
Submitted:
21 June 2023
Posted:
21 June 2023
You are already at the latest version
Abstract
Due to the influence of bio-geographical and environmental factors, as well as anthropogenic landscape features, organism shows different reproductive strategies among different populations. There is lack of detailed information on the reproductive biology of Kentish plover Charadrius al-exandrinus in arid lands in Central Asia. In this study, we summarized the characteristics of the reproductive biology of three geographically distinct plover populations in Aibi Lake in the Northwestern Xinjiang, Taitema Lake in the Southern Xinjiang and artificial reservoirs around Urumqi City in Northern Xinjiang respectively, based on 440 eggs of 158 nests observed and ana-lyzed from April to July of 2019 and 2020. We find that there is no significant difference in clutch size among the three populations. However, the egg size of Taitema Lake population is signifi-cantly larger than those of the other two populations, whilst egg volume and clutch volume of ar-tificial reservoirs population are significantly larger than that of Aibi Lake. With the postpone-ment of laying date, the northern and northwestern populations respectively show the character-istics of small clutch size and relatively small egg size, and decrease of reproductive outputs. The heavier female plovers in Taitema Lake lays eggs earlier, and there was significantly positive cor-relation between the female body mass and the clutch size and egg size. The tarsometatarsus length of the female plovers was significantly positively correlated with the reproductive output in all three populations. The model selection results show that female body size and ambient tem-perature restrict the egg size and reproductive output of Kentish plovers, which is consistent with the upper limit hypothesis of the maternal condition and the maternal constraint. Our data sup-port that Kentish plover show distinct flexibility in breeding strategies to cope with the harsh nat-ural environment in arid lands of Xinjiang, China. The results of relatively high average clutch size and average egg size imply that saline wetlands in Western China are important breeding habitats for Kentish plover.
Keywords:
Kentish plover
; life-history
; reproductive output
; breeding behavior
; inland arid area
1. Introduction
The reproductive life history represents the overall trade-off between investment in egg and clutch and timing of reproduction of important events in the reproductive process of biological organisms, with a particular focus on the mechanism of why different species in the same environment or the same species in different environments may develop different reproduction strategies [1,2,3,4,5,6,7,8,9,10,11]. Reproduction, the most concerned content of ornithologists, is the fundamental process for life history, and reproductive strategy is the decisive factor that affects the fitness of animals, directly through individual survival and reproduction, population dynamics and continuity [3,10,12,13,14,15,16]. Large numbers of studies have described the diversity of reproductive life history strategies of birds due to variation of environmental conditions and individual morphology [16,17,18,19,20,21,22,23,24,25,26,27,28,29]. Additionally, the body size and reproductive characteristics in populations of the same species is also influenced by environmental and phylogenetic factors [30,31,32]. Reproductive life history characteristics of birds, such as egg-laying time, egg size, clutch size and clutch volume, are usually considered to be the main indicators to measure reproductive output [10,27,28,29,30,31,32,33]. These indicators can effectively reveal the different reproductive traits of birds in different environments due to adaptive trade-off and constraint of physiological mechanisms, which can be interpreted as different reproductive life history strategies [27,28,29,30,31,32,33,34]. For example, egg size and clutch size of birds can be adjusted according to the changes of egg-laying date, ambient temperature, food resources and female physical condition [4,28,35,36].
Life history theory seeks to explain how natural selection and other evolutionary forces shape organisms to optimize their survival and reproduction in the face of ecological challenges posed by the environment [1,2]. The trade-off between egg size and clutch size is one of the core principles of life history evolution, in which numerous studies have reported a rather positive covariance between egg size and clutch size: good-quality females may lay more and larger eggs compared to low-quality females [1,36,37,38]; adequate food resources usually lead to early laying of eggs and larger clutch sizes, instead of larger eggs [6,39,40]. Theoretically, the reproductive output of parent birds that lay eggs at a fixed number can be increased by increasing the size of a single egg, but the physiological constraint hypothesis predicts that the reproductive output will be constrained by the maternal condition, which is specifically manifested in that larger individuals lay larger eggs, while smaller individuals lay smaller eggs due to insufficient energy storage in their bodies [41]. The body size can partly represent the female's energy storage, so as the body mass increases, the female can supply more energy (egg mass) or lay more eggs [42]. The size of egg has costs and benefits, and individuals might balance these costs and benefits when allocating resources to the size of an egg [43]. According to life history theory, females should have some flexibility in the allocation of resources on eggs, including laying the optimal egg size and small clutch size in the case of abundant resources, and laying relatively small egg size but large clutch size in the case of limited resources [2,29,44,45]. However, the constant number of offspring has been found in some groups. Birds provide care to their offspring during laying, incubating and brooding. As the incubation capacity hypothesis predicts that the maximum clutch size that females can lay is constrained by the incubating capacity (e.g. incubating spots size), this may explain why most species of shorebirds have a relatively invariant clutch size [46,47]. Thus, most plovers vary their reproductive output by varying their egg size rather than clutch size [6].
On the other hand, the morphological constraint hypothesis holds that the egg length or width was constrained by the size of female partial morphological features. When the egg width is constrained, females can increase their energy investment by laying longer egg, for example, lizards can lay longer eggs to increase their energy supply to eggs as the tail base width of female lizards constrains the egg width [48]. In the light of this phenomenon, some scholars have put forward the concept of egg shape allometry which states that with the increase of egg size, the growth rate of egg length and egg width is different, and the pattern of allometry in egg shape can be checked by comparing the regression slope between egg length and egg width [49] Therefore, the size of a single egg can be increased indefinitely. The upper limit hypothesis of maternal constraint predicts that the egg width is physically constrained by the cloaca of bird, whilst the increase of egg size is mainly achieved by increasing the egg length, which is manifested as the egg length allometry [45,49].
The Kentish plover Charadrius alexandrinus is a small shorebird (with a body length of 14~17.5cm) with an extremely large range and multiple geographical races which live in both temperate and subtropical climate zones in four continents. Kentish plovers often inhabit the open and flat coastal beaches, the bare land around saltwater lakes and inland lakes [50,51]. The breeding period of Kentish plover is usually from March to August every year with certain variations in different regions. They usually lay 3 oval eggs per nest, accidentally 2 to 4 eggs, with length of 29~35 mm and width of 22~25 cm, mass of 8~11 g [9]. The female bird lays an egg with 2 day interval until the incubation starts after the last egg is laid. The incubation period is approximately 23 to 29 days [9,50]. In China, Kentish plovers breed in coastal areas, inland lakes and near reservoirs in the north, and inhabit all Provinces during migration, and can be seen throughout most of the southeastern coast during non-breeding periods [54,55]. Because of their wide distribution, the breeding strategies of Kentish plovers are diverse and have strong plasticity, which attracts the attention of scholars in animal ecology, behavior and evolution [46]. Variations in breeding strategies are common phenomena in birds living in different distribution ranges [10,46,56,57]. For example, reproductive characteristics such as egg length and width, clutch size, laying date, incubation rate, growth rate, brood desertion and adult survival often differ distinctively between northern and southern bird species [58,59,60]. Several studies have investigated how reproductive traits vary within a single species as elevation increases and climate becomes more severe [21,61,62,63]. We hypothesize that, as far as the avian life history variation along altitudinal gradients is concerned, breeding in high-elevation habitat results in shift to shorter life-history strategy within a single species.
At present, the reproductive biology of Kentish plover has been studied mainly in coastal areas, and most of the data come from Europe and North Africa [50,64,65,66]. In China, the study of Kentish plover reproductive biology mainly focuses on the populations in Bohai Bay [67], Qinghai Lake [68]. The populations in inland arid areas of Western China have drawn less attention [69]. Due to the influence of geography, temperature and environment, organism shows different reproductive strategies among different populations [45,47,68,70]. In view of the harsh natural environment in inland arid areas of Xinjiang, we hypothesis that Kentish plover may show flexibility in breeding strategies to cope with the changeable environment. Here, we focus on the reproductive traits of Kentish plover for revealing the reproductive strategy of this bird adapting to different localities in arid desert areas of Xinjiang (Figure 1), where the precipitations vary greatly, but the temperature differences are not significant (Figure 2). During the early stage of breeding season the climate of Taitema Lake is extremely dry with frequent dust storms. Hence, we hypothesis that, the plovers of Taitema Lake adapt to the poor climate conditions by laying larger eggs and shortening incubation period. According to the physiological constraint hypothesis, we hypothesis that female traits such as body mass or tarsometatarsus length determine reproductive output which is manifested in different egg size and clutch size in different populations. According to the upper limit hypothesis, we hypothesis that larger egg show a more positive allometry in egg shape. Therefore, the primary objective of this study was to provide baseline information about the breeding strategies of the Kentish plover populations in inner arid regions of Xinjiang, one of the important breeding areas for this species in China.
2. Materials and Methods
2.1. Study area
The current study was conducted, during the breeding season of Kentish plover, from April to July of 2019 and 2020. We collected and compared a variety of reproductive traits among three geographically distinct Kentish plover populations of Xinjiang, including Aibi Lake (AL) population in the Northwestern Xinjiang, Taitema Lake (TL) population in the Southern Xinjiang and artificial reservoir group (AR) population around Urumqi City in Northern Xinjiang (Figure 1).
