
Submitted:
21 June 2023
Posted:
25 June 2023
You are already at the latest version
Abstract
Filamentous manganese (Mn) oxide particles, which occur in the suboxic zone of stratified waterbodies, are important drivers of diverse elemental cycles. Although these particles are considered to be bacteriogenic, the environmental factor responsible for their formation has not been identified. Based on this study of laboratory cultures of a model Mn(II)-oxidizing bacterium, the supply of algal extracellular polymeric substances (EPS) was shown to stimulate Mn(II) oxidation and thus the production of filamentous Mn oxide particles. This observation was consistent with results obtained for naturally occurring particles collected from a near-bottom layer (depth of ~90 m) in the northern basin of Lake Biwa, Japan, that is, most Mn particles resembling delta-MnO2 were associated with an EPS-like gelatinous matrix, which contained dead algal cells and was lectin-stainable. In the lake water column, EPS produced by photosynthesis sank to the bottom layer. The analysis of the quality of water samples, which have been collected from the study site for 18 years, reveals that the annual average total phytoplankton biovolume in the surface layer correlates with the density of filamentous Mn particles in the near-bottom layer. Among different phytoplankton species, green algae appeared to be the key species. The results of this study suggest that algal EPS production induces the formation of filamentous Mn oxide particles in the near-bottom layer of the northern basin of Lake Biwa.
Keywords:
biogenic manganese oxides
; manganese(II)-oxidizing bacterium
; Metallogenium-like particles
; total polysaccharide content
; transparent exopolymer particles
1. Introduction
The manganese (Mn) redox process occurring at the oxic-anoxic interface of stratified waterbodies drives the biogeochemical cycles of numerous elements dissolved in water. Once dissolved Mn(II) ions diffuse from the anoxic layer or sediment to the upper suboxic layer, they are oxidized to form particulate Mn(III, IV) oxides. The Mn oxides serve as scavengers of numerous metal ions and as oxidants of organic and inorganic substances [1,2]. When Mn oxide particles sink to the anoxic layer, they are reduced to Mn(II) ion by an inorganic reductant (i.e., sulfide) or microorganisms with a dissimilatory Mn-reducing activity. Thus, the Mn cycling at the oxic-anoxic interface affects both chemical and biological processes in the water environment (e.g., [3,4,5,6,7,8,9]).
The Mn oxide particles in the oxic layer often have filamentous structures, which are referred to as “Metallogenium”-like particles [10,11,12,13,14]. Despite the importance of biogeochemical implications, the particle formation mechanism remains an enigma [15]. The results of a recent study demonstrated that a Mn(II)-oxidizing alphaproteobacterium, Bosea sp. BIWAKO-01, produces filamentous Mn particles under laboratory culture conditions [13]. The Mn(II) oxidation and filament formation occurred in static cultures and proceeded faster under CO2-rich, low-O2 (5%-10% in air) conditions. The CO2 dependency resulted from the decrease in the cultural pH below 6.0 via carbonation, yielding an optimum pH for Mn(II) oxidation by Bosea sp. BIWAKO-01 [13]. Interestingly, a relatively low content of agar (e.g., 500 mg L-1) is needed for microbial Mn(II) oxidation. Similar results were reported for other Mn(II)-oxidizing bacteria [16]. It remains unknown whether these bacteria serve as producers of filamentous Mn particles in the water environment. However, the results obtained for these bacterial cultures indicate that environmental factors trigger the bacterial production of particles.
Given that a low concentration of polysaccharides stimulate the Mn(II) oxidation in certain bacterial cultures [13,16], such substances may be responsible for the occurrence of Mn particles in the environment. Diverse phytoplankton species produce extracellular polymeric substances (EPS), which mainly consist of acidic polysaccharides [17]. Such gelatinous matter is ubiquitous in form of transparent exopolymer particles (TEP) in aquatic environments [18]. The results of previous research indicated their significant contribution to the organic carbon pool in aquatic environments [18,19]. These exopolymers provide microhabitats for the colonization by bacteria, leading to high metabolic activity at microparticle surfaces [20,21,22,23]. The effects of phytoplankton or EPS on the formation of filamentous Mn particles in stratified waterbodies have not been considered, although laboratory culture experiments [13,16] suggest their contribution.
The results of several studies that were carried out in both the southern and northern basins of Lake Biwa, Japan, indicated the presence of filamentous Mn-rich particles in hypoxic layers [24,25,26]. Our working hypothesis is that algal EPS produced in the epilimnion reach the lower layer by sinking and induce the microbial formation of filamentous Mn particles. In this study, the significance of algal EPS for the Mn particle formation was investigated using laboratory cultures of Mn(II)-oxidizing bacterium Bosea sp. BIWAKO-01. Microscopic analysis was used to determine the structural features of naturally occurring Mn filaments ,which were collected at the Imazuoki-chuo point in the northern basin of Lake Biwa. The spatial and temporal distribution of polysaccharides in the water column was monitored at the study site to show that algal EPS are present in the near-bottom layer in which filamentous Mn particles occur. Furthermore, we used water quality data that have been obtained in the past 18 years at the study site to examine whether the particle formation correlates with the growth of phytoplankton in the surface layer. The results of this study support the significance of algal EPS.
2. Materials and Methods
2.1. Culture experiments
Cultures of a Mn(II)-oxidizing alphaproteobacterium, Bosea sp. strain BIWAKO-01, were used as a laboratory model system to yield filamentous Mn particles. Strain BIWAKO-01 was statically cultured in petri dishes in the dark at 10% O2 and 20 °C, as previously described [13]. The M3 liquid culture medium [16] contained 100 mg L-1 malt extract, 40 mg L-1 yeast extract, 0.5 mM NaHCO3, and 500 mg L-1 agar. Filter-sterilized MnSO4 solution was added to the medium at 0.1 or 2 mM.
To examine the effect of the addition of algal biomass on bacterial Mn(II) oxidation, the 20 mL M3 medium from which agar was omitted was inoculated with BIWAKO-01 at 1.6 × 106 CFU (CFU: colony forming unit), along with green algal species, Staurastrum arctiscon or S. dorsidentiferum, which were isolated from the water of Lake Biwa and stored at the Lake Biwa Environmental Research Institute (LBERI; Shiga, Japan). These green algae were grown in CT medium [27] (Table S1) at 20 °C using 12 h light-dark cycles. The light intensity was set to 60 µmol m-2 s-1. After three weeks, the cell densities of S. arctiscon and S. dorsidentiferum reached 1.6 × 104 and 1.5 × 104 cells mL-1, respectively, and 1 mL of these cultures were transferred into 20 mL of BIWAKO-01 cultures. In this experiment, 2 mM MnSO4 was added to the culture in the beginning. For the removal of extracellular mucilage sheaths from algal cells, S. arctiscon and S. dorsidentiferum cells were harvested by filtration using glass fiber filters (Whatman GF/B; GE Healthcare, Buckinghamshire, UK) and then repeatedly washed by injecting sterile distilled water using a spray bottle [28] until the extracellular sheath became invisible under the light microscope when stained with India ink (see below). The cells on each filter were rinsed and resuspended in CT medium to obtain a cell density of 1.5 × 104 cells mL-1. The culture supernatant fluid was removed by centrifugation at 10,000 × g for 10 min at 4 °C and the oxidized Mn was determined spectrophotometrically using leucoberbelin blue [29]. For this assay, KMnO4 solution was used as standard.
2.2. Electron and light microscopy
Filamentous Mn oxide particles were filtered through a carbon-coated membrane filter (Nisshin EM, Tokyo, japan) and vacuum dried. The particles were analyzed with a transmission electron microscope (TEM; JEM-2100F, JEOL, Tokyo, Japan) at 200 kV and selected area electron diffraction (SAED).
Suspended solids containing gelatinous organic substances were stained with India ink and observed using a differential interference microscope (Eclipse 80i, Nikon, Tokyo, Japan). Under the microscope, transparent gelatinous substances became visible; after staining, they appeared white with respect to the background. The densities of the filamentous Mn particles in the water samples were measured with the microscope at 100-200× magnification. Lectin staining and fluorescence microscopy were used to detect polysaccharidic substances, as described previously [13,30]. This assay was conducted with a BX60 epifluorescence microscope equipped with a DP70 charge coupled device camera (Olympus, Tokyo, Japan). The suspended matter including filamentous Mn particles collected in the near-bottom layer of Lake Biwa was gently rinsed with deionized water and stained for 15 min at 23 °C with fluorescein-conjugated Lycopersicon esculentum lectin (LEL), Lens culinaris agglutinin (LCA), Phaseolus vulgaris erythroagglutinin (PHA-E), soy bean agglutinin (SBA), or peanut agglutinin (PNA). These lectins were purchased from Vector Laboratories (Burlingame, CA, USA). Epifluorescence microscopy was conducted at an excitation wavelength of 470 to 490 nm.
