
Submitted:
26 June 2023
Posted:
27 June 2023
You are already at the latest version
Abstract
Abstract: OTUD6A is a deubiquitinase that plays crucial roles in various human diseases. However, the precise regulatory mechanism of OTUD6A remains unclear. In this study, we found that OTUD6A significantly inhibited the production of type I interferon. Consistently, peritoneal macrophages and bone marrow-derived macrophages from Otud6a-/- mice produced more type I interferon after virus infection compared to cells from WT mice. Otud6a-/- mice also exhibited increased resistance to lethal HSV-1 and VSV infections, as well as LPS attack due to decreased inflammatory responses. Mechanistically, mass spectrometry results revealed that UBC13 was an OTUD6A-interacting protein, and the interaction significantly enhanced after HSV-1 stimulation. Taken together, our findings suggest that OTUD6A plays a crucial role in the innate immune response and may serve as a potential therapeutic target for infectious disease.
Keywords:
OTUD6A
; IFNβ
; Ubc13
; NF-κB
; innate immunity
1. Introduction
The ubiquitin-proteasome pathway (UPP) is the most broadly and effective mechanism for modulating in a broad range of cellular physiology, including antigen presentation[1,2], homeostasis[3,4], circadian rhythm[5,6], signal transduction[7,8], protein trafficking[9,10], cell differentiation[11,12], immune defense[13,14] and inflammation[15]. The Ubiquitin–Proteasome System (UPS) is composed of ubiquitin (Ub), ubiquitin-activating enzyme (E1), ubiquitin-binding enzyme (E2), ubiquitin-ligase (E3)[16,17,18], 26S proteasome[19], deubiquitinating enzymes (DUBs)[20,21,22] and target proteins. The E1→E2→E3 enzymatic cascade covalently attaches the ubiquitin C-terminus to target-specific proteins. This cascade is initiated by E1 and forms a thioester bond between the carboxyl group of the terminal glycine residue of ubiquitin in an ATP dependent manner[23]. The activated ubiquitin is then transferred to the Cys residue of the E2 enzyme via transesterification, and E2 transfers ubiquitin to the substrate protein via E3. Ultimately, the 26S proteasome recognizes and degraded the labeled Ub-protein conjugates[24,25] , removing most short-lived and aberrant peptides and proteins from eukaryotic cells.
Ubiquitination is involved in regulating signal transduction in antiviral immune responsesby precisely activating downstream proteins through pattern recognition receptor signaling, which initiates the innate immune response[26].Ubiquitylation is a reversible covalent modification that regulates the stability, activity and localization of target proteins. Deubiquitylating enzymes (DUBs), also known as deubiquitinases, are proteases that remove ubiquitin conjugates from diverse substrate proteins[27]. The human genome contains sixteen ovarian tumor domain-containing 1 (OTU) family members, making them the second largest DUB subfamily and they have emerged as regulators of important signal cascades[28,29].
OTUD1 functions as a metastasis suppressive factor by repressing the metastasis of breast cancer through the deubiquitination of SMAD7[30]. Additionally, OTUD1 downregulates the induction of Type I IFN by cleaving the viral infection-induced atypical K6-linked ubiquitination of IRF3[31]. A study has shown that OTUB1, a Lys48-specific deubiquitinating enzyme, preferentially binds to the Ubc13~Ub thiolester, which is regulated by free ubiquitin. Free ubiquitin can bind to a second site in OTUB1 and increase the affinity with UBC13, preventing ubiquitin transfer and suppressing the accumulation of polyubiquitin[32,33,34]. Another study has shown that OTUD3, an acetylation-dependent deubiquitinase, shuts down the innate antiviral immune response by directly hydrolyzing lysine 63 (Lys63)-linked polyubiquitination of MAVS[14].