The Aibi Lake Basin is a closed basin located in the inland area of the Junggar basin in Xinjiang in northwest China (43°380-45°520 N, 79°530-85°020E, with average altitude of 200 m) [71]. It has a total area of 50621 km2 where plains make up 25762 km2. The Aibi Lake, one of the important breeding habitats and temporary rest stations for migratory birds in western China, is under the jurisdiction of the Xinjiang Aibi Lake Wetland National Nature Reserve. The Lake, characterized by its dry and hot weather, high temperature low precipitation and little huaman disturbance, is the largest salt lake in western China, with a water area of 520 km2. The average annual temperature is 8.3 °C, and the average annual precipitation is 90.9 mm. The average annual precipitation on the surface of the lake is about 95 mm, with an the annual evaporation can reach as high as 1315 mm. Strong winds with a maximum wind speed of 55 m/s are more likely occur from April to June Kentish plover population there usually nesting on saline-alkali land, sand land and gravel substrates in the region [72].
The reservoir group including Liuchengzi Reservoir and Wushihua Reservoir around Urumqi City is located at the north edge of Boghda Mountain. They are typical plain reservoirs mainly used for aquaculture and irrigation. The specific study sites are located on the southeast bank of Liuchengzi Reservoir (N44.255835°, E87.885218°; with average altitude of 480 m) and the east of Wushihua Reservoir (N44.196667°, E87.741022°; with altitude of about 475 m). Since the distance between the two sites is only 13 km, and there is no difference in female morphology and egg characteristics between the populations form these two sites, we combined those two populations as an artificial reservoir population. This area is close to villages and towns, mostly surrounded by reclaimed farmland or industrial parks with strong human disturbance, such as livestock and road construction which accidentally cause the failure of nests.
Taitema Lake (TL) with a water surface of 300 km2, is located in the southeast margin of Taklimakan Desert, about 50 km north of Ruoqiang County, Bayingolin Mongol Autonomous Prefecture. It is the terminal lake of three river systems: the Tarim River, the Cherchen River, and the rivers on the northern slope of the Altun Mountains [68,73]. The Taitema Lake region has an extreme continental climate, making it extremely arid. The average annual precipitation is 17.4–42 mm, the average annual evaporation is 2500–3000 mm, and the extreme maximum temperature is 43.6℃. Furthermore, above-ground vegetation is sparse. The study area is located on the southeastern shore of the lake (N39.412433°, E88.517855°; with average altitude of 800 m). The breeding habitat of plovers is basically harsh desert composed of desert grassland, sand land and sand dunes which is well-preserved with little human disturbance [73,74].
2.2. Data Source
In this study, the regional meteorological data (Figure 2) came from the National Meteorological Information Center (https:// www.nmic.gov.cn/ en/) of the China Meteorological Administration, with data on temperature and precipitation obtained from the weather station, which is closest to the study region. We calculated the average value every ten days, including daily average temperature (AT), daily maximum temperature (DTmax), daily minimum temperature (DTmin), daily temperature difference (DD) and sum of rainfall (RF) during the breeding season from March 27 to July 24, each year in 2019 and 2020.
2.3. Data collection and traits measurement
We find nests by watching birds with a telescope, binoculars and by searching on foot inside the nesting grounds during the breeding season [75]. We monitored nests every 3–4 days in each year during the breeding season, until the hatching of eggs or the failure of the breeding attempt. The morphological traits of female parent and its eggs size characteristics (length and width) were measured with a digital caliper (0.01mm) and the weighed with a digital scale (0.01 g) [75]. GPS data of the nests were recorded. We captured breeding female parent with a walk-in funnel trap placed over the nests that had been incubated for at least seven days [68]. Female body mass (FBM, g) is considered to be the female's weight after laying all eggs. Female tarsometatarsus length (FTL, mm) is the tarsometatarsus length on the right side of the female. Egg mass (OEM, single egg weight, g), clutch size (CS, the total number of eggs laid in a single breeding period), egg length (EL, linear length at the longest end of the egg, mm), egg width (EW, linear length at the maximum width of the egg, mm), egg shape (ES, ratio of egg width and egg length) were measured for analyzing allometry in egg shape.
We also calculated egg volume (EV, cm3): EV =Kv×L×W2, where Kv=0.5236-(0.5236×2×(L/W)/100), L=egg length, W=egg width, and clutch volume (CV, the sum of the volumes of all eggs in a single breeding period, cm3) [76]. We estimated laying date (LD) through observation and floating the eggs in luke-warm water [75]. We used the Julian day, which we calculated as the number of days between the 1st April (i.e. 1) of each year, as the egg-laying date. The time interval between the beginning and the end of hatching of the plover was taken as incubation period (IP). The monitoring of AL population is too difficult and the number of nests was too small, so the data of incubation period could not be obtained by continuous monitoring.
2.4. Statistical analysis
The collected data is sorted out with Excel data. Non-normally distributed data was log10 converted to meet the assumption, i.e. the Kolmogorov-Smirnov normality test and Levene’s variance homogeneity test. We used the One-way ANOVA to make multiple comparisons on the female body mass, tarsometatarsus length, egg size, clutch volume and other traits of the three populations with post hoc Tukey’s tests. For the data that do not meet the above assumptions, we use the nonparametric Kruskal-Wallis test to examine data differences between the three populations with post hoc Steel-Dwass tests (multiple comparisons). We used analysis of independent t-test to examine the annual differences in reproductive traits of each population, the results showed no significant annual differences on reproductive traits among them, so two years of reproductive traits data from each population were combined for subsequent analysis. Model II regression in “lmodel2” package was used to make regression analysis between egg size, clutch size, clutch volume and laying time one by one, as well between egg size, clutch size, clutch volume and the body mass and tarsometatarsus length of their female parent one by one, so as to verify the effects of laying time and female morphology on egg size and clutch size.
For the egg shape allometry, we first make the regression between egg length and egg width. If the slope is greater than 1, there is exists egg shape allometry. Then, residual on the egg length, egg width with clutch size were extracted respectively to eliminate the effects of the clutch size on the egg length and width [45], and regression was made between the residual egg length and the residual egg width to test whether there is allometry for the egg shape after removing the effects of the clutch size.
Model Selection Analysis was conducted to test the effect of environmental factors and female morphological traits on egg volume and clutch volume. Daily average temperature, daily temperature difference (DD), daily maximum temperature, daily rainfall, laying date, clutch size, the female body mass and female tarsometatarsus length were taken as independent variables to carry out the Multinomial Logistic Model analysis. Populations were included as a fixed index, and the parameter years was entered as a random effect in order to control the interpopulation phylogenetic relatedness.
Akaike information Criterion (AIC) was used to compare models and determine two principles of the best model screening: (1) the minimum AIC value is required; (2) the Dealt value of the model < 2. All data are listed in the form of mean ± standard error (Mean ± SE), and all operations are completed in R.v.3.6.3 software, using the packages ‘‘ggplot’’ and ‘‘gplots’’. Differences were considered significant when P< 0.05.
3. Results
3.1. Female reproductive traits among populations
A total of 440 eggs from 158 nests of the Kentish plovers in three geographically distinct populations were measured from April to July of 2019 and 2020. Among them, there are 54 eggs from 20 nests in AL, 175 from 61 nests in TL and 211 from 77 nests in AR.
The results show that there is no significant difference in the female body mass among the three populations, while the female tarsometatarsus length of AL population was significantly longer than that of the other two populations. There was also no significant difference in clutch size among populations, but significant differences in egg volume and clutch volume in TL population, which were larger than those of the other two populations. Additionally, the egg width, egg volume and clutch volume of AR population were significantly larger than those of AL population (Table 1).
3.2. Laying date and reproductive traits
According to the observation, the AR population is strongly affected by the interference of artificial water level control and regional grazing. The first laying peak appears at the early stage of breeding season, while the second laying peak immediately follows the expansion of suitable nesting area in early June, when the water level has retreats to its lowest point. In addition, the average incubation period (IP) of the plover population in AR is significantly longer than that of TL population (Table 1).
Among the three populations, there was a significant negative correlation between the female body mass and laying date of the plover in TL (Figure 3A), which showed that the heavier females laid eggs earlier. There was also a significant negative correlation between the laying date and clutch size, clutch volume of the AL population (Figure 3B,F). The laying date of AR population significantly restricts the egg volume (Figure 3C–E) and clutch volume (Figure 3F). With the postponement of laying date, the plover there produces relatively small eggs representing less reproductive output.
3.3. Reproductive output and female traits
The results show that heavier and larger females lay larger eggs. Among the three populations, the female body mass in TL was significantly positively correlated with clutch size (Figure 4A), egg width, egg volume (Figure 4B,C) and clutch volume (Figure 4D). There was an extremely significant positive correlation between the female tarsometatarsus length and egg length (Figure 5A), clutch volume (Figure 5B) in AR population.
3.4. Model selection
In the egg size and clutch volume model reveal that that body mass is again a significant predictor of the variation in egg size. The results of egg size model selection shows that the best model for egg volume of three populations of birds includes the female body mass, female tarsometatarsus length and average temperature. The AICc value of the model is -197.50 and the weight is 0.948 (Table 2). The best model of the clutch volume of three populations of birds includes female body mass, egg length and average temperature. The AICc value of the model is -197.00 and the weight is 0.611 (Table 2).
