
Submitted:
26 June 2023
Posted:
27 June 2023
You are already at the latest version
Abstract
Bacterial Extracellular Vesicles (BEVs) generated from the bacteria has high feasibility of intracellular interactions with other cells, can be used as a cargo to deliver any therapeutic substances like monoclonal antibodies, proteins, plasmids, siRNA and small molecules for the treatment of neurodegenerative diseases (NDs). BEVs have a high ability for delivering therapeutic molecules across the blood-brain barrier to treat Alzheimer's Disease (AD) via various mechanisms. In this review, we have sum up in-depth research details of the role and advancement of BEVs in NDs, AD and its Therapeutics. Moreover, details about offensive and defensive functions of BEVs in ND pathogenesis and examine the critical BEV networks in the microbiome-gut-brain axis and their role in neurodegenerative disorders. Additionally determining the roles of BEVs in the neuroimmune system and their interaction with neurodegenerative disorders and exploring the risk factors of BEVs in the autophagy-lysosomal pathway and their possible effects on neurodegenerative disorders are summarised in detail. In conclusion, this review seeks to contribute to a better understanding of the potential function of BEVs in NDs and to identify new therapeutic intervention strategies.
Keywords:
Bacterial Extracellular Vesicles
; Therapeutics
; Neurodegenerative Disease
; Alzheimer's Disease
; Nanocarriers
1. Introduction
Neurodegenerative diseases (NDs) are a group of progressive disorders that affect the central nervous system and result in the gradual degeneration and mortality of neurons [1]. Alzheimer's disease, Parkinson's disease, Huntington's disease, and amyotrophic lateral sclerosis (ALS) are among these conditions of NDs [1]. Alzheimer's Disease International estimates that 50 million people worldwide are afflicted by dementia alone [2]. Consequently, neurodegenerative disorders (NDs) pose a significant threat to public health and a significant burden on society [2,3]. Despite significant efforts to develop treatments, no known cures for NDs are presently available [4].
Researchers have begun to investigate the function of bacterial extracellular vesicles (BEVs) in the pathogenesis of NDs in recent years [5,6]. BEVs are small, membrane-bound particles released by bacteria (Figure 1) that contain a variety of bioactive molecules, such as proteins, lipids, and nucleic acids [5,6]. It has been demonstrated that these vesicles serve an essential role in bacterial communication, adaptation, and virulence [7]. In addition, recent studies indicate that BEVs may also add to the development and progression of NDs, potentially presenting novel therapeutic targets [6,7].
The potential function of BEVs in NDs has been the subject of intensive research over the past few years [8]. According to studies, BEVs can play both offensive and defensive functions in the context of NDs, including regulation of immune responses, modulation of the microbiome-gut-brain axis, and induction of autophagy-lysosomal pathway dysfunction [6,8,9]. In addition, recent advancements in BEV isolation, characterization, and engineering have paved the way for the development of novel therapeutic strategies [5,9].
This review's objective is to provide an overview of the current literature on the role of BEVs in NDs and to investigate recent developments in the field of BEV therapeutics. Specifically, this analysis seeks to:
Focus on the offensive and defensive functions of BEVs in ND pathogenesis.
Examine the critical BEV networks in the microbiome-gut-brain axis and their role in neurodegenerative disorders.
Determine the roles of BEVs in the neuroimmune system and their interaction with neurodegenerative disorders.
Explore the risk factors of BEVs in the autophagy-lysosomal pathway and their possible effects on neurodegenerative disorders.
Emphasize the potential uses of BEVs in developing new drug for neurodegenerative diseases.
Suggest future prospect of the study of the role of BEVs in neurodegenerative diseases.
This review seeks to contribute to a better understanding of the potential function of BEVs in NDs and to identify new therapeutic intervention strategies.
1. Offensive and Defensive roles Bacterial Extracellular Vesicles in Neurodegenerative Disease
Latest research has shown that bacterial extracellular vesicles [10], also known as BEVs, can cause neuroinflammation and affect neuronal function [11], which points to the potential neurotoxicity of these particles in the context of neurodegenerative disorders [11,12]. (Kim et al., 202; Kumar et al., 2020). BEVs tend to yield the inflammatory cytokines and chemokines after activating microglia and astrocytes with virulence factors such lipopolysaccharides, peptidoglycans, and proteins [13,14,15]. Alzheimer's disease (AD), Parkinson's disease (PD), and multiple sclerosis (MS), among others, have all been associated to the activation of neuroinflammatory pathways [12,15].
BEVs generated from Pseudomonas aeruginosa were discovered in a recent study (Figure 2) to induce inflammation and mortality of dopaminergic neurons in the substantia nigra [11,12,16]. Inflammation and mortality of dopaminergic neurons in the substantia nigra is a defining hallmark of Parkinson's disease (PD) [11,15]. It was discovered that BEVs generated from Escherichia coli caused neuronal death and impaired memory in an Alzheimer's disease mice model [15,16]. By inducing neuroinflammation and impairing neuronal function, BEVs may contribute to the pathogenesis of NDs, according to these findings [16,17].
In addition to their potential neurotoxicity [17], it has been demonstrated that BEVs serve a protective role in NDs [17]. Several studies have demonstrated, for instance, that BEVs can exert neuroprotective and immunomodulatory effects [17,18]. Specifically, it has been demonstrated that BEVs from commensal gut bacteria enhance cognitive function and reduce neuroinflammation in mouse models of NDs [18,19]. These results suggest that BEVs may also have therapeutic applications for the treatment of NDs [18,19]. The neurotoxicity of BEVs necessitates caution in their use as therapeutics, but their neuroprotective and immunomodulatory properties present opportunities for the development of novel treatments for NDs [10,18]. To thoroughly comprehend the mechanisms underlying the offensive and defensive roles of BEVs in NDs, additional research is required [10,17].
Recent research suggests that BEVs can also play a defensive function in NDs due to their neuroprotective and immunomodulatory properties [17,18]. Multiple studies have demonstrated, for instance, that BEVs derived from specific bacterial strains can improve neuronal survival and function in neurodegenerative conditions [19]. In the mouse model of Alzheimer’s disease, Haney et al. [20] found that BEVs from the probiotic Lactobacillus rhamnosus GG could reduce amyloid-beta (A) deposition and enhance cognitive function [21]. Wang et al. [21] demonstrated that BEVs from Bifidobacterium infantis could reduce inflammation and oxidative stress in a mouse model of Parkinson's disease [22], resulting in enhanced motor function [18,19,22].
BEVs have been shown to possess immunomodulatory properties in the context of NDs, in addition to their neuroprotective effects [15,22]. By modulating the gut-brain axis [23], found that BEVs from Akkermansia muciniphila could reduce neuroinflammation and enhance cognitive function in a mouse model of Parkinson's disease [15,24]. Similarly, demonstrated that BEVs from Lactobacillus plantarum PS128 modulated microglial activity to enhance cognitive function and reduce neuroinflammation in an Alzheimer's disease mouse model [22,25]. These studies suggest that BEVs may serve a dual role in NDs by possessing both offensive and defensive characteristics [15,25]. Some BEVs can induce neuroinflammation and impede neuronal function, whereas others can prevent neurodegenerative processes and modulate the immune system [18,21]. To thoroughly comprehend the mechanisms underlying the defensive properties of BEVs and to investigate their potential as therapeutic agents for NDs, additional research is required.
1. Critical Networks of Bacterial Extracellular Vesicles in the microbiome-gut-brain axis
The microbiome-gut-brain axis (MGBA) is a complex network of bidirectional communication between the gastrointestinal tract, the central nervous system (CNS), and the gut microbiota [19,26]. Recent evidence suggests that this axis regulates a variety of physiological and pathological processes, such as neuroinflammation and neurodegeneration [26,27]. The gut microbiota has a vast array of microorganisms inhabiting the human gastrointestinal tract, and it has been shown to influence brain function and behaviour via multiple mechanisms [27]. These include the production of neurotransmitters and short-chain fatty acids, modulation of the immune system, and regulation of the hypothalamic-pituitary-adrenal axis [28,29]. Multiple NDs, such as Parkinson's [28], Alzheimer's [29,30], and multiple cerebral sclerosis (Table 1), have been linked to abnormalities in the gastrointestinal microbiota in their pathogenesis [30,31,32]. Additionally, it has been demonstrated that bacterial extracellular vesicles (BEVs) produced by intestinal microbiota can cross the blood-brain barrier and directly affect the CNS function [22,31]. BEVs from the gut commensal Bacteroides fragilis have been shown to facilitate the differentiation and maturation of oligodendrocytes, which are essential for the production of myelin in the CNS [6,33]. In a mouse model of Alzheimer's disease, BEVs from Akkermansia muciniphila have been shown to protect against neuroinflammation and cognitive decline [23,24,33].
The above discussed results point out that the microbiota-gut-brain axis (MGBA) plays an important role in the pathogenesis of NDs, and that BEVs produced by intestinal microbiota may represent a novel drug delivery system for such conditions [34]. It has also been demonstrated that the gut microbiome can influence brain function and behaviour via multiple mechanisms, including the production of neurotransmitters, regulation of the immune system, and modulation of the gut-brain axis signalling pathways [35]. Bacterial extracellular vesicles (BEVs), that are produced by numerous bacteria in the microbiome of the gut, have been identified as potential mediators of this communication between the gut and the brain [6,35]. The effects of BEVs on the microbiome-gut-brain axis and their potential function in NDs have been studied and discussed in animal models [18]. In cell cultures and mouse models, BEVs from the gastrointestinal microbiome of Parkinson's disease (PD) patients were able to induce alpha-synuclein aggregation, which is a hallmark of PD pathology [15,22]. In a mouse model of Alzheimer's disease (AD), It was observed that BEVs from a specific gut bacterium, Akkermansia muciniphila, reduced neuroinflammation and enhance cognitive function [36].