OTUD6A is a member of OTU domain family DUBs. Previous studies have shown that OTUD6A regulates cell proliferation and tumorigenesis. For instance, OTUD6A deubiquitinates and stabilizes Drp1 to regulate mitochondrial morphology in colon cancer, and it plays a critical role in promoting tumor cell resistance to chemoradiotherapy by deubiquitinating and stabilizing TopBP1[35,36]. OTUD6A specifically promotes prostate tumorigenesis by stabilizing Brg1, AR and c-Myc through the removal of Brg1, AR and c-Myc polyubiquitination, respectively[37,38]. Ubiquitination- dependent processes are critical for producing type I interferon in the antiviral signaling pathway[39,40]. However, whether OTUD6A affects the innate immune response remains unknown.
In this study, we used a library of DUBs expression plasmids to screen for DUBs that specifically regulate classical natural immune signaling pathways and found that overexpression of OTUD6A suppressed type I interferon expression. It was found that OTUD6A interacts with Ubc13 and not only participates in the regulation of innate immunity but also regulates the inflammatory response. In conclusion, our findings suggest that OTUD6A plays a dual role in regulating innate immunity and inflammatory responses and could provide a potential therapeutic target for related immune diseases.
2. Materials and Methods
2.1. Cell culture
HEK293T cells, Hela cells, iBMDM cells, primary bone marrow derived macrophage (BMDM) cells and peritoneal macrophage (PM) cells were cultured in DMEM medium supplemented with 10% FBS (Gibco, Rockville, MD, USA) and 100 U/ml penicillin and 100 mg/ml streptomycin at 37°C under 5% CO2 atmosphere. HEK293T cells, Hela cells, iBMDM cells were maintained in our laboratory. For the isolation of mouse primary peritoneal macrophages (PMs), cells were collected from the lavage of the peritoneal cavity from mice that were pre-stimulated with thioglycolate (TG) for 3 days. For BMDMs, bone marrow cells were isolated from femurs and tibiae of 8 to 12-week-old Otud6a-/- mice and WT mice. Cells were cultured with 20 ng/ml recombinant murine GM-CSF (315-03; Peprotech) in a 10 cm dish for 7 days before experiments.
2.2. Reagents and antibodies
Polyethylenimine(PEI) (764582, Sigma-Aldrich) and jetPRIME (114-15, Polyplus) were used for transfection. DSS (Cat# D8906) was purchased from Sigma-Aldrich. Listeria monocytogenes were provided by the laboratory of Zengfan Jiang, Peking University. The antibodies used in this study were as follows: Anti-OTUD6A (Proteintech, Cat# 24486-1-AP); Anti-NF-κB P65(CST, Cat# 8242); Anti-p-p65 (Ser536) (93H1)( CST, Cat# 3033); Anti-IκBα(CST, Cat# 9242); Anti-p-IκBα(CST, Cat# 5209); Goat anti-mouse IgG, HRP conjugate (Proteintech, Cat# SA00001-1); Goat anti-rabbit IgG, HRP conjugate(Proteintech, Cat# SA00001-2).
2.3. Plasmid construction
The plasmids used in this study were prepared using standard molecular biology techniques, and coding sequences completely verified. Ifnβ promoter luciferase reporter plasmids (IFN-β–luc) and mammalian expression plasmids for human STING, MAVS, TBK1, IRF3, TRIF and RIG-1 were previously described [41].All truncations deletions and mutants were constructed using standard molecular biology techniques. Each truncation, deletion and mutant were confirmed by sequencing.
2.4. Luciferase Reporter Assay
293T cells were seeded onto 24-well plates and transfected with the luciferase reporter plasmid, along with the specified expression plasmids or an empty vector (VEC). After 24 hours, luciferase activity was measured using the Dual-Luciferase Reporter Assay System (Promega) following the manufacturers’ instructions.
2.5. RNA Extraction and Real-Time PCR (qRT-PCR)
Total RNA from cells, blood and tissues was extracted using Trizol reagent (Invitrogen, Waltham, MA, USA) and then reverse transcribed into cDNA using HiScript II Q RT SuperMix (Vazyme, Nanjing, China). Quantitative Real-time PCR was performed using the ChamQ Universal SYBR qPCR Master Mix (Vazyme, Nanjing, China) on a 7500 Fast real-time PCR machine (Applied Biosystems, Waltham, MA, USA). The qRT-PCR Data were analyzed using the Livak method (2−ΔΔCt) using GraphPad Prism Version 8.0 (GraphPad Software, La Jolla, CA). The data are presented as relative abundance normalized to GAPDH mRNA levels.
