
Submitted:
26 June 2023
Posted:
26 June 2023
You are already at the latest version
Abstract
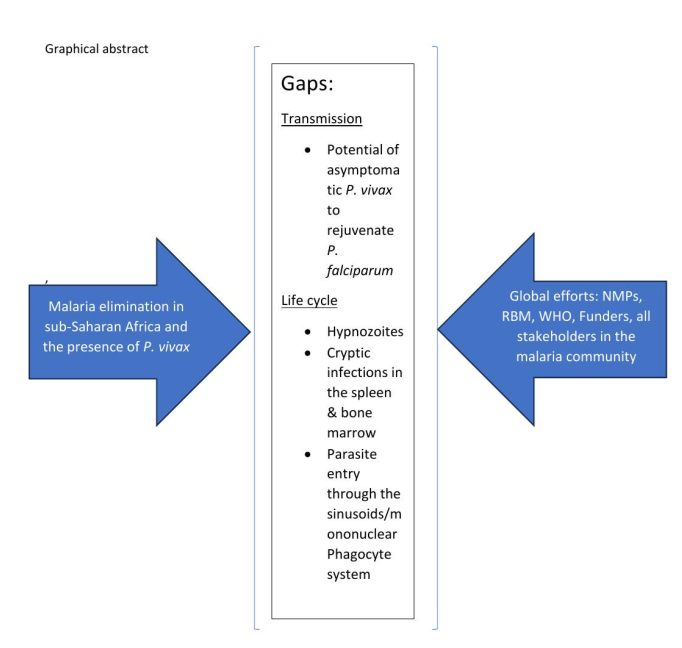
The global malaria community has picked up the theme of malaria elimination in greater than 90 % of the world’s population in the next decade. The recent reports of P. vivax in all sub-Saharan Africa, including Duffy negative individuals, threaten the efforts to achieve elimination. This is not only in view of strategies that are tailored only to P. falciparum elimination, but also due to currently revealed biological characteristics of P. vivax concerning the relapse patterns of hypnozoites and conservation of a large biomass at cryptic sites in the bone marrow and spleen. A typical scenario has been observed in Botswana from the period between 2008 to 2018, which palpably projects how P. vivax could endanger efforts at malaria elimination where the two parasites co-exist. The need for the global malaria community, NMPs, funding agencies and relevant stakeholders to engage in a forum to discuss and recommend clear pathways for sub-Saharan Africa regarding malaria elimination including P. vivax, is warranted.
Keywords:
P. vivax
; Botswana
; Malaria eliminatio
; P. vivax biology
; malaria and sinusoids
1. Introduction
The World Health Organization (WHO) Global Technical Strategy (GTS) for malaria is elimination by 2030 in 35 countries while a target of global eradication by 2040 has recently been announced (https://www.who.int/publications/i/item/WHO-CDS-GMP-2019.10, [1]. The new affirmation follows on the previous initiative of eradication launched in 2007 followed by the malERA initiative in 2011 [2,3,4]. The malERA elimination and eradication agenda has highlighted key challenges that need to be overcome to achieve elimination and eradication by the global community. These include a). Prioritizing how to detect the foci of the burden of disease and foci of drug resistance, and how they spread. b). Ensuring a clear understanding of the operational process of surveillance and response, based on scientific evidence. c). Ensuring that the biology of Plasmodium vivax (P. vivax) is well understood and constitutes part of the strategies for elimination in all countries including sub-Saharan Africa. d). Sustaining efforts at capacity building and engagement between scientists, policy makers and local actors so that the focus on elimination and eradication is clear and upheld [2,3,5].
In all sub-Saharan Africa, the current strategy on surveillance involves using either the Malaria Indicator Survey (MIS), Demographic and Health Indicator Survey (DHIS) or the Multiple Indicator Cluster Survey (MICS) as a sampling framework [6,7]. These surveys are modeled to obtain household information on malaria intervention coverage, prevalence of passive malaria cases, anemia and antimalaria drug use in children under 5 years of age and in pregnant women. The surveys are managed centrally and done every 2 - 4 years because of the expense involved, which makes it unrealistic to do it more frequently. In addition, the passive surveys miss out on asymptomatic infections that silently increase transmission and sustain it [8]. They are also not precisely standardized with respect to detection. This approach is palpably insufficient to provide the information on disease burden and transmission, yet such information is needed to implement a targeted intervention. In Africa, while these problems exist, an additional barrier to elimination is the fact that African countries have held on to the perception that the burden of non-falciparum malaria is minimal, which has obliquely caused strategies for elimination to be focused mainly on falciparum malaria. There is clear and established evidence of P. vivax and non-falciparum malaria presence in Africa [9,10,11]. In addition, P. vivax has been reported to cause severe malaria disease that is comparable to that in P. falciparum across all age groups [12]. It is time for sub-Saharan Africa NMPs, to rethink strategies on elimination to include P. vivax and non-falciparum malaria. The good news is that for elimination, WHO recommends active case detection (ACD) to be implemented [13]. This recommendation makes it easier for policy implementers in Africa to carefully think through malaria elimination strategies that include non-falciparum malaria, particularly P. vivax [14]. The focus of the current article is to project rethinking strategies based on current information on P. vivax biology and data obtained over six years in active surveys in Botswana on P. vivax transmission.
2. Human Plasmodium parasites life cycle and their recently found tissue niches
2.1. Sporozoite inoculation and invasion of tissues
The epidemiology of malaria depends on a complex interplay between the intermediate host, parasite, definitive vectors, and the environment [15,16]. These factors differ from one geographic region to another, so the epidemiology differs accordingly. There are five Plasmodium species of the phylum Apicomplexa, order Haemosporidae, that cause malaria in humans: P. falciparum, P. vivax, P. ovale curtisi, P. ovale wallikeri, (P. ovale), P. malariae and P. knowlesi [17]. An infection is initiated when a female anopheline mosquito infected with the parasite, inoculates sporozoites estimated to be between 15-123 under the skin of an individual, while probing for a blood vessel to take a meal [18,19,20,21,22]. From under the skin, the sporozoites get activated based on the environmental milieu, express proteins required for gliding motility and cell traversal, and travel through the blood to the liver using the sinusoids as entry points to hepatocytes [23,24,25,26]. The sinusoids are lined with resident macrophages, the Kupffer cells, which aid in the transmigration of the sporozoites by a gliding motility [27], through the sinusoids to reach hepatocytes, [28,29]. Proteins for cell traversal enable entry through the hepatocytes until one is selected for development. Alternatively, the sporozoites may also use the lymphatic system to enter tissues with the help of the sinus lining monocyte macrophage system [30].
2.2. Entry into extrahepatic tissues/cryptic sites
It has recently been established, that sporozoites can access and develop in extrahepatic tissues of the body, including the spleen, the bone marrow, and lungs [19,31,32,33,34]. This raises the question of how the parasites enter these non-hepatic tissues or cryptic sites. In some vertebrates (birds and lizards), sporozoites can enter tissues and differentiate further, using the monocyte-macrophage phagocytic system including in the skin [27]. The Kupffer cells which are the monocyte macrophage system in the liver, are known to be deficient in the production of reactive oxygen species (ROS), fail to respond to interferon-gamma in an infection, are defective in secreting microbicidal molecules and serve as permissive host for intracellular parasites [35]. This characteristic has also been seen in spleen red pulp macrophages [36]. Comparatively, it may be that the culpable cells permitting intracellular parasite migration in non-hepatic tissues, are the sinus lining monocyte macrophages in the bone marrow, Langerhans cells in the skin, and alveolar macrophages in lung cells (Figure 1). These require further interrogation, as they have a bearing on the role of relapses of hypnozoites in P. vivax and P. ovale malaria transmission [34] and asymptomatic infections. These could also provide a new avenue for inhibiting malaria transmission by blocking, using monoclonal antibodies.
2.3. Development of asexual and sexual forms
The development within the tissues is impacted by the specific tissue environment. Rapid transcriptional changes occur in each parasite depending on the tissue milieu that determines the transmission success [23]. In the liver, the sporozoites grow into liver schizonts that undergo schizogony and transform into merozoites, which are the red blood cell (rbc) infective forms [37]. The merozoites exit hepatocytes through the sinusoids and Kupffer cells, to infect rbcs in the form of merosomes, which are merozoites enclosed in a membrane [38]. The merozoites use gliding motility similarly as sporozoites, for entry into rbcs [39].
In P. vivax and P. ovale, some merozoites cease replicating and enter a dormant hypnozoite stage, which can relapse upon a trigger to enter a new replication mode to generate new merozoites [40]. Recent reports reveal that P. vivax hypnozoites express genes towards gametocyte stage early in their development and seem programmed to commit towards gametocytogenesis upon activation [41]. In this way, they serve as parasite reservoirs for propagation. In the rbcs, the merozoites go through early ring forms, late trophozoites and blood schizonts which burst the rbcs as they expand to initiate a new cycle of infection as merozoites (Figure 1A). P. vivax merozoites are restricted to invade reticulocytes while P. falciparum parasites are not restricted although they show preference for younger rbcs in an invasion [42]. The schizonts contain about 16 – 32 merozoites [43]. The cycle of rbc invasion by merozoites leads to a paroxysm of fever which repeats every 24 hours in P. knowlesi, 48 hours in P. falciparum, P. vivax and P. ovale, and 72 hours in P. malariae [43]. These paroxysms give the respective descriptors of quotidian, tertian and quartian malaria respectively, to the disease. During the asexual rbc cycle, some of the merozoites develop into male and female immature gametes called gametocytes, which mature to initiate the sexual cycle [44]. The immature forms which are not visible in the peripheral circulation are present in the bone marrow and spleen where they mature and re-enter into circulation [37]. In P. falciparum, five developmental forms have been observed and characterized as stages 1-V, although only the stage V forms are seen in the peripheral circulation [45]. In P. vivax however, all the asexual and sexual stages of the parasite can be seen in the peripheral circulation [46]. The mature gametocytes are taken up during feeding by anopheline mosquitoes and develop through gametocytogenesis to produce male microgametes and a female macrogamete. The microgametes undergo exflagellation before fertilization with the macrogamete into zygotes [47] in the mosquito midgut (Figure 1A). There is genetic recombination during this period leading to a diploid genome. The zygotes further develop into motile ookinetes in the midgut. Subsequently, the ookinetes move from the midgut epithelium to the basal lamina and develop into oocysts in a sporogonic cycle. In the oocysts, the sporogony continues, generating numerous sporozoites [48,49]. When mature, the sporozoites are released from the oocysts after rupture into the hemolymph and migrate into the salivary gland ready for a new cycle of inoculation and infection in humans [50] (Figure 1A). The genes expressed in the developmental cycle of the parasite appear to be species specific, based on the host and vectors that the parasite interacts with during the life cycle [50]. These differences are very important in the life cycle of P. vivax and P. ovale that have hypnozoite stages. The gene expression pattern of the parasites is modulated based on the host factors to allow early release and viability of the gametocytes and sustain transmission (46). P. vivax is also established to adapt very well to climate variations and multiple vectors, which adds to the difficulty associated with eliminating the parasite [49] (Cogswell fb, 1992) [51].
