Submitted:
21 June 2023
Posted:
25 June 2023
You are already at the latest version
Abstract
Salinity stress affect plant growth and development by causing osmotic stress, and nutrient imbalances through excess Na+, K+ and Cl- ion accumulations that induce toxic effects during germination, seedling development, vegetative growth, flowering, and fruit set. However, the effects of salt stress on such processes, especially in polyploidised leguminous plants remains unexplored and scantly reported compared to their diploid counterparts. This paper discusses the physiological and molecular response of legumes towards salinity stress based osmotic and ionic imbalances in plant cells. A multigenic response involving various compatible solutes, osmolytes, ROS, polyamines and antioxidant activity, together with genes encoding proteins involved in the signal transduction, regulation and response mechanisms to this stress were identified and discussed. This discussion reaffirms polyploidisation as the driving force in plant evolution and adaptation to environmental stress constraints such as drought, feverish temperatures, and in particular, salt stress. As a result, thorough physiological and molecular elucidation of the role of gene duplication through induced autopolyploidisation and possible mechanisms regulating salinity stress tolerance in grain legumes must be further studied.
Keywords:
Antioxidant activity
; autopolyploidy
; legumes
; salinity stress
; reactive oxygen species.
1. Introduction
Leguminous crops of the Fabaceae family such as chickpea (Cicer arietinum), cowpea (Vigna unguiculata), kidney bean (Phaseolus vulgaris), lentils (Lens culinaris), pea (Pisum sativum) and soybean (Glycine max L.) are considered the most valuable grain crops playing a key role in the diet of many people throughout the world. Grain legumes, also referred to as pulses, serve as excellent sources of essential proteins, dietary fibre, carbohydrates, mineral nutrients, and phytochemicals with very minimum-to-no fat content [1,2]. These grain crops have formed the basis of early farming culture systems, together with cereal crops centuries ago [3]. They are harvested as both dry seeds and vegetables used for human nutrition and livestock feeding. Apart from their nutritional aspect, species of the Leguminosae remain ecologically dominant across many vegetations [4], fixing atmospheric nitrogen (N) through an association with symbiotic rhizobial bacteria [5] and reducing the need for inorganic N fertilisers. Consequently, whole genome duplication events during the Cretaceous-Paleogene around 66 million years ago played a significant role in legume survival and diversification. This evolutionary process transformed legume wild taxa to forms which enabled sustainable human gains through domestication and numerous breeding programmes [3].
The domestication and production of legumes through agriculture was crucial in safeguarding food and nutritional security while enhancing environmental nitrogen cycle in terrestrial ecosystems [6,7]. Although, N-fixing microbials in association with nodulating plant species mostly grow under arid conditions as companion or dominant species, the degree in which these plants, particularly, polyploidised legumes exhibited tolerance to abiotic stress factors such as drought and salinity stress still requires further investigations. According to Rao et al. [8], the variations in gene composition and expression patterns found in homologous polyploid plants resulted in different beneficial physiological responses enhancing tolerance and optimum growth and yield under abiotic stress conditions. The study indicated that autotetraploids of Lycium ruthenicum revealed an increased abscisic acid (ABA) content of 78.4% under both normal and severe drought stress conditions compared with their diploid counterparts.
Nevertheless, molecular, and physiological variations between polyploids and diploids were mainly related to the biosynthesis and signal transduction pathways of ABA. This sesquiterpenoid hormone plays an important role in many cellular processes such as seed development, dormancy, germination, growth, and development of plants, as well as stress responses, and is subjected to complex regulations that involve the actions of a small family of 9-cisepoxycarotenoid dioxygenase (NCED) genes that indirectly produce xanthoxin, the main precursor of ABA [9]. The new challenge, however, is to understand the regulation of these biosynthetic genes and their pathways which predominantly lead to the stimulation of ABA biosynthesis and signal transduction during salinity stress.
Several studies have investigated the effects of abiotic stresses in various crop species of legumes, especially in diploid lines than polyploidised genotypes. As such, more studies must unravel how ABA induces the expression of osmotic proteins, especially in increasing stress tolerance of plants from genome to translational level as alluded by Rao et al. [8]. Under salinity stress, plants generally experience reduced growth, impaired ion or osmotic homeostasis and disruptions of metabolic processes subsequently leading to losses in crop growth and yields. This stress serves as one of the major limiting abiotic factors in agriculture, threatening food, and nutritional security. The adverse effects of this stress constraint in plant cells, organs and whole plant level take place during any stage of growth, including germination, seedling development, vegetative growth, and reproduction of plants [10]. Furthermore, salinity stress largely influences legume growth and yield by disturbing photosynthesis, hormonal regulation, nutritional imbalances, cause ion toxicity and osmotic stress effects. As reported by Farooq et al. [11], unravelling the response mechanisms and tolerance, as well as available management strategies to this abiotic stress in grain legumes may help in understanding the performance of these plants under salinity stress.
In this paper, the metabolic and genomic effects of salt stress, including tolerance mechanisms of polyploidised plants, in contrast with their diploid counterpart are discussed. This review analysed current knowledge on the application of induced autopolyploidisation to enhance plants’ adaptive traits against salinity through a multigenic response involving various compatible solutes, osmolytes, ROS, polyamines, and antioxidant activity, together with genes encoding proteins involved in the signal transduction, regulation, and response mechanisms. Altogether, the review examined whether synthetic polyploids of grain legumes exhibited enhanced response mechanisms to prevent toxic accumulation of Na+ and Cl- ion levels in the cell cytosol, through a more complete genomic and physiological understanding which remains a precondition for the development of new and improved salt-tolerant leguminous varieties.
2. Role of Autopolyploidy on Legume Response to Salinity Stress
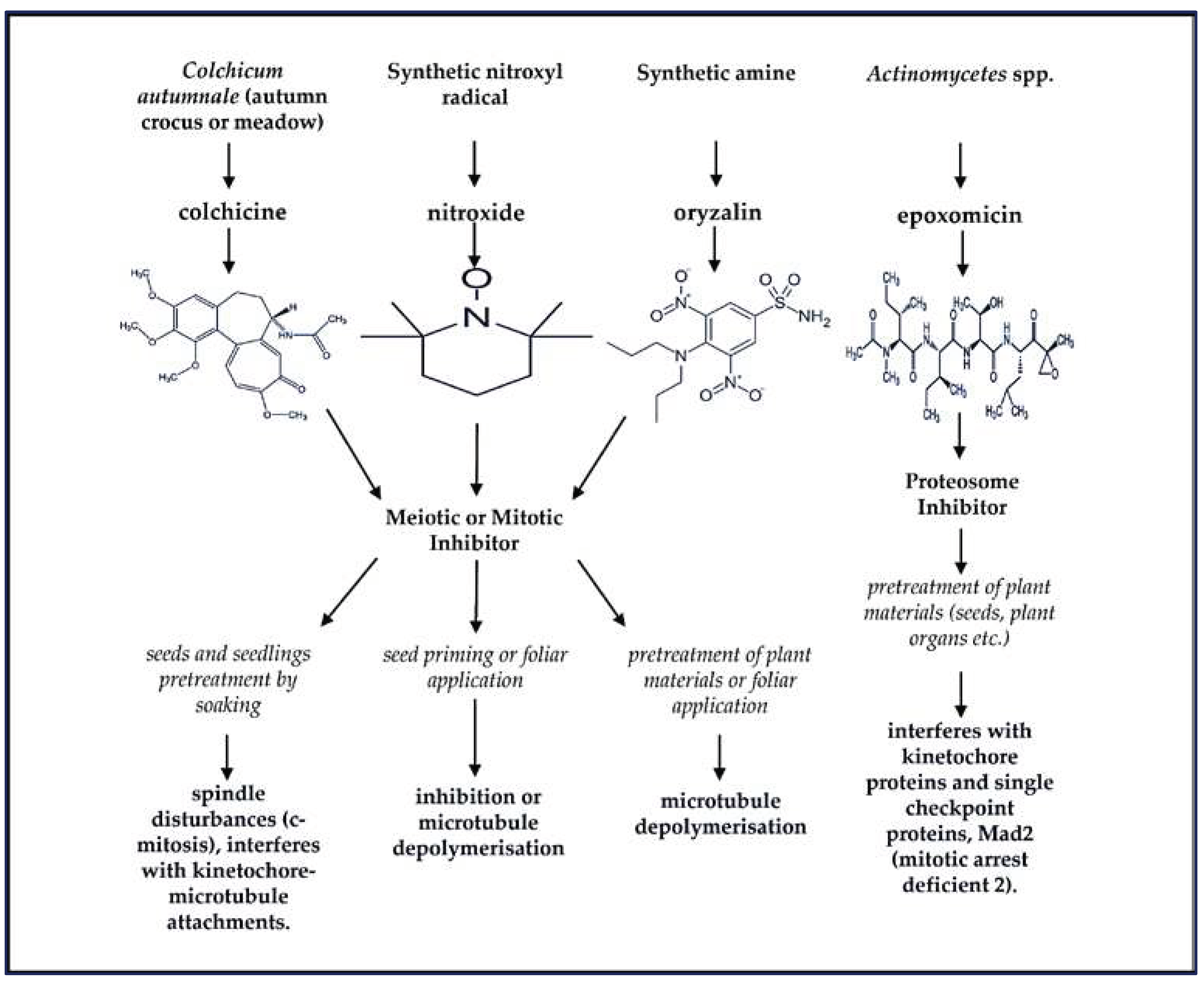
Plant ploidy level, which refers to the number of copies of the entire genome in a plant cell has implications for both physiological and molecular responses to both biotic and abiotic stresses. This process involves additional rounds of one or more genome duplications without cytokinesis to generate polyploidised mutant plants. The ploidy level can also be carried through into the germ line (gametes) to form multiple generations of polyploids [12]. Naturally, autopolyploidy or allopolyploidy can take place due to incomplete meiosis through gametogenesis and formation of unreduced chromosome number by microtubule inhibition within diploid cells [13]. Both meiotic and mitotic cell errors occur spontaneously in plants through variable frequencies depending on the species. This is a widespread phenomenon, occurring naturally and very gradually in flowering plants, but can also be rapidly synthetically induced using mutagenic chemicals such as colchicine, oryzalin, epoxomicin and nitroxide as described in Figure 1 [14,15,16].
These antimitotic agents are widely reported and comparatively evaluated in literature for their efficiency in inducing chromosome doubling through in vitro and in vivo regeneration of varying plant species [17,18,19,20,21,22,23]. As indicated on Figure 1, the influence of these chemicals on the formation of more than one haploid genomes in a single cell nucleus of plants occur as a result of the interference with kinetochore-microtubule attachments or what is widely known as microtubule depolymerisation which is extensively reported [18,19,23,24]. However, during the production of synthetic polyploids, the efficiency of genome replication depends on the concentration of the antimitotic agent and the duration of pretreatment, especially when mutagenic chemicals such as colchicine and oryzalin are applied for DNA duplication [18,19]. If successfully achieved, the marked effects of genome doubling could ameliorate unfavourable external pressures such as drought and salinity stress, including deterring pests as well as viral and bacterial induced diseases.
