.3 log2(TPM+1) higher in normal tissue than in tumor tissue, and TIMP-4 is 4.8 of log2(TPM+1) higher in normal tissue compared tumor tissue (Fig 1a b). We observed that expression levels in breast cancer were significantly increased (p < 0.05) compared to normal tissue for TIMP-1. In contrast, the expression levels of TIMP-3 and TIMP-4 significantly increased in normal tissue compared to tumor tissue. TIMP-2 levels increased in normal tissue compared to tumor tissue, although the data were not statistically significant (Figure 1a).
Breast cancer can progress rapidly from an early stage to an advanced stage; the earliest stage is stage 0 (carcinoma in situ) and ranges from stage I to stage IV; at the latter stage, the breast cancer has metastasized to other organs [24]. The rapid progression of breast cancer makes early diagnosis and monitoring of cancer progression difficult [25]. Therefore, we performed a correlation analysis in GEPIA of the expression levels of TIMPs and pathological stages of breast cancer. We observed that TIMP-4 expression differences were statistically significant p=0.0306 at stages I, II, III, IV, and V, indicating that TIMP-4 may be related to essential roles in the occurrence and development of breast cancer (Figure 1c). In contrast, the differences in expression of TIMP-1, TIMP-2, and TIMP-3 were greater p=0.05, i.e., there is no variability in cancer stages. This indicates that further studies are required to determine whether there is a relationship between their expression and breast cancer development (Figure 1c). These results suggest that TIMP-4 levels in patient biopsies could be considered a potential biomarker of breast cancer progression.
Figure 1.
TIMP-1, TIMP-2, TIMP-3, and TIMP-4 mRNA expression levels in breast cancer, normal breast tissue, and breast cancer stages according to TNM system. a) Comparison of TIMP-1, TIMP-2, TIMP-3, and TIMP-4 expression levels in breast cancer and normal tissue data were significant in TIMP-1, TIMP-3, and TIMP-4 (p-value <0.05). b) comparison of TIMPs expression in breast cancer with normal tissue. c) Correlations between TIMP1, TIMP-2, TIMP-3 and TIMP-4 expression and tumor stage in breast cancer patients. Data were generated in the GEPIA database; we used log2 (TPM+1) for the logarithmic scale. BRCA= Breast cancer.
Figure 1.
TIMP-1, TIMP-2, TIMP-3, and TIMP-4 mRNA expression levels in breast cancer, normal breast tissue, and breast cancer stages according to TNM system. a) Comparison of TIMP-1, TIMP-2, TIMP-3, and TIMP-4 expression levels in breast cancer and normal tissue data were significant in TIMP-1, TIMP-3, and TIMP-4 (p-value <0.05). b) comparison of TIMPs expression in breast cancer with normal tissue. c) Correlations between TIMP1, TIMP-2, TIMP-3 and TIMP-4 expression and tumor stage in breast cancer patients. Data were generated in the GEPIA database; we used log2 (TPM+1) for the logarithmic scale. BRCA= Breast cancer.
3.2. Survival analysis of TIMPs expression in breast cancer patients
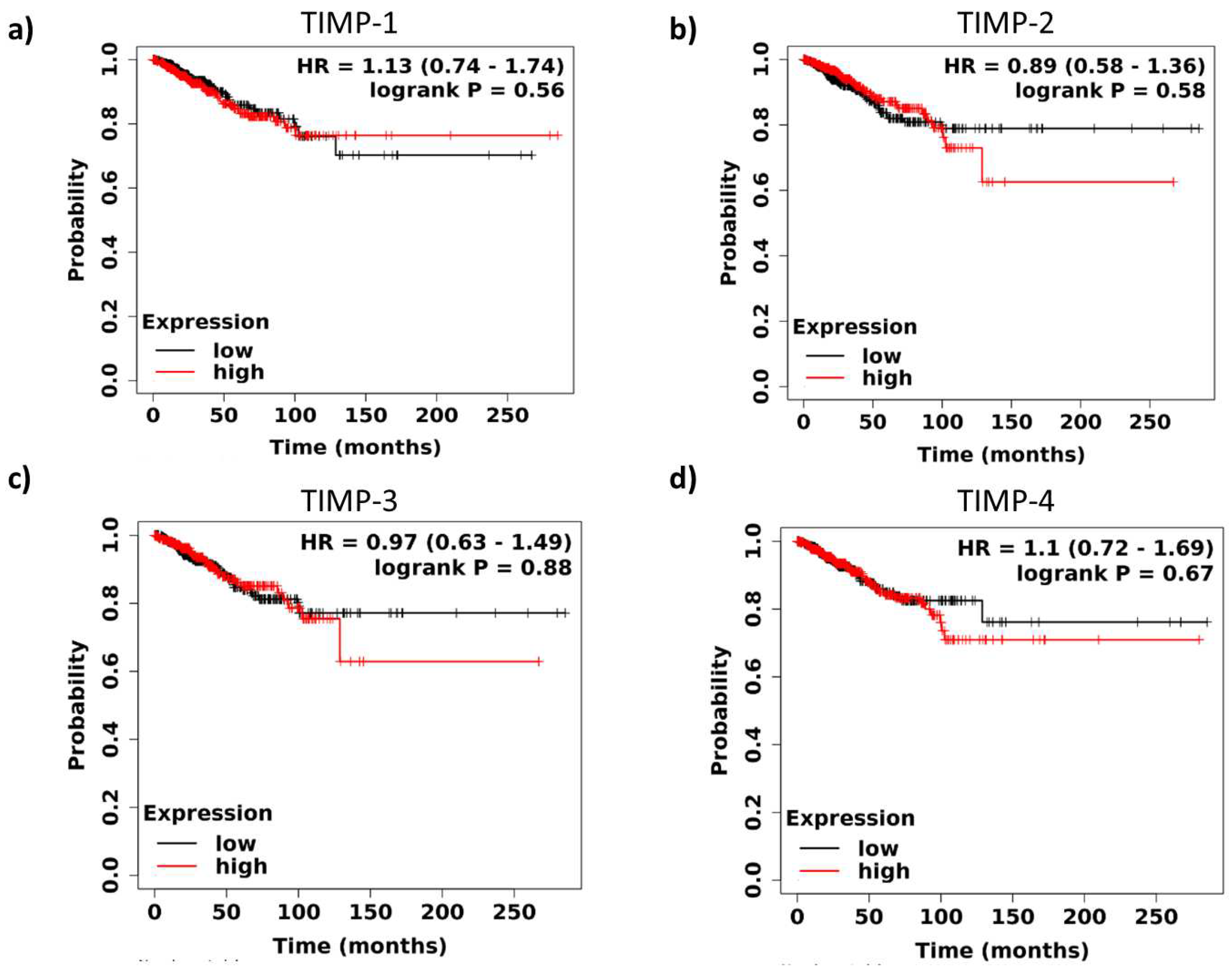
The role of TIMPs in breast cancer is very controversial; there are reports describing that TIMPs may have both a protumor and antitumor effect. It has been suggested that elevated levels of TIMPs are associated with early relapse in breast cancer patients [26]. Considering that we observed that TIMP-1 increases its expression in breast cancer, unlike TIMP-2, TIMP-3, and TIMP-4, whose expression decreases, we decided to perform an analysis in the Kaplan-Meier Plotter database (https://kmplot.com/analysis/) (Figure 2). We observed that high TIMP-1 mRNA levels negatively correlate with recurrence-free survival (RFS) of breast cancer patients. In contrast, patients with low TIMP-2, TIMP-3, and TIMP-4 mRNA expression showed a positive correlation of RFS in breast cancer patients. TIMP-1, TIMP-2, TIMP-3, and TIMP-4 mRNA levels were not statistically significant (p<0.05).
Figure 2.
Expression levels of TIMP-1, TIMP-2, TIMP-3, TIMP-4 and their correlation with survival in cancer patients. Lines in red represent high levels, and lines in black represent low levels of expression. The Y-axis represents the probability of survival, and the X-axis represents the follow-up time of the patients. A representative p-value is represented in each graph. The data were obtained from the Kaplan-Meier Plotter (https://kmplot.com/analysis/).
Figure 2.
Expression levels of TIMP-1, TIMP-2, TIMP-3, TIMP-4 and their correlation with survival in cancer patients. Lines in red represent high levels, and lines in black represent low levels of expression. The Y-axis represents the probability of survival, and the X-axis represents the follow-up time of the patients. A representative p-value is represented in each graph. The data were obtained from the Kaplan-Meier Plotter (https://kmplot.com/analysis/).