In the extrahepatic tissues (bone marrow and spleen), all the blood stages of the P. vivax, have been observed to be present. The immature gametocytes are reported to be enriched in the bone marrow parenchyma and the spleen [52,53,54], while the spleen is further enriched with late-stage asexual forms, the trophozoites and schizonts [34]. Schizogony in the bone marrow and spleen can also generate merozoites additionally, for rbc infection [34]. Schizonts have also been seen to accumulate in the lungs and adipose tissues [53,54]. These extrahepatic sites seem to be reservoirs for transmission and sources of sustenance of transmission, as they are protected from immune attack, and drug treatment and occur in asymptomatic individuals [27,34,55]. Whether these are synchronous with the liver stages in the release of merozoites into circulation, is currently unknown.
2.4. The reticuloendothelial (RES)/mononuclear phagocyte system (MPS) in Plasmodium life cycle
The term ‘RES´ was originally coined by Karl Albert Ludwig Aschoff in 1924 [56] to reference cells involved in phagocytosis. The Reticulo referred to the tendency of these large cells to connect by cytoplasmic projections to form a network or reticulum, while endothelial referred to the closeness to the vascular endothelium. In later years, the RES was renamed as cells of the mononuclear phagocyte system (MPS) [57] based on their function to phagocytose, morphology and kinetics. Currently, they are known to be made up of three key cells: monocytes, macrophages and dendritic cells based on function and phenotype [58]. The cells may be resident in their tissues or recruited, lining the sinusoids of their respective tissues. Macrophages are derived from embryonic progenitors starting from the yolk sac and fetal monocytes, which are subsequently distributed throughout the developing tissue through the peripheral circulation [59,60]. These cells subsequently self-renew in the absence of adult hematopoiesis [61]. Monocytes on the other hand, are derived from monocyte progenitor cells and dendritic cells from a dendritic cell precursor (adult hematopoietic stem cell precursors) [61]. Based on this classification, the previously classified RES cells: Kupffer, microglia, alveolar macrophages, splenic red pulp macrophages, Langerhans cells and lymphatic cells are all considered as belonging to the macrophage type of cells as they are embryonically derived [61] (Figure 1B). An important point though is that while their origins are the same, the gene expression patterns are dependent on the tissues in which they are resident [62]. Interestingly, in all the indicated tissues, Plasmodium sexual and asexual parasites have been observed as discussed below, but much more studied in P. vivax.
2.5. RES/MPS as host sites of cryptic infections by Plasmodium parasites
Blood and Liver: Sporozoites enter the blood stream in the peripheral circulation using a gliding motility [39]; a process that is unique to apicomplexan parasites. The parasites stay in the blood, until they reach liver sinusoids where they are initially sequestered. By interaction with heparan sulphate proteoglycans from Kupffer and stellate cells, they enter hepatocytes enclosed in a parasitophorous vacuole of host origin, by gliding motility through the Kupffer cells [28,49,63] (Figure 1B). They finally settle in one hepatocyte to signal an end to the migration and continue development into schizonts. It has been suggested that cell entry within a parasitophorous vacuole limits cellular damage during migration, reduces the risk of an inflammatory response to the parasite and is used by the parasite for nutrient intake and efflux of waste materials [26,64]. Sinusoids present in tissues of the RES have been reported to harbor sexual and asexual forms of the Plasmodium parasites through human autopsy studies and rodent experiments [53,54]. A question that needs further interrogation is: do Plasmodium asexual and sexual forms have preference for the use of sinusoids to enter tissues because of the peculiar role played by macrophages in sinusoid entry?
Bone marrow and the spleen: Bone marrow is the major site for erythropoiesis while the spleen filters blood to remove old and damaged erythrocytes, particles, and pathogens [65,66]. The spleen can contribute to erythropoiesis, in a time of need such as blood loss or trauma. During erythropoiesis and in the filtering function of the spleen, the macrophages of the sinusoids and fenestrated endothelial cells play key roles [67]. Bone marrow macrophages modulate erythropoiesis by close apposition to the developing erythroblasts, while in blood filtration in the spleen, the macrophages recognize pathogens from pattern recognition receptors and pathogen associated molecular patterns for removal, during the open and slow microcirculation through the sinusoids [68]. Parasite entry into the bone marrow and spleen is also suggested to be by gliding motility, like that of leukocytes during their transit to sites of acute inflammation, aided by tissue sentinel macrophages and molecular signals from the pathogens and damaged cells [69,70,71,72,73]. Therefore, a role of macrophages assisting in parasites entry as mentioned previously, would be highly significant in the life cycle of the parasite. It has also been observed that merozoite infected rbcs could home into the bone marrow and spleen in a receptor mediated process, due to vascular leakages which provide signals for invasion [54]. These observations were seen in both the rodent P. berghei parasite and human P. falciparum, indicating that the bone marrow and spleen are major sites for parasite development in the Plasmodium life cycle. The unique biology of P. vivax and P. ovale in having hypnozoite stages presents an even more intriguing scenario considering that this stage can cause relapse after several years [74,75]. Although hypnozoites have been attributed to a source from the liver, the current understanding of the life cycle may point to sources of cryptic relapses outside of the liver. A large proportion (> 90%) of chronic P. vivax infections are asymptomatic, subpatent and submicroscopic [74]. These non-tangible infections form the largest biomass of P. vivax infections, and have been observed to be present in the spleen accounting for close to 98% (34) of the biomass. In addition, P. vivax genes involved in sequestration and cyto-adherence have been found to be spleen dependent [76,77]. The spleen and bone marrow therefore serve as important sites in the life cycle of P. vivax particularly. These cryptic infections must be considered carefully in surveillance strategies as they would ensure that tools for intervention are optimal. Clearly, the use of passive case detection would fail to ensure malaria elimination as are done in most malaria endemic sub-Saharan African countries, focusing only on P. falciparum malaria.
Lymphatic system: The lymphatic system maintains tissue fluid homeostasis and coordinates the transport of immune cells to the tissues [78]. The system has been observed to harbor asexual plasmodium parasites of all stages including merozoites [20,79]. The source of the parasites could be from sporozoites at the time of inoculation or directly from infected rbcs in the circulation [20], considering that the lymphatic system is closely linked with the hepatic sinusoids [80]. In the lymphatic system parasite development could occur leading to the generation of merozoites, which can stay latent for some time before joining the peripheral circulation to invade new rbcs, sustaining the life cycle [79]. It is reported that the subscapular macrophages of the lymph nodes could be the main cells permitting intracellular pathogens entry, which have been seen for viruses as well [81].
2.6. Plasmodium vivax conundrum and strategic activities in an infection
A pathogen’s ability to enable easy uptake by host phagocytes, is described as the Trojan Horse model [82]. It appears that cells of the MPS are frequently used as Trojan Horses for entry into host tissues by intracellular parasites [83]. Prolonged environmental changes and interactions of pathogens with host cells are key drivers of adaptation and enablers of spread or resilience in an infection [84,85]. These changes ensure some level of disengagement with or manipulation of the host immune system to ensure survival. The Plasmodium sporozoite has three main biological characteristics: to be motile, traverse cells and invade host cells [86]. For these functions, the sporozoites and merozoites have characteristic secretory granules which are dense granules for modifying host cells, rhoptries for forming the parasitophorous vacuoles and micronemes for rbc invasion [87]. The sporozoites have been definitively seen with rhoptries and micronemes, while merozoites have all three granules. Gliding motility is the main enabler of the three characteristics [87]. These characteristics are shared by the Plasmodium parasites. These biological characteristics and cryptic infections will be discussed considering data obtained from Botswana from 2012 to 2018 and malaria elimination.
3. Trends in malaria transmission in Botswana from the year 2008 - 2018
The defined malaria elimination and eradication strategy is for countries to identify the foci of Plasmodium infections, treat the population and sustain surveillance to track any local transmission. From 2012 to 2018, we carried out systematic active studies on malaria transmission profile in Botswana, in collaboration with the National Malaria Program (NMP), to understand epidemiological outbreaks and provide the data that could allow the NMP to target hotspots of infection covering all Plasmodium species. We present the findings together with the epidemiological outbreaks of malaria documented by the NMP from 2008 to 2012, and the predicted outlook in 2018. The findings from these studies reveal a profoundly complex transmission pattern of P. vivax and its interaction with P. falciparum for survival [88,89]. The observations are important to be considered and factored into planning strategies for malaria elimination [16].
Botswana is a member of the Elimination 8 countries in Southern Africa and is part of the front-line countries that targeted malaria elimination in 2015, after concerted efforts to reduce malaria disease burden to < 1 per 1000 population by 2012 [90,91].