According to Corneillie et al. [24], morphometric analysis of polyploidised Arabidopsis plants revealed that their epidermal tissues contained increased cell sizes and reduced cell number per mesophyll compared to diploid plants. In line with these observations, the various polyploidised varieties of Arachis, Glycine wightii, Medicago sativa and Phaseolus aureus also demonstrated morphological similarities to Arabidopsis polyploids [25]. In view of the abovementioned reports, many studies presented the idea that polyploids offer adaptive advantages to environmental stress than the diploid progenitors even if the mechanism underlying stress resistance remains elusive. This somewhat holds significant truth since some of polyploids frequently showed vigorous growth, higher yield, and resistance under stressful conditions than their parent diploid species. Tetraploid plants of Medicago truncatula Gaertn and Medicago scutellate (L.) Mill. recorded up to 100% greater autumn biomass and 58% larger seed weights compared to their diploid parent lines under field conditions [26].
The M. truncatula Gaertn and M. scutellate (L.) Mill are well known to produce high protein forage content for livestock and a significant fix of atmospheric nitrogen, especially under rainfed farming systems without severe exposure to drought, heat, or salinity stress [27]. However, it is still not yet clear, especially because of complex regulatory mechanisms involved in plant development and response to environmental stresses, whether there is a common set of physiological and molecular strategies that polyploidised plants use to mitigate against abiotic stress. Some of the physiological changes that have been observed in polyploidised mutant plants compared to diploid lines as illustrated on Figure 2 include increased adaptability to environmental stress. Rao et al. [8] and Tossi et al. [28] reported that the positive impact on plant growth and net yield production of crops are due to the diversification of gene functions accompanying this evolutionary process. As expected, chromosome duplication would trigger major physiological changes that lead to notable resistance to stress, including the changing of the morphology of plant organs that confers stress tolerance.
Figure 1.
Chemical structures of naturally and synthetically produced mutagenic compounds that are used to achieve the multiplication of complete sets of chromosomes and production of random mutations in the genetic materials of plant species, giving rise to the birth of new varieties. These multiple sets of chromosomes and nucleotide alterations (particularly, substitution of guanine (G)- cytosine (C) to adenine (A) alkylation [G:C to A] induced by ethyl methane sulfonate), may take place in one nucleus, and can be stably inherited to progenies [12,14,16].
Figure 1.
Chemical structures of naturally and synthetically produced mutagenic compounds that are used to achieve the multiplication of complete sets of chromosomes and production of random mutations in the genetic materials of plant species, giving rise to the birth of new varieties. These multiple sets of chromosomes and nucleotide alterations (particularly, substitution of guanine (G)- cytosine (C) to adenine (A) alkylation [G:C to A] induced by ethyl methane sulfonate), may take place in one nucleus, and can be stably inherited to progenies [12,14,16].
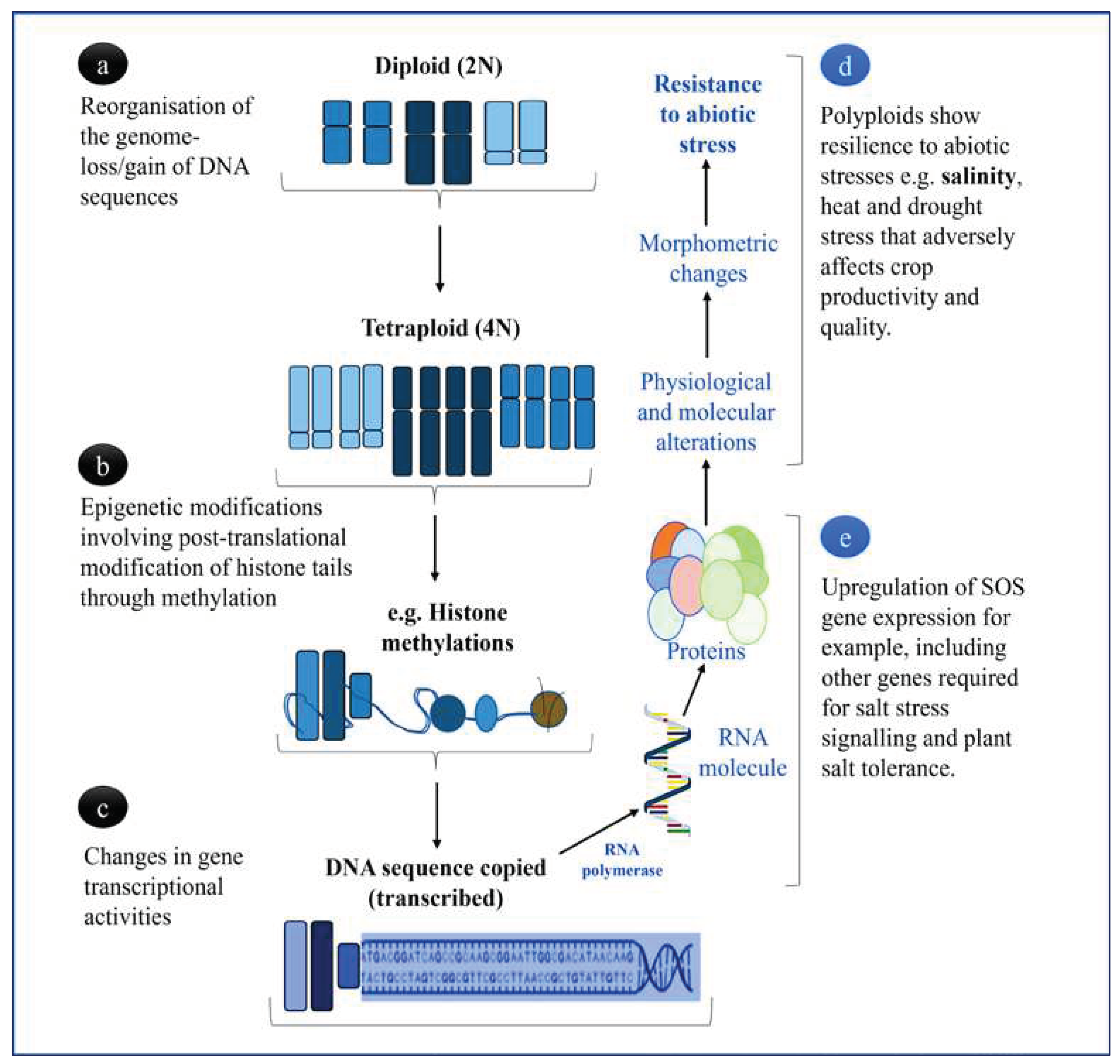
3. Physiological and Molecular Status of Leguminous Polyploids
In legumes, polyploidy serves as a major genetic driver in regulating growth and development processes, including the legume-rhizobium symbiosis which plays an important role on global nutrient cycle and diversification of plant species as previously mentioned [29]. Thus, the metaphase inhibition illustrated in Figure 2, which may be induced through efficient, reliable, low-cost, and safe pretreatments of plant materials such as cells, organs, or seeds with the different doses of mutagenic chemicals (also demonstrated on Figure 1) for chromosome doubling could result in altered gene transcriptional activities (Figure 2a–c). Accordingly, such interactions may have further consequences that include differential expression of common and distinct gene families depending on the strength and activity of promoter sequences. Some of these expressed genes may encode proteins that are responsible for significant notable physiological and phenotypic changes that enhance the resistance of leguminous plants to abiotic stresses such as salinity, drought, and high temperature. However, at this molecular level, salinity influences the expression of genes related to the biochemical parameters associated with salt tolerance. For instance, in non-leguminous species, such as Steria rebaudiana Bertoni (an important medicinal plant native to the northeastern Paraguay and southern Brazil), the overexpression of peroxidase encoding genes (POD4, POD6 and POD9) was reported [30].
The expression of these genes virtually influenced all metabolic reactions and physiological processes, specifically, the functional pathways, nutritional imbalances and hormonal regulations eventually getting all these impeded. Although, unlike the widely reported drought triggered effects on gene regulation and expression, the effects of salinity are more physiologically pronounced, involving major biochemical metabolite synthesis, enzyme induction and membrane transport than the upregulation and downregulation of plant genes. Razzaque et al. [31] reported allelic diversity at several genetic loci that have been found in genotypes that show tolerance to salt stress, influencing growth and nutritive value of leguminous crops. However, the marked physiological effects in polyploids because of genome doubling and altered gene functions revealed increased adaptability to environmental variability. Genome duplication may lead to the improvement and/or divergence of an entire duplicated pathway among others, resulting in noticeable changes on the physiological traits related to water use efficiency and photosynthesis. In Rao et al. [8], chlorophyll and hydrogen peroxide contents, together with abscisic acid (ABA) were used as physiological indicators to judge the response of polyploidised L. ruthenicum plants against environmental stress. Under reduced soil water content, which also led to osmotic stress from dehydration, the diploids showed inhibition of chlorophyll synthesis, and excessive increase in the concentration of hydrogen peroxide (H2O2). As indicated by Slesak et al. [32], H2O2 is increased during environmental stress, interacting with thiol-containing proteins, and activating the different signalling pathways, including transcriptional factors (TFs) involved in the regulation of gene expression and cell-cycle processes in plants.
Figure 2.
Summary of polyploidisation events of genome in plants and their role in abiotic stress tolerance enhanced by chromosome duplication. (a) Genome reorganisation facilitating increased allo- or auto- polyploidisation, either leading to extensive gene duplication or fractionation (loss) following polyploidy induction, (b) DNA modifications that regulate whether genes are turned on or off after polyploidisation, reprogramming gene expression networks, and (c) ploidy-related modifications on gene structure and function as a result of the depolymerisation of microtubule activities causing great alteration in the expression patterns of RNA-mediated polypeptides (d, e), which are also influenced by external stimuli such as environmental stress (d) [18,21,32].
Figure 2.
Summary of polyploidisation events of genome in plants and their role in abiotic stress tolerance enhanced by chromosome duplication. (a) Genome reorganisation facilitating increased allo- or auto- polyploidisation, either leading to extensive gene duplication or fractionation (loss) following polyploidy induction, (b) DNA modifications that regulate whether genes are turned on or off after polyploidisation, reprogramming gene expression networks, and (c) ploidy-related modifications on gene structure and function as a result of the depolymerisation of microtubule activities causing great alteration in the expression patterns of RNA-mediated polypeptides (d, e), which are also influenced by external stimuli such as environmental stress (d) [18,21,32].