3.3. Interaction of TIMPs with their target proteins and functions
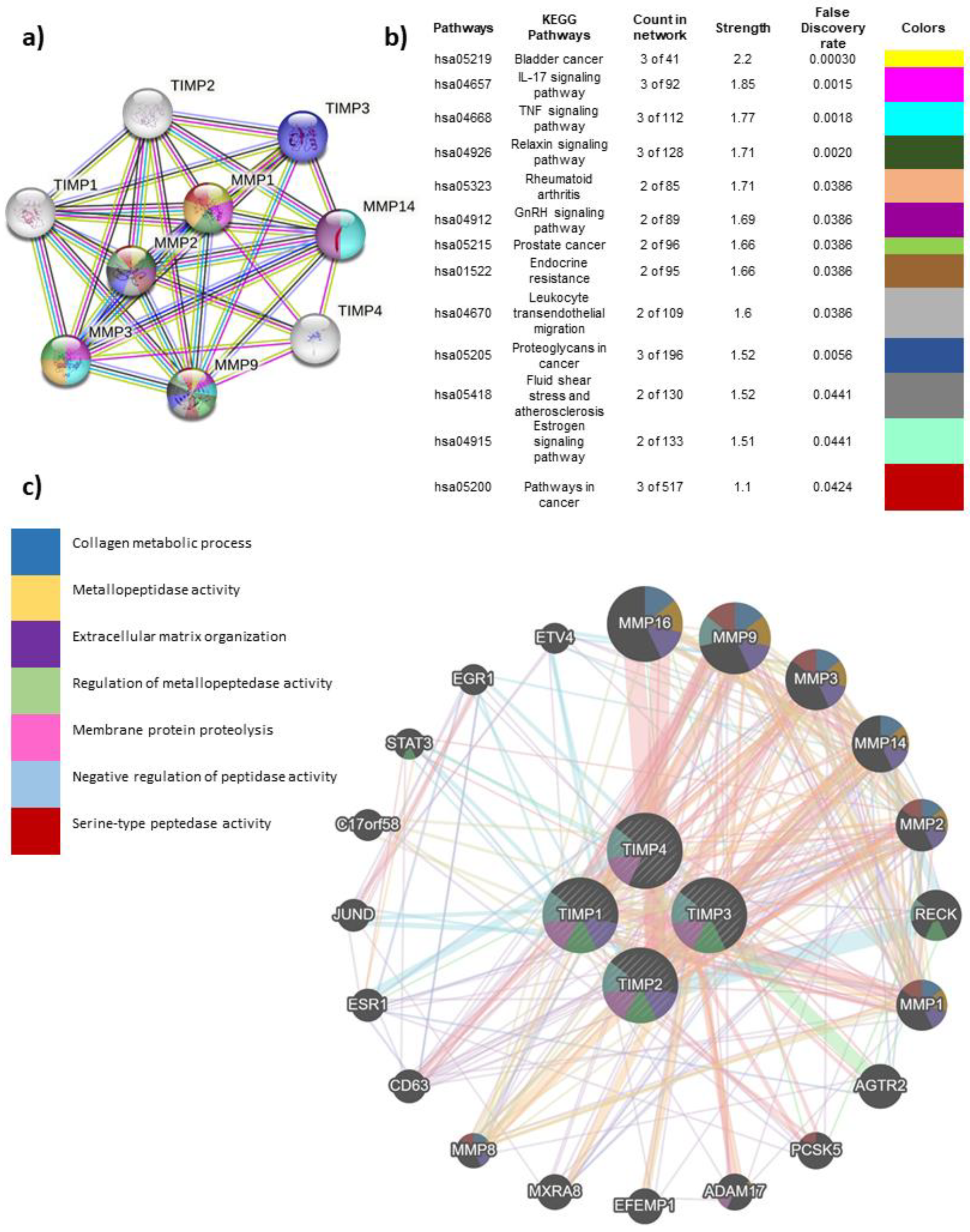
Breast cancer is associated with molecular alterations in protein networks that interact to regulate mechanisms related to proliferation, migration, invasion, metastasis, and drug resistance [27]. To understand the protein-protein interaction (PPI) mechanisms, we decided to perform an interaction and function analysis in the STRING (https://string-db.org/) and the Metascape (https://metascape.org/gp/index.html#/main/step1) database to explain the cooperative interaction in TIMPs: TIMP-1, TIMP-2, TIMP-3 and TIMP-4 and for physical interactions with other proteins, as well as to explain the functions of TIMPs in breast cancer development (Figure 3). The species set for the analysis was "Homo sapiens"; the other values were kept as default settings for the PPI analysis. The results in the PPI network showed 9 nodes, the number of edges was 27, the expected number of nodes was 5, and the average node degree was 6. The average local clustering coefficient was 0.659. The p-value of the PPI enrichment was 1.4e-11. Mutual interaction analysis between TIMPs showed that TIMP-1 interacts with TIMP-2 and TIMP-3. However, there is no interaction of TIMP-4 with the rest of the TIMPs (Figure 3a). KEGG pathway enrichment analysis obtained from the STRING database (Figure 3b) showed enrichment in 4 cancer-related functions, bladder cancer, prostate cancer, proteolysis in cancer, and global cancer processes, including tumor progression and maintenance. We also used the GeneMANIA database to analyze the functions and interaction of TIMPs with other target proteins essential in the maintenance and incision of the extracellular matrix. The results show that TIMPs interact together with MMP-16, MMP-9, MMP-8, MMP-3, MMP-14, MMP-2, MMP-1, RECK (reversion inducing cysteine-rich protein with kazal motifs), AGTR2 (angiotensin II receptor type 2), PCSK5 (proprotein convertase subtilisin/kexin type 5), ADAM17 (Disintegrin and metalloproteinase domain-containing protein 17), EFEMP1 (EGF-containing fibulin-like extracellular matrix protein 1), MXRA8 (Matrix remodeling-associated protein 8), CD63 (CD63 antigen), ESR1 (estrogen receptor 1), JUND (Transcription factor JunD), AP-1 transcription factor, C17orf58 (Chromosome 17 Open Reading Frame 58), STAT3 (Signal transducer and activator of transcription 3), ETV4 ETS variant transcription factor 4 (Figure 3c). Our results show that the interaction of TIMPs and their target proteins MMP-16, -14, -9, -8, -3, -2, and -1 are related to metabolic processes, collagen degradation, and metalloproteinase activity. Interestingly, MMP-1, -2, -3, -8, -9, and PCK5, through TIMPs, regulate the functions of extracellular matrix organization, regulation of metallopeptidase activity, membrane proteolysis proteins, peptidase inhibition activity and serine-type peptidase activity (Figure 3c).
Figure 3.
Mutual protein-protein interaction and cooperation analysis of TIMP-1, TIMP-2, TIMP-3, and TIMP-4. a) Protein-protein interaction and cooperativity analysis of TIMPs (STRIG analysis). b) Enrichment of the KEGG pathway of TIMPs (STRING). The colors indicate the cellular functions in which each TIMP in panel a participates. c) Diagram of protein-protein interaction analysis of TIMPs and their related molecules (GeneMANIA) in maintaining the extracellular matrix. On the left side, colors represent the functions of TIMPs and their target proteins represented in each node, respectively. The brown nodes represent other unrelated functions in cancer.
Figure 3.
Mutual protein-protein interaction and cooperation analysis of TIMP-1, TIMP-2, TIMP-3, and TIMP-4. a) Protein-protein interaction and cooperativity analysis of TIMPs (STRIG analysis). b) Enrichment of the KEGG pathway of TIMPs (STRING). The colors indicate the cellular functions in which each TIMP in panel a participates. c) Diagram of protein-protein interaction analysis of TIMPs and their related molecules (GeneMANIA) in maintaining the extracellular matrix. On the left side, colors represent the functions of TIMPs and their target proteins represented in each node, respectively. The brown nodes represent other unrelated functions in cancer.
We observed that TIMP-1 interacts physically through prediction and co-expression data with MMP-1, MMP-2, MMP-3, MMP-8, MMP-9, and MMP-14. Interestingly, in prediction and co-expression data, we observed that TIMP-2 shows physical interaction with MMP-1, MMP-2, MMP-3, MMP-8, MMP-9, MMP-14, and MMP-16. We also observed that there is co-expression of TIMP-2 with TIMP-1. We observed that TIMP-3 interacts physically and in co-expression with MMP-1, MMP-2, MMP-3, MMP-9, and MMP-14. Finally, we observed that TIMP-4 interacts directly with MMP-1, MMP-2, MMP-3, MMP-14 and MMP-16. In addition, we observed that there is co-expression of TIMP-4 and TIMP-3. We observed the interaction of TIMPs and target proteins such as MMP-1, MMP-2, MMP-3, MMP-8, MMP-9, MMP-14, and MMP-16. The low expression of TIMPs in breast cancer could be related to the progression of cancer to more aggressive stages as a consequence of the non-inhibition of the function of MMPs, including MMP-1, -2, -3, -8, -9, -14, and -16. Furthermore, we propose that there may be a cooperative effect between TIMP-1, TIMP-2, and TIMP-3 as an opportunity to treat breast cancer.
