
Submitted:
21 June 2023
Posted:
22 June 2023
You are already at the latest version
Abstract
Trichoderma sp. is a fungus in the genus Trichoderma, ubiquitous in soils, on plant roots and in decaying plant residues. Due to its competitiveness and mycoparasitic potential against other microorganisms, particular strains of Trichoderma sp. are used in agriculture as biocontrol agents against plant pathogens. However, Trichoderma afroharzianum. has been recently reported as a pathogen causing ear rot disease on maize in Germany, France and Italy leading to massive infections on maize cobs. This raised the question, whether and to what extend Trichoderma can infect other cereal crops than maize and cause disease symptoms and yield losses. To address this question, two varieties of wheat, barley and sorghum millet were grown in the greenhouse and artificially inoculated with T. afroharzianum by both spray and point inoculation at the time of flowering. Disease severity was scored weekly and thousand-kernel weight and colonization rate were determined after harvest. As early as 14 days after inoculation, the first visual symptoms appeared on wheat and barley as tan or brown discoloration of the base of a floret within the spikelets. After spray inoculation, clear discolorations of the entire ear were seen, while point inoculation only showed symptoms at the injection site and above. No visible symptoms were observed on sorghum millet. The colonization rate on wheat and barley grains was significantly increased compared to the control while thousand kernel weights (TKW) were significantly reduced. No differences in colonization rate and TKW compared to the control were observed in sorghum. This is the first report of Trichoderma afroharzianum infecting wheat and barley, causing diseases symptoms and significantly reducing thousand-kernel weights.
Keywords:
Trichoderma
; maize
; wheat
; barley
; host range
1. Introduction
Cereals belong to the family of Poaceae and are broadly used as a staple food for humans and livestock around the world. Among the most important cereals are wheat, barley and sorghum, which are grown and consumed globally. Wheat is one of the most widely grown cereals and provides food for a large part of the world's population. Global wheat production is estimated at 772 million tons in 2021 [1]. Barley is another important cereal used as animal feed and for human consumption . While wheat and barley are mainly produced under moderate climate conditions, sorghum is adapted to warmer climates and is one of the most important crops for human consumption and animal feed in tropical and subtropical regions. In 2018, sorghum was the world's fifth largest crop - after wheat, corn, rice and barley [1].
Increases in the cultivation of cereals within corn rotations, combined with higher summer temperatures and increasing drought, have increased problems from infections with fungal pathogens [2,3]. Trichoderma spp. are members of the filamentous fungi and are found worldwide in soils, on plant roots, on decaying plant debris and on wood. They are considered competent substrate colonizers and antagonists of other microorganisms and are therefore used for biological control of plant diseases worldwide [4,5,6]. Trichoderma spp. may promote plant health by either directly countering pathogens through mycoparasitism, competitive inhibition, and antibiosis or indirectly via enhanced plant growth and stimulating systemic plant defenses [7,8]. Trichoderma showed positive effects on plant growth and suppression of important cereal diseases. It has been shown to be effective in controlling Fusarium head blight, Fusarium crown rot and dumping off by producing enzymes and secondary metabolites that inhibit the growth and reduce the levels of mycotoxins in wheat grains [9]. Thus, it had positive effects on suppressing leaf diseases like tan spot disease caused by Pyrenophora tritici-repentis [10] and rust diseases caused by Puccinia spp. on wheat and barley [11,12,13]. Manzar et al. (2021) indicated that T. asperellum is a potential biocontrol agent for managing anthracnose in sorghum [14]. In addition, the root colonization promotes root growth and enhancing nutrient uptake, which can improve the overall health and productivity of cereal crops [13,15,16].
Though widely considered beneficial, members of this fungal genus have been reported to be pathogenic on commercially important plants. In 2020, Trichoderma afroharzianum was reported for the first time in Europe as an ear rot disease in maize [17]. In this report, T. afroharzianum ear rot was characterized by massive production of gray-green spores in the inter-kernel regions and on the outer surface of the husks causing a significant reduction of dry matter content and premature germination of kernels [17]. Since then, the pathogen has also been reported from France and Italy [18].
The recent spread of Trichoderma ear rot in maize raised the question whether T. afroharzianum can also infect other cereal species which are commonly grown in crop rotation cycles with maize. Therefore, barley (Hordeum vulgare), wheat (Triticum aestivum) and sorghum millet (Sorghum bicolor) were cultivated in the greenhouse and inoculated at the time point of flowering with a mixture of pathogenic T. afroharzianum strains. Visual disease symptoms were assessed weekly and thousand-kernel weight and colonization rate were determined after harvest.
2. Materials and Methods
Figure 1.
Method of pathogenicity testing with Trichoderma in wheat, sorghum and barley.