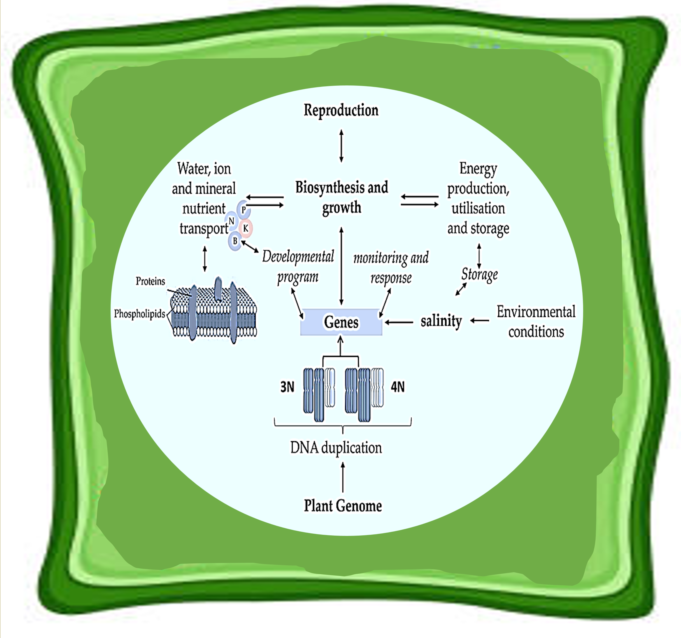
4. Ionic and Osmotic Stress Signalling Responses in Polyploidised Plants
To cope with salinity stress, plants have developed several adaptive strategies such as enhanced enzymatic and physiological reactions, as well as higher levels of metabolite expression such as phenols that are involved in antioxidant activities of the plant. This stress affects vital metabolic functions whereby Na+ and Cl- ions interferes with several nutrients such as those highlighted in Figure 3. Salt stress affects competitive nutrient uptake, accumulation, and transport in plants [33], consequently disturbing nutritional balance, hormone interactions, causing osmotic stress effects and specific ion toxicity [34]. In soybean, the ratio of gibberellin (GA) and abscisic acid (ABA) was negatively regulated by NaCl during seed germination and post-germinative growth [35]. This report further indicated that NaCl considerably down-regulated GA1, GA3 and GA4, while regulating the ABA content. As outlined in Figure 3, high soil salt content affect crops’ growth and development processes by influencing water availability in the cells, ion toxicity, oxidative stress, cell membrane organization, meiotic/ mitotic processes and causing nutritional disorders, among others.
In terms of inorganic ions, essential elements (N, P, K, S, Mg etc.) are considered intrinsic components of the structure and metabolism in plants [6,39]. The severe abnormalities that may arise due to salinity stress will result in symptoms of specific nutritional deficiencies such as chlorosis and necrosis. It remains common knowledge that salt stress influences the transfer of inorganic nutrients from the environment into a plant [40], but further insights are still required on how nonspecific osmotic stress and specific ion effects are regulated at both physiological and molecular level. Even though, the mechanisms through which legumes with altered ploidy adapted to salinity stress still need to be investigated, insights on other non-leguminous crops demonstrated novel traits and changes in existing physiological processes because of polyploidization.
As previous reported, polyploidised mutants always showed improved biosynthetic, carbolic, and signalling transduction pathways under environmental stress as indicated in Figure 2 and Figure 3. In the general scheme of signal transduction, polyploidised plants would likely show rapid environmental and developmental signal perception by specialised receptors, and effective activation of signalling cascade involving secondary messengers leading to an efficient response by the plant cells (Figure 3) [31,38]. Part of polyploids success is signalling between plants and microbes during symbiotic nitrogen fixation (SNF) which influences microbial infection of plant cell, cell division leading to nodulation, autoregulation of nodule development and intracellular accommodation of bacteria, metabolism and transport supporting SNF to prevent nodule senescence [41].
Salinity stress increases the accumulation of reactive oxygen species (ROS), however, total phenolic content, ascorbic acid content and antioxidant activity traits were enhanced in 320 tetraploids compared to 84 diploid genotypes of Solanum tuberosum L. [42]. According to the study revelations, the concentration of antioxidant compounds correlated positively with the skin tuber colour and ploidy levels. In Citrus limon (L.) Osbeck, Bhuvaneswari et al. [43] indicated that cyclic monoterpene, limonene, had significantly increased in tetraploids induced by 0.025% colchicine for 24 hours. Limonene is biosynthesised from the precussor geranyl disphosphate (GPP) by enzymatic biotransformation with d-or-l-limonene synthetase. The production of limonene monoterpenes is completed by methyl group deprotonation in the α-terpinyl cation, and this occurs in a wide variety of species including Conifers, Lamiales and Pinales, of which some are tetraploids [44,45].
Moreover, overexpression of terpenoid biosynthesis gene in soybean influenced the nodulation signalling pathways. The results indicated that the six terpenoid synthesis gene (SoTPS6, SoNEOD, SoLINS, SoSABS, SoGPS, and SoCINS) in soybean hairy roots increased nodule number, nodule root length and fresh root weight [46]. As reported in other studies, the expression of terpene in the roots was highly coordinated and cell specific. The abovementioned observations demonstrated the role of ploidy on this largest class of specialized metabolites found within different structures in plants. According to Pott et al. [47], monoterpenes like limonene plays a key role in plant’s interaction with the environment, especially in protecting plants against biotic and abiotic stresses.
Generally, polyploid plants often have improved traits than their diploid relatives and may have greater advantage in regulating both osmotic and ionic stress. The improved traits or maintained homeostasis plays an important role in many of the functions occurring in polyploidised plant cells. The accepted explanation of the influence of salinity stress is that imbalances in ion content in plant cells affect plant fitness, especially by affecting plant nutritional status, cell water potential and cause toxic effects through accumulation of ions, which disturb nutrient acquisition and result in cytotoxicity [48,49]. Under the stress, polyploidised plants show increased capacity to maintain unequal concentrations of sodium (Na+) and potassium (K+) ions both inside and outside the cells. Such improved responses occur due to tolerance genes and TFs regulating ion Na+/K+ pump and exclusion, through selective permeability of plasma membrane proteins (PMP), high sodium affinity transporter (HKT) and Na+/H+ exchangers that help to alleviate ion toxicity in the cells [50,51].
Although, information regarding polyploidised leguminous crops’ reaction to toxic ions is very scant, monocotyledous crops such as hexaploidy wheat spp. (Triticum aestivum genome BBAADD) demonstrated improved ion toxicity alleviation than both diploid counterparts and tetraploid wheat progenitors (T. turgidum) or durum wheat (T. durum). In polyploidised rice (HN2026-4x and Nipponbare-4x), Tu et al. [52] reported improved regulation of Na+ content and H+ proton content flux at root tips, with decreased Na+ and increased H+ efflux in the roots. In legume, the physiological mechanisms explaining the improvement of salt tolerance with increasing levels of ploidy remains under researched. However, genome duplication has been widely suggested to improve salinity stress resistance through enhanced proton transport pump and homeostasis in various monocot and dicot vegetable crop species [51,52,53].
5. Metabolic Changes of Legume Polyploids Exposed to Salinity Stress
Salinity stress limits crop growth and metabolism, posing major impacts on agricultural productivity. As previously mentioned, over-irrigation systems and drought stress continue to cause oversalinisation in many areas used for planting of agroeconomically important legume crops. Salinity stress also known as hyperionisation as indicated previously by Gupta and Huang [54] pose serious effects on metabolic and growth processes of plants through excessive accumulation of ions as previously indicated. Numerous reports have elaborated on NaCl effects on growth, mineral composition, proline content and antioxidant enzyme activity, which dramatically reduces the productivity of many crops. As highlighted in Figure 3, this negatively influences energy production, utilization and storage in plants generating ROS, reducing cell growths, assimilates production, and causing membrane dysfunctions [55], consequently disrupting growth, reproduction, and yield in many crop species. Hamayun et al. [56] reported significant decreases in endogenous levels of secondary metabolites, GA, jasmonic acid, salicylic acid (SA) and ABA due to salinity stress. However, abrupt changes in the osmotic and ionic state of the plant causes negative effects on crop’s metabolism and growth.
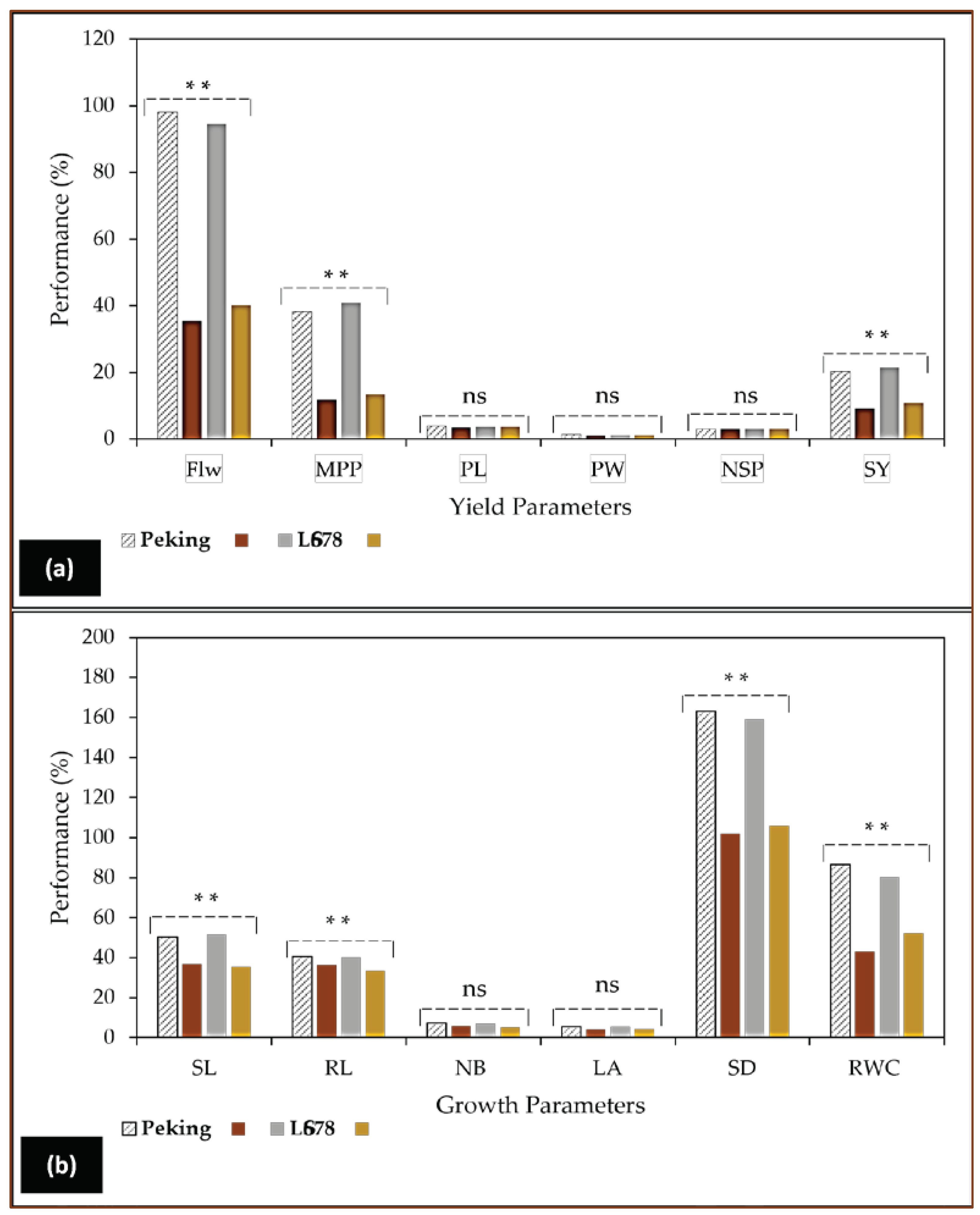
These depressive effects of NaCl were also demonstrated in a previous study conducted in our laboratory (Figure 4) where diploid plants showed inhibition to soybean, cultivar LS678 and Peking growth and yield characteristics [57]. The soybean plants were subjected to two levels of salinity stress. Similar findings were earlier reported by Amirjani [58] and Hamayun et al. [56] in soybean, cultivar Hwangkeumkong. The observations made in Figure 4a indicated that Peking and LS678 soybean plants exhibited noticeable morphological variations, particularly, in their root and stomatal density compared to the marginal but significant reductions observed in shoot length, number of branches and leaf area. The salt-stressed plants clearly indicated that yield components in both cultivars were decisively determined by the stress (Figure 5, a), especially, flowering and the number of pods produced per plant. As a consequence of the stress, lower flowering percentages and pod numbers were recorded for LS678 and Peking at 40.1%, 13.4 and 35.5%, 11.8, respectively [57]. In contrast, polyploidised plants gave improved results of 64.9% of flowering and 37.38% pods per plant using 0.8% and 0.6% EMS, respectively in mungbean (Vigna radiata L.) according to Roychowdhury et al. [59].