2.3. Study site and available water quality data
Lake Biwa is a warm monomictic lake in the Shiga Prefecture, Japan, with an area of 670 km2 and maximum depth of 104 m. The study site was the Imazuoki-chuo point in the northern basin of Lake Biwa (35°23′41″ N, 136°07′57″ E). For the fixed-point study, the water quality was bimonthly monitored by the LBERI at depths of 0.5, 5, 10, 15, 20, 30, 40, 60, 80, 85, and 90 m. Because the water depth in this area is ~90 m, the 90-m sample was collected 1 m above the bottom of lake. We monitored the vertical distribution of polysaccharides at the study site from August 2014 to February 2020. The water samples collected at the above-mentioned depths, except for the 30-m sample, were provided by the LBERI.
To investigate the environmental factors affecting the occurrence of filamentous Mn particles, LBERI’s database was used. The database contains water quality data obtained on samples that were collected twice a month at the study site from April 2002 to February 2020 including the pH and dissolved oxygen (DO), total organic carbon (TOC), total nitrogen (TN), total phosphorus (TP), and chlorophyll a (Chl.a) contents. These data are partially available at a website [31]. In addition, 18-year data of the abundance of filamentous Mn particles observed at a depth of 90 m and the biovolume of phytoplankton at 0.5 m were used. The total biovolume was expressed as the sum of the biovolumes of Cyanophyceae (cyanobacteria), Chlorophyceae (including Charophyceae; green algae), Bacillariophyceae (diatoms), Chrysophyceae, Dinophyceae, and Cryptophyceae, which were calculated from the cellular or colony sizes, as described elsewhere (Kishimoto et al. 2013). The biovolume data obtained from 2002 to 2009 at the Imazuoki-chuo point were published previously [32,33].
The water stratification that develops during the summer season at the study site results in the depletion of DO in the near-bottom layer, which is placed under hypoxic or subhypoxic conditions in the fall and winter months (Figure 1). During this period, Mn2+ ions are eluted from lake sediments rich in Mn oxides and diffuse to and are reoxidized in the oxic or suboxic near-bottom layer [34]. Subsequently, the stratified water column overturns usually until late winter, as represented by the bottom-layer DO content in Figure 1. Therefore, the Mn particle density decreases to a nondetectable level via dispersion. The annual cycle of filamentous Mn particle formation in the near-bottom layer proceeds during the period of the complete overturn to the next complete overturn. In this study, the annual average values of the density of filamentous Mn particles and other water quality parameters were calculated using data obtained during one annual cycle of Mn particle formation (counted from February or March of the present year to the next; Figure 1).
2.4. Analysis of polysaccharides
For the analysis of the polysaccharides, 250 to 300 mL of the water samples were freeze-dried. The total solids were suspended in 10 mL of methanol to concentrate the contents 25 to 30 times. A portion of the methanol suspension was transferred into a glass tube and dried under a gentle N2 stream. The residual solids were then resuspended in 2.0 mL of deionized water. The total polysaccharide concentrations were determined with an anthrone reagent [35]. Briefly, the concentrated suspension (2 mL) in a glass tube was placed in an ice bath for 5 min and 4 mL of an anthrone reagent containing 0.2 g anthrone in 100 mL of 95% sulfuric acid was slowly added to the glass tube and then mixed vigorously. The glass tube was heated for 10 min using a boiling water bath and allowed to cool at room temperature. The absorbance at 625 nm was determined with a spectrophotometer. The measurements were conducted in triplicate. Glucose solution was used as standard.
The neutral monosaccharide composition was analyzed using a previously described procedure [36]. Briefly, the water samples were freeze-dried and the solids were hydrolyzed with 4 M trifluoroacetic acid at 100 °C for 3 h. The alditol acetates were separated and quantified using the Development and Assessment of Sustainable Humanosphere system (DASH) at Kyoto University, that is, a gas chromatograph-mass spectrometer system QP2010 (Shimadzu, Kyoto, Japan) equipped with a capillary column (SP-2330, 0.25 mm × 0.2 μm × 15 m; Sigma-Aldrich, St. Louis, MO, USA). The alditol acetates were identified based on their retention times and mass spectra.
2.5. Statistical analysis
Principal component analysis (PCA) was performed to identify the variables that positively correlate with the Mn particles. The annual average values of water quality parameters, including the pH, DO, TOC, TN, TP, and algal volume, were used for the first PCA and the biovolume of each algal species was utilized for the second PCA. The analysis was conducted using PRIMER 6 software (Primer-E, Plymouth, UK).
3. Results
3.1. Filamentous Mn particle formation in laboratory model system
The Mn(II)-oxidizing bacterium Bosea sp. BIWAKO-01was cultured in a medium containing agar to which 0.1 mM dissolved Mn(II) was intermittently added for 14 days to obtain a total dose of 0.7 mM (Figure S1). The cumulative formation of oxidized Mn due to the addition of Mn(II) indicates that the dissolved Mn(II) ions in the medium are mostly oxidized and maintained at low concentrations. The TEM observations reveal that the filamentous particles consist of nano-sized sheets, which are consistent with the structure of the biogenic layer-type Mn oxide mineral δ-MnO2 (Figure 2).
The results showed that the strain BIWAKO-01 cannot oxidize Mn(II) in the absence of agar during the culture period, as previously reported [13]. However, this strain oxidized Mn(II) in the agar-free medium when supplemented with a green algal species, that is, S. dorsidentiferum or S. arctiscon. When Mn(II) was added at 2 mM, the oxidized Mn content reached 1.2 ± 0.3 and 0.7 ± 0.2 mM (mean ± standard deviation; n = 3) in the presence of S. dorsidentiferum and S. arctiscon, respectively (Figure 3). A large proportion of the Mn particles was in localized contact with extracellular mucilage sheaths of algal cells, which appeared white with respect to the background after India ink staining. The content of oxidized Mn was below the detection level (< 0.01 mM) in the cultures with naked algal cells whose extracellular mucilage sheaths were extensively removed by spraying with deionized water. Filamentous particles were not observed in these cultures, demonstrating that the significance of EPS produced by algal cells for the Mn(II) oxidation and production of filamentous Mn particles by the bacterium.
3.2. Filamentous Mn particles collected in the near-bottom layer of Lake Biwa
The sizes of naturally occurring filamentous particles collected from the near-bottom layer ranged from ~5 to 20 μm (Figure 4). The particle suspensions reacted with leucoberbelin blue, indicating the presence of oxidized Mn. The TEM and SAED analyses reveal that the Mn mineral is δ-MnO2 with d values of 0.253 and 0.148 nm (Figure S2). The filamentous particles coexisted with aggregates including fine solids and gelatinous substances, which became visible after India ink staining (Figure 4). Several dead cells of green algae and diatoms (frustules) were observed in the aggregates. Staining with various lectins including LEL (recognizing N-acetylglucosamine; Figure 4), LCA (α-mannose and α-glucose; Figure S3), PHA-E and SBA (N-acetylgalactosamine), and PNA (galactose) suggests that the organic matrix of the aggregates contains these polysaccharidic moieties. These lectins also bind to the filaments.
3.3. Distribution of polysaccharides in the water column of Lake Biwa
The spatiotemporal distribution of total polysaccharides in the water column was monitored from August 2014 to March 2020. The data are shown in Figure 5 along with the Chl.a concentrations and filamentous Mn particle densities derived from LBERI’s database. During the study period, the total polysaccharide concentration at a depth of 0.5 m was 0.51 ± 0.15 mg L-1 (n = 14) in the spring season (March to May), 0.76 ± 0.29 mg L-1 (n = 16) in the summer season (June to August), 0.60 ± 0.12 mg L-1 (n = 18) in the fall season (September to November), and 0.45 ± 0.18 mg L-1 (n = 18) in the winter season (December to February). From August 2014 to February 2020, the total polysaccharide concentration at a depth of 0.5 m strongly correlated with the biovolume of phytoplankton cells (r = 0.772, P < 0.001; Figure 6) and less but significantly correlated with the Chl.a concentration (r = 0.377, P = 0.002). Among the phytoplankton species, green algae were the most dominant with respect to the total biovolume and significantly correlated with the total polysaccharides (r = 0.723, P < 0.001; Figure 6), suggesting that these species mainly contribute to the production of polysaccharides in the surface layer. The profiles in Figure 5 also show that a certain part of polysaccharides in the surface layer sinks to the near-bottom layer. During the monitoring term, the total polysaccharide concentration at a depth of 90 m ranged from 0.17 to 0.64 mg L-1 (0.36 ± 0.10 mg L-1; n = 68).