Table 1.
A summary of research findings in relation to the neuro-immune system affecting different types of NDs and their key conclusions of the study.
Table 1.
A summary of research findings in relation to the neuro-immune system affecting different types of NDs and their key conclusions of the study.
| NDs | Key Finding | Source |
|---|---|---|
| Alzheimer's Disease | Microglial activation contributes to amyloid beta deposition and neuronal damage | [37] |
| Parkinson's disease | T cells infiltrate the substantia nigra and promote neuroinflammation | [38,39] |
| Multiple sclerosis | Dysbiosis and gut-derived molecules contribute to neuroinflammation and disease progression | [40,41,42] |
Another study identified a group of BEVs produced by gut bacteria that could cross the blood-brain barrier as a result, penetrating the brain, modulating the immune system [43], and potentially playing a significant role in NDs [17,27]. These studies indicate that BEVs can play a significant role in the communication between the gastrointestinal microbiome and the brain, and that their dysregulation may contribute to the development and progression of NDs [36,43]. The mechanisms underlying the effects of BEVs on the microbiome-gut-brain axis and their potential as therapeutic targets for NDs require additional study [27,36].
1. Functions of Bacterial Extracellular Vesicles in Neuro immune system and their crosstalk
The neuro-immune system, which is made up of interactions between the neurological system and the immune system, is critical in neurodegenerative diseases (NDs) [38,44]. This neuro-immune systems help keep the homeostasis in balance. If this balance is distraught, it can lead to chronic inflammation, damage to neurons, and eventually NDs [45,46]. In terms of NDs, the neuro-immune system is made up of immune cells like microglia and astrocytes that become active when there is neuroinflammation [47]. When these cells become active, they release cytokines and chemokines that cause more inflammation and damage to neurons [47,48]. Peripheral immune cells, such as T cells and monocytes, can also promote neuroinflammation by crossing the blood-brain barrier and entering the central nervous system [48,49].
Recent studies have shown how important the microbiome-gut-brain axis is in NDs and how it affects the neuroimmune system [54]. Dysbiosis, which is an imbalance in the gut microbiome, has been linked to the development of NDs [54,55]. This could be because of small molecules from the gut, like lipopolysaccharides, affect the immune system [13,45]. Latest studies have looked at how BEVs and the neuro-immune system interact, which shows how BEVs might be able to change the immune response in NDs [13,17,43]. For example, BEVs made from the gut bacteria Bacteroides fragilis suppressed the immune response in a mouse model of multiple sclerosis [56]. It was found that the BEVs helped regulatory T cells grow (Table 2), regulatory T cells are very important for calming down immune responses and preventing autoimmunity [50].
BEVs from the gut bacteria Akkermansia muciniphila were demonstrated to diminish neuroinflammation in a mouse model of Parkinson's disease [23,24]. It was found that the BEVs decreased the amount of pro-inflammatory cytokines in the brain and increased the number of anti-inflammatory cytokines [13,15]. This suggests that the BEVs have a neuroprotective effect. A study looked at how BEVs from the gut bacteria Bifidobacterium bifidum might affect the immune system in an Alzheimer's disease animal model [21,53]. The researchers found that giving the BEVs to the mice led to less inflammation in the brain and better brain function [53]. These studies show that BEVs may be able to change the immune response in NDs, which means they may be a good way to treat these diseases [17,54]. But more research is needed to fully understand the mechanisms behind these effects and to figure out the best ways to use BEVs as medicines.
1. Risk factors of Bacterial Extracellular Vesicles in Autophagy Lysosomal Pathway
The autophagy-lysosomal pathway (ALP) (Figure 3) is a complicated cellular process that gets rid of damaged organelles, misfolded proteins, and invading pathogens by breaking them down and recycling them [57,58,59,60]. Dysregulation of the ALP has been linked to neurodegenerative diseases like Alzheimer's, Parkinson's, and Huntington's [58,61,62,63].
The ALP has three main types: macroautophagy, microautophagy, and chaperone-mediated autophagy [64,65]. Autophagy or macroautophagy, has been studied the most and it involves the formation of autophagosomes that engulf cytoplasmic parts and fuse with lysosomes to break them down [66,67].
ALP is very important in removing toxic proteins, like alpha-synuclein in PD, Aβ and phospho tau in AD, from building up in the brain and trying to clear this toxic proteins when activated [68,69,70,71]. Also, abnormal regulation of the ALP has been linked to the activation of inflammatory pathways and oxidative stress, both of which contribute to the development of NDs [49,72,73]. BEVs may be able to promote the ALP in different NDs, such as PD and AD [13,15,17]. Recent studies have shown that by controlling or promoting the ALP using BEVs [18], suggesting that BEVs could be a promising therapeutic target for treating NDs (Figure 4) focussing on macro autophagy, chaperone-mediated autophagy and micro autophagy [52,53,74].
The ALP is very important for keeping cells in balance, and its malfunction has been linked to the development of several NDs [68,75,76]. More research is needed to fully understand how the ALP works in NDs and to look into the possibility that BEVs could be used as a therapy to control the ALP [65,77,78]. As per the previous studies, BEVs may have an effect on the autophagy-lysosomal pathway, in turn helping or keeping cells in balance by getting rid of damaged organelles and protein clusters [79,80]. Autophagy is a strictly regulated process that involves the creation of autophagosomes, which are double-membrane vesicles [58,68]. These vesicles take in the toxic cytoplasmic materials and send it to lysosomes by fusing to form autolysosome to clear or broken down the engulfed proteins or organelles [81,82,83]. There are many hydrolytic enzymes in lysosomes that can break down the contents of autophagosomes into nutrients that can be used to make energy and change the shape of cells [84,85].
Studies have shown that BEVs can mess up the autophagy-lysosomal pathway, which makes it harder for cells to get rid of waste and causes toxic aggregates to build up [85,86]. For example, a recent study showed that BEVs made from Porphyromonas gingivalis, a pathogenic oral bacterium linked to Alzheimer's disease, could stop autophagy by stopping lysosomes from becoming acidic and stopping autophagosomes from breaking down [86,87,88]. BEVs made from Bacteroides fragilis, a common gut bacterium that can change the immune system, could stop autophagy in dendritic cells by stopping the fusion of autophagosomes and lysosomes [27,53,88].
On the other hand, some studies have shown that BEVs may actually be playing a protective role in the autophagy-lysosomal pathway by assisting to make new lysosomes and speeding up autophagic flux [53,84,86]. For example, a recent study showed that BEVs made from Lactobacillus acidophilus, a probiotic bacterium with anti-inflammatory properties, could improve autophagy flux by increasing lysosomal biogenesis and promoting lysosomal acidification [89,90]. It was also found that BEVs made from Akkermansia muciniphila linked to better metabolic health, could speed up the removal of misfolded proteins in a mouse model of Parkinson's disease by activating the autophagy-lysosomal pathway [24,90,91]. In neurodegenerative diseases, the connection between BEVs and the autophagy-lysosomal pathway is complicated and needs to be investigated more [24,90,92]. The possibility that BEVs could interfere with or improve this important way for cells to get rid of waste could have big effects on how neurodegenerative diseases start and how they can be treated.
1. Prospective applications of Bacterial Extracellular Vesicles in Therapeutics
In East Asian countries traditional Chinese Medicine (TCM), makes use of herbal concoctions to treat NDs such as AD and PD for many years [60,93,94]. However, BEV can become an effective alternative for such treatment [78,87,90]. Depending on their source and the surrounding environment, exogenous vesicles (EVs) have the potential to both trigger immune responses and induce anti-tumor responses; as a result, they may offer useful tools for the development of innovative cancer treatments [87,95].
Since BEVs can cross the blood-brain barrier, several recent studies have looked into how BEVs could be used to treat NDs [89]. They can change immune responses and protect neurons from damage [77,96]. Evidently, BEVs have the potential to be used as a drug delivery system to treat NDs [89]. BEVs can carry a wide range of cargo, such as proteins, nucleic acids, and small molecules. This makes them an ideal vehicle to deliver active agents to specific cells or parts of the brain [97]. In fact, a number of studies have shown that therapeutic cargo can be delivered by BEVs in nonclinical models of NDs [97,98,99]. For example, one research study used BEVs made from mesenchymal stem cells to deliver miR-133b to neurons in a mouse model of Alzheimer's disease [97,98] and this study illustrated the tremendous output in cognitive function and lowered the levels of amyloid beta [97,98,99].
It has been repeatedly found that BEV can not only change the immune system but also protect nerve cells, which makes them a very useful drug delivery system for treating NDs [99]. Reducing neuroinflammation, a characteristic feature of many NDs, has been demonstrated as an effect of BEVs on the brain's immune system [90,98]. Another very interesting and important factor is that BEVs protect neurons from damage by lowering oxidative stress and helping neurons stay alive [90,98].
It's important to remember that while using BEVs to treat NDs has a lot of potentials, there are also risks and challenges. For example, BEVs may interfere with the way lysosomes work and slow down the waste removal system of the cell, which can certainly cause toxic proteins and other cellular debris to build up and cause different kinds of issues [100,101]. Another challenge is the lack of information. While the research has shown some promising results, there is a major possibility that the lack of substantial information about the working of the pharmacokinetics, its safety, and effectiveness of BEVs in humans, especially when it comes to long-term use and possible immune responses can produce some side effects [78,92,101].