Primer sequences were listed as follows:
hIfnβ-F: 5’-AGGACAGGATGAACTTTGAC-3’
hIfnβ-R: 5’-TGATAGACATTAGCCAGGAG-3’
hGapdh-F: 5’- GAAGGTGAAGGTCGGAGTC-3’
hGapdh-R: 5’- GAAGATGGTGATGGGATTTC-3’
mIfnb-F: 5’-TCCGAGCAGAGATCTTCAGGAA-3’
mIfnb-R: 5’-TGCAACCACCACTCATTCTGAG-3’
mGapdh-F: 5’- TCACCACCATGGAGAAGGC-3’
mGapdh-R: 5’- GCTAAGCAGTTGGTGGTGCA-3’
mIfit1-F: 5’- CAAGGCAGGTTTCTGAGGAGT-3’
mIfit1-R: 5’- ACCATCAGCATTCTCTCCCAT-3’
mIfit2-F:5’- AGTACAACGAGTAAGGAGTCACT-3’
mIfit2-R: 5’-AGGCCAGTATGTTGCACATGG-3’
mCxcl2-F: 5’-GGGCGGTCAAAAAGTTTGC-3’
mCxcl2-R: 5’-GTTAGCCTTGCCTTTGTTCAGTATC-3’
mCxcl10-F: 5’-CCAAGTGCTGCCGTCATTTTC-3’
mCxcl10-R: 5’-GGCTCGCAGGGATGATTTCAA-3’
mifna1-F: 5’-GCCTTGACACTCCTGGTACAAATGAG-3’
mifna1-R: 5’-CAGCACATTGGCAGAGGAAGACAG-3’
mifna2-F: 5’-CTCTGTGCTTTCCTCGTGATGC-3’
mifna2-R: 5’-CCTCCTCATCTGTGCCAGGACC-3’
mifna4-F: 5’-CCTGTGTGATGCAGGAACC-3’
mifna4-R: 5’-TCACCTCCCAGGCACAGA-3’
mifna5’-F: 5’-TTTGGATTCCCACAGGAGAAGGTGG-3’
mifna5’-R: 5’-TCATTGAGCTGCTGATGGACTT-3’
mIL1b-F: 5’-CAACCAACAAGTGATATTCTCCATG-3’
mIL1b-R: 5’-GATCCACACTCTCCAGCTGCA-3’
mIL6-F: 5’-TCTGCAAGAGACTTCCATCCAGTTGC-3’
mIL6-R: 5’-AGCCTCCGACTTGTGAAGTGGT-3’
mISG15’-F: 5’-TGAGGTCTTTCTGACGCAGACTGT-3’
mISG15’-R: 5’-TCAGGCGCAAATGCTTGATCACTG-3’
mTNFα-F: 5’-ACAGAAAGCATGATCCGCG-3’
mTNFα-R: 5’-GCCCCCCATCTTTTGGG-3’
mIL12b-F: 5’-TGGGAGTACCCTGACTCCTG-3’
mIL12b-R: 5’-AGGAACGCACCTTTCTGGTT-3’
VSV-G-F: 5’-CAAGTCAAAATGCCCAAGAGTCACA-3’
VSV-G-R: 5’-TTTCCTTGCATTGTTCTACAGATGG-3’
HSV-1-F: 5’-GCAGAAGGTCTCCGGTAAT-3’
HSV-1-R: 5’-CGCTACGATACGACACCAA-3’
2.6. RNA sequencing (RNA-seq)
Whole RNA of cells with specific treatment were purified using RNeasy Mini Kit (QIAGEN NO. 74104). The transcriptome library for sequencing was generated using VAHTSTM mRNA-seq v2 Library Prep Kit for Illumina (Vazyme Biotech, Nanjing, China) according to the manufacturer’s recommendations. Following library preparation, the libraries were sequenced on Illumina Hiseq X Ten platform using (2 × 150 bp) paired-end module. The raw images were transformed into raw reads by base calling using CASAVA softwares (https://support.illumina.com.cn/sequencing/sequencing_software/casava.html). Subsequently, the raw reads in a Fastq format were processed using in-house PERL scripts. Clean reads were obtained by removing reads with adapters, reads with more than 5% unknown bases and low-quality reads (defined as reads with more than 50% low-quality bases, where a low-quality base has a sequencing quality below 10). Additionally, the Q20 and Q30 scores, as well as the GC content of the clean data, were calculated (Vazyme Biotech, Nanjing, China). The original data of the RNA-seq has been uploaded to the GEO DataSets.