3.5. Allometry
The regression slope (slope = 1.625, 95% CI: 1.327~1.990, Figure 6A) between egg length and egg width was significantly larger than 1 (P < 0.0001), indicating that there was allometry of egg shape, and the growth rate of egg length was significantly faster than that of egg width; After extracting the regression residuals of egg length, egg width and clutch size respectively, the regression slope between egg length residuals and egg width residuals (slope = 2.277, 95% CI: 1:858~2.790, Figure 6B) is also significantly larger than 1 (P < 0.0001), indicating that after eliminating the effect of clutch size, the egg shape allometry still exists, and the growth rate of egg length is significantly faster than that of egg width.
4. Discussion
4.1. Differences of reproduction traits among populations
The average clutch size in the three populations (2.86±0.35~2.96±0.35) was similar to that in other sites [47,50,51,52,53]. The proportion of nests with three eggs in the three populations in the study area is higher than 85%, which is higher than about 70% reported in other places [9,50]. Compared with other sites, the average egg volume is also larger [50,75,77]. The clutch size and the egg volume are generally considered to be related to food supply [40]. Larger eggs have been proved to improve the hatching success rate and increase the initial energy reserve of nestling [39,43]. For example, the TL population with relatively harsh environment and high daily average temperature produces larger eggs relative to body size, which strongly support our hypothesis that the plovers of TL adapt to the poor climate conditions by laying larger eggs and shortening incubation period.
4.2. Relationship between reproductive traits and laying date
All three populations began to incubate in April, which was later than that in European and Northern African populations [50,65]. The shorter breeding season may be related to the late start of laying, which may be limited by climatic conditions (mainly low temperature in early spring) [78]. Secondly, the laying data is a life history characteristics, which depends on habitat conditions, including altitudinal gradients. Compared with other populations, TL population has a poor early climate conditions due to a higher altitude (850m), which makes shorter female tarsometatarsus length, later laying date and shorter incubation period, resulting in the shortening of the breeding season in TL [79], which strongly support our hypothesis that breeding in high-elevation habitat results in shift to shorter life-history strategy within a single species. Since the interference of livestock in the late breeding season is the main reason for the nesting failure of AR population (unpublished data), the laying begins earlier, which may be affected by temperature, resulting in a long incubation period. The results of our another manuscript in preparation indicated that there were differences in nest survival rates among populations, in which the rate of the TL population was the highest (0.702, n=61 nests), and that of AR population was the lowest (0.296, n=77 nests) due to predation, parental desertion and the higher possibility of being trampled by livestock. We also discovered that the nest survival rate of the AR population decreased with the postponement of breeding season, and the female there would lay relatively small eggs resulted in less reproductive output. These results testified that the AR population manifested relatively flexible reproductive strategies, such as trade off between egg size and clutch size, in response to the lower survival rate. Additionally, higher level of human interference such as artificial water level control, regional grazing, road construction as well as predation risks also affect the end of reproduction by causing birds to leave early which led to the higher nesting failure in AR population. The later start of laying and shorter laying time of TL population may be the adaptation strategy of the population to severe environmental condition, which is consistent with the reproductive limit hypothesis in high altitude [79,80].
There was a significant negative correlation between laying date and egg volume, clutch volume of birds in AR, which showed that egg size and clutch volume decreased with the passage of laying date. There may be four reasons for this phenomenon. Firstly, individuals which lay eggs early may be older females. They arrive early and start breeding early. Usually, young females tend to lay eggs lately, and lay smaller eggs [6]. Secondly, according to the embryonic temperature hypothesis [80], larger eggs can obviously reduce the mortality of offspring in cold environment [81]. At the early stage of reproduction, the temperature is low, and it is beneficial for hatching to produce larger eggs. Thirdly, it is also possible that the eggs produced in the later stage belong to the second clutch, which is due to the compensatory laying after the failure of hatching in the first clutch. Usually the eggs in the second clutch are smaller. Finally, in inland arid areas where water resources are relatively scarce, the change of water level directly affects the potential habitat of nesting. For example, after the water level retreats due to artificial control in the middle and late breeding period, a large area of ideal nesting areas created, resulting in explosive nesting and laying in AR.
4.3. Egg size and reproductive output are affected by both maternal and environment conditions
There was a significant positive correlation between the clutch size, egg size, clutch volume and bird postpartum weight in TL. In the face of local harsh climate, larger females will produce a relatively large number of eggs to improve the hatching success rate and the initial energy reserve of nestling [39,52,82]. It has been suggested that the egg size has a significant impact on the performance of nestling [43]. The tarsometatarsus length is an important identification indicator of bird age or quality. Our results show that there is a significant positive correlation between reproductive output and the female tarsometatarsus length in AR. In addition, the food supplementation usually leads to early laying and larger clutch size rather than larger eggs, which is common in birds [39,40].
As the parent birds that lay eggs at a fixed number, Kentish plover can increase the reproductive output by increasing the size of a single egg, but it is limited by maternal and environmental conditions. According to the model selection results (Table 2), the egg volume and clutch volume of the three plover populations are affected by the female conditions, including the female body mass and female tarsometatarsus length, which is consistent with our hypothesis that female traits such as body mass or tarsometatarsus length determine reproductive output which is manifested in different egg size and clutch size in different populations. The reproductive output of individuals with larger body size is also larger, which conforms to the prediction of physiological constraint hypothesis [41]. Additionally, the clutch volume of the plover is also affected by the egg length, which may be due to the limitation of its hard shell by the birth cloaca, and the size of a single egg can only be increased by increasing the egg length, which supports the upper limit hypothesis of maternal constraint [45,49].
4.4. The allometry of egg shape
The egg shape, the same as the egg size, is a highly variable characteristic in the life history of birds [2,34,51,83]. However, little was known about the adaptive significance of bird egg shape and how the differences arise and evolve [84]. We found that the growth rate of egg length of Kentish plover is significantly faster than that of egg width (Figure 6), which confirms the prediction of the upper limit hypothesis and is consistent with the results of the model selection results. Due to the intermittent laying mode of birds (such as laying every other day), only one egg is laid at a time. Theoretically, the size of a single egg will not be limited by the volume of abdominal cavity. At the same time, the egg width of birds will not be limited due to the open pelvis (some birds lay nearly round eggs). Therefore, in addition to the limitation of the birth cloaca, the explanation of the egg length allometry may be related to other factors, such as nest size, structure, shape and climate [85]. Further research is needed to determine the underlying mechanism.
5. Conclusions
The present study strongly supported our hypothesis that Kentish plover may show flexibility in breeding strategies to cope with the changeable environment. The three populations of plovers have precocial chicks and an invariant clutch size. The laying date of the northern Xinjiang populations was earlier than that of the southern Xinjiang populations. The environment in TL is the most barren and the climate is the harshest among the three, and the female tarsometatarsus length is the smallest, but the egg size and the clutch volume are significantly larger than those of AR and AL populations. Female body mass has a significant positive correlation with egg size, clutch size and clutch volume in all the three populations. The egg width, egg volume and clutch volume of the AR population were significantly larger than those of AL population, and the female tarsometatarsus length of the population has a significant positive correlation with the egg length, clutch volume. The single egg size and clutch volume of the three bird populations are affected by morphological constraint and maternal constraint. In total, our observation results of relatively high average egg size and clutch size implies that saline wetlands in Western China is an important breeding habitat for Kentish plover. Finally, future research will be recommended to further clarify the underlying mechanism of reproductive strategies of the species in response to regional environmental modification resulted by anthropogenic landscape features (human activities), altitudinal gradients and global climate changes.
Author Contributions
Conceptualization, Tamás Székely; Data curation, Peng Ding and Zitan Song; Formal analysis, Peng Ding; Investigation, Peng Ding and Zitan Song; Writing – original draft, Peng Ding, Lei Shi and Mardan Turghan; Writing – review & editing, Yang Liu and Lei Shi. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (No. 31660613), Wildlife and Plant Background Survey of Urumqi City (2022-2024) and Wildlife Investigation of Bortala Mongol Autonomous Prefecture (2021-2023) from Xinjiang Forestry and Grass Bureau.