3.4. Global interactome of TIMPs in breast cancer
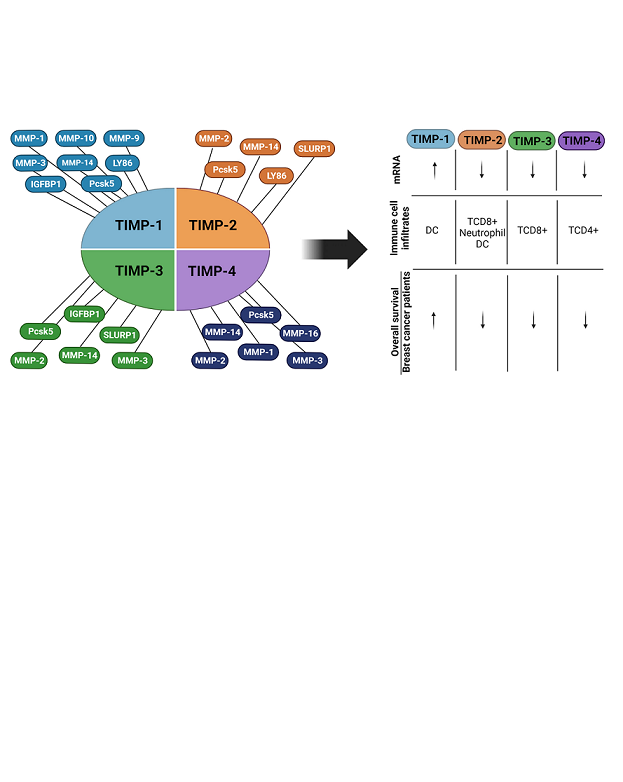
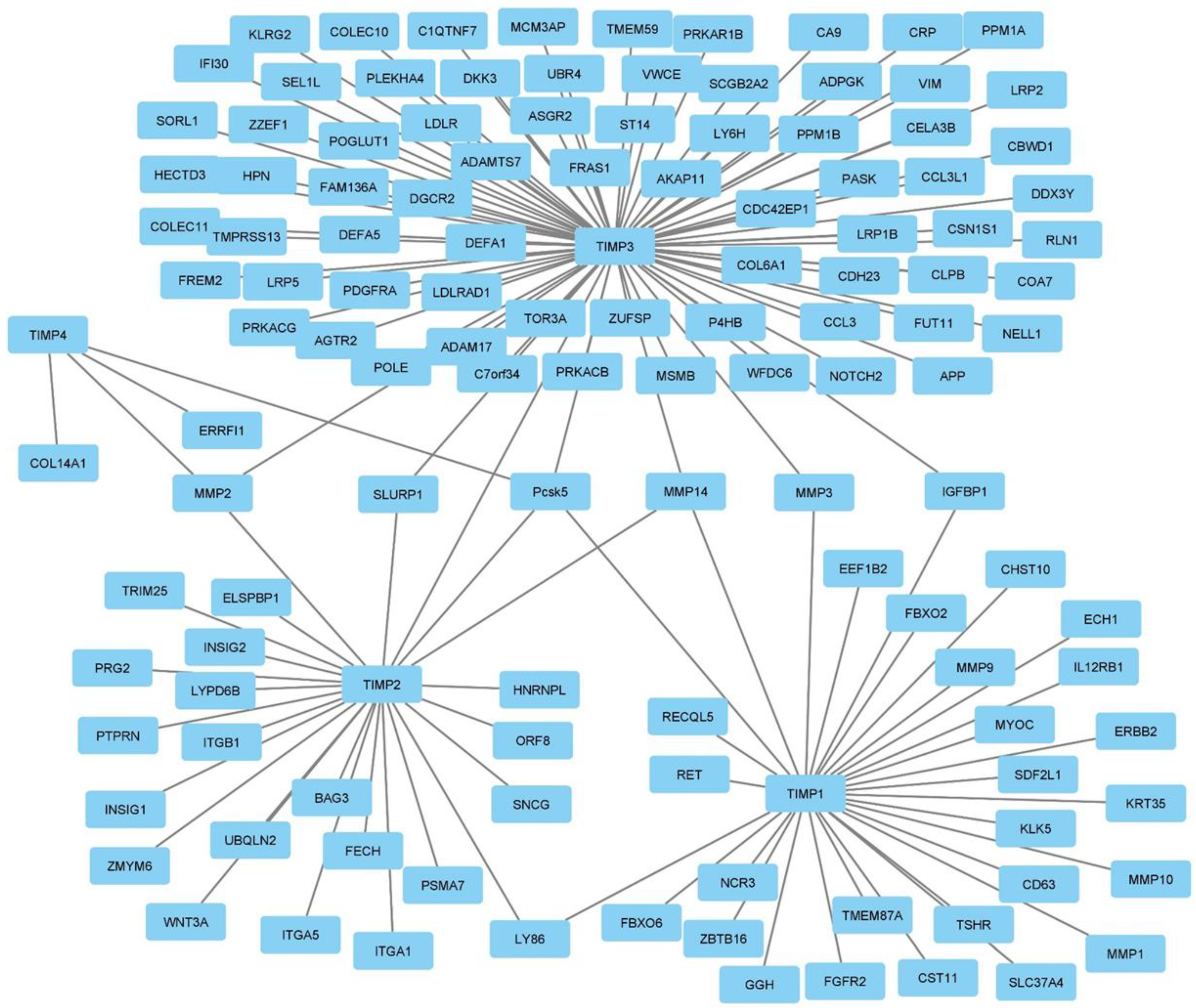
To determine the cooperation of TIMPs with their target proteins, the four interactomes were analyzed in Cytoscape to generate the global interactome (Figure 4). We observed that SLURP1 (secreted LY6/PLAUR domain containing 1) interacts with TIMP-2 and TIMP-3. MMP-3 and IGFBP1 (insulin-like growth factor binding protein 1) interact with TIMP-1 and TIMP-3. LY86 (lymphocyte antigen 86) interacts with TIMP-1 and TIMP-2. In contrast, MMP-2 interacts with TIMP-1, TIMP-2 and TIMP-4 and MMP-14 interacts with TIMP-1, TIMP-2 and TIMP3. While the four TIMPs, TIMP-1, TIMP-2, TIMP-3, and TIMP-4, interact with Pcsk5 (proprotein convertase subtilisin/kexin type 5).
Figure 4.
Global interactome of TIMPs. TIMP-1, TIMP-2, TIMP-3, and TIMP-4 are shown in the center of each interactome. The proteins that have interaction with each of the TIMPs are observed. The lines represent the interaction of the protein with each TIMPs. The interactome was created in Cytoscape software.
Figure 4.
Global interactome of TIMPs. TIMP-1, TIMP-2, TIMP-3, and TIMP-4 are shown in the center of each interactome. The proteins that have interaction with each of the TIMPs are observed. The lines represent the interaction of the protein with each TIMPs. The interactome was created in Cytoscape software.
Interaction analyses revealed that TIMP-1 interacts with MMP-1, MMP-2, MMP-3, MMP-8, MMP-9 and MMP-14. TIMP-2 interacts with MMP-1, MMP-2, MMP-3, MMP-8, MMP-9, MMP-14 and MMP-16. TIMP-3 interacts with MMP-1, MMP-2, MMP-3, MMP-9 and MMP-14. Finally, we observed that TIMP-4 interacts with MMP-1, MMP-2, MMP-3, MMP-14 and MMP-16.