Even with these certain and uncertain challenges, the possible uses of BEVs in NDs are an interesting and intriguing area of research that needs more study and development [78,90]. With continued progress in our understanding of how NDs work and how BEVs affect these processes, BEVs may end up being a useful addition to our arsenal of neuroprotective and disease-modifying therapies [84]. Bacterial extracellular vesicles (BEVs) are starting to look like a good way to treat diseases that damage nerve cells (NDs) [90]. Recent preclinical studies on mouse models of NDs have shown that BEVs can change the immune response, lower neuroinflammation, and improve cognitive function [90,99]. Also, BEVs have been shown to protect neurons in the lab by preventing the buildup of misfolded proteins, encouraging autophagy, and improving lysosomal function [68,81,84].
On a practical note, several preclinical studies have looked into the possibility that BEVs could be used to deliver drugs to treat NDs [78,89,98]. BEVs used as drug delivery agents in model organisms have depicted some promising results. For example, BEVs were allowed to carry therapeutic agents like siRNA and drugs to specific brain cells [77,89,98]. The results showed that BEVs made from mesenchymal stem cells can deliver active siRNA that targets the tau protein in the brain and reduce tau phosphorylation and clumping in a mouse model of Alzheimer's disease [77,98]. The use of BEVs to treat NDs in a clinical setting is still in its early stages [101]. Clinical trials and several other studies are being conducted and reports are mostly positive.
1. Future insights of Bacterial Extracellular Vesicles
In recent years, there has been a lot of interest in the new field of research on the role of bacterial extracellular vesicles (BEVs) in neurodegenerative diseases (NDs). Even though we know more about how BEVs might be used in therapy, there is still a lot to learn about how they work. One important area of future research is to learn more about how BEVs interact with the autophagy-lysosomal pathway, as this may be a key part of how they affect NDs. Also, there needs to be more research on the possible risks of using BEVs as therapeutic agents, such as the possibility of bad immune responses and effects on other biological systems that were not intended.
The development of better and more targeted ways to get BEVs to the brain is another important area of future research. At the moment, not much is known about the factors that affect how BEVs are taken up and distributed in the brain, which may be important for their therapeutic effectiveness. Advances in nanotechnology and other delivery technologies may open up new ways to improve the targeting and delivery of BEVs to certain parts of the brain. Another big problem is that there aren't any standard protocols for isolating and describing BEVs. There are a lot of different ways to isolate BEVs, which can change the purity and make-up of the vesicles that are isolated. Also, because there is no agreement on how to define and group BEVs, it is hard to compare the results of different studies.
Another problem is that we don't know enough about how BEVs affect NDs and how they work. Several studies have shown that BEVs can change how the immune system responds, improve the survival of neurons, and help get rid of misfolded proteins. However, the exact molecular processes that cause these effects are not well understood. This makes it harder to improve BEV-based treatments for NDs. Concerns have also been raised about the ability of BEV to cause immunogenicity and toxicity, as well as effects that are not intended. Before BEVs can be used in clinical settings, these problems show how important it is to test their safety and make sure they are safe.
1. Conclusions
Clearly, bacterial extracellular vesicles (BEVs) have the potential to play an important role in the field of neurodegenerative diseases (NDs) and their therapeutics. According to the studies discussed in this thread, BEVs have immunomodulatory and neuroprotective properties that may be useful in the treatment of NDs. BEVs have demonstrated promise as prospective drug delivery systems for the treatment of neurodegenerative diseases (NDs) due to their ability to cross the blood-brain barrier (BBB) and target specific brain cells. However, there are obstacles and limitations that must be taken into account when researching and implementing BEVs in NDs. A lack of standardisation in BEV isolation and characterization methods, for instance, can result in inconsistent results and hinder the comparability of studies. In addition, a greater understanding of the mechanisms underlying the effects of BEVs on NDs requires more exhaustive research.
To completely realise the potential of BEVs in NDs, future research should focus on addressing these challenges and limitations. To ascertain the safety and efficacy of these therapies in humans, it is necessary to investigate the optimal dose, frequency, and duration of BEV treatment. In addition, additional research is required to better comprehend the underlying mechanisms by which BEVs exert their effects and to identify the BEVs most likely to be effective in treating NDs. In conclusion, BEVs hold great promise as a potential therapeutic approach for NDs, but additional research is required to completely comprehend their potential and overcome their limitations of use. The prospective benefits of BEVs in NDs make them an intriguing research topic that warrants further investigation.
Author Contributions
The research study was conceived and conceptualized by: M.L. and A.I.; drafting the manuscript, literature collection, methodology and editing the manuscript: A.I., K.L., X.-J.G., R.J., K.V., J.P, and M.L. Helped organize the manuscript and provided the technical support A.I., K.L., X.-J.G., R.J., K.V. and J.P.; Reviewed and edited the paper A.I., K.L., X.-J.G., R.J., K.V. and J.P.; funding acquisition and resources M.L. and A.I. All authors have read and agreed to the published version of the manuscript.
Acknowledgments
This study was supported by the Research Committee of Hong Kong Baptist University (HKBU-MPCF-003-2022-23), Hong Kong Health and Medical Research Fund (HMRF/17182541, HMRF/17182551, HMRF/09203776) and the General Research Fund from Research Grant Council (HKBU 12302620, 1230202, and 12101022) of Hong Kong Government. We would like to thank Dr. Carol Chu for her assistance in managing the laboratory and procuring the requirements for the experiments, and Mrs. Shilpi Shikha for providing picture design and technical writing. We would like to thank Dr. Martha Dahlen for her English editing of this manuscript.
Conflicts of interest: There are no conflict to declare.
References
- Iyaswamy, A.; Vasudevan, K.; Jayaraman, S.; Jaganathan, R.; Thakur, A.; Chang, R.C.; Yang, C. Editorial: Advances in Alzheimer's disease diagnostics, brain delivery systems, and therapeutics. Front Mol Biosci 2023, 10, 1162879. [Google Scholar] [CrossRef] [PubMed]
- Teunissen, C.E.; Verberk, I.M.W.; Thijssen, E.H.; Vermunt, L.; Hansson, O.; Zetterberg, H.; van der Flier, W.M.; Mielke, M.M.; Del Campo, M. Blood-based biomarkers for Alzheimer's disease: towards clinical implementation. Lancet Neurol 2022, 21, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S.J.; Fulton-Howard, B.; Goate, A. Interpretation of risk loci from genome-wide association studies of Alzheimer's disease. Lancet Neurol 2020, 19, 326–335. [Google Scholar] [CrossRef]
- Veitch, D.P.; Weiner, M.W.; Aisen, P.S.; Beckett, L.A.; DeCarli, C.; Green, R.C.; Harvey, D.; Jack, C.R., Jr.; Jagust, W.; Landau, S.M.; et al. Using the Alzheimer's Disease Neuroimaging Initiative to improve early detection, diagnosis, and treatment of Alzheimer's disease. Alzheimers Dement 2022, 18, 824–857. [Google Scholar] [CrossRef] [PubMed]
- Cuesta, C.M.; Guerri, C.; Urena, J.; Pascual, M. Role of Microbiota-Derived Extracellular Vesicles in Gut-Brain Communication. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Diaz-Garrido, N.; Badia, J.; Baldoma, L. Microbiota-derived extracellular vesicles in interkingdom communication in the gut. J Extracell Vesicles 2021, 10, e12161. [Google Scholar] [CrossRef]
- Zhao, G.; Jones, M.K. Role of Bacterial Extracellular Vesicles in Manipulating Infection. Infect Immun 2023, 91, e0043922. [Google Scholar] [CrossRef]