Institutional Review Board Statement
The study was approved by the Animal Welfare and Ethics Committee of Xinjiang Agricultural University (protocol code: 2018015; date: 2018.7.13).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
Special appreciation goes to Dr. Tao Liang of Nanjing Forestry University for his guidance regarding statistical methods, and Dr. Naerhulan Halimubieke of University of Bath for providing professional advice regarding phrasing and for useful help at the beginning of the study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Roff, D.A. The evolution of life histories: Theory and analysis; Chapman & Hall: New York, USA, 1992. [Google Scholar]
- Stearns, S.C. The Evolution of Life Histories; Oxford University Press: Oxford, UK, 1992. [Google Scholar]
- Newton, I. Lifetime Reproduction in Birds; Academic Press: London, UK, 1989. [Google Scholar]
- Xu, F.; Yang, W.K.; Li, Y.M. Enlarged egg size increases offspring fitness of a frog species on the Zhoushan archipelago of China. Sci. Rep. 2019, 9, 11653. [Google Scholar] [CrossRef]
- Armstrong, T.; Robertson, R.J. Parental investment based on clutch value: Nest desertion in response to partial clutch loss in dabbling ducks. Anim. Behav. 1988, 36, 941–943. [Google Scholar] [CrossRef]
- Verhoeven, M.A.; Loonstra, A.H.; McBride, A.D.; Tinbergen, J.M.; Kentie, R.; Hooijmeijer, J.C.; Both, C.; Senner, N.R.; Piersma, T. Variation in egg size of Black-tailed Godwits. Ardea 2019, 107, 291–302. [Google Scholar] [CrossRef]
- Ghalambor, C.K.; Martin, T.E. Fecundity-survival trade-offs and parental risk-taking in birds. Science 2001, 292, 494–497. [Google Scholar] [CrossRef]
- Martin, T.E. Age-related mortality explains life history strategies of tropical and temperate songbirds. Science 2015, 349, 966–970. [Google Scholar] [CrossRef] [PubMed]
- Szekely, T.; Karsai, I. , Williams, T.D. Determination of clutch size in the Kentish Plover Charadrius alexandrinus. Ibis 1994, 136, 341–348. [Google Scholar] [CrossRef]
- Tian, L.; Liu, Y.; Zhou, Z.; Zhou, H.; Lu, S.; Zhang, Z. Reproductive success of a tropical barn swallow Hirundo rustica population is lower than that in temperate regions. Animals 2023, 13, 62. [Google Scholar] [CrossRef] [PubMed]
- Szekely, T.; Cuthill, I.C. Trade-off between mating opportunities and parental care: Brood desertion by female Kentish Plovers. P ROY SOC B-BIOL SCI 2000, 267, 2087–2092. [Google Scholar] [CrossRef]
- Zheng, G.M. Ornithology, 2nd ed.; Beijing Normal University Press: Beijing, China, 2012; pp. 260–276. [Google Scholar]
- Monaghan, P. Resource allocation and life history strategies in birds. Acta Zool. Sin. 2004, 50, 942–947. [Google Scholar]
- Karagicheva, J.; Rakhimberdiev, E.; Saveliev, A.; Piersma, T. Annual chronotypes functionally link life histories and life cycles in birds. Funct. Ecol. 2018, 32, 2369–2379. [Google Scholar] [CrossRef]
- Weiser, E.L.; Brown, S.C.; Lanctot, R.B.; Gates, H.R.; Abraham, K.F.; Bentzen, R.L.; Bêty, J.; Boldenow, M.L.; Brook, R.W.; Donnelly, T.F.; English, W.B.; Flemming, S.A.; Franks, S.E.; Gilchrist, H.G.; Giroux, M.; Johnson, A.; Kennedy, L.V.; Koloski, L.; Kwon, E.; Lamarre, J.; Lank, D.B.; Lecomte, N.; Liebezeit, J.R.; McKinnon, L.; Nol, E.; Perz, J.; Rausch, J.; Robards, M.; Saalfeld, S.T.; Senner, N.R.; Smith, P.A.; Soloviev, M.; Solovyeva, D.; Ward, D.H.; Woodard, P.F.; Sandercock, B.K. Life-history tradeoffs revealed by seasonal declines in reproductive traits of Arctic-breeding shorebirds. J. Avian Biol. 2018, 49, e01531. [Google Scholar] [CrossRef]
- Lu, X.; Ke, D.; Zeng, X.; Yu, T. Reproductive ecology of two sympatric Tibetan snowfinch species at the edge of their altitudinal range: Response to more stressful environments. J Arid Environ 2009, 73, 1103–1108. [Google Scholar] [CrossRef]
- Luo, R.S.; Su, T.P.; Niu, N.; Liang, W. Effect of temperature on timing of reproduction and nesting success in a tropical population of Barn Swallows in south China. Chin. J. Ecol. 2016, 8, 2159–2163. [Google Scholar] [CrossRef]
- Remeš, V.; Matysioková, B.; Vrána, J. Adaptation and constraint shape the evolution of growth patterns in passerine birds across the globe. Front. Zool. 2020, 17, 29. [Google Scholar] [CrossRef] [PubMed]
- Hille, S.M.; Cooper, C.B. Elevational trends in life histories: Revising the pace-of-life framework. Biol. Rev. 2015, 90, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Lu, X. Reproductive ecology of blackbirds (Turdus merula maximus) in a high-altitude location, Tibet. J Ornithol. 2015, 146, 72–78. [Google Scholar] [CrossRef]
- Lu, X. , Gong, G., Zeng, X. Reproductive ecology of Brown-cheeked Laughing Thrushes (Garrulus henrici) in Tibet. J Field Ornithol. 2008, 79, 152–158. [Google Scholar] [CrossRef]
- Boyle, A.W. , Sandercock, B.K., Martin, K. Patterns and drivers of intraspecific variation in avian life history along elevational gradients: A meta-analysis. Biological Reviews 2015, 91, 469–482. [Google Scholar] [CrossRef]
- Deeming, D.C. Importance and evolution of incubation in avian reproduction. — In: Avian incubation: Behaviour, environment, and evolution (Deeming, D.C., ed.). Oxford University Press: Oxford, 2002.
- Colwell, M.A. B.; Millett, J.J.; Meyer, J.N.; Hall, S.J.; Hurley, S.E.; Mcallister, A.; Transou, R.R. LeValley. Snowy plover reproductive success in beach and river habitats. J. Field Ornith. 2005; 76, 373–382. [Google Scholar] [CrossRef]
- Chalfoun, A.D., K. A. Schmidt. Adaptive breeding habitat selection: Is it for the birds? Auk 2012, 129, 589–599. [Google Scholar] [CrossRef]
- Dunn, P. Breeding dates and reproductive performance. Advances in Ecological Research, 2004; 35, 69–87. [Google Scholar] [CrossRef]
- Caro, T. Antipredator defenses in birds and mammals. Chicago: University of Chicago Press; 2005.
- Luo, R.S.; Su, T.P.; Niu, N.; Liang, W. Effect of temperature on timing of reproduction and nesting success in a tropical population of Barn Swallows in south China. Chin. J. Ecol. 2016, 8, 2159–2163. [Google Scholar] [CrossRef]
- Song, S.; Chen, J.N.; Jiang, B.; Liu, N.F. Variation in egg and clutch size of the Black Redstart (Phoenicurus ochruros) at the northeastern edge of the Qinghai-Tibetan Plateau. Avian Res. 2016, 7, 20. [Google Scholar] [CrossRef]
- Boyle, A.W.; Sandercock, B.K.; Martin, K. Patterns and drivers of intraspecific variation in avian life history along elevational gradients: A meta-analysis. Biol. Rev. 2016, 91, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Xiao, H.; Han, H.; Wei, W.; Zhou, H.; Zhang, Z. Comparison of breeding strategies of two sympatric thrush species in an alpine environment. Front. Ecol. Evol. 2022, 10, 1049983. [Google Scholar] [CrossRef]
- Hille, S.M.; Cooper, C.B. Elevational trends in life histories: Revising the pace-of-life frame. Biol. Rev. 2015, 90, 204–213. [Google Scholar] [CrossRef]
- Jetz, W.; Sekercioglu, C.H.; Bohning-Gaese, K. The worldwide variation in avian clutch size across species and space. PLoS Biol. 2008, 12, 2650–2657. [Google Scholar] [CrossRef] [PubMed]
- Stoddard, M.C.; Yong, E.H.; Akkaynak, D.; Sheard, C.; Tobias, J.A.; Mahadevan, L. Avian egg shape: Form, function, and evolution. Science 2017, 356, 1249–1254. [Google Scholar] [CrossRef]
- Du, B.; Liu, C.J.; Yang, M.; Bao, S.J.; Guan, M.M.; Liu, N.F. Horned larks on the Tibetan Plateau adjust the breeding strategy according to the seasonal changes in the risk of nest predation and food availability. J. Avian Biol. 2014, 45, 466–474. [Google Scholar] [CrossRef]
- Hasegawa, M.; Arai, E. Convergent evolution of the tradeoff between egg size and tail fork depth in swallows and swifts. J. Avian Biol. 2018, 49, e01684. [Google Scholar] [CrossRef]
- Charnov, E.L.; Downhower, J.F.; Brown, L.P. Optimal offspring sizes in small litters. Evol. Ecol. 1995, 9, 57–63. [Google Scholar] [CrossRef]
- Hargitai, R.; Török, J.; Tóth, L.; Hegyi, G.; Rosivall, B.; Szigeti, B.; Szöllősi, E. Effects of environmental conditions and parental quality on inter- and intraclutch egg-size variation in the Collared Flycatcher (Ficedula albicollis). Auk 2005, 122, 509–522. [Google Scholar] [CrossRef]
- Christians, J.K. Avian egg size: Variation within species and inflexibility within individuals. Biol. Rev. 2002, 77, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Ruffino, L.; Salo, P.; Koivisto, E.; Banks, P.B.; Korpimaki, E. Reproductive responses of birds to experimental food supplementation: A meta-analysis. Front. Zool. 2014, 11, 80. [Google Scholar] [CrossRef]
- Hallmann, K.; Griebler, E.M. An exploration of differences in the scaling of life history traits with body mass within reptiles and between amniotes. Ecol. Evol. 2018, 8, 5480–5494. [Google Scholar] [CrossRef] [PubMed]
- Yanagi, S.; Tuda, M. Female size constrains egg size via the influence of reproductive organ size and resource storage in the seed beetle Callosobruchus chinensis. J. Insect Physiol. 2012, 58, 1432–1437. [Google Scholar] [CrossRef] [PubMed]
- Krist, M. Egg size and offspring quality: A meta-analysis in birds. Biol. Rev. 2011, 86, 692–716. [Google Scholar] [CrossRef]
- Bouakkaz, A.; Belhassini, T.; Bensouilah, M.A. Bensouilah & M. Houhamdi. Breeding behaviour of the Kentish Plover (Charadrius alexandrinus) in a salt marsh frothe Eastern High Plateaux, northeast Algeria. Journal of King Saud University-Science, 2017; 29, 291–301. [Google Scholar] [CrossRef]
- Liang, T.; Zhou, L.; He, W.F.; Shi, L. Variations in the reproductive strategies of three populations of Phrynocephalus helioscopus in China. Peer J 2018, 6, e5705. [Google Scholar] [CrossRef]
- Székely, T. Why study plovers? The significance of non-model organisms in avian ecology, behaviour and evolution. J. Ornithol. 2019, 160, 923–933. [Google Scholar] [CrossRef]
- Colwell, M.A.; Haig, S.M. The population ecology and conservation of Charadrius Plovers. Taylor & Francis Group: Florida, USA, 2019; pp. 91–127.