These efforts included increased awareness campaigns towards uptake of the use of Long-Lasting Insecticide Treated Nets (LLINs), Indoor Residual Spraying (IRS), increased use of RDTs for diagnosis [91] and introduction of artemisinin combination therapy (ACT) for treatment. Unfortunately, the projected target for elimination could not be achieved, because of challenges in detecting asymptomatic infections and the sporadic cases of malaria localized in the north, eastern, southeastern and parts of the southern regions [92] (Figure 2). The failure was partly because the surveillance strategy had been based on passive case detection which did not account for asymptomatic infections and non-falciparum malaria [93]. The burden of malaria disease has traditionally been in the northern part of the country which is wetter and warmer compared to the southern part of the country. Occasionally the relatively wetter parts to the eastern, southeastern, and southern parts of the country also experience sporadic transmission, depending on the rainfall patterns [92]. Rainfall is seasonal and occurs between November and May [92,94] so the transmission season occurs within the same period.
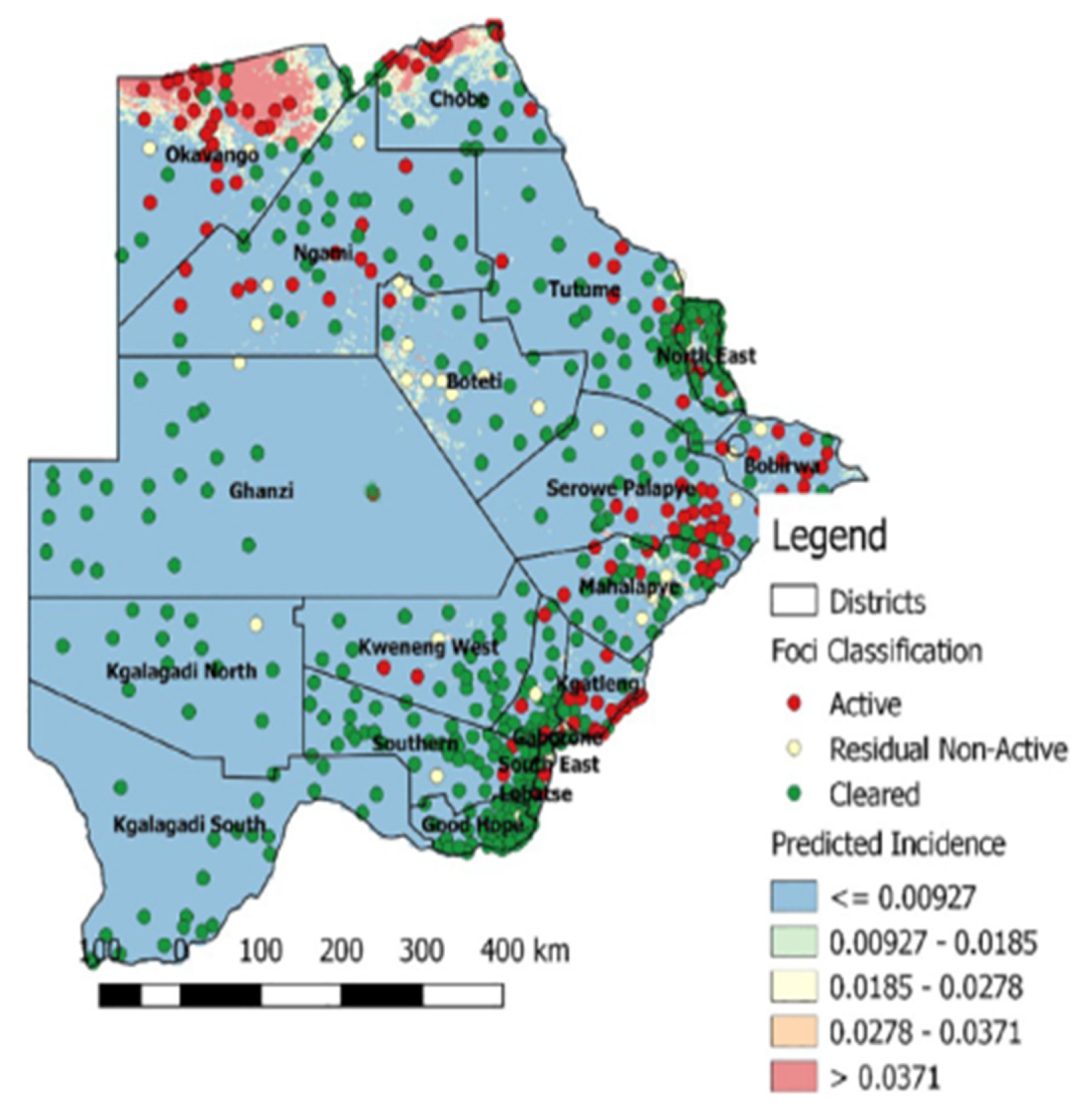
In 2008 and 2009, the foci of malaria disease burden were in three districts: Okavango, Chobe, and Ngami in the north (Figure 2). By 2010, the burden in Okavango had diminished a bit with the foci localized in Ngami and Chobe (Figure 2). At the same time a small node of transmission was beginning from Ngami towards Boteti. This became significant in 2011 (Figure 2). By 2012, the entire stretch of land from the northeastern section towards the southeastern border regions were experiencing significant and sporadic malaria outbreaks. These outbreaks were seen despite the general reduction in malaria disease burden <1 per 1000 populations, with reference to P. falciparum as the main causative parasite. It was a puzzling occurrence and after several discussions on the way forward, it was decided that an active survey was needed to unravel some elements of the puzzle. In 2012, considering that the target for malaria elimination was 2015, an active survey was done across the country, within the transmission period and species-specific nested PCR was used for detection of all Plasmodium species and asymptomatic infections. The result of this survey was profound as published previously [88]. We affirmed that indeed P. falciparum was mainly localized in the Kavango and Ngami regions (no samples were collected in Chobe at the time), with some asymptomatic infections. However, we observed for the first time that P. vivax asymptomatic infections were present and localized at hotspots across the country [88]. Areas where the asymptomatic P. vivax burden were found mirrored the observed outbreaks, in a way that suggested that they were part of the epidemic outbreaks that were recorded. In addition, areas in the South (Kweneng), that had seen some occasional outbreaks but deemed not to be important (Figure 2), were discovered as major foci of P. vivax asymptomatic infections. These findings corroborated the observed characteristics of P. vivax transmission, which is that P. vivax asymptomatic infections significantly contribute to transmission and account for the majority in low endemic settings, as observed elsewhere [95,96]. In addition, they were highest in areas where P. falciparum were not detected by nested PCR, indicating a reciprocal interaction between the two parasites as have been observed generally. Since samples were collected in the transmission season, we could not make a claim to relapsing P. vivax hypnozoites. Two questions were on our minds: if these were partly hypnozoites, that relapsed a. what caused the triggers for activation; and b. were these relapses from liver hypnozoites only? Since the outbreaks were spread out over a large area, within the same period, we wondered if the trigger/triggers were something in the environment that led to a physiological response and activation or due to an innate biological clock that spontaneously induced an activation. We also wondered whether the parasites responses to a trigger were spontaneous within a giving geographical area (what we would term a ‘Spoke wheel effect’) (Figure 3) or relayed. Following the extension of the elimination agenda to 2018, in 2016, to insight into the nature of the observed P. vivax parasites, we again, we embarked on another survey at the same sites as we did previously and included Chobe that we had previously missed. This time we collected samples all year round from August 2016 to October 2017, and added qPCR to increase the sensitivity of detection. The findings were particularly interesting. We noted that not only was P. vivax still present in the sites we had previously seen, but also for some unknown reasons P. falciparum had re-appeared in all the places that P. vivax had been seen previously. In addition, with the return of P. falciparum, the burden of P. vivax diminished but did not disappear, allowing for coexistence within the communities. Interestingly, we identified P. vivax in the non-transmission season, indicating a role of hypnozoites relapses in the population. The active survey reflected in the epidemic outbreak for the 2016/2017 period, showing an 80% increase in malaria transmission for that period [97], (WHO, world Malaria report, 2018). In 2018, the NMP mapped out areas of active malaria transmission across the country, using a modelling approach. It was evident that the predicted active places were consistently in areas where P. vivax asymptomatic infections had been detected. Not surprisingly, the elimination date was revised to 2025, following a review in 2021 by the Malaria Elimination Oversight Committee (MEOC) of the WHO, to include Botswana in the Elimination 2025 (E-2025) countries [98].
Figure 2.
Annual patterns of malaria disease prevalence in Botswana: from 2008 to 2012 (Source: NMP Botswana).
Figure 2.
Annual patterns of malaria disease prevalence in Botswana: from 2008 to 2012 (Source: NMP Botswana).