- Raghav, A.; Singh, M.; Jeong, G.B.; Giri, R.; Agarwal, S.; Kala, S.; Gautam, K.A. Extracellular vesicles in neurodegenerative diseases: A systematic review. Front Mol Neurosci 2022, 15, 1061076. [Google Scholar] [CrossRef]
- Yang, Y.; Boza-Serrano, A.; Dunning, C.J.R.; Clausen, B.H.; Lambertsen, K.L.; Deierborg, T. Inflammation leads to distinct populations of extracellular vesicles from microglia. J Neuroinflammation 2018, 15, 168. [Google Scholar] [CrossRef]
- Choi, J.; Kim, Y.K.; Han, P.L. Extracellular Vesicles Derived from Lactobacillus plantarum Increase BDNF Expression in Cultured Hippocampal Neurons and Produce Antidepressant-like Effects in Mice. Exp Neurobiol 2019, 28, 158–171. [Google Scholar] [CrossRef]
- Jones, L.B.; Kumar, S.; Bell, C.R.; Peoples, V.A.; Crenshaw, B.J.; Coats, M.T.; Scoffield, J.A.; Rowe, G.C.; Sims, B.; Matthews, Q.L. Effects of Pseudomonas aeruginosa on Microglial-Derived Extracellular Vesicle Biogenesis and Composition. Pathogens 2019, 8. [Google Scholar] [CrossRef]
- Lee, E.J.; Choi, Y.; Lee, H.J.; Hwang, D.W.; Lee, D.S. Human neural stem cell-derived extracellular vesicles protect against Parkinson's disease pathologies. J Nanobiotechnology 2022, 20, 198. [Google Scholar] [CrossRef]
- Vanaja, S.K.; Russo, A.J.; Behl, B.; Banerjee, I.; Yankova, M.; Deshmukh, S.D.; Rathinam, V.A.K. Bacterial Outer Membrane Vesicles Mediate Cytosolic Localization of LPS and Caspase-11 Activation. Cell 2016, 165, 1106–1119. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.; Wolf, J.M.; Prados-Rosales, R.; Casadevall, A. Through the wall: extracellular vesicles in Gram-positive bacteria, mycobacteria and fungi. Nat Rev Microbiol 2015, 13, 620–630. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Li, X.D.; Zhang, S.M.; Wang, H.W.; Wang, Y.L. Extracellular vesicles in neurodegenerative diseases: Insights and new perspectives. Genes Dis 2021, 8, 124–132. [Google Scholar] [CrossRef]
- Aires, I.D.; Ribeiro-Rodrigues, T.; Boia, R.; Ferreira-Rodrigues, M.; Girao, H.; Ambrosio, A.F.; Santiago, A.R. Microglial Extracellular Vesicles as Vehicles for Neurodegeneration Spreading. Biomolecules 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Wang, Y.; Zheng, J.C. Extracellular vesicles, from the pathogenesis to the therapy of neurodegenerative diseases. Transl Neurodegener 2022, 11, 53. [Google Scholar] [CrossRef]
- Yang, Z.; Gao, Z.; Yang, Z.; Zhang, Y.; Chen, H.; Yang, X.; Fang, X.; Zhu, Y.; Zhang, J.; Ouyang, F.; et al. Lactobacillus plantarum-derived extracellular vesicles protect against ischemic brain injury via the microRNA-101a-3p/c-Fos/TGF-beta axis. Pharmacol Res 2022, 182, 106332. [Google Scholar] [CrossRef]
- Kang, C.S.; Ban, M.; Choi, E.J.; Moon, H.G.; Jeon, J.S.; Kim, D.K.; Park, S.K.; Jeon, S.G.; Roh, T.Y.; Myung, S.J.; et al. Extracellular vesicles derived from gut microbiota, especially Akkermansia muciniphila, protect the progression of dextran sulfate sodium-induced colitis. PLoS One 2013, 8, e76520. [Google Scholar] [CrossRef]
- Haney, M.S.; Bohlen, C.J.; Morgens, D.W.; Ousey, J.A.; Barkal, A.A.; Tsui, C.K.; Ego, B.K.; Levin, R.; Kamber, R.A.; Collins, H.; et al. Identification of phagocytosis regulators using magnetic genome-wide CRISPR screens. Nat Genet 2018, 50, 1716–1727. [Google Scholar] [CrossRef]
- Konieczna, P.; Groeger, D.; Ziegler, M.; Frei, R.; Ferstl, R.; Shanahan, F.; Quigley, E.M.; Kiely, B.; Akdis, C.A.; O'Mahony, L. Bifidobacterium infantis 35624 administration induces Foxp3 T regulatory cells in human peripheral blood: potential role for myeloid and plasmacytoid dendritic cells. Gut 2012, 61, 354–366. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Lee, E.J.; Bae, I.H.; Myoung, K.; Kim, S.T.; Park, P.J.; Lee, K.H.; Pham, A.V.Q.; Ko, J.; Oh, S.H.; et al. Lactobacillus plantarum-derived extracellular vesicles induce anti-inflammatory M2 macrophage polarization in vitro. J Extracell Vesicles 2020, 9, 1793514. [Google Scholar] [CrossRef]
- Chelakkot, C.; Choi, Y.; Kim, D.K.; Park, H.T.; Ghim, J.; Kwon, Y.; Jeon, J.; Kim, M.S.; Jee, Y.K.; Gho, Y.S.; et al. Akkermansia muciniphila-derived extracellular vesicles influence gut permeability through the regulation of tight junctions. Exp Mol Med 2018, 50, e450. [Google Scholar] [CrossRef] [PubMed]
- Ashrafian, F.; Shahriary, A.; Behrouzi, A.; Moradi, H.R.; Keshavarz Azizi Raftar, S.; Lari, A.; Hadifar, S.; Yaghoubfar, R.; Ahmadi Badi, S.; Khatami, S.; et al. Akkermansia muciniphila-Derived Extracellular Vesicles as a Mucosal Delivery Vector for Amelioration of Obesity in Mice. Front Microbiol 2019, 10, 2155. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.J.; Chen, J.L.; Liao, J.F.; Chen, Y.H.; Chieu, M.W.; Ke, Y.Y.; Hsu, C.C.; Tsai, Y.C.; Hsieh-Li, H.M. Lactobacillus plantarum PS128 prevents cognitive dysfunction in Alzheimer's disease mice by modulating propionic acid levels, glycogen synthase kinase 3 beta activity, and gliosis. BMC Complement Med Ther 2021, 21, 259. [Google Scholar] [CrossRef]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The gut-brain axis: interactions between enteric microbiota, central and enteric nervous systems. Ann Gastroenterol 2015, 28, 203–209. [Google Scholar]
- Wang, H.; Long, T.; You, J.; Li, P.; Xu, Q. Bibliometric Visualization Analysis of Microbiome-Gut-Brain Axis from 2004 to 2020. Med Sci Monit 2022, 28, e936037. [Google Scholar] [CrossRef]
- Claudino Dos Santos, J.C.; Lima, M.P.P.; Brito, G.A.C.; Viana, G.S.B. Role of enteric glia and microbiota-gut-brain axis in parkinson disease pathogenesis. Ageing Res Rev 2023, 84, 101812. [Google Scholar] [CrossRef] [PubMed]
- Kargbo, R.B. Microbiome-Gut-Brain Axis Modulation: New Approaches in Treatment of Neuropsychological and Gastrointestinal Functional Disorders. ACS Med Chem Lett 2023, 14, 692–695. [Google Scholar] [CrossRef]
- Cattaneo, A.; Cattane, N.; Galluzzi, S.; Provasi, S.; Lopizzo, N.; Festari, C.; Ferrari, C.; Guerra, U.P.; Paghera, B.; Muscio, C.; et al. Association of brain amyloidosis with pro-inflammatory gut bacterial taxa and peripheral inflammation markers in cognitively impaired elderly. Neurobiol Aging 2017, 49, 60–68. [Google Scholar] [CrossRef]
- Sharon, G.; Sampson, T.R.; Geschwind, D.H.; Mazmanian, S.K. The Central Nervous System and the Gut Microbiome. Cell 2016, 167, 915–932. [Google Scholar] [CrossRef]
- Jung, J.H.; Kim, G.; Byun, M.S.; Lee, J.H.; Yi, D.; Park, H.; Lee, D.Y.; Group, K.R. Gut microbiome alterations in preclinical Alzheimer's disease. PLoS One 2022, 17, e0278276. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.Q.; Yu, L.L.; Qi, G.Y.; Mi, Y.S.; Wu, W.Q.; Lee, Y.K.; Zhai, Q.X.; Tian, F.W.; Chen, W. Can dietary patterns prevent cognitive impairment and reduce Alzheimer's disease risk: Exploring the underlying mechanisms of effects. Neurosci Biobehav Rev 2022, 135, 104556. [Google Scholar] [CrossRef] [PubMed]
- Sorboni, S.G.; Moghaddam, H.S.; Jafarzadeh-Esfehani, R.; Soleimanpour, S. A Comprehensive Review on the Role of the Gut Microbiome in Human Neurological Disorders. Clin Microbiol Rev 2022, 35, e0033820. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, P.J.; Cryan, J.F.; Dinan, T.G.; Clarke, G. Kynurenine pathway metabolism and the microbiota-gut-brain axis. Neuropharmacology 2017, 112, 399–412. [Google Scholar] [CrossRef]
- Ou, Z.; Deng, L.; Lu, Z.; Wu, F.; Liu, W.; Huang, D.; Peng, Y. Protective effects of Akkermansia muciniphila on cognitive deficits and amyloid pathology in a mouse model of Alzheimer's disease. Nutr Diabetes 2020, 10, 12. [Google Scholar] [CrossRef]
- McQuade, A.; Blurton-Jones, M. Microglia in Alzheimer's Disease: Exploring How Genetics and Phenotype Influence Risk. J Mol Biol 2019, 431, 1805–1817. [Google Scholar] [CrossRef]
- Brochard, V.; Combadiere, B.; Prigent, A.; Laouar, Y.; Perrin, A.; Beray-Berthat, V.; Bonduelle, O.; Alvarez-Fischer, D.; Callebert, J.; Launay, J.M.; et al. Infiltration of CD4+ lymphocytes into the brain contributes to neurodegeneration in a mouse model of Parkinson disease. J Clin Invest 2009, 119, 182–192. [Google Scholar] [CrossRef]
- Bhattarai, Y.; Kashyap, P.C. Parkinson's disease: Are gut microbes involved? Am J Physiol Gastrointest Liver Physiol 2020, 319, G529–G540. [Google Scholar] [CrossRef]