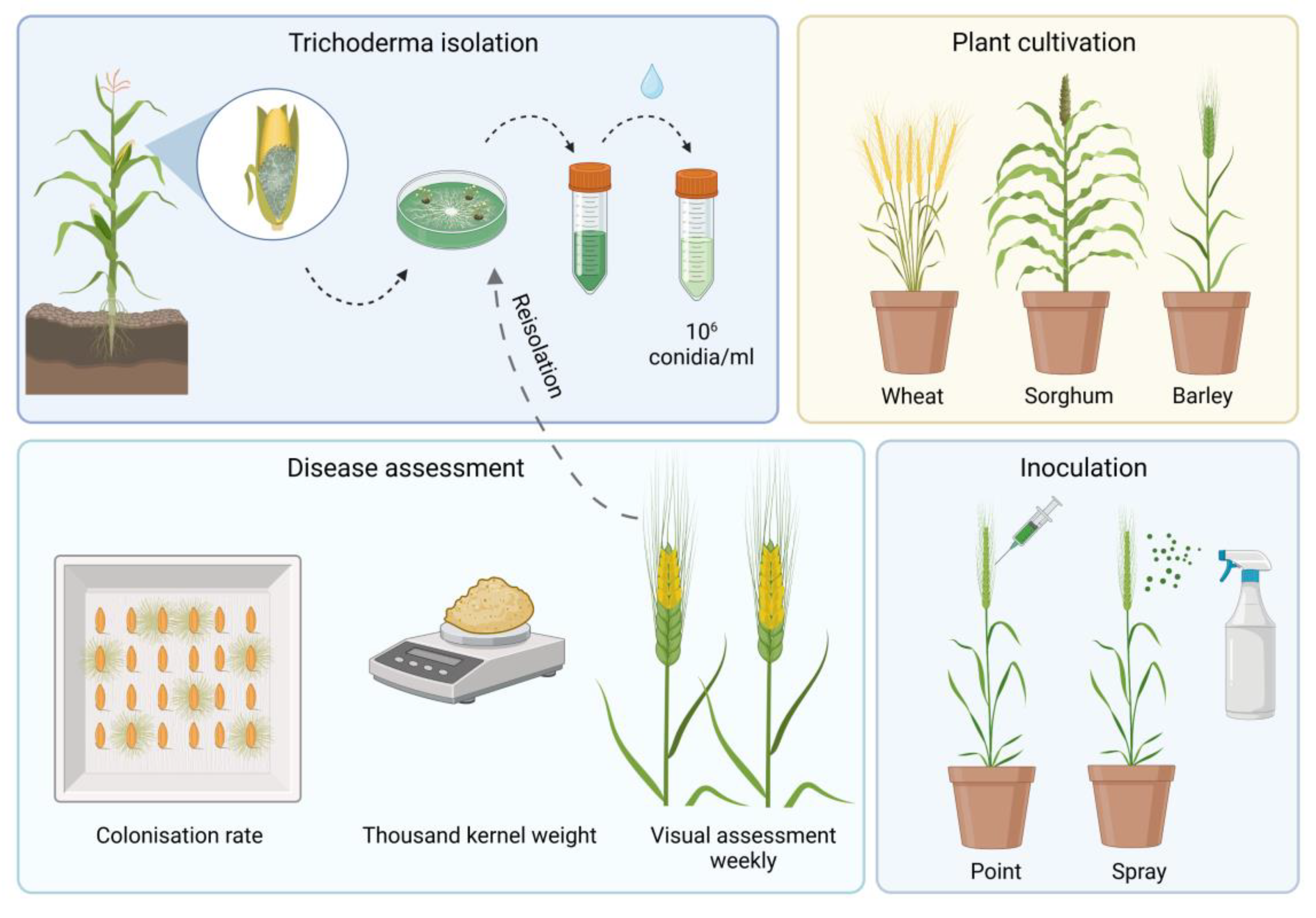
2.1. Fungal Cultivation and Inoculum Preparation
Trichoderma strains were collected from naturally infected silage and grain maize in Germany in 2018 and 2019 (Table 1) [17]. Spore suspension was produced as described by Pfordt et al. (2019) and diluted to 106 conidia/ml. Tri1, Tri2, and Tri3 were mixed in the same ratio to prepare one mixed isolate (TriMix). Plants were inoculated with one pathogenic T. afroharzianum strain (TriMix) and the apathogenic T. afroharzianum type strain from the Westerdiijk Institute CBS124620 (CBS, Utrecht, NL). Control plants were inoculated with water (control).
2.2. Plant Cultivation and Inoculation Procedure
Two varieties of wheat (Triticum aestivum), barley (Hordeum vulgare) and sorghum (Sorghum bicolor) were chosen, respectively, for pathogenicity testing in the greenhouse. One winter wheat variety and one spring wheat variety, one spring malt barley and one winter barley and two sorghum varieties (silo and grain type) were used.
Kernels of winter wheat and winter barley were vernalized for seven weeks at 4°C and planted in 7 cm diameter pots filled with a mixture of potting soil, sand and compost (3x1x3). Pots were placed in the greenhouse at 22°C and seasonal light cycle. Ten plants in two repetitions were inoculated with Trichoderma and five plants per repetition were inoculated with sterile water, which served as control. Inoculation was conducted with two different inoculation methods at full anthesis. Point inoculation was carried out by syringe injecting 25 μl of spore suspension into the center of two florets. Spray inoculation was conducted by spraying 2 ml spore suspension from two sides on cereal heads. Ears were covered with plastic bags for 48 hours post inoculation.
2.4. Disease Assessment
Severity of infection was visually scored weekly as percentage (0-100%) of diseased ear tissue. After harvest, kernels were counted, weighed and the weight extrapolated to 1,000 grains to calculate thousand kernel weight. To investigate the colonization rate, harvested seeds were surface disinfected for ten minutes with 0.25% silver nitrate and placed on filter paper in growth chambers under sterile conditions. After 3 days, kernels with Trichoderma outgrowth were counted and colonization rate was assessed by the number of infected kernels divided by the total number of analyzed kernels.
2.3. Germination Rate in Pots
Twenty seeds per treatment of previously inoculated grains were planted in pots in the greenhouse. Seeds were surface disinfected for ten minutes with 0.25% silver nitrate and planted in 4 cm diameter pots filled with a mixture of potting soil, sand and compost. Pots were placed in the greenhouse at 22°C with 12h day-/night light cycle and watered as needed. Four weeks after planting the emergence rate was assessed.
2.5. Statistical Analysis
Statistical analysis was conducted using STATISTICA version 13 (Statistica GmbH, Germany). Differences between means of colonization rate and thousand-kernel weight was analyzed using parametric ANOVA by 5% probability. Analysis of variance (ANOVA) was carried out by Tukey-HSD-test at 5% probability.
3. Results
3.1. Visual Disease Symptoms
First symptoms of Trichoderma infection occurred two weeks after inoculation. Distinct symptoms were observed as tan or brown discoloration of the base of a floret within the spikelets. Later, the base of the infected spikelets and parts of the rachis developed a dark brown color. The browning started at the injection side around the inoculated spikelet and spread upwards the ear until all spikelets above the infection showed discolorations. After spray inoculation, browning started at several spikelets at the whole ear and merged until the whole ear was discolored. As the infection progressed, infected spikelets bleached to become light tan. Infected kernels became shriveled, chalky, and kernels were light weighted (Figure 2A,B).
Symptoms in barley were much less distinct. The brownish discoloration occurred three weeks after inoculation at the site of point inoculation. Trichoderma infection appeared as premature bleaching of individual or several spikelets with tan discolorations at the base. The seeds in blighted heads did not fill properly and appear shriveled and bleached (Figure 2C).
No visible disease symptoms were observed in sorghum (Figure 2D).