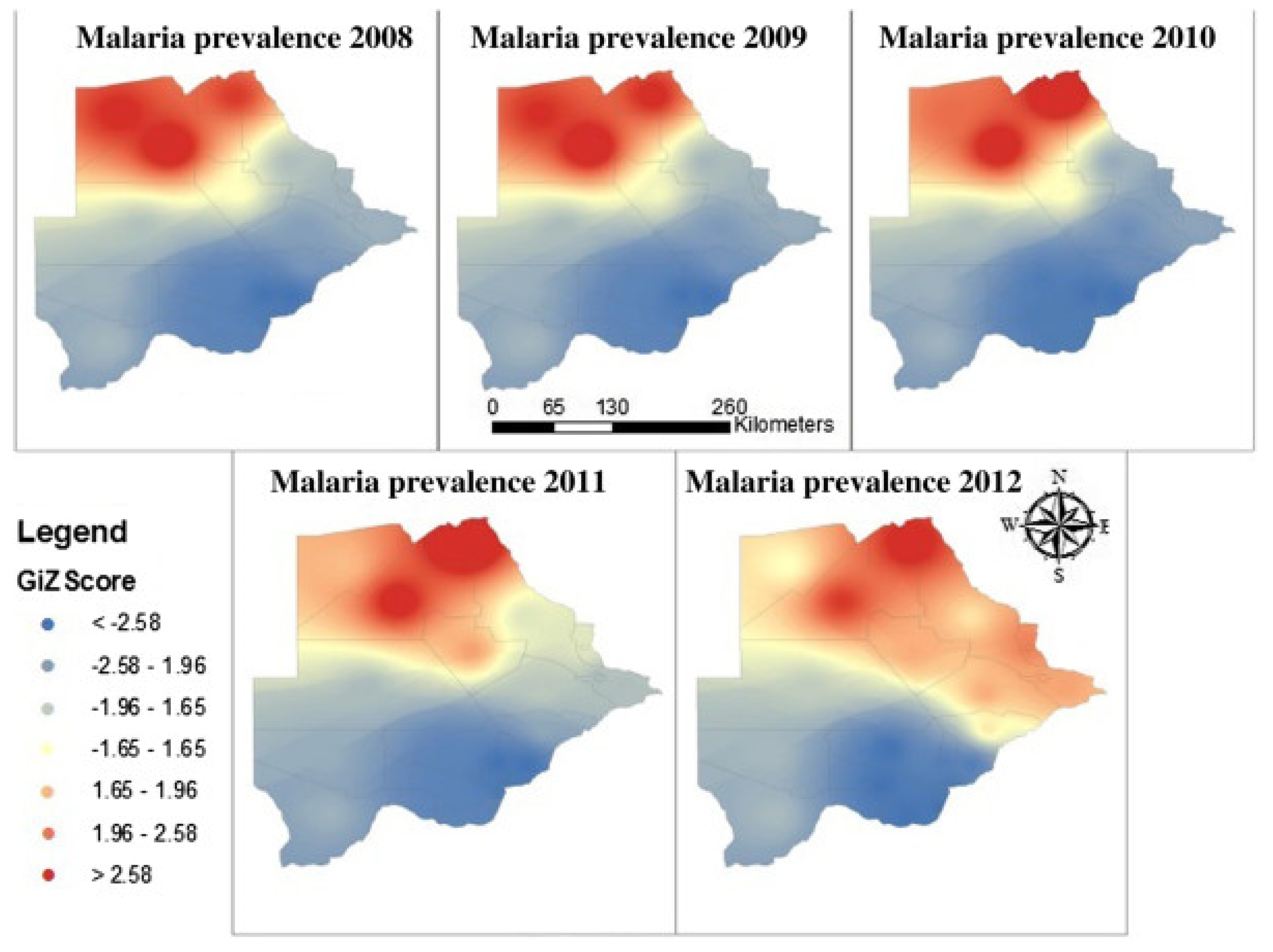
Figure 3.
Asymptomatic Plasmodium infections in 2012 and 2016-2017.
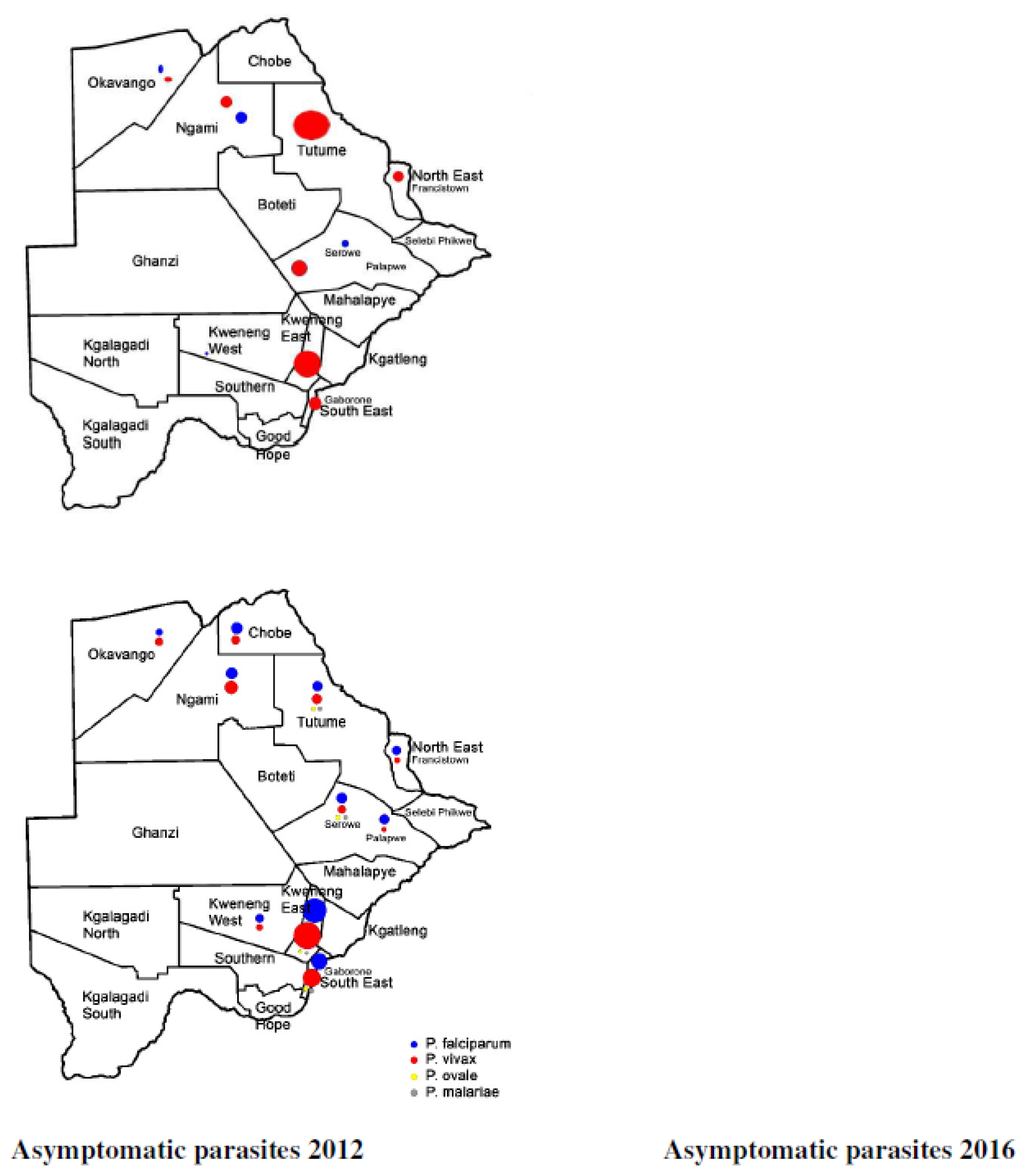
Figure 4.
Predicted malaria prevalence 2018 (Source: NMP Botswana).
4. Discussion
We have presented the pattern of malaria disease outbreaks in Botswana and their congruence with foci of P. vivax infection across the country [88,89] This pattern has not been previously seen, which raises the need for critical thinking of trends of malaria disease outbreaks in Botswana and areas in Africa with low endemicity, taking into consideration the probable role of P. vivax. We have shown here that active malaria survey under low transmission or elimination agenda, is a powerful tool that can presage the scenario for epidemic outbreaks. In all the surveys presented here, the asymptomatic parasites detected from the active survey precisely mirrored the epidemic outbreaks that followed. This affirmed previous observations that P. vivax asymptomatic infections contribute to malaria disease transmission [74,99,100,101]. We also observed for the first time that P. vivax asymptomatic infections can drive the re-emergence of P. falciparum, in an area, although the mechanism by which this occurs is currently unknown. It is clear that, P. vivax parasites once established in an area seem entrenched and will require vigorous test and treat approach using anti-blood stage and tissue stage drugs to eliminate them. A P. vivax strategy using a sero-diagnostic tool as proposed recently, will be needed in the elimination agenda [102]. Considering that all African countries that have reported the presence of P. vivax have also reported on the presence of Duffy negative infected individuals, the road to malaria elimination in Africa will not be easy [9]. This demands rethinking strategies to consider all malaria species as targets for elimination as a must do activity [14].
The new biological characteristics identified in P. vivax life cycle with cryptic infections detected in extrahepatic sites, (spleen, bone marrow and the lymphatic system) present a new conundrum. It is of interest that in all the cryptic sites, the sinusoidal system is used by the parasites to enter tissues. The sinusoids have a unique anatomical structure of having the monocyte macrophage system present in addition to the fenestrated endothelium [38]. All the tissues are therefore involved in the defense of the body, particularly the spleen, where the largest biomass of P. vivax infection has been found. Could it be that the Trojan Horse model approach described previously, is a parasite strategy for infection that has not been duly considered [82]? If the parasites adapt successfully to using the defense tools in tissues as a way of escape, then clearly effective counter measures such as anti-blood stage and tissue drugs are needed that would not be restricted as with the 8-aminoquinolines in G6PD deficiency. Leishmania donovani and related species that cause visceral leishmaniasis, are known to spread throughout the body using the mononuclear phagocyte system [103,104]. In these parasites the growth in the MPS is vigorous leading to the destruction of the tissues and dissemination in the body. In the spleen splenomegaly is seen before the tissue destruction [105]. The initial immune response to an infection in mice is TH1 dependent which subsides from TH2 responses to reduce inflammation but leads to a chronic infection and tissue destruction [106,107]. In humans it appears the TH1/TH2 balance fails such that growth continues vigorously, leading to death [108]. It is interesting that the two tissues in which high numbers of P. vivax parasite biomass have been seen (spleen and bone marrow) are also hematopoietic. When one considers that P. vivax is restricted to the use of reticulocytes, it may be that the parasites modulate the hematopoietic tissues to increase erythropoiesis, to enable them to thrive. This mechanism of infection has been demonstrated in L. donovani infections in hamsters [109]. The spleen is known to supplement hematopoiesis under trauma and stress [67]. In a study done in Colombia, under low endemic settings where P. vivax and P. falciparum exist, all patients with uncomplicated malaria seen with hepato-splenomegaly were P. vivax infected [110]. The frequency of pallor and hemoglobinuria were higher in P. vivax than in P. falciparum infections [110]. These clinical signals in P. vivax uncomplicated malaria should be documented in clinical presentations to facilitate diagnosis of P. vivax infections under low endemic settings. The mechanism of infection of P. vivax parasites in extra-hepatic tissue warrants thorough investigations.