The total polysaccharides obtained at depths of 0.5 and 90 m included rhamnose, fucose, arabinose, xylose, mannose, galactose, and glucose (Figure S4). The compositions of these saccharides temporally changed but insignificantly differed spatially, although two 90-m samples from July 2015 and December 2016 contained higher glucose concentrations. In general, the saccharide compositions at 0.5 m were similar to those at 90 m. Significant correlations were observed for rhamnose (r = 0.561; P = 0.001), fucose (r = 0.456; P = 0.010), arabinose (r = 0.360; P = 0.046), xylose (r = 0.797; P < 0.001), and mannose (r = 0.503; P = 0.004).
Typically, the densities of filamentous Mn particles in the near-bottom layer are high from late fall to winter due to the DO depletion (Figure 1 and Figure 5). This period is inconsistent with that during which high total polysaccharide concentrations were observed in the bottom layer (Figure 5). The correlation between the density of filamentous Mn particles and the total polysaccharide concentration at 90 m was insignificant (P > 0.1).
3.4. Correlation between filamentous Mn particles and the phytoplankton biovolume in the past 18 years
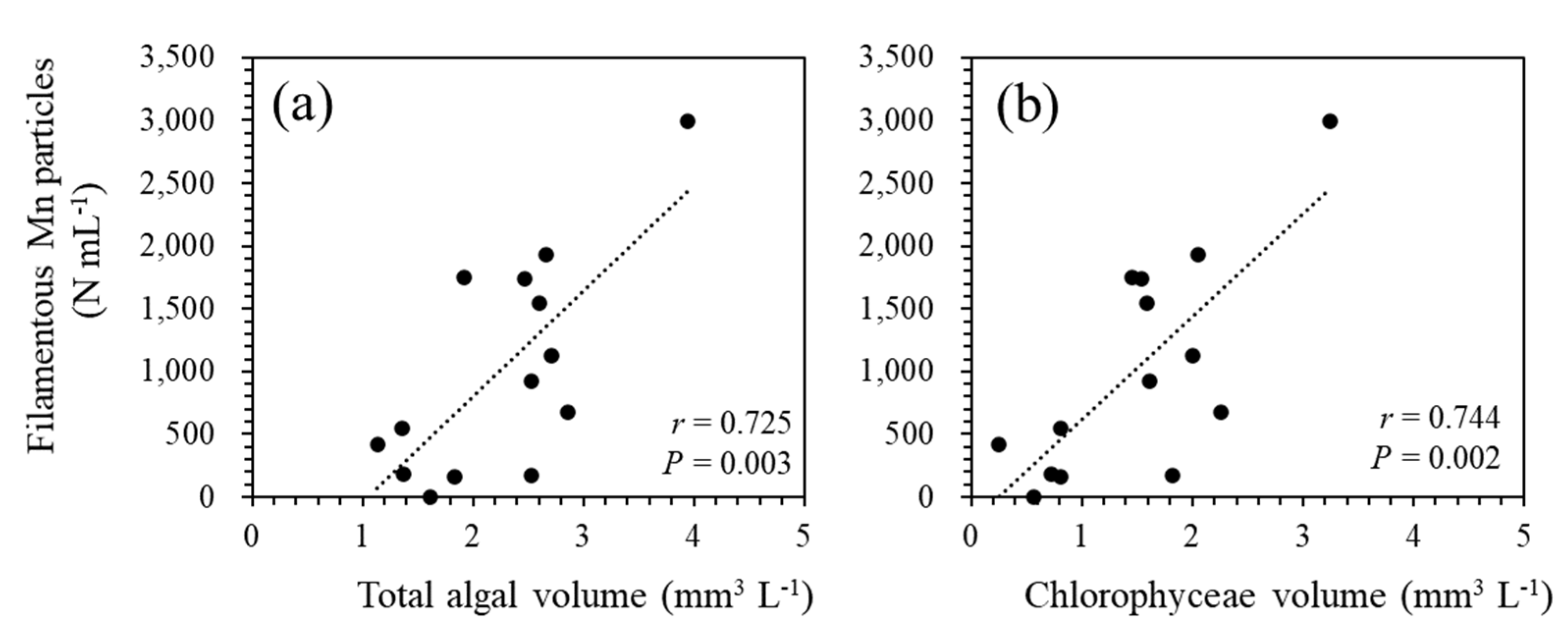
Among the 18-year water quality data, three periods (i.e., March 2009-February 2010, March 2010-February 2011, and February 2013-February 2014; Figure 1) were excluded from the calculation because of the lack of more than three data points during the emergence of filamentous Mn particles. The period June 2019-March 2020 was also excluded because of the incomplete overturn of the water column (Figure 1), which resulted in the chronic formation of particles in the near-bottom layer. The PCA of annual average data (n = 14; Figure 7) reveals the relationship between the density of filamentous Mn particles at 90 m and the total algal biovolume at 0.5 m. Among different algal species, this relationship was observed for the biovolume of green algae (Figure 7). The coefficients of the correlations between Mn particles and the total algal biovolume and green algal biovolume were determined to be 0.725 (P = 0.003) and 0.744 (P = 0.002), respectively (Figure 8). These results are consistent with the contribution of phytoplankton (green algae) in the surface layer to the formation of filamentous Mn particles. The correlations with other water quality parameters, including the pH, DO, TOC, TN, and TP at 90 m, were insignificant.
4. Discussion
The results of a previous study of Bosea sp. BIWAKO-01 cultures demonstrated that the addition of a low concentration of gelatinous polysaccharides, such as agar and agarose, stimulates the bacterial oxidation of Mn(II) [13]. In the presence of polysaccharides, the BIWAKO-01 cell extends a few filaments from the cell surface and becomes encapsulated with the increase in the number of filaments. Oxidized Mn phases include γ-MnOOH, a needle-type Mn(III) oxide [13], which likely forms via the reduction of thin sheet-like δ-Mn(IV)O2 by excess dissolved Mn2+ ions. Thus, the dissolved Mn2+ remaining in the culture fluid controls the mineralogy of the final product [37]. In this study, a low concentration of dissolved Mn2+ (0.1 mM) was added intermittently, which hindered the formation of Mn(III) oxide, maintaining the sheet-like structures of δ-MnO2 (Figure S1 and Figure 2). Filamentous Mn particles from Lake Biwa had similar structures resembling to that of δ-MnO2 (Figure S2), as widely reported for biogenic or naturally occurring Mn oxide phases [1,2].
The addition of polysaccharides to BIWAKO-01 cultures might be relevant to the static conditions affecting the cellular motility in the slightly viscous medium or polymeric matrix [13], although its role remains unknown. In this study, a similar effect was observed for EPS-bearing cells of green algae, S. dorsidentiferum and S. arctiscon (Figure 3). Diverse microalgae, including green algae, diatoms, and cyanobacteria, nonenzymatically and enzymatically oxidize Mn(II) [38,39,40,41]. The results of this study show that the Mn(II) oxidation depends on the presence of BIWAKO-01. Green algae could not oxidize Mn(II) in cultures without BIWAKO-01. The Mn oxide particles were in localized contact with the EPS matrix (Figure 3), suggesting that BIWAKO-01 cells grow on the EPS and produce the Mn oxide particles. This phenomenon is consistent with observations made on natural Mn particles in the near-bottom layer of Lake Biwa, which are in contact with organic aggregates containing gelatinous, lectin-stainable substances and dead algal cells (Figure 4 and Figure S3). The reactivity of the filaments during staining with the lectins PHE-A, SBA, and PNA was relatively high, suggesting that the filaments also contain lectin-stainable polysaccharides. This was the case for the filaments produced by BIWAKO-01; the filament structures consist of lectin-stainable acidic polysaccharides and Mn oxide phases [13]. Our observations are similar to those of previous studies. In the hypolimnion of a freshwater reservoir, filamentous Mn particles were present in a mixture of algae, diatoms, and amorphous material [42]. The results of several studies demonstrated the presence of Mn precipitates associated with diatom frustules in freshwater biofilms [43,44]. In the Gotland Basin, Baltic Sea, 68% of all Mn-rich particles observed in the stratified column were associated with large organic aggregates with radii ranging from 10 to 54 μm [5]. The Mn particle-associated polysaccharidic substances are reported to serve as microbial hotspots and affect their stationary sinking velocity by altering the particle density [45]. The results of the present study suggest the significant contribution of such organic material to the biogenic formation of Mn oxide particles.