The first disease symptoms on spring wheat were observed 14 days after inoculation with TriMix, while disease symptoms on winter wheat occurred later, after 21 days (Figure 3). No symptoms were visible on control plants and CBS inoculated spring wheat plants, however, slight symptoms were observed in winter wheat. Symptoms on wheat quickly developed to progressive bleaching of spikes and spikelets until 35 dpi (Figure 3). Later, the further disease progress could no longer be followed visually because heads turned yellow due to repining. Disease severity was highest after spray inoculation with TriMix leading to more than 20% in both wheat varieties. Point inoculation with syringe lead to more than 15% disease severity in both varieties. Both TriMix treatments showed significantly higher disease levels than water inoculated control plants. However, no disease symptoms were observed after inoculation with the CBS strain.
Winter barley showed first disease symptoms 21 days after point inoculation with TriMix. Infection lead to 32% of disease severity at 28 dpi. No visible symptoms were observed after spray inoculation in both varieties. Neither the water inoculated control plants nor CBS inoculated plants showed any disease symptoms after inoculation with both methods.
3.2. Colonization Rate and Thousand-Kernel Weight
After maturity, whole ears were harvested, threshed, weighed and thousand kernel weight was calculated. Afterwards, hundred kernels were placed on filter paper and colonization rate was assessed. Colonization rate after TriMix inoculation was significantly higher of both wheat and barley varieties than water and CBS inoculated plants. Colonization rate in winter barley after TriMix inoculation was the highest (47% of infected kernels) followed by spring barley with 20% (Figure 4). CBS also led to a slight colonization of kernels of winter wheat, however, it did not differ significantly from control plants. The thousand-kernel weight was significantly reduced after TriMix inoculation in both wheat varieties in comparison to water inoculated control plants. Both wheat varieties showed similarly high colonization rates with 48.6% and 49.3% of infected kernels after TriMix inoculation, which was significantly higher than CBS and water inoculated plants (Figure 5). Thousand kernel weight was not affected by TriMix inoculation overall both inoculation methods. However, when the two inoculation methods are considered separately, it is shown that point inoculation resulted in a significant reduction in thousand kernel weight in both varieties (Table 2).
After artificial inoculation in sorghum, neither colonization rate, nor thousand-kernel weight was affected by the inoculation of pathogenic TriMix isolate (Figure 6).
There were significant differences found in the colonization rate and the thousand-kernel weight in relation to the inoculation method used, which was consistent for barley, wheat and sorghum (Table 2). Inoculation with TriMix resulted in significantly higher colonization rates and a significant reduction of thousand-kernel weight in winter barley, while spring barley showed no significant differences compared to control plants. Both inoculation methods resulted in higher colonization rates compared to control plants. Spray inoculation lead to higher colonization rate of 36.4% than point inoculation 30.2%. However, only spray inoculation reduced the thousand-kernel weight significantly. Both varieties of wheat were highly infected with, respectively 48.6% and 49.3% colonization rate. Similar to barley, spray inoculation led to a higher colonization rate (59.3%) than spray inoculation (38.4%). However, point inoculation significantly reduced thousand-kernel weight in comparison to water inoculated control plants.
No significant differences were observed between the inoculation method in sorghum after TriMix inoculation (Table 2).
3.3. Germination Rate
Germination rates in different crops were reduced after TriMix infection, however no significant differences could be analyzed due to low treatment number. 83% of water inoculated control kernels of wheat were germinated, however germination rate after TriMix inoculation was reduced to 75% over all repetitions (Figure 7).
4. Discussion
The results of the present study demonstrate that Trichoderma can infect wheat and barley and causes visual disease symptoms as well as a reduction of thousand kernel weight after artificial inoculation in the greenhouse. This is the first report describing Trichoderma afroharzianum as an ear-infecting pathogen in wheat and barley. Symptoms of infection were determined as tan and brownish discoloration of spikelets, which spread and coalesce until the entire kernels are discolored. Finally, grains shriveled and bleached. Apparently, water and nutrient supply are cut off reducing kernel size and shriveling of kernels. These symptoms can be confused with extended symptoms of other wheat and barley diseases like Fusarium head blight, spot blotch or kernel smudge. This could be the reason why the disease has not yet been described in the field.
Severity of visual disease symptoms, in general, corresponds with colonization rate. High disease severity in wheat and barley after spray inoculation (25%) led to high colonization rates (50%), in contrast to low symptom severity after point inoculation (15%) which resulted in lower colonization rates (35%). However, colonization rate was higher as visual disease severity indicating that more kernels were infected without showing symptoms. Moreover, no symptoms were observed in winter barley, although assessment of colonization rate showed 19.6% infected kernels. Visual ratings tended to underestimate the actual disease severity, probably because infected seeds partially appeared symptomless.
Two inoculation methods were used in these investigations to study the ability of Trichoderma to infect and spread within the ears of cereal crops [19]. Both inoculation methods led to severe infections, however, point inoculation caused visible symptoms only at the site of inoculation and above, while spray inoculation led to severe infections on the whole ear. Point inoculation is a useful technique to investigate fungal growth and spread from the initial inoculation point within the head. In contrast, spray inoculation is mainly used to study the ability of the pathogen to initially infect the tissue without previous injuries [20]. The point inoculation method injects conidia via syringe directly inside a floret, which not only causes an artificial wound for fungal entry, but also provides optimal conditions for fungal colonization in the spike tissues. This inoculation method does not determine the fungal ability to cause initial infection but provides information about the ability of the fungus to spread inside the ear between the florets [21]. In contrast, the spray inoculation with conidia suspensions without wounding spike tissues, can assess the ability of the fungus to cause initial infection like it is happening under natural infection in the field [22]. This study successfully demonstrates that Trichoderma can initially infect wheat and barley after spray inoculation and spread between the florets. The results provide evidence that Trichoderma can infect cereal plant heads under controlled conditions and produce a head blight-like type of disease. It remains to be investigated whether this disease also occurs naturally under field conditions.
Although, visual symptoms and colonization rates were higher after spray inoculation, point inoculation significantly reduced the thousand-grain weight in wheat and barley. The reduced kernel size may result from the premature ripening, apparently induced by shortage of water and food supply due to a cut-off of transport vessels in the point-inoculated florets as it is known from Fusarium head blight [19]. It is not known yet, how Trichoderma enters the florets and infects the interior floret tissue after spray inoculation and how this happens under natural field conditions. It can be assumed that Trichoderma, similar to Fusarium either directly penetrates the epidermal tissue of the inner palea and lemma, enters via natural crevices by hyphal growth, or uses the anthers as primary tissue for the initiation of colonization [19].