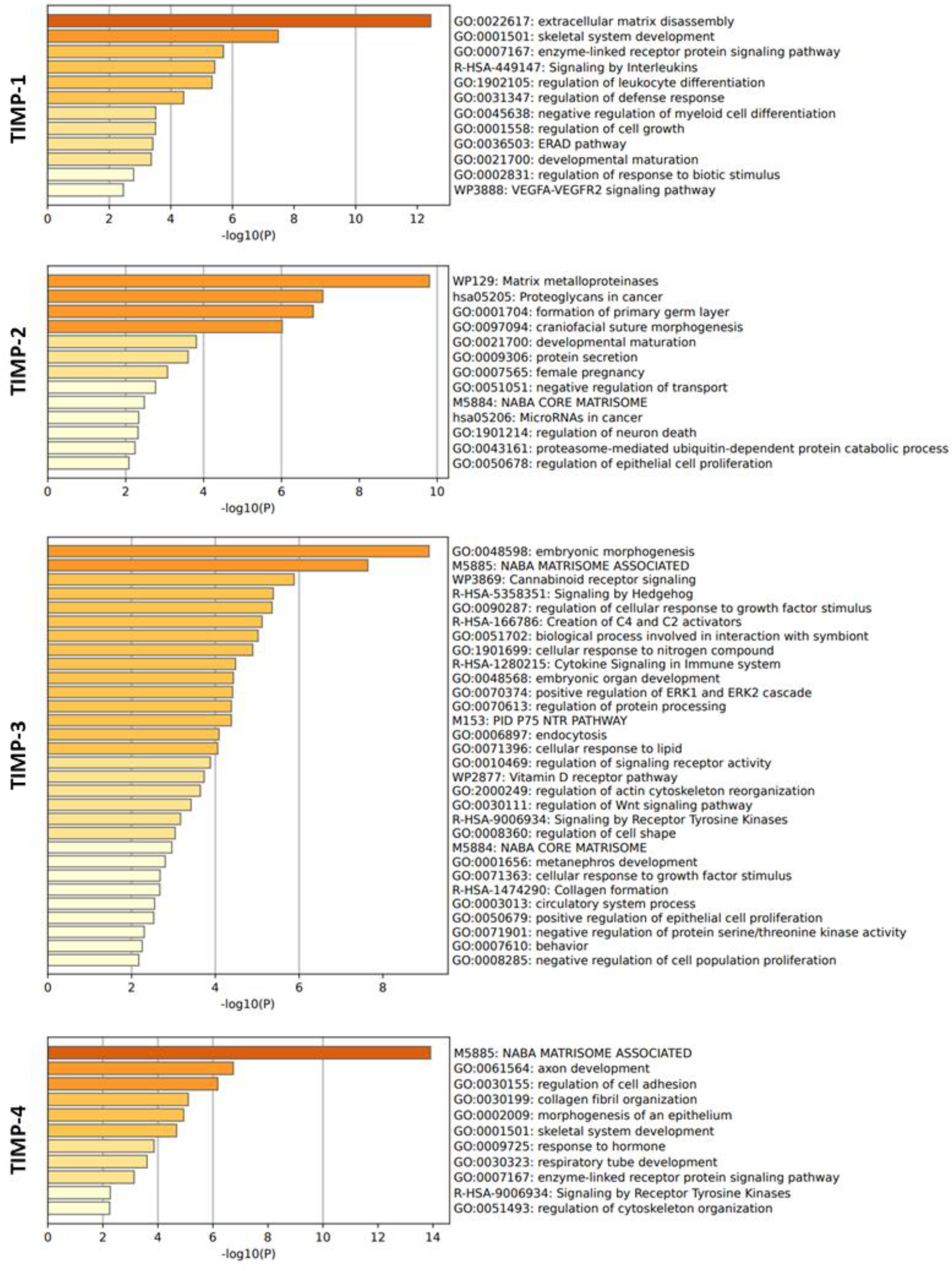
To identify the cellular functions where TIMP-1, TIMP-2, TIMP-3, and TIMP-4 participate, a Metascape pathway function enrichment analysis (https://metascape.org/gp/index.html#/main/step1) from proteins that physically interact with TIMP-1, 2, 3, and 4. The functions regulating the different TIMPs are shown in Figure 5. For each TIMP, the -log10(p) function enrichment data greater than 4 were as follows. TIMP-1 shows higher function enrichment in the GO:0022617: extracellular matrix disassembly and GO:0001501: skeletal system development pathways. TIMP-2 shows higher enrichment of functions in the pathway of WP129: Matrix metalloproteinases, hsa05205: Proteoglycans in cancer, GO:0001704: formation of the primary germ layer, GO:0097094: craniofacial suture morphogenesis. TIMP-3 has further enrichment in GO:0048598: embryonic morphogenesis, M5885: NABA matrisome associated pathways, WP3869: Cannabinoid receptor signaling, R-HSA-5358351: Signaling by Hedgehog, GO:0090287: regulation of cellular response to growth factor stimulus, R-HSA-166786: Creation of C4 and C2 activators, R-HSA-166786: Creation of C4 and C2 activators, GO:0051702: the biological process involved in interaction with symbiont, GO:1901699: cellular response to nitrogen compound, R-HSA-1280215: Cytokine Signaling in Immune system, GO:0048568: embryonic organ development, GO:0070374: positive regulation of ERK1 and ERK2 cascade, GO:0070613: regulation of protein processing, M153: PID P75 NTR PATHWAY, GO:0006897: endocytosis, GO:0006897: endocytosis and GO:0071396: cellular response to lipid. In contrast, TIMP-4 shows greater enrichment with functions in the pathways of M5885: Naba matrisome associated, GO:0061564: axon development, and GO:0030155: regulation of cell adhesion.
Figure 5.
Function enrichment analysis of TIMP-1, TIMP-2, TIMP-3, and TIMP-4. GO and function terms are plotted. The colors indicate the functions that show the highest enrichment for the values of –log10(p).
Figure 5.
Function enrichment analysis of TIMP-1, TIMP-2, TIMP-3, and TIMP-4. GO and function terms are plotted. The colors indicate the functions that show the highest enrichment for the values of –log10(p).
3.5. Correlation analysis of TIMP-1, TIMP-2, TIMP-3 and TIMP-4 with MMPs
TIMPs have been associated with breast cancer invasion and metastasis due to their ability to bind to MMPs and prevent their activation process [10]. We performed a correlation analysis to evaluate the effect of TIMPs on the expression levels of MMPs in breast cancer (Table 1). In breast cancer patients, TIMP-1 has a weak positive correlation with MMP-1, MMP-2, MMP-3, MMP-8, MMP-9, and MMP-14; the results were not statistically significant for MMP-1. In breast cancer samples, TMP-2 shows a weak positive correlation with MMP-1, MMP-8, and MMP-9, a moderate positive correlation with MMP-3 and MMP-16, and a strong positive correlation with MMP-2 and MMP-14, all data were statistically significant except for MMP-2. On the other hand, in breast cancer samples, TIMP-3 showed a weak positive correlation with MMP-1, MMP-3, and MMP-9 and a moderate positive correlation with MMP-2 and MMP-14; all data were statistically significant. TIMP-4 showed a weak positive correlation with MMP-2, MMP-3, MMP-14, and MMP-16 and a weak negative correlation with MMP-1; all values were statistically significant. Correlation analyses of TIMP-1, TIMP-2, TIMP-3, and TIMP-4 with MMPs in breast cancer patient samples show that as TIMP levels increase, MMP levels also increase, but not in the case of TIMP-4 and MMP-1.
In normal tissue, we observed that TIMP-1 has a weak positive correlation with MMP-1, MMP-2, MMP-8, MMP-9, and MMP-14, while a moderate positive correlation was observed with MMP-3; all data were significant except for MMP-8. TIMP-2 shows strong positive correlation with MMP2, weak positive correlation with MMP-3, MMP-8, MMP-9, MMP-14 and MMP-16. We observed a weak negative correlation between TIMP-2 and MMP-1, but they were not significant as with MMP-3. TIMP-3 shows a weak negative correlation with MMP-1, MMP-3, MMP-9, and MMP-14; only a weak positive correlation was observed with MMP-2. The data were statistically significant, except for MMP-9 and MMP-14. Finally, we observed that TIMP-4 has a weak negative correlation with MMP-1, MMP-3, MMP-14, and MMP-16; only a weak positive correlation was observed with MMP-2. All data were significant except for MMP-14. In normal breast tissue, we observed a weak negative correlation of TIMPs with MMPs; the data indicate that when TIMP levels are low, MMP levels increase.

Table 1.
Correlation analysis of TIMP-1, TIMP-2, TIMP-3, and TIMP-4 with MMPs in the GEPIA database.
Table 1.
Correlation analysis of TIMP-1, TIMP-2, TIMP-3, and TIMP-4 with MMPs in the GEPIA database.
| TIMPs | MMPs | Tissue | |||
| Tumor | Normal | ||||
| R | P | R | P | ||
|
TIMP-1 |
MMP-1 | 0.023 | 0.46 | 0.39 | *** |
| MMP-2 | 0.21 | *** | 0.38 | *** | |
| MMP-3 | 0.14 | *** | 0.6 | *** | |
| MMP-8 | 0.062 | * | 0.16 | 0.085 | |
| MMP-9 | 0.17 | *** | 0.45 | *** | |
| MMP-14 | 0.14 | *** | 0.47 | *** | |
|
TIMP-2 |
MMP-1 | 0.32 | *** | -0.0038 | 0.97 |
| MMP-2 | 0.87 | 0 | 0.76 | *** | |
| MMP-3 | 0.54 | *** | 0.016 | 0.87 | |
| MMP-8 | 0.4 | *** | 0.3 | *** | |
| MMP-9 | 0.25 | *** | 0.19 | * | |
| MMP-14 | 0.85 | *** | 0.24 | ** | |
| MMP-16 | 0.63 | *** | 0.19 | * | |
|
TIMP-3 |
MMP-1 | 0.15 | *** | -0.24 | * |
| MMP-2 | 0.58 | *** | 0.43 | *** | |
| MMP-3 | 0.29 | *** | -0.29 | ** | |
| MMP-9 | 0.11 | *** | -0.044 | 0.64 | |
| MMP-14 | 0.55 | *** | -0.14 | 0.14 | |
|
TIMP-4 |
MMP-1 | -0.22 | *** | -0.26 | ** |
| MMP-2 | 0.22 | *** | 0.3 | ** | |
| MMP-3 | 0.19 | *** | -0.31 | *** | |
| MMP-14 | 0.072 | * | -0.17 | 0.072 | |
| MMP-16 | 0.25 | *** | -0.38 | *** | |
BRCA, Breast invasive carcinoma. Tumor, correlation analysis in breast cancer tissue from TCGA database. Normal, correlation analysis in normal tissue from TCGA database. Correlation (R) and P-Value (P). Statistical significance *P < 0.05, **P< 0.01, ***P< 0.001.