The presence of dead algal cells in the near-bottom layer (Figure 4) indicates that photosynthetic products, including algal biomass, sink to the lake bottom. At the study site, the distribution of total polysaccharides in the water column was widespread but varied spatiotemporally (Figure 5). The higher concentration gradient in the upper layer and correlations with the biovolume of phytoplankton cells or Chl.a concentration indicate that these substances originate from phytoplankton in the upper layer (Figure 6). Green algae are considered to be responsible for the total polysaccharide concentration at the study site. In an oligotrophic reservoir, the TEP produced in the surface layer contribute to the carbon export to the hypolimnion at 30 m, with values of 0.02% to 31% [46]. Although our observations provide indirect evidence for the origin of gelatinous polysaccharides in the near-bottom layer, the decomposition, utilization, and transformation of photosynthetic EPS by heterotrophic microorganisms during sinking cannot be excluded, as previously reported [18,22,47]. The results of a previous study in the northern basin of Lake Biwa showed that HCl-hydrolyzable saccharides in the epilimnion (2.5 m) and hypolimnion (40 m) consist of relatively large quantities of rhamnose, fucose, arabinose, and xylose [48]. This is consistent with our observations (Figure S4). The high activity of heterotrophic consumers in the lake water observed in previous studies suggests that polysaccharides, which are photosynthetically produced in the epilimnion, are modified by the heterotrophs and the remaining refractory polysaccharides reach the hypolimnion [48,49].
Although the results obtained for laboratory microbial cultures suggest the involvement of algal EPS in the formation of filamentous Mn particles (Figure 3), the variation of filamentous Mn particles in the near-bottom layer cannot be explained by the total polysaccharide concentration. This might result from the different timing of the DO depletion and elevation of polysaccharides in the environment. Therefore, we assume that lectin-positive gelatinous substances are supplied by the sinking of phytoplankton cells throughout the year and remain, at least partly, on the bottom. We examined the correlations among water quality data based on average values obtained during an annual cycle of filamentous Mn particle formation. The analysis of 18-year water quality data reveals a positive correlation between the annual average density of filamentous Mn particles at 90 m and total algal (or green algal) biovolume at 0.5 m (Figure 7 and Figure 8). It is likely that a higher primary production leads to an increase in the DO consumption by heterotrophic consumers in the hypolimnion, accelerating the Mn particle formation. However, the abundance of Mn particles cannot be explained by the fluctuation of the DO or TOC concentration. This suggests that the (green) algal biomass directly contributes rather to the formation of Mn particles in the near-bottom layer.
Regarding the long-term trend of the total biovolume of phytoplankton cells in the northern basin of Lake Biwa (0.5 m), research showed that the average total biovolume ranged from 1.5 to 1.8 mm3 L-1 from 2000-2009, corresponding to 63% to 85% of the levels observed from 1980-1989 [33]. Although the total biovolume decreases, the total volume of EPS surrounding the phytoplankton cells (16 to 21 mm3 L-1 during 2000 and 2009) reaches more than twice the levels of 1980-1989 and 1990-1999, suggesting the increase in phytoplankton species with increasing EPS content. Such a long-term succession of phytoplankton species likely affects the supply of EPS to the near-bottom layer and thus the Mn cycle. In recent years, Lake Biwa has experienced incomplete overturns of the water column by lake warming [31]. It has been argued that the resultant DO depletion seriously affects the fate of Mn and other elements such as arsenic in the bottom environment [34,50]. It has been shown that the redox cycle of Mn in Lake Biwa affects the dynamics of metal ions [34,51,52]. Thus, the phytoplankton succession may interfere with the DO depletion-induced dynamics of Mn and other metal ions in the bottom environment because algal EPS might force back metal ions dissolved in water via the induction of microbial Mn(II) oxidation.
5. Conclusions
In this study, we provide several lines of evidence that algal EPS production is a key environmental factor directly inducing the formation of filamentous Mn oxide particles in the near-bottom layer of the northern basin of Lake Biwa. This implies that EPS drive the geochemical cycles of Mn and other elements. Such an EPS-dependent mechanism may be ubiquitous in nature because Mn particles associated with algal EPS or EPS-like matrix have been reported elsewhere. Further studies are needed to clarify the molecular mechanism of filamentous Mn oxide particle formation by Mn(II)-oxidizing bacteria in the presence of polysaccharidic substances.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Mn(II) oxidation by Bosea sp. BIWAKO-01 in agar-containing cultures; Figure S2: TEM image of filamentous Mn oxide particle collected at a depth of 90 m at the study site of Lake Biwa; Figure S3: differential interference contrast and epifluorescence images of aggregates collected at a depth of 90 m at the study site of Lake Biwa; Figure S4: neutral monosaccharides detected in the total polysaccharides collected from the lake waters at 0.5 m and 90 m during 2014-2017; Table S1: composition of CT medium.
Author Contributions
Conceptualization, S.F., H.I., M.F., K.O., S.I. and N.M.; methodology, S.F., H.I., M.F., H.H., S.S., S.I. and N.M.; validation, M.F. and N.M.; investigation, S.F., H.I., H.H., S.S., S.I. and N.M.; resources, S.F. and S.I.; data curation, M.F.; writing—original draft preparation, N.M.; writing—review and editing, S.F., H.I., M.F., H.H., S.S., K.O. and S.I.; visualization, F.M. and N.M.; supervision, S.I. and N.M.; project administration, N.M.; funding acquisition, N.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Japan Society for the Promotion of Science (JSPS) KAKENHI Grant Number JP26281042 and JP17H01898.
Data Availability Statement
All datasets generated for this study are included in the article.
Acknowledgments
We thank Professor Jun Takada at Okayama University for useful discussions and advice regarding the TEM work and Ayami Matsuzaki, Takuya Matsumoto, and Ayako Oishi at Akita Prefectural University for assistance with the culture work and polysaccharide analysis. The GC-MS measurements were conducted by using the DASH at the Research Institute for Sustainable Humanosphere, Kyoto University. The article is dedicated to the memory of Toru Suzuki, who was a Professor of Gifu University and contributed greatly to the progress of this research project.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tebo, B.M.; Bargar, J.R.; Clement, B.G.; Dick, G.J.; Murray, K. J.; Parker, D.; Verity, R.; Webb, S.M. Biogenic manganese oxides: properties and mechanisms of formation. Annu Rev Earth Planet Sci 2004, 32, 287–328. [Google Scholar] [CrossRef]
- Huang, Y.; Huangfu, X.; Ma, C.; Liu, Z. Sequestration and oxidation of heavy metals mediated by Mn(II) oxidizing microorganisms in the aquatic environment. Chemosphere 2023, 329, 138594. [Google Scholar] [CrossRef]
- Lienemann, C.P.; Taillefert, M.; Perret, D.; Gaillard, J.F. Association of cobalt and manganese in aquatic systems: chemical and microscopic evidence. Geochim Cosmochim Acta 1997, 61, 1437–1446. [Google Scholar] [CrossRef]
- Taillefert, M.; Macgregor, B.J.; Gaillard, J.F.; Lienemann, C.P.; Stahl, D.A. Evidence for a dynamic cycle between Mn and Co in the water column of a stratified lake. Environ Sci Technol 2002, 36, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Neretin, L.N.; Pohl, C.; Jost, G.; Leipe, T.; Pollehne, F. Manganese cycling in the Gotland Deep, Baltic Sea. Mar Chem 2003, 82, 125–143. [Google Scholar] [CrossRef]
- Dellwig, O.; Leipe, T.; März, C.; Glockzin, M.; Pollehne, F.; Schnetger, B.; Yakushev, E.V.; Böttcher, M.E.; Brumsack, H.J. A new particulate Mn-Fe-P-shuttle at the redoxcline of anoxic basins. Geochim Cosmochim Acta 2010, 74, 7100–7115. [Google Scholar] [CrossRef]
- Bauer, S.; Blomqvist, S.; Ingri, J. Distribution of dissolved and suspended particulate molybdenum, vanadium, and tungsten in the Baltic Sea. Mar Chem 2017, 196, 135–147. [Google Scholar] [CrossRef]
- Ossa, O. F.; Hofmann, A.; Wille, M.; Spangenberg, J.E.; Bekker, A.; Poulton, S.W.; Eickmann, B.; Schoenberg, R. Aerobic iron and manganese cycling in a redox-stratified Mesoarchean epicontinental sea. Earth Planet Sci Lett 2018, 500, 28–40. [Google Scholar] [CrossRef]
- Henkel, J.V.; Schulz-Vogt, H.N.; Dellwig, O.; Pollehne, F.; Schott, T.; Meeske, C.; Beier, S.; Jürgens, K. Biological manganese-dependent sulfide oxidation impacts elemental gradients in redox-stratified systems: indications from the Black Sea water column. ISME J 2022, 16, 1523–1533. [Google Scholar] [CrossRef]