Trichoderma species live endophytically in soils of all ecosystems and different agroclimatic zones [23,24]. Their rapid growth, the ability to parasitize other fungi, their high adaptability and competitiveness have rendered them worldwide importance as microorganisms in biological plant protection and bio stimulant products [25,26,27]. They are widely used to control fungal diseases and promote plant growth in cereal crops like wheat and barley [16,28,29]. Whether the use of biological fungicides and biostimulants poses a risk to cereal crops cannot yet be determined, since the routes of natural infection of Trichoderma in the field have not yet been adequately explored. Those biocontrol products can be applied on soil as liquid or granulate, foliar sprayed per handhold, tractor or even through irrigation water. However, our results suggest that especially spray inoculation in cereal cultivation caused infections of Trichoderma in wheat and barley which resulted in high colonization rates and reduction of thousand kernel weight. We conclude that if spores of pathogenic Trichoderma species get dispersed on the spikes during flowering, the fungus can initially infect and spread in the cereal spikes.
Overall, the results show that cereals such as wheat and barley are potential hosts enabling or even enhancing survival and spread of Trichoderma in cereal crop rotations with maize. This risk situation is comparable to Fusarium ear rot, which can infect both maize and wheat via plant residues that represent a source of inoculum and greatly increase the risk of disease [20,30].
This is the first report describing T. afroharzianum as causal agent in wheat and barley leading to visible disease symptoms and reduction of thousand-kernel weight. The information provided in this work is important to better understand the epidemiology and disease cycle of T. afroharzianum within arable cropping systems comprising small grain cereals and maize. Hence, the development of management practices to reduce the amount of inoculum transmitted by crop residues appears a relevant measure also for Trichoderma epidemics. Further research is necessary to monitor whether pathogenic strains of Trichoderma from maize are in fact infecting wheat or barley under field conditions and to explore under which conditions such host leaps may happen.
Author Contributions
Conceptualization, A.P. ; methodology, P.G.; validation, A.P, and P.G..; investigation, P.G.; resources, A.P.; data curation, A.P.; writing—original draft preparation, A.P.; writing—review and editing, A.v.T.; visualization, A.P.; supervision, A.v.T.; project administration, A.P.; funding acquisition, A.P and A.v.T. All authors have read and agreed to the published version of the manuscript.”.
Funding
This research was funded by Federal Ministry for Food and Agriculture, grant number 2221NR014A.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Written informed consent has been obtained from the patient(s) to publish this paper.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Manuela Mücke for her excellent technical assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- FAOSTAT. Food And Agricultural Data; Food And Agriculture Organization Of The United Nation, 2021.
- Juroszek, P.; Tiedemann, A. Von. Climatic Changes And The Potential Future Importance Of Maize Diseases: A Short Review. Journal Of Plant Diseases And Protection 2013, 49–56.
- Nnadi, N.E.; Carter, D.A. Climate Change And The Emergence Of Fungal Pathogens. Plos Pathog. 2021, 17, E1009503. [Google Scholar] [CrossRef]
- Kubicek, C.P.; Komon-Zelazowska, M.; Druzhinina, I.S. Fungal Genus Hypocrea/Trichoderma: From Barcodes To Biodiversity. J. Zhejiang Univ. Sci. B 2008, 9, 753–763. [Google Scholar] [CrossRef] [PubMed]
- Jaklitsch, W.M.; Voglmayr, H. Biodiversity Of Trichoderma (Hypocreaceae) In Southern Europe And Macaronesia. Stud. Mycol. 2015, 80, 1–87. [Google Scholar] [CrossRef] [PubMed]
- Ghazanfar, M. U. , Raza, M., Raza, W., Qamar, M. I. Trichoderma As Potential Biocontrol Agent, Its Exploitation In Agriculture: A Review. Plant Protection 2018, 2. [Google Scholar]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Sharma, A. Trichoderma: The "Secrets" Of A Multitalented Biocontrol Agent. Plants (Basel) 2020, 9. [Google Scholar] [CrossRef]
- Ferreira, F.V.; Musumeci, M.A. Trichoderma As Biological Control Agent: Scope And Prospects To Improve Efficacy. World J. Microbiol. Biotechnol. 2021, 37, 90. [Google Scholar] [CrossRef]
- Bouanaka, H.; Bellil, I.; Harrat, W.; Boussaha, S.; Benbelkacem, A.; Khelifi, D. On The Biocontrol By Trichoderma Afroharzianum Against Fusarium Culmorum Responsible Of Fusarium Head Blight And Crown Rot Of Wheat In Algeria. Egypt J Biol Pest Control 2021, 31, 1–13. [Google Scholar] [CrossRef]
- Perelló, A.; Mónaco, C.; Simón, M.; Sisterna, M.; Bello, G. Biocontrol Efficacy Of Trichoderma Isolates For Tan Spot Of Wheat In Argentina. Crop Protection 2003, 22, 1099–1106. [Google Scholar] [CrossRef]
- El-Sharkawy, H.H.A.; Tohamey, S.; Khalil, A.A. Combined Effects Of Streptomyces Viridosporus And Trichoderma Harzianum On Controlling Wheat Leaf Rust Caused By Puccinia Triticina. Plant Pathology Journal (Faisalabad) 2015, 14, 182–188. [Google Scholar]