Additionally, the mean number of seeds per pod and 100-seed weight were increased in all the mutagenic treatments involving gamma rays and EMS in mungbean, urdbean varieties T-9 and Pant U-30 [60]. These mutation-based improvements conferred increased genetic diversity required to achieve economically important traits in both leguminous and non-leguminous crops wherein the selected mutant lines successfully contributed to diversity of the crop’s genetic base which is highly required for growth improvement against environmental stresses such as drought and salinity. Furthermore, the genetic variability induced through mutagenesis in treated populations also demonstrated some level of genotype dependency as reported by Laskar et al. [61] in lentil (Lens culinaris Medik). The genome structure in polyploids also have implications for both physiology and stress responses during vegetative and reproductive growths. As reports showed that polyploidy has occurred at least once in angiosperm lineages, the significantly altered phenotypes often influence the interactions of polyploids towards both biotic and abiotic stress constraints. According to Forrester and Ashman [29], plant-biotic interaction involving legume-rhizobia mutualism serve as an important interaction which regulates nutrient cycle, in addition to its indirect impact on vegetative (number of trifoliate leaves, leaf size, shoot height, root length, branch number etc.) and reproductive growth parameters (flower number, fruit pods, seed number and 100-seed weight) as indicated in Figure 4a,b.
Figure 4.
The reproductive yield (a) and vegetative growth (V) (b) changes caused by the increase in substrate salinity (NaCl) in soybean (Glycine max L. Merr.), cultivar LS678 and Peking [57]. Soybean plants were cultivated in 25 cm plastic pots containing pre-sterilised vermiculite, and then irrigated weekly with 250 mM NaCl from V2 until V3 stage under in vivo conditions. This study revealed that soybean plants subjected to salinity stress exhibited highly reduced growth [shoot length (SL), root length (RL), number of branches (NB), leaf area (LA), stomatal density (SD) and relative water content RWC)] and yield parameters [flower number (Flw), mean pods per plant (MPP), pod length (PL), pod weight (PW), number of seeds per pod (NSP) and seed yield (SY)], in addition to biochemical features such as photosynthetic pigments, carbohydrates, phenolic content, flavonoids, and antioxidant capacity as reported in this cited report [57]. .
Figure 4.
The reproductive yield (a) and vegetative growth (V) (b) changes caused by the increase in substrate salinity (NaCl) in soybean (Glycine max L. Merr.), cultivar LS678 and Peking [57]. Soybean plants were cultivated in 25 cm plastic pots containing pre-sterilised vermiculite, and then irrigated weekly with 250 mM NaCl from V2 until V3 stage under in vivo conditions. This study revealed that soybean plants subjected to salinity stress exhibited highly reduced growth [shoot length (SL), root length (RL), number of branches (NB), leaf area (LA), stomatal density (SD) and relative water content RWC)] and yield parameters [flower number (Flw), mean pods per plant (MPP), pod length (PL), pod weight (PW), number of seeds per pod (NSP) and seed yield (SY)], in addition to biochemical features such as photosynthetic pigments, carbohydrates, phenolic content, flavonoids, and antioxidant capacity as reported in this cited report [57]. .
Although, some researchers found that the notion that autopolyploids are larger than their diploid progenitors does not always hold true, such as the comparison of Arabidopsis and Musa plants, wherein obtained variations brought from haploidy to diploidy showed increases in growth parameters, but then decreased with the increase in ploidy levels in both species. Such findings were reported by Corneillie et al. [62] in A. thaliana and Brisibe and Ekanem [63] in Musa species. Quantitatively and qualitatively studying these differences in mutant plants can allow scientists to infer the functions of duplicated genes or map certain mutations on the chromosomes. Furthermore, such studies must continue selectively breeding crops to produce varieties that have higher yields, or varieties adapted to specific environmental conditions and are resistant to plant pathogens. As it is apparent from Figure 4, polyploidisation could substantially improve tolerance of legumes to salinity stress even though, this may result to losses of certain important characters. Leguminous crops such as soybean, chickpea, dry bean, cowpea, lentil, and peas are important sources of proteins and oils, so their potential improvement via ploidy duplication must rather be associated with the improvement of these useful economic traits.
3. Gene Expression in Polyploids Under Salinity Conditions
Although, detailed understanding of the molecular and physiological mechanisms explaining the improvement of salinity stress tolerance with increasing levels of ploidy in leguminous crops remain scant. Tossi et al. [28] reported the change in some leaf traits that favoured abiotic stress tolerance in Arachis duranensis x A. ipaensis synthetic allotetraploids. In urdbean (Vigna mungo (L.) Hepper) mutagenised genetic variability contributed to improved phenotypic traits in M2 and M3 generations [60]. Chao et al. [64] earlier reported improved salinity stress resistance in autopolyplodised Arabidopsis thaliana through enhanced potassium (K) accumulation in root and mesophyll tissues. The study revealed that ploidy level was a significant determinant of leaf K concentration attributed to the 57.2% variations observed in tetraploids constituting a 32% K-level increase in these plants compared to the diploid lines. Furthermore, the findings showed that autotetraploid accession Wa-1 (Warsaw, Poland) contributed additional alleles for increased leaf K with no obvious traits found in their diploid counterparts. According to Munns and Tester [65] increased K/Na ratio enhanced tolerance to osmotic and ionic components of salinity stress as similarly reported by Chao et al. [64].
Isayenkov and Maathuis [66] also reported the adaptation to adverse environmental salinity stress conditions in polyploidised plants driven by genes related to the efficient operation of NADPH-dependent ‘ROS-Ca2+ hub’. This multigenic family of calcium-dependent protein kinase encodes structurally conserved unimolecular calcium sensors or protein kinase effector proteins [67]. As reported in previous studies, Ca2+-dependent protein kinases diversity could be amplified by splicing and post-translational modifications [68], with a proposition that these protein kinases follow a common regulatory mechanism through phosphorylation [69] to confer tolerance to salinity stress. Hexaploidy lines of Ipomoea trifida reportedly exhibited increased K+ retention while excluding Na+, presenting highly reduced sensitivity of plasma membrane K+-permeability channels in both mature and elongation root zones of plants [70]. The above-mentioned ion channel activation occurs as a result of ROS-induced protein modification/ membrane-dependent NADPH oxidase system, regulated by the expression of respiratory burst oxidase homolog (Rboh) genes [71,72]. The retainment of high K+ levels and exclusion/ accumulation of low Na+ concentrations in the cytosol confers salinity stress tolerance in plants [70].
In many legumes, specific redox-dependent signalling pathways involving ROS, reactive nitrogen species (RNS) and reactive sulfur species (RSS) play a key role in acclimation and tolerance to environmental stress. According to Matamoros and Becana [73], ROS-induced modifications in legumes occur through oxidation, S-nitrosylasion, S-glutathionylation or persulfidation of cysteine thiol group, oxidation of methionine residue, nitration of tyrosine residue and lysine/arginine residue carbonylation. Molecular analysis of redox-based post translational modification of proteins revealed functional implications of these alterations on nodulation and whole plant growth and development under adverse conditions. Even though, polyploidy is a major genetic driver of ecological and evolutional diversity in plants, its effects on legume interactions remains partially explored on mutualistic relationship of legumes with microbes. Such studies, like that of Forrester and Ashman [25] reported that Medicago sativa subsp. caerulea autotetraploid plants produced larger nodules with larger nitrogen fixation zones than diploid plants using two strains of rhizobia, Sinorhizobium meiloti and Sinorhizobium medica. A focal microscopy was used to quantify the internal traits of nodule formation in both diploid and neotetraploid M. sativa plants.

Table 1.
