
Submitted:
24 June 2023
Posted:
25 June 2023
You are already at the latest version
Abstract
NAD+ is known classically as a metabolite that participates in catabolic and anabolic pathways throughout the metabolism that is taught to students in introductory biochemistry courses. However, non-classical studies starting over a decade ago found that NAD+ is also involved in higher order functions, in part because of its involvement in the activation of SIRTs and the support of the mitochondrial unfolded protein response. Many studies since have suggested that NAD+ is involved in an extensive spectrum of human pathologies, including neurodegenerative disorders, cardiomyopathy, obesity, and diabetes, Further, healthy aging and longevity appear to be closely related to NAD+ and its related metabolites, including NAR and NMN. Together, these studies show that this system has value as a dietary supplement to improve general health overall, as well as mitigating specific disease conditions. Accordingly, many are now recommending the consumption of materials in this system as dietary supplements. Nicotinamide riboside (NAR) appears to have special value in this regard. It appears to be better tolerated than other molecules in this system, as well as better pharmacodynamics and greater potency. Unfortunately, NAR is a reactive molecule, often unstable during its manufacturing, transport, and storage. Indeed, HPLC analyses of many commercial samples of NAR shows that they contain substantial amounts of material that are not, in fact, NAR. In some of these commercial preparations, NAR is a minority component. Therefore, more stable derivatives of NAR that are easily converted upon consumption into NAR are therefore desired. Recently work related to prebiotic chemistry provided the borate derivative of NAR. NARB is considerably more stable than NAR itself. However, immediately upon consumption, the borate dissociates from NARB, and is lost in the body through dilution and binding to other species, notably carbohydrates such as fructose and glucose. The NAR left behind is expected to behave pharmacologically in ways identical to NAR itself. This review provides a comprehensive summary (through Q1 of 2023) of literature that makes the case for the consumption of NAR as a dietary supplement. It then summarizes the challenges of delivering quality NAR to consumers using standard synthesis, manufacture, shipping, and storage approaches. It concludes by outlining the advantages of NAR-borate in these processes.
Keywords:
nicotinamide riboside
; vitamin B3 derivative
; efficacy
; safety
; healthy aging
; longevity
1. Introduction
Nicotinamide riboside (NAR) is widely used as a dietary supplement, Structurally, it is an advanced form of vitamin B3 (nicotinic acid), incorporating into its structure more elements of nicotinamide adenine dinucleotide (in its oxidized form, NAD+) [1]. NAR influences, in particular, energy metabolism and neuroprotection [2,3,4].
From a non-medical perspective, as a part of the nicotinamide adenine dinucleotide (NAD+) cofactor, it also inferred to be a vestige of the “ribonucleic acid (RNA) world” [5], an episode of life on early Earth where RNA was the only encoded component of both metabolism and genetics, and this inference is supported by the ubiquity of NAD+ in all branches of tree of modern life [6].
NAR is, however, a rather reactive molecule. Its glycosidic bond joins a positively charged pyridinium heterocycle to a carbohydrate. This bond is therefore especially unstable to cleavage, making NAR difficult to synthesize, store, and transport. This creates broad utility for any method for the synthesis of NAR, as well as broad utility for derivatives of NAR that are more stable.
Stable forms of NAR are especially important today. Since July 2013, NAR became accessible in dietary supplement form of NAR chloride (NARC), NAR is sold and widely consumed as a dietary supplement, under various trademarks, including Tru Niagen™ and Niagen® (produced by ChromaDex). Other products containing NARC have been commercially available since [7,8]. Interesting, analysis of some of these products at various times has shown that they do not contain pure NAR, but also products undoubtedly arising from the reactivity of NAR.
Notwithstanding, commercial NAR products are suggested to elevate the level of oxidized NAD (NAD+) in those who consume it [8]. Studies report that chronic NAR supplementation (NARS) is well tolerated and elevates NAD+ in healthy middle-aged and older adults [2,3]. Other studies suggest NAR oral bioavailability in mice and humans [9]. NAR as a dietary supplement has also been the target of clinical studies [2,3]. These have proven its effect to boost NAD+ in its consumers [8,10]. These facts make it timely to review the current state of the art.
NAR, a pyridine-nucleoside form of vitamin B3, consists of nicotinamide (NA) and ribose as its fundamental components. It can be found in various sources such as milk, yeast, beer, as well as bacteria and mammals.
NAR-enriched foodstuffs per se are not well established. Presumably, products that contain yeast are excellent natural sources for the compound [11,12]. However, dairy products have also been noted to contain NAR [9,13]. The amounts of NAR in foodstuffs are extremely low, most likely at micromolar levels.
NAR shares biological properties with other NAD+ precursors, specifically NA and nicotinic acid (NAc, niacin). Indeed, these are old or “classical” vitamins B3. Unlike NAR, the NA and niacin have disadvantages when consumed. Hepatotoxicity or flushing can be potential side effects associated with NA, whereas a recent preclinical study indicates that NA remains in the rat body for a shorter duration when compared to NAR.
Niacin taken in large amounts is associated with adverse effects, including cutaneous flushing when administered in an immediate release formulation. Sustained release formulations may cause hepatotoxicity.
A precursor that is structurally more advanced towards NAD+ is nicotinamide mononucleotide (NMN). NMN has been recently discovered to be converted extracellularly to NAR, which is transported into cells [11,14,15]. Since NMN is converted to NAR in the body and the price of NMN is half of the price of NAR, technology producing NAR rather than NMN is especially needed [16].
Among the NAD+ precursors, NAR is exceptional, as it produces fewer reported unfavorable side effects [7].
In its classical metabolic roles, NAD+ is a versatile receiver of hydride equivalents, becoming NAD (NADH). Analogous chemistry is seen with its phosphorylated derivative, nicotinamide adenine dinucleotide phosphate (NADP and NADPH). Traditionally, NAD+ and its derivatives function as coenzymes for dehydrogenases and oxidoreductases, playing crucial roles in fundamental energy metabolism processes such as glycolysis, the citric acid cycle, and mitochondrial (MT) electron transport. In addition to its classical functions, NAD+ also serves as a vital substrate for signaling enzymes, e.g., poly [adenosine diphosphate (ADP)-ribosyl] polymerases, sirtuins (SIRTs), and ADP-ribosyl-transferases. These enzymes are referred to as “NAD+ consumers” [17,18,19,20,21].
Throughout classical understanding as well as recent discoveries, NAD+ has been recognized as a crucial and abundant metabolite present in all mammalian cells. It participates in a wide array of cellular mechanisms, including essential processes such as metabolism and cell signaling that are vital for survival. However, the limited presence of NAR in food sources (with some quantitative studies available) and the challenges associated with obtaining significant quantities of pure NAR as a dietary supplement have hindered extensive research on the effects of NAR on cells and tissues [22,23]. Nevertheless, recently, there have been notable advancements in the synthesis techniques for producing NAR, resulting in new and refined approaches [24]. However, these new technologies were developed for cell-based investigations and animal feeding tests, and not for human consumption [25,26].
Non-classical roles also exist for NAD+. For example, NAD+ also activates SIRTs and supports the mitochondrial response to unfolded proteins. Pursuing these non-classical roles, NAD+ metabolism is seen to be involved in an extensive spectrum of pathologies, including cancer, neurodegenerative disorders, cardiomyopathy, obesity, diabetes, and even hearing loss [4,27,28,29,30,31].
In many tissues (e.g., brain, muscle, skin, liver, pancreas, and adipose tissue), the level of NAD+ decreases with age. A separate decrease in the NAD+/NADH ratio affects the cellular redox state, highlighted by anaerobic glycolysis and oxidative phosphorylation (OXPHOS). This decreases, it is thought, the capacity of cells to generate adenosine triphosphate (ATP) [32]. Consequently, healthy aging and longevity are thought to be related to NAD+ metabolism, mainly through NAR and NMN, two essential and well-studied NAD+ derivatives. One argument for NA-related dietary supplements is that these can have prophylactic and therapeutic impact on functional decline, improving age-associated neurodegenerative, cardiovascular (CV), and metabolic diseases and conditions, and promoting the beneficial effects of calorie restriction (CR) [4,33,34,35,36].
As a precursor of NAD+, NAR is also proposed to be important in regulating oxidative stress (OXS), inhibiting oxidative injury and inflammatory response, with beneficial effects in the treatment of sepsis [37], systemic lupus erythematosus (SLE) [38], and necrotizing enterocolitis (NEC) [39]; furthermore, NAR showed significant protective effect on lung injury caused by paraquat (PQ) in mice [40].
Separately, sepsis-caused multiple organ failure is the major risk of morbidity and mortality in intensive care units. In experimental models of mouse sepsis induced by lipopolysaccharide (LPS) injection, feces injection in peritoneum or by cecal ligation and puncture (CLP), NAR inhibited plasma high mobility group box 1 (HMGB1) release, OXS, and tissue infiltration, increased endogenous antioxidant ability, prevented lung and heart injury and improved survival [37,41]. This is mediated via NAD+/SIRT1 signaling.
NAR could be a potential adjuvant for SLE treatment: in vitro testing in monocytes from patients with SLE showed that NAR restricted autophagy and attenuated interferon-beta (IFN-β) release in an NAD+-dependent manner but also through inosine signaling [38]. Also, in the mouse experimental NEC model, NAR administration alleviated OXS, increased NAD+ levels and intestinal microcirculatory perfusion, and relieved signs of endothelial dysfunction by modulating the SIRT1-associated endothelial nitric oxide synthase (eNOS) acetylation/deacetylation pathway [39,42].
Compared with control (CON) group, which received saline by one-time gavage, intraperitoneal (i.p.) injection of 300 mg/kg NAR solution led to the inhibition of the inflammatory response, peroxidation injury and apoptosis at the lung level, and to the survival time prolongation of the PQ intoxicated mice, mainly by upregulating SIRT1 and nuclear factor erythroid 2 (NF-E2)-related factor 2 (NRF2) protein expression [40].
We recently reported an inexpensive synthesis of borate-stabilized NAR based on models for how nucleosides might have been formed on a prebiotic Earth. Unlike many commercially available NAR products, borate-stabilized NAR can be delivered in pure form, exhibiting high stability against thermal and chemical decomposition. Thus, NAR borate (NARB) is a promising nutraceutical to replace NARC, as currently sold on the US market. This paper reviews the current research supporting the value of NAR for healthy aging and longevity, the perspectives of senotherapeutic NAD+ supplementation, and thus forms of NAR that can be made, shipped, and stored without decomposition.
2. Effects of Nicotinamide Riboside on Several Organs and Systems
2.1. Nicotinamide Riboside and the Nervous System
The effects of NAR on energy metabolism and neuroprotection were highlighted with the first research about this compound. In yeast and mammals, NAR is metabolized by two main pathways: (i) degradative processes from which NA results; (ii) synthetic processes exploiting NAR kinases (NRK1, NRK2), resulting in the increasing of tissue NAD+. This increase is reported to increase insulin sensitivity, mitochondrial biogenesis, and SIRT functions.
Thus, in experimental models of Alzheimer’s disease (AD), NAR in larger amounts than is found naturally in food leads to brain protective effects by the stimulation of NAD+ anabolism [7]. NAR is a neuroprotective factor that improves cognition after brain injuries, such as acute ischemia. In an experimental model, 300 mg/kg NARC was i.p. administered, 20 minutes after reperfusion, in mice with middle cerebral artery (MCA) occlusion. The NARC-treated group exhibited better memory function and recovery of learning in the Morris water maze test.
Following acute treatment with NARC, apoptosis in the hippocampus, neuronal loss, and hippocampal infarct volume were decreased. NARC also increased the measured amounts of NAD+ and ATP, and stimulated adenosine 5’-monophosphate (AMP)-activated protein kinase and SIRT1, as observed by high-performance liquid chromatography (HPLC) and Western blot assays [43]. Moreover, the clinical phenotype and T-cell survival/function were improved by S-adenosylmethionine (SAM) and NAR co-administration for patients with phosphoribosyl pyrophosphate synthetase 1 (PRPS1) deficiency (Arts syndrome, manifested by serious neurological and immunological deficiencies especially in males) [44].
NAR is reported to be beneficial in other neurological syndromes. For example, Gulf War illness (GWI) is a chronic neuropsychiatric disorder characterized by neurocognitive deficits arising with OXS, neuroinflammation and neuronal damage. Currently, no effective treatment of GWI is known [45]. However, in a GWI experimental model, NAR-mediated SIRT1 activation restored brain MT bioenergetics dysfunction (following astroglia activation) and reduced neuroinflammation. Here, NAR was administered as a dietary supplement, 100 μg/kg daily for two months. In the brain of NAR-treated GWI mice, a decrease of lipid peroxidation and proinflammatory cytokines was closely related with the increase of deacetylation of the peroxisome proliferator-activated receptor gamma coactivator-1alpha (PGC-1α) and nuclear factor-kappaB (NF-κB) p65 subunit [46].
Many central nervous system (CNS) disorders arise from dysfunctionalities in the gut microbiota–brain axis. In the mice brain, NARS provided protection against alcohol-induced depressive behavior and decreased the level of anti-inflammatory [interleukin (IL)-10 and transforming growth factor (TGF)-β] and proinflammatory [IL-1β, IL-6, and tumor necrosis factor (TNF)-α] cytokines. In the hippocampus, NAR significantly reduced the number of activated microglia and the inhibition of the protein kinase B (AKT)/glycogen synthase kinase 3 beta (GSK3β)/β-catenin signaling pathway and increased the brain-derived neurotrophic factor (BDNF). Similar with donor mice, in recipient mice, fecal microbiota transplantation (FMT) improved the microglial activity, the level of BDNF and cytokines, the activation of AKT/GSK3β/β-catenin signaling pathway, and cognitive behavior [47].
Intracortical administration of NAR protects against excitotoxicity-induced axonal degeneration (AxD) and decreases the brain damage provoked by injection of N-methyl-D-aspartate (NMDA). Both NAR and NAD+ prevented neuronal death due to the axonal stress. However, NAR exhibited better neuroprotection than NAD+ at the level of cortical neurons [48].
Moreover, in rats with TNF-induced optic nerve degeneration, intravitreal injection of NAR showed significant axonal protection. In retina and optic nerve, NAR upregulated the levels of SIRT1 autophagy pathway, decreased the p62 protein levels and increased the levels of microtubule-associated protein 1A/1B-light chain 3-II (LC3-II) that localizes with the mitochondrial inner membrane. The presence of NRK1, upregulated by NAR administration, was also evidenced in the retinal and optic nerve fibers and in the retinal ganglion cells (RGCs) [49,50].
This research continues. For example, recently, Sun et al. (2023) proved that NAD+ depletion mediates cytotoxicity in human neurons with autophagy (ATG) deficiency, leading to cytotoxicity and neurodegeneration. The research was conducted on autophagy-deficient (ATG5–/–) human embryonic stem cells (hESCs) representing a so-called ‘human neuronal platform’. In ATG5–/– neurons, NAR treatment improved cell viability by the restoration of MT bioenergetics and proteostasis [51].
In experimental (preclinical) models and also in clinical studies, NAR improved ataxia scores and immunoglobulin G (IgG) levels in ataxia telangiectasia (A–T). In ATM-deficient mice, which is a model for the A–T phenotype in humans, NAR administration improved motor function and prevented neuroinflammation, neurodegeneration, MT dysfunction and cellular senescence [52]. NAR dietary supplementation was well tolerated by A–T patients; ataxia, dysarthria, and the quality of life (QoL) were all reportedly improved without adverse effects [53,54].
In cluster of differentiation (CD) 157 knockout (KO) male mice, oral administration of NAR increased the levels of oxytocin (OT) in cerebrospinal fluid, stimulating the release of this anxiolytic factor during stress, and corrected the fearful and anxiety-like behaviors and the social deficits [55]. A recent study highlighted the remission of social behavior impairment by oral gavage administration of NAR for 12 days in CD157, but not in CD38 KO mice [56].
In cell culture (in vitro) and in mouse models of amyotrophic lateral sclerosis (ALS), NAR and pterostilbene (PT) supplementation cooperatively delayed motor neuron failure, decreased the levels of neuroinflammation markers in the spinal cord, influenced the muscle metabolism, and increased to a small extent the survival of hSOD1G93A transgenic mice [57]. Further, in SOD1G93A transgenic and wild-type (WT) mice, starting at 50 days of age, oral administration of NAR (20 mg/mL) improved MT proteostasis and adult neurogenesis. In the brain of SOD1G93A mice, neural stem cells (NSCs)/neuronal precursor cells (NPCs) proliferation and migration were enhanced following the NARS [58]. A recent clinical trial evidenced that NAR and pterostilbene (PT) co-administration proved to be effective in inhibiting OXS-induced cellular damage, the major pathophysiological mechanism of ALS [59,60].
In mouse models for Cockayne syndrome (CS) (CSA–/– and CSBm/m, modeling human CS), NAR is reported to protect from noise-induced hearing loss (NIHL). The main mechanism of action involves the activation of NAD+/SIRT3 pathway, which contributes to the reduction of spiral ganglia neurite degeneration caused by intensive noise exposure (NE). NAD+ supplementation could prove beneficial for the treatment of CS-related hearing loss at a young age and age-related hearing loss (ARHL) affecting elderly individuals [61,62,63].
In Sprague–Dawley female rats, oral administration of NAR (200 mg/kg) for four weeks relieved the nociceptive and aversive dimensions of intravenous (i.v.) paclitaxel (PTX)-induced peripheral neuropathy. In addition, NARS increased NAD+ plasma level by 50% and did not influence the myelosuppressive properties of PTX, the adverse locomotor effects not being observed [64].
In male rats, NAR oral treatment (500 mg/kg) alleviated PTX-induced corneal and somatic hypersensitivity to tactile stimuli, with no suppression of basal tear production/chemosensitivity and without altering the corneal afferent density. NAR also reversed the PTX-induced hindpaw hypersensitivity to cool and tactile stimuli, with no inverting of the non-peptidergic intraepidermal nerve fiber (IENF) loss [65].
In the recent studies regarding neuroprotective therapy for glaucoma, in a mouse model of acute retinal ganglion cell (RGC) damage caused by optic nerve crush (ONC), and of chronic RGC degeneration obtained by intracameral injection of microbeads that induced ocular hypertension, orally administered NAR removed retinal inflammation, as shown by immunofluorescence staining for glial fibrillary acidic protein (GFAP). It also enhanced survival and preserved the function of RGC [66,67]. In a mouse model of light-induced retinal degeneration, i.p. injected NAR increased the NAD+ level and had a protective effect on retinal function through the restoration of the photoreceptor cell layers, by the reduction of the inflammation and by the limitation of the consequences of apoptosis process [68].
In 125 patients with primary open-angle glaucoma (POAG), a randomized double-blind, placebo (PLA)-controlled trial showed an effect of NAR (300 mg NAR for 24 months) in slowing down optic nerve degeneration. Compared with PLA, NAR patients had a lower degree of progressive retinal nerve fiber layer (RNFL) thinning and visual field (VF) loss [69,70].
The protective effects of NAR were also investigated on hydrogen peroxide-induced oxidative damage in lens epithelial cells. In vitro, on SRA01/04 cell line, NAR significantly reduced apoptosis and the generation of reactive oxygen species (ROS), increased cell viability, and improved levels of superoxide dismutase (SOD), catalase (CAT), total glutathione (GSH), and MT membrane potential. NAR was found to reduce OXS damage through the targeting of mitogen-activated protein kinase (MAPK) and Janus kinase 2 (JAK2)/signal transducer and activator of transcription 3 (STAT3) pathways [71].
Pang et al. (2021) reported that some aberrant metabolic pathways during pregnancy, such as NAD+, OXPHOS, tricarboxylic acid (TCA) cycle and tryptophan metabolism, underlie Zika virus (ZIKV)-induced microcephaly in newborns. Preclinical experiments suggested that in ZIKV-infected mice, NARS improved survival, reduced the apoptosis, and increased the thickness of the cerebral cortex [72].
2.2. Nicotinamide Riboside and the Cardiovascular System
Dietary supplementation by NAD+ precursors (e.g., NAR) appears to have useful impact in many areas of cardiovascular diseases (CVDs). Here, the goal is to improve overall cardiometabolic health, by increasing OXPHOS capacity and mitophagy, removing of mitochondrial deoxyribonucleic acid (mtDNA) mutations, and for cardioprotective effects in ischemia–reperfusion (I–R) injury, arrhythmias, heart failure (HF), myocardial infarction (MI), and high blood pressure [73,74,75,76].