- El-Sharkawy, H.H.; Rashad, Y.M.; Ibrahim, S.A. Biocontrol Of Stem Rust Disease Of Wheat Using Arbuscular Mycorrhizal Fungi And Trichoderma Spp. Physiological And Molecular Plant Pathology 2018, 103, 84–91. [Google Scholar] [CrossRef]
- Hýsek, J.; Vach, M.; Brožová, J.; Sychrová, E.; Civínová, M.; Neděelník, J.; Hrubý, J. The Influence Of The Application Of Mineral Fertilizers With The Biopreparation Supresivit (Trichoderma Harzianum) On The Health And The Yield Of Different Crops. Archives Of Phytopathology And Plant Protection 2002, 35, 115–124. [Google Scholar] [CrossRef]
- Manzar, N.; Singh, Y.; Kashyap, A.S.; Sahu, P.K.; Rajawat, M.V.S.; Bhowmik, A.; Sharma, P.K.; Saxena, A.K. Biocontrol Potential Of Native Trichoderma Spp. Against Anthracnose Of Great Millet (Sorghum Bicolour L.) From Tarai And Hill Regions Of India. Biological Control 2021, 152, 104474. [Google Scholar] [CrossRef]
- Sharma, P.; Nath Patel, A.; Kumar Saini, M.; Deep, S. Field Demonstration Of Trichoderma Harzianum As A Plant Growth Promoter In Wheat (Triticum Aestivum L). JAS 2012, 4, 65–73. [Google Scholar] [CrossRef]
- Mahato, S.; Bhuju, S.; Shrestha, J. [Importieren] Effect Of Trichoderma Viride As Biofertilizer On Growth And Yield Of Wheat. Malays. J. Sustain. Agric. 2018, 2, 1–5. [Google Scholar] [CrossRef]
- Pfordt, A.; Schiwek, S.; Karlovsky, P.; Tiedemann, A. Von. Trichoderma Afroharzianum Ear Rot–A New Disease On Maize In Europe. Front. Agron. 2020, 2. [Google Scholar] [CrossRef]
- Sanna, M.; Pugliese, M.; Gullino, M.L.; Mezzalama, M. First Report Of Trichoderma Afroharzianum Causing Seed Rot On Maize In Italy. Plant Dis. 2022, PDIS12212697PDN. [Google Scholar] [CrossRef]
- Fusarium Head Blight Of Wheat And Barley; Leonard, K. J.; Bushnell, W.R., Ed.; APS Press: St. Paul, 2005; ISBN 089054302X. [Google Scholar]
- Dill-Macky, R.; Jones, R.K. The Effect Of Previous Crop Residues And Tillage On Fusarium Head Blight Of Wheat. Plant Dis. 2000, 84, 71–76. [Google Scholar] [CrossRef]
- Mesterhazy, A. Types And Components Of Resistance To Fusarium Head Blight Of Wheat. Plant Breeding 1995, 114, 377–386. [Google Scholar] [CrossRef]
- Miedaner, T. Breeding Wheat And Rye For Resistance To Fusarium Diseases. Plant Breeding 1997, 116, 201–220. [Google Scholar] [CrossRef]
- Röhrich, C.R.; Iversen, A.; Jaklitsch, W.M.; Voglmayr, H.; Vilcinskas, A.; Nielsen, K.F.; Thrane, U.; Döhren, H. Von; Brückner, H.; Degenkolb, T. Screening The Biosphere: The Fungicolous Fungus Trichoderma Phellinicola, A Prolific Source Of Hypophellins, New 17-, 18-, 19-, And 20-Residue Peptaibiotics. Chem. Biodivers. 2013, 10, 787–812. [Google Scholar] [CrossRef]
- Trichoderma: Host Pathogen Interactions And Applications; Sharma, A. K.; Sharma, P., Eds., 1st Ed. 2020; Springer Singapore; Imprint Springer: Singapore, 2020; ISBN 978-981-15-3321-1. [Google Scholar]
- Chaverri, P.; Branco-Rocha, F.; Jaklitsch, W.; Gazis, R.; Degenkolb, T.; Samuels, G.J. Systematics Of The Trichoderma Harzianum Species Complex And The Re-Identification Of Commercial Biocontrol Strains. Mycologia 2015, 107, 558–590. [Google Scholar] [CrossRef] [PubMed]
- Ghazanfar, M.U.; Raza, M.; Raza, W.; Qama, M.I. Trichoderma As Potential Biocontrol Agent, Its Exploitation In Agriculture, A Review. Plant Protection 2018, 103–135. [Google Scholar]
- Gupta, R.; Bar, M. Plant Immunity, Priming, And Systemic Resistance As Mechanisms For Trichoderma Spp. Biocontrol. In Trichoderma; Sharma, A.K., Sharma, P., Eds.; Springer Singapore: Singapore, 2020; pp. 81–110. ISBN 978-981-15-3320-4. [Google Scholar]
- Abdelzaher, H.M.A. Occurrence Of Damping-Off Of Wheat Caused By Pythium Diclinum Tokunaga In El-Minia, Egypt And Its Possible Control By Gliocladium Roseum And Trichoderma Harzianum. Archives Of Phytopathology And Plant Protection 2004, 37, 147–159. [Google Scholar] [CrossRef]
- Abo-Elnaga, H.I.G. Biological Control Of Damping Off And Root Rot Of Wheat And Sugar Beet With Trichoderma Harzianum. Plant Pathology Journal (Faisalabad) 2012, 11, 25–31. [Google Scholar] [CrossRef]
- Blandino, M.; Pilati, A.; Reyneri, A.; Scudellari, D. Effect Of Maize Crop Residue Density On Fusarium Head Blight And On Deoxynivalenol Contamination Of Common Wheat Grains. Cereal Research Communications 2010, 38, 550–559. [Google Scholar] [CrossRef]
Figure 2.
Disease symptoms in wheat at 21, 28, 35, 42 and 49 days post inoculation (A), close up of symptoms on wheat ears (B), lacking disease symptoms in inoculated sorghum (C), infected kernels of barley (D).
Figure 2.
Disease symptoms in wheat at 21, 28, 35, 42 and 49 days post inoculation (A), close up of symptoms on wheat ears (B), lacking disease symptoms in inoculated sorghum (C), infected kernels of barley (D).

Figure 3.
Disease severity at 7,14, 21, 28 and 35 days post inoculation (dpi) at winter wheat (left) and spring wheat (right) after spray and point inoculation with TriMix, CBS and water control.
Figure 3.
Disease severity at 7,14, 21, 28 and 35 days post inoculation (dpi) at winter wheat (left) and spring wheat (right) after spray and point inoculation with TriMix, CBS and water control.