3.6. Immune cell infiltrates by expression of TIMP-1, TIMP-2, TIMP-3 and TIMP-4 in breast cancer patients and subtypes
Immune cells are essential components of normal breast tissue and act in innate and adaptive immunity. During breast cancer progression, there is an increase in immune cell infiltrates in the tumor parenchyma and stroma, such as granzyme B+ CD4+ and CD8+ cytotoxic T lymphocytes, B lymphocytes, macrophages, and dendritic cells [28]. These infiltrating lymphocytes could function as prognostic indicators for response to chemotherapy and survival of breast cancer patients [29].
Given the importance of immune cell infiltrates in breast cancer development, we decided to perform an analysis in the TIMER2.0 database to analyze the correlation between infiltrating B cells, CD8+ T cells, CD8+ T cells, CD4+ T cells, macrophages, neutrophils and dendritic cells (DC) in breast cancer and the expression of TIMP-1, TIMP-2, TIMP-3, and TIMP-4 (Table 2). We analyzed 1100 breast cancer samples, type Basal 191, HER-2 82, BRCA-LumA 568, and BRCA-LumB 219. Correlation data were adjusted with Rho value and purity adjustment using Spearman correlation. TIMP1 has a positive correlation with dendritic cell infiltrates and a negative correlation with B-cell infiltrates. The data were not significant in TIMP-1 expression with CD8+ T cell infiltrates, CD4+ T cell infiltrates, macrophages, and neutrophils. TIMP-2 expression correlated positively with CD8+ T cell infiltrates, Neutrophils, and DCs. We observed a negative correlation between TIMP-2 expression and B-cell infiltrates; no statistically significant data were observed for CD4+ T-cell and macrophage infiltrates. TIMP-3 expression levels show a positive correlation with CD8+ T-cell infiltrates and a negative correlation with B-cell infiltrates. We did not observe a significant correlation with CD4+ T-cell, macrophage, neutrophil, and DC infiltrates. Finally, TIMP-4 expression levels show a positive correlation with CD4+ T cell infiltrates and a negative correlation with B cell infiltrates, macrophages, and neutrophils but no significant correlation with CD8+ T cell infiltrates and DCs.
Table 2.
Correlation between TIMPs and immune cell infiltration in breast cancer patients.
| TIMP | B cells | CD8+ T cells | CD4+ T cells | Macrophages | Neutrophils | DC |
| TIMP-1 | -0.136 | 0.043 | -0.045 | 0.063 | -0.029 | 0.149 |
| TIMP-2 | -0.237 | 0.406 | -0.004 | 0.087 | 0.317 | 0.279 |
| TIMP-3 | -0.135 | 0.311 | 0.087 | -0.048 | 0.054 | 0.05 |
| TIMP-4 | -0.11 | 0.072 | 0.143 | -0.108 | -0.139 | 0.03 |
Data in red indicate positive correlation (p<0.05, p>0), data in blue indicate negative correlation (p<0.05, p>0) and data in gray indicate non-significant data (p>0.05).
4. Discussion
Worldwide, breast cancer is the most common malignancy and the leading cause of cancer-related deaths in women [1]. Despite advances in breast cancer therapy, it is crucial to identify new biomarkers related to the subtype, grade, and size of breast tumors. TIMPs could be considered therapeutic targets involved with breast cancer progression and development. An imbalance between the expression of TIMPs and MMPs has been described as an important factor in cancer development by affecting the integrity of the extracellular matrix [14], inducing apoptosis, and promoting breast cancer migration, invasion, and angiogenesis [8].
TIMP-1 is a multifunctional protein that regulates the activation of different metalloproteinases, including MMPs and ADAMs. It also plays a central role in carcinogenesis, controlling the development and progression of breast cancer [30,31]. Our results showed an increase in TIMP-1 mRNA expression in breast cancer relative to normal tissue and an increase in the overall survival of patients. Interestingly, TIMP-1 presented the highest physical interaction and positively correlated with proteins involved in extracellular matrix disassembly, such as MMP-1, MMP-2, MMP-3, MMP-8, MMP-9, and MMP-14, regulating their activity. In addition, TIMP-1 is found to positively regulate the expression of skeletal system proteins and proteins that regulate signaling pathways that promote breast cancer progression. In this context, previous studies have reported that TIMP-1 regulates EMT [32], in MCF10A mammary epithelial cells through Twist regulation, which suggests TIMP-1 is upregulated in breast cancer, increasing survival in breast cancer patients [33]. However, it has been reported that high serum and tissue TIMP-1 levels in breast cancer patients are positively associated with lymph node metastasis, lower overall, and relapse-free survival [34]. In addition, high levels of TIMP-1 are associated with an increased risk of relapse in patients with primary breast carcinoma [35]. After a 3-year follow-up of breast cancer patients, high levels of TIMP-1 and MMP-9 are significantly associated with bone metastasis [36]. High levels of TIMP-1 in breast cancer have been reported to promote growth and inhibit apoptosis through activation of FAK, PI3K, Akt, and Bcl-2 signaling [37,38]. In addition, high levels of TIMP-1 in tumor tissue and serum of breast cancer patients are associated with a low response to chemotherapy and endocrine therapy [39,40]. TIMP-1 also induces the expression of transcription factors that promote EMT, such as SLUG, TWIST, ZEB1, and ZEB2, thereby inhibiting the expression of epithelial markers and promoting the expression of mesenchymal markers [32]. A previous report showed no association between TIMP-1 polymorphism and breast cancer; however, patients with C/C genotypes report elevated TIMP-1 levels, suggesting that the C allele may influence the level of TIMP-1 expression in serum [41]. All these data demonstrate that TIMP-1 expression is directly related to markers of tumor progression in breast cancer.
Regarding TIMP-2 participates in the remodeling of the extracellular matrix and can regulate the proliferation, invasion, and chemoresistance of tumor cells [42,43]. Our study found that the mRNA expression level of TIMP-2 was higher in normal tissue to breast cancer, whereas low TIMP-2 levels are directly related to lower overall survival. The primary function of TIMP-2 is to regulate the activation of MMPs, and we found that it interacts directly with MMP-1, MMP-2, MMP-3, MMP-8, MMP-9, MMP-14, and MMP-16. Previous reports show that nude mice transfected with TIMP-2 decreased tumor growth and invasion of mammary cancer cells [44]. Peney et al. report that TIMP-2 inhibits TNBC growth and metastasis in a murine model through modulation of epithelial-mesenchymal transition (EMT) by decreased expression of SNAIL, SLUG, ZEB1, and vimentin, as well as inhibition of PI3K and p27 [45]. However, Ree et al. report that TIMP-2 overexpression correlates with breast cancer progression; they found that high TIMP-2 mRNA levels promote lymph node metastasis in breast cancer patients [46].
Furthermore, high protein levels TIMP-2 in breast cancer tissues correlate with lower overall survival and recurrence-free survival in patients [47]. Interestingly, this dual effect of TIMP-2 is regulated at different stages of carcinogenesis, and the effect varies according to the tumor microenvironment; therefore, it is suggested that TIMP-2, TIMP-4, MMP-14 and MMP-2 levels should be determined together for prognostic evaluation [48]. In addition, TIMP-2 is a determinant in regulating MMP-2 activation, where TIMP-2 binds to proMMP-2 and forms a complex with MMP-14, which cleaves MMP-2, thus initiating its activation process [49]. These findings suggest that TIMP-2 may have a dual role in mammary carcinogenesis, promoting or inhibiting tumor progression in breast cancer.
TIMP-3 has been described to have anticancer effects, induce apoptosis, and inhibit angiogenesis and metastasis [50,51]. Interestingly, it has been described that it may be a biomarker and is the only TIMP that has been proposed as a therapeutic target in cancer [51,52,53]. On the other hand, plasma from patients with oral head and neck squamous cell carcinoma has been found to have decreased TIMP3 mRNA levels compared to healthy patients [53]. It has also been reported that overexpression of TIMP-3 inhibits in vitro invasion and promotes apoptosis of HeLa cervical cancer cells and HT1080 fibrosarcoma [54]. We found that TIMP-3 interacts physically and in co-expression with MMP-1, MMP-2, MMP-3, MMP-9, and MMP-14. Our analysis also showed that TIMP-3 regulates more functions and the highest number of protein interactions concerning the other TIMPs. Previous reports describe that the decrease in TIMP-3 levels is due to hypermethylation of the TIMP3 gene promoter and transcriptional repression, preventing TIMP-3 inhibits the activity of MMPs and consequently avoiding a malignant and invasive phenotype of cancer cells [55,56]. Bian, J., et al. in 1996 reported that overexpression of TIMP-3 in colon carcinoma DLD-1 cells inhibited cell growth and in nude mice inhibited tumor-forming capacity [57]. In another study, they report that decreased TIMP-3 expression increases interleukin-6 (IL-6) production, which promotes cell growth and invasion, decreased overall survival, and relapse-free survival periods in HPV-infected non-small cell lung cancer patients [58].