For example, in an experimental model of cafeteria (CAF) diet-induced obesity (DIO), oral administration of 400 mg/kg of NAR combined with CR (-62% kcal) for 28 days decreased the weight of obese male Wistar rats, their visceral and subcutaneous adiposity, their triglyceride (TG)/high-density lipoprotein (HDL) ratio, and their heart size. Also, insulin resistance and antioxidant capacity (glutathione peroxidase (GPx) and CAT enzymatic markers of cardiac OXS) were improved by NARS supplements [77]. In a mouse model, NAR treatment for 30 days led to the NAD+/NADH ratio balancing, lowering acetylation level, improving mitochondrial function and HF with preserved ejection fraction (HFpEF) phenotypes [78]. In a murine model of myocardial hypertrophy induced by transverse aortic constriction (TAC) surgery, NAR dietary supplementation reduced levels of inflammatory cytokines (IL-1β, TNF-α), mitigated the NLR family pyrin domain containing 3 (NLRP3) inflammasome activation, the elevation of myocardial NAD+ level, and the improving of cardiac dysfunctions and morpho-functional changes (myocardial hypertrophy). The research led to the observation that the NAD+/SIRT3/manganese superoxide dismutase (MnSOD) signaling pathway was also regulated by NARS [79].
Moreover, in mice, NAR administration inhibited TAC-induced endothelial-to-mesenchymal transition of endothelial cells (ECs) and promoted MT unfolded protein response leading to the improvement of prohibiting proteins’ expression and to the decrease of cardiac fibrosis (CF) progression [80].
Recent clinical trials highlighted the cardioprotective role of oral NARS (usually 1 g twice daily, a well-tolerated dose) for ischemic heart disease patients diagnosed with atrial fibrillation (AF) [81], HF patients with MT dysfunction and peripheral blood mononuclear cells (PBMCs) inflammatory activation [82] or clinically stable HF patients with reduced ejection fraction (EF) [83].
In murine models, boosting NAD+ levels using dietary NARS appeared to reduce the development of aortic aneurysms and sudden death by aortic ruptures. Acute aortic aneurysms and lethal ruptures were induced by angiotensin II (Ang II) administration on apolipoprotein E (ApoE)-deficient mice fed throughout the entire experiment with a Western diet (WD). Fatal aortic ruptures caused by atherosclerosis and hypertension are reported to be prevented by boosting mitochondrial respiration [84].
Additional studies in this space have accumulated. Thus, NRK2 is reported to limit dilated cardiomyopathy (DCM) in mice with chronic pressure overload (PO). In addition, Ang II-induced cardiomyocyte death is reported to be mitigated by NRK2 overexpression [85]. NAR administration preserves cardiac function in a mouse model of DCM and TAC-induced cardiac hypertrophy, alleviating HF development by stabilizing NAD+ levels in the myocardial tissue.
NAc adenine dinucleotide, methylnicotinamide (MeNA), and N1-methyl-4-pyridone-5-carboxamide represent the three biomarkers with increased myocardial levels due to NARS [86]. In murine models, NAR was i.v. administered to assess its ability to mitigate doxorubicin (DOX)-induced cardiomyopathy. OXS, autolysosomes accumulation and autophagic flux blockade because of DOX cardiotoxicity were all prevented by NAR via NAD+/SIRT1 signaling pathway [87,88]. For the specific pathogen-free (SPF) male Wistar rats, the protective effect exhibited significant levels through preventive use of NAR [89].
NAR also attenuated cardiac and I–R injury. In the ischemic heart, NRK2 regulates metabolic adaptation and MT function post-MI [90]. In a cardiac I–R injury experimental (mouse) model, orally administered NAR and resveratrol (RSV) nanocrystal self-assembled microspheres (NAR/RSVms) for eight hours decreased MI, with no important adverse effects on internal organs [91]. Also, in a fentanyl–midazolam anesthetized I–R injury rat model, NAR was i.v. administered as a bolus before the ischemia started. It was found that NAR could target cardiac I–R injury as a promising cardioprotective natural compound [92]. Moreover, in a mouse model of superior mesenteric artery ischemia, NARS improved microcirculation and mesenteric vessels relaxation, and protected the intestinal wall against I–R injury [93].
In human aortic endothelial cells (HAECs) and in murine aortic rings, NAR acts as vasoprotective agent at the endothelial level by modulation of intracellular NAD+ and inhibition of inflammation, as evidenced by decreased expression of von Willebrand factor (vWF) and ICAM-1 [94].
2.3. Nicotinamide Ribose and the Digestive System
In mice subjected to partial hepatectomy, boosting NAD+ by putting NARS in the drinking water promoted liver regeneration. This was interpreted as increasing DNA biosynthesis at the hepatocyte level and amelioration of HS [95]. In an HFD-fed murine model, NAR treatment enhanced hepatic MT function and NAD+ levels and prevented lipid accumulation in the liver [96]. In vitro, HS was induced in AML12 mouse hepatocytes treated with 250 μM palmitic acid (PA) for 48 hours. Cell exposure to NAR (10 μM and 10 mM) for 24 hours did not affect morphology nor viability, instead it was observed a decreasing of TNF-α and IL-6 proinflammatory markers and an increasing of SIRT1 gene activity, PGC-1α, transcription factor A, carnitine palmitoyltransferase 1, uncoupling protein 2, mtDNA and MT biogenesis [97].
Supplementation with NAD+ precursors (NAR, NMN) can prevent liver injury. Proposals to treat non-alcoholic fatty liver disease (NAFLD). NRK1 by targeting for hepatic steatosis (HS) are based in the discovery that in aged mice or in mice treated with high-fat diet (HFD), NRK1 overexpressed by adenovirus mediated gene transduction regulates lipid metabolism (mainly TG level) and NAD+ biosynthesis in the liver, insulin sensitivity and glucose tolerance [98]. In aged mice with moderate NAFLD-like phenotype, NARS (2.5 g/kg food for three months) significantly reduced the levels of total cholesterol (TC), TG, aspartate aminotransferase (AST), and alanine aminotransferase (ALT), and increased the amount of liver NAD+, diminished inflammatory infiltration, and mitigated HS and liver fibrosis (LF) [99,100,101]. In a diet-induced NAFLD mice model, supplementation with a multi-ingredient mixture of betaine, N-acetylcysteine, L-carnitine, and NAR slowed NAFLD progression and influenced the gut–liver axis through the correction of intestinal microbiota dysbiosis and modulation of short-chain fatty acids (SCFAs) levels in feces [102].
Daily NAR and PT (NARPT, Basis™) co-administration in 111 adults with NAFLD was assessed in a six-month prospective, randomized, double-blind, PLA-controlled clinical trial. Here, NARPT treatment was well tolerated, and significantly decreased serum levels of ceramide 14:0 (a toxic lipid), ALT, and gamma-glutamyltransferase (GGT) [103].
In an HFD-induced male C57BL/6 J mice model of LF, NAR administration (400 mg/kg/day for 20 weeks) attenuated the development of LF. The body weight (b.w.), the amount of collagen in the liver, and the activation of hepatic stellate cells (HepSCs) were significantly reduced by NARS, compared to liver inflammation, HS, and ALT levels which have not been mitigated [104]. In a similar context, NAR exhibited a protective effect against LF induced by carbon tetrachloride in mice through the increasing of SIRT1 gene activity, modulation of Smads signaling pathway acetylation, and suppression of TGF-β-induced HepSCs activation [105].
NAR treatment (400 mg/kg·b.w. for 16 days) attenuated liver injuries induced by alcohol in C57BL/6 J mice fed with Lieber–DeCarli ethanol liquid diet. The protective effect of NARS involves SIRT1 activation, increasing hepatic NAD+ levels and PGC-1α deacetylation, enhancing MT biogenesis/functionality, and decreasing OXS status and ethanol-induced lipid accumulation [106].
A recent in vitro study, in a human hepatic (HepG2) cell line model of citrin deficiency highlighted the potential use of NARS for balancing of dysregulated glycolysis and fatty acid β-oxidation (FAO) [107].
NARS has even been suggested as being useful to manage the sequelae of infectious diseases. For example, as part of research aimed at new treatments for severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) disease, it was highlighted that NARS significantly inhibited the murine hepatitis virus (MHV) replication by increasing the level of NAD+, activation of the TCA cycle and of MT metabolism [108].
2.4. Nicotinamide Riboside and the Urinary System
In the United States hospitals, about 3–10% of the adults suffer from acute kidney injury (AKI). NAD+ supplementation, by oral administration of NA, NAc, NAR, dihydronicotinamide riboside (reduced NAR – NARH) and NAM, and SIRT activation may be useful in the prophylaxis and treatment of AKI, if some recent studies on MT metabolism are developed [109,110,111,112,113]. For example, in an experimental model of newborn (one day) mice from mothers fed a low protein diet, NARS restored the SIRT3 expression, by the induction of PGC-1α, improved MT and cellular protection against OXS, increasing the density of nephrons, renal capillaries, and glomerular podocytes [114].
The effect of four NARPT doses (NAR/PT – 1.25 g/50 mg, 2.5 g/100 mg, 3.75 g/150 mg, 4 g/200 mg), administered twice a day for two days, was tested in a randomized, double-blind, PLA-controlled study in 24 hospitalized AKI patients. Serum NAD+ levels were increased by NARPT treatment. Safety was assessed by various assays, such as estimated glomerular filtration rate (eGFR), creatinine levels, electrolyte levels, hepatic function parameters, and blood count; these all remained unchanged. Minor gastrointestinal (GI) side effects were reported only by three patients. NARPT supplementation up to 1 g/200 mg twice a day were reported as being safe and well tolerated in hospitalized AKI patients [110].
In another recent preclinical study, in rats, NAR prophylactic supplementation did not mitigate tubular impairment and induction of profibrotic genes neither in bilateral I–R injury-induced long-term AKI nor in chronic kidney disease (CKD) experimental model, even though an increased NAD+ blood levels were evidenced [115]. However, Doke et al. (2023), a study in male mice with cisplatin- or I–R-induced AKI suggested that NAR and NMN supplementation restored NAD+ serum levels and improved MT metabolism and kidney functionality [116].
A randomized PLA-controlled, double blind, cross-over trial was conducted for six weeks to determine the impact of NAR (1 g/day) and coenzyme Q10 (CoQ10, 1.2 g/day) on exercise tolerance and metabolic profile in 25 CKD patients with eGFR of <60 mL/min/1.73 m2. Compared with PLA, only systemic MT metabolism and lipid profile (e.g., TG, free fatty acids, ceramides) were improved by NAR and CoQ10 co-administration, but not aerobic capacity (VO2 peak) or total work efficiency [117].
2.5. Nicotinamide Riboside and the Musculoskeletal System
Numerous preclinical studies have suggested that NAR and NMN dietary supplements increase NAD+ levels in the skeletal muscle and help protect musculoskeletal system from age-related metabolic dysfunction. NAD+ biosynthesis pathways/homeostasis and its bioavailability in skeletal muscle cells are influenced by NA phosphoribosyltransferase (NAMPT), NRK1, and NRK2 enzymes [118,119]. After the application of NARS in hexose-6-phosphate dehydrogenase (H6PD) KO (H6PDKO) mice, NAD+/NADH ratio was elevated but without any effect on insulin sensitivity, MT respiratory dysfunctions (MT acylcarnitine), acetyl coenzyme A (Ac-CoA) metabolism and endoplasmic reticulum (ER) oxidative stress [120,121].
In a double-blind, PLA-controlled, randomized, crossover trial for 21 days, 12 aged men were supplemented with 1 g/day NAR. It was found that both MT bioenergetics and skeletal muscle NAD+ metabolism were stimulated by NARS; at the same time the amounts of serum inflammatory cytokines were reduced [122]. Moreover, in a randomized, double-blinded, PLA-controlled, crossover intervention study in 13 healthy obese humans, NARS (1 g daily for six weeks) induced some minor modifications in body composition and sleeping metabolic rate, increased the level of NAD+ metabolites such as NAc adenine dinucleotide and MeNA, and altered the acetylcarnitine concentrations in the skeletal muscle [123].
Recent studies showed that, in healthy humans, oral NAD+ supplementation (NAR, NMN) does not alter whole-body or skeletal muscle metabolic responses to an endurance exercise [124,125]. During endurance exercises, one week of NARS (1 g daily) for eight young males did influence neither skeletal muscle MT respiration nor auto-PARylation of poly ADP-ribose polymerase 1 (PARP1), acetylation of tumor protein 53 (p53) Lys382 and MnSODLys122, or overall acetylation [126]. A recent randomized controlled trial testing of in-home aerobic and exercise training and 16 weeks NARS targeted the skeletal muscle MT OXPHOS and improved the muscle mass and fitness in adolescent and young adult (AYA) survivors of hematopoietic cell transplantation (HemSCT) [127].
However, some preclinical studies showed that, compared to the CON group, chronic NARS (300 mg/kg b.w./day for 21 days by oral gavage) decreases the exercise/swimming performance in Wistar rats. The redox features and pleiotropic metabolism of NAD+ and NADP+ could be an explanation for the potential inhibition of oral NAR treatment on physical performance of rats [128,129,130].
NAD+ repletion improves MT activity and muscle stem cells (MuSCs) function and enhances life span in aged mice. In an experimental model of muscular dystrophy, NARS prevented the senescence of MuSC in the mdx (C57BL/10ScSn-Dmd(mdx)/J) mice [131,132]. Also, seeking treatment of MT myopathy, orally administered NAR was found to induce MT unfolded protein response, MT biogenesis in BAT and skeletal muscle, prevent abnormalities of MT ultrastructure, and prevent deletion of mtDNA [133]. In this regard, NARS increased the levels of NAD+ in liver and skeletal muscle, exercise capacity and MT respiration, alleviating the exercise intolerance in adenine nucleotide translocator 1 (ANT1)-deficient mice [134]. In the recent study on 20 body mass index (BMI)-discordant monozygotic twin pairs, NARS (250 mg to 1 g daily for five months) improved muscle MT biogenesis, satellite cell differentiation, and gut microbiota [135].
NAD+ boosting was preclinically characterized as a promising therapeutic approach for rheumatoid arthritis (RA) patients. NAR and NMN treatments were ex vivo tested on RA–PBMCs, thus highlighting NAD+ amount increasing via NAMPT and nicotinamide mononucleotide adenylyl transferase (NMNAT), and reducing the pro-inflammatory, pro-oxidative and pro-apoptosis in RA patients [136].
The influence of NAR (250 mM, 500 mM, 1 M) on the development and growth of pectoralis major muscle (PMM) was examined on fertilized Cobb 500 broiler eggs. NARS introduced by injection into the yolk and albumen influenced in ovo broiler myogenesis in a fashion correlated with the increasing of PMM weight, length, and fiber density [137,138,139].
3. Nicotinamide Riboside as a Tool to Mitigate Metabolic Disorders
3.1. Obesity
Among NAD+ precursors, NAR is often suggested as a dietary supplement to enhance oxidative metabolism and protect against HFD-induced obesity and deficient MT functionality for age-related diseases [25,140]. In aging mice, NAD+ supplementation via NAR rejuvenated intestinal stem cells (ISCs) improved the repairing capacity of gut damages by increasing the activity of SIRT1 and mammalian target of rapamycin complex 1 (mTORC1) [141]. In experimental models of mildly obese male mice (C57BL/6N, C57BL/6J), NARS for eight weeks increased MT respiration. However, it had minimal impact on energy metabolism, without influencing b.w., internal organs weight, glucose metabolism, liver lipids amount, and metabolic adaptability [142].
NAR- and NMN-conditioned microbiota reduced HFD-induced weight gain in C57BL/6J male mice by increasing their energy consumption. Here, butyrate-producing Firmicutes were enriched by FMT from NAR-supplemented donors to HFD fed naïve mice [143,144]. Alterations in intestinal brush border membrane functionality and bacterial populations following intra-amniotic administration (to Gallus gallus) were reported for 30 mg/mL doses of NAR and its derivatives, water-soluble NAR tributyrate chloride (NARTBC) and oil-soluble NAR trioleate chloride (NARTOC). Bifidobacterium, Clostridium, Lactobacillus, and Escherichia coli populations were significantly increased by NAR treatment [145]. Further, oral supplementation of NAR for 12 weeks alters gut microbial composition (Erysipelotrichaceae, Lachnospiraceae and Ruminococcaceae families) in HFD-fed rats and mice, but not in humans [146].
NARS exerts an anti-obesity effect and prevents inflammation and liver fibrosis (LF) in white adipose tissue (WAT) of eight weeks (young) and 16 weeks (old) female C57BL/6J DIO mice fed an HFD/high-sucrose diet/high-cholesterol diet or HFD combined with 400 mg/kg/day NAR for 20 weeks. Weight and size of gonadal WAT (gWAT) adipocytes of old mice females were decreased by NARS [147]. Similar results were seen in recent research concerning NAR and CR effects on gut microbiota and liver inflammatory and morphological markers in CAF DIO in adult male Wistar rats [148]. The effect of NAR on lipid metabolism and gut microflora–bile acid axis was studied in alcohol-exposed mice. NARS increased the NAD+/NADH ratio, deoxycholic acid and hyocholic acid levels, and decreased the activation of the protein phosphatase 1 (PP1) signaling pathway, chenodeoxycholic acid, TG and total bile acid levels, and lipid accumulation [149].
Oral NARS also appeared to confer metabolic benefits in obese mice. Thus, NAR induces a thermogenic response in the brown adipose tissue (BAT) of lean mice. Over five weeks, male C57BL/67 mice were supplemented with NAR (400 mg/kg/day), giving a reduction of abdominal visceral fat depots and an increase in body temperature [150,151]. Also, compared with controls, NARS conferred marginal metabolic benefits, augmenting MT functionality in C57BL/6NJ obese mice without remodeling the skeletal muscle acetyl-proteome [152]. NARS given daily to suckling male mice improved lipid and energy metabolism in skeletal muscle and liver in adulthood. Correlated with an upregulation of SIRT1 and AMP-dependent protein kinase signaling pathways [153]. In obese rats under CR, NARS (400 mg/kg for four weeks) neutralized hypothalamic inflammation by reverting high levels of TNF-α and increases weight loss without altering skeletal muscle mass [154]. Further, high doses (9 g/kg diet) of NARS for 18 weeks induced glucose intolerance and WAT dysfunction in male C57BL/6JRccHsd mice fed a mildly obesogenic diet (MOD) containing low but adequate tryptophan [155,156,157].
In a HFD mouse model, a reduction of obesity and insulin resistance was seen by dual targeting of visceral adipose tissue (VAT) and BAT using a novel combination of metabolic cofactors (NAR, N-acetylcysteine, betaine, L-carnitine) orally administered for four weeks [158]. NARS (400 mg/kg daily) ameliorates high-fructose-induced lipid metabolism disorder in C57BL/6J mice via improving fibroblast growth factor 21 (FGF21) resistance in the liver and WAT. NAR treatment upregulated the SIRT1–NF-κB pathway, decreasing inflammatory processes and increasing NAD+/NADH ratios [159]. Another study concluded that DNA methylation changes are associated with the programming of WAT browning features by RSV and NAR neonatal supplementations in mice [160]. A positive signal was seen with a combined treatment of L-carnitine (0.4%, w/w) and NAR (0.3%, w/w). This evidently improved hepatic metabolism and attenuated obesity and liver steatosis in HFD-fed Ldlr–/– Leiden mice [161].
As a mechanism to treat metabolic disorders, NAR may shift the differentiation of human primary white adipocytes to beige adipocytes. This may impact substrate preference and uncouple respiration through SIRT1 activation and MT-derived reactive species production [162]. In a randomized, double blinded, PLA-controlled, crossover study in human volunteers (45–65 years, BMI 27–35 kg/m2), NAR (1 g daily for six weeks) enhanced in vitro beta-adrenergic BAT activity and did not alter cold-induced thermogenesis [163]. A randomized, PLA-controlled, double-blind, clinical trial in 40 healthy obese (BMI>30 kg/m2) insulin-resistant men looked at safety, insulin-sensitivity, and lipid-mobilizing effects of NAR (1000 mg twice daily) dietary supplementation for 12 weeks. It was found that NARS was safe and well tolerated, but did not improve either endogenous glucose generation, oxidation, or elimination, and did not improve insulin sensitivity [2]. Further, similar impacts of NARS were seen on endocrine pancreatic function and incretin hormones in non-diabetic men with obesity [164]. In a follow up study by the same authors, NARS does not alter MT respiration, content, or morphology in skeletal muscle in middle-aged, obese, and insulin-resistant men [165].