- Elgar, M.A.; Heaphy, L.J. Covariation between clutch size, egg weight and egg shape: Comparative evidence for chelonians. J. Zool. 1989, 219, 137–152. [Google Scholar] [CrossRef]
- Kratochvíl, L.; Frynta, D. Egg shape and size allometry in geckos (Squamata: Gekkota), lizards with contrasting eggshell structure: Why lay spherical eggs? [J]. Journal of Zoological Systematics and Evolutionary Research 2010, 44, 217–222. [Google Scholar] [CrossRef]
- Fraga, R.M.; Amat, J.A. Breeding biology of a Kentish plover (Charadrius alexandrinus) population in an inland saline lake. Ardeola 1996, 43, 69–85. [Google Scholar]
- Fojt, E.; Triplet, P.; Robert, J.C.; Stillman, R.A. Comparison of the breeding habitats of Little Ringed Plover Charadrius dubius and Kentish plover Charadrius alexandrinus on a shingle bed. Bird Study 2000, 47, 8–12. [Google Scholar] [CrossRef]
- del Hoyo, J.; Wiersma, P.; Kirwan, G.M. Kentish plover (Charadrius alexandrinus), version 1.1. In Birds of the World; Keeney, B.K. Cornell Lab of Ornithology: Ithaca, NY, USA, 2021. [Google Scholar] [CrossRef]
- Alrashidi, M.; Kosztolanyi, A.; Shobrak, M.; Kupper, C.; Szekely, T. Parental cooperation in an extreme hot environment: Natural behaviour and experimental evidence. Anim. Behav. 2011, 82, 235–243. [Google Scholar] [CrossRef]
- Zheng, G.M. A Checklist on the Classification and Distribution of the Birds of China, 3rd ed.; Science Press: Beijing, China, 2017; p. 68. [Google Scholar]
- Wang, D.; Zheng, S.C.; Wang, P.; Matsiko, J.; Sun, H.Z.; Hao, Y.F.; Li, Y.M.; Zhang, Z.W.; Que, P.J.; Meng, D.R.; Zhang, Q.H.; Jiang, G.B. Effects of migration and reproduction on the variation in persistent organic pollutant levels in Kentish plovers from Cangzhou Wetland, China. Sci. Total Environ. 2019, 670, 122–128. [Google Scholar] [CrossRef]
- Zeraoula, A.; Bensouilah, T.; Brahmia, H.; Bouslama, Z.; Houhamdi, M. Breeding biology of the European Blackbird Turdus merula in orange orchards. J King Saud Univ Sci 2015, S1018364715000981. [Google Scholar] [CrossRef]
- Potapov, R.L. Adaptation of birds to life in high mountains in Eurasia. Acta Zool Sin 2004, 50, 970–977. [Google Scholar]
- Ghalambor, C.K.; Martin, T.E. Fecundity-survival trade-offs and parental risk-taking in birds. Science 2001, 292, 494–497. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.E. Age-related mortality explains life history strategies of tropical and temperate songbirds. Science 2015, 349, 966–970. [Google Scholar] [CrossRef]
- Martin, K. and Wiebe, K.L. Coping mechanisms of alpine and arctic breeding birds: Extreme weather and limitations to reproductive resilience. Integr Comp Biol 2004, 44, 177–185. [Google Scholar] [CrossRef]
- Badyaev, A.V.; Ghalambor, C. K. 2001. Evolution of life histories along elevational gradients: Trade-off between parental care and fecundity. Ecology 2001, 8282, 2948–2960. [Google Scholar] [CrossRef]
- Johnson, L.S.; Ostlind, E.; Brubaker, J.L.; Balenger, S.L.; Johnson, B.G.P.; Golden, H. Changes in egg size and clutch size with elevation in a Wyoming population of Mountain Bluebirds. Condor 2006, 108, 591. [Google Scholar] [CrossRef]
- Bears, H.; Martin, K.; White, G.C. Breeding in high-elevation habitat results in shift to slower life-history strategy within a single species. J. Anim. Ecol. 2009, 78, 365–375. [Google Scholar] [CrossRef]
- Amat, J.A.; Fraga, R.M.; Arroyo, G.M. Intraclutch egg-size variation and offspring survival in the Kentish plover Charadrius alexandrinus. Ibis 2001, 143, 17–23. [Google Scholar] [CrossRef]
- Rocha, A.D.; Fonseca, D.; Masero, J.A. Coastal saltpans are a good alternative breeding habitat for Kentish plover Charadrius alexandrinus when umbrella species are present. J. Avian Biol. 2016, 47, 824–833. [Google Scholar] [CrossRef]
- Bouakkaz, A.; Belhassini, K.; Bensouilah, T.; Bensouilah, M.A.; Houhamdi, M. Breeding behaviour of the Kentish plover (Charadrius alexandrinus) in a salt marsh from the Eastern High Plateaux, northeast Algeria. J. King Saud Univ. Sci. 2017, 29, 291–301. [Google Scholar] [CrossRef]
- Que, P.; Chang, Y.J.; Eberhart-phillips, L.J.; Liu, Y.; Székely, T. Low nest survival of a breeding shorebird in Bohai Bay, China. J. Ornithol. 2015, 156, 297–307. [Google Scholar] [CrossRef]
- Song, Z.T.; Lin, X.; Que, P.J.; Halimubieke, N.; Huang, Q.J.; Zhang, Z.W.; Székely, T.; Liu, Y. The allocation between egg size and clutch size depends on local nest survival rate in a mean of bet-hedging in a shorebird. Avian Res. 2020, 11, 40. [Google Scholar] [CrossRef]
- Ma, M. A Checklist on the Distribution of the Birds in Xinjiang. Science Press: Beijing, China, 2011; pp. 49–50.
- Vincze, O.; Kosztolányi, A.; Barta, Z.; Küpper, C.; Alrashidi, M.; Amat, J.A. Parental cooperation in a changing climate: Fluctuating environments predict shifts in care division. Global Ecol. Biogeogr. 2017, 26, 347–358. [Google Scholar] [CrossRef]
- Abuduwaili, J. , Zhang, Z.Y., Jiang, F.Q. Assessment of the distribution, sources and potential ecological risk of heavy metals in the dry surface sediment of Aibi Lake in Northwest China. PLoS ONE 2015, 10, e0120001. [Google Scholar] [CrossRef]
- Ma, M.; Bayahen, K.; Li, F.; Hu, B.W.; Wu, J.Q.; Douglas, M.; Gao, X.; Amanjiang, R.; Chen, Y.; Mei, Y.; Ding, P. List of birds and count of Autumn migration in Ebinur Wetland Nature Reserve. Sichuan, J. Zool. 2010, 6, 912–918. [Google Scholar]
- Ye, Z.; Chen, S.; Zhang, Q.; Liu, Y.; Zhou, H. Ecological water demand of Taitema Lake in the lower reaches of the Tarim River and the Cherchen River. Remote Sens. 2022, 14, 832. [Google Scholar] [CrossRef]
- Zuo, T.; Chen, Y.; Ding, J. Research on vegetation coverage dynamics and prediction in the Taitema Lake region. Water 2022, 14, 725. [Google Scholar] [CrossRef]
- Szekely, T.; Kosztolanyi, A.; Kupper, C. Practical guide for investigating breeding ecology of Kentish plover Charadrius alexandrinus. University of Bath: Bath, UK, 2008.