- Gregory, E.; Perry, R.S.; Staley, J.T. Characterization, distribution, and significance of Metallogenium in Lake Washington. Microb Ecol 1980, 6, 125–140. [Google Scholar] [CrossRef]
- Maki, J.S.; Tebo, B.M.; Palmer, F.E.; Nealson, K.H.; Staley, J.T. The abundance and biological activity of manganese-oxidizing bacteria and Metallogenium-like morphotypes in Lake Washington, USA. FEMS Microbiol Ecol 1987, 45, 21–29. [Google Scholar] [CrossRef]
- Sternbeck, J. Manganese cycling in a eutrophic lake -rates and pathways. Aquat Geochem 1996, 1, 399–426. [Google Scholar] [CrossRef]
- Furuta, S.; Ikegaya, H.; Hashimoto, H.; Ichise, S.; Kohno, T.; Miyata, N.; Takada, J. Formation of filamentous Mn oxide particles by the alphaproteobacterium Bosea sp. strain BIWAKO-01. Geomicrobiol J 2015, 32, 666–676. [Google Scholar] [CrossRef]
- Zakharova, Y.R.; Parfenova, V.V.; Granina, L.Z.; Kravchenko, O.S.; Zemskaya, T.I. Distribution of iron- and manganese-oxidizing bacteria in the bottom sediments of Lake Baikal. Inland Water Biol, 2010; 3, 313–321. [Google Scholar] [CrossRef]
- Ehrlich, H.L.; Newman, D.K. Geomicrobiology of manganese. In Geomicrobiology, 5th ed.; CRC Press: Boca Raton, USA, 2009; pp. 347–420. [Google Scholar]
- Miyajima, T. Production of Metallogenium-like particles by heterotrophic manganese-oxidizing bacteria collected from a lake. Arch Microbiol 1992, 158, 100–106. [Google Scholar] [CrossRef]
- Leppard, G.G. The characterization of algal and microbial mucilages and their aggregates in aquatic ecosystems. Sci Total Environ 1995, 165, 103–131. [Google Scholar] [CrossRef]
- Passow, U. Transparent exopolymer particles (TEP) in aquatic environments. Prog Oceanogr 2002, 55, 287–333. [Google Scholar] [CrossRef]
- Mari, X.; Passow, U.; Migon, C.; Burd, A.B.; Legendre, L. Transparent exopolymer particles: effects on carbon cycling in the ocean. Prog Oceanogr 2017, 151, 13–37. [Google Scholar] [CrossRef]
- Berman, T.; Viner-Mozzini, Y. Abundance and characteristics of polysaccharide and proteinaceous particles in Lake Kinneret. Aquat Microb Ecol 2001, 24, 255–264. [Google Scholar] [CrossRef]
- Azam, F.; Malfatti, F. Microbial structuring of marine ecosystems. Nat Rev Microbiol 2007, 5, 782–791. [Google Scholar] [CrossRef]
- Callieri, C.; Corno, G.; Contesini, M.; Fontaneto, D.; Bertoni, R. Transparent expolymer particles (TEP) are driven by chlorophyll a and mainly confined to the euphotic zone in a deep subalpine lake. Inland Waters 2017, 7, 118–127. [Google Scholar] [CrossRef]
- Cai, Y.-M. Non-surface attached bacterial aggregates: a ubiquitous third lifestyle. Front Microbiol 2020, 11, 557035. [Google Scholar] [CrossRef]
- Miyajima, T. Biological manganese oxidation in a lake. I. Occurrence and distribution of Metallogenium sp., and its kinetic properties. Arch Hydrobiol 1992, 124, 317–325. [Google Scholar] [CrossRef]
- Furuta, S.; Yoshida, M.; Okamoto, T.; Wakabayashi, T.; Ichise, S.; Aoki, S.; Kono, T.; Miyajima, T. Morphological variations of a manganese-oxidizing microorganism Metallogenium observed in the developmental process of cultures collected from Lake Biwa waters. Jpn J Limnol 2007, 68, 433–441. [Google Scholar] [CrossRef]
- Ishikawa, K.; Furuta, S.; Nakajima, T. Microbes as indicator species in low oxygen environments. In Lake Biwa. Interactions between nature and people; Kawanabe, H., Nishino, M., Maehata, M., Eds.; Springer: Dordrecht, Netherlands, 2012; pp. 231–233. [Google Scholar] [CrossRef]
- Watanabe, M.; Ichimura, T. Fresh- and salt-water forms of Spirulina platensis in axenic cultures. Bull Jpn Soc Phycol 1977, 25, 371–377. [Google Scholar]
- Giroldo, D.; Vieira, A.A.H.; Paulshn, B.S. Microbial degradation of extracellular polysaccharides released by a tropical strain of Staurastrum orbiculare (Zygnematophyceae). Phycologia 2005, 44, 671–677. [Google Scholar] [CrossRef]
- Boogerd, F.C.; de Vrind, J.P.M. Manganese oxidation by Leptothrix discophora. J Bacteriol 1987, 169, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Ikegaya, H.; Nakase, T.; Iwata, K.; Tsuchida, H.; Sonobe, S.; Shimmen, T. Studies on conjugation of Spirogyra using monoclonal culture. J Plant Res 2012, 125, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Shiga Prefecture (2023) Annual Reports on the Environment. Shiga Prefecture, Japan. Available on https://www.pref.shiga.lg.jp/ippan/kankyoshizen/kankyou/11319.html.
- Kishimoto, N.; Ichise, S.; Suzuki, K.; Yamamoto, C. Analysis of long-term variation in phytoplankton biovolume in the northern basin of Lake Biwa. Limnology 2013, 14, 117–128. [Google Scholar] [CrossRef]
- Ichise, S.; Ikegaya, H.; Furuta, S.; Fujiwara, N.; Ikeda, S.; Kishimoto, N.; Nishimura, O. Analysis of long-term variation of phytoplankton biovolume and gelatinous sheath volume in Lake Biwa. Jpn J Water Treat Biol 2013, 49, 65–74. [Google Scholar] [CrossRef]
- Nakashima, Y.; Shimizu, A.; Maruo, M.; Sohrin, Y. Trace elements influenced by environmental changes in Lake Biwa: I Seasonal variations under suboxic hypolimnion conditions during 2007 and 2009. Limnology 2016, 17, 151–162. [Google Scholar] [CrossRef]
- Scott, T.A.; Melvin, E.H. Determination of dextran with anthrone. Anal Chem 1953, 25, 1656–1661. [Google Scholar] [CrossRef]
- Hayashi, T. Measuring β-glucan deposition in plant cell walls. In Plant fibers. Modern methods of plant analysis; Linskens, H.F., Jackson, J.F., Eds.; Springer: Berlin, Germany, 1989; pp. 138–160. [Google Scholar]
- Lefkowitz, J.P. , Rouff, A.A.; Elzinga, E.J. Influence of pH on the reductive transformation of birnessite by aqueous Mn(II). Environ Sci Technol 2013, 47, 10364–10371. [Google Scholar] [CrossRef]
- Richardson, L.L.; Aguilar, C.; Nealson, K.H. Manganese oxidation in pH and O2 microenvironments produced by phytoplankton. Limnol Oceanogr 1988, 33, 352–363. [Google Scholar] [CrossRef]
- Knauer, K.; Jabusch, T.; Sigg, L. Manganese uptake and Mn(II) oxidation by the alga Scenedesmus subspicatus. Aquat Sci 1999, 61, 44–58. [Google Scholar] [CrossRef]
- Wang, R.; Wang, S.; Tai, Y.; Tao, R.; Dai, Y.; Guo, J.; Yang, Y.; Duan, S. Biogenic manganese oxides generated by green algae Desmodesmus sp. WR1 to improve bisphenol A removal. J Hazard Mater 2017, 339, 310–319. [Google Scholar] [CrossRef]
- Chaput, D.L.; Fowler, A.J.; Seo, O.; Duhn, K.; Hansel, C.M.; Santelli, C.M. Mn oxide formation by phototrophs: spatial and temporal patterns, with evidence of an enzymatic superoxide-mediated pathway. Sci Rep 2019, 9, 18244. [Google Scholar] [CrossRef] [PubMed]
- Stein, L.Y.; Jones, G.; Alexander, B.; Elmund, K.; Wright-Jones, C.; Nealson, K.H. Intriguing microbial diversity associated with metal-rich particles from a freshwater reservoir. FEMS Microbiol Ecol 2002, 42, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Robbins, E.I.; Corley, T.L. Microdynamics and seasonal changes in manganese oxide epiprecipitation in Pinal Creek, Arizona. Hydrobiologia 2005, 534, 165–180. [Google Scholar] [CrossRef]
- Keim, C.N.; Nalini Jr., H. A.; de Lena, J.C. Manganese oxide biominerals from freshwater environments in Quadrilatero Ferrifero, Minas Gerais, Brazil. Geomicrobiol J 2015, 32, 549–559. [Google Scholar] [CrossRef]
- Glockzin, M.; Pollehne, F.; Dellwig, O. Stationary sinking velocity of authigenic manganese oxides at pelagic redoxclines. Mar Chem 2014, 160, 67–74. [Google Scholar] [CrossRef]
- De Vicente, I.; Ortega-Retuerta, E.; Romera, O.; Morales-Baquero, R.; Reche, I. Contribution of transparent exopolymer particles to carbon sinking flux in an oligotrophic reservoir. Biogeochemistry 2009, 96, 13–23. [Google Scholar] [CrossRef]
- Ortega-Retuerta, E.; Duarte, C.M.; Reche, I. Significance of bacterial activity for the distribution and dynamics of transparent exopolymer particles in the Mediterranean Sea. Microb Ecol 2010, 59, 808–818. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, K. Seasonal variations and dynamics of dissolved carbohydrates in Lake Biwa. Org Geochem 2004, 35, 169–179. [Google Scholar] [CrossRef]
- Urabe, J.; Nakanishi, M.; Kawabata, K. Contribution of metazoan plankton to the cycling of nitrogen and phosphorus in Lake Biwa. Limnol Oceanogr 1995, 40, 232–241. [Google Scholar] [CrossRef]
- Sohrin, Y.; Nakashima, Y.; Maruo, M. Trace elements influenced by environmental changes in Lake Biwa: II. Chemical variations in the hypolimnion over the last half-century. Limnology 2016, 17, 163–173. [Google Scholar] [CrossRef]
- Takamatsu, T.; Kawashima, M.; Koyama, M. The role of Mn2+-rich hydrous manganese oxide in the accumulation of arsenic in lake sediments. Water Res 1985, 19, 1029–1032. [Google Scholar] [CrossRef]
- Sugiyama, M.; Hori, T.; Kihara, S.; Matsui, M. Geochemical behavior of trace elements in Lake Biwa. Limnology 2005, 6, 117–130. [Google Scholar] [CrossRef]
Figure 1.