3.2. Nicotinamide Riboside and Diabetes
NARS can be used to mitigate type 2 diabetes (T2D) and neuropathy in HFD-fed prediabetic and diabetic male C57BL/6J mice. NAR treatment significantly restored liver NADP+ and NADPH amounts and reduced weight gain, blood glucose and HS [166]. NAD+ precursors (NMN, NAR) improved MT function in diabetes and prevented experimental diabetic peripheral neuropathy (DPN). The experiments were carried out in streptozotocin (STZ)-induced diabetic rats or mice. NMN was administered for two months (50 or 100 mg/kg b.w. i.p. injection on alternate days). HFD-fed mice were supplemented by NAR at 150 or 300 mg/kg for two months. NAR treatment in HFD-fed mice led to the normalization of adult dorsal root ganglion (DRG) neurons’ functionality [167]. From the point of view of cellular and molecular mechanisms, a recent study in NRK1 KO mice showed that NRK1 protected against HFD diet and age-induced pancreatic β-cell failure [168]. Also, NARS promoted mitofusin 2 (MFN2)-mediated MT fusion in diabetic hearts through the SIRT1–PGC-1α–peroxisome proliferator-activated receptor alpha (PPARα) pathway [169].
The effects appear to be broad. Supplementation with NAR reduces brain inflammation and improves cognitive function in diabetic mice. NAR treatment for six weeks of HFD-fed diabetic Institute of Cancer Research (ICR) male mice significantly reduced amyloidogenesis (Aβ precursor protein, presenilin 1) and neuroinflammation (IL-1, IL-6, and TNF-α markers) [10]. In STZ-induced diabetic rats, i.p. administered NAR every 48 hours for six weeks improved enteric neuropathy through myenteric plexus neuroprotection [170].
Other benefits of NARS in experimental murine diabetes refer mainly to the (i) alleviation of hepatic metaflammation by modulating NLRP3 inflammasome in a rodent (8-week-old KK/HlJ male mice) model of T2D [171], (ii) enhancement of endothelial progenitor cell (EPC) function to promote refractory healing of diabetic wounds through mediating the SIRT1–AMP-activated protein kinase (AMPK) pathway [172], and (iii) improvement of fetal growth under hypoglycemia previously induced in pregnant mice [173].
4. Nicotinamide Riboside for Healthy Aging and Longevity
4.1. Nicotinamide Riboside for Healthy Aging
NAD+ metabolism/homeostasis is important in aging and disease [174,175]. Declining NAD+ levels induce the augmentation of HIF-1α and a pseudohypoxic state disrupting PGC-1α/-1β-independent nuclear–MT communication during aging [176].
Thus, in recent preclinical studies, managing NAD+ deficits with NA, NAc, NAR, and NMN provided a healthy aging [177]. In Caco-2 cell line and NIAAA mouse model, NAD+ supplementation by NAR alleviated intestinal barrier injury induced by ethanol via protecting epithelial MT function [178]. NARS increased the formation of human leukocytes from hCD34+ progenitors in immunodeficient mice model. Thus, through increased MT clearance, NAR potently stimulated the hematopoiesis [179] and the lymphoid potential of Atm–/– and old mice HSCs [180,181]. Moreover, NMN can be used to activate SIRT1 and improve the pathophysiology of diet- and age-induced diabetes in mice [182].
Likewise, reduced NMN is a new and potent NAD+ precursor in mammalian cells and mice blood and internal organs [183]. In a randomized, double-blind, PLA-controlled, clinical trials in healthy middle-aged and older adults, chronic NARS (500 mg twice daily) is safe and well tolerated, and it is recommended for (i) increasing NAD+ level [8] and (ii) treating elevated systolic blood pressure (SBP) and arterial stiffness having initial above-normal (120–159 mmHg) SBP [184].
In aging and disease, a close connection relates NAD+ levels and the activation of SIRTs, as seen in murine models: (i) NAR alleviates cisplatin (CPT)-induced peripheral neuropathy (CIPN) and neuronal death via SIRT2 activation and enhancement of nucleotide excision repair in Lewis lung carcinoma model [185]; (ii) SIRT3 is required for regeneration of WT or mutant liver but not for the beneficial effect of NAR [186]; (iii) SIRT3 deficiency aggravates contrast-induced acute kidney injury (CIAKI) in vitro (HK-2 cells) and in vivo (WT and SIRT3 KO mice) [187]; (iv) NAR attenuates inflammation and OXS by activation of SIRT1 and normalization of NAD+/NADH ratio in alcohol-stimulated RAW 264.7 macrophages and in mouse BM-derived macrophages [188].
NARS could be a missing piece in the puzzle of exercise therapy for older adults. Short-term (ST) oral NAR treatment, 300 mg/kg or 600 mg/kg daily, improves muscle quality and function in middle-aged male C57BL/6J mice and increases cellular energetics and differentiating capacity of myogenic progenitors [189]. A randomized, PLA-controlled trial of safe and well-tolerated daily NARPT supplementation (1 g NAR and 0.2 g PT) improved skeletal muscle regeneration after experimental muscle injury in 23 elderly individuals (55–80 years) [190]. Moreover, in a double-blind, cross-over study, acute NARS improved redox homeostasis and exercise performance in 12 old individuals compared with the same number of young men [191].
Recent studies have highlighted that senotherapeutic NARS could contribute to healthy aging and longevity. As a sample of results: (i) the effects of senolytic drugs, including NAR, were tested on human mesenchymal stromal cells (MSCs) with no significant action on molecular markers for replicative senescence [192]; (ii) the senotherapeutic NAR triflate improved the NAD+ levels of buffy coat-derived platelet concentrates, but cannot prevented storage lesion for 23 days [193]; (iii) 17-α-estradiol late in life extends lifespan in aging UM-HET3 heterogeneous male mice, but NARS does not affect lifespan in either sex [194].
In this regard, NAD+ supplementation (NAR, NA, NAc) exhibited emerging roles in replicative and chronological aging in fungi and mammals. For example: (i) Saccharomyces cerevisiae YOR071C gene encodes the high affinity NAR transporter Nrt1 polypeptide [195]; (ii) S. cerevisiae unicellular organism probably represents one of the most recognized experimental aging models for the study of replicative lifespan (RLS, proliferating cells) and chronological lifespan (CLS, non-proliferating cells) [196,197,198].
4.2. Brain Aging, Cognitive Impairment and Neurodegenerative Diseases
Supplements of NAD+ precursors’ (NAR, NARH, NMN) are a potential way to prevent cognitive decline within aging-associated diseases, such as neurodegenerative disorders [199,200,201]. For example, in models of murine dementia, NARS decreased neuroinflammation, DNA damage, and apoptosis, while contributing to maintaining synaptic plasticity, integrity of blood-brain barrier (BBB) and gut microbiota functionality. It also improved hippocampal synaptic plasticity, learning and memory in Alzheimer’s disease (AD). Amyloid beta (Aβ) forming in the brain can be prevented by NARS partly through the upregulation of ubiquitination and proteasomal degradation of PGC-1α-mediated beta-secretase 1 (BACE1) [202,203].
These studies have extended for over a decade. For example, in Tg2576 mice, NAR dietary supplementation (250 mg/kg/day) for three months significantly alleviated cognitive damage, increased the NAD+/NADH ratio in the cerebral cortex and reduced Aβ production [26]. A further study examines DNA repair-deficient 3xTgAD/Polβ+/– mice that exhibited cognitive impairment, synaptic dysfunction, phosphorylated tau (p-tau) pathologies, and neuronal death, the main characteristics of human AD. Here, NARS supplements decreased neuroinflammation, DNA damage, and hippocampal neurons’ apoptosis, increased SIRT3 activity in the brain, improved cognitive function, and restored the hippocampal synaptic plasticity [204]. NARS supplements (2.5 g/kg in food for three months) in APP/PS1 transgenic AD and aged mice inhibited serum NAMPT elevation, astrocyte activation, neuroinflammation, senescence, Aβ accumulation, and astrocyte migration to Aβ, as well as improving locomotor activity, cognitive function, behavior, and dementia progression [205,206]. Inhibition of CD38 and NARS alleviated LPS-induced microglial and astrocytic neuroinflammation/neurodegeneration by increasing NAD+ levels in mice brain and by suppressing of NF-κB signaling pathway at the microglia level [207]. In APP/PS1 transgenic (AD) mice, NARS for eight weeks normalized gut dysbiosis for Adlercreutzia, Akkermansia, Bacteroides, Bifidobacterium, Butyricicoccus, Desulfovibrio, Lactobacillus, Olsenella, and Oscillospira microbiota species [208]. In late-onset AD (LOAD) patients, NAR and caffeine co-administration partially restores diminished NAD+ availability but did not alter bioenergetic metabolism [209]. In a randomized, double-blinded, PLA-controlled, phase II clinical trial, combined metabolic activators (CMAs) administered in a single dose during the first 28 days and twice daily between days 28–84 significantly increased the cognitive capacity and alleviated NAD+ plasma levels and GSH metabolism of AD patients. CMA complex included 12.35 g L-serine (61.75%), 3.73 g L-carnitine tartrate (18.65%), 2.55 g N-acetyl-L-cysteine (12.75%), and 1 g NAR (5%) [210].
NAR food supplementation for 28 days rescues Ang II-induced cerebral small vessel disease (CSVD) in C57BL/6 mice. NARS significantly reduced glial activation, neuroinflammation, and white matter injury (WMI) that is associated with cognitive dysfunction. It also supported BBB integrity and vascular remodeling, and improved Ang II-induced CSVD [211].
Further, treatment with NAD+ precursor NAR rescues MT defects in induced pluripotent stem cells (iPSCs) and aging-associated dopaminergic neuronal loss and motor decline and Drosophila models of glucocerebrosidase (GBA)-related Parkinson’s disease (PD) [212]. In a randomized, PLA-controlled, phase I clinical trial in 30 newly diagnosed, treatment-naïve PD patients (NADPARK study), oral NARS 1 g for 30 days significantly increased the level of NAD+ and its related metabolites in the cerebrospinal fluid (CSF) and decreased the inflammatory cytokines amounts also in serum and CSF [213]. In a recent randomized, PLA-controlled, crossover trial in 22 healthy older adults, oral NARS (500 mg, twice daily, six weeks), the levels of NAD+ in neuronal-origin enriched plasma extracellular vesicles (NEVs) were increased, and levels of kinases (Aβ42, pJNK, pERK1/2) implicated in neuroinflammation and insulin resistance pathways were inhibited [214]. Moreover, in a double-blind, PLA-controlled trial of 29 PD patients, including cases with common pathogenic mutations in the methylenetetrahydrofolate reductase (MTHFR) gene, it was found that high-dose NARS for 30 days was not associated with altered DNA methylation homeostasis [215].
4.3. Aging and Cancer
NAD+ metabolism is a major feature for cancer pathogenesis (tumorigenesis), being closely related to genome integrity provided by efficient redox homeostasis, MT metabolism and signal transduction [23]. The anti-tumoral effect of NAR was studied in experimental models of hepatocellular carcinoma (HCC), such as the subcutaneous transplantation of tumor in Balb/c nude mice (xenograft) and C57BL/6J mice (allograft), and hematogenous metastatic tumor in nude mice. Daily administration of NAR (400 mg/kg) by oral gavage extended the overall survival of HCC mice and decreased the size of allografted tumors and lung, liver, and bone metastases. Also, in vitro TGF-β-induced migration/invasion of HepG2 cells was inhibited by NARS [216].
In a mouse model of C26 adenocarcinoma, NAR pellet dietary supplementation significantly improved cancer cachexia and inflammation through the inhibition of specific molecular markers, such as IL-6, TNF-α, PCG-1α, and muscle-specific ubiquitin-proteasome ligases (e.g., mitofusin-2, atrogin-1, muscle RING-finger protein-1 (MuRF-1)) [217]. Oral administration of 200 mg/kg NAR in female tumor-bearing rats, in a preclinical model of mammary gland cancer induced by N-methyl-nitrosourea (MNU) and previously treated with PTX i.v. injections (three doses of 6.6 mg/kg) led to the decreasing of tumor growth and the Ki67 index of tumoral cells, and to the improvement of peripheral neuropathy symptoms [218]. In high-risk skin cancer patients, boosting NAD+ with NAR, MNM and NA p.o. supplements decreased the incidence of keratinocyte carcinoma (KC). This presumably occurred through its cellular protective effects mainly targeting DNA repair and prevention of possible activation of oncogenic mutations [219].
5. Safety and Bioavailability of Nicotinamide Riboside
5.1. Nicotinamide Riboside Safety
The safety of NAR chloride (NARC) was assessed by biological assays, including bacterial reverse mutagenesis tests (Ames test), in vitro chromosome aberration test, in vivo micronucleus tests, and toxicological studies in rats. These covered acute (24-hour), sub-acute (14-day) and chronic (90-day) administration. Genotoxicity of NARC was not observed, and lethality was absent even at a dose of 5000 mg/kg (p.o.). NARC had the lowest-observed-adverse-effect level (LOAEL) and the no-observed-adverse-effect level (NOAEL) at the dose of 1000 mg/kg/day and 300 mg/kg/day, respectively. Here, liver, kidney, ovary, and testicles were the target organs for oral toxicity assays [220].
A randomized, double-blind, PLA-controlled clinical trial for eight weeks was performed in overweight healthy women and men who received different doses of NARC (orally 100 mg, 300 mg, 1000 mg). Between NARC and PLA groups or between different NARC groups neither important differences nor adverse effect were observed. However, within two weeks of NARC supplementation, significant increases in serum levels of NAD+ and NAD+ metabolites were recorded [3].
In a randomized, double-blind, PLA-controlled study, the safety and efficacy of NARPT (Basis™) was assessed for eight weeks in 120 healthy adult volunteers (60–80 years old). In addition to the significant increase of NAD+ blood levels, repeated doses of NARPT supplementation did not cause any adverse effects [221]. The European Food Safety Authority (EFSA) Panel on Nutrition, Novel Foods and Allergens (NDA) of the European Commission (EC) concluded that NAR and NARC are “safe for the healthy adult population, excluding pregnant and lactating women, and that an intake of up to 230 mg/day is safe for pregnant and lactating women”. Moreover, for the two active compounds, there are no concerns about genotoxicity and their safety/tolerability in human studies [222,223,224]. Three more international leading regulatory authorities, including the US Food and Drug Administration (FDA), Health Canada (HC), and the Therapeutic Goods Administration (TGA) of Australia concluded that NAR is safer for human use than other NAD+ metabolites [225].
Similar studies were performed on NAR-related compounds. A non-randomized study of single oral supplementation within the daily tolerable upper levels of NA affects blood NA and NAD+ amounts in six healthy humans. NA daily intake up to 200 mg found, at 0.5 hours, a 30-fold increase of its maximum amount in the whole blood of male volunteers. At 12 hours, NAD+ serum concentrations have reached the maximum level. Metabolomic variations due to NA dietary supplementation lasted one day and then after two days they returned to the baseline [226].
Moreover, in another study, oral supplementation of NA (500 mg) significantly increased blood NAD+ levels after 12 hours in a cohort of five healthy adult subjects [227]. A toxicological study assessed the safety of Restorin® NMN, a direct NAD+ precursor (β-NMN) in a high purity formulation, by oral administration at 500, 1000, and 2000 mg/kg/day in Sprague–Dawley rats over three months followed by a recovery period of two weeks. No adverse effects were observed at the dose of 500 mg/kg/day. Restorin® NMN has a LOAEL of 2000 mg/kg/day and a NOAEL of 500 mg/kg/day in male rats and 1000 mg/kg/day in female rats [228].
In a double blind, PLA-controlled, randomized clinical trial, oral administration of NMN (250 mg/day for 12 weeks) proved to be safe with increasing the level of blood NAD+ in 30 healthy volunteers. A significant increase in the level of nicotinic acid mononucleotide (NAcMN) rather than in NMN amount was also seen [229]. MIB-626, an oral formulation of a microcrystalline unique polymorph of β-NMN, increased circulating NAD+ levels in a double-blind, PLA-controlled trial in 32 overweight or obese middle-aged and older adults (55–80 years). Once or twice daily 1000 mg MIB-626 supplementation proved to be safe and well-tolerated. Age, gender or BMI did not influence the variations in serum amounts of NMN or NAD+ [230].
5.2. Nicotinamide Riboside Bioavailability
In mice and humans, NAR is the most orally bioavailable of NAD+precursors. Amounts of NAD+ in mouse liver were elevated by oral NARS with better pharmacokinetics (PK) than seen with NAc and NA. In one healthy volunteer, a pilot study showed that a single dose of oral NARS elevated serum NAD+ levels as much as 2.7-fold. The first clinical trial of NAR PK in healthy humans highlighted that 100, 300 and 1000 mg of NAR single dose increased the NAD+ metabolome in a dose-dependent manner [231].
An open-label, non-randomized study of the PK of the NAR nutritional supplement and its effects on blood NAD+ levels was performed in eight healthy volunteers. Oral NAR (250 mg) was administered daily on the first two days. Then, the maximum dose was increased to 1000 mg twice daily on days 7 and 8. NARS did not induce adverse effects and was well tolerated. In NAR and NAD+ levels, starting from baseline to the ninth day, the absolute changes were highly correlated [232,233].
BM stromal cell antigen 1 (BST1) regulates NAR metabolism via its dual roles in glycohydrolase and base-exchange enzyme activities. Two different pathways contribute to the increasing of NAD+ level after oral NAR treatment: (i) NAR salvage pathway in early phase of NAR direct absorption, and (ii) Preiss–Handler pathway in the late phase of NAR hydrolysis to NA by BST1 followed by transformation to NAc by the distal gut microbiome [234,235].
Pre-steady state and steady state kinetic analyses were made for the N-ribosyl hydrolase activity of hCD157. Cell-surface hCD157 binds and catalyzes the slow hydrolysis of NAR and NAcR after the formation of an initial complex with NAD+ derivatives [236].
The import of NAR and NAcR into human cells is mediated by the equilibrative nucleoside transporter (ENT) family of proteins, ENT1, ENT2, and ENT4. After entering the HEK293 cells, NAR is activated by NRK phosphorylation in the cytosol and then transformed to give NA [237]. NMN and NAR metabolism in mammalian cells is predominantly controlled by NRK1 and purine nucleoside phosphorylase (PNP). Simultaneous PNP downregulation could improve the health advantages of NARS [14,238].
Kulikova et al. (2015) studied the generation, release, and uptake of the NAD+ precursor NAcR by human cells. Under normal culture conditions, untransfected HeLa cells generated and released a sufficient level of NAcR and NAR as whole compounds derived from NAD+ metabolism [239].
A pilot study investigated changes in the human plasma and urine NAD+ metabolome during a 6-hour i.v. infusion of 3 μM/min NAD+. No changes were seen after 2 hours in the levels of plasma NAD+ or itsmetabolites, such as NA, MeNA, adenosine diphosphate ribose (ADPR) and NMN. After six hours, an increased urinary excretion of NAD+ and MeNA was seen; at the same time, NA urinary level was not increased [240].
In a simulated intestinal fluid using porcine pancreatin, NAR trioleate chloride (NARTOCl), a new synthesized hydrophobic NAR derivative, was partly digestible and easily released NAR [241].
NARH is a potent enhancer of NAD+ levels in vitro in mammalian cells, significantly increasing NAD+/NADH ratio, as well as in vivo after i.p. injection to C57BL/6J mice [242]. Although there are minor structural dissimilarities, NARH defines a new pathway for NAD+ biosynthesis, independent of NRK1, and acts as an orally bioavailable NAD+ precursor also in mammalian cells and mouse tissues [243]. In a recent study, it was found that NAR and dihydronicotinic acid riboside synergistically act to increase NAD+ intracellular level by generating NARH [244].
6. Borate-Stabilized Nicotinamide Riboside
This collection of research results is cited to encourage the consumption of NAR as a dietary supplement. Indeed, several companies now offer NAR in pill form for this purpose. Unfortunately, NAR is a reactive species, often unstable during manufacture, transport, and storage. Accordingly, HPLC analysis of many of the commercially available products found them to not be pure NAR, but rather mixtures of NAR and various other materials. Often, the NAR is present in small amounts.
These facts led us to develop NAR borate, and less expensive routes to make it. These are reviewed below.