5. Conclusion
We have shown here that P. vivax presence in the low transmission environment in Botswana appears entrenched. This could be attributed to the complex biology of the parasite in the human host. The Global Health community needs to have a discourse on this and provide guidelines for NMPs to follow. Without such an input for Africa NMPs, the path to elimination will be formidable. This engagement should also consider available diagnostic options, active case detection and treatment with 8-aminoquinolines. Bottle necks and challenges that program implementors face to put into practice WHO drug recommendations for radical cure of P. vivax infections, must be assessed and recommendations provided.
Funding
This work was Funded by Merck Global Health Institute.
Acknowledgments
The authors acknowledge the support of the staff of the National Malaria Program Botswana and the Pan African Vivax and Ovale Network (PAVON).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Feachem, R.G.A.; Chen, I.; Akbari, O.; Bertozzi-Villa, A.; Bhatt, S.; Binka, F.; Boni, M.F.; Buckee, C.; Dieleman, J.; Dondorp, A.; et al. Malaria eradication within a generation: Ambitious, achievable, and necessary. Lancet 2019, 394, 1056–1112. [Google Scholar] [CrossRef] [PubMed]
- Rabinovich, R.N.; Drakeley, C.; Djimde, A.A.; Hall, B.F.; Hay, S.I.; Hemingway, J.; Kaslow, D.C.; Noor, A.; Okumu, F.; Steketee, R.; et al. malERA: An updated research agenda for malaria elimination and eradication. PLoS Med. 2017, 14, e1002456. [Google Scholar] [CrossRef] [PubMed]
- malERA Refresh Consultative Panel on Tools for Malaria Elimination. malERA: An updated research agenda for diagnostics, drugs, vaccines, and vector control in malaria elimination and eradication. PLoS Med. 2017, 14, e1002455. [Google Scholar] [CrossRef] [PubMed]
- Dowdle, W.R.; Cochi, S.L. The principles and feasibility of disease eradication. Vaccine 2011, 29 (Suppl. S4), D70–D73. [Google Scholar] [CrossRef]
- malERA Refresh Consultative Panel on Health Systems and Policy Research. malERA: An updated research agenda for health systems and policy research in malaria elimination and eradication. PLoS Med. 2017, 14, e1002454. [Google Scholar] [CrossRef]
- malERA Refresh Consultative Panel on Characterising the Reservoir and Measuring Transmission. malERA: An updated research agenda for characterising the reservoir and measuring transmission in malaria elimination and eradication. PLoS Med. 2017, 14, e1002452. [Google Scholar] [CrossRef]
- Alegana, V.A.; Wright, J.; Bosco, C.; Okiro, E.A.; Atkinson, P.M.; Snow, R.W.; Tatem, A.J.; Noor, A.M. Malaria prevalence metrics in low- and middle-income countries: An assessment of precision in nationally-representative surveys. Malar J. 2017, 16, 475. [Google Scholar] [CrossRef]
- malERA Refresh Consultative Panel on Characterising the Reservoir and Measuring Transmission. malERA: An updated research agenda for characterising the reservoir and measuring transmission in malaria elimination and eradication. PLoS Med. 2017, 14, e1002452. [Google Scholar] [CrossRef]
- Quaye, I.K.; Aleksenko, L.; Oeuvray, C.; Yewhalaw, D.; Duah, N.; Gyan, B.; Haiyambo, D.H.; Dongho, G.B.D.; Torgby, R.A.; Amoah, L.; et al. The Pan African Vivax and Ovale Network (PAVON): Refocusing on Plasmodium vivax, ovale and asymptomatic malaria in sub-Saharan Africa. Parasitol. Int. 2021, 84, 102415. [Google Scholar] [CrossRef]
- Yman, V.; Wandell, G.; Mutemi, D.D.; Miglar, A.; Asghar, M.; Hammar, U.; Karlsson, M.; Lind, I.; Nordfjell, C.; Rooth, I.; et al. Persistent transmission of Plasmodium malariae and Plasmodium ovale species in an area of declining Plasmodium falciparum transmission in eastern Tanzania. PLoS Negl Trop. Dis. 2019, 13, e0007414. [Google Scholar] [CrossRef]
- Ruas, R.; Pinto, A.; Nuak, J.; Sarmento, A.; Abreu, C. Non-falciparum malaria imported mainly from Africa: A review from a Portuguese hospital. Malar J. 2017, 16, 298. [Google Scholar] [CrossRef]
- Naing, C.; Whittaker, M.A.; Nyunt Wai, V.; Mak, J.W. Is Plasmodium vivax malaria a severe malaria?: A systematic review and meta-analysis. PLoS Negl Trop. Dis. 2014, 8, e3071. [Google Scholar] [CrossRef]
- Moonen, B.; Cohen, J.M.; Snow, R.W.; Slutsker, L.; Drakeley, C.; Smith, D.L.; Abeyasinghe, R.R.; Rodriguez, M.H.; Maharaj, R.; Tanner, M.; et al. Operational strategies to achieve and maintain malaria elimination. Lancet 2010, 376, 1592–1603. [Google Scholar] [CrossRef] [PubMed]
- Lover, A.A.; Baird, J.K.; Gosling, R.; Price, R.N. Malaria Elimination: Time to Target All Species. Am. J. Trop. Med. Hyg. 2018, 99, 17–23. [Google Scholar] [CrossRef]
- Frost, W.H. Some conceptions of epidemics in general by Wade Hampton Frost. Am. J. Epidemiol. 1976, 103, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Coosemans, M.; Wery, M.; Mouchet, J.; Carnevale, P. Transmission factors in malaria epidemiology and control in Africa. Mem. Inst. Oswaldo Cruz 1992, 87 (Suppl. S3), 385–391. [Google Scholar] [CrossRef]
- Sutherland, C.J.; Tanomsing, N.; Nolder, D.; Oguike, M.; Jennison, C.; Pukrittayakamee, S.; Dolecek, C.; Hien, T.T.; do Rosario, V.E.; Arez, A.P.; et al. Two nonrecombining sympatric forms of the human malaria parasite Plasmodium ovale occur globally. J. Infect. Dis. 2010, 201, 1544–1550. [Google Scholar] [CrossRef]
- Ponnudurai, T.; Lensen, A.H.; van Gemert, G.J.; Bensink, M.P.; Bolmer, M.; Meuwissen, J.H. Sporozoite load of mosquitoes infected with Plasmodium falciparum. Trans. R. Soc. Trop. Med. Hyg. 1989, 83, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Tavares, J.; Formaglio, P.; Medvinsky, A.; Menard, R.; Amino, R. Imaging sporozoite cell traversal in the liver of mice. Methods Mol. Biol. 2013, 923, 401–410. [Google Scholar] [CrossRef]
- Amino, R.; Thiberge, S.; Shorte, S.; Frischknecht, F.; Menard, R. Quantitative imaging of Plasmodium sporozoites in the mammalian host. C. R. Biol. 2006, 329, 858–862. [Google Scholar] [CrossRef]
- Vanderberg, J.P. Development of infectivity by the Plasmodium berghei sporozoite. J. Parasitol. 1975, 61, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Sidjanski, S.; Vanderberg, J.P. Delayed migration of Plasmodium sporozoites from the mosquito bite site to the blood. Am. J. Trop. Med. Hyg. 1997, 57, 426–429. [Google Scholar] [CrossRef]
- Roth, A.; Adapa, S.R.; Zhang, M.; Liao, X.; Saxena, V.; Goffe, R.; Li, S.; Ubalee, R.; Saggu, G.S.; Pala, Z.R.; et al. Unraveling the Plasmodium vivax sporozoite transcriptional journey from mosquito vector to human host. Sci. Rep. 2018, 8, 12183. [Google Scholar] [CrossRef] [PubMed]
- Mota, M.M.; Pradel, G.; Vanderberg, J.P.; Hafalla, J.C.; Frevert, U.; Nussenzweig, R.S.; Nussenzweig, V.; Rodriguez, A. Migration of Plasmodium sporozoites through cells before infection. Science 2001, 291, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Loubens, M.; Marinach, C.; Paquereau, C.; Hamada, S.; Hoareau-Coudert, B.; Akbar, D.; Franetich, J.; Silvie, O. The claudin-like apicomplexan microneme protein is required for gliding motility and infectivity of Plasmodium sporozoites. PLoS Pathog. 2023, 19, e1011261. [Google Scholar] [CrossRef] [PubMed]
- Pradel, G.; Frevert, U. Malaria sporozoites actively enter and pass through rat Kupffer cells prior to hepatocyte invasion. Hepatology 2001, 33, 1154–1165. [Google Scholar] [CrossRef]