Long-term variations of filamentous Mn particle densities and DO concentrations at a depth of 90 m at the study site. Vertical lines represent complete overturns of the water column.
Figure 1.
Long-term variations of filamentous Mn particle densities and DO concentrations at a depth of 90 m at the study site. Vertical lines represent complete overturns of the water column.
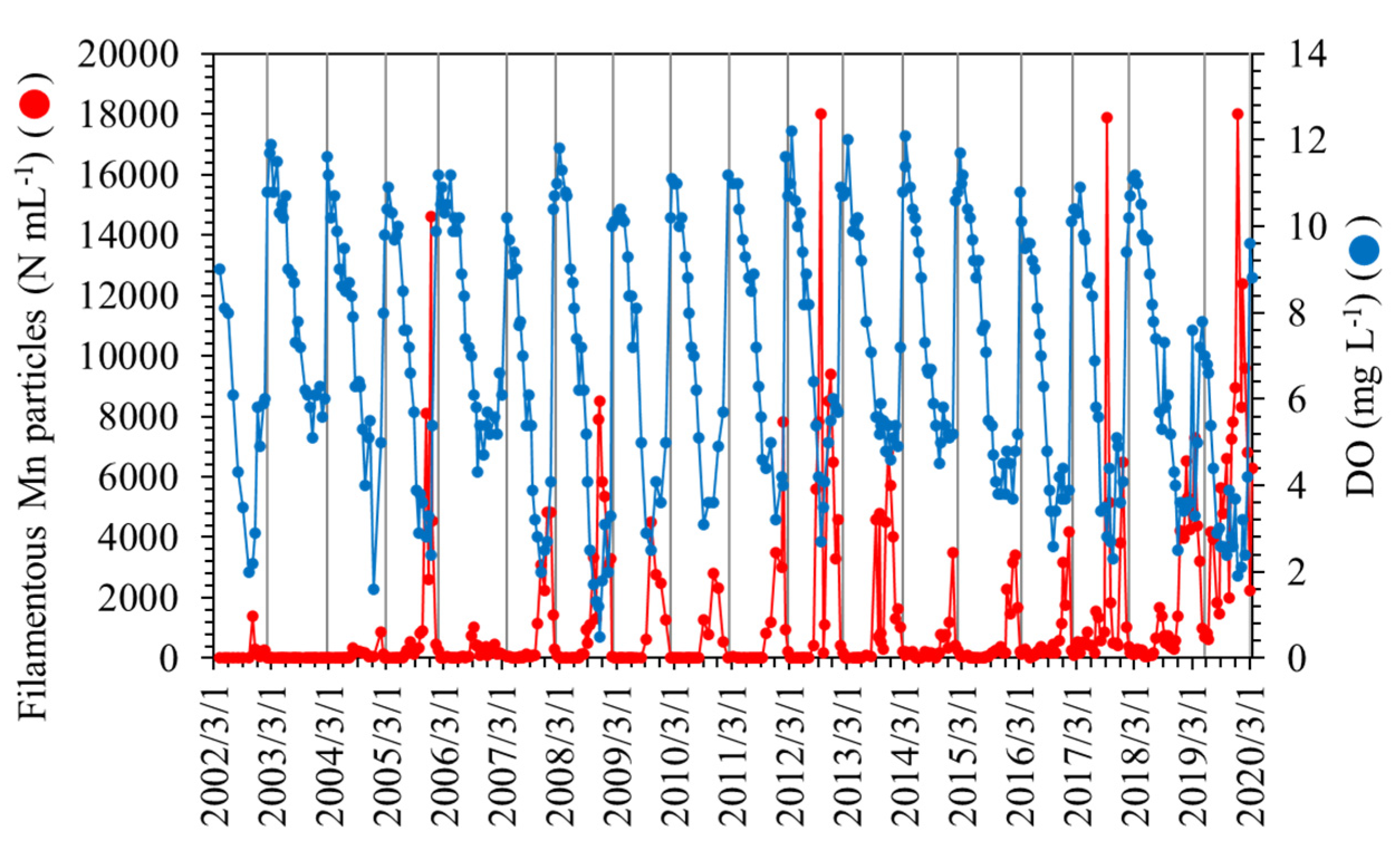
Figure 2.
TEM image of filamentous Mn oxide particle collected from BIWAKO-01 culture at day 21. The high magnification image shows that the filaments consist of sheet-type structure. Bar: 100 nm.
Figure 2.
TEM image of filamentous Mn oxide particle collected from BIWAKO-01 culture at day 21. The high magnification image shows that the filaments consist of sheet-type structure. Bar: 100 nm.