6.1. Prebiotic Synthesis
While this is irrelevant to the use of NAR as a dietary supplement, it is of conceptual interest to note that the synthesis of NAR-borate was grounded in considerations of prebiotic chemistry. The analysis began with the observation that the chemical and biological syntheses of NAR are extremely challenging, with the challenge centered on the bond that joins the nicotinamide ring to the ribose ring.
Thus, while several methods of chemical and biological syntheses of NAR are reported [245], these are rather expensive. At the time of this writing, only one United States company (ChromaDex) produces NARC [3,220].
Further, NAR is itself unstable. When analyzed by HPLC and other standard analytical methods, many samples of commercial NAR are in fact mixtures of products that indicate decomposition of the material [246].
Prebiotic chemistry was applied to this problem. Here, Kim & Benner discovered that NAR phosphate emerges in stable form by direct reaction of ribose-1,2-cyclic phosphate with nicotinamide [247,248]. Ribose-1,2-cyclic phosphate is available inexpensively from ribose and amidotriphosphate, which is available from the very inexpensive cyclic trimetaphosphate and ammonia. The phosphorylated NAR product is then enzymatically dephosphorylated by a phosphatase [249]. In the final stage, NAR is stabilized by borate, resulting NAR borate (NARB) [250].
Borate-stabilized NAR was found to have a high stability against thermal and chemical degradation [250,251]. This mitigates problems in currently commercial NAR.
The borate might be seen as simply a stabilizing species. However, recent research suggests that borate itself might be a key element in NAD+ metabolism [252,253]. Borate released by the ribose–borate complex by hydrolysis could be a reservoir of physiological boron (B). In addition, borate esters can remain undigested in the upper gastric system and thus be more accessible to the microbiota [254]. According to new insights into the essentiality of B in the healthy symbiosis between the microbiota and the human host, NARB becomes an interesting prebiotic that delivers B that is essential to bacteria for the synthesis of autoinducer-2 (AI-2) and for strengthening the colonic mucus [254,255,256].
The prebiotic synthesis and borate stabilization of NAR had two very important advantages: (i) it is more cost effective due to less expensive starting materials; (ii) NAR is more stable, mitigating the fact that many samples of commercial NARC are mixtures of products that indicate decomposition of the material (Figure 1).
6.2. Solubility and Degradation Kinetics of Nicotinamide Riboside Borate
The possibility that NARB provides, at last, a stable form of NAR should motivate the study of its effect in pre-clinical and clinical studies. Since borate esters are dynamic, with borate dissociating from NAR upon dilution, we expect that the effects of NARB should closely parallel the effects of NAR.
The solubility and degradation kinetics of NARB were examined in various physiological media, including 0.1 N hydrochloric acid (pH 1.5), sodium acetate buffer (pH 5.0), water (pH 7.0), and phosphate-buffered saline (pH 7.4), using HPLC analysis. Furthermore, stability studies were conducted to investigate how different pH levels and temperatures influenced the degradation kinetics of NARB. The results revealed significant variations in the solubility and stability of NARB within the tested biological solutions. The solubility of NARB was found to be pH-dependent, increasing as the pH level rose [257].
Using the shake-flask method at room temperature, the solubility of NARB was evaluated. The measurements showed that NARB exhibited solubilities of 1972.7±15.4 mg/mL, 1060.5±31.0 mg/mL, and 926.0±34.4 mg/mL at pH 1.5, 5.0, and 7.4, respectively. These findings indicate that NARB demonstrated high solubility in all three solutions with varying pH levels, meeting the standards outlined in the US Pharmacopoeia recommendations [257] (Figure 2).
To investigate the spontaneity of NARB degradation in various pH solutions, the Gibbs free energy (ΔG) was calculated. The ΔG value obtained for NARB was 2.43 kcal/mol [257].
When examining NARB as a nutritional supplement, it is important to investigate the generation of NA and the ribose–borate complex, which acts as a physiological reservoir of B, as degradation byproducts. It is understood that the existence of anionic borate can impede the conversion of NAR into NA and trigger the activation of SIRT. While there is limited scientific data on slight degradation of NAR in simulated gastric fluids, substantial degradation seems to occur in the intestine due to its elevated pH levels [233]. The stability of NARB is significantly influenced by neutral conditions, exemplified by the pH 7.4 employed in this study. In the Arrhenius model of NARB degradation, the rate of degradation is higher at pH 7.4 compared to pH 1.5 and pH 5.0, supporting the notion that the degradation of NARB is influenced by the concentration of HO- ions in the solution. It is crucial to exercise caution when using an aqueous solution of NARB, particularly at high pH values where an excess of HO- ions can accelerate the degradation process due to hydrolysis. Our findings suggest that there may be a slightly distinct mechanism of degradation for NAR and NARB under acidic conditions compared to neutral or alkaline conditions. Consequently, this model can serve as a general reference for comprehending the stability of NARB in various buffered solutions. Additionally, temperature plays a role in the content of NARB in solution, with a 10°C increase approximately doubling its degradation rate under any pH condition.
The estimation of logP is crucial for predicting the permeability of NARB. It is well-established that compounds with estimated logP values below 1.72 exhibit low permeability. According to the Biopharmaceutical Classification System (BCS), NARB is classified as Class III due to its high solubility but anticipated low permeability, as indicated by a logP value of -4.17. This physiological characteristic may be linked to the proportion of released degradation products. Ultra-high-performance liquid chromatography (UHPLC) spectra have revealed that the degradation products of NARB in aqueous solutions are NA and the ribose–borate complex. When NARB degrades, each molecule gives rise to one molecule of NAM and one molecule of ribose–borate. Consequently, the concentration of NA can serve as a marker for the degradation of both NARB and NARC, the commercially available form. In this study, a developed high-performance thin-layer chromatography (HPTLC) method was employed to separate and quantify NAR and NA peaks with a remarkable separation resolution (R=1.8), surpassing the acceptable resolution (R>1) recommended by the US Food and Drug Administration [257]. This method offers a rapid means of detecting NAR, its degradation product NA, and the ribose–borate complex. Our research data presents, for the first time, a definitive upper temperature limit for the processing of NARB and its products, which can impact the production of supplements at various stages. The optimization of NARB administration will have a significant influence on its efficacy. Our study aimed to uncover the mechanism by which NARB degradation produces a physiological product when exposed to simulated physiological fluids in vitro. The development of an industrial pilot NARB opens up possibilities for further research, which could have a profound impact on its potential therapeutic applications. Consequently, under these conditions, NARB solutions and formulations have the potential for high stability. The findings of this study will prove valuable for pharmaceutical scientists involved in manipulating NARB working solutions and designing appropriate formulations for NARB delivery. We observed a clear dependence of NARB solubility on pH. NARB has exhibited extreme instability at pH levels greater than 8.0, while remaining stable in both acidic and neutral environments.
As a result, NARB may offer advantages over NARC as a nutritional supplement, primarily due to the boric acid (BA) residue's ability to block the glycosidic bond between the pyrimidine base and ribose. The study has shed light on the key physicochemical properties that should be considered when utilizing NARB in both in vitro and in vivo experiments. The described HPTLC method has potential for distinguishing between NARB and its degradation products in environments with pH conditions similar to those found in the gastrointestinal (GI) tract. The obtained data also holds significance for the processing, production, and storage of this supplement.
In addition, the greatest direction of use of NARB could be as a prebiotic compound, the latest research shows that the presence of NAR in the colon has a very high nutritional value and through its ester form of BA it can become a nutraceutical with a role in healthy symbiosis as an essential B element as well as an essential nutrient in the health metabolism of the human microbiome [143,144].
7. Conclusions and Perspectives
NAD+ is known classically as a metabolite that stands astride both catabolic and anabolic pathways throughout the metabolism that is taught in introductory biochemistry courses. However, non-classical studies starting over a decade ago found that it is also involved in higher order functions, in part because of its involvement in the activation of SIRTs and the support of the MT unfolded protein response. Many studies have suggested that NAD+ is involved in an extensive spectrum of pathologies, including neurodegenerative disorders, cardiomyopathy, obesity, and diabetes. Further, healthy aging and longevity appear to be closely related to NAD+ and its related metabolites, including through NAR and NMN. This system appears to have prophylactic and therapeutic value improving age-associated neurodegenerative, CV, and metabolic diseases and conditions.
Accordingly, many are now recommending the consumption of materials in this system as dietary supplements, hoping to achieve overall improvements in human health. Among NAD+ precursors, nicotinamide ribose (NAR) appears to have special values. These include better tolerance, better uptake, and overall greater potency.
Unfortunately, NAR is a reactive molecule, often unstable during its manufacturing, transport, and storage. Indeed, HPLC analyses of many commercial samples of NAR shows that they contain substantial amounts of material that are not, in fact, NAR. More stable derivatives of NAR that are easily converted upon consumption into NAR are therefore desired.
Recently work related to prebiotic chemistry provided the borate derivative of NAR. NARB is considerably more stable than NAR itself. However, immediately upon consumption, the borate dissociates from NARB, and is lost in the body through dilution and binding to other species, notably carbohydrates such as fructose and glucose. The NAR left behind is expected to behave pharmacologically in ways identical to NAR itself.
This review provides a comprehensive uncritical summary through Q1 of 2023 of literature that makes the case for the consumption of NAR. It then summarizes the challenges of delivering quality NAR to consumers using standard synthesis, manufacture, shipping, and storage approaches. It concludes by outlining the advantages of NARB in these processes.
Author Contributions
Conceptualization, A.B., I.R.S., G.D.M. and S.A.B.; methodology, M.V.C., O.E.N., A.S.P., G.R. and C.B.; writing—original draft preparation, I.R.S., G.D.M. and L.E.B.; writing—review and editing, A.B., I.R.S., G.D.M. and S.A.B.; supervision, A.B., I.R.S., G.D.M. and S.A.B.; funding acquisition, A.B., I.R.S and J.N. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant of the Romanian Ministry of Education and Research, CNFIS–UMFCV, project number 2629c.
Institutional Review Board Statement
The manuscript does not contain experiments on laboratory animals.
Informed Consent Statement
The manuscript does not contain clinical studies or patient data.
Conflicts of Interest
The authors declare no conflict of interest. The funder had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Penberthy WT, Kirkland JB (2012) Chapter 19: Niacin. In: Erdman JW Jr, MacDonald IA, Zeisel SH (eds). Present knowledge in nutrition. 10th edition, International Life Sciences Institute (ILSI), Wiley–Blackwell, John Wiley & Sons, Ames, Iowa, USA, 293–306. [CrossRef]
- Dollerup OL, Christensen B, Svart M, Schmidt MS, Sulek K, Ringgaard S, Stødkilde-Jørgensen H, Møller N, Brenner C, Treebak JT, Jessen N (2018) A randomized placebo-controlled clinical trial of nicotinamide riboside in obese men: safety, insulin-sensitivity, and lipid-mobilizing effects. Am J Clin Nutr 108(2):343–353. [CrossRef]
- Conze D, Brenner C, Kruger CL (2019) Safety and metabolism of long-term administration of NIAGEN (nicotinamide riboside chloride) in a randomized, double-blind, placebo-controlled clinical trial of healthy overweight adults. Sci Rep 9(1):9772. [CrossRef]
- Mehmel M, Jovanović N, Spitz U (2020) Nicotinamide riboside – the current state of research and therapeutic uses. Nutrients 12(6):1616. [CrossRef]
- Gilbert W (1986) Origin of life: the RNA world. Nature 319(6055):618. [CrossRef]
- White HB 3rd (1976) Coenzymes as fossils of an earlier metabolic state. J Mol Evol 7(2):101–104. [CrossRef]
- Chi Y, Sauve AA (2013) Nicotinamide riboside, a trace nutrient in foods, is a vitamin B3 with effects on energy metabolism and neuroprotection. Curr Opin Clin Nutr Metab Care 16(6):657–661. [CrossRef]
- Martens CR, Denman BA, Mazzo MR, Armstrong ML, Reisdorph N, McQueen MB, Chonchol M, Seals DR (2018) Chronic nicotinamide riboside supplementation is well-tolerated and elevates NAD+ in healthy middle-aged and older adults. Nat Commun 9(1):1286. [CrossRef]
- Trammell SAJ, Yu L, Redpath P, Migaud ME, Brenner C (2016) Nicotinamide riboside is a major NAD+ precursor vitamin in cow milk. J Nutr 146(5):957–963. [CrossRef]
- Lee HJ, Yang SJ (2019) Supplementation with nicotinamide riboside reduces brain inflammation and improves cognitive function in diabetic mice. Int J Mol Sci 20(17):4196. [CrossRef]
- Bogan KL, Brenner C (2008) Nicotinic acid, nicotinamide, and nicotinamide riboside: a molecular evaluation of NAD+ precursor vitamins in human nutrition. Annu Rev Nutr 28:115–130. [CrossRef]
- Belenky P, Stebbins R, Bogan KL, Evans CR, Brenner C (2011) Nrt1 and Tna1-independent export of NAD+ precursor vitamins promotes NAD+ homeostasis and allows engineering of vitamin production. PLoS One 6(5):e19710. [CrossRef]
- Bieganowski P, Brenner C (2004) Discoveries of nicotinamide riboside as a nutrient and conserved NRK genes establish a Preiss–Handler independent route to NAD+ in fungi and humans. Cell 117(4):495–502. [CrossRef]
- Ratajczak J, Joffraud M, Trammell SA, Ras R, Canela N, Boutant M, Kulkarni SS, Rodrigues M, Redpath P, Migaud ME, Auwerx J, Yanes O, Brenner C, Cantó C (2016) NRK1 controls nicotinamide mononucleotide and nicotinamide riboside metabolism in mammalian cells. Nat Commun 7:13103. [CrossRef]
- Schmidt MS, Brenner C (2019) Absence of evidence that Slc12a8 encodes a nicotinamide mononucleotide transporter. Nat Metab 1(7):660–661. [CrossRef]
- Canto C (2022) NAD+ precursors: a questionable redundancy. Metabolites 12(7):630. [CrossRef]
- Cantó C, Auwerx J (2011) NAD+ as a signaling molecule modulating metabolism. Cold Spring Harb Symp Quant Biol 76:291–298. [CrossRef]
- Imai S, Guarente L (2014) NAD+ and sirtuins in aging and disease. Trends Cell Biol 24(8):464–471. [CrossRef]
- Zhang N, Sauve AA (2018) Regulatory effects of NAD+ metabolic pathways on sirtuin activity. Prog Mol Biol Transl Sci 154:71–104. [CrossRef]