- Sampson, T.R.; Debelius, J.W.; Thron, T.; Janssen, S.; Shastri, G.G.; Ilhan, Z.E.; Challis, C.; Schretter, C.E.; Rocha, S.; Gradinaru, V.; et al. Gut Microbiota Regulate Motor Deficits and Neuroinflammation in a Model of Parkinson's Disease. Cell 2016, 167, 1469–1480 e1412. [Google Scholar] [CrossRef]
- Ordonez-Rodriguez, A.; Roman, P.; Rueda-Ruzafa, L.; Campos-Rios, A.; Cardona, D. Changes in Gut Microbiota and Multiple Sclerosis: A Systematic Review. Int J Environ Res Public Health 2023, 20. [Google Scholar] [CrossRef] [PubMed]
- Freedman, S.N.; Shahi, S.K.; Mangalam, A.K. The "Gut Feeling": Breaking Down the Role of Gut Microbiome in Multiple Sclerosis. Neurotherapeutics 2018, 15, 109–125. [Google Scholar] [CrossRef] [PubMed]
- Roig-Carles, D.; Willms, E.; Fontijn, R.D.; Martinez-Pacheco, S.; Mager, I.; de Vries, H.E.; Hirst, M.; Sharrack, B.; Male, D.K.; Hawkes, C.A.; et al. Endothelial-Derived Extracellular Vesicles Induce Cerebrovascular Dysfunction in Inflammation. Pharmaceutics 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Cantoni, C.; Lin, Q.; Dorsett, Y.; Ghezzi, L.; Liu, Z.; Pan, Y.; Chen, K.; Han, Y.; Li, Z.; Xiao, H.; et al. Alterations of host-gut microbiome interactions in multiple sclerosis. EBioMedicine 2022, 76, 103798. [Google Scholar] [CrossRef] [PubMed]
- Rutsch, A.; Kantsjo, J.B.; Ronchi, F. The Gut-Brain Axis: How Microbiota and Host Inflammasome Influence Brain Physiology and Pathology. Front Immunol 2020, 11, 604179. [Google Scholar] [CrossRef]
- Sterling, K.G.; Dodd, G.K.; Alhamdi, S.; Asimenios, P.G.; Dagda, R.K.; De Meirleir, K.L.; Hudig, D.; Lombardi, V.C. Mucosal Immunity and the Gut-Microbiota-Brain-Axis in Neuroimmune Disease. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Thakur, S.; Dhapola, R.; Sarma, P.; Medhi, B.; Reddy, D.H. Neuroinflammation in Alzheimer's Disease: Current Progress in Molecular Signaling and Therapeutics. Inflammation 2023, 46, 1–17. [Google Scholar] [CrossRef]
- Patrycy, M.; Chodkowski, M.; Krzyzowska, M. Role of Microglia in Herpesvirus-Related Neuroinflammation and Neurodegeneration. Pathogens 2022, 11. [Google Scholar] [CrossRef]
- Zhang, F.; Jiang, L. Neuroinflammation in Alzheimer's disease. Neuropsychiatr Dis Treat 2015, 11, 243–256. [Google Scholar] [CrossRef]
- Eskandari, S.K.; Sulkaj, I.; Melo, M.B.; Li, N.; Allos, H.; Alhaddad, J.B.; Kollar, B.; Borges, T.J.; Eskandari, A.S.; Zinter, M.A.; et al. Regulatory T cells engineered with TCR signaling-responsive IL-2 nanogels suppress alloimmunity in sites of antigen encounter. Sci Transl Med 2020, 12. [Google Scholar] [CrossRef]
- Ferreira, T.G.; Trindade, C.; Bell, P.; Teixeira-Ferreira, A.; Perales, J.E.; Vommaro, R.C.; Domingues, R.; Ferreira, E.O. Identification of the alpha-enolase P46 in the extracellular membrane vesicles of Bacteroides fragilis. Mem Inst Oswaldo Cruz 2018, 113, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Zhang, Y.; Chen, S.; Zeng, Y.; Fu, X.; Chen, T.; Luo, S.; Zhang, X. The role of the probiotic Akkermansia muciniphila in brain functions: insights underpinning therapeutic potential. Crit Rev Microbiol 2023, 49, 151–176. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Guo, M.; Zhao, J.; Zhang, H.; Wang, G.; Chen, W. Bifidobacterium breve intervention combined with environmental enrichment alleviates cognitive impairment by regulating the gut microbiota and microbial metabolites in Alzheimer's disease mice. Front Immunol 2022, 13, 1013664. [Google Scholar] [CrossRef] [PubMed]
- Haas-Neill, S.; Forsythe, P. A Budding Relationship: Bacterial Extracellular Vesicles in the Microbiota-Gut-Brain Axis. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Bicknell, B.; Liebert, A.; Borody, T.; Herkes, G.; McLachlan, C.; Kiat, H. Neurodegenerative and Neurodevelopmental Diseases and the Gut-Brain Axis: The Potential of Therapeutic Targeting of the Microbiome. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Reparaz, J.; Mielcarz, D.W.; Wang, Y.; Begum-Haque, S.; Dasgupta, S.; Kasper, D.L.; Kasper, L.H. A polysaccharide from the human commensal Bacteroides fragilis protects against CNS demyelinating disease. Mucosal Immunol 2010, 3, 487–495. [Google Scholar] [CrossRef]
- Zhu, Z.; Yang, C.; Iyaswamy, A.; Krishnamoorthi, S.; Sreenivasmurthy, S.G.; Liu, J.; Wang, Z.; Tong, B.C.; Song, J.; Lu, J.; et al. Balancing mTOR Signaling and Autophagy in the Treatment of Parkinson's Disease. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- Feng, Y.; He, D.; Yao, Z.; Klionsky, D.J. The machinery of macroautophagy. Cell Res 2014, 24, 24–41. [Google Scholar] [CrossRef]
- Song, J.X.; Malampati, S.; Zeng, Y.; Durairajan, S.S.K.; Yang, C.B.; Tong, B.C.; Iyaswamy, A.; Shang, W.B.; Sreenivasmurthy, S.G.; Zhu, Z.; et al. A small molecule transcription factor EB activator ameliorates beta-amyloid precursor protein and Tau pathology in Alzheimer's disease models. Aging Cell 2020, 19, e13069. [Google Scholar] [CrossRef]
- Iyaswamy, A.; Krishnamoorthi, S.K.; Zhang, H.; Sreenivasmurthy, S.G.; Zhu, Z.; Liu, J.; Su, C.F.; Guan, X.J.; Wang, Z.Y.; Cheung, K.H.; et al. Qingyangshen mitigates amyloid-beta and Tau aggregate defects involving PPARalpha-TFEB activation in transgenic mice of Alzheimer's disease. Phytomedicine 2021, 91, 153648. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, C.; Liu, J.; Chun-Kit Tong, B.; Zhu, Z.; Malampati, S.; Gopalkrishnashetty Sreenivasmurthy, S.; Cheung, K.H.; Iyaswamy, A.; Su, C.; et al. A Curcumin Derivative Activates TFEB and Protects Against Parkinsonian Neurotoxicity in Vitro. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Iyaswamy, A.; Wang, X.; Krishnamoorthi, S.; Kaliamoorthy, V.; Sreenivasmurthy, S.G.; Kumar Durairajan, S.S.; Song, J.X.; Tong, B.C.; Zhu, Z.; Su, C.F.; et al. Theranostic F-SLOH mitigates Alzheimer's disease pathology involving TFEB and ameliorates cognitive functions in Alzheimer's disease models. Redox Biol 2022, 51, 102280. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Zhu, Z.; Tong, B.C.; Iyaswamy, A.; Xie, W.J.; Zhu, Y.; Sreenivasmurthy, S.G.; Senthilkumar, K.; Cheung, K.H.; Song, J.X.; et al. A stress response p38 MAP kinase inhibitor SB202190 promoted TFEB/TFE3-dependent autophagy and lysosomal biogenesis independent of p38. Redox Biol 2020, 32, 101445. [Google Scholar] [CrossRef] [PubMed]
- Fung, T.Y.; Iyaswamy, A.; Sreenivasmurthy, S.G.; Krishnamoorthi, S.; Guan, X.J.; Zhu, Z.; Su, C.F.; Liu, J.; Kan, Y.; Zhang, Y.; et al. Klotho an Autophagy Stimulator as a Potential Therapeutic Target for Alzheimer's Disease: A Review. Biomedicines 2022, 10. [Google Scholar] [CrossRef]
- Guan, X.; Iyaswamy, A.; Sreenivasmurthy, S.G.; Su, C.; Zhu, Z.; Liu, J.; Kan, Y.; Cheung, K.H.; Lu, J.; Tan, J.; et al. Mechanistic Insights into Selective Autophagy Subtypes in Alzheimer's Disease. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Menzies, F.M.; Fleming, A.; Rubinsztein, D.C. Compromised autophagy and neurodegenerative diseases. Nat Rev Neurosci 2015, 16, 345–357. [Google Scholar] [CrossRef]
- Yang, C.; Su, C.; Iyaswamy, A.; Krishnamoorthi, S.K.; Zhu, Z.; Yang, S.; Tong, B.C.; Liu, J.; Sreenivasmurthy, S.G.; Guan, X.; et al. Celastrol enhances transcription factor EB (TFEB)-mediated autophagy and mitigates Tau pathology: Implications for Alzheimer's disease therapy. Acta Pharm Sin B 2022, 12, 1707–1722. [Google Scholar] [CrossRef]
- Wong, E.; Cuervo, A.M. Autophagy gone awry in neurodegenerative diseases. Nat Neurosci 2010, 13, 805–811. [Google Scholar] [CrossRef]
- Selvarasu, K.; Singh, A.K.; Iyaswamy, A.; Gopalkrishnashetty Sreenivasmurthy, S.; Krishnamoorthi, S.; Bera, A.K.; Huang, J.D.; Durairajan, S.S.K. Reduction of kinesin I heavy chain decreases tau hyperphosphorylation, aggregation, and memory impairment in Alzheimer's disease and tauopathy models. Front Mol Biosci 2022, 9, 1050768. [Google Scholar] [CrossRef]
- Sreenivasmurthy, S.G.; Iyaswamy, A.; Krishnamoorthi, S.; Reddi, R.N.; Kammala, A.K.; Vasudevan, K.; Senapati, S.; Zhu, Z.; Su, C.F.; Liu, J.; et al. Bromo-protopine, a novel protopine derivative, alleviates tau pathology by activating chaperone-mediated autophagy for Alzheimer's disease therapy. Front Mol Biosci 2022, 9, 1030534. [Google Scholar] [CrossRef]
- Zhu, Z.; Liu, L.F.; Su, C.F.; Liu, J.; Tong, B.C.; Iyaswamy, A.; Krishnamoorthi, S.; Sreenivasmurthy, S.G.; Guan, X.J.; Kan, Y.X.; et al. Corynoxine B derivative CB6 prevents Parkinsonian toxicity in mice by inducing PIK3C3 complex-dependent autophagy. Acta Pharmacol Sin 2022, 43, 2511–2526. [Google Scholar] [CrossRef]