- Frischknecht, F.; Matuschewski, K. Plasmodium Sporozoite Biology. Cold Spring Harb Perspect. Med. 2017, 7, a025478. [Google Scholar] [CrossRef]
- Frevert, U.; Usynin, I.; Baer, K.; Klotz, C. Nomadic or sessile: Can Kupffer cells function as portals for malaria sporozoites to the liver? Cell. Microbiol. 2006, 8, 1537–1546. [Google Scholar] [CrossRef]
- Wake, K. Cell-cell organization and functions of 'sinusoids' in liver microcirculation system. J. Electron. Microsc. (Tokyo) 1999, 48, 89–98. [Google Scholar] [CrossRef]
- Krettli, A.U.; Dantas, L.A. Which routes do Plasmodium sporozoites use for successful infections of vertebrates? Infect. Immun. 2000, 68, 3064–3065. [Google Scholar] [CrossRef]
- Baer, K.; Roosevelt, M.; Clarkson, A.B.J.; van Rooijen, N.; Schnieder, T.; Frevert, U. Kupffer cells are obligatory for Plasmodium yoelii sporozoite infection of the liver. Cell. Microbiol. 2007, 9, 397–412. [Google Scholar] [CrossRef]
- Baer, K.; Klotz, C.; Kappe, S.H.I.; Schnieder, T.; Frevert, U. Release of hepatic Plasmodium yoelii merozoites into the pulmonary microvasculature. PLoS Pathog. 2007, 3, e171. [Google Scholar] [CrossRef]
- Malleret, B.; Renia, L.; Russell, B. The unhealthy attraction of Plasmodium vivax to reticulocytes expressing transferrin receptor 1 (CD71). Int. J. Parasitol. 2017, 47, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Kho, S.; Qotrunnada, L.; Leonardo, L.; Andries, B.; Wardani, P.A.I.; Fricot, A.; Henry, B.; Hardy, D.; Margyaningsih, N.I.; Apriyanti, D.; et al. Hidden Biomass of Intact Malaria Parasites in the Human Spleen. N. Engl. J. Med. 2021, 384, 2067–2069. [Google Scholar] [CrossRef] [PubMed]
- Lepay, D.A.; Nathan, C.F.; Steinman, R.M.; Murray, H.W.; Cohn, Z.A. Murine Kupffer cells. Mononuclear phagocytes deficient in the generation of reactive oxygen intermediates. J. Exp. Med. 1985, 161, 1079–1096. [Google Scholar] [CrossRef]
- Nusrat, A.R.; Wright, S.D.; Aderem, A.A.; Steinman, R.M.; Cohn, Z.A. Properties of isolated red pulp macrophages from mouse spleen. J. Exp. Med. 1988, 168, 1505–1510. [Google Scholar] [CrossRef] [PubMed]
- Venugopal, K.; Hentzschel, F.; Valkiunas, G.; Marti, M. Plasmodium asexual growth and sexual development in the haematopoietic niche of the host. Nat. Rev. Microbiol. 2020, 18, 177–189. [Google Scholar] [CrossRef]
- Sturm, A.; Amino, R.; van de Sand, C.; Regen, T.; Retzlaff, S.; Rennenberg, A.; Krueger, A.; Pollok, J.; Menard, R.; Heussler, V.T. Manipulation of host hepatocytes by the malaria parasite for delivery into liver sinusoids. Science 2006, 313, 1287–1290. [Google Scholar] [CrossRef]
- Yahata, K.; Hart, M.N.; Davies, H.; Asada, M.; Wassmer, S.C.; Templeton, T.J.; Treeck, M.; Moon, R.W.; Kaneko, O. Gliding motility of Plasmodium merozoites. Proc. Natl. Acad. Sci. USA 2021, 118, e2114442118. [Google Scholar] [CrossRef] [PubMed]
- White, N.J. Determinants of relapse periodicity in Plasmodium vivax malaria. Malar J. 2011, 10, 297. [Google Scholar] [CrossRef]
- Mancio-Silva, L.; Gural, N.; Real, E.; Wadsworth, M.H.; Butty, V.L.; March, S.; Nerurkar, N.; Hughes, T.K.; Roobsoong, W.; Fleming, H.E.; et al. A single-cell liver atlas of Plasmodium vivax infection. Cell. Host Microbe 2022, 30, 1048–1060.e5. [Google Scholar] [CrossRef] [PubMed]
- Hentzschel, F.; Gibbins, M.P.; Attipa, C.; Beraldi, D.; Moxon, C.A.; Otto, T.D.; Marti, M. Host cell maturation modulates parasite invasion and sexual differentiation in Plasmodium berghei. Sci. Adv. 2022, 8, eabm7348. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Zhang, C.; Joy, D.A. Host-Malaria Parasite Interactions and Impacts on Mutual Evolution. Front. Cell. Infect. Microbiol. 2020, 10, 587933. [Google Scholar] [CrossRef] [PubMed]
- Thomson, J.G. Malaria in Nyasaland: (Section of Tropical Diseases and Parasitology). Proc. R. Soc. Med. 1935, 28, 391–404. [Google Scholar] [PubMed]
- Hawking, F.; Wilson, M.E.; Gammage, K. Evidence for cyclic development and short-lived maturity in the gametocytes of Plasmodium falciparum. Trans. R. Soc. Trop. Med. Hyg. 1971, 65, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Bantuchai, S.; Imad, H.; Nguitragool, W. Plasmodium vivax gametocytes and transmission. Parasitol. Int. 2022, 87, 102497. [Google Scholar] [CrossRef]
- Enger, A.; Strand, O.A.; Ranheim, T.; Hellum, K.B. Exflagellation of microgametocytes in Plasmodium vivax malaria: A diagnostic conundrum. Med. Princ Pract. 2004, 13, 298–300. [Google Scholar] [CrossRef]
- Baton, L.A.; Ranford-Cartwright, L.C. Do malaria ookinete surface proteins P25 and P28 mediate parasite entry into mosquito midgut epithelial cells? Malar J. 2005, 4, 15. [Google Scholar] [CrossRef]
- Prudencio, M.; Rodriguez, A.; Mota, M.M. The silent path to thousands of merozoites: The Plasmodium liver stage. Nat. Rev. Microbiol. 2006, 4, 849–856. [Google Scholar] [CrossRef]
- Real, E.; Howick, V.M.; Dahalan, F.A.; Witmer, K.; Cudini, J.; Andradi-Brown, C.; Blight, J.; Davidson, M.S.; Dogga, S.K.; Reid, A.J.; et al. A single-cell atlas of Plasmodium falciparum transmission through the mosquito. Nat. Commun. 2021, 12, 3196. [Google Scholar] [CrossRef]
- Bray, R.S.; Garnham, P.C. The life-cycle of primate malaria parasites. Br. Med. Bull. 1982, 38, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Smalley, M.E.; Abdalla, S.; Brown, J. The distribution of Plasmodium falciparum in the peripheral blood and bone marrow of Gambian children. Trans. R. Soc. Trop. Med. Hyg. 1981, 75, 103–105. [Google Scholar] [CrossRef] [PubMed]
- Joice, R.; Nilsson, S.K.; Montgomery, J.; Dankwa, S.; Egan, E.; Morahan, B.; Seydel, K.B.; Bertuccini, L.; Alano, P.; Williamson, K.C.; et al. Plasmodium falciparum transmission stages accumulate in the human bone marrow. Sci. Transl. Med. 2014, 6, 244re5. [Google Scholar] [CrossRef] [PubMed]
- De Niz, M.; Meibalan, E.; Mejia, P.; Ma, S.; Brancucci, N.M.B.; Agop-Nersesian, C.; Mandt, R.; Ngotho, P.; Hughes, K.R.; Waters, A.P.; et al. Plasmodium gametocytes display homing and vascular transmigration in the host bone marrow. Sci. Adv. 2018, 4, eaat3775. [Google Scholar] [CrossRef]
- Lee, R.S.; Waters, A.P.; Brewer, J.M. A cryptic cycle in haematopoietic niches promotes initiation of malaria transmission and evasion of chemotherapy. Nat. Commun. 2018, 9, 1689. [Google Scholar] [CrossRef]
- Wake, K.; Kawai, Y.; Smedsrod, B. Re-evaluation of the reticulo-endothelial system. Ital. J. Anat. Embryol. 2001, 106, 261–269. [Google Scholar]
- van Furth, R.; Cohn, Z.A.; Hirsch, J.G.; Humphrey, J.H.; Spector, W.G.; Langevoort, H.L. Mononuclear phagocytic system: New classification of macrophages, monocytes and of their cell line. Bull. World Health Organ. 1972, 47, 651–658. [Google Scholar]
- Yona, S.; Gordon, S. From the Reticuloendothelial to Mononuclear Phagocyte System - The Unaccounted Years. Front. Immunol. 2015, 6, 328. [Google Scholar] [CrossRef]
- Ginhoux, F.; Jung, S. Monocytes and macrophages: Developmental pathways and tissue homeostasis. Nat. Rev. Immunol. 2014, 14, 392–404. [Google Scholar] [CrossRef]
- Guilliams, M.; Ginhoux, F.; Jakubzick, C.; Naik, S.H.; Onai, N.; Schraml, B.U.; Segura, E.; Tussiwand, R.; Yona, S. Dendritic cells, monocytes and macrophages: A unified nomenclature based on ontogeny. Nat. Rev. Immunol. 2014, 14, 571–578. [Google Scholar] [CrossRef]