- Hoyt, D.F. Practical methods of estimating volume and fresh weight of bird eggs. Auk 1979, 96, 73–77. [Google Scholar] [CrossRef]
- Norte, A.C.; Ramos, J.A. Nest-site selection and breeding biology of Kentish plover Charadrius alexandrinus on sandy beaches of the Portuguese west coast. Ardeola 2004, 51, 255–268. [Google Scholar]
- Weggler, M. Constraints on, and determinants of, the annual number of breeding attempts in the multi-brooded Black Redstart Phoenicurus ochruros. Ibis 2006, 148, 273–284. [Google Scholar] [CrossRef]
- Bears, H.; Martin, K.; White, G.C. Breeding in high-elevation habitat results in shift to slower life-history strategy within a single species. J. Anim. Ecol. 2009, 78, 365–375. [Google Scholar] [CrossRef]
- Martin, T.E. , 2008. Egg size variation among tropical and temper-ate songbirds: An embryonic temperature hypothesis. Proc. Natl Acad. Sci. 2008, 105, 9268–9271. [Google Scholar] [CrossRef]
- Yampolsky, L.Y.; Scheiner, S.M. Why larger offspring at lower temperatures? A demographic approach. Am. Nat. 1996, 147, 86–100. [Google Scholar] [CrossRef]
- Boulton, R.L.; Cassey, P. How avian incubation behaviour influences egg surface temperatures: Relationships with egg position, development and clutch size. J. Avian Biol. 2012, 43, 289–296. [Google Scholar] [CrossRef]
- Cooney, C.R.; Sheard, C.; Clark, A.D.; Healy, S.D.; Liker, A.; Street, S.E.; Troisi, C.A.; Thomas, G.H.; Székely, T.; Hemmings, N.; Wright, A.E. Ecology and allometry predict the evolution of avian developmental durations. Nat. Commun. 2020, 11, 2383. [Google Scholar] [CrossRef]
- Barta, Z.; Szekely, T. The optimal shape of avian eggs. Funct. Ecol. 1997, 11, 656–662. [Google Scholar] [CrossRef]
- Duursma, D.E.; Gallagher, R.V.; Price, J.J.; Griffith, S.C. Variation in avian egg shape and nest structure is explained by climatic conditions. Sci. Rep. 2018, 8, 4141. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Map of the study areas of three populations of Charadrius alexandrinus in Xinjiang.
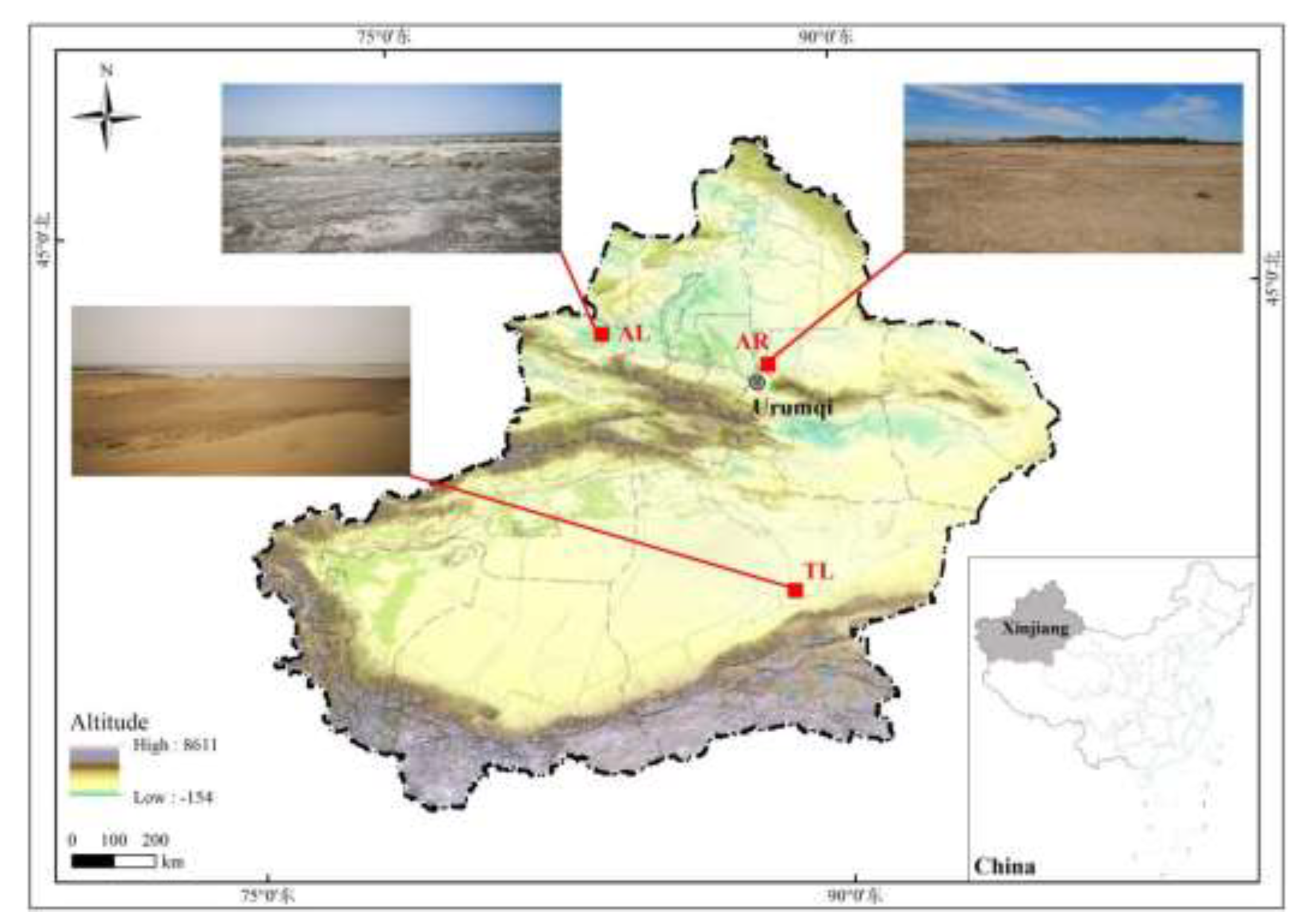
Figure 2.
Temperature and precipitation in the three regions (DTmax: Daily Maximum Temperature, DTmin: Daily Minimum Temperature, AT: Average Temperature, RF: Rainfall. Data were averages for 2019 and 2020).
Figure 2.
Temperature and precipitation in the three regions (DTmax: Daily Maximum Temperature, DTmin: Daily Minimum Temperature, AT: Average Temperature, RF: Rainfall. Data were averages for 2019 and 2020).
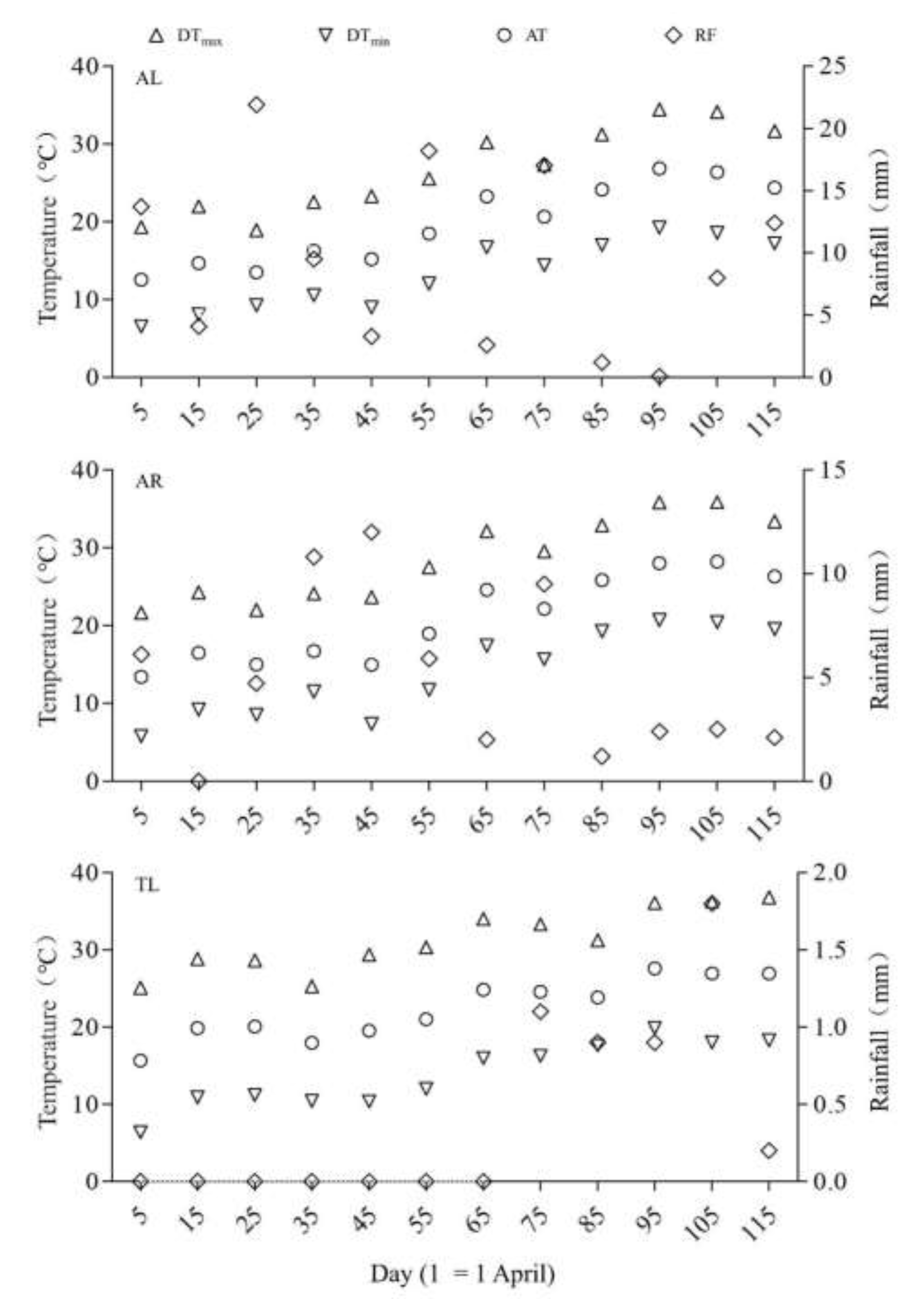
Figure 3.