Candidate genes conferring salinity stress tolerance in leguminous crops.
| Common name | Species name | Gene family/ name | Function | Reference |
|---|---|---|---|---|
| Peanut | Arachis hypogeal L. | ABSCISIC ACID INSENSITITVE 4S (AhAB14s) | Gene downregulation enhanced survival rate, biomass accumulation and root/shoot ratio of seedlings | Luo et al. [74] |
| Chickpea | Cicer arietinum | CaRab-GTP (CaRabA2, CaRab-B, CabRab-C, CabRab-D, Cab-Rab-E, and CabRab-H | Regulate Na+ accumulation in leaves | Sweetman et al. [38] |
| Pigeon pea | Cajanus cajan L. | GAPDH, UBC and HSP90 | Housekeeping genes carrying out cellular maintenance through stable expression irrespective of internal/ external signals | Sinha et al. [75] |
| Soybean | Glycine max L. Merrill. | GmSALT3 and GmSALT18 | Regulate Na+ accumulation in leaf mesophylls via root-based mechanisms | Guo et al. [76] |
| Birdsfoot trefoil | Lotus japonicus | LjLTP10 | Gene preventing osmotic stress by managing water deficit through structural architecture such as cuticular composition. | Tapia et al. [77] |
| Barrel medic | Medicago truncatula | MtDef4.3 | Defense gene against infectious microbes, including rhizobia species. | Gao et al. [78] |
| Lupin | Lipinus angustifolius | Aldehyde dehydrogenase (ALDH) | Catalyse irreversible oxidation of aldehyde molecules to non-toxic carboxylic acids | Jimencz-Lopez [79] |
| Common bean | Phaseolus vulgaris | PvDREB | Circumvent salinity stress by mechanic and osmotic adjustment | Konzen et al. [80] |
| Barrel medic | M. truncatula | Nodule-specific Cysteine Rich (NCR) | Control nodule organogenesis and gene duplication. | Guefrachi et al. [81] |
| Cowpea | Vigna unguiculata ssp. sesquipedalis (asparagus bean) | Root-derived DEGs (GmsSOS1, SOS2 and OsNHX2) | Genes involved in redox reactions, enhancing antioxidant enzymes activity and reduces Na+ accumulation under salt stress | Pan et al. [82] |
| Mung bean | Vigna radiata L. Wilczek | VrKUP, VrHAK and VrKT | Regulate influx and efflux of K+ through K+ transporters and channels | Azzam et al. [3], Ragel et al. [83] |
| Red clover | Trifolium spp. (T. pratense L.) | NHX1 | Na+/H+ antiporter in plants, predominantly, Trifolium spp. | Delormel [84] |
| Pea | Pisum sativum | PsRPL3OE, PsChla/bBP and PsFDH | Regulate protein synthesis, photosynthesis, and long chain lipid synthesis in the mesophylls | Joshi et al. [85] |
| Wild peanut | Arachis duranensis | AdWRKYs (AdWRKY18, AdWRKY40, AdWRKY42, AdWRKY56 and AdWRKY64) | Genes regulating ABA accumulation, stomatal aperture, and root development | Zhang et al. [86] |
There was a strong direct effect on nodulation and nodule traits such as N-fixation zone, nodule area and bacteriods containing symbiosomes due to increased ploidy level. These nodule features and other polyploidy-induced unique characteristics involving activation of nodule-specific transcriptome, node genes within mobile plasmids of rhizobial genomes and ample of nodule-specific cysteine-rich (NRC) genes (Table 1). NCR gene family is extensively large in Medicago spp. with about 600 of other genes [81,87]. The wider availability of such genome sequence information of Medicago and several other legume species will boost genomic research and breeding for yield improvement and adaptation of plants to adverse conditions. RNA sequencing transcriptome analyses in polyploids provided information into the transcriptional mechanisms, molecular mediation of various cellular processes, genome reprogramming and gene function as indicated in Table 1. As far as a decade ago, transcriptome analysis has been performed in several diploid legumes such as Glycine max (L.) Merrill, M. truncatula Guertn, Lotus corniculatus L. var. japonicus Ragel and Cicer arietinum L. to unravel the overall and specific transcriptional activities of genes in both diploid and polyploidised lines [88]. In shoots and roots of M. truncatula, transcriptional responses to treatment with rhizobia detected varied gene expression patterns shifts under different conditions [78].
Some of the genes, like leginsulin, defensins, root transporters, nodule-related genes and circadian clock genes are widely explored across legume species in relation to biological symbiotic nodulation system (Table 1) [89,90,91]. Currently, sufficient information is available in legumes that is required to elucidate responsible genes and cellular processes involved in plant-microbe symbiosis, however, many genes in the nodulation, plant growth and response to abiotic stress such as salt and drought stress still need to be investigated. Furthermore, not all genes are upregulated during symbiosis or the period of stress. Genes such as MtDef4.3 (Table 1) of plant defensin genes in M. truncatula get downregulated under rhizobia-induced nodulation for N-fixation [78]. Most of these expressed genes contribute to the morphology, physiology, and developmental attributes, also providing housekeeping functions necessary to normalize expression, regulation, and response of plants to environmental instabilities.
The WRKY genes encoding WRKY proteins containing conserved sequence WRKYGQK heptapeptide at the N-terminal end have also been widely implicated in leguminous and non-leguminous crop tolerance to biotic and abiotic stress. Although, WRKY evolution in legumes is still unknown, Song et al. [92] reported that this family comprise a class of TFs involved in physiological changes that enhances plant responses under abiotic stress such as drought and salinity stress constraints. As alluded by various other researchers, natural and induced polyploidisation via mutagenesis can unleash new alleles of WRKY genes and others that control traits required for salinity stress tolerance.
4. Discussion and Prospects
Salinity stress has caused adverse negative impacts on plants for decades, limiting growth, reproduction, and survival. The kind of pressures inflicted by this stress during plant growth and development include osmotic stress, ion toxicity and reduced cell water potential, that constitutes water deficit stress. According to Zhao et al. [93], salt stress severely affects plants throughout their life cycle, hindering seed germination, growth and development, flowering, and fruiting. In addition to its toxicity, salt stress limit nutrient uptake by roots. Stress-induced Na+ ions also function as signalling molecules of this abiotic stress and drought, especially after perception by non-selective cation channels [31]. Non-selective cation channels are multi protein complexes found within cellular membranes and they give high permeability to ions, mainly Na+, K+, Ca2+ and Cl- through polarisation of the plasma membranes [94]. In cowpea, Na+, K+, K/Na ratio, seedling heights and chlorophyll were significantly influenced by salt stress [95]. Mungbean plants also exposed to saline conditions exhibited reduced growth and yield, unable to take up sufficient water required for metabolic processes and failed to maintain turgidity due to low osmotic potential in the cells [96]. Other leguminous crops such as soybean, lupin, pea, peanut, and pigeon pea are also seriously affected by salinity stress, however, their intrinsic tolerance mechanism and agro-physiological characteristics still need to be explored.
As alluded by HanumanthaRao et al. [96], salinity tolerance comprises multifaceted responses at the molecular and physiological levels, but much remains to be studied from the natural and adaptive tolerance to this stress in both diploid and polyploidised leguminous plants. It is evident that ploidy level particularly, in leguminous crops that are widely studied/ serving as industrial crops and essential to ensure food security is poorly correlated with salinity stress and any other abiotic stress resistance. This poor correlation may be due to the overwhelming number of duplicated sets of genes that are involved at different developmental stages in coping with salt stress. It is, therefore, important to comparatively evaluate diploids and polyploids stress tolerance traits and further explore these traits in breeding programs to obtain new lines for durable tolerance. As HanumanthaRao et al. [96] reported, tolerance in salinity stress is achievable in legumes via the overexpression of housekeeping differentially expressed genes such as SOS1 involved in the exclusion of toxic Na+ ions by plasma membrane Na+/H+ antiporter, sequestration of Na+ into the vacuoles to reduce ion toxicity and encoding compatible osmolytes to protect cell proteins against salinity-induced inactivation and denaturation. Such osmolytes, predominantly amino acids, amino acid derivatives, sugars and polyols as reported by Kurz [97] are synthesised or taken up by plants for adaptation purposes under extreme environmental conditions.
Generally, during abiotic stress, osmolytes encoded by the raffinose family oligosaccharides (RFO) genes produce enzymes that convert sucrose into RFOs which establish cell stability and offer protection against abiotic stress such as salinity, drought, and cold stress [98]. These candidate RFO genes were quantitatively analysed in Phaseolus vulgaris and Glycine max by the above report, revealing that reduction of RFO content in the seeds could improve the nutritional quality of these beans without compromising normal plant developmental and functional characteristics, especially under abiotic stress conditions. The abovementioned intrinsic mechanism of stress response in plants exhibit enhanced stress tolerance in polyploid plants relative to their diploid counterparts. Wei et al. [99] reported that autotetraploid trifoliate orange (Poncirus trifoliata (L.) Raf.) accumulated higher levels of soluble osmolyte sugars and proline under salt stress compared to the diploids. In addition, polyploids exhibited enhanced salt tolerance by displaying specific enrichment of DEG genes with varying gene expression patterns in both roots and shoots. As indicated in many other studies, genes related to hormone signal transduction, starch and sucrose metabolism, proline biosynthesis, antioxidant enzyme activity, and several specific genes encoding transcriptional factors get over expressed in polyploid plants [20,32,43,72,100].
5. Conclusion
Studying the response of leguminous crops under different stresses and environments at the physiological and molecular level and comparing the gathered information between diploids and polyploids will enable us to understand and measure the beneficial activities of majority of genes found in these plants. This will also provide further insights on gene regulation under different biological contexts, identify genes that are important for the various plant growth and developmental processes, and strengthen in-depth investigations and physio-molecular understanding underlying salinity stress tolerance in leguminous crops. Understanding of the physiological and molecular responses of polyploidy induced tolerance to abiotic constraints such as salinity and drought stress could provide correlative insights into the legume biology and enhance our understanding and acceleration towards the establishment of efficacious breeding approaches. As succinctly reviewed in this paper, polyploidy remains the driving force in the evolutionary adaptation and success of leguminous crops, thus artificially induced autopolyploidy using chemical agents such as colchicine and oryzalin generated anatomical, morphological, physiological, and molecular changes increase tolerance to abiotic and biotic stresses, which positively impact of plant growth and yield productivity.
Author Contributions
Conceptualization, M.P.; writing—original draft preparation, M.P.; writing—review and editing, M.P.; visualization, M.P. The author has read and agreed to the published version of the manuscript.
Funding
This research received no external funding. The APC was funded by DEPARTMENT OF RESEARCH, DEVELOPMENT AND ADMINISTRATION, UNIVERSITY OF LIMPOPO.
Acknowledgments
The author thanks the Department of Biodiversity and Faculty of Science and Agriculture at the University of Limpopo.
Conflicts of Interest
The authors declare no conflict of interest.