- Okabe K, Yaku K, Tobe K, Nakagawa T (2019) Implications of altered NAD metabolism in metabolic disorders. J Biomed Sci 26(1):34. [CrossRef]
- Buonvicino D, Ranieri G, Pittelli M, Lapucci A, Bragliola S, Chiarugi A (2021) SIRT1-dependent restoration of NAD+ homeostasis after increased extracellular NAD+ exposure. J Biol Chem 297(1):100855. [CrossRef]
- Carpi FM, Cortese M, Orsomando G, Polzonetti V, Vincenzetti S, Moreschini B, Coleman M, Magni G, Pucciarelli S (2018) Simultaneous quantification of nicotinamide mononucleotide and related pyridine compounds in mouse tissues by UHPLC–MS/MS. Sep Sci Plus 1(1):22–30. [CrossRef]
- Demarest TG, Babbar M, Okur MN, Dan X, Croteau DL, Fakouri NB, Mattson MP, Bohr VA (2019) NAD+ metabolism in aging and cancer. Annu Rev Cancer Biol 3:105–130. [CrossRef]
- Yang T, Chan NYK, Sauve AA (2007) Syntheses of nicotinamide riboside and derivatives: effective agents for increasing nicotinamide adenine dinucleotide concentrations in mammalian cells. J Med Chem 50(26):6458–6461. [CrossRef]
- Cantó C, Houtkooper RH, Pirinen E, Youn DY, Oosterveer MH, Cen Y, Fernandez-Marcos PJ, Yamamoto H, Andreux PA, Cettour-Rose P, Gademann K, Rinsch C, Schoonjans K, Sauve AA, Auwerx J (2012) The NAD+ precursor nicotinamide riboside enhances oxidative metabolism and protects against high-fat diet-induced obesity. Cell Metab 15(6):838–847. [CrossRef]
- Gong B, Pan Y, Vempati P, Zhao W, Knable L, Ho L, Wang J, Sastre M, Ono K, Sauve AA, Pasinetti GM (2013) Nicotinamide riboside restores cognition through an upregulation of proliferator-activated receptor-γ coactivator 1α regulated β-secretase 1 degradation and mitochondrial gene expression in Alzheimer’s mouse models. Neurobiol Aging 34(6):1581–1588. [CrossRef]
- Cantó C, Menzies KJ, Auwerx J (2015) NAD+ metabolism and the control of energy homeostasis: a balancing act between mitochondria and the nucleus. Cell Metab 22(1):31–53. [CrossRef]
- Conlon N, Ford D (2022) A systems-approach to NAD+ restoration. Biochem Pharmacol 198:114946. [CrossRef]
- Sharma C, Donu D, Cen Y (2022) Emerging role of nicotinamide riboside in health and diseases. Nutrients 14(19):3889. [CrossRef]
- Helman T, Braidy N (2023) Importance of NAD+ anabolism in metabolic, cardiovascular and neurodegenerative disorders. Drugs Aging 40(1):33–48. [CrossRef]
- Fang J, Wu H, Zhang J, Mao S, Shi H, Yu D, Chen Z, Su K, Xing Y, Dong H, Shi H (2022) A reduced form of nicotinamide riboside protects the cochlea against aminoglycoside-induced ototoxicity by SIRT1 activation. Biomed Pharmacother 150:113071. [CrossRef]
- She J, Sheng R, Qin ZH (2021) Pharmacology and potential implications of nicotinamide adenine dinucleotide precursors. Aging Dis 12(8):1879–1897. [CrossRef]
- Belenky P, Bogan KL, Brenner C (2007) NAD+ metabolism in health and disease. Trends Biochem Sci 32(1):12–19. Erratum in: Trends Biochem Sci, 2008, 33(1):1. [CrossRef]
- Belenky P, Racette FG, Bogan KL, McClure JM, Smith JS, Brenner C (2007) Nicotinamide riboside promotes Sir2 silencing and extends lifespan via Nrk and Urh1/Pnp1/Meu1 pathways to NAD+. Cell 129(3):473–484. [CrossRef]
- Yoshino J, Baur JA, Imai SI (2018) NAD+ intermediates: the biology and therapeutic potential of NMN and NR. Cell Metab 27(3):513–528. [CrossRef]
- Stock AJ, Ayyar S, Kashyap A, Wang Y, Yanai H, Starost MF, Tanaka-Yano M, Bodogai M, Sun C, Wang Y, Gong Y, Puligilla C, Fang EF, Bohr VA, Liu Y, Beerman I (2023) Boosting NAD ameliorates hematopoietic impairment linked to short telomeres in vivo. Geroscience Feb 24. [CrossRef]
- Hong G, Zheng D, Zhang L, Ni R, Wang G, Fan GC, Lu Z, Peng T (2018) Administration of nicotinamide riboside prevents oxidative stress and organ injury in sepsis. Free Radic Biol Med 123:125–137. [CrossRef]
- Wu J, Singh K, Lin A, Meadows AM, Wu K, Shing V, Bley M, Hassanzadeh S, Huffstutler RD, Schmidt MS, Blanco LP, Tian R, Brenner C, Pirooznia M, Kaplan MJ, Sack MN (2022) Boosting NAD+ blunts TLR4-induced type I IFN in control and systemic lupus erythematosus monocytes. J Clin Invest 132(5):e139828. [CrossRef]
- Zhang X, Tian B, Deng Q, Cao J, Ding X, Liu Q, Zhang Y, Ye C, Deng C, Qiu L, Guo C (2022) Nicotinamide riboside relieves the severity of experimental necrotizing enterocolitis by regulating endothelial function via eNOS deacetylation. Free Radic Biol Med 184:218–229. [CrossRef]
- Fan XK, Xu CQ, Cao KQ, Zhao GJ, Hong GL, Lu ZQ (2022) [Study on the protective effect and mechanism of nicotinamide riboside on lung injury in paraquat intoxicated mice]. Zhonghua Lao Dong Wei Sheng Zhi Ye Bing Za Zhi (Chin J Ind Hyg Occup Dis) 40(8):561–567. [CrossRef]
- Selli J, Vural Keles D, Keles ON, Celik M, Yetim Z (2023) Nicotinamide riboside preserves ovarian injury in experimental sepsis model in rats. Eurasian J Med Jan 17. [CrossRef]
- DiNicolantonio JJ, McCarty MF, O’Keefe JH (2022) Nutraceutical activation of Sirt1: a review. Open Heart 9(2):e002171. [CrossRef]
- Cheng YH, Zhao JH, Zong WF, Wei XJ, Xu Z, Yuan Y, Jiang YF, Luo X, Wang W, Qu WS (2022) Acute treatment with nicotinamide riboside chloride reduces hippocampal damage and preserves the cognitive function of mice with ischemic injury. Neurochem Res 47(8):2244–2253. [CrossRef]
- Lenherr N, Christodoulou J, Duley J, Dobritzsch D, Fairbanks L, Datta AN, Filges I, Gürtler N, Roelofsen J, van Kuilenburg ABP, Kemper C, West EE, Szinnai G, Huemer M (2021) Co-therapy with S-adenosylmethionine and nicotinamide riboside improves t-cell survival and function in Arts syndrome (PRPS1 deficiency). Mol Genet Metab Rep 26:100709. [CrossRef]
- Kodali M, Jankay T, Shetty AK, Reddy DS (2023) Pathophysiological basis and promise of experimental therapies for Gulf War Illness, a chronic neuropsychiatric syndrome in veterans. Psychopharmacology (Berl) 240(4):673–697. [CrossRef]
- Joshi U, Evans JE, Pearson A, Saltiel N, Cseresznye A, Darcey T, Ojo J, Keegan AP, Oberlin S, Mouzon B, Paris D, Klimas N, Sullivan K, Mullan M, Crawford F, Abdullah L (2020) Targeting sirtuin activity with nicotinamide riboside reduces neuroinflammation in a GWI mouse model. Neurotoxicology 79:84–94. [CrossRef]
- Jiang Y, Liu Y, Gao M, Xue M, Wang Z, Liang H (2020) Nicotinamide riboside alleviates alcohol-induced depression-like behaviours in C57BL/6J mice by altering the intestinal microbiota associated with microglial activation and BDNF expression. Food Funct 11(1):378–391. [CrossRef]
- Vaur P, Brugg B, Mericskay M, Li Z, Schmidt MS, Vivien D, Orset C, Jacotot E, Brenner C, Duplus E (2017) Nicotinamide riboside, a form of vitamin B3, protects against excitotoxicity-induced axonal degeneration. FASEB J 31(12):5440–5452. [CrossRef]
- Kitaoka Y, Sase K, Tsukahara C, Fujita N, Arizono I, Takagi H (2020) Axonal protection by nicotinamide riboside via SIRT1-autophagy pathway in TNF-induced optic nerve degeneration. Mol Neurobiol 57(12):4952–4960. [CrossRef]
- Chen A, Kristiansen CK, Hong Y, Kianian A, Fang EF, Sullivan GJ, Wang J, Li X, Bindoff LA, Liang KX (2021) Nicotinamide riboside and Metformin ameliorate mitophagy defect in induced pluripotent stem cell-derived astrocytes with POLG mutations. Front Cell Dev Biol 9:737304. [CrossRef]
- Sun C, Seranova E, Cohen MA, Chipara M, Roberts J, Astuti D, Palhegyi AM, Acharjee A, Sedlackova L, Kataura T, Otten EG, Panda PK, Lara-Reyna S, Korsgen ME, Kauffman KJ, Huerta-Uribe A, Zatyka M, Silva LFSE, Torresi J, Zhang S, Hughes GW, Ward C, Kuechler ER, Cartwright D, Trushin S, Trushina E, Sahay G, Buganim Y, Lavery GG, Gsponer J, Anderson DG, Frickel EM, Rosenstock TR, Barrett T, Maddocks ODK, Tennant DA, Wang H, Jaenisch R, Korolchuk VI, Sarkar S (2023) NAD depletion mediates cytotoxicity in human neurons with autophagy deficiency. Cell Rep 42(5):112372. [CrossRef]
- Yang B, Dan X, Hou Y, Lee JH, Wechter N, Krishnamurthy S, Kimura R, Babbar M, Demarest T, McDevitt R, Zhang S, Zhang Y, Mattson MP, Croteau DL, Bohr VA (2021) NAD+ supplementation prevents STING-induced senescence in ataxia telangiectasia by improving mitophagy. Aging Cell 20(4):e13329. [CrossRef]
- Veenhuis SJG, van Os NJH, Janssen AJWM, van Gerven MHJC, Coene KLM, Engelke UFH, Wevers RA, Tinnevelt GH, Ter Heine R, van de Warrenburg BPC, Weemaes CMR, Roeleveld N, Willemsen MAAP (2021) Nicotinamide riboside improves ataxia scores and immunoglobulin levels in ataxia telangiectasia. Mov Disord 36(12):2951–2957. [CrossRef]
- Steinbruecker K, Tiefenthaler E, Schernthaler EM, Jungwirth J, Wortmann SB (2022) Nicotinamide riboside for ataxia telangiectasia: report of an early treated individual. Neuropediatrics Oct 12. [CrossRef]
- Gerasimenko M, Cherepanov SM, Furuhara K, Lopatina O, Salmina AB, Shabalova AA, Tsuji C, Yokoyama S, Ishihara K, Brenner C, Higashida H (2020) Nicotinamide riboside supplementation corrects deficits in oxytocin, sociability and anxiety of CD157 mutants in a mouse model of autism spectrum disorder. Sci Rep 10(1):10035. [CrossRef]
- Gerasimenko M, Higashida H (2023) Remission of social behavior impairment by oral administration of a precursor of NAD in CD157, but not in CD38, knockout mice. Front Immunol 14:1166609. [CrossRef]
- Harlan BA, Killoy KM, Pehar M, Liu L, Auwerx J, Vargas MR (2020) Evaluation of the NAD+ biosynthetic pathway in ALS patients and effect of modulating NAD+ levels in hSOD1-linked ALS mouse models. Exp Neurol 327:113219. [CrossRef]
- Zhou Q, Zhu L, Qiu W, Liu Y, Yang F, Chen W, Xu R (2020) Nicotinamide riboside enhances mitochondrial proteostasis and adult neurogenesis through activation of mitochondrial unfolded protein response signaling in the brain of ALS SOD1G93A mice. Int J Biol Sci 16(2):284–297. Erratum in: Int J Biol Sci, 2022, 18(5):2181-2183. [CrossRef]
- Obrador E, Salvador R, Marchio P, López-Blanch R, Jihad-Jebbar A, Rivera P, Vallés SL, Banacloche S, Alcácer J, Colomer N, Coronado JA, Alandes S, Drehmer E, Benlloch M, Estrela JM (2021) Nicotinamide riboside and pterostilbene cooperatively delay motor neuron failure in ALS SOD1G93A mice. Mol Neurobiol 58(4):1345–1371. [CrossRef]
- Freeberg KA, Udovich CAC, Martens CR, Seals DR, Craighead DH (2023) Dietary supplementation with NAD+-boosting compounds in humans: current knowledge and future directions. J Gerontol A Biol Sci Med Sci Apr 17:glad106. [CrossRef]
- Brown KD, Maqsood S, Huang JY, Pan Y, Harkcom W, Li W, Sauve A, Verdin E, Jaffrey SR (2014) Activation of SIRT3 by the NAD⁺ precursor nicotinamide riboside protects from noise-induced hearing loss. Cell Metab 20(6):1059–1068. [CrossRef]
- Han S, Du Z, Liu K, Gong S (2020) Nicotinamide riboside protects noise-induced hearing loss by recovering the hair cell ribbon synapses. Neurosci Lett 725:134910. [CrossRef]
- Okur MN, Mao B, Kimura R, Haraczy S, Fitzgerald T, Edwards-Hollingsworth K, Tian J, Osmani W, Croteau DL, Kelley MW, Bohr VA (2020) Short-term NAD+ supplementation prevents hearing loss in mouse models of Cockayne syndrome. NPJ Aging Mech Dis 6:1. [CrossRef]
- Hamity MV, White SR, Walder RY, Schmidt MS, Brenner C, Hammond DL (2017) Nicotinamide riboside, a form of vitamin B3 and NAD+ precursor, relieves the nociceptive and aversive dimensions of paclitaxel-induced peripheral neuropathy in female rats. Pain 158(5):962–972. [CrossRef]
- Hamity MV, Kolker SJ, Hegarty DM, Blum C, Langmack L, Aicher SA, Hammond DL (2022) Nicotinamide riboside alleviates corneal and somatic hypersensitivity induced by paclitaxel in male rats. Invest Ophthalmol Vis Sci 63(1):38. [CrossRef]
- Pîrvu AS, Andrei AM, Stănciulescu EC, Baniţă IM, Pisoschi CG, Jurja S, Ciuluvica R (2021) NAD+ metabolism and retinal degeneration (Review). Exp Ther Med 22(1):670. [CrossRef]
- Zhang X, Zhang N, Chrenek MA, Girardot PE, Wang J, Sellers JT, Geisert EE, Brenner C, Nickerson JM, Boatright JH, Li Y (2021) Systemic treatment with nicotinamide riboside is protective in two mouse models of retinal ganglion cell damage. Pharmaceutics 13(6):893. [CrossRef]
- Zhang X, Henneman NF, Girardot PE, Sellers JT, Chrenek MA, Li Y, Wang J, Brenner C, Nickerson JM, Boatright JH (2020) Systemic treatment with nicotinamide riboside is protective in a mouse model of light-induced retinal degeneration. Invest Ophthalmol Vis Sci 61(10):47. [CrossRef]
- Leung CKS, Ren ST, Chan PPM, Wan KHN, Kam AKW, Lai GWK, Chiu VSM, Ko MWL, Yiu CKF, Yu MCY (2022) Nicotinamide riboside as a neuroprotective therapy for glaucoma: study protocol for a randomized, double-blind, placebo-control trial. Trials 23(1):45. Erratum in: Trials, 2022, 23(1):134. [CrossRef]
- Silva SGN, Occhiutto ML, Costa VP (2023) The use of nicotinamide and nicotinamide riboside as an adjunct therapy in the treatment of glaucoma. Eur J Ophthalmol Mar 14:11206721231161101. [CrossRef]
- Zhou B, Zhao G, Zhu Y, Chen X, Zhang N, Yang J, Lin H (2021) Protective effects of nicotinamide riboside on H2O2-induced oxidative damage in lens epithelial cells. Curr Eye Res 46(7):961–970. [CrossRef]
- Pang H, Jiang Y, Li J, Wang Y, Nie M, Xiao N, Wang S, Song Z, Ji F, Chang Y, Zheng Y, Yao K, Yao L, Li S, Li P, Song L, Lan X, Xu Z, Hu Z (2021) Aberrant NAD+ metabolism underlies Zika virus-induced microcephaly. Nat Metab 3(8):1109–1124. [CrossRef]
- Matasic DS, Brenner C, London B (2018) Emerging potential benefits of modulating NAD+ metabolism in cardiovascular disease. Am J Physiol Heart Circ Physiol 314(4):H839–H852. [CrossRef]
- Ahmad F, Tomar D, Aryal A C S, Elmoselhi AB, Thomas M, Elrod JW, Tilley DG, Force T (2020) Nicotinamide riboside kinase-2 alleviates ischemia-induced heart failure through P38 signaling. Biochim Biophys Acta Mol Basis Dis 1866(3):165609. [CrossRef]
- Blanco-Vaca F, Rotllan N, Canyelles M, Mauricio D, Escolà-Gil JC, Julve J (2022) NAD+-increasing strategies to improve cardiometabolic health? Front Endocrinol (Lausanne) 12:815565. [CrossRef]
- Liu Y, Huang Y, Xu C, An P, Luo Y, Jiao L, Luo J, Li Y (2022) Mitochondrial dysfunction and therapeutic perspectives in cardiovascular diseases. Int J Mol Sci 23(24):16053. [CrossRef]
- de Castro JM, Assumpção JAF, Stein DJ, Toledo RS, da Silva LS, Caumo W, Carraro CC, da Rosa Araujo AS, Torres ILS (2020) Nicotinamide riboside reduces cardiometabolic risk factors and modulates cardiac oxidative stress in obese Wistar rats under caloric restriction. Life Sci 263:118596. [CrossRef]
- Liu X, Zhang Y, Deng Y, Yang L, Ou W, Xie M, Ding L, Jiang C, Yu H, Li Q, Li T (2022a) Mitochondrial protein hyperacetylation underpins heart failure with preserved ejection fraction in mice. J Mol Cell Cardiol 165:76–85. [CrossRef]
- Ma S, Feng J, Lin X, Liu J, Tang Y, Nie S, Gong J, Wang L (2021) Nicotinamide riboside alleviates cardiac dysfunction and remodeling in pressure overload cardiac hypertrophy. Oxid Med Cell Longev 2021:5546867. [CrossRef]
- Zhang M, Weng H, Zheng J (2019) NAD+ repletion inhibits the endothelial-to-mesenchymal transition induced by TGF-β in endothelial cells through improving mitochondrial unfolded protein response. Int J Biochem Cell Biol 117:105635. [CrossRef]
- Pool L, Knops P, Manintveld OC, Brugts JJ, Theuns DAMJ, Brundel BJJM, de Groot NMS (2022) The HF-AF ENERGY Trial: nicotinamide riboside for the treatment of atrial fibrillation in heart failure patients. Cardiovasc Drugs Ther Oct 13. [CrossRef]
- Zhou B, Wang DD, Qiu Y, Airhart S, Liu Y, Stempien-Otero A, O’Brien KD, Tian R (2020) Boosting NAD level suppresses inflammatory activation of PBMCs in heart failure. J Clin Invest 130(11):6054–6063. [CrossRef]
- Wang DD, Airhart SE, Zhou B, Shireman LM, Jiang S, Melendez Rodriguez C, Kirkpatrick JN, Shen DD, Tian R, O’Brien KD (2022) Safety and tolerability of nicotinamide riboside in heart failure with reduced ejection fraction. JACC Basic Transl Sci 7(12):1183-1196. [CrossRef]
- Oller J, Gabandé-Rodríguez E, Roldan-Montero R, Ruiz-Rodríguez MJ, Redondo JM, Martín-Ventura JL, Mittelbrunn M (2022) Rewiring vascular metabolism prevents sudden death due to aortic ruptures – brief report. Arterioscler Thromb Vasc Biol 42(4):462–469. [CrossRef]