Regressions of female body mass (A), clutch size (B), egg length (C), egg width (D), egg volume (E), clutch volume (F) and laying date from three populations of C. alexandrinus.
Figure 3.
Regressions of female body mass (A), clutch size (B), egg length (C), egg width (D), egg volume (E), clutch volume (F) and laying date from three populations of C. alexandrinus.
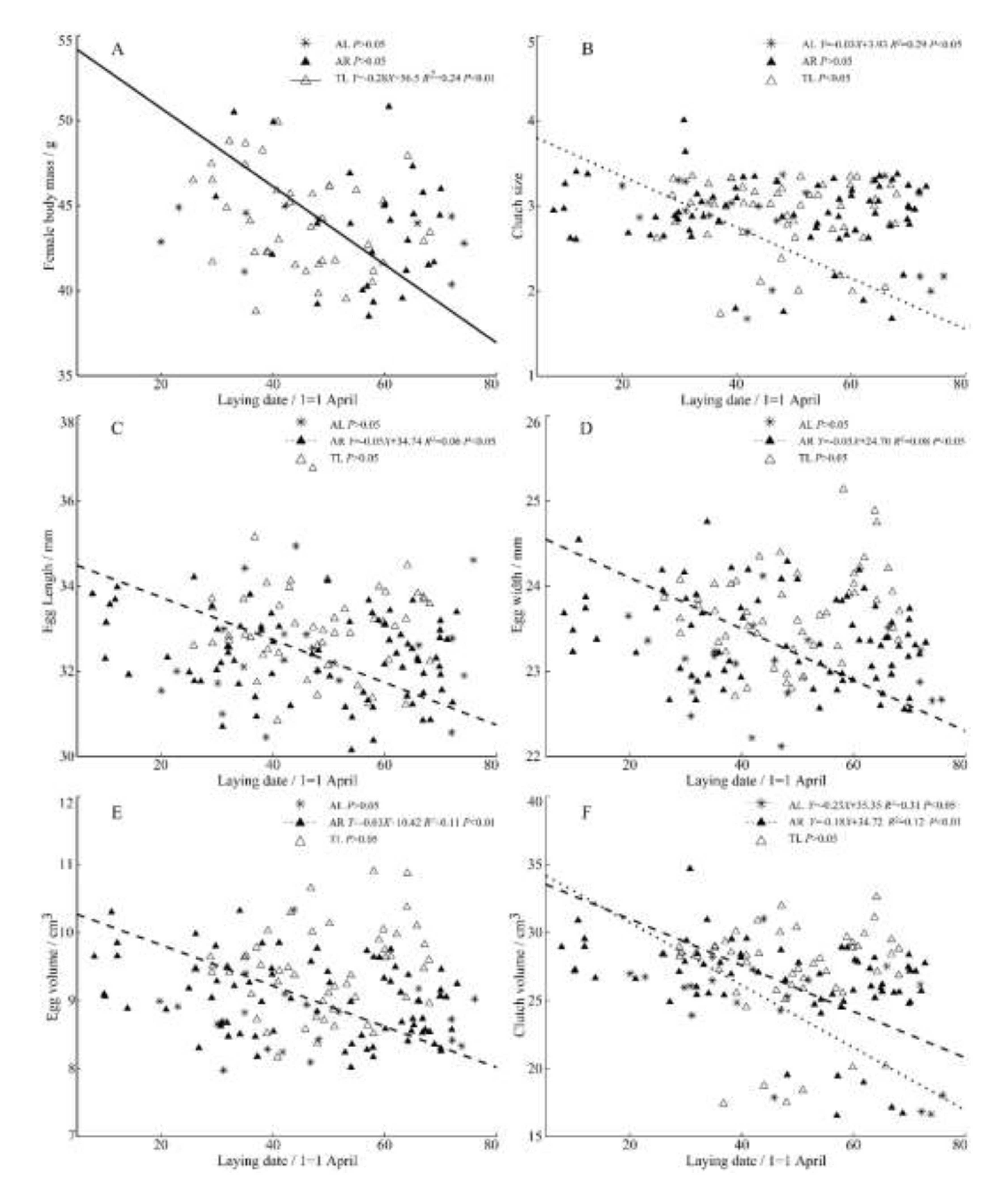
Figure 4.
Regressions of female body mass and clutch size (A), egg width (B), egg volume (C), clutch volume (D) from three populations of C. alexandrinus.
Figure 4.
Regressions of female body mass and clutch size (A), egg width (B), egg volume (C), clutch volume (D) from three populations of C. alexandrinus.
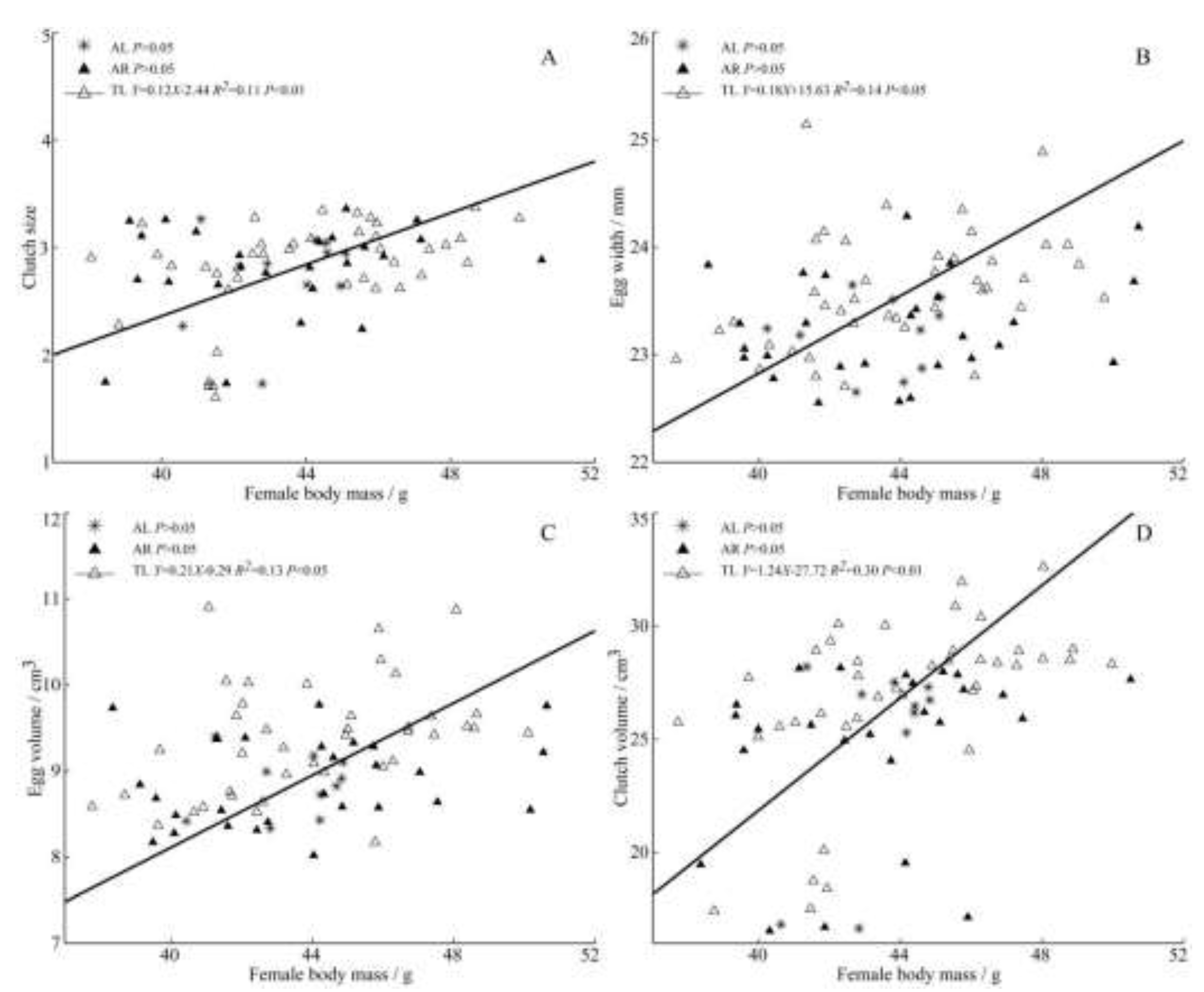
Figure 5.
Regressions of female tarsometatarsus length and egg length (A), clutch volume (B) from three populations of C. alexandrinus.
Figure 5.
Regressions of female tarsometatarsus length and egg length (A), clutch volume (B) from three populations of C. alexandrinus.