On the other hand, in an in vivo study using mice, they reported that silencing of TIMP-3 resulted in the suppression of mammary tumors and suggested that it was under the regulation of the Tnf or Tnfr1 genes [59]. It has also been reported that breast cancer patients with high TIMP-3 mRNA levels are associated with a better response to endocrine therapy with tamoxifen [60]. Interestingly, in biopsies of patients with head and neck carcinoma, overexpression of TIMP-3 mRNA is associated with poorer survival compared to those with low mRNA levels [61]. Interestingly, results found in other types of cancer show that TIMP-3 overexpression is associated with shorter survival. In contrast, in our study, we report that lower TIMP-3 expression is associated with decreased survival in breast cancer patients.
TIMP-4 is the least studied member of the TIMPs; however, elevated levels of TIMP-4 have been found in breast, ovarian, cervical, prostate, brain, colon, endometrial, and renal papillary tumors. In contrast, down-regulation was observed in pancreatic and renal clear cell tumors [62]. However, our results showed that TIMP-4 is downregulated in breast cancer patients relative to healthy patients, and this was associated with shorter overall survival. TIMP-4 showed indirect interaction with MMP-1, MMP-2, MMP-3, MMP-14, and MMP-16 and had the lowest number of protein interactions for the other TIMPs. Background reports that in nude mice with cervical cancer cell xenografts, overexpression of TIMP-4 induces tumor formation. In vitro, assays promote the enrichment of the cancer stem cell population through NFκB signaling [63]. On the other hand, they report that in patients with estrogen receptor-negative infiltrating ductal breast cancer, overexpression of TIMP-4 is associated with malignant progression and correlates with a low probability of long-term disease-free survival (more than 3 years) [64]. TIMP-4 also inhibits apoptosis of MDA-MB-435 breast cancer cells in vitro and in vivo employing a nude mouse model by regulating the expression of TIMP-4, which upregulates the expression of Bcl-2 and Bcl-xL [65].
Recently, research on the immune microenvironment of breast cancer has increased because immune infiltration is highly correlated with the diagnosis, progression, prognosis, and response to the treatment of breast cancer patients [66].
It has been shown that there is an increase in tumor-infiltrating DCs in both primary and metastatic lesions. It is associated with increased overall survival and decreased recurrence rate in breast cancer patients and was positively correlated with CD4+ and CD8+ T lymphocyte infiltration [67].
For this reason, we determined the relationship between TIMPs expression and immune infiltration of breast cancer. We found that TIMP-1 expression correlated positively with dendritic cell infiltrates and negatively correlated with B-cell infiltrates. In other cancer types, a positive relationship between TIMP-1 and dendritic cells has been reported in glioblastoma, adenocarcinoma of the stomach, and squamous cell carcinoma of the head and neck [68,69]. On the other hand, our results show that TIMP-2 expression correlates positively with the infiltration of CD8+ T cells, Neutrophils, and DCs. Interestingly, in stomach adenocarcinoma and colon adenocarcinoma, we also report that TIMP-2 was positively correlated with CD4+ T cells, CD8+ T cells, macrophages, neutrophils, and dendritic cells [70]. We also found that TIMP-3 expression levels positively correlate with CD8+ T-cell infiltrates. While in glioblastoma patients, TIMP-3 expression was associated with B cells, CD4+ T cells, macrophages, and neutrophils [68]. Finally, TIMP-4 expression levels positively correlate with cellular infiltrates of CD4+ T cells. However, in patients with glioblastoma, TIMP-4 expression was positively associated with B-cell, CD8+ T-cell, and macrophage infiltration [68]. These findings and previous reports describe the importance of immune system infiltration in prognosis and response to treatment in breast cancer.
5. Conclusions
The expression of different TIMPs is associated with shorter or longer overall survival of breast cancer patients. Our results show that overexpression of TIMP-1 is associated with longer patient survival, while low TIMP-2, TIMP-3, and TIMP-4 levels are associated with shorter survival of breast cancer patients. TIMPs were shown to directly interact with MMPs and proteins involved in functions that regulate breast cancer progression. Interestingly, TIMP-3 showed the highest interactions and regulation of cancer-related functions. The correlation between immune cell infiltrates and the expression of TIMPs is of great importance as they provide information on the diagnosis, prognosis, progression, and response to the treatment of breast cancer. Of the four TIMPs, TIMP-2 showed the highest positive correlation with infiltration of CD8+ immune cells, neutrophils, and dendritic cells; therefore, patients with overexpression of TIMP-2 have a better prognosis and greater response to treatment. TIMP-4 showed a lower correlation with immune infiltrates and thus would be associated with a lower response to treatment.
6. Significance of the study
TIMPs have a dual role in breast carcinogenesis, overexpression of TIMP-1 is associated with breast cancer progression and low patient survival. In contrast, low TIMP-2, TIMP-3, and TIMP-4 expression correlate with low patient survival. We propose TIMP-3 as a potential biomarker of breast cancer initiation and progression because of the number of interactions with target proteins related to carcinogenesis. Overexpression of TIMP-4 correlates with breast cancer dissemination. Direct interaction of TIMPs with MMPs regulates breast cancer dissemination in patients. The correlation between TIMP-2 expression and immune cell infiltrates is associated with better prognosis and response to treatment of breast cancer patients. On the other hand, low correlation of TIMP-4 with immune cell infiltrates is associated with low response to treatment.
Author Contributions
Conceptualization, formal analysis, Writing original draft, Writing, review and edition: L.C.S., A.A.I., and N.N.T., methodology, formal analysis and investigation: D.A.N.T., E.H.L., C.W.O., J.O.O, M.A.L.V., and M.A.M.C.