- Shahzadi SK, Marzook H, Qaisar R, Ahmad F (2022) Nicotinamide riboside kinase-2 inhibits JNK pathway and limits dilated cardiomyopathy in mice with chronic pressure overload. Clin Sci (Lond) 136(2):181–196. [CrossRef]
- Diguet N, Trammell SAJ, Tannous C, Deloux R, Piquereau J, Mougenot N, Gouge A, Gressette M, Manoury B, Blanc J, Breton M, Decaux JF, Lavery GG, Baczkó I, Zoll J, Garnier A, Li Z, Brenner C, Mericskay M (2018) Nicotinamide riboside preserves cardiac function in a mouse model of dilated cardiomyopathy. Circulation 137(21):2256–2273. Erratum in: Circulation, 2018, 137(21):e690. [CrossRef]
- Zheng D, Zhang Y, Zheng M, Cao T, Wang G, Zhang L, Ni R, Brockman J, Zhong H, Fan GC, Peng T (2019) Nicotinamide riboside promotes autolysosome clearance in preventing doxorubicin-induced cardiotoxicity. Clin Sci (Lond) 133(13):1505–1521. [CrossRef]
- Podyacheva E, Toropova Y (2021) Nicotinamide riboside for the prevention and treatment of doxorubicin cardiomyopathy. Opportunities and prospects. Nutrients 13(10):3435. [CrossRef]
- Podyacheva E, N Yu N, V A V, Mukhametdinova D, Goncharova I, Zelinskaya I, Sviridov E, Martynov M, Osipova S, Toropova Y (2022) Intravenous nicotinamide riboside administration has a cardioprotective effect in chronic doxorubicin-induced cardiomyopathy. Int J Mol Sci 23(21):13096. [CrossRef]
- Marzook H, Gupta A, Tomar D, Saleh MA, Patil K, Semreen MH, Hamoudi R, Soares NC, Qaisar R, Ahmad F (2023) Nicotinamide riboside kinase-2 regulates metabolic adaptation in the ischemic heart. J Mol Med (Berl) 101(3):311–326. [CrossRef]
- Nie H, Zhang Y, Yu H, Xiao H, Li T, Yang Q (2021) Oral delivery of carrier-free dual-drug nanocrystal self-assembled microspheres improved NAD+ bioavailability and attenuated cardiac ischemia/reperfusion injury in mice. Drug Deliv 28(1):433–444. [CrossRef]
- Xiao Y, Phelp P, Wang Q, Bakker D, Nederlof R, Hollmann MW, Zuurbier CJ (2021) Cardioprotecive properties of known agents in rat ischemia-reperfusion model under clinically relevant conditions: only the NAD precursor nicotinamide riboside reduces infarct size in presence of Fentanyl, Midazolam and Cangrelor, but not Propofol. Front Cardiovasc Med 8:712478. [CrossRef]
- Toropova YG, Pechnikova NA, Zelinskaya IA, Zhuravsky SG, Kornyushin OV, Gonchar AI, Ivkin DY, Leonova YV, Karev VE, Karabak IA (2018) Nicotinamide riboside has protective effects in a rat model of mesenteric ischaemia–reperfusion. Int J Exp Pathol 99(6):304–311. [CrossRef]
- Mateuszuk Ł, Campagna R, Kutryb-Zając B, Kuś K, Słominska EM, Smolenski RT, Chlopicki S (2020) Reversal of endothelial dysfunction by nicotinamide mononucleotide via extracellular conversion to nicotinamide riboside. Biochem Pharmacol 178:114019. [CrossRef]
- Mukherjee S, Chellappa K, Moffitt A, Ndungu J, Dellinger RW, Davis JG, Agarwal B, Baur JA (2017) Nicotinamide adenine dinucleotide biosynthesis promotes liver regeneration. Hepatology 65(2):616–630. Erratum in: Hepatology, 2017, 65(4):1427. [CrossRef]
- Dall M, Trammell SAJ, Asping M, Hassing AS, Agerholm M, Vienberg SG, Gillum MP, Larsen S, Treebak JT (2019) Mitochondrial function in liver cells is resistant to perturbations in NAD+ salvage capacity. J Biol Chem 294(36):13304–13326. [CrossRef]
- Lee HJ, Yang SJ (2019) Nicotinamide riboside regulates inflammation and mitochondrial markers in AML12 hepatocytes. Nutr Res Pract 13(1):3–10. [CrossRef]
- Fan R, Cui J, Ren F, Wang Q, Huang Y, Zhao B, Wei L, Qian X, Xiong X (2018) Overexpression of NRK1 ameliorates diet- and age-induced hepatic steatosis and insulin resistance. Biochem Biophys Res Commun 500(2):476–483. [CrossRef]
- Han X, Bao X, Lou Q, Xie X, Zhang M, Zhou S, Guo H, Jiang G, Shi Q (2019) Nicotinamide riboside exerts protective effect against aging-induced NAFLD-like hepatic dysfunction in mice. PeerJ 7:e7568. [CrossRef]
- Bock KW (2020) Modulation of aryl hydrocarbon receptor (AHR) and the NAD+-consuming enzyme CD38: Searches of therapeutic options for nonalcoholic fatty liver disease (NAFLD). Biochem Pharmacol 175:113905. [CrossRef]
- Dall M, Hassing AS, Niu L, Nielsen TS, Ingerslev LR, Sulek K, Trammell SAJ, Gillum MP, Barrès R, Larsen S, Poulsen SS, Mann M, Ørskov C, Treebak JT (2021) Hepatocyte-specific perturbation of NAD+ biosynthetic pathways in mice induces reversible nonalcoholic steatohepatitis-like phenotypes. J Biol Chem 297(6):101388. [CrossRef]
- Quesada-Vázquez S, Bone C, Saha S, Triguero I, Colom-Pellicer M, Aragonès G, Hildebrand F, Del Bas JM, Caimari A, Beraza N, Escoté X (2022) Microbiota dysbiosis and gut barrier dysfunction associated with non-alcoholic fatty liver disease are modulated by a specific metabolic cofactors’ combination. Int J Mol Sci 23(22):13675. [CrossRef]
- Dellinger RW, Holmes HE, Hu-Seliger T, Butt RW, Harrison SA, Mozaffarian D, Chen O, Guarente L (2022) Nicotinamide riboside and pterostilbene reduces markers of hepatic inflammation in NAFLD: a double-blind, placebo-controlled clinical trial. Hepatology Sep 9. [CrossRef]
- Pham TX, Bae M, Kim MB, Lee Y, Hu S, Kang H, Park YK, Lee JY (2019) Nicotinamide riboside, an NAD+ precursor, attenuates the development of liver fibrosis in a diet-induced mouse model of liver fibrosis. Biochim Biophys Acta Mol Basis Dis 1865(9):2451–2463. [CrossRef]
- Jiang R, Zhou Y, Wang S, Pang N, Huang Y, Ye M, Wan T, Qiu Y, Pei L, Jiang X, Huang Y, Yang H, Ling W, Li X, Zhang Z, Yang L (2019) Nicotinamide riboside protects against liver fibrosis induced by CCl4 via regulating the acetylation of Smads signaling pathway. Life Sci 225:20–28. [CrossRef]
- Wang S, Wan T, Ye M, Qiu Y, Pei L, Jiang R, Pang N, Huang Y, Liang B, Ling W, Lin X, Zhang Z, Yang L (2018) Nicotinamide riboside attenuates alcohol induced liver injuries via activation of SirT1/PGC-1α/mitochondrial biosynthesis pathway. Redox Biol 17:89–98. [CrossRef]
- Yau WW, Chen GB, Zhou J, Francisco JC, Thimmukonda NK, Li S, Singh BK, Yen PM (2023) Nicotinamide riboside rescues dysregulated glycolysis and fatty acid β-oxidation in a human hepatic cell model of citrin deficiency. Hum Mol Genet 32(11):1922–1931. [CrossRef]
- Lee SR, Roh JY, Ryu J, Shin HJ, Hong EJ (2022) Activation of TCA cycle restrains virus-metabolic hijacking and viral replication in mouse hepatitis virus-infected cells. Cell Biosci 12(1):7. [CrossRef]
- Parikh SM (2019) Metabolic stress resistance in acute kidney injury: evidence for a PPAR-gamma-coactivator-1 alpha-nicotinamide adenine dinucleotide pathway. Nephron 143(3):184–187. [CrossRef]
- Simic P, Vela Parada XF, Parikh SM, Dellinger R, Guarente LP, Rhee EP (2020) Nicotinamide riboside with pterostilbene (NRPT) increases NAD+ in patients with acute kidney injury (AKI): a randomized, double-blind, placebo-controlled, stepwise safety study of escalating doses of NRPT in patients with AKI. BMC Nephrol 21(1):342. [CrossRef]
- Lin W, Wu X, Wen J, Fei Y, Wu J, Li X, Zhang Q, Dong Y, Xu T, Fan Y, Wang N (2021) Nicotinamide retains Klotho expression and ameliorates rhabdomyolysis-induced acute kidney injury. Nutrition 91–92:111376. [CrossRef]
- Piedrafita A, Balayssac S, Mayeur N, Gazut S, Grossac J, Buleon M, Alves M, Klein J, Minville V, Marcheix B, Schanstra JP, Faguer S (2021) The tryptophan pathway and nicotinamide supplementation in ischaemic acute kidney injury. Clin Kidney J 14(12):2490–2496. [CrossRef]
- Fontecha-Barriuso M, Lopez-Diaz AM, Carriazo S, Ortiz A, Sanz AB (2021) Nicotinamide and acute kidney injury. Clin Kidney J 14(12):2453–2462. [CrossRef]
- Pezzotta A, Perico L, Morigi M, Corna D, Locatelli M, Zoja C, Benigni A, Remuzzi G, Imberti B (2022) Low nephron number induced by maternal protein restriction is prevented by nicotinamide riboside supplementation depending on sirtuin 3 activation. Cells 11(20):3316. [CrossRef]
- Morevati M, Egstrand S, Nordholm A, Mace ML, Andersen CB, Salmani R, Olgaard K, Lewin E (2021) Effect of NAD+ boosting on kidney ischemia–reperfusion injury. PLoS One 16(6):e0252554. [CrossRef]
- Doke T, Mukherjee S, Mukhi D, Dhillon P, Abedini A, Davis JG, Chellappa K, Chen B, Baur JA, Susztak K (2023) NAD+ precursor supplementation prevents mtRNA/RIG-I-dependent inflammation during kidney injury. Nat Metab 5(3):414–430. [CrossRef]
- Ahmadi A, Begue G, Valencia AP, Norman JE, Lidgard B, Bennett BJ, Van Doren MP, Marcinek DJ, Fan S, Prince DK, Gamboa JL, Himmelfarb J, de Boer IH, Kestenbaum BR, Roshanravan B (2023) Randomized crossover clinical trial of coenzyme Q10 and nicotinamide riboside in chronic kidney disease. JCI Insight May 9:e167274. [CrossRef]
- Fletcher RS, Ratajczak J, Doig CL, Oakey LA, Callingham R, Da Silva Xavier G, Garten A, Elhassan YS, Redpath P, Migaud ME, Philp A, Brenner C, Canto C, Lavery GG (2017) Nicotinamide riboside kinases display redundancy in mediating nicotinamide mononucleotide and nicotinamide riboside metabolism in skeletal muscle cells. Mol Metab 6(8):819–832. [CrossRef]
- Sonntag T, Ancel S, Karaz S, Cichosz P, Jacot G, Giner MP, Sanchez-Garcia JL, Pannérec A, Moco S, Sorrentino V, Cantó C, Feige JN (2022) Nicotinamide riboside kinases regulate skeletal muscle fiber-type specification and are rate-limiting for metabolic adaptations during regeneration. Front Cell Dev Biol 10:1049653. [CrossRef]
- Doig CL, Zielinska AE, Fletcher RS, Oakey LA, Elhassan YS, Garten A, Cartwright D, Heising S, Alsheri A, Watson DG, Prehn C, Adamski J, Tennant DA, Lavery GG (2020) Induction of the nicotinamide riboside kinase NAD+ salvage pathway in a model of sarcoplasmic reticulum dysfunction. Skelet Muscle 10(1):5. [CrossRef]
- Damgaard MV, Nielsen TS, Basse AL, Chubanava S, Trost K, Moritz T, Dellinger RW, Larsen S, Treebak JT (2022) Intravenous nicotinamide riboside elevates mouse skeletal muscle NAD+ without impacting respiratory capacity or insulin sensitivity. iScience 25(2):103863. [CrossRef]
- Elhassan YS, Kluckova K, Fletcher RS, Schmidt MS, Garten A, Doig CL, Cartwright DM, Oakey L, Burley CV, Jenkinson N, Wilson M, Lucas SJE, Akerman I, Seabright A, Lai YC, Tennant DA, Nightingale P, Wallis GA, Manolopoulos KN, Brenner C, Philp A, Lavery GG (2019) Nicotinamide riboside augments the aged human skeletal muscle NAD+ metabolome and induces transcriptomic and anti-inflammatory signatures. Cell Rep 28(7):1717–1728.e6. [CrossRef]
- Remie CME, Roumans KHM, Moonen MPB, Connell NJ, Havekes B, Mevenkamp J, Lindeboom L, de Wit VHW, van de Weijer T, Aarts SABM, Lutgens E, Schomakers BV, Elfrink HL, Zapata-Pérez R, Houtkooper RH, Auwerx J, Hoeks J, Schrauwen-Hinderling VB, Phielix E, Schrauwen P (2020) Nicotinamide riboside supplementation alters body composition and skeletal muscle acetylcarnitine concentrations in healthy obese humans. Am J Clin Nutr 112(2):413–426. [CrossRef]
- Custodero C, Saini SK, Shin MJ, Jeon YK, Christou DD, McDermott MM, Leeuwenburgh C, Anton SD, Mankowski RT (2020) Nicotinamide riboside – a missing piece in the puzzle of exercise therapy for older adults? Exp Gerontol 137:110972. [CrossRef]
- Campelj D, Philp A (2022) NAD+ therapeutics and skeletal muscle adaptation to exercise in humans. Sports Med 52(Suppl 1):91–99. [CrossRef]
- Stocks B, Ashcroft SP, Joanisse S, Dansereau LC, Koay YC, Elhassan YS, Lavery GG, Quek LE, O’Sullivan JF, Philp AM, Wallis GA, Philp A (2021) Nicotinamide riboside supplementation does not alter whole-body or skeletal muscle metabolic responses to a single bout of endurance exercise. J Physiol 599(5):1513–1531. [CrossRef]
- Song M, Armenian SH, Bhandari R, Lee K, Ness K, Putt M, Lindenfeld L, Manoukian S, Wade K, Dedio A, Guzman T, Hampton I, Lin K, Baur J, McCormack S, Mostoufi-Moab S (2022) Exercise training and NR supplementation to improve muscle mass and fitness in adolescent and young adult hematopoietic cell transplant survivors: a randomized controlled trial {1}. BMC Cancer 22(1):795. [CrossRef]
- Kourtzidis IA, Stoupas AT, Gioris IS, Veskoukis AS, Margaritelis NV, Tsantarliotou M, Taitzoglou I, Vrabas IS, Paschalis V, Kyparos A, Nikolaidis MG (2016) The NAD+ precursor nicotinamide riboside decreases exercise performance in rats. J Int Soc Sports Nutr 13:32. [CrossRef]
- Kourtzidis IA, Dolopikou CF, Tsiftsis AN, Margaritelis NV, Theodorou AA, Zervos IA, Tsantarliotou MP, Veskoukis AS, Vrabas IS, Paschalis V, Kyparos A, Nikolaidis MG (2018) Nicotinamide riboside supplementation dysregulates redox and energy metabolism in rats: implications for exercise performance. Exp Physiol 103(10):1357–1366. [CrossRef]
- Deloux R, Tannous C, Ferry A, Li Z, Mericskay M (2018) Aged nicotinamide riboside kinase 2 deficient mice present an altered response to endurance exercise training. Front Physiol 9:1290. [CrossRef]
- Zhang H, Ryu D, Wu Y, Gariani K, Wang X, Luan P, D’Amico D, Ropelle ER, Lutolf MP, Aebersold R, Schoonjans K, Menzies KJ, Auwerx J (2016) NAD+ repletion improves mitochondrial and stem cell function and enhances life span in mice. Science 352(6292):1436–1443. [CrossRef]
- Moon J, Kim HR, Shin MG (2018) Rejuvenating aged hematopoietic stem cells through improvement of mitochondrial function. Ann Lab Med 38(5):395–401. [CrossRef]
- Khan NA, Auranen M, Paetau I, Pirinen E, Euro L, Forsström S, Pasila L, Velagapudi V, Carroll CJ, Auwerx J, Suomalainen A (2014) Effective treatment of mitochondrial myopathy by nicotinamide riboside, a vitamin B3. EMBO Mol Med 6(6):721–731. [CrossRef]
- Schaefer PM, Huang J, Butic A, Perry C, Yardeni T, Tan W, Morrow R, Baur JA, Wallace DC (2022) Nicotinamide riboside alleviates exercise intolerance in ANT1-deficient mice. Mol Metab 64:101560. [CrossRef]
- Lapatto HAK, Kuusela M, Heikkinen A, Muniandy M, van der Kolk BW, Gopalakrishnan S, Pöllänen N, Sandvik M, Schmidt MS, Heinonen S, Saari S, Kuula J, Hakkarainen A, Tampio J, Saarinen T, Taskinen MR, Lundbom N, Groop PH, Tiirola M, Katajisto P, Lehtonen M, Brenner C, Kaprio J, Pekkala S, Ollikainen M, Pietiläinen KH, Pirinen E (2023) Nicotinamide riboside improves muscle mitochondrial biogenesis, satellite cell differentiation, and gut microbiota in a twin study. Sci Adv 9(2):eadd5163. [CrossRef]
- Perez-Sanchez C, Escudero-Contreras A, Cerdó T, Sánchez-Mendoza LM, Llamas-Urbano A, Arias-de la Rosa I, Pérez-Rodriguez M, Muñoz-Barrera L, Abalos-Aguilera MDC, Barbarroja N, Calvo J, Ortega-Castro R, Ruiz-Vilches D, Moreno JA, Burón MI, González-Reyes JA, Collantes-Estevez E, Lopez-Pedrera C, Villalba JM (2023) Preclinical characterization of pharmacological NAD+ boosting as a promising therapeutic approach in rheumatoid arthritis. Arthritis Rheumatol Apr 24. [CrossRef]
- Gonzalez JM, Jackson AR (2020) In ovo feeding of nicotinamide riboside affects broiler pectoralis major muscle development. Transl Anim Sci 4(3):txaa126. [CrossRef]
- Xu X, Jackson AR, Gonzalez JM (2021) The effects of in ovo nicotinamide riboside dose on broiler myogenesis. Poult Sci 100(3):100926. [CrossRef]
- Xu X, Alcocer HM, Gravely ME, Jackson AR, Gonzalez JM (2022) Effects of in ovo injection of nicotinamide riboside on high-yield broiler myogenesis. J Anim Sci 100(8):skac203. [CrossRef]
- Sambeat A, Ratajczak J, Joffraud M, Sanchez-Garcia JL, Giner MP, Valsesia A, Giroud-Gerbetant J, Valera-Alberni M, Cercillieux A, Boutant M, Kulkarni SS, Moco S, Canto C (2019) Endogenous nicotinamide riboside metabolism protects against diet-induced liver damage. Nat Commun 10(1):4291. [CrossRef]
- Igarashi M, Miura M, Williams E, Jaksch F, Kadowaki T, Yamauchi T, Guarente L (2019) NAD+ supplementation rejuvenates aged gut adult stem cells. Aging Cell 18(3):e12935. [CrossRef]
- Cartwright DM, Oakey LA, Fletcher RS, Doig CL, Heising S, Larner DP, Nasteska D, Berry CE, Heaselgrave SR, Ludwig C, Hodson DJ, Lavery GG, Garten A (2021) Nicotinamide riboside has minimal impact on energy metabolism in mouse models of mild obesity. J Endocrinol 251(1):111–123. [CrossRef]