- Sarkar, S.; Malovic, E.; Harischandra, D.S.; Ngwa, H.A.; Ghosh, A.; Hogan, C.; Rokad, D.; Zenitsky, G.; Jin, H.; Anantharam, V.; et al. Manganese exposure induces neuroinflammation by impairing mitochondrial dynamics in astrocytes. Neurotoxicology 2018, 64, 204–218. [Google Scholar] [CrossRef]
- Tong, B.C.; Wu, A.J.; Huang, A.S.; Dong, R.; Malampati, S.; Iyaswamy, A.; Krishnamoorthi, S.; Sreenivasmurthy, S.G.; Zhu, Z.; Su, C.; et al. Lysosomal TPCN (two pore segment channel) inhibition ameliorates beta-amyloid pathology and mitigates memory impairment in Alzheimer disease. Autophagy 2022, 18, 624–642. [Google Scholar] [CrossRef] [PubMed]
- Gaurav, I.; Thakur, A.; Kumar, G.; Long, Q.; Zhang, K.; Sidu, R.K.; Thakur, S.; Sarkar, R.K.; Kumar, A.; Iyaswamy, A.; et al. Delivery of Apoplastic Extracellular Vesicles Encapsulating Green-Synthesized Silver Nanoparticles to Treat Citrus Canker. Nanomaterials (Basel) 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Tong, B.C.; Huang, A.S.; Wu, A.J.; Iyaswamy, A.; Ho, O.K.; Kong, A.H.; Sreenivasmurthy, S.G.; Zhu, Z.; Su, C.; Liu, J.; et al. Tetrandrine ameliorates cognitive deficits and mitigates tau aggregation in cell and animal models of tauopathies. J Biomed Sci 2022, 29, 85. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.B.; Liu, J.; Tong, B.C.; Wang, Z.Y.; Zhu, Z.; Su, C.F.; Sreenivasmurthy, S.G.; Wu, J.X.; Iyaswamy, A.; Krishnamoorthi, S.; et al. TFEB, a master regulator of autophagy and biogenesis, unexpectedly promotes apoptosis in response to the cyclopentenone prostaglandin 15d-PGJ2. Acta Pharmacol Sin 2022, 43, 1251–1263. [Google Scholar] [CrossRef] [PubMed]
- Panaro, M.A.; Benameur, T.; Porro, C. Extracellular Vesicles miRNA Cargo for Microglia Polarization in Traumatic Brain Injury. Biomolecules 2020, 10. [Google Scholar] [CrossRef]
- Go, V.; Bowley, B.G.E.; Pessina, M.A.; Zhang, Z.G.; Chopp, M.; Finklestein, S.P.; Rosene, D.L.; Medalla, M.; Buller, B.; Moore, T.L. Extracellular vesicles from mesenchymal stem cells reduce microglial-mediated neuroinflammation after cortical injury in aged Rhesus monkeys. Geroscience 2020, 42, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Sil, S.; Singh, S.; Chemparathy, D.T.; Chivero, E.T.; Gordon, L.; Buch, S. Astrocytes & Astrocyte derived Extracellular Vesicles in Morphine Induced Amyloidopathy: Implications for Cognitive Deficits in Opiate Abusers. Aging Dis 2021, 12, 1389–1408. [Google Scholar] [CrossRef]
- Jones, E.; Stentz, R.; Telatin, A.; Savva, G.M.; Booth, C.; Baker, D.; Rudder, S.; Knight, S.C.; Noble, A.; Carding, S.R. The Origin of Plasma-Derived Bacterial Extracellular Vesicles in Healthy Individuals and Patients with Inflammatory Bowel Disease: A Pilot Study. Genes (Basel) 2021, 12. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, C.; Sun, J.; Shen, H.M.; Wang, J.; Yang, C. Impairment of the autophagy-lysosomal pathway in Alzheimer's diseases: Pathogenic mechanisms and therapeutic potential. Acta Pharm Sin B 2022, 12, 1019–1040. [Google Scholar] [CrossRef] [PubMed]
- Iyaswamy, A.; Krishnamoorthi, S.K.; Liu, Y.W.; Song, J.X.; Kammala, A.K.; Sreenivasmurthy, S.G.; Malampati, S.; Tong, B.C.K.; Selvarasu, K.; Cheung, K.H.; et al. Yuan-Hu Zhi Tong Prescription Mitigates Tau Pathology and Alleviates Memory Deficiency in the Preclinical Models of Alzheimer's Disease. Front Pharmacol 2020, 11, 584770. [Google Scholar] [CrossRef] [PubMed]
- Sreenivasmurthy, S.G.; Iyaswamy, A.; Krishnamoorthi, S.; Senapati, S.; Malampati, S.; Zhu, Z.; Su, C.F.; Liu, J.; Guan, X.J.; Tong, B.C.; et al. Protopine promotes the proteasomal degradation of pathological tau in Alzheimer's disease models via HDAC6 inhibition. Phytomedicine 2022, 96, 153887. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Shen, Y.; Zhang, S.; Xiao, Y.; Shi, S. Salmonella Interacts With Autophagy to Offense or Defense. Front Microbiol 2020, 11, 721. [Google Scholar] [CrossRef] [PubMed]
- Keller, M.D.; Torres, V.J.; Cadwell, K. Autophagy and microbial pathogenesis. Cell Death Differ 2020, 27, 872–886. [Google Scholar] [CrossRef]
- Villageliu, D.N.; Samuelson, D.R. The Role of Bacterial Membrane Vesicles in Human Health and Disease. Front Microbiol 2022, 13, 828704. [Google Scholar] [CrossRef]
- Chen, M.F.; Lu, M.S.; Hsieh, C.C.; Chen, W.C. Porphyromonas gingivalis promotes tumor progression in esophageal squamous cell carcinoma. Cell Oncol (Dordr) 2021, 44, 373–384. [Google Scholar] [CrossRef]
- Di Gioia, S.; Daniello, V.; Conese, M. Extracellular Vesicles' Role in the Pathophysiology and as Biomarkers in Cystic Fibrosis and COPD. Int J Mol Sci 2022, 24. [Google Scholar] [CrossRef]
- Li, M.; Lee, K.; Hsu, M.; Nau, G.; Mylonakis, E.; Ramratnam, B. Lactobacillus-derived extracellular vesicles enhance host immune responses against vancomycin-resistant enterococci. BMC Microbiol 2017, 17, 66. [Google Scholar] [CrossRef]
- Choi, J.H.; Moon, C.M.; Shin, T.S.; Kim, E.K.; McDowell, A.; Jo, M.K.; Joo, Y.H.; Kim, S.E.; Jung, H.K.; Shim, K.N.; et al. Lactobacillus paracasei-derived extracellular vesicles attenuate the intestinal inflammatory response by augmenting the endoplasmic reticulum stress pathway. Exp Mol Med 2020, 52, 423–437. [Google Scholar] [CrossRef]
- Kim, D.K.; Lee, J.; Kim, S.R.; Choi, D.S.; Yoon, Y.J.; Kim, J.H.; Go, G.; Nhung, D.; Hong, K.; Jang, S.C.; et al. EVpedia: a community web portal for extracellular vesicles research. Bioinformatics 2015, 31, 933–939. [Google Scholar] [CrossRef]
- Cadwell, K. Crosstalk between autophagy and inflammatory signalling pathways: balancing defence and homeostasis. Nat Rev Immunol 2016, 16, 661–675. [Google Scholar] [CrossRef] [PubMed]
- Iyaswamy, A.; Krishnamoorthi, S.K.; Song, J.X.; Yang, C.B.; Kaliyamoorthy, V.; Zhang, H.; Sreenivasmurthy, S.G.; Malampati, S.; Wang, Z.Y.; Zhu, Z.; et al. NeuroDefend, a novel Chinese medicine, attenuates amyloid-beta and tau pathology in experimental Alzheimer's disease models. J Food Drug Anal 2020, 28, 132–146. [Google Scholar] [CrossRef] [PubMed]
- Ng, R.C.; Jian, M.; Ma, O.K.; Bunting, M.; Kwan, J.S.; Zhou, G.J.; Senthilkumar, K.; Iyaswamy, A.; Chan, P.K.; Li, M.; et al. Chronic oral administration of adipoRon reverses cognitive impairments and ameliorates neuropathology in an Alzheimer's disease mouse model. Mol Psychiatry 2021, 26, 5669–5689. [Google Scholar] [CrossRef] [PubMed]
- Mombelli, L.; Fidanza, M.A.; Roncari, G. [Evaluation of a new method of determination of glomerular filtration rate by use of radioactive methylglucamine iothalamate]. Acta Isot (Padova) 1968, 8, 21–29. [Google Scholar]
- Lee, J.Y.; Kim, H.S. Extracellular Vesicles in Neurodegenerative Diseases: A Double-Edged Sword. Tissue Eng Regen Med 2017, 14, 667–678. [Google Scholar] [CrossRef]
- Keighron, C.N.; Avazzadeh, S.; Goljanek-Whysall, K.; McDonagh, B.; Howard, L.; Ritter, T.; Quinlan, L.R. Extracellular Vesicles, Cell-Penetrating Peptides and miRNAs as Future Novel Therapeutic Interventions for Parkinson's and Alzheimer's Disease. Biomedicines 2023, 11. [Google Scholar] [CrossRef]
- Reza-Zaldivar, E.E.; Hernandez-Sapiens, M.A.; Gutierrez-Mercado, Y.K.; Sandoval-Avila, S.; Gomez-Pinedo, U.; Marquez-Aguirre, A.L.; Vazquez-Mendez, E.; Padilla-Camberos, E.; Canales-Aguirre, A.A. Mesenchymal stem cell-derived exosomes promote neurogenesis and cognitive function recovery in a mouse model of Alzheimer's disease. Neural Regen Res 2019, 14, 1626–1634. [Google Scholar] [CrossRef]
- Ruan, J.; Miao, X.; Schluter, D.; Lin, L.; Wang, X. Extracellular vesicles in neuroinflammation: Pathogenesis, diagnosis, and therapy. Mol Ther 2021, 29, 1946–1957. [Google Scholar] [CrossRef]
- Yuan, Y.; Sun, J.; You, T.; Shen, W.; Xu, W.; Dong, Q.; Cui, M. Extracellular Vesicle-Based Therapeutics in Neurological Disorders. Pharmaceutics 2022, 14. [Google Scholar] [CrossRef]
- Cano, A.; Ettcheto, M.; Bernuz, M.; Puerta, R.; Esteban de Antonio, E.; Sanchez-Lopez, E.; Souto, E.B.; Camins, A.; Marti, M.; Pividori, M.I.; et al. Extracellular vesicles, the emerging mirrors of brain physiopathology. Int J Biol Sci 2023, 19, 721–743. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Bacterial Extracellular Vesicles (BEVs) structure and its components used for its therapeutic function and defensive mechanisms.