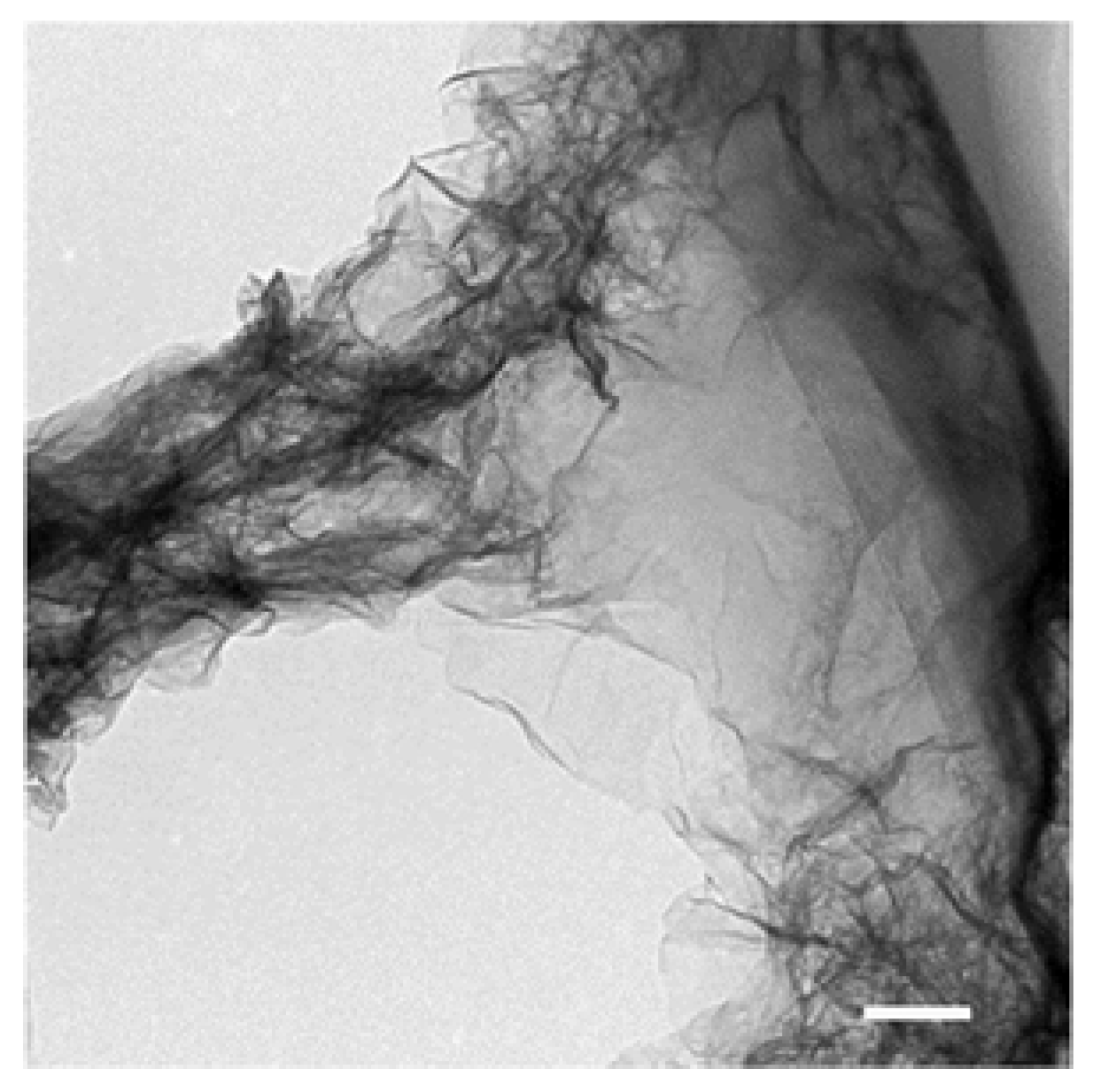
Figure 3.
Microscopic images of filamentous Mn oxide particles that formed in laboratory cultures of Bosea sp. BIWAKO-01. The bacterial strain was grown in the presence of green algae: S. dorsidentiferum intact (a) and EPS-removed (b) cells; S. arctiscon intact (c) and EPS-removed (d) cells. Images were obtained on India ink-stained specimens. The arrowheads indicate filamentous Mn particles. Bar: 30 μm.
Figure 3.
Microscopic images of filamentous Mn oxide particles that formed in laboratory cultures of Bosea sp. BIWAKO-01. The bacterial strain was grown in the presence of green algae: S. dorsidentiferum intact (a) and EPS-removed (b) cells; S. arctiscon intact (c) and EPS-removed (d) cells. Images were obtained on India ink-stained specimens. The arrowheads indicate filamentous Mn particles. Bar: 30 μm.

Figure 4.
Microscopic images of filamentous Mn oxide particles collected at a depth of 90 m at the study site. (a) India ink-stained aggregates including filamentous Mn particles (arrowhead 1), dead algal cells (arrowhead 2), and gelatinous substances (arrowhead 3). Bar: 20 μm. (b) Unstained aggregates including numerous Mn particles and dead algal cells (arrowheads 4 and 5). Bar: 100 μm. (c, d) Differential interference contrast and epifluorescence images of aggregates stained with fluorescein-conjugated LEL. Gelatinous substances are indicated with arrowheads. Bar: 10 μm.
Figure 4.
Microscopic images of filamentous Mn oxide particles collected at a depth of 90 m at the study site. (a) India ink-stained aggregates including filamentous Mn particles (arrowhead 1), dead algal cells (arrowhead 2), and gelatinous substances (arrowhead 3). Bar: 20 μm. (b) Unstained aggregates including numerous Mn particles and dead algal cells (arrowheads 4 and 5). Bar: 100 μm. (c, d) Differential interference contrast and epifluorescence images of aggregates stained with fluorescein-conjugated LEL. Gelatinous substances are indicated with arrowheads. Bar: 10 μm.

Figure 5.
Seasonal and vertical variations of Chl.a (a), total polysaccharide (b) concentrations, and filamentous Mn particle densities (c) at the study site.
Figure 5.
Seasonal and vertical variations of Chl.a (a), total polysaccharide (b) concentrations, and filamentous Mn particle densities (c) at the study site.
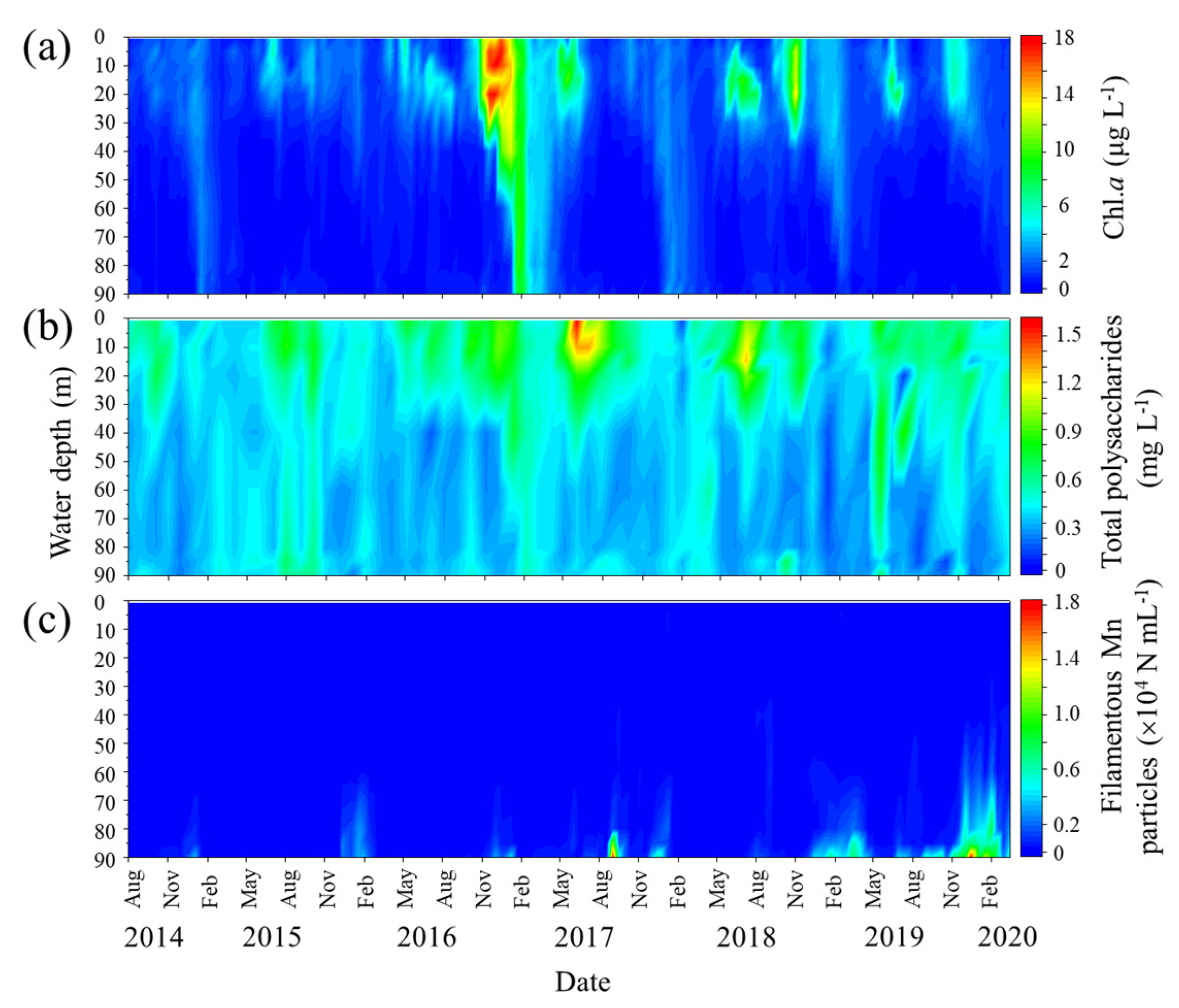
Figure 6.
Correlations between the total polysaccharide and Chl.a concentrations (a), total phytoplankton biovolume (b), Chlorophyceae biovolume (c), Cyanobacteria biovolume (d), Bacillariophyceae biovolume (e), and Chrysophyceae biovolume (f). All the data were obtained at a depth of 0.5 m at the study site of Lake Biwa.
Figure 6.