- Lozada-Fernández VV, deLeon O, Kellogg SL, Saravia FL, Hadiono MA, Atkinson SN, Grobe JL, Kirby JR (2022) Nicotinamide riboside-conditioned microbiota deflects high-fat diet-induced weight gain in mice. mSystems 7(1):e0023021. [CrossRef]
- Sauve AA (2022) Metabolic disease, NAD metabolism, nicotinamide riboside, and the gut microbiome: connecting the dots from the gut to physiology. mSystems 7(1):e0122321. [CrossRef]
- Kolba N, Zarei A, Cheng J, Agarwal N, Dadmohammadi Y, Khazdooz L, Abbaspourrad A, Tako E (2022) Alterations in intestinal brush border membrane functionality and bacterial populations following intra-amniotic administration (Gallus gallus) of nicotinamide riboside and its derivatives. Nutrients 14(15):3130. [CrossRef]
- Peluso AA, Lundgaard AT, Babaei P, Mousovich-Neto F, Rocha AL, Damgaard MV, Bak EG, Gnanasekaran T, Dollerup OL, Trammell SAJ, Nielsen TS, Kern T, Abild CB, Sulek K, Ma T, Gerhart-Hines Z, Gillum MP, Arumugam M, Ørskov C, McCloskey D, Jessen N, Herrgård MJ, Mori MAS, Treebak JT (2023) Oral supplementation of nicotinamide riboside alters intestinal microbial composition in rats and mice, but not humans. NPJ Aging 9(1):7. [CrossRef]
- Kim MB, Pham TX, vanLuling M, Kostour V, Kang H, Corvino O, Jang H, Odell W, Bae M, Park YK, Lee JY (2022) Nicotinamide riboside supplementation exerts an anti-obesity effect and prevents inflammation and fibrosis in white adipose tissue of female diet-induced obesity mice. J Nutr Biochem 107:109058. [CrossRef]
- Longo L, de Castro JM, Keingeski MB, Rampelotto PH, Stein DJ, Guerreiro GTS, de Souza VEG, Cerski CTS, Uribe-Cruz C, Torres ILS, Álvares-da-Silva MR (2023) Nicotinamide riboside and dietary restriction effects on gut microbiota and liver inflammatory and morphologic markers in cafeteria diet-induced obesity in rats. Nutrition 110:112019. [CrossRef]
- Yu X, Xue M, Liu Y, Zhou Z, Jiang Y, Sun T, Liang H (2020) Effect of nicotinamide riboside on lipid metabolism and gut microflora–bile acid axis in alcohol-exposed mice. Food Sci Nutr 9(1):429–440. [CrossRef]
- Crisol BM, Veiga CB, Lenhare L, Braga RR, Silva VRR, da Silva ASR, Cintra DE, Moura LP, Pauli JR, Ropelle ER (2018) Nicotinamide riboside induces a thermogenic response in lean mice. Life Sci 211:1–7. [CrossRef]
- Crisol BM, Veiga CB, Braga RR, Lenhare L, Baptista IL, Gaspar RC, Muñoz VR, Cordeiro AV, da Silva ASR, Cintra DE, Moura LP, Pauli JR, Ropelle ER (2020) NAD+ precursor increases aerobic performance in mice. Eur J Nutr 59(6):2427–2437. [CrossRef]
- Williams AS, Koves TR, Pettway YD, Draper JA, Slentz DH, Grimsrud PA, Ilkayeva OR, Muoio DM (2021) Nicotinamide riboside supplementation confers marginal metabolic benefits in obese mice without remodeling the muscle acetyl-proteome. iScience 25(1):103635. [CrossRef]
- Serrano A, Palou A, Bonet ML, Ribot J (2022) Nicotinamide riboside supplementation to suckling male mice improves lipid and energy metabolism in skeletal muscle and liver in adulthood. Nutrients 14(11):2259. [CrossRef]
- de Castro JM, Stein DJ, Medeiros HR, de Oliveira C, Torres ILS (2021) Nicotinamide riboside neutralizes hypothalamic inflammation and increases weight loss without altering muscle mass in obese rats under calorie restriction: a preliminary investigation. Front Nutr 8:648893. [CrossRef]
- Shi W, Hegeman MA, van Dartel DAM, Tang J, Suarez M, Swarts H, van der Hee B, Arola L, Keijer J (2017) Effects of a wide range of dietary nicotinamide riboside (NR) concentrations on metabolic flexibility and white adipose tissue (WAT) of mice fed a mildly obesogenic diet. Mol Nutr Food Res 61(8):1600878. [CrossRef]
- Shi W, Hegeman MA, Doncheva A, Bekkenkamp-Grovenstein M, de Boer VCJ, Keijer J (2019) High dose of dietary nicotinamide riboside induces glucose intolerance and white adipose tissue dysfunction in mice fed a mildly obesogenic diet. Nutrients 11(10):2439. [CrossRef]
- Shi W, Hegeman MA, Doncheva A, van der Stelt I, Bekkenkamp-Grovenstein M, van Schothorst EM, Brenner C, de Boer VCJ, Keijer J (2019) Transcriptional response of white adipose tissue to withdrawal of vitamin B3. Mol Nutr Food Res 63(13):e1801100. [CrossRef]
- Quesada-Vázquez S, Antolín A, Colom-Pellicer M, Aragonès G, Herrero L, Del Bas JM, Caimari A, Escoté X (2022) Reduction of obesity and insulin resistance through dual targeting of VAT and BAT by a novel combination of metabolic cofactors. Int J Mol Sci 23(23):14923. [CrossRef]
- Zhao H, Tian Y, Zuo Y, Zhang X, Gao Y, Wang P, Sun L, Zhang H, Liang H (2022) Nicotinamide riboside ameliorates high-fructose-induced lipid metabolism disorder in mice via improving FGF21 resistance in the liver and white adipose tissue. Food Funct 13(23):12400–12411. [CrossRef]
- Serrano A, Asnani-Kishnani M, Couturier C, Astier J, Palou A, Landrier JF, Ribot J, Bonet ML (2020) DNA methylation changes are associated with the programming of white adipose tissue browning features by resveratrol and nicotinamide riboside neonatal supplementations in mice. Nutrients 12(2):461. [CrossRef]
- Salic K, Gart E, Seidel F, Verschuren L, Caspers M, van Duyvenvoorde W, Wong KE, Keijer J, Bobeldijk-Pastorova I, Wielinga PY, Kleemann R (2019) Combined treatment with L-carnitine and nicotinamide riboside improves hepatic metabolism and attenuates obesity and liver steatosis. Int J Mol Sci 20(18):4359. [CrossRef]
- Nagy L, Rauch B, Szerafin T, Uray K, Tóth A, Bai P (2022) Nicotinamide-riboside shifts the differentiation of human primary white adipocytes to beige adipocytes impacting substrate preference and uncoupling respiration through SIRT1 activation and mitochondria-derived reactive species production. Front Cell Dev Biol 10:979330. [CrossRef]
- Nascimento EBM, Moonen MPB, Remie CME, Gariani K, Jörgensen JA, Schaart G, Hoeks J, Auwerx J, van Marken Lichtenbelt WD, Schrauwen P (2021) Nicotinamide riboside enhances in vitro beta-adrenergic brown adipose tissue activity in humans. J Clin Endocrinol Metab 106(5):1437–1447. [CrossRef]
- Dollerup OL, Trammell SAJ, Hartmann B, Holst JJ, Christensen B, Møller N, Gillum MP, Treebak JT, Jessen N (2019) Effects of nicotinamide riboside on endocrine pancreatic function and incretin hormones in nondiabetic men with obesity. J Clin Endocrinol Metab 104(11):5703–5714. [CrossRef]
- Dollerup OL, Chubanava S, Agerholm M, Søndergård SD, Altıntaş A, Møller AB, Høyer KF, Ringgaard S, Stødkilde-Jørgensen H, Lavery GG, Barrès R, Larsen S, Prats C, Jessen N, Treebak JT (2020) Nicotinamide riboside does not alter mitochondrial respiration, content or morphology in skeletal muscle from obese and insulin-resistant men. J Physiol 598(4):731–754. [CrossRef]
- Trammell SAJ, Weidemann BJ, Chadda A, Yorek MS, Holmes A, Coppey LJ, Obrosov A, Kardon RH, Yorek MA, Brenner C (2016) Nicotinamide riboside opposes type 2 diabetes and neuropathy in mice. Sci Rep 6:26933. [CrossRef]
- Chandrasekaran K, Najimi N, Sagi AR, Yarlagadda S, Salimian M, Arvas MI, Hedayat AF, Kevas Y, Kadakia A, Russell JW (2022) NAD+ precursors repair mitochondrial function in diabetes and prevent experimental diabetic neuropathy. Int J Mol Sci 23(9):4887. [CrossRef]
- Cercillieux A, Ratajczak J, Joffraud M, Sanchez-Garcia JL, Jacot G, Zollinger A, Métairon S, Giroud-Gerbetant J, Rumpler M, Ciarlo E, Valera-Alberni M, Sambeat A, Canto C (2022b) Nicotinamide riboside kinase 1 protects against diet and age-induced pancreatic β-cell failure. Mol Metab 66:101605. [CrossRef]
- Hu L, Guo Y, Song L, Wen H, Sun N, Wang Y, Qi B, Liang Q, Geng J, Liu X, Fu F, Li Y (2022) Nicotinamide riboside promotes Mfn2-mediated mitochondrial fusion in diabetic hearts through the SIRT1–PGC1α–PPARα pathway. Free Radic Biol Med 183:75–88. [CrossRef]
- Costa CJ, Cohen MW, Goldberg DC, Mellado W, Willis DE (2023) Nicotinamide riboside improves enteric neuropathy in streptozocin-induced diabetic rats through myenteric plexus neuroprotection. Dig Dis Sci Mar 15. [CrossRef]
- Lee HJ, Hong YS, Jun W, Yang SJ (2015) Nicotinamide riboside ameliorates hepatic metaflammation by modulating NLRP3 inflammasome in a rodent model of type 2 diabetes. J Med Food 18(11):1207–1213. [CrossRef]
- Wang ZH, Bao XG, Hu JJ, Shen SB, Xu GH, Wu YL (2021) Nicotinamide riboside enhances endothelial precursor cell function to promote refractory wound healing through mediating the Sirt1/AMPK pathway. Front Pharmacol 12:671563. [CrossRef]
- Lee SR, Jeong SH, Mukae M, Kim SY, Ko JW, Kwun HJ, Hong EJ (2023) Dietary supplementation with nicotinamide riboside improves fetal growth under hypoglycemia. J Nutr Biochem 116:109310. [CrossRef]
- Chu X, Raju RP (2022) Regulation of NAD+ metabolism in aging and disease. Metabolism 126:154923. [CrossRef]
- Ji LL, Yeo D (2022) Maintenance of NAD+ homeostasis in skeletal muscle during aging and exercise. Cells 11(4):710. [CrossRef]
- Gomes AP, Price NL, Ling AJY, Moslehi JJ, Montgomery MK, Rajman L, White JP, Teodoro JS, Wrann CD, Hubbard BP, Mercken EM, Palmeira CM, de Cabo R, Rolo AP, Turner N, Bell EL, Sinclair DA (2013) Declining NAD+ induces a pseudohypoxic state disrupting nuclear–mitochondrial communication during aging. Cell 155(7):1624–1638. [CrossRef]
- Cercillieux A, Ciarlo E, Canto C (2022) Balancing NAD+ deficits with nicotinamide riboside: therapeutic possibilities and limitations. Cell Mol Life Sci 79(8):463. [CrossRef]
- Li W, Zhou Y, Pang N, Hu Q, Li Q, Sun Y, Ding Y, Gu Y, Xiao Y, Gao M, Ma S, Pan J, Fang EF, Zhang Z, Yang L (2022) NAD supplement alleviates intestinal barrier injury induced by ethanol via protecting epithelial mitochondrial function. Nutrients 15(1):174. [CrossRef]
- Vannini N, Campos V, Girotra M, Trachsel V, Rojas-Sutterlin S, Tratwal J, Ragusa S, Stefanidis E, Ryu D, Rainer PY, Nikitin G, Giger S, Li TY, Semilietof A, Oggier A, Yersin Y, Tauzin L, Pirinen E, Cheng WC, Ratajczak J, Canto C, Ehrbar M, Sizzano F, Petrova TV, Vanhecke D, Zhang L, Romero P, Nahimana A, Cherix S, Duchosal MA, Ho PC, Deplancke B, Coukos G, Auwerx J, Lutolf MP, Naveiras O (2019) The NAD-booster nicotinamide riboside potently stimulates hematopoiesis through increased mitochondrial clearance. Cell Stem Cell 24(3):405–418.e7. [CrossRef]
- Zong L, Tanaka-Yano M, Park B, Yanai H, Turhan FT, Croteau DL, Tian J, Fang EF, Bohr VA, Beerman I (2021) NAD+ augmentation with nicotinamide riboside improves lymphoid potential of Atm-/- and old mice HSCs. NPJ Aging Mech Dis 7(1):25. [CrossRef]
- Sun X, Cao B, Naval-Sanchez M, Pham T, Sun YBY, Williams B, Heazlewood SY, Deshpande N, Li J, Kraus F, Rae J, Nguyen Q, Yari H, Schröder J, Heazlewood CK, Fulton M, Hatwell-Humble J, Das Gupta K, Kapetanovic R, Chen X, Sweet MJ, Parton RG, Ryan MT, Polo JM, Nefzger CM, Nilsson SK (2021) Nicotinamide riboside attenuates age-associated metabolic and functional changes in hematopoietic stem cells. Nat Commun 12(1):2665. [CrossRef]
- Yoshino J, Mills KF, Yoon MJ, Imai S (2011) Nicotinamide mononucleotide, a key NAD+ intermediate, treats the pathophysiology of diet- and age-induced diabetes in mice. Cell Metab 14(4):528–536. [CrossRef]
- Zapata-Pérez R, Tammaro A, Schomakers BV, Scantlebery AML, Denis S, Elfrink HL, Giroud-Gerbetant J, Cantó C, López-Leonardo C, McIntyre RL, van Weeghel M, Sánchez-Ferrer Á, Houtkooper RH (2021) Reduced nicotinamide mononucleotide is a new and potent NAD+ precursor in mammalian cells and mice. FASEB J 35(4):e21456. [CrossRef]
- Freeberg KA, Craighead DH, Martens CR, You Z, Chonchol M, Seals DR (2022) Nicotinamide riboside supplementation for treating elevated systolic blood pressure and arterial stiffness in midlife and older adults. Front Cardiovasc Med 9:881703. [CrossRef]
- Acklin S, Sadhukhan R, Du W, Patra M, Cholia R, Xia F (2022) Nicotinamide riboside alleviates cisplatin-induced peripheral neuropathy via SIRT2 activation. Neurooncol Adv 4(1):vdac101. [CrossRef]
- Mukherjee S, Mo J, Paolella LM, Perry CE, Toth J, Hugo MM, Chu Q, Tong Q, Chellappa K, Baur JA (2021) SIRT3 is required for liver regeneration but not for the beneficial effect of nicotinamide riboside. JCI Insight 6(7):e147193. [CrossRef]
- Zhang Q, Liu X, Li N, Zhang J, Yang J, Bu P (2018) Sirtuin 3 deficiency aggravates contrast-induced acute kidney injury. J Transl Med 16(1):313. Erratum in: J Transl Med, 2022, 20(1):46. [CrossRef]
- Kang H, Park YK, Lee JY (2021) Nicotinamide riboside, an NAD+ precursor, attenuates inflammation and oxidative stress by activating sirtuin 1 in alcohol-stimulated macrophages. Lab Invest 101(9):1225–1237. [CrossRef]
- Seldeen KL, Shahini A, Thiyagarajan R, Redae Y, Leiker M, Rajabian N, Dynka A, Andreadis ST, Troen BR (2021) Short-term nicotinamide riboside treatment improves muscle quality and function in mice and increases cellular energetics and differentiating capacity of myogenic progenitors. Nutrition 87–88:111189. [CrossRef]
- Jensen JB, Dollerup OL, Møller AB, Billeskov TB, Dalbram E, Chubanava S, Damgaard MV, Dellinger RW, Trošt K, Moritz T, Ringgaard S, Møller N, Treebak JT, Farup J, Jessen N (2022) A randomized placebo-controlled trial of nicotinamide riboside and pterostilbene supplementation in experimental muscle injury in elderly individuals. JCI Insight 7(19):e158314. [CrossRef]
- Dolopikou CF, Kourtzidis IA, Margaritelis NV, Vrabas IS, Koidou I, Kyparos A, Theodorou AA, Paschalis V, Nikolaidis MG (2020) Acute nicotinamide riboside supplementation improves redox homeostasis and exercise performance in old individuals: a double-blind cross-over study. Eur J Nutr 59(2):505–515. [CrossRef]
- Grezella C, Fernandez-Rebollo E, Franzen J, Ventura Ferreira MS, Beier F, Wagner W (2018) Effects of senolytic drugs on human mesenchymal stromal cells. Stem Cell Res Ther 9(1):108. [CrossRef]
- Delabie W, Maes W, Devloo R, Van den Hauwe MR, Vanhoorelbeke K, Compernolle V, Feys HB (2020) The senotherapeutic nicotinamide riboside raises platelet nicotinamide adenine dinucleotide levels but cannot prevent storage lesion. Transfusion 60(1):165–174. [CrossRef]
- Harrison DE, Strong R, Reifsnyder P, Kumar N, Fernandez E, Flurkey K, Javors MA, Lopez-Cruzan M, Macchiarini F, Nelson JF, Markewych A, Bitto A, Sindler AL, Cortopassi G, Kavanagh K, Leng L, Bucala R, Rosenthal N, Salmon A, Stearns TM, Bogue M, Miller RA (2021) 17-α-Estradiol late in life extends lifespan in aging UM-HET3 male mice; nicotinamide riboside and three other drugs do not affect lifespan in either sex. Aging Cell 20(5):e13328. Erratum in: Aging Cell, 2022, 21(11):e13672. [CrossRef]
- Belenky PA, Moga TG, Brenner C (2008) Saccharomyces cerevisiae YOR071C encodes the high affinity nicotinamide riboside transporter Nrt1. J Biol Chem 283(13):8075–8079. [CrossRef]
- Orlandi I, Alberghina L, Vai M (2020) Nicotinamide, nicotinamide riboside and nicotinic acid-emerging roles in replicative and chronological aging in yeast. Biomolecules 10(4):604. [CrossRef]
- Odoh CK, Guo X, Arnone JT, Wang X, Zhao ZK (2022) The role of NAD and NAD precursors on longevity and lifespan modulation in the budding yeast, Saccharomyces cerevisiae. Biogerontology 23(2):169–199. [CrossRef]
- Belenky P, Christensen KC, Gazzaniga F, Pletnev AA, Brenner C (2009) Nicotinamide riboside and nicotinic acid riboside salvage in fungi and mammals. Quantitative basis for Urh1 and purine nucleoside phosphorylase function in NAD+ metabolism. J Biol Chem 284(1):158–164. Erratum in: J Biol Chem, 2009, 284(12):8208. [CrossRef]
- Braidy N, Poljak A, Grant R, Jayasena T, Mansour H, Chan-Ling T, Guillemin GJ, Smythe G, Sachdev P (2014) Mapping NAD+ metabolism in the brain of ageing Wistar rats: potential targets for influencing brain senescence. Biogerontology 15(2):177–198. [CrossRef]
- Verdin E (2015) NAD⁺ in aging, metabolism, and neurodegeneration. Science 350(6265):1208–1213. [CrossRef]
- Reiten OK, Wilvang MA, Mitchell SJ, Hu Z, Fang EF (2021) Preclinical and clinical evidence of NAD+ precursors in health, disease, and ageing. Mech Ageing Dev 199:111567. [CrossRef]