Figure 1.
Bacterial Extracellular Vesicles (BEVs) structure and its components used for its therapeutic function and defensive mechanisms.
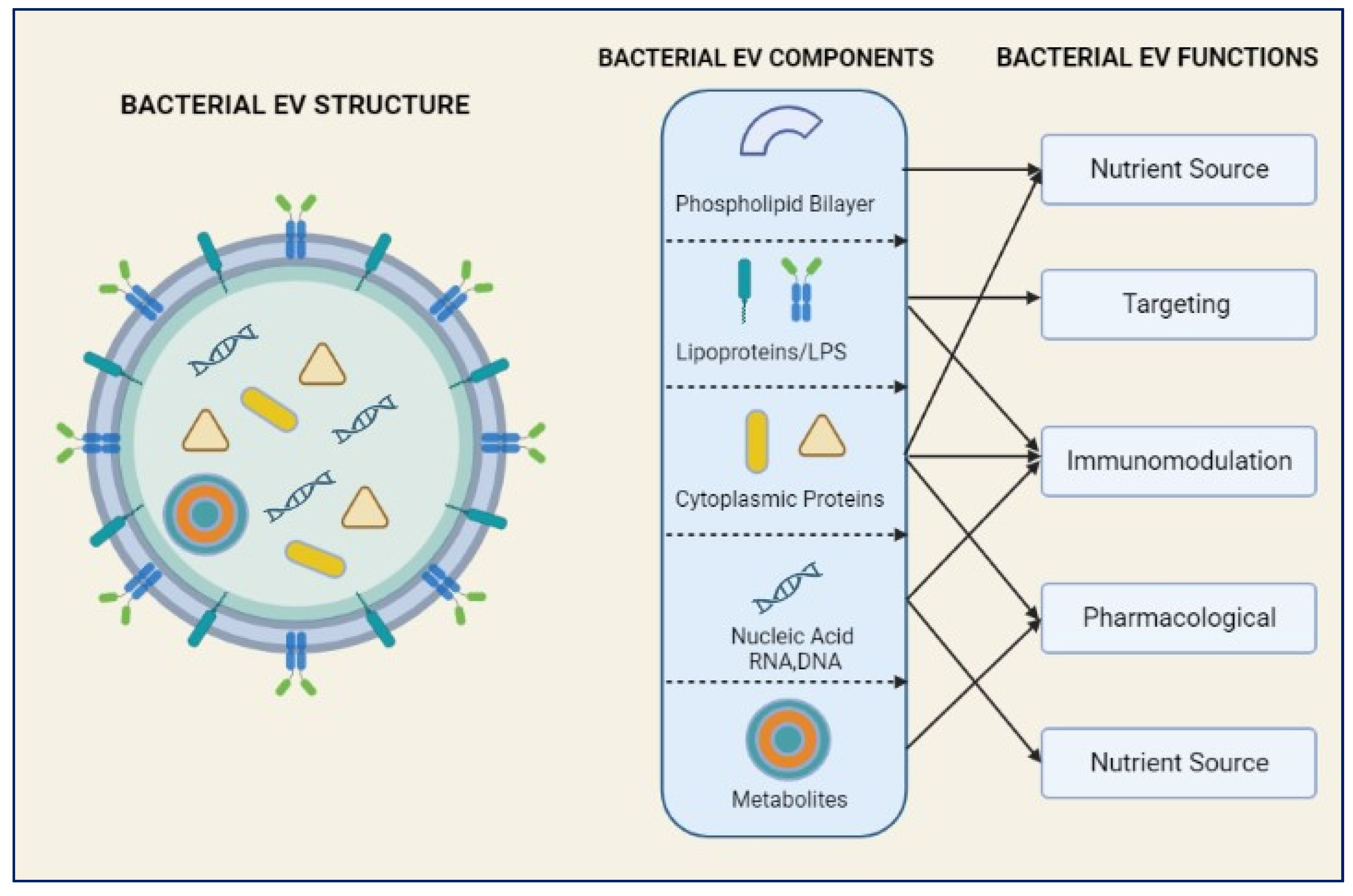
Figure 2.
Offensive and Defensive roles Bacterial Extracellular Vesicles in Neurodegenerative Disease and its prospective applications in therapeutics.
Figure 2.
Offensive and Defensive roles Bacterial Extracellular Vesicles in Neurodegenerative Disease and its prospective applications in therapeutics.
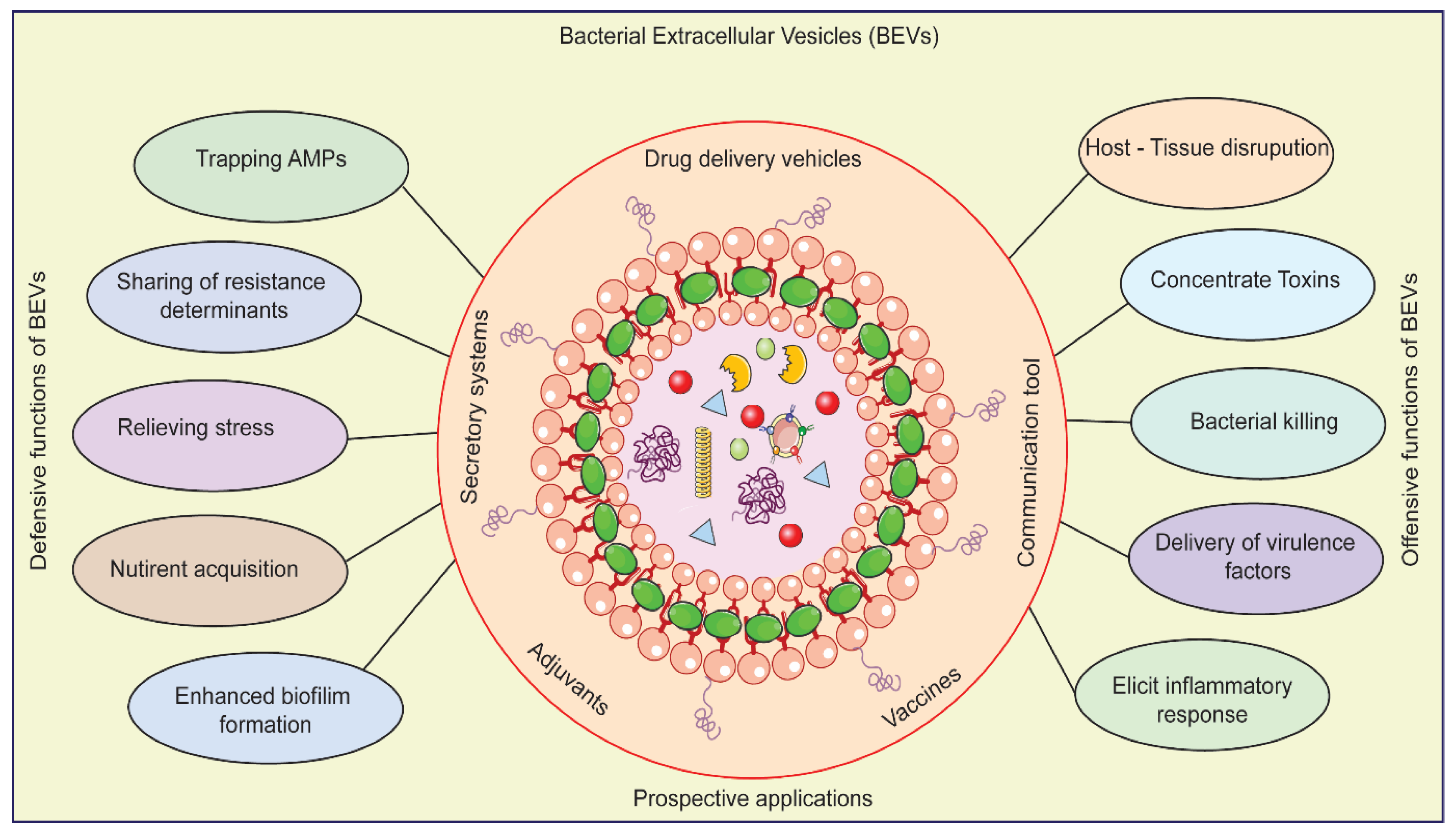
Figure 3.