References
- de Jager, I.; Borgonjen-van den Berg, K.J.; Giller, K.E.; Brouwer, I.D. Current and potential role of grain legumes on protein and micronutrient adequacy of the diet of rural Ghanaian infants and young children: using linear programming. Nutr J. 2019, 18, 12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Mascher, M.; Abbo, S.; Jayakodi, M. Advancing grain legumes domestication and evolution studies with genomics. Plant Cell Physiol 2022, 63, 1540–1553. [Google Scholar] [CrossRef] [PubMed]
- Koenen, E.J.M.; Ojeda, D.I.; Blakker, F.T.; Wieringa, J.J.; Kidner, C.; Hardy, O.J.; Pennington, R.T.; Herendeen, P.S.; Bruneau, A.; Hughes, C.E. The origin of the legumes is a complex paleopolyploid phylogenomic tangle closely associated with the Cretaceous-Paleogene (K-Pg) mass extinction event. Syst Biol 2021, 70, 508–526. [Google Scholar] [CrossRef] [PubMed]
- Wagner, T.C.; Richter, J.; Jobert, D.F.; Ficher, C. A dominance shift in arid savanna: An herbaceous legume outcompetes local C4 grasses. Ecol Evol 2018, 8, 6729–6787. [Google Scholar] [CrossRef] [PubMed]
- Xu-Ri, Dai, D.; Xu, X. The symbiotic nitrogen fixation by legumes in a legume-companion and a legume-dominant alpine steppe on the central Tibetan plateau. Ecol Res 2021, 36, 545–555. [CrossRef]
- Zhang, P.; Dumroese, R.K.; Pinto, J.R. Organic or inorganic nitrogen and rhizobia inoculation provide synergistic growth response of a leguminous forb and tree. Front Plant Sci 2019, 10, 1308. [Google Scholar] [CrossRef]
- Ullah, A.; Bano, A.; Khan, N. Climate change and salinity effects on crops and chemical communication between plants and plant growth-promoting microorganisms under stress. Front. Sustain. Food Syst. 2021, 5, 618092. [Google Scholar] [CrossRef]
- Rao, S.; Tian, Y.; Xia, X.; Li, Y.; Chen, J. Chromosome doubling mediate superior drought tolerance in Lycium ruthenicum via abscisic acid signaling. Hortic Res 2020, 7, 40. [Google Scholar] [CrossRef]
- Xiong, L.; Zhu, J.-K. Regulation of abscisic acid biosynthesis. Plant Physiol 2003, 133, 29–36. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, Y. Unravelling salt stress signalling in plants. J Int Plant Biol 2018, 60, 796–804. [Google Scholar] [CrossRef]
- Farooq, M.; Gogoi, N.; Hassain, M.; Barthakar, S.; Paul, S.; Bharadwaj, N. ’ Migdadi, H.M.; Alghamdi, S.S.; Siddique, K.H.M. Effects, tolerance mechanisms and management of salt stress in grain legumes. Plant Physiol Biochem 2017, 118, 199–217. [Google Scholar] [CrossRef]
- Pelé, A.; Rousseau-Gueutin, M.; Chèvre, A.-M. Speciation success of polyploid plants closely relates to the regulation of meiotic recombination. Front. Plant Sci. 2018, 9, 907. [Google Scholar] [CrossRef]
- Farhadi, N.; Panahandeh, J.; Motallebi-Azar, A.; Molchtarzadeh, S. Production of autotetaploid plants by in vitro chromosome engineering in Allium hirtifolium. Hort Plant J In Press. 2022, 1–13. [Google Scholar] [CrossRef]
- Meng, L.; Mohan, R.; Kwok, B.A.B.; Elofsson, M.; Sin, N.; Crews, C.M. Epoxomicin, a potent and selective proteasome inhibitor, exhibits in vivo anti-inflammatory activity. Proc Natl Acad Sci 1999, 96, 10403–10408. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, S.; Akutsu, M.; Okazaki, K. Mechanisms of action of nitrous oxide gas applied as a polyplodizing agent during meiosis in lilies. Sex Plant Reprod 2008, 22, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Chaparro, A.F.; Halder, S.; Trader, D.J. Synthesis and application of a clickable epoxomicin-based probe for proteasome activity analysis. Curr Prot 2022, 2, e490. [Google Scholar] [CrossRef]
- Sbrana, I.; Sibio, A.D.; Lomi, A.; Scarcell, V. C-mitosis and numerical chromosome aberration analyses in human lymphocytes: 10 known or suspected spindle poisons. Mutat Res-Fund Mol M. 1993, 287, 57–70. [Google Scholar] [CrossRef]
- Giani, S.; Campanoni, P.; Breviario, D. A dual effect on protein synthesis and degradation modulates the tubulin level in rice cells treated with oryzalin. Planta 2002, 214, 837–847. [Google Scholar] [CrossRef] [PubMed]
- Manzoor, A.; Ahmad, T.; Bashir, M.A.; Hafiz, I.A.; Silvestri, C. Studies on colchicine induced chromosome doubling for enhancement of quality traits in ornamental plants. Plants 2019, 8, 194. [Google Scholar] [CrossRef]
- Madani, H.; Escrich, A.; Hossani, B.; Sanchez-Munoz, R.; Khojasteh, A.P. Effect of polyploidy induction on natural metabolite production in medicinal plants. Biomolecules 2021, 11, 899. [Google Scholar] [CrossRef]
- Kozgunova, E.; Nishina, M.; Goshima, G. Kinetochore protein depletion underlies cytokinesis failure and somatic polyploidization to the moss Physcomitrella patens. eLife 2019, 8, e43652. [Google Scholar] [CrossRef] [PubMed]
- Tahjib-Ul-Arif, M.; Wei, X.; Jahan, I.; Hasanuzzaman, M.; Sabuj, Z.H.; Zulfiqar, F.; Chen, J.; Iqbal, R.; Dastogeer, K.M.G.; Sohag, A.A.M.; Tonny, S.H.; Hamid, I.; Al-Ashkar, I.; Mirzapour, M.; El Sabagh, A.; Murata, Y. Exogenous nitric oxide promotes salinity tolerance in plants: A meta-analysis. Front Plant Sci 2022, 13, 957735. [Google Scholar] [CrossRef] [PubMed]
- Fatima, A.; Husain, T.; Suhel, M.; Prasad, S.M.; Singh, V.P. Implication of nitric oxide under salinity stress: The possible interaction with other signaling molecules. J Plant Growth Regul. 2022, 41, 163–177. [Google Scholar] [CrossRef]
- Corneillie, S.; Storme, N.D.; Acker, R.; Fangel, J.U.; Bruyne, M.; Rycke, R.; Geelen, D.; Willats, W.G.T.; Vauholme, B.; Booerjan, W. Polyploidy affects plant growth and alters cell wall composition. Plant Physiol 2019, 179, 74–87. [Google Scholar] [CrossRef] [PubMed]
- Forester, N.J.; Ashman, T-L. The direct effects of plant polyploidy on the legume-rhizobia mutualism. Ann Bot 2018, 121, 209–220. [Google Scholar] [CrossRef]
- Innes, L.A.; Denton, M.D.; Dundas, I.S.; Peck, D.M.; Humphries, A.W. The effect of ploidy number on vigor, productivity, and potential adaptation to climate change in annual Medicago species. Crop Sci. 2021, 61, 89–103. [Google Scholar] [CrossRef]
- Kanatas, P.; Gozoulis, I.; Travlos, I. Irrigation timing as a practice of effective weed management in established alfalfa (Medicago sativa L.) crop. Agron 2021, 11, 550. [Google Scholar] [CrossRef]
- Tossi, V.E.; Martínez Tosar, L.J.; Laino, L.E.; Iannicelli, J.; Regalado, J.J.; Escandón, A.S.; Baroli, I.; Causin, H.F.; Pitta-Álvarez, S.I. Impact of polyploidy on plant tolerance to abiotic and biotic stresses. Front Plant Sci 2022, 13, 869423. [Google Scholar] [CrossRef]
- Forrester, N.J.; Ashman, T.-L. Autopolyploidy alters nodule-level interactions in the legume-rhizobium mutualism. Am J Bot 2020, 107, 179–185. [Google Scholar] [CrossRef]
- Azzam, C.R.; Al-Taweel, S.K.; Abdel-Aziz, R.M.; Rabea, K.M.; Abou-Sreea, A.I.B.; Rady, M.M.; Ali, E.F. Salinity effects on gene expression, morphological and physio-biochemical responses of Stevia rebaudiana Bertoni in vitro. Plants 2021, 10, 820. [Google Scholar] [CrossRef]
- Razzaque, S.; Elias, S.M.; Haque, T.; Biswas, S.; Jewel, G.M.N.A.; Rahman, S.; Weng, X.; Ismail, A.M.; Juenger, T.E.; Seraj, Z.I. Gene expression analysis associated with salt stress in reciprocally crossed rice population. Sci Rep 2019, 9, 8249. [Google Scholar] [CrossRef] [PubMed]
- Slesak, I.; Libilc, M.; Karpinska, B.; Karpinski, S.; Miszalski, Z. The role of hydrogen peroxide in regulation of plant metabolism and cellular signalling in response to environmental stresses. Acta Biochim Pol 2007, 54, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Nadeen, M.; Li, J.; Yahya, M.; Wang, M.; Ali, A.; Cheng, A.; Wang, X.; Ma, C. Grain legumes and fear of salt stress. Focus on mechanisms and management strategies. Int J Mol Sci 2019, 20, 799. [Google Scholar] [CrossRef]
- He, Y.; Fu, J.; Yu, C.; Wang, X.; Jiang, Q.; Hong, J.; Lu, K.; Xue, G.; Yan, C.; James, A.; Xu, L.; Chen, J.; Jiang, D. Increasing cyclic electron flow is related to Na+ sequestration into vacuoles for salt tolerance in soybean. J. Exp. Bot. 2015, 66, 6877–6889. [Google Scholar] [CrossRef]
- Shu, K.; Qi, Y.; Chen, F.; Meng, Y.; Luo, X.; Shuai, H.; Zhou, W.; Ding, J.; Du, J.; Liu, J.; Yang, F.; Wang, Q.; Liu, W.; Yong, T.; Wang, X.; Feng, Y.; Yang, W. Salt stress represses soybean seed germination by negatively regulating GA biosynthesis while positively mediating ABA biosynthesis. Front Plant Sci 2017, 8, 1372. [Google Scholar] [CrossRef]
- Kang, Z.; Xiaowu, W.; Feng, C. Plant polyploidy: Origin, evolution, and its influence on crop domestication. Hort Plant J 2019, 5, 237–239. [Google Scholar] [CrossRef]