- Guilliams, M.; De Kleer, I.; Henri, S.; Post, S.; Vanhoutte, L.; De Prijck, S.; Deswarte, K.; Malissen, B.; Hammad, H.; Lambrecht, B.N. Alveolar macrophages develop from fetal monocytes that differentiate into long-lived cells in the first week of life via GM-CSF. J. Exp. Med. 2013, 210, 1977–1992. [Google Scholar] [CrossRef]
- Okabe, Y.; Medzhitov, R. Tissue-specific signals control reversible program of localization and functional polarization of macrophages. Cell 2014, 157, 832–844. [Google Scholar] [CrossRef]
- Pradel, G.; Garapaty, S.; Frevert, U. Proteoglycans mediate malaria sporozoite targeting to the liver. Mol. Microbiol. 2002, 45, 637–651. [Google Scholar] [CrossRef]
- Hussain, T.; Linera-Gonzalez, J.; Beck, J.M.; Fierro, M.A.; Mair, G.R.; Smith, R.C.; Beck, J.R. The PTEX Pore Component EXP2 Is Important for Intrahepatic Development during the Plasmodium Liver Stage. mBio 2022, 13, e0309622-22. [Google Scholar] [CrossRef] [PubMed]
- Comazzetto, S.; Shen, B.; Morrison, S.J. Niches that regulate stem cells and hematopoiesis in adult bone marrow. Dev. Cell. 2021, 56, 1848–1860. [Google Scholar] [CrossRef] [PubMed]
- Groom, A.C.; Schmidt, E.E.; MacDonald, I.C. Microcirculatory pathways and blood flow in spleen: New insights from washout kinetics, corrosion casts, and quantitative intravital videomicroscopy. Scanning Microsc. 1991, 5, 159–173, discussion 173-4. [Google Scholar] [PubMed]
- Steiniger, B.; Barth, P.; Hellinger, A. The perifollicular and marginal zones of the human splenic white pulp : Do fibroblasts guide lymphocyte immigration? Am. J. Pathol. 2001, 159, 501–512. [Google Scholar] [CrossRef]
- Buffet, P.A.; Safeukui, I.; Deplaine, G.; Brousse, V.; Prendki, V.; Thellier, M.; Turner, G.D.; Mercereau-Puijalon, O. The pathogenesis of Plasmodium falciparum malaria in humans: Insights from splenic physiology. Blood 2011, 117, 381–392. [Google Scholar] [CrossRef]
- Johnston, B.; Butcher, E.C. Chemokines in rapid leukocyte adhesion triggering and migration. Semin. Immunol. 2002, 14, 83–92. [Google Scholar] [CrossRef]
- Nakhaei-Nejad, M.; Hussain, A.M.; Zhang, Q.; Murray, A.G. Endothelial PI 3-kinase activity regulates lymphocyte diapedesis. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H3608–H3616. [Google Scholar] [CrossRef]
- Hliscs, M.; Millet, C.; Dixon, M.W.; Siden-Kiamos, I.; McMillan, P.; Tilley, L. Organization and function of an actin cytoskeleton in Plasmodium falciparum gametocytes. Cell. Microbiol. 2015, 17, 207–225. [Google Scholar] [CrossRef] [PubMed]
- Itkin, T.; Gur-Cohen, S.; Spencer, J.A.; Schajnovitz, A.; Ramasamy, S.K.; Kusumbe, A.P.; Ledergor, G.; Jung, Y.; Milo, I.; Poulos, M.G.; et al. Distinct bone marrow blood vessels differentially regulate haematopoiesis. Nature 2016, 532, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Vestweber, D. How leukocytes cross the vascular endothelium. Nat. Rev. Immunol. 2015, 15, 692–704. [Google Scholar] [CrossRef]
- Angrisano, F.; Robinson, L.J. Plasmodium vivax - How hidden reservoirs hinder global malaria elimination. Parasitol. Int. 2022, 87, 102526. [Google Scholar] [CrossRef] [PubMed]
- Commons, R.J.; Simpson, J.A.; Watson, J.; White, N.J.; Price, R.N. Estimating the Proportion of Plasmodium vivax Recurrences Caused by Relapse: A Systematic Review and Meta-Analysis. Am. J. Trop. Med. Hyg. 2020. [Google Scholar] [CrossRef]
- Del Portillo, H.A.; Ferrer, M.; Brugat, T.; Martin-Jaular, L.; Langhorne, J.; Lacerda, M.V.G. The role of the spleen in malaria. Cell. Microbiol. 2012, 14, 343–355. [Google Scholar] [CrossRef]
- Lapp, S.A.; Korir-Morrison, C.; Jiang, J.; Bai, Y.; Corredor, V.; Galinski, M.R. Spleen-dependent regulation of antigenic variation in malaria parasites: Plasmodium knowlesi SICAvar expression profiles in splenic and asplenic hosts. PLoS ONE 2013, 8, e78014. [Google Scholar] [CrossRef]
- Betterman, K.L.; Harvey, N.L. The lymphatic vasculature: Development and role in shaping immunity. Immunol. Rev. 2016, 271, 276–292. [Google Scholar] [CrossRef]
- Landau, I.; Chabaud, A.G.; Vuong, P.N.; Deharo, E.; Gautret, P. Circulation in the lymphatic system and latency of Plasmodium merozoites. Preliminary note. Parasite 1995, 2, 185–186. [Google Scholar] [CrossRef]
- Iwakiri, Y. The lymphatic system: A new frontier in hepatology. Hepatology 2016, 64, 706–707. [Google Scholar] [CrossRef]
- Gray, E.E.; Cyster, J.G. Lymph node macrophages. J. Innate Immun. 2012, 4, 424–436. [Google Scholar] [CrossRef] [PubMed]
- Giusiano, G. The Trojan Horse Model in Paracoccidioides: A Fantastic Pathway to Survive Infecting Human Cells. Front. Cell. Infect. Microbiol. 2021, 10, 605679. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.M.; Oghumu, S.; Gupta, G.; McGwire, B.S.; Drew, M.E.; Satoskar, A.R. Mechanisms of cellular invasion by intracellular parasites. Cell Mol. Life Sci. 2014, 71, 1245–1263. [Google Scholar] [CrossRef] [PubMed]
- Kronstad, J.W.; Hu, G.; Choi, J. The cAMP/Protein Kinase A Pathway and Virulence in Cryptococcus neoformans. Mycobiology 2011, 39, 143–150. [Google Scholar] [CrossRef]
- May, R.C.; Stone, N.R.H.; Wiesner, D.L.; Bicanic, T.; Nielsen, K. Cryptococcus: From environmental saprophyte to global pathogen. Nat. Rev. Microbiol. 2016, 14, 106–117. [Google Scholar] [CrossRef]
- Arredondo, S.A.; Schepis, A.; Reynolds, L.; Kappe, S.H.I. Secretory Organelle Function in the Plasmodium Sporozoite. Trends Parasitol. 2021, 37, 651–663. [Google Scholar] [CrossRef]
- Cowman, A.F.; Tonkin, C.J.; Tham, W.; Duraisingh, M.T. The Molecular Basis of Erythrocyte Invasion by Malaria Parasites. Cell. Host Microbe 2017, 22, 232–245. [Google Scholar] [CrossRef] [PubMed]
- Motshoge, T.; Ababio, G.K.; Aleksenko, L.; Read, J.; Peloewetse, E.; Loeto, M.; Mosweunyane, T.; Moakofhi, K.; Ntebele, D.S.; Chihanga, S.; et al. Molecular evidence of high rates of asymptomatic P. vivax infection and very low P. falciparum malaria in Botswana. BMC Infect. Dis. 2016, 16, 1–8. [Google Scholar] [CrossRef]
- Motshoge, T.; Haiyambo, D.H.; Ayanful-Torgby, R.; Aleksenko, L.; Ntebela, D.; Malleret, B.; Renia, L.; Peloewetse, E.; Paganotti, G.M.; Quaye, I.K. Recent Molecular Assessment of Plasmodium vivax and Plasmodium falciparum Asymptomatic Infections in Botswana. Am. J. Trop. Med. Hyg. 2021. [Google Scholar] [CrossRef]
- Moonasar, D.; Maharaj, R.; Kunene, S.; Candrinho, B.; Saute, F.; Ntshalintshali, N.; Morris, N. Towards malaria elimination in the MOSASWA (Mozambique, South Africa and Swaziland) region. Malar J. 2016, 15, 419. [Google Scholar] [CrossRef]
- Chihanga, S.; Haque, U.; Chanda, E.; Mosweunyane, T.; Moakofhi, K.; Jibril, H.B.; Motlaleng, M.; Zhang, W.; Glass, G.E. Malaria elimination in Botswana, 2012-2014: Achievements and challenges. Parasit. Vectors 2016, 9, 99. [Google Scholar] [CrossRef] [PubMed]
- Thomson, M.C.; Mason, S.J.; Phindela, T.; Connor, S.J. Use of rainfall and sea surface temperature monitoring for malaria early warning in Botswana. Am. J. Trop. Med. Hyg. 2005, 73, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Simon, C.; Moakofhi, K.; Mosweunyane, T.; Jibril, H.B.; Nkomo, B.; Motlaleng, M.; Ntebela, D.S.; Chanda, E.; Haque, U. Malaria control in Botswana, 2008-2012: The path towards elimination. Malar J. 2013, 12, 458. [Google Scholar] [CrossRef]