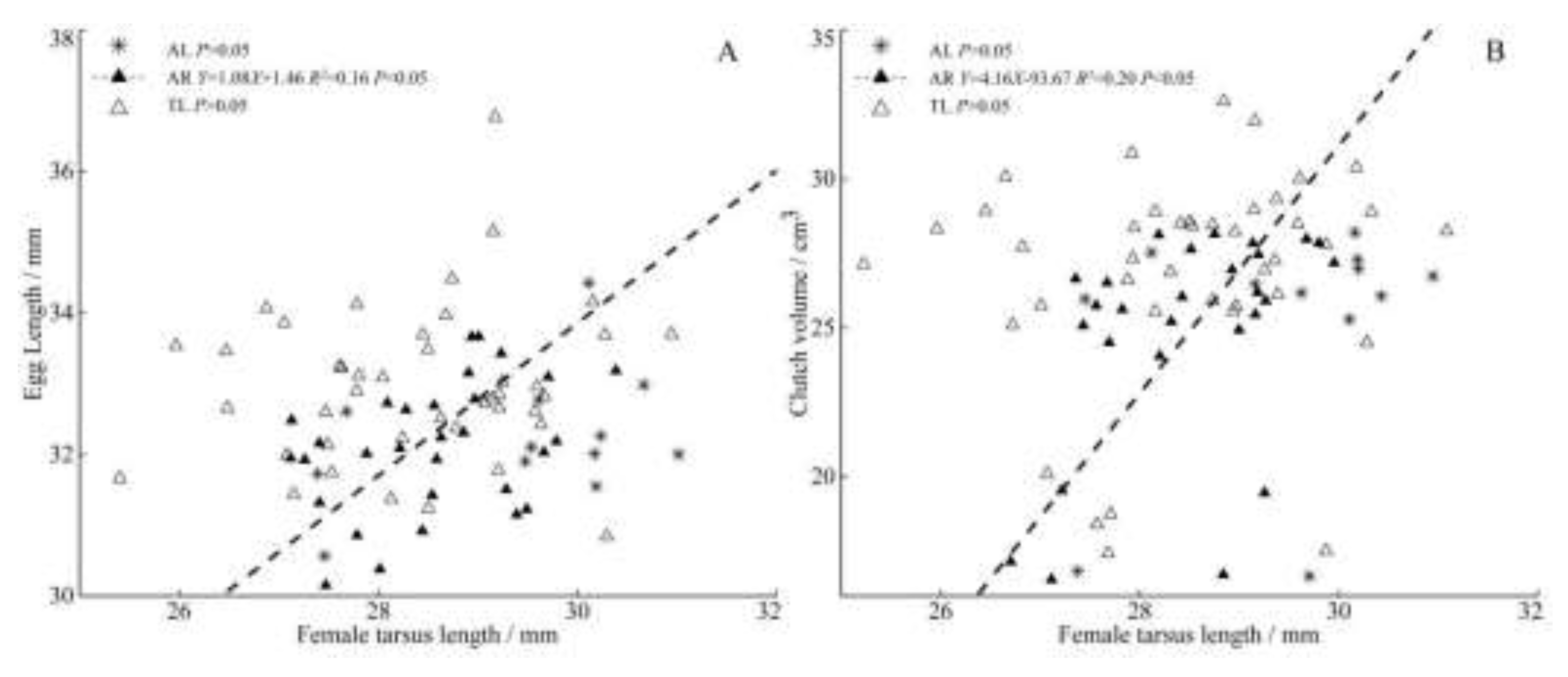
Figure 6.
Allometry of egg shape of C. alexandrinus (A-egg shape allomery, B-egg shape allometry by residuals).
Figure 6.
Allometry of egg shape of C. alexandrinus (A-egg shape allomery, B-egg shape allometry by residuals).
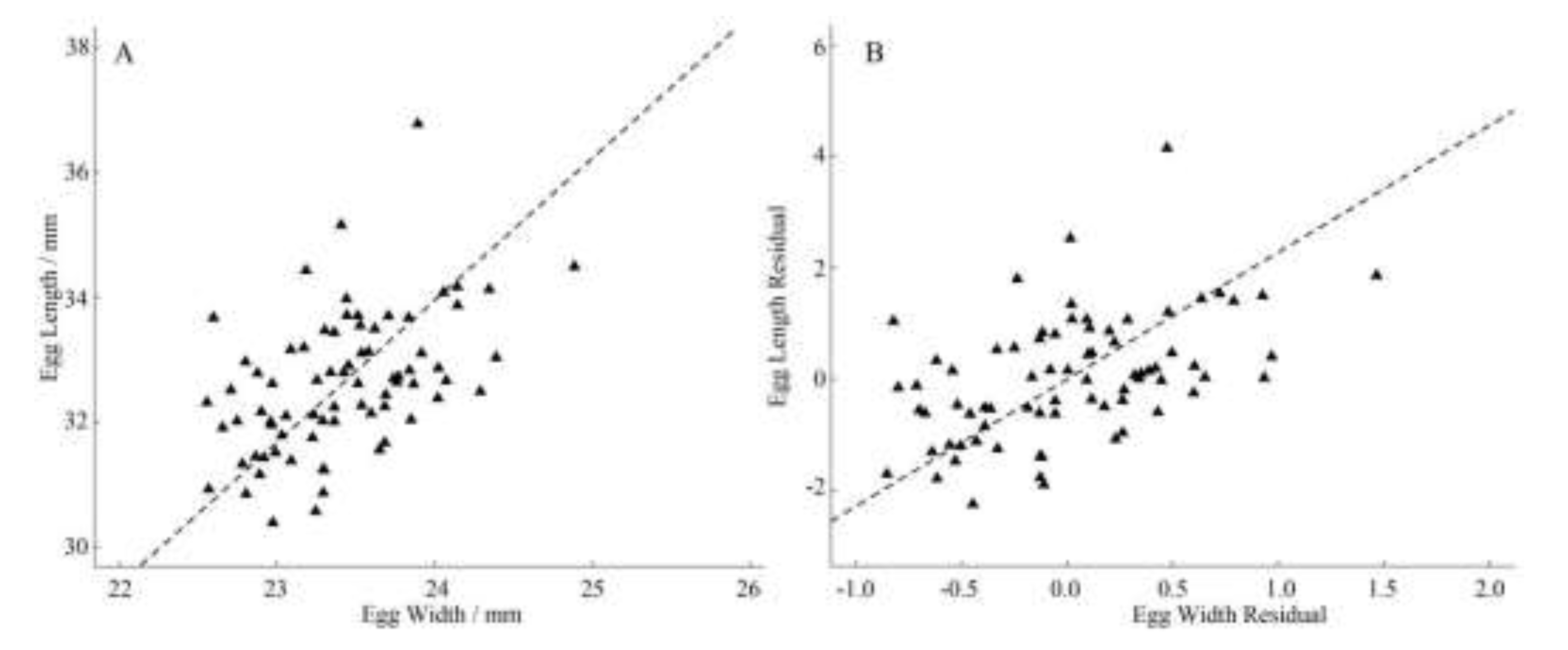

Table 1.
Descriptive statistics of female reproductive traits in three populations of C.alexandrinus.
Table 1.
Descriptive statistics of female reproductive traits in three populations of C.alexandrinus.
| Traits | AL(n=20) | AR(n=77) | TL(n=61) | F-level and | P-value |
|---|---|---|---|---|---|
| Female body mass *#, g | 43.42±1.52 | 43.79±3.37 | 43.95±2.93 | F2, 75=0.127, | P=0.881 |
| Female tarsometatarsus length *#, mm | 29.47±1.12 a | 28.51±0.84 b | 28.38±1.23 b | F2, 81=4.602, | P=0.012 |
| Egg length †, mm | 32.38±1.30 B | 32.36±1.08 B | 33.01±1.24 A | F2, 437=15.810, | P<0.001 |
| Egg width #, mm | 23.10±0.55 Bb | 23.38±0.55 Ba | 23.64±0.54 A | F2, 436=23.070, | P<0.001 |
| Egg shape # | 0.71±0.03 | 0.72±0.03 | 0.72±0.03 | F2, 436=3.961 | P=0.020 |
| Egg volume #, cm3 | 8.80±0.60 Bb | 9.02±0.58 Ba | 9.40±0.64 A | F2, 436=28.790 | P<0.001 |
| Clutch size † | 2.75±0.43 | 2.96±0.31 | 2.87±0.34 | χ2=5.929 | P=0.052 |
| Clutch volume †, cm3 | 24.57±4.02 Bb | 26.39±3.19 a | 27.07±3.50 A | χ2=12.249 | P=0.002 |
| Incubation period &, day | / | 26±1.33 a | 25.09±0.51 b | U=54.50 | P=0.037 |
Note: Different capital letters indicate significance at the P<0.01 level; Different lowercase letters indicate significance at the P<0.05 level. The monitoring of AL population is too difficult and the number of nests was too small, so the data of incubation period could not be obtained by continuous monitoring. *AL n = 12; AR n = 30; TL n = 42. # ANOVA; †Nonparametric Kruskal-Wallis test; &Nonparametric Mann-Whitney U test.
Table 2.
Model selection for reproductive output of C.alexandrinus.
| Parameter | Optimization model | |
|---|---|---|
| Egg volume | Clutch volume | |
| Clutch size | ||
| Female body mass | + | + |
| Female tarsometatarsus length | + | |
| Laying date | ||
| Egg length | + | |
| Egg width | ||
| Average temperature | + | + |
| Daily temperature difference | ||
| Daily maximum temperature | ||
| Rainfall | ||
| Population | ||
| AICc | -197.50 | -197.00 |
| Weight | 0.948 | 0.611 |
Note: +indicates that the selected parameters of the optimal model; Weight was the proportion of the selected parameters in all parameters of the optimal model.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