Acknowledgments
L.C.S. (No. 829247), A.A.I. (No. 666320) and D.A.N.T. (1043993) thank CONACYT for the scholarship of the doctoral students and Dr. E.H.L. for the postdoctoral fellowship.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sung, H., Ferlay, J., Siegel, R. L., Laversanne, M., Soerjomataram, I., Jemal, A., & Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA: A Cancer Journal for Clinicians 2021, 71, 209–249. [CrossRef]
- Siegel, R. L., Miller, K. D., Wagle, N. S., & Jemal, A. Cancer statistics, 2023. CA: A Cancer Journal for Clinicians 2023, 73, 17–48. [CrossRef]
- Yin, L., Duan, J. J., Bian, X. W., & Yu, S. C. Triple-negative breast cancer molecular subtyping and treatment progress. Breast Cancer Research 2020. [CrossRef]
- Schettini, F., Brasó-Maristany, F., Kuderer, N. M., & Prat, A. A perspective on the development and lack of interchangeability of the breast cancer intrinsic subtypes. npj Breast Cancer 2022, 8, 1–4. [CrossRef]
- Yersal, O., & Barutca, S. Biological subtypes of breast cancer: Prognostic and therapeutic implications. World Journal of Clinical Oncology 2014, 5, 412–424. [CrossRef]
- Min, S. K., Lee, S. K., Woo, J., Jung, S. M., Ryu, J. M., Yu, J., … Nam, S. J. Relation between tumor size and lymph node metastasis according to subtypes of breast cancer. Journal of Breast Cancer 2021. [CrossRef]
- Raeeszadeh-Sarmazdeh, M., Greene, K. A., Sankaran, B., Downey, G. P., Radisky, D. C., & Radisky, E. S. Directed evolution of the metalloproteinase inhibitor TIMP-1 reveals that its N- and C-terminal domains cooperate in matrix metalloproteinase recognition. Journal of Biological Chemistry 2019. [CrossRef]
- Cayetano-Salazar, L., Nava-Tapia, D. A., Astudillo-Justo, K. D., Arizmendi-Izazaga, A., Sotelo-Leyva, C., Herrera-Martinez, M., … Navarro-Tito, N. Flavonoids as regulators of TIMPs expression in cancer: Consequences, opportunities, and challenges. Life Sciences 2022. [CrossRef]
- Vandenbroucke, R. E., & Libert, C. Is there new hope for therapeutic matrix metalloproteinase inhibition? Nature Reviews Drug Discovery 2014, 13, 904–927. [CrossRef]
- Cabral-Pacheco, G. A., Garza-Veloz, I., Rosa, C. C. D. La, Ramirez-Acuña, J. M., Perez-Romero, B. A., Guerrero-Rodriguez, J. F., … Martinez-Fierro, M. L. The roles of matrix metalloproteinases and their inhibitors in human diseases. International Journal of Molecular Sciences 2020, 21, 1–53. [CrossRef]
- Jackson, H. W., Defamie, V., Waterhouse, P., & Khokha, R. TIMPs: Versatile extracellular regulators in cancer. Nature Reviews Cancer 2017, 17, 38–53. [CrossRef]
- Wang, X., & Khalil, R. A. Matrix Metalloproteinases, Vascular Remodeling, and Vascular Disease. In Advances in Pharmacology, 1st ed.; Elsevier Inc., 2018; Vol. 81. [CrossRef]
- Bourboulina, D., & Stetler-Stevenson, W. G. Matrix MetalloProteinases (MMPs) andTissue Inhibitors of MetalloProteinases (TIMPs): positive and negative regulators intumor cell adhesion. Semin Cancer Biol 2011, 20, 161–168. [CrossRef]
- Shimoda, M. Extracellular vesicle-associated MMPs: A modulator of the tissue microenvironment. In Advances in Clinical Chemistry, 1st ed.; Elsevier Inc., 2019; Vol. 88. [Google Scholar] [CrossRef]
- Wang, D. D., Xu, W. X., Chen, W. Q., Li, L., Yang, S. J., Zhang, J., & Tang, J. H. Identification of TIMP2 as a Prognostic Biomarker and Its Correlation with Tumor Immune Microenvironment: A Comprehensive Pan-Cancer Analysis. Journal of Oncology 2022, 2022. [CrossRef]
- Chen, W. Q., Yang, S. J., Xu, W. X., Deng, F., Wang, D. D., Tang, J. H., & Ding, J. Bioinformatics analysis revealing prognostic significance of TIMP2 gene in breast cancer. Medicine (United States) 2021, 100, E27489. [CrossRef]
- Tang, Z., Li, C., Kang, B., Gao, G., Li, C., & Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Research 2017, 45, W98–W102. [CrossRef]
- Lánczky, A., & Győrffy, B. Web-based survival analysis tool tailored for medical research (KMplot): Development and implementation. Journal of Medical Internet Research 2021, 23, 1–7. [CrossRef]
- Oughtred, R., Stark, C., Breitkreutz, B. J., Rust, J., Boucher, L., Chang, C., … Tyers, M. The BioGRID interaction database: 2019 update. Nucleic Acids Research 2019, 47, D529–D541. [CrossRef]
- Paul Shannon, 1, Andrew Markiel, 1, Owen Ozier, 2 Nitin S. Baliga, 1 Jonathan T. Wang, 2 Daniel Ramage, 2, Nada Amin, 2, Benno Schwikowski, 1, 5 and Trey Ideker2, 3, 4, 5, 山本隆久, … 大森敏行. Cytoscape: A Software Environment for Integrated Models. Genome Research 1971, 13, 426.
- Warde-Farley, D., Donaldson, S. L., Comes, O., Zuberi, K., Badrawi, R., Chao, P., … Morris, Q. The GeneMANIA prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Research 2010, 38 SUPPL. 2, 214–220. [CrossRef]
- Li, T., Fan, J., Wang, B., Traugh, N., Chen, Q., Liu, J. S., … Liu, X. S. TIMER: A web server for comprehensive analysis of tumor-infiltrating immune cells. Cancer Research 2017, 77, e108–e110. [CrossRef]
- Chen, X., Zhong, S. L., Lu, P., Wang, D. D., Zhou, S. Y., Yang, S. J., … Tang, J. H. miR-4443 participates in the malignancy of breast cancer. PLoS ONE 2016. [CrossRef]
- Harbeck, N., Penault-Llorca, F., Cortes, J., Gnant, M., Houssami, N., Poortmans, P., … Cardoso, F. Breast cancer. Nature Reviews Disease Primers 2019, 5. [CrossRef]
- Feng, Y., Spezia, M., Huang, S., Yuan, C., Zeng, Z., Zhang, L., … Ren, G. Breast cancer development and progression: Risk factors, cancer stem cells, signaling pathways, genomics, and molecular pathogenesis. Genes and Diseases 2018, 5, 77–106. [CrossRef] [PubMed]
- Dechaphunkul, A., Phukaoloun, M., Kanjanapradit, K., Graham, K., Ghosh, S., Santos, C., & Mackey, J. R. Prognostic Significance of Tissue Inhibitor of Metalloproteinase-1 in Breast Cancer. International Journal of Breast Cancer 2012. [CrossRef]
- Guda, P., Chittur, S. V., & Guda, C. Comparative Analysis of Protein-Protein Interactions in Cancer-Associated Genes. Genomics, Proteomics and Bioinformatics 2009. [CrossRef]
- Goff, S. L., & Danforth, D. N. The Role of Immune Cells in Breast Tissue and Immunotherapy for the Treatment of Breast Cancer. Clinical Breast Cancer 2021. [CrossRef]
- Gao, Z. hua, Li, C. xin, Liu, M., & Jiang, J. yuan. Predictive and prognostic role of tumour-infiltrating lymphocytes in breast cancer patients with different molecular subtypes: a meta-analysis. BMC Cancer 2020. [CrossRef]
- Song, G., Xu, S., Zhang, H., Wang, Y., Xiao, C., Jiang, T., … Wang, X. TIMP1 is a prognostic marker for the progression and metastasis of colon cancer through FAK-PI3K/AKT and MAPK pathway. Journal of Experimental & Clinical Cancer Research 2016, 35, 148. [CrossRef]
- Cheng, G., Fan, X., Hao, M., Wang, J., Zhou, X., & Sun, X. Higher levels of TIMP-1 expression are associated with a poor prognosis in triplenegative breast cancer. Molecular Cancer 2016. [CrossRef]
- Jung, Y. S., Liu, X. W., Chirco, R., Warner, R. B., Fridman, R., & Kim, H. R. C. TIMP-1 induces an EMT-like phenotypic conversion in MDCK cells independent of its MMP-inhibitory domain. PLoS ONE 2012, 7. [CrossRef]
- D’Angelo, R. C., Liu, X. W., Najy, A. J., Jung, Y. S., Won, J., Chai, K. X., … Kim, H. R. C. TIMP-1 via TWIST1 induces EMT phenotypes in human breast epithelial cells. Molecular Cancer Research 2014, 12, 1324–1333. [CrossRef]
- Wu, Z. S., Wu, Q., Yang, J. H., Wang, H. Q., Ding, X. D., Yang, F., & Xu, X. C. Prognostic significance of MMP-9 and TIMP-1 serum and tissue expression in breast cancer. International Journal of Cancer 2008, 122, 2050–2056. [CrossRef]
- Talvensaari-Mattila, A., & Turpeenniemi-Hujanen, T. High preoperative serum TIMP-1 is a prognostic indicator for survival in breast carcinoma. Breast Cancer Research and Treatment 2005, 89, 29–34. [CrossRef]
- Voorzanger-Rousselot, N., Juillet, F., Mareau, E., Zimmermann, J., Kalebic, T., & Garnero, P. Association of 12 serum biochemical markers of angiogenesis, tumour invasion and bone turnover with bone metastases from breast cancer: A crossectional and longitudinal evaluation. British Journal of Cancer 2006, 95, 506–514. [CrossRef]
- Würtz, S., Schrohl, A. S., Mouridsen, H., & Brünner, N. TIMP-1 as a tumor marker in breast cancer - An update. Acta Oncologica 2008, 47, 580–590. [CrossRef]
- Liu, X. W., Bernardo, M. M., Fridman, R., & Kim, H. R. C. Tissue inhibitor of metalloproteinase-1 protects human breast epithelial cells against intrinsic apoptotic cell death via the focal adhesion kinase/phosphatidylinositol 3-kinase and MAPK signaling pathway. Journal of Biological Chemistry 2003, 278, 40364–40372. [CrossRef]
- Klintman, M., Würtz, S. Ø., Christensen, I. J., Hertel, P. B., Fernö, M., Malmberg, M., … Brünner, N. Association between tumor tissue TIMP-1 levels and objective response to first-line chemotherapy in metastatic breast cancer. Breast Cancer Research and Treatment 2010, 121, 365–371. [CrossRef]
- Schrohl, A. S., Meijer-Van Gelder, M. E., Holten-Andersen, M. N., Christensen, I. J., Look, M. P., Mouridsen, H. T., … Foekens, J. A. Primary tumor levels of tissue inhibitor of metalloproteinases-1 are predictive of resistance to chemotherapy in patients with metastatic breast cancer. Clinical Cancer Research 2006. [CrossRef]
- Balkhi, S., Mashayekhi, F., Salehzadeh, A., & Saeidi Saedi, H. TIMP1 and TIMP3 circulating levels and promoter polymorphisms in breast cancer. British Journal of Biomedical Science 2021, 78, 236–238. [CrossRef]
- Escalona, R. M., Bilandzic, M., Western, P., Kadife, E., Kannourakis, G., Findlay, J. K., & Ahmed, N. TIMP-2 regulates proliferation, invasion and STAT3-mediated cancer stem cell-dependent chemoresistance in ovarian cancer cells. BMC Cancer 2020, 20, 960. [CrossRef]
- Wang, W., Li, D., Xiang, L., Lv, M., Tao, L., Ni, T., … Zhou, Y. TIMP-2 inhibits metastasis and predicts prognosis of colorectal cancer via regulating MMP-9. Cell Adhesion and Migration 2019, 13, 273–284. [CrossRef]
- Imren, S., Kohn, D. B., Shimada, H., Blavier, L., & DeClerck, Y. A. Overexpression of tissue inhibitor of metalloproteinases-2 retroviral-mediated gene transfer in vivo inhibits tumor growth and invasion. Cancer research 1996, 56, 2891–5.