- Bouyer, D.; Geier, F.; Kragler, F.; Schnittger, A.; Pesch, M.; Wester, K.; Balkundle, R.; Timner, J.; Fleck, C.; Hulskamp, M. Two-dimensional patterning by a trapping depletion mechanism. The role of TTG1 and GL3 in Arabidopsis trichome formation. PLoS Biol 2008, 6, e141. [Google Scholar] [CrossRef] [PubMed]
- Sweetman, C.; Khassanova, G.; Miller, T.K.; Booth, N.J.; Kurishbayev, A.; Jatayev, S.; Gupta, N.K.; Langridge, P.; Jenkins, C.L.D.; Soole, K.L.; Day, D.A.; Shavrukov, Y. Salt-induced expression of intracellular vesicle trafficking genes, CaRab-GTP, and their association with Na+ accumulation in leaves of chickpea (Cicer arietinum L.). BMC Plant Biol 2020, 20 (Suppl. 1), 183. [Google Scholar] [CrossRef] [PubMed]
- Kocira, A.; Staniak, M.; Tomaszewska, M.; Koprnas, R.; Cymerman, J.; Panasiewicz, K.; Lipinska, H. Legume cover crops as one of the elements of strategic weed management and soil quality improvement. A review. Agric 2020, 10, 394. [Google Scholar] [CrossRef]
- Magadlela, A.; Perz-Fernandez, A.; Kleinert, A.; Dreyer, L.L.; Valentine, A.J. Source of inorganic N affects the cost of growth in a legume tree species (Virginia divaricate) from the Mediterrean-type Fynbos ecosystem. J Plant Ecol 2016, 9, 752–761. [Google Scholar] [CrossRef]
- Roy, S.; Liu, W.; Nandety, R.S.; Crook, A.; Mysore, K.S.; Pislariu, C.I.; Frugoli, J.; Dickstein, R.; Udvardi, M.K. Celebrating 20 years of genetic discoveries in legume nodulation and symbiotic nitrogen fixation. Plant Cell 2020, 32, 15–41. [Google Scholar] [CrossRef] [PubMed]
- Berdugo-Cely, J.A.; Céron-Lasso, MdS.; Yockteng, R. Phenotypic and molecular analyses in diploid and tetraploid genotypes of Solanum tuberosum L. reveal promising genotypes and candidate genes associated with phenolic compounds, ascorbic acid contents, and antioxidant activity. Front Plant Sci 2023, 13, 1007104. [Google Scholar] [CrossRef]
- Bhuvaneswari, G.; Thirugnanasampandan, R.; Gogulramnath, M. Effect of colchicine induced tetraploidy on morphology, cytology, essential oil composition, gene expression and antioxidant activity of Citrus limon (L.) Osbeck. Physiol Mol Biol Plants 2020, 26, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Hyatt, D.C.; Youn, B.; Zhao, Y.; Santhamma, B.; Coates, R.M.; Croteau, R.B.; Kang, C.H. Structure of limonene synthase, a simple model for terpenoid cyclase catalysis. Proc Natl Acad Sci USA 2007, 104, 5360–5365. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Sharma, R.A. Plant terpenes: Defense responses, phylogenetic analysis, regulation and clinical applications. 3 Biotech 2015, 5, 129–151. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Miao, L.; Hou, Q.; Darwish, D.B.; Alrdahe, S.S.; Ali, A.; Benedito, V.A.; Tadege, M.; Wang, X.B.; Zhao, J. Overexpression of terpenoid biosynthesis genes from garden sage (Salvia officinalis) modulates rhizobia interaction and nodulation in soybean. Front Plant Sci 2021, 12, 783269. [Google Scholar] [CrossRef]
- Pott, D.M.; Osorio, S.; Vallarino, J.G. From central to specialized metabolism: an overview of some secondary compounds derived from the primary metabolism for their role in conferring nutritional and organoleptic characteristics to fruit. Front Plant Sci 2019, 10, 835. [Google Scholar] [CrossRef]
- Kalia, V.C.; Gong, C.; Patel, S.K.S.; Lee, J.-K. Regulation of plant mineral nutrition by signal molecules. Microorganisms 2021, 9, 774. [Google Scholar] [CrossRef]
- Jekabsone, A.; Andersone-Ozola, U.; Karlsons, A.; Romanovs, M.; Levinsh, G. Effects of salinity on growth, ion accumulation and mineral nutrition of different accessions of a crop wild relative legume species, Trifolium fragiferum. Plants 2022, 11, 797. [Google Scholar] [CrossRef]
- Devkar, V.; Thirumalaikumar, V.P.; Xue, G.-P.; Vallarino, J.G.; Turechova, V.; Strnad, M.; Fernie, A.R.; Hoefgen, R.; Mueller-Roeber, B.; Balazadeh, S. Multifaceted regulatory function of tomato SITAFI in the response to salinity stress. New Physiol 2019, 225, 1681–1698. [Google Scholar] [CrossRef]
- Amin, I.; Rasool, S.; Mir, M.A.; Wani, W.; Masoodi, K.Z. Ion homeostasis for salinity tolerance in plants: A molecular approach. Physiol Plant 2021, 171, 578–594. [Google Scholar] [CrossRef]
- Tu, Y.; Jiang, A.; Gan, L.; Hossan, M.; Zhang, J.; Peng, B.; Xiong, Y.; Song, Z.; Cui, D.; Xu, W.; Zhang, J.; He, Y. Genome duplication improves rice root resistance to salt stress. Rice (NY) 2014, 7, 15. [Google Scholar] [CrossRef]
- Yang, C.; Zhao, L.; Zhang, H.; Liu, B. Evolution of physiological responses to salt stress in hexaploidy wheat. PNAS 2014, 111, 11882–11887. [Google Scholar] [CrossRef] [PubMed]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: physiological, biochemical, and molecular characterization. Int J Genomics 2014, 701596. [Google Scholar] [CrossRef] [PubMed]
- Machado, R.M.A.; Serralheiro, R.P. Soil salinity: Effect on vegetable crop growth. Management practices to prevent and mitigate soil salinization. Hortic 2017, 3, 30. [Google Scholar] [CrossRef]
- Hamayun, M.; Hussain, A.; Khan, S.A.; Kim, H.-Y.; Khan, A.L.; Waqas, M.; Irshad, M.; Iqbal, A.; Rehman, G.; Jan, S.; Lee, I.-J. Khan, A.L.; Waqas, M.; Irshad, M.; Iqbal, A.; Rehman, G.; Jan, S.; Lee, I-J. Gibberellins producing endophytic fungus Porostereum spadiceum AGH786 rescues growth of salt affected soybean. Front Microbiol 2017, 8, 686. [Google Scholar] [CrossRef]
- Mangena, P. Role of benzyladenine seed priming on growth and physiological and biochemical response of soybean plants grown under high salinity stress conditions. Int J Agron 2020, 8847098. [Google Scholar] [CrossRef]
- Amirjani, M.R. Effect of salinity stress on growth, mineral composition, proline content, antioxidant enzymes of soybean. Amer J Plant Physiol, 2010, 5, 350–360. [Google Scholar] [CrossRef]
- Roychowdhury, R.; Datta, S.; Gupta, P.; Tah, J. Analysis of genetic parameters on mutant populations of mungbean (Vigna radiata L.) after ethyl methane sulphonate treatment. Not Sci Biol 2010, 4, 137–143. [Google Scholar] [CrossRef]
- Goyal, S.; Wani, M.R.; Raina, A.; Laskar, R.A.; Khan, S. Phenotypic diversity in mutagenized population of urdbean (Vigna mungo (L.) Hepper). Heliyon 2021, 7, e06356. [Google Scholar] [CrossRef] [PubMed]
- Laskar, R.A.; Laskar, A.A.; Raina, A.; Khan, S.; Younus, H. Induced mutation analysis with biochemical and molecular characterization of high yielding lentil mutant lines. Int J Biol Macromol 2018, 109, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Corneillie, S.; De Storme, N.D.; van Acker, R.; Fangel, J.U.; De Bryne, M.; De Rycke, R.; Geelen, D.; Willats, W.G.T.; Vanholme, B.; Boerjan, W. Polyploidy affects plant growth and alters cell wall composition. Plant Physiol 2019, 179, 74–87. [Google Scholar] [CrossRef] [PubMed]
- Brisibe, E.A.; Ekanem, N.G. Ploidy level and nucleotide variations in inflorescence dichotomous cultivars of plantain (Musa spp. AAB genome). BMC Genomics 2019, 20, 713. [Google Scholar] [CrossRef] [PubMed]
- Chao, D.-Y.; Dilkes, B.; Luo, H.; Douglas, A.; Yakubova, E.; Lahner, B.; Salt, D.E. Polyploids exhibit higher potassium uptake and salinity tolerance in Arabidopsis. Sci 2013, 341, 658–659. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu Rev Plant Biol 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant salinity stress: Many unanswered questions remain. Front Plant Sci 2019, 10, 80. [Google Scholar] [CrossRef]
- Schulz, P.; Herde, M.; Romeis, T. Calcium-dependent protein kinase: Hubs in plant stress signaling and development. Plant Physiol 2013, 163, 523–530. [Google Scholar] [CrossRef]
- Almadanim, M.C.; Concalves, N.M.; Rosa, M.T.G.; Alexandre, B.M.; Cordeiro, A.M.; Rodriques, M.; Saibo, N.J.M.; Soares, C.M.; Romao, C.V.; Oliveira, M.M.; Abreu, I.A. The rice cold-responsive calcium-dependent protein kinase OsCPK17 is regulated by alternative splicing and post-translational modifications. Biochim et Biophysic Acta Mol Cell Res 2018, 1865, 231–246. [Google Scholar] [CrossRef]
- Demirkol, G. PopW. improves salt stress tolerance of red clover (Trifolium pratense L.) via activating phytohormones and salinity related genes. Biologia 2023, 78, 979–991. [Google Scholar] [CrossRef]
- Liu, Y.; Yu, Y.; Sun, J.; Cao, Q.; Tang, Z.; Liu, M.; Xu, T.; Ma, D.L.Z.; Sun, J. Root-zone-specific sensitivity of K+- and Ca2+-permeable chemicals to H2O2 determines ion homeostasis in salinized diploid and hexaploidy Ipomoea trifida. J Exp Bot 2019, 70, 1389–1405. [Google Scholar] [CrossRef]