- Braidy N, Liu Y (2020) Can nicotinamide riboside protect against cognitive impairment? Curr Opin Clin Nutr Metab Care 23(6):413–420. [CrossRef]
- Birkisdóttir MB, van Galen I, Brandt RMC, Barnhoorn S, van Vliet N, van Dijk C, Nagarajah B, Imholz S, van Oostrom CT, Reiling E, Gyenis Á, Mastroberardino PG, Jaarsma D, van Steeg H, Hoeijmakers JHJ, Dollé MET, Vermeij WP (2022) The use of progeroid DNA repair-deficient mice for assessing anti-aging compounds, illustrating the benefits of nicotinamide riboside. Front Aging 3:1005322. Erratum in: Front Aging, 2022, 3:1086552. [CrossRef]
- Hou Y, Lautrup S, Cordonnier S, Wang Y, Croteau DL, Zavala E, Zhang Y, Moritoh K, O’Connell JF, Baptiste BA, Stevnsner TV, Mattson MP, Bohr VA (2018) NAD+ supplementation normalizes key Alzheimer’s features and DNA damage responses in a new AD mouse model with introduced DNA repair deficiency. Proc Natl Acad Sci U S A 115(8):E1876–E1885. [CrossRef]
- Xie X, Gao Y, Zeng M, Wang Y, Wei TF, Lu YB, Zhang WP (2019) Nicotinamide ribose ameliorates cognitive impairment of aged and Alzheimer’s disease model mice. Metab Brain Dis 34(1):353–366. [CrossRef]
- Larrick JW, Mendelsohn AR (2021) Modulation of cGAS-STING pathway by nicotinamide riboside in Alzheimer’s disease. Rejuvenation Res 24(5):397–402. [CrossRef]
- Roboon J, Hattori T, Ishii H, Takarada-Iemata M, Nguyen DT, Heer CD, O’Meally D, Brenner C, Yamamoto Y, Okamoto H, Higashida H, Hori O (2021) Inhibition of CD38 and supplementation of nicotinamide riboside ameliorate lipopolysaccharide-induced microglial and astrocytic neuroinflammation by increasing NAD. J Neurochem 158(2):311–327. [CrossRef]
- Chu X, Hou Y, Meng Q, Croteau DL, Wei Y, De S, Becker KG, Bohr VA (2022) Nicotinamide adenine dinucleotide supplementation drives gut microbiota variation in Alzheimer’s mouse model. Front Aging Neurosci 14:993615. [CrossRef]
- Ryu WI, Shen M, Lee Y, Healy RA, Bormann MK, Cohen BM, Sonntag KC (2022) Nicotinamide riboside and caffeine partially restore diminished NAD availability but not altered energy metabolism in Alzheimer’s disease. Aging Cell 21(7):e13658. [CrossRef]
- Yulug B, Altay O, Li X, Hanoglu L, Cankaya S, Lam S, Velioglu HA, Yang H, Coskun E, Idil E, Nogaylar R, Ozsimsek A, Bayram C, Bolat I, Oner S, Tozlu OO, Arslan ME, Hacimuftuoglu A, Yildirim S, Arif M, Shoaie S, Zhang C, Nielsen J, Turkez H, Borén J, Uhlén M, Mardinoglu A (2023) Combined metabolic activators improve cognitive functions in Alzheimer’s disease patients: a randomised, double-blinded, placebo-controlled phase-II trial. Transl Neurodegener 12(1):4. [CrossRef]
- Li CC, Chen WX, Wang J, Xia M, Jia ZC, Guo C, Tang XQ, Li MX, Yin Y, Liu X, Feng H (2020) Nicotinamide riboside rescues angiotensin II-induced cerebral small vessel disease in mice. CNS Neurosci Ther 26(4):438–447. [CrossRef]
- Schöndorf DC, Ivanyuk D, Baden P, Sanchez-Martinez A, De Cicco S, Yu C, Giunta I, Schwarz LK, Di Napoli G, Panagiotakopoulou V, Nestel S, Keatinge M, Pruszak J, Bandmann O, Heimrich B, Gasser T, Whitworth AJ, Deleidi M (2018) The NAD+ precursor nicotinamide riboside rescues mitochondrial defects and neuronal loss in iPSC and fly models of Parkinson’s disease. Cell Rep 23(10):2976–2988. [CrossRef]
- Brakedal B, Dölle C, Riemer F, Ma Y, Nido GS, Skeie GO, Craven AR, Schwarzlmüller T, Brekke N, Diab J, Sverkeli L, Skjeie V, Varhaug K, Tysnes OB, Peng S, Haugarvoll K, Ziegler M, Grüner R, Eidelberg D, Tzoulis C (2022) The NADPARK study: a randomized phase I trial of nicotinamide riboside supplementation in Parkinson’s disease. Cell Metab 34(3):396–407.e6. [CrossRef]
- Vreones M, Mustapic M, Moaddel R, Pucha KA, Lovett J, Seals DR, Kapogiannis D, Martens CR (2023) Oral nicotinamide riboside raises NAD+ and lowers biomarkers of neurodegenerative pathology in plasma extracellular vesicles enriched for neuronal origin. Aging Cell 22(1):e13754. [CrossRef]
- Gaare JJ, Dölle C, Brakedal B, Brügger K, Haugarvoll K, Nido GS, Tzoulis C (2023) Nicotinamide riboside supplementation is not associated with altered methylation homeostasis in Parkinson’s disease. iScience 26(3):106278. [CrossRef]
- Pang N, Hu Q, Zhou Y, Xiao Y, Li W, Ding Y, Chen Y, Ye M, Pei L, Li Q, Gu Y, Sun Y, Fang EF, Chen M, Zhang Z, Yang L (2023) Nicotinamide adenine dinucleotide precursor suppresses hepatocellular cancer progression in mice. Nutrients 15(6):1447. [CrossRef]
- Park JM, Han YM, Lee HJ, Park YJ, Hahm KB (2021) Nicotinamide riboside vitamin B3 mitigated C26 adenocarcinoma-induced cancer cachexia. Front Pharmacol 12:665493. [CrossRef]
- Hamity MV, White SR, Blum C, Gibson-Corley KN, Hammond DL (2020) Nicotinamide riboside relieves Paclitaxel-induced peripheral neuropathy and enhances suppression of tumor growth in tumor-bearing rats. Pain 161(10):2364–2375. [CrossRef]
- Kahn B, Borrelli M, Libby T (2022) A narrative review of nicotinamide adenine dinucleotide (NAD)+ intermediates nicotinamide riboside and nicotinamide mononucleotide for keratinocyte carcinoma risk reduction. J Drugs Dermatol 21(10):1129–1132. [CrossRef]
- Conze DB, Crespo-Barreto J, Kruger CL (2016) Safety assessment of nicotinamide riboside, a form of vitamin B3. Hum Exp Toxicol 35(11):1149–1160. Erratum in: Hum Exp Toxicol, 2018, 37(4):448. [CrossRef]
- Dellinger RW, Santos SR, Morris M, Evans M, Alminana D, Guarente L, Marcotulli E (2017) Repeat dose NRPT (nicotinamide riboside and pterostilbene) increases NAD+ levels in humans safely and sustainably: a randomized, double-blind, placebo-controlled study. NPJ Aging Mech Dis 3:17. Erratum in: NPJ Aging Mech Dis, 2018, 4:8. [CrossRef]
- EFSA Panel on Nutrition, Novel Foods and Food Allergens (NDA); Turck D, Castenmiller J, de Henauw S, Hirsch-Ernst KI, Kearney J, Maciuk A, Mangelsdorf I, McArdle HJ, Naska A, Pelaez C, Pentieva K, Siani A, Thies F, Tsabouri S, Vinceti M, Cubadda F, Engel KH, Frenzel T, Heinonen M, Marchelli R, Neuhäuser-Berthold M, Pöting A, Poulsen M, Sanz Y, Schlatter JR, van Loveren Agnès de Sesmaisons-Lecarré H, Germini A, Knutsen HK (2019) Safety of nicotinamide riboside chloride as a novel food pursuant to Regulation (EU) 2015/2283 and bioavailability of nicotinamide from this source, in the context of Directive 2002/46/EC. EFSA J 17(8):e05775. [CrossRef]
- Marinescu AG, Chen J, Holmes HE, Guarente L, Mendes O, Morris M, Dellinger RW (2020) Safety assessment of high-purity, synthetic nicotinamide riboside (NR-E) in a 90-day repeated dose oral toxicity study, with a 28-day recovery arm. Int J Toxicol 39(4):307–320. Erratum in: Int J Toxicol, 2022, 41(1):47. [CrossRef]
- EFSA Panel on Nutrition, Novel Foods and Food Allergens (NDA); Turck D, Bohn T, Castenmiller J, De Henauw S, Hirsch-Ernst KI, Maciuk A, Mangelsdorf I, McArdle HJ, Naska A, Pelaez C, Pentieva K, Siani A, Thies F, Tsabouri S, Vinceti M, Cubadda F, Frenzel T, Heinonen M, Prieto Maradona M, Marchelli R, Neuhäuser-Berthold M, Poulsen M, Schlatter JR, van Loveren H, Albert O, de Sesmaisons Lecarré A, Knutsen HK (2021) Extension of use of nicotinamide riboside chloride as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J 19(11):e06843. [CrossRef]
- Sun P, Qie S, Pan B (2022) Nicotinamide riboside will play an important role in anti-aging therapy in humans, especially in the face skin anti-aging treatment. Aesthetic Plast Surg 46(Suppl 1):192–194. [CrossRef]
- Ito TK, Sato T, Hakamata A, Onoda Y, Sato S, Yamazaki F, Horikawa M, Takahashi Y, Kitamoto T, Suzuki M, Uchida S, Odagiri K, Setou M (2020) A nonrandomized study of single oral supplementation within the daily tolerable upper level of nicotinamide affects blood nicotinamide and NAD+ levels in healthy subjects. Transl Med Aging 4:45–54. [CrossRef]
- Ito TK, Sato T, Takanashi Y, Tamannaa Z, Kitamoto T, Odagiri K, Setou M (2021) A single oral supplementation of nicotinamide within the daily tolerable upper level increases blood NAD+ levels in healthy subjects. Transl Med Aging 5:43–51. [CrossRef]
- Turner J, Licollari A, Mihalcea E, Tan A (2021) Safety evaluation for Restorin® NMN, a NAD+ precursor. Front Pharmacol 12:749727. [CrossRef]
- Okabe K, Yaku K, Uchida Y, Fukamizu Y, Sato T, Sakurai T, Tobe K, Nakagawa T (2022) Oral administration of nicotinamide mononucleotide is safe and efficiently increases blood nicotinamide adenine dinucleotide levels in healthy subjects. Front Nutr 9:868640. [CrossRef]
- Pencina K, Lavu S, Dos Santos M, Beleva YM, Cheng M, Livingston D, Bhasin S (2022) MIB-626, an oral formulation of a microcrystalline unique polymorph of β-nicotinamide mononucleotide, increases circulating nicotinamide adenine dinucleotide and its metabolome in middle-aged and older adults. J Gerontol A Biol Sci Med Sci Feb 19:glac049. [CrossRef]
- Trammell SAJ, Schmidt MS, Weidemann BJ, Redpath P, Jaksch F, Dellinger RW, Li Z, Abel ED, Migaud ME, Brenner C (2016) Nicotinamide riboside is uniquely and orally bioavailable in mice and humans. Nat Commun 7:12948. [CrossRef]
- Airhart SE, Shireman LM, Risler LJ, Anderson GD, Nagana Gowda GA, Raftery D, Tian R, Shen DD, O’Brien KD (2017) An open-label, non-randomized study of the pharmacokinetics of the nutritional supplement nicotinamide riboside (NR) and its effects on blood NAD+ levels in healthy volunteers. PLoS One 12(12):e0186459. [CrossRef]
- Campbell MTD, Jones DS, Andrews GP, Li S (2019) Understanding the physicochemical properties and degradation kinetics of nicotinamide riboside, a promising vitamin B3 nutritional supplement. Food Nutr Res 63:3419. [CrossRef]
- Yaku K, Palikhe S, Izumi H, Yoshida T, Hikosaka K, Hayat F, Karim M, Iqbal T, Nitta Y, Sato A, Migaud ME, Ishihara K, Mori H, Nakagawa T (2021) BST1 regulates nicotinamide riboside metabolism via its glycohydrolase and base-exchange activities. Nat Commun 12(1):6767. [CrossRef]
- Chellappa K, McReynolds MR, Lu W, Zeng X, Makarov M, Hayat F, Mukherjee S, Bhat YR, Lingala SR, Shima RT, Descamps HC, Cox T, Ji L, Jankowski C, Chu Q, Davidson SM, Thaiss CA, Migaud ME, Rabinowitz JD, Baur JA (2022) NAD precursors cycle between host tissues and the gut microbiome. Cell Metab 34(12):1947–1959.e5. [CrossRef]
- Preugschat F, Carter LH, Boros EE, Porter DJ, Stewart EL, Shewchuk LM (2014) A pre-steady state and steady state kinetic analysis of the N-ribosyl hydrolase activity of hCD157. Arch Biochem Biophys 564:156–163. [CrossRef]
- Kropotov A, Kulikova V, Nerinovski K, Yakimov A, Svetlova M, Solovjeva L, Sudnitsyna J, Migaud ME, Khodorkovskiy M, Ziegler M, Nikiforov A (2021) Equilibrative nucleoside transporters mediate the import of nicotinamide riboside and nicotinic acid riboside into human cells. Int J Mol Sci 22(3):1391. [CrossRef]
- Kropotov A, Kulikova V, Solovjeva L, Yakimov A, Nerinovski K, Svetlova M, Sudnitsyna J, Plusnina A, Antipova M, Khodorkovskiy M, Migaud ME, Gambaryan S, Ziegler M, Nikiforov A (2022) Purine nucleoside phosphorylase controls nicotinamide riboside metabolism in mammalian cells. J Biol Chem 298(12):102615. [CrossRef]
- Kulikova V, Shabalin K, Nerinovski K, Dölle C, Niere M, Yakimov A, Redpath P, Khodorkovskiy M, Migaud ME, Ziegler M, Nikiforov A (2015) Generation, release, and uptake of the NAD precursor nicotinic acid riboside by human cells. J Biol Chem 290(45):27124–27137. [CrossRef]
- Grant R, Berg J, Mestayer R, Braidy N, Bennett J, Broom S, Watson J (2019) A pilot study investigating changes in the human plasma and urine NAD+ metabolome during a 6 hour intravenous infusion of NAD. Front Aging Neurosci 11:257. [CrossRef]
- Zarei A, Khazdooz L, Madarshahian S, Enayati M, Mosleh I, Lin T, Yan B, Ufheil G, Wooster TJ, Abbaspourrad A (2021) Synthesis, stability, and bioavailability of nicotinamide riboside trioleate chloride. Nutrients 14(1):113. [CrossRef]
- Yang Y, Mohammed FS, Zhang N, Sauve AA (2019) Dihydronicotinamide riboside is a potent NAD+ concentration enhancer in vitro and in vivo. J Biol Chem 294(23):9295–9307. [CrossRef]
- Giroud-Gerbetant J, Joffraud M, Giner MP, Cercillieux A, Bartova S, Makarov MV, Zapata-Pérez R, Sánchez-García JL, Houtkooper RH, Migaud ME, Moco S, Canto C (2019) A reduced form of nicotinamide riboside defines a new path for NAD+ biosynthesis and acts as an orally bioavailable NAD+ precursor. Mol Metab 30:192–202. [CrossRef]
- Ciarlo E, Joffraud M, Hayat F, Giner MP, Giroud-Gerbetant J, Sanchez-Garcia JL, Rumpler M, Moco S, Migaud ME, Cantó C (2022) Nicotinamide riboside and dihydronicotinic acid riboside synergistically increase intracellular NAD+ by generating dihydronicotinamide riboside. Nutrients 14(13):2752. [CrossRef]
- Makarov MV, Migaud ME (2019) Syntheses and chemical properties of β-nicotinamide riboside and its analogues and derivatives. Beilstein J Org Chem 15:401–430. [CrossRef]
- Neamţu AS, Biţă A, Scorei IR, Rău G, Bejenaru LE, Bejenaru C, Rogoveanu O, Oancea CN, Radu A, Pisoschi CG, Neamţu J, Mogoşanu GD (2020) Simultaneous quantitation of nicotinamide riboside and nicotinamide in dietary supplements via HPTLC–UV with confirmation by online HPTLC–ESI–MS. Acta Chromatogr 32(2):128–133. [CrossRef]
- Kim HJ, Benner SA (2017) Prebiotic stereoselective synthesis of purine and noncanonical pyrimidine nucleotide from nucleobases and phosphorylated carbohydrates. Proc Natl Acad Sci U S A 114(43):11315–11320. [CrossRef]
- Kim HJ, Benner SA (2018) A direct prebiotic synthesis of nicotinamide nucleotide. Chemistry 24(3):581–584. [CrossRef]
- Kim HJ, Benner SA, Scorei IR (2018) Synthesis and stabilization of nicotinamide ribose and its derivatives. United. States Patent and Trademark Office (USPTO), Provisional.
- Kim HJ, Furukawa Y, Kakegawa T, Bita A, Scorei R, Benner SA (2016) Evaporite borate-containing mineral ensembles make phosphate available and regiospecifically phosphorylate ribonucleosides: borate as a multifaceted problem solver in prebiotic chemistry. Angew Chemie Int Ed Engl 55(51):15816–15820. [CrossRef]
- Scorei R (2012) Is boron a prebiotic element? A mini-review of the essentiality of boron for the appearance of life on Earth. Orig Life Evol Biosph 42(1):3–17. [CrossRef]
- Nielsen FH (2018) Boron in aging and longevity. In: Malavolta M, Mocchegiani E (eds). Trace elements and minerals in health and longevity. Book Series “Healthy Ageing and Longevity (HAL)”, vol. 8, Springer, Cham, Switzerland, 163–177. [CrossRef]
- Hunter JM, Nemzer BV, Rangavajla N, Biţă A, Rogoveanu OC, Neamţu J, Scorei IR, Bejenaru LE, Rău G, Bejenaru C, Mogoşanu GD (2019) The fructoborates: part of a family of naturally occurring sugar–borate complexes – biochemistry, physiology, and impact on human health: a review. Biol Trace Elem Res 188(1):11–25. [CrossRef]
- Biţă A, Scorei IR, Bălşeanu TA, Ciocîlteu MV, Bejenaru C, Radu A, Bejenaru LE, Rău G, Mogoşanu GD, Neamţu J, Benner SA (2022) New insights into boron essentiality in humans and animals. Int J Mol Sci 23(16):9147. [CrossRef]
- Biţă A, Scorei IR, Rangavajla N, Bejenaru LE, Rău G, Bejenaru C, Ciocîlteu MV, Dincă L, Neamţu J, Bunaciu A, Rogoveanu OC, Pop MI, Mogoşanu GD (2023) Diester chlorogenoborate complex: a new naturally occurring boron-containing compound. Inorganics 11(3):112. [CrossRef]
- Mitruţ I, Scorei IR, Manolea HO, Biţă A, Mogoantă L, Neamţu J, Bejenaru LE, Ciocîlteu MV, Bejenaru C, Rău G, Mogoşanu GD (2022) Boron-containing compounds in Dentistry: a narrative review. Rom J Morphol Embryol 63(3):477–483. [CrossRef]
- Neamţu AS, Biţă A, Scorei IR, Mogoşanu GD, Pisoschi CG (2021) Solubility, stability and degradation kinetics of nicotinamide riboside borate, a vitamin B3 derivative, in physiological solutions. Fiziologia (Physiology) 31(2):33–42. Available online: http://revista_fiziologia.umft.ro/archives/Fiziologia_2021_31_2102_Final.pdf.
Figure 1.
NARB mass spectrum: 11B (m/z 281); 10B (m/z 280).
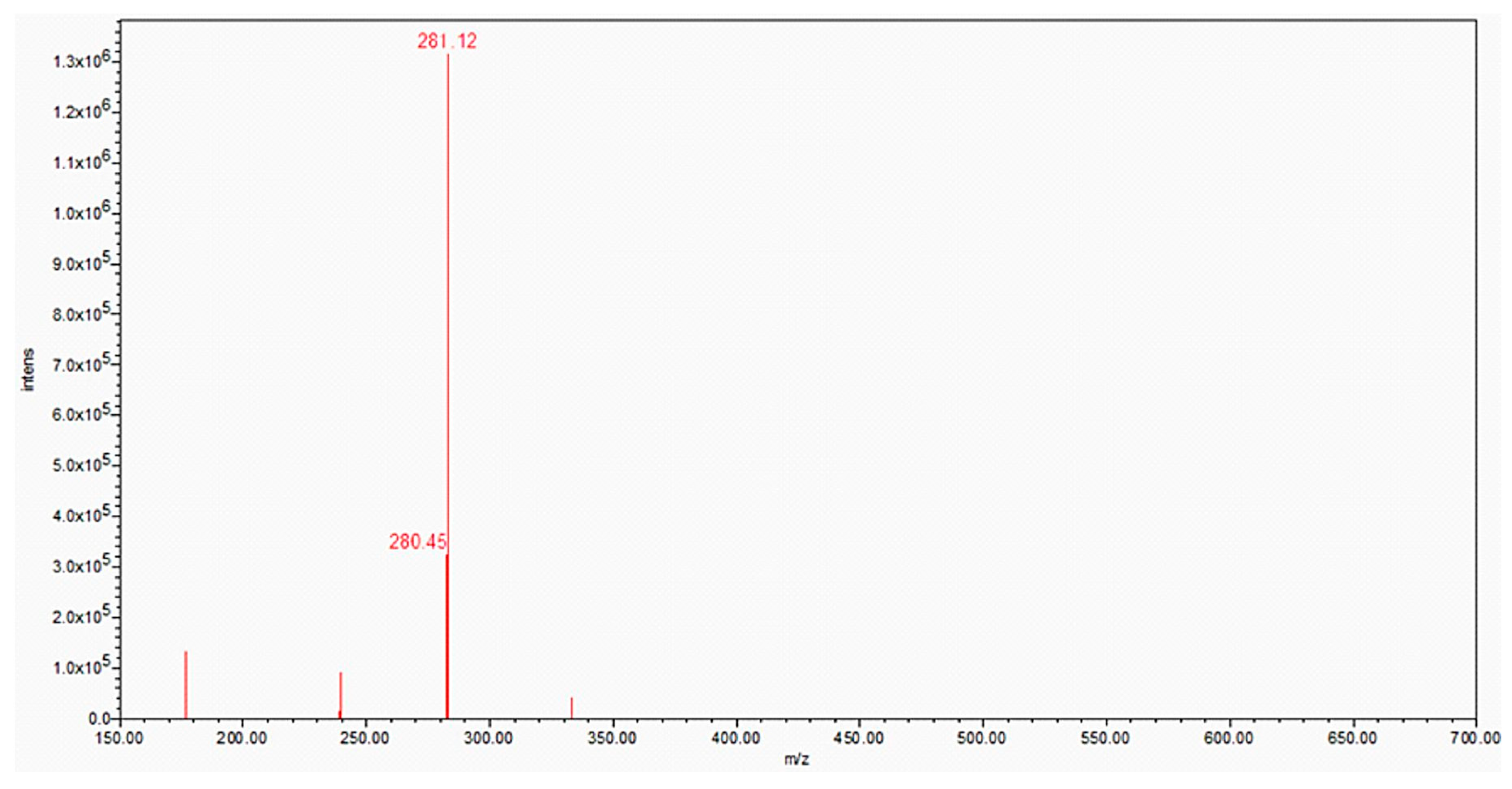
Figure 2.
pH stability for NARB.
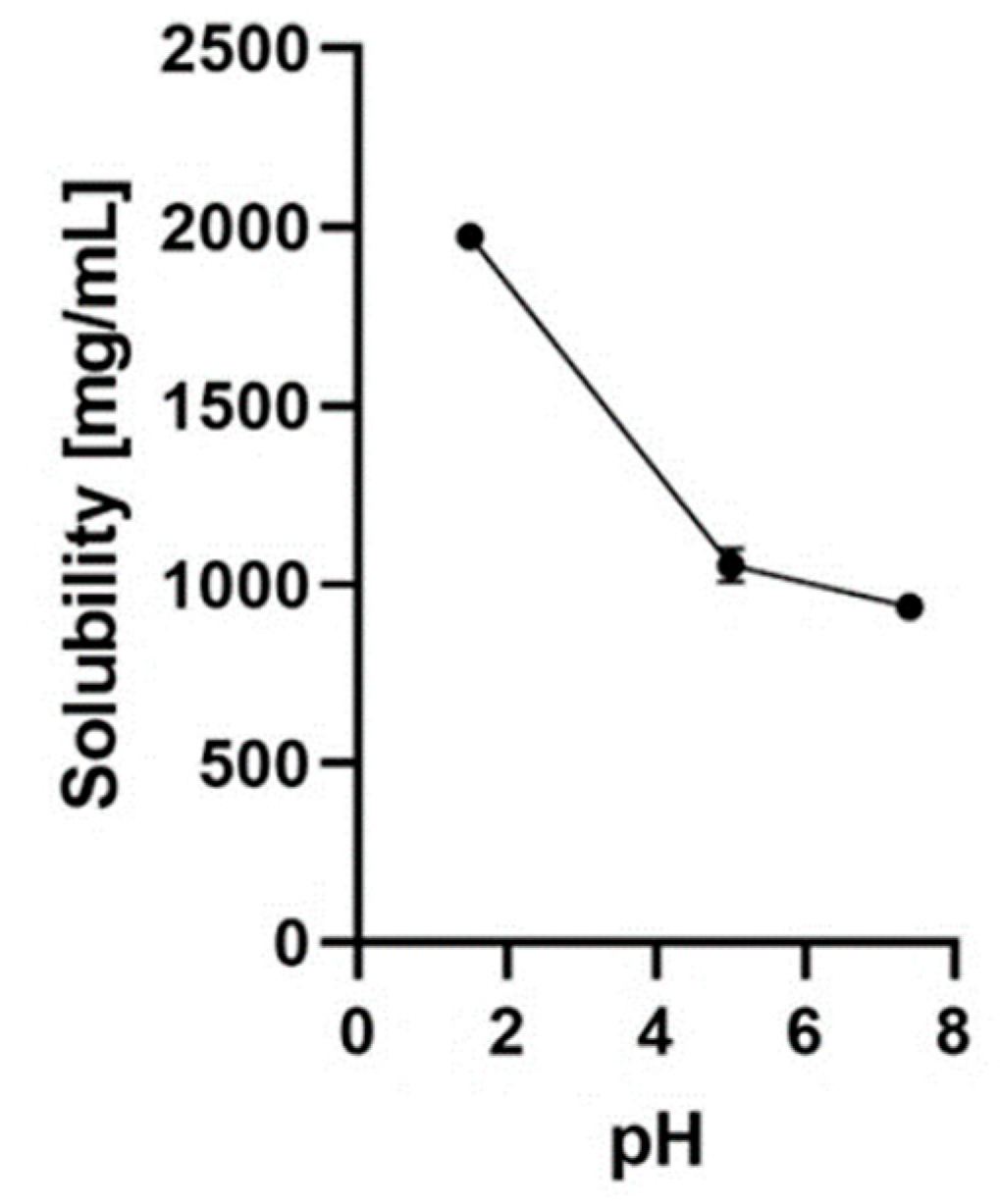
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