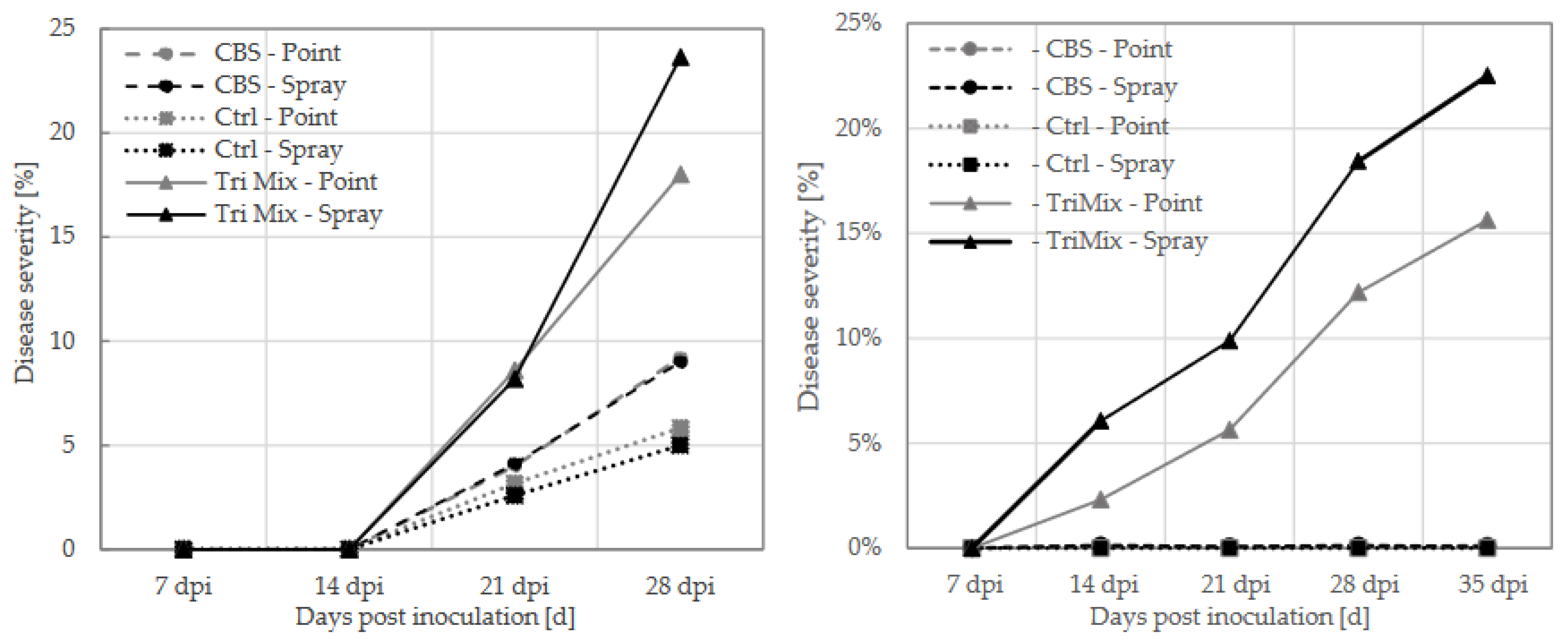
Figure 4.
Colonization rate (%) (left) and thousand kernel weight (g) (right) of winter and spring barley varieties after control (Ctrl), TriMix and CBS inoculation (means of both inoculation methods). Different letters indicate significant differences between treatments (Tukey-test, p ≤ 0.05). Error bar represent standard error.
Figure 4.
Colonization rate (%) (left) and thousand kernel weight (g) (right) of winter and spring barley varieties after control (Ctrl), TriMix and CBS inoculation (means of both inoculation methods). Different letters indicate significant differences between treatments (Tukey-test, p ≤ 0.05). Error bar represent standard error.
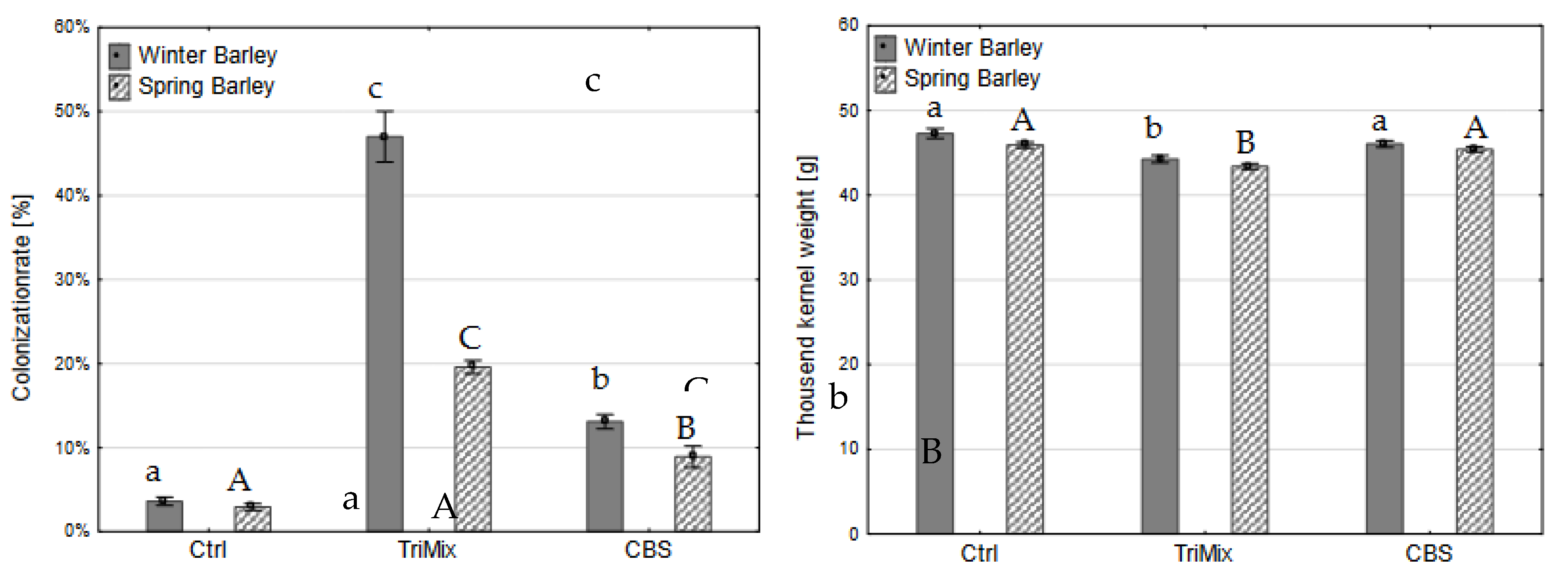
Figure 5.