Correlations between the total polysaccharide and Chl.a concentrations (a), total phytoplankton biovolume (b), Chlorophyceae biovolume (c), Cyanobacteria biovolume (d), Bacillariophyceae biovolume (e), and Chrysophyceae biovolume (f). All the data were obtained at a depth of 0.5 m at the study site of Lake Biwa.
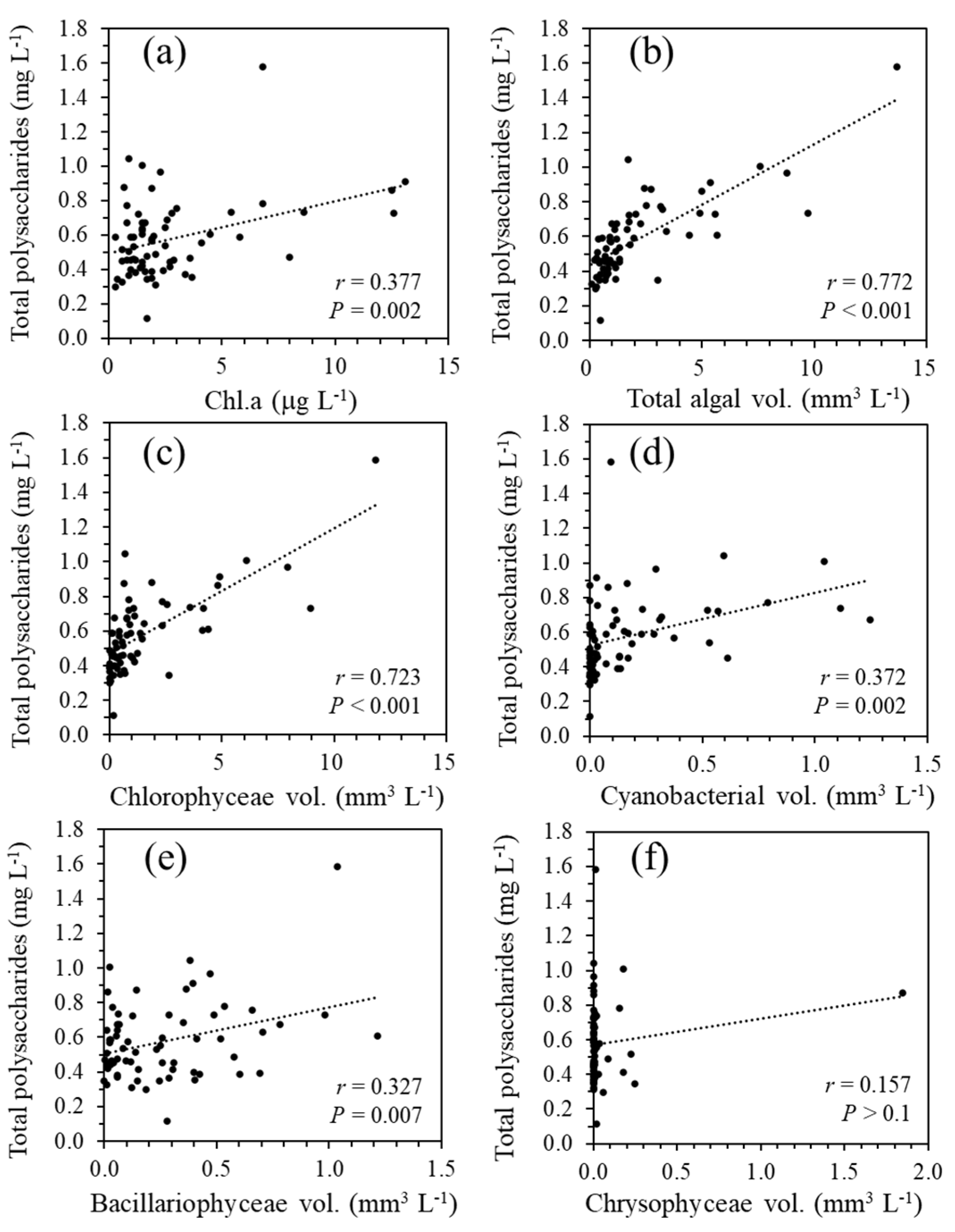
Figure 7.
PCA biplot for the annual average values of filamentous Mn particle densities and water quality data (a) and biovolume of algal species (b) collected over 18 years at the study site.
Figure 7.
PCA biplot for the annual average values of filamentous Mn particle densities and water quality data (a) and biovolume of algal species (b) collected over 18 years at the study site.
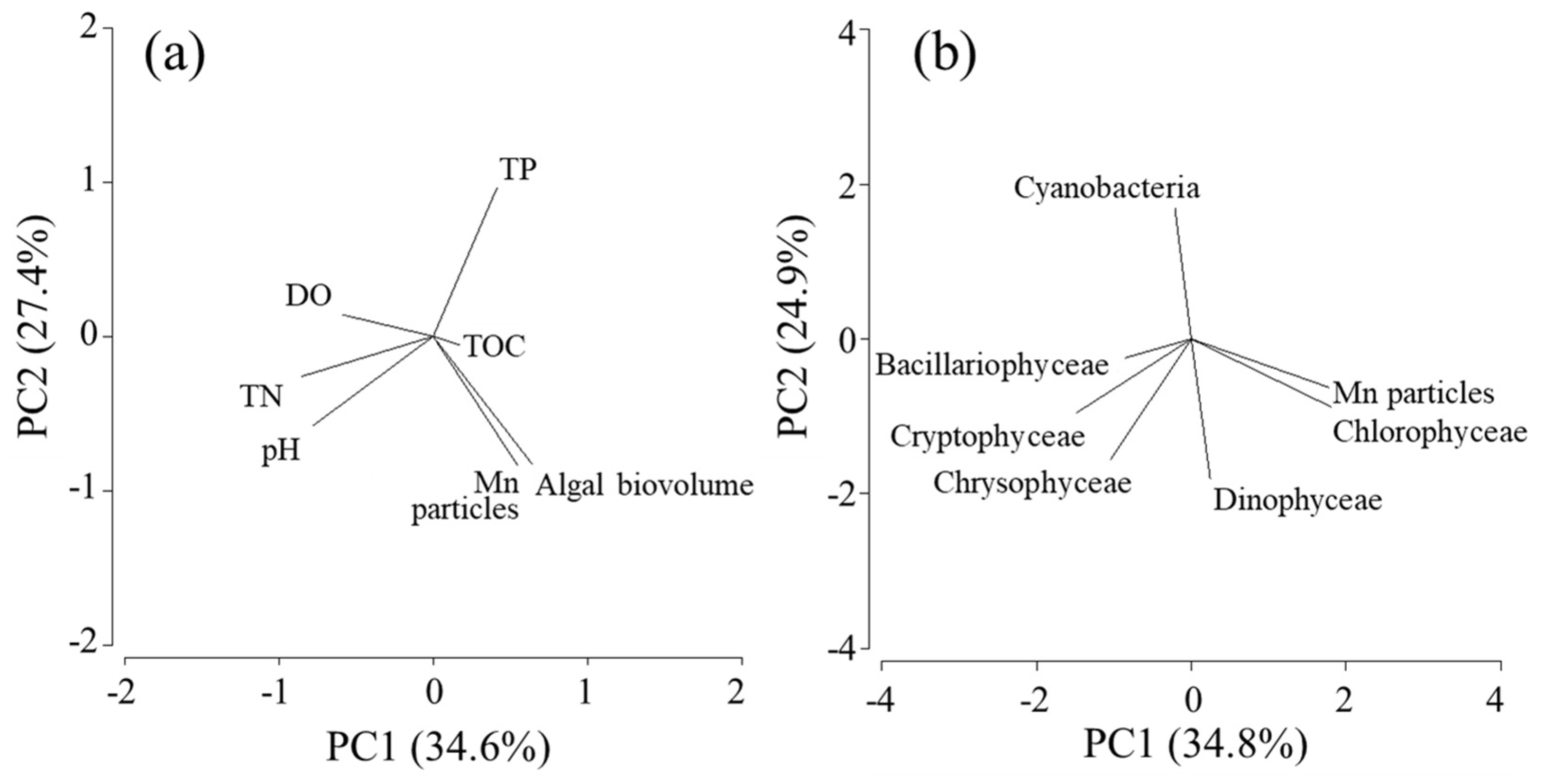
Figure 8.
Correlations between the filamentous Mn particle density at 90 m and total phytoplankton biovolume (a) and Chlorophyceae biovolume (b) at 0.5 m. The plots represent the annual average data collected for 18 years.
Figure 8.
Correlations between the filamentous Mn particle density at 90 m and total phytoplankton biovolume (a) and Chlorophyceae biovolume (b) at 0.5 m. The plots represent the annual average data collected for 18 years.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