- Craig, M.H.; Sharp, B.L.; Mabaso, M.L.; Kleinschmidt, I. Developing a spatial-statistical model and map of historical malaria prevalence in Botswana using a staged variable selection procedure. Int. J. Health. Geogr. 2007, 6, 44. [Google Scholar] [CrossRef] [PubMed]
- Roucher, C.; Rogier, C.; Sokhna, C.; Tall, A.; Trape, J. A 20-year longitudinal study of Plasmodium ovale and Plasmodium malariae prevalence and morbidity in a West African population. PLoS ONE 2014, 9, e87169. [Google Scholar] [CrossRef]
- Thomson-Luque, R.; Bautista, J.M. Home Sweet Home: Plasmodium vivax-Infected Reticulocytes-The Younger the Better? Front. Cell. Infect. Microbiol. 2021, 11, 675156. [Google Scholar] [CrossRef]
- Bango, Z.A.; Tawe, L.; Muthoga, C.W.; Paganotti, G.M. Past and current biological factors affecting malaria in the low transmission setting of Botswana: A review. Infect. Genet. Evol. 2020, 85, 104458. [Google Scholar] [CrossRef]
- Moakofhi, K.; Edwards, J.K.; Motlaleng, M.; Namboze, J.; Butt, W.; Obopile, M.; Mosweunyane, T.; Manzi, M.; Takarinda, K.C.; Owiti, P. Advances in malaria elimination in Botswana: A dramatic shift to parasitological diagnosis, 2008-2014. Public. Health. Action 2018, 8, S34–S38. [Google Scholar] [CrossRef]
- Saenz, F.E.; Arevalo-Cortes, A.; Valenzuela, G.; Vallejo, A.F.; Castellanos, A.; Poveda-Loayza, A.C.; Gutierrez, J.B.; Alvarez, A.; Yan, Y.H.; Benavides, Y.; et al. Malaria epidemiology in low-endemicity areas of the northern coast of Ecuador: High prevalence of asymptomatic infections. Malar J. 2017, 16, 300. [Google Scholar] [CrossRef]
- Lover, A.A.; Dantzer, E.; Hongvanthong, B.; Chindavongsa, K.; Welty, S.; Reza, T.; Khim, N.; Menard, D.; Bennett, A. Prevalence and risk factors for asymptomatic malaria and genotyping of glucose 6-phosphate (G6PD) deficiencies in a vivax-predominant setting, Lao PDR: Implications for sub-national elimination goals. Malar J. 2018, 17, 218. [Google Scholar] [CrossRef]
- Niang, M.; Diop, F.; Niang, O.; Sadio, B.D.; Sow, A.; Faye, O.; Diallo, M.; Sall, A.A.; Perraut, R.; Toure-Balde, A. Unexpected high circulation of Plasmodium vivax in asymptomatic children from Kedougou, southeastern Senegal. Malar J. 2017, 16, 497. [Google Scholar] [CrossRef] [PubMed]
- Obadia, T.; Nekkab, N.; Robinson, L.J.; Drakeley, C.; Mueller, I.; White, M.T. Developing sero-diagnostic tests to facilitate Plasmodium vivax Serological Test-and-Treat approaches: Modeling the balance between public health impact and overtreatment. BMC Med. 2022, 20, 98. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.; Russell, D.G. The interaction of Leishmania species with macrophages. Adv. Parasitol. 1992, 31, 175–254. [Google Scholar] [CrossRef]
- Murray, H.W.; Berman, J.D.; Davies, C.R.; Saravia, N.G. Advances in leishmaniasis. Lancet 2005, 366, 1561–1577. [Google Scholar] [CrossRef] [PubMed]
- Lafuse, W.P.; Story, R.; Mahylis, J.; Gupta, G.; Varikuti, S.; Steinkamp, H.; Oghumu, S.; Satoskar, A.R. Leishmania donovani infection induces anemia in hamsters by differentially altering erythropoiesis in bone marrow and spleen. PLoS ONE 2013, 8, e59509. [Google Scholar] [CrossRef] [PubMed]
- Miralles, G.D.; Stoeckle, M.Y.; McDermott, D.F.; Finkelman, F.D.; Murray, H.W. Th1 and Th2 cell-associated cytokines in experimental visceral leishmaniasis. Infect. Immun. 1994, 62, 1058–1063. [Google Scholar] [CrossRef]
- Engwerda, C.R.; Kaye, P.M. Organ-specific immune responses associated with infectious disease. Immunol. Today 2000, 21, 73–78. [Google Scholar] [CrossRef]
- Requena, J.M.; Soto, M.; Doria, M.D.; Alonso, C. Immune and clinical parameters associated with Leishmania infantum infection in the golden hamster model. Vet. Immunol. Immunopathol. 2000, 76, 269–281. [Google Scholar] [CrossRef]
- Liu, Y.; Pop, R.; Sadegh, C.; Brugnara, C.; Haase, V.H.; Socolovsky, M. Suppression of Fas-FasL coexpression by erythropoietin mediates erythroblast expansion during the erythropoietic stress response in vivo. Blood 2006, 108, 123–133. [Google Scholar] [CrossRef]
- Arevalo-Herrera, M.; Lopez-Perez, M.; Medina, L.; Moreno, A.; Gutierrez, J.B.; Herrera, S. Clinical profile of Plasmodium falciparum and Plasmodium vivax infections in low and unstable malaria transmission settings of Colombia. Malar J. 2015, 14, 154. [Google Scholar] [CrossRef]
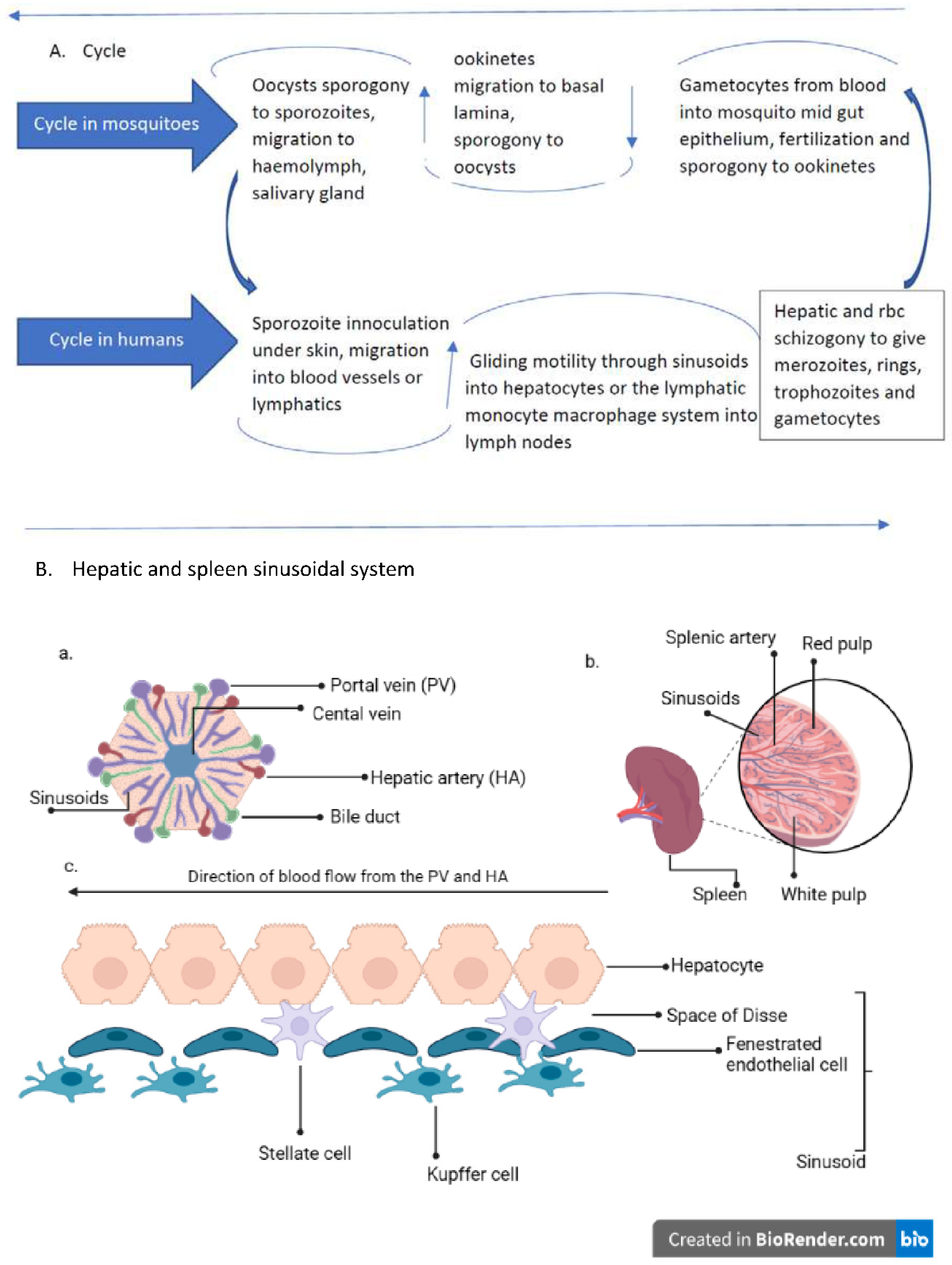
Figure 1.
A. Plasmodium life cycle; A. description of plasmodium life cycle from the sexual stage oocysts through sporogony to sporozoites, and migration into the salivary gland till inoculation into human intermediary host during feeding, through the asexual stage in host tissues and back to the sexual stages B. a liver lobule showing the triads and the hepatic sinus lining monocyte macrophage system (sinusoid). Parasites seeking entry, interact with the Kupffer cells and stellate cells through proteoglycans. Cells arrive at the sinusoids from blood flow coming through the intersection of Portal vein and Hepatic Artery in the liver. B. The spleen and a transverse section showing the areas of sinusoidal system: In the spleen, blood flows through the splenic artery into the central artery, either through the perifollicular zone and then through the venous sinusoids in the fast circulation, or the red pulp of the cords (consisting of fibroblasts and reticular fibers, but without endothelial cells) in a slow microcirculation. The slow circulation enables the mononuclear phagocyte system to remove particulate matter and generate the requisite immune response. From the red pulp, the blood squeezes through the venous sinusoids which filters the blood further removing infected rbcs or intraerythrocytic bodies and exit through the splenic veins to the portal vein. c. a hepatic sinusoidal system.
Figure 1.
A. Plasmodium life cycle; A. description of plasmodium life cycle from the sexual stage oocysts through sporogony to sporozoites, and migration into the salivary gland till inoculation into human intermediary host during feeding, through the asexual stage in host tissues and back to the sexual stages B. a liver lobule showing the triads and the hepatic sinus lining monocyte macrophage system (sinusoid). Parasites seeking entry, interact with the Kupffer cells and stellate cells through proteoglycans. Cells arrive at the sinusoids from blood flow coming through the intersection of Portal vein and Hepatic Artery in the liver. B. The spleen and a transverse section showing the areas of sinusoidal system: In the spleen, blood flows through the splenic artery into the central artery, either through the perifollicular zone and then through the venous sinusoids in the fast circulation, or the red pulp of the cords (consisting of fibroblasts and reticular fibers, but without endothelial cells) in a slow microcirculation. The slow circulation enables the mononuclear phagocyte system to remove particulate matter and generate the requisite immune response. From the red pulp, the blood squeezes through the venous sinusoids which filters the blood further removing infected rbcs or intraerythrocytic bodies and exit through the splenic veins to the portal vein. c. a hepatic sinusoidal system.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