Colonization rate (%) (left) and thousand kernel weight (g) (right) of winter and spring wheat varieties after control (Ctrl), TriMix and CBS inoculation (means of both inoculation methods). Different letters indicate significant differences between treatments (Tukey-test, p ≤ 0.05). Error bar represent standard error.
Figure 5.
Colonization rate (%) (left) and thousand kernel weight (g) (right) of winter and spring wheat varieties after control (Ctrl), TriMix and CBS inoculation (means of both inoculation methods). Different letters indicate significant differences between treatments (Tukey-test, p ≤ 0.05). Error bar represent standard error.
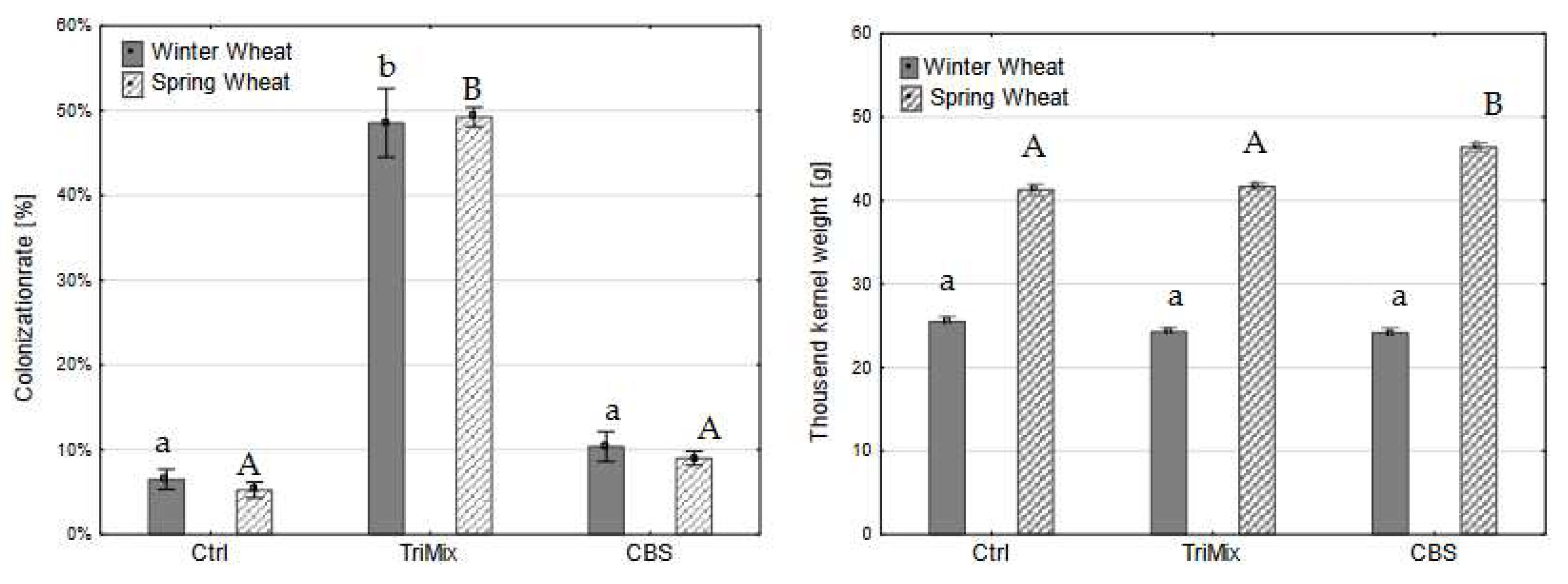
Figure 6.
Colonization rate (%) (left) and thousand kernel weight (g) (right) of silo type and grain type varieties sorghum after control (Ctrl), TriMix and CBS inoculation (means of both inoculation methods). Error bar represent standard error.
Figure 6.
Colonization rate (%) (left) and thousand kernel weight (g) (right) of silo type and grain type varieties sorghum after control (Ctrl), TriMix and CBS inoculation (means of both inoculation methods). Error bar represent standard error.
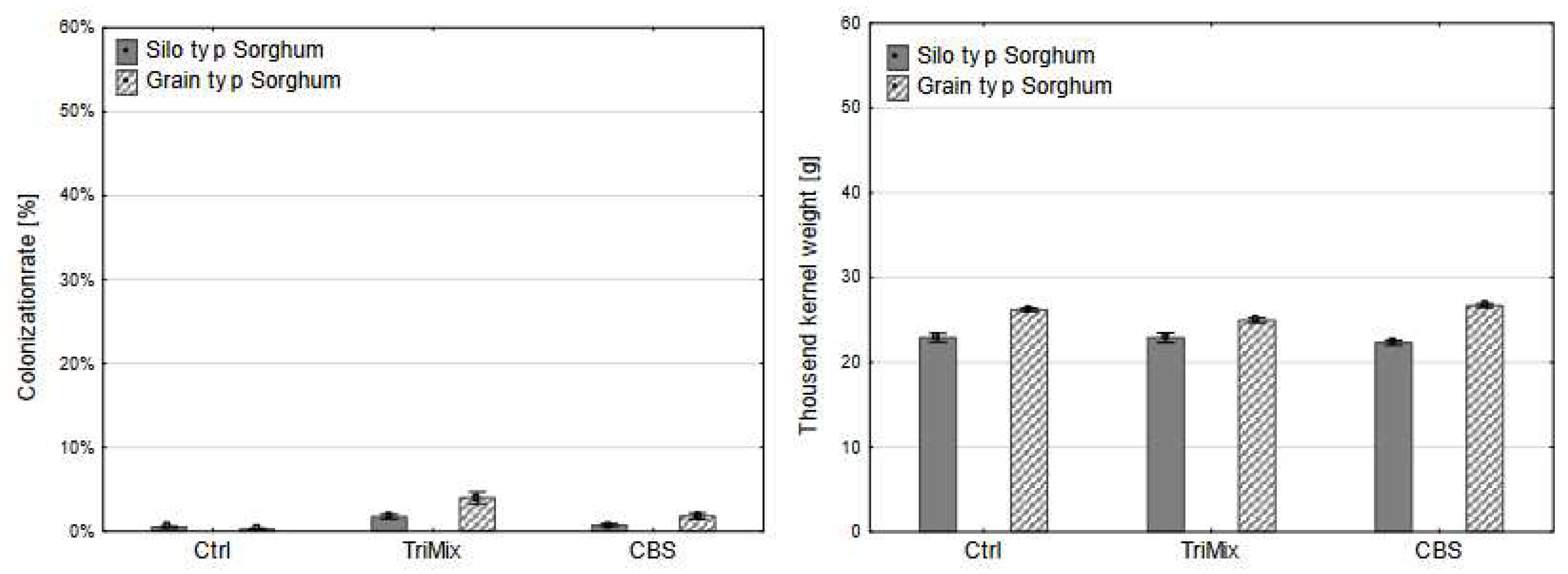
Figure 7.