- Huang, H.; Ullah, F.; Zhou, D.-X.; Yi, M.; Zhao, Y. Mechanisms of ROS Regulation of Plant Development and Stress Responses. Front Plant Sci 2019, 10, 800. [Google Scholar] [CrossRef]
- Gui, T.-Y.; Gao, D-H.; Ding, H-C.; Yan, X.-H. Identification of Respiratory Burst Oxidase Homolog (Rboh) family genes from Pyropia yezoensis and their correlation with archeospore release. Front Plant Sci 2022, 13, 929299. [Google Scholar] [CrossRef]
- Matamoros, M.A.; Becana, M. Molecular responses of legumes to abiotic stress: Post-translational modifications of proteins and redox signaling. J Exp Bot 2021, 72, 5876–5892. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Wan, Q.; Zhang, K.; Zhang, X.; Guo, R.; Wang, C.; Zheng, C.; Liu, F.; Ding, Z.; Wan, Y. AhABI4s negatively regulate salt-stress response in peanut. Front Plant Sci 2021, 12, 741641. [Google Scholar] [CrossRef]
- Sinha, P.; Saxena, R.K.; Singh, V.K.; Krishnamurthy, L.; Varshney, R.K. Selection and validation of housekeeping genes as reference for gene expression studies in pigeonpea (Cajanus cajan) under heat and salt stress conditions. Front Plant Sci 2015, 6, 1071. [Google Scholar] [CrossRef]
- Guo, X.; Jiang, J.; Liu, Y.; Yu, L.; Chang, R.; Guan, R.; Qiu, L. Identification of a novel salt tolerance-related locus in wild soybean (Glycine soja Sieb. & Zucc.). Front Plant Sci 2021, 12, 791175. [Google Scholar] [CrossRef]
- Tapia, G.; Morales-Quintana, L.; Parra, C.; Berbel, A.; Alcorta, M. Study of nsLTPs in Lotus japonicus genome reveal a specific epidermal cell member (LjLTP10) regulated by drought stress in aerial organs with a putative role in cutin formation. Plant Mol Biol 2013, 8, 485–501. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Selee, B.; Schnabel, E.L.; Poehlman, W.L.; Chavan, S.A.; Frugoli, J.A.; Feltus, F.A. time series transcriptome analysis in Medicago truncatula shoot and root tissue during early nodulation. Front Plant Sci 2022, 13, 861639. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Lopez, J.C. Narrow-leafed lupin (Lupinus angustifolius L.) functional identification and characterization of the aldehyde dehydrogenase (ALDH) gene superfamily. Plant Gene 2016, 6, 67–76. [Google Scholar] [CrossRef]
- Konzen, E.R.; Recchia, G.H.; Cassieri, F.; Caldas, D.G.G.; yTeran, J.C.B.M.; Gepts, P.; Tsai, S.M. DREB genes from common bean (Phaseolus vulgaris L.) show broad to specific abiotic stress responses and distinct levels of nucleotide diversity. Int J Genomics 2019, 9520642. [Google Scholar] [CrossRef]
- Guefrachi, I.; Nagymihaly, M.; Pislariu, C.I.; Van de Velde, W.; Ratet, P.; Mars, M.; Udvardi, M.K.; Kondorosi, E.; Mergaert, P.; Alunni, B. Extreme specificity of NCR gene expression in Medicago truncatula. BMC Genomics 2014, 15, 712. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Yu, X.; Shao, J.; Liu, Z.; Gao, T.; Zheng, Y.; Zeng, C.; Liang, C.; Chen, C. Transcriptomic profiling and analysis of differentially expressed genes in asparagus bean (Vigna unguiculata ssp. sesquipedalis) under salt stress. PLoS ONE 2019, 14, e0219799. [Google Scholar] [CrossRef] [PubMed]
- Ragel, P.; Raddatz, N.; Leidi, E.O.; Quintero, F.J.; Pardo, J.M. Regulation of K+ nutrition in plants. Front Plant Sci 2019, 10, 281. [Google Scholar] [CrossRef] [PubMed]
- Delormel, T.Y.; Boudsocq, M. Properties and functions of calcium-dependent protein kinases and their relatives in Arabidopsis thaliana. New Physiol 2019, 224, 585–604. [Google Scholar] [CrossRef]
- Joshi, A.; Dang, H.Q.; Vaid, N.; Tuteja, N. Isolation of high salinity stress tolerant gene from Pisum sativum by random overexpression in Escherichia coli and their functional validation. Plant Signal Behav 2009, 4, 400–412. [Google Scholar] [CrossRef]
- Zhang, Y.; Du, P.; Xiong, F.; Zhang, X.; Song, H. WRKY Genes improve drought tolerance in Arachis duranensis. Front Plant Sci 2022, 13, 910408. [Google Scholar] [CrossRef]
- Mergaert, P.; Kereszt, A.; Kondorosi, E. Gene expression in nitrogen-fixing symbiotic nodule cells in Medicago truncatula and other nodulating plants. Plant Cell 2020, 32, 42–68. [Google Scholar] [CrossRef]
- Garg, R.; Jain, M. Transcriptome analyses in legumes: A resource for functional genomics. Plant Genomics 2013, 6, 1–9. [Google Scholar] [CrossRef]
- Sprent, J.I.; Ardley, J.; James, E.K. Biogeography of nodulated legumes and their nitrogen-fixing symbionts. New Phytol. 2017, 215, 40–56. [Google Scholar] [CrossRef]
- Trujillo, D.I.; Silverstein, K.A.; Young, N.D. Nodule-specific plat domain proteins are expanded in the Medicago lineage and required for nodulation. New Phytol 2019, 222, 1538–1550. [Google Scholar] [CrossRef]
- Poehlman, W.L.; Schnabel, E.L.; Chavan, S.A.; Frugoli, J.A.; Feltus, F.A. Identifying temporally regulated root nodulation biomarkers using time series gene co-expression network analysis. Front Plant Sci 2019, 10, 1409. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Sun, W.; Yang, G.; Sun, G. WRKY transcription factors in legumes. BMC Plant Biol 2018, 18, 243. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of plant responses to salt stress. Int J Mol Sci 2021, 22, 4609. [Google Scholar] [CrossRef] [PubMed]
- Tyerman, S.D. Nonselective cation channels. Multiple functions and commonalities. Plant Physiol 2002, 128, 327–328. [Google Scholar] [CrossRef]
- Taffouo, V.D.; Kouamou, J.K.; Ngalangue, L.M.T.; Ndjeudji, B.A.N.; Akoa, A. Effects of salinity stress on growth, ions partitioning and yield of some cowpea (Vigna unguiculata L. Walp.) cultivars. Int J Bot 2009, 5, 135–143. [Google Scholar] [CrossRef]
- HanumanthaRao, B.; Nair, R.M.; Nayyar, H. Salinity and high temperature tolerance in mungbean [Vigna radiata (L.) Wilczek] from a physiological perspective. Front Plant Sci 2016, 7, 957. [Google Scholar] [CrossRef]
- Kurz, M. Compatible solute influence on nucleic acids: Many questions but few answers. Saline Syst 2008, 4, 6. [Google Scholar] [CrossRef]
- Koning, R.; Kiekens, R.; Toili, M.E.M.; Angenon, G. Identification and expression analysis of the genes involved in the raffinose family oligosaccharides pathways of Phaseolus vulgaris and Glycine max. Plants 2021, 10, 1465. [Google Scholar] [CrossRef]
- Wei, T.; Wang, Y.; Liu, J.H. Comparative transcriptome analysis reveals synergistic and disparate defense pathways in the leaves and roots of trifoliate orange (Poncirus trifoliata) autotetraploids with enhanced salt tolerance. Hortic Res 2020, 7, 88. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, Y. Unraveling salt stress signaling in plants. J Int Plant Biol 2018, 60, 796–804. [Google Scholar] [CrossRef]
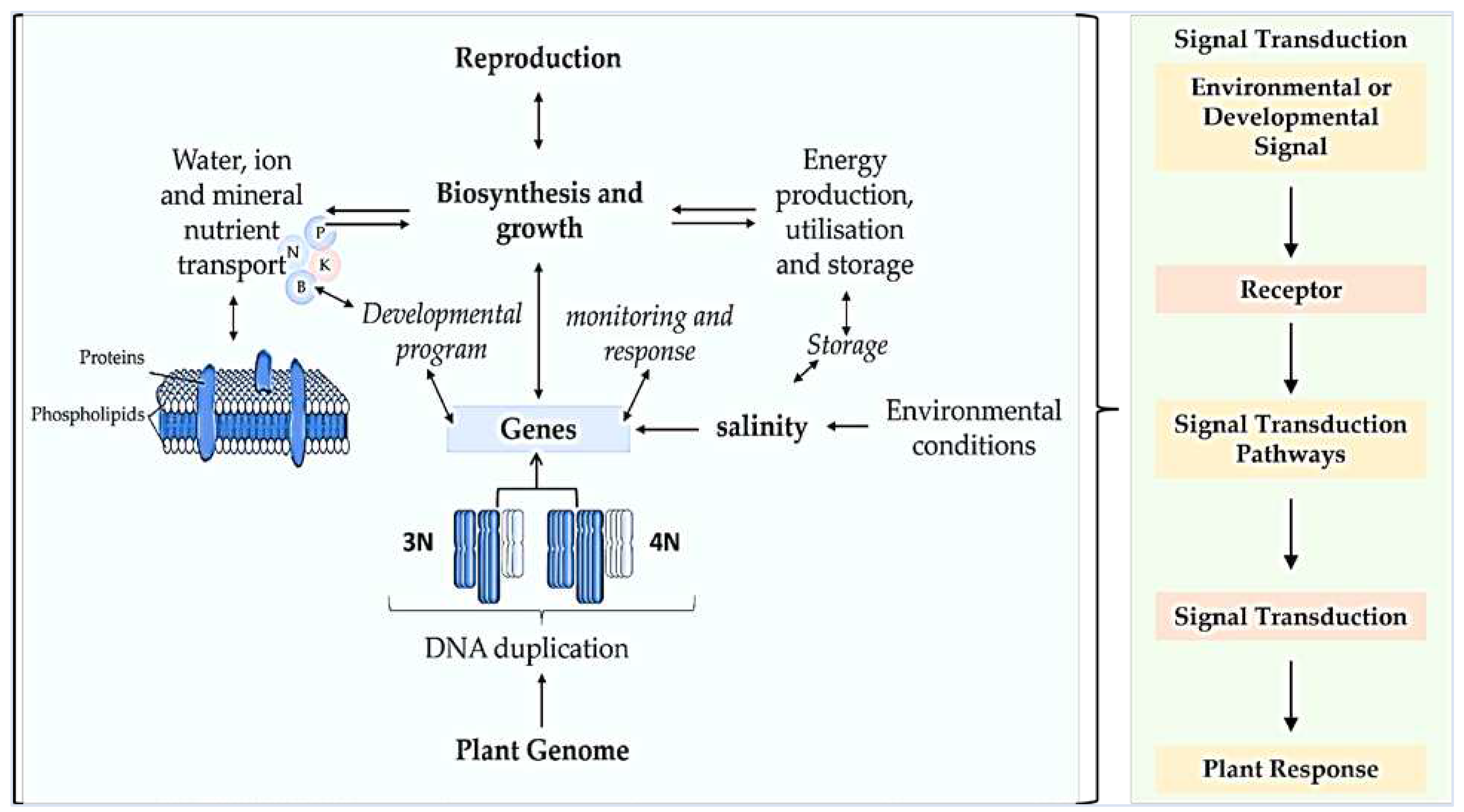
Figure 3.
Biochemical and physiological events in the sensing and response of plants to salinity stress. The prevalence and recurrence of polyploidization in plant species make it one of the most important evolutionary events [36,37] in diversification of species through DNA duplication, altered developmental and monitoring responses, energy production, utilisation and storage during abiotic stress and evolution of genes, as well as high reproduction. Environmental or developmental signals include light, temperature, nutrients etc., receptors such as kinases, ion channels and G-protein-coupled-receptors; repressor protein degradation and protein phosphorylation; hormonal transport etc.; and responses by transcriptional/ gene expression and post-translational modifications were improved in polyploidised plants [31,38].
Figure 3.
Biochemical and physiological events in the sensing and response of plants to salinity stress. The prevalence and recurrence of polyploidization in plant species make it one of the most important evolutionary events [36,37] in diversification of species through DNA duplication, altered developmental and monitoring responses, energy production, utilisation and storage during abiotic stress and evolution of genes, as well as high reproduction. Environmental or developmental signals include light, temperature, nutrients etc., receptors such as kinases, ion channels and G-protein-coupled-receptors; repressor protein degradation and protein phosphorylation; hormonal transport etc.; and responses by transcriptional/ gene expression and post-translational modifications were improved in polyploidised plants [31,38].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