- Peeney, D., Jensen, S. M., Castro, N. P., Kumar, S., Noonan, S., Handler, C., … Stetler-Stevenson, W. G. TIMP-2 suppresses tumor growth and metastasis in murine model of triple-negative breast cancer. Carcinogenesis 2020, 41, 313–325. [CrossRef]
- Ree, A. H., Flørenes, V. A., Berg, J. P., Mælandsmo, G. M., Nesland, J. M., & Fodstad, Ø. High levels of messenger RNAs for tissue inhibitors of metalloproteinases (TIMP-1 and TIMP-2) in primary breast carcinomas are associated with development of distant metastases. Clinical Cancer Research 1997.
- Remacle, A., McCarthy, K., Noël, A., Maguire, T., McDermott, E., O’Higgins, N., … Duffy, M. J. High levels of TIMP-2 correlate with adverse prognosis in breast cancer. International Journal of Cancer 2000, 86, 118–121.
- Shen, Q., Lee, E. S., Pitts, R. L., Wu, M. H., & Yuan, S. Y. Tissue inhibitor of metalloproteinase-2 regulates matrix metalloproteinase-2-mediated endothelial barrier dysfunction and breast cancer cell transmigration through lung microvascular endothelial cells. Molecular Cancer Research 2010, 8, 939–951. [CrossRef]
- Noel, A., Maillard, C., Rocks, N., Jost, M., Chabottaux, V., Sounni, N. E., … Foidart, J. M. Membrane associated proteases and their inhibitors in tumour angiogenesis. Journal of Clinical Pathology 2004, 57, 577–584. [CrossRef]
- Gu, X., Fu, M., Ding, Y., Ni, H., Zhang, W., Zhu, Y., … Zhu, J. TIMP-3 expression associates with malignant behaviors and predicts favorable survival in HCC. PLoS ONE 2014, 9, 1–8. [CrossRef]
- Su, C. W., Su, B. F., Chiang, W. L., Yang, S. F., Chen, M. K., & Lin, C. W. Plasma levels of the tissue inhibitor matrix metalloproteinase-3 as a potential biomarker in oral cancer progression. International Journal of Medical Sciences 2017, 14, 37–44. [CrossRef]
- Edwards, D.R. TIMP-3 and endocrine therapy of breast cancer: An apoptosis connection emerges. Journal of Pathology 2004, 202, 391–394. [Google Scholar] [CrossRef]
- Su, C. W., Lin, C. W., Yang, W. E., & Yang, S. F. TIMP-3 as a therapeutic target for cancer. Therapeutic Advances in Medical Oncology 2019. [CrossRef]
- Baker, A. H., George, S. J., Zaltsman, A. B., Murphy, G., & Newby, A. C. Inhibition of invasion and induction of apoptotic cell death of cancer cell lines by overexpression of TIMP-3. British Journal of Cancer 1999, 79, 1347–1355. [CrossRef]
- Kang, S. H., Choi, H. H., Kim, S. G., Jong, H. S., Kim, N. K., Kim, S. J., & Bang, Y. J. Transcriptional inactivation of the tissue inhibitor of metalloproteinase-3 gene by DNA hypermethylation of the 5’-CpG island in human gastric cancer cell lines. International Journal of Cancer 2000, 86, 632–635.
- Ninomiya, I., Kawakami, K., Fushida, S., Fujimura, T., Funaki, H., Takamura, H., … Ohta, T. Quantitative detection of TIMP-3 promoter hypermethylation and its prognostic significance in esophageal squamous cell carcinoma. Oncology Reports 2008. [CrossRef]
- Bian, J., Wang, Y., Smith, M. R., Kim, H., Jacobs, C., Jackman, J., … Sun, Y. Suppression of in vivo tumor growth and induction of suspension cell death by tissue inhibitor of metalloproteinases (TIMP)-3. Carcinogenesis 1996, 17, 1805–1811. [CrossRef]
- Wu, D. W., Tsai, L. H., Chen, P. M., Lee, M. C., Wang, L., Chen, C. Y., … Lee, H. Loss of TIMP-3 promotes tumor invasion via elevated IL-6 production and predicts poor survival and relapse in HPV-infected non-small cell lung cancer. American Journal of Pathology 2012, 181, 1796–1806. [CrossRef]
- Jackson, H. W., Hojilla, C. V., Weiss, A., Sanchez, O. H., Wood, G. A., & Khokha, R. Timp3 deficient mice show resistance to developing breast cancer. PLoS ONE 2015, 10, 1–14. [CrossRef]
- Span, P. N., Lindberg, R. L. P., Manders, P., Tjan-Heijnen, V. C. G., Heuvel, J. J. T. M., Beex, L. V. A. M., & Sweep, C. G. J. F. Tissue inhibitors of metalloproteinase expression in human breast cancer: TIMP-3 is associated with adjuvant endocrine therapy success. Journal of Pathology 2004, 202, 395–402. [CrossRef]
- Kornfeld, J. W., Meder, S., Wohlberg, M., Friedrich, R. E., Rau, T., Riethdorf, L., … Riethdorf, S. Overexpression of TACE and TIMP3 mRNA in head and neck cancer: Association with tumour development and progression. British Journal of Cancer 2011, 104, 138–145. [CrossRef] [PubMed]
- Melendez-zajgla, J., Pozo, L. Del, Ceballos, G., & Maldonado, V. Tissue Inhibitor of Metalloproteinases-4. The road less traveled. Molecular Cancer 2008, 11, 1–11. [CrossRef]
- Lizarraga, F., Espinosa, M., Ceballos-Cancino, G., Vazquez-Santillan, K., Bahena-Ocampo, I., Schwarz-Cruz y Celis, A., … Melendez-Zajgla, J. Tissue inhibitor of metalloproteinases-4 (TIMP-4) regulates stemness in cervical cancer cells. Molecular Carcinogenesis 2016, 55, 1952–1961. [CrossRef] [PubMed]
- Liss, M., Sreedhar, N., Keshgegian, A., Sauter, G., Chernick, M. R., Prendergast, G. C., & Wallon, U. M. Tissue inhibitor of metalloproteinase-4 is elevated in early-stage breast cancers with accelerated progression and poor clinical course. American Journal of Pathology 2009, 175, 940–946. [CrossRef]
- Jiang, Y., Wang, M., Çeliker, M. Y. Ç., Liu, Y. E., Sang, Q. X. A., Goldberg, I. D., & Shi, Y. E. Stimulation of mammary tumorigenesis by systemic tissue inhibitor of matrix metalloproteinase 4 gene delivery. Cancer Research 2001.
- Dieci, M. V., Miglietta, F., & Guarneri, V. Immune infiltrates in breast cancer: Recent updates and clinical implications. Cells 2021, 10, 1–27. [CrossRef]
- La Rocca, G., Anzalone, R., Corrao, S., Magno, F., Rappa, F., Marasà, S., … Cappello, F. CD1a down-regulation in primary invasive ductal breast carcinoma may predict regional lymph node invasion and patient outcome. Histopathology 2008, 52, 203–212. [CrossRef]
- Han, J., Jing, Y., Han, F., & Sun, P. Comprehensive analysis of expression, prognosis and immune infiltration for TIMPs in glioblastoma. BMC Neurology 2021, 21, 1–15. [CrossRef]
- Liu, L., Yang, S., Lin, K., Yu, X., Meng, J., Ma, C., … Jin, D. Sp1 induced gene TIMP1 is related to immune cell infiltration in glioblastoma. Scientific Reports 2022, 12, 1–18. [CrossRef]
- Jian, F., Yanhong, J., Limeng, W., Guoping, N., Yiqin, T., Hao, L., & Zhaoji, P. TIMP2 is associated with prognosis and immune infiltrates of gastric and colon cancer. International Immunopharmacology 2022, 110. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