Germination rate (%) of wheat, barley and sorghum after inoculation with TriMix and water (Control). Error bar represent standard error.
Figure 7.
Germination rate (%) of wheat, barley and sorghum after inoculation with TriMix and water (Control). Error bar represent standard error.
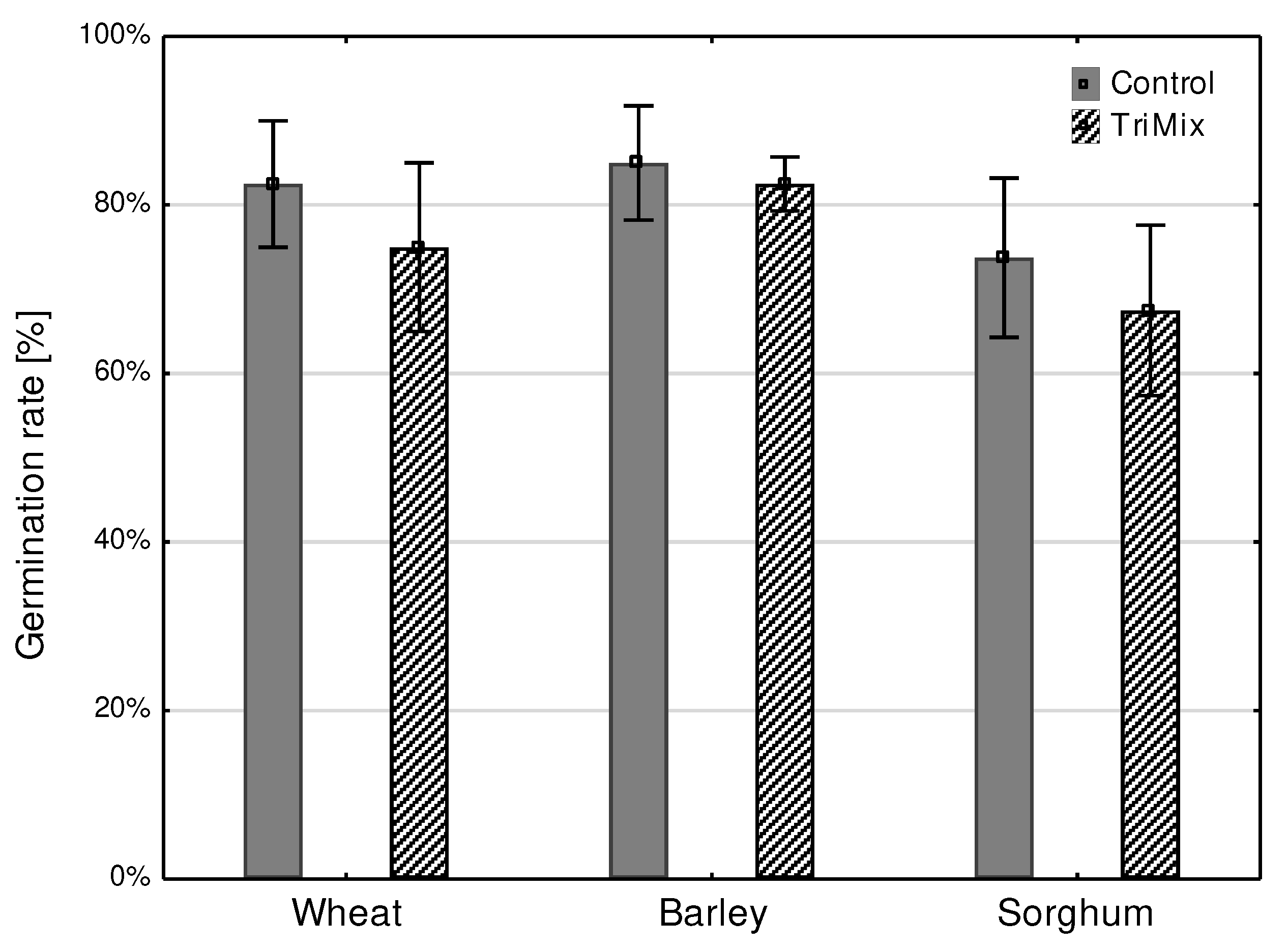

Table 1.
Origin of strains used in this study.
| Species | Isolat | Origin | Host |
|---|---|---|---|
| T. afroharzianum | CBS1246201 | type strain | |
| T. afroharzianum | TriMix | ||
| -Tri1 | Croix de Pardie | Maize | |
| - Tri2 | Künzing | Maize | |
| -Tri3 | Pocking | Maize |
1 Westerdijk Fungalbio Diversity Insitute, CBS, Utrecht, NL.
Table 2.
Colonization rate and thousand-kernel weight (TKW) for barley, wheat and sorghum at different varieties and inoculation methods.
Table 2.
Colonization rate and thousand-kernel weight (TKW) for barley, wheat and sorghum at different varieties and inoculation methods.
| Crop | Methode | Treatment | Colonization rate [%] | TKW[g] |
|---|---|---|---|---|
| Barley | Spray | TriMix | 36.4 a | 43.0 a |
| Control | 4.8 b | 47.9 b | ||
| Point | TriMix | 30.2 a | 44.7 a | |
| Control | 1.8 b | 45.3 a | ||
| Wheat | Spray | TriMix | 59.5 a | 33.1 a |
| Control | 4.0 b | 33.7 a | ||
| Point | TriMix | 38.4 a | 33.4 a | |
| Control | 7.7 b | 34.8 a | ||
| Sorghum | Spray | TriMix | 2.8 a | 22.3 a |
| Control | 0.5 a | 24.0 a | ||
| Point | TriMix | 3.0 a | 25.5 a | |
| Control | 0.3 a | 25.1 a |
Different letters indicate significant differences between TriMix and Control (α≤0.05, Tukey test).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