The process of initiation, nucleation and maturation of autophagy including the autophagosome formation and autophagy flux for the formation of autolysosome.
Figure 3.
The process of initiation, nucleation and maturation of autophagy including the autophagosome formation and autophagy flux for the formation of autolysosome.
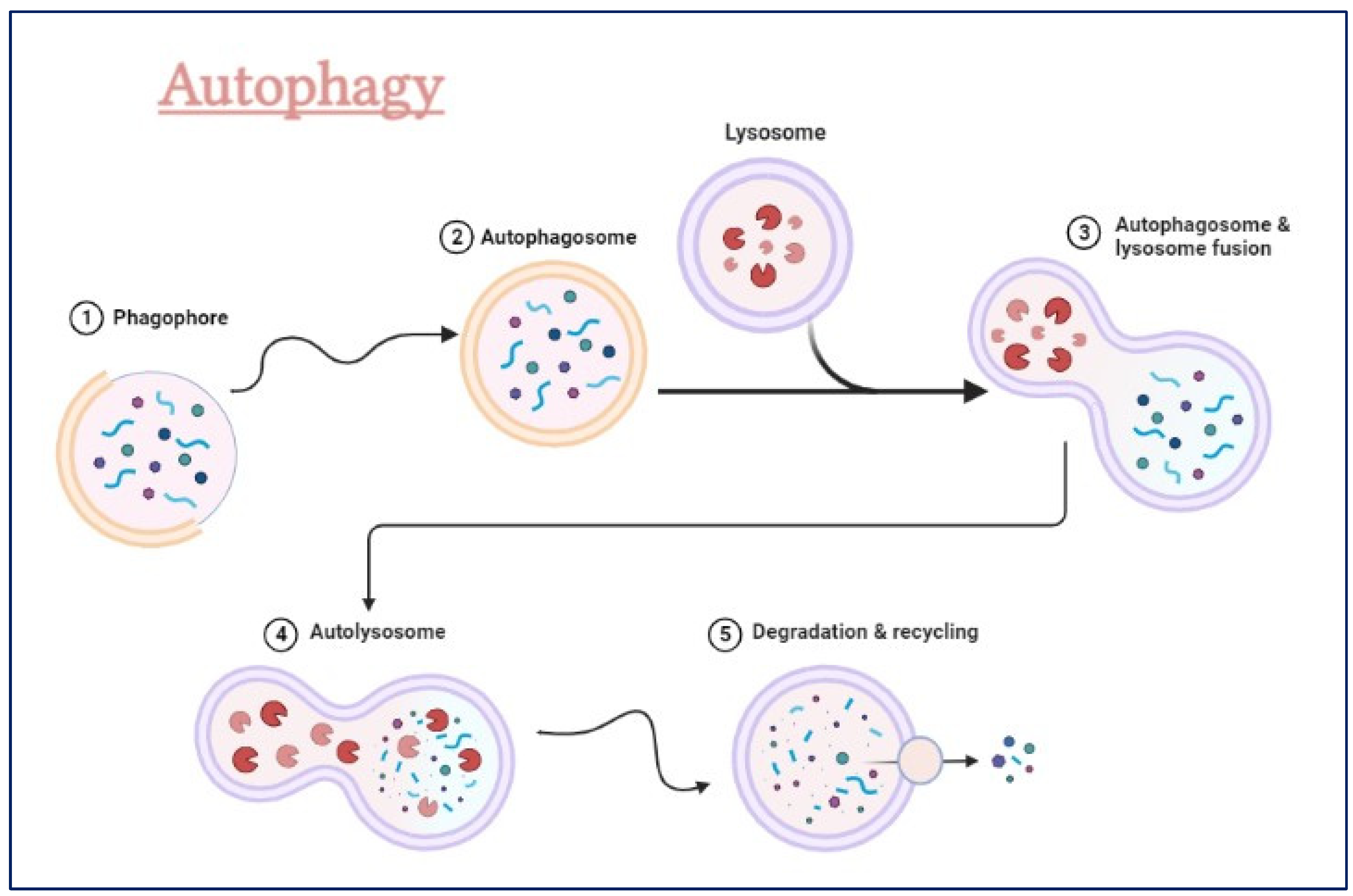
Figure 4.
Three major types of autophagy lysosomal pathways in NDs and other brain diseases are macro autophagy, chaperone-mediated autophagy and micro autophagy.
Figure 4.
Three major types of autophagy lysosomal pathways in NDs and other brain diseases are macro autophagy, chaperone-mediated autophagy and micro autophagy.
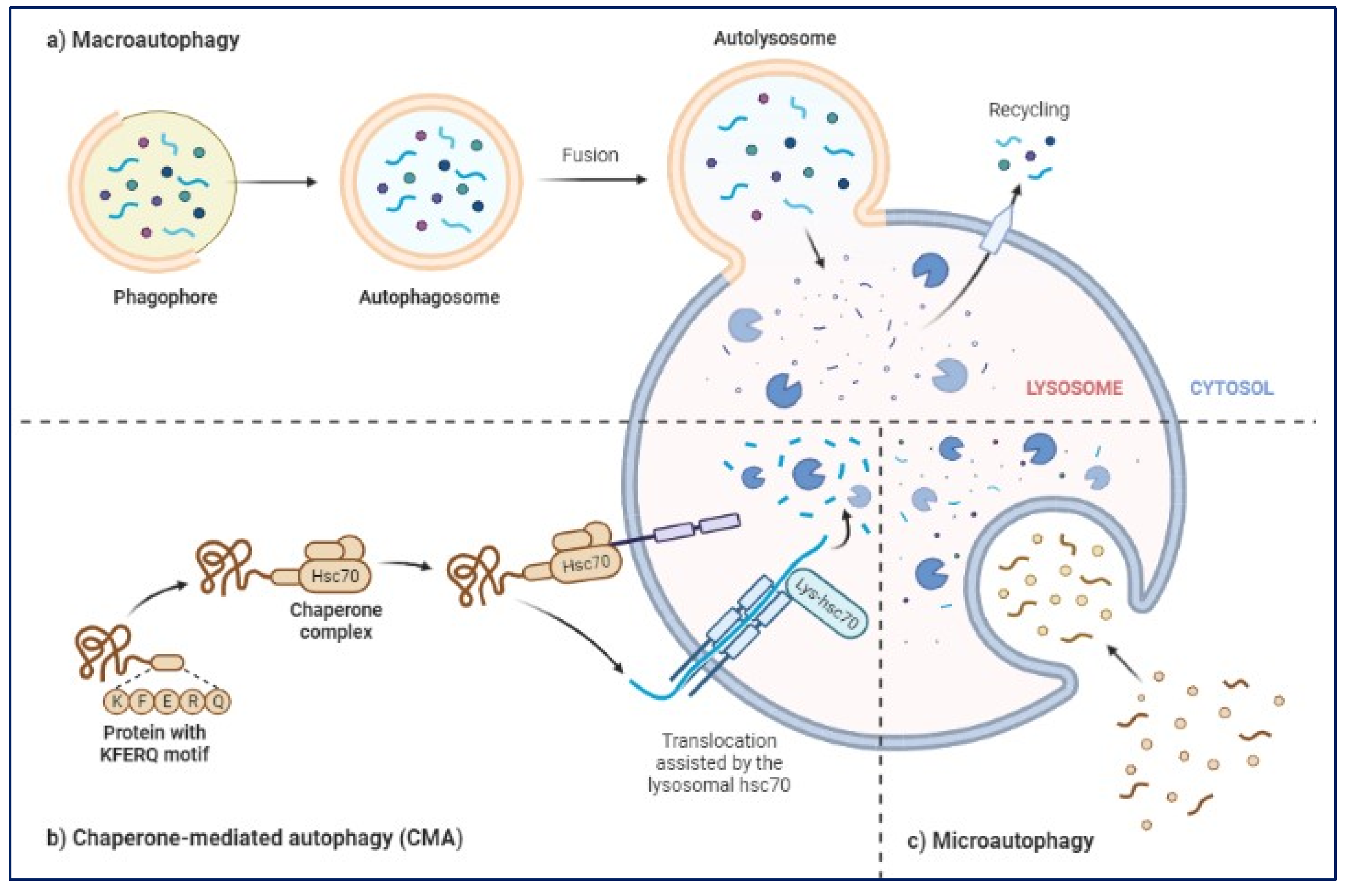
Table 2.
Studies that describing how BEVs and the neuro-immune system interact in the pathogenesis and therapeutics of NDs.
Table 2.
Studies that describing how BEVs and the neuro-immune system interact in the pathogenesis and therapeutics of NDs.
| Study | BEV Source | Model/System | Key Findings |
|---|---|---|---|
| [21,50,51] | Bacteroides fragilis | Mouse model of MS | BEVs promoted the expansion of regulatory T cells, dampening immune responses and preventing autoimmunity |
| [23,24,52] | Akkermansia muciniphila | Mouse model of PD | BEVs reduced neuroinflammation by decreasing pro-inflammatory cytokine levels while raising anti-inflammatory cytokine levels. |
| [21,22,53] | Bifidobacterium bifidum | Mouse model of AD | BEVs reduced neuroinflammation and improved cognitive function |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