2.7. Virus Infection in cell and mice
Wild-type C57BL/6 mice were obtained from the Department of Laboratory Animal Science of Peking University Health Science Center, Beijing. The mice were bred and housed under specific pathogen-free conditions. For all in vivo experiments, age- and sex-matched mice were used, unless otherwise specified. Six-week-old mice were infected with HSV-1 virus (5 × 107 plaque-forming unit (PFU) virus) or VSV (3×108 PFU per mouse). At the indicated times after infection, the mice were sacrificed, and blood samples were collected. The expression of cell cytokine and viral gene were analyzed using RT- qPCR.
All animals care and mouse experiments were conducted in accordance with ethical guidelines and approved by the Animal Care and Use Committee of Peking University. The project was assigned the license number is LA2016239. Otud6a-/- mice were purchased from Shanghai Model Organisms Center, Shanghai.
2.8. Mass Spectrometry
To identify proteins that potentially interact with OTUD6A, purified Flag-OTUD6A samples from 293T and Hela cells were subjected to mass spectrometry analysis. The purification was performed using Flag antibody prior to the mass spectrometry experiments.
2.9. Coimmunoprecipitation and immunoblot assays
HEK293 cells and Hela cells were seeded on 10 cm dishes (1×107 cells/dish). The cells were then transfected with a total of 10 μg of Flag-OTUD6A expression plasmids or an empty plasmid. After 24 hours of transfection, the cells were lysed in lysis buffer (0.5% Triton X-100, 20mM HEPES (pH 7.4), 150mM NaCl, 12.5mM β-glycerolphosphate, 1.5mM MgCl2, 2mM EGTA, 10mM NaF, 1mM Na3VO4, 2 mM DTT) supplemented with protease inhibitors. The lysates were centrifuged, and the supernatant was incubated with anti-Flag antibodies at 4°C overnight. The following day, prewashed protein A/G beads (Pierce) were added to the lysates and incubated at 4°C for 4 hours. The beads were then washed four times with cold PBS and eluted with SDS sample buffer containing DTT by boiling for 10 minutes, in preparation for western blotting.
2.10. Statistical Analysis
Differences between experimental and control groups were evaluated using Student t-test when comparing two groups of data, or one-way ANOVA analysis when comparing more than two groups of data. Statistical analysis and data visualization were performed using GraphPad Prism 8.0 software. The data were presented as mean ± standard deviation (SD). The p values were as follows: ns p>0.05, *p< 0.05, **p< 0.01, ***p< 0.001 and ****p<0.0001.
3. Results
3.1. OTUD6A overexpression inhibits the production of type I IFN
To investigate the impact of the OTUDs family on the innate immune response, we constructed all the human OTUDs expression plasmids. In addition to transfecting deubiquitinases, we co-transfected STING and cGAS in 293T cells to activate the expression of Ifnβ. Luciferase reporter assays demonstrated that among the OTUDs tested, OTUD6A exhibited the most significant inhibition of Ifnβ expression (Figure 1a), indicating its potential as a negative regulator of the cGAS-STING signaling pathway. To further assess its impact on other canonical pattern recognition receptors(PRRs) such as RIG-I and innate immunity downstream proteins like MAVS, TBK1, IRF3, TRIF, and STING, we co-transfected OTUD6A and observed that it significantly inhibited the induction of Ifnβ by RIG-I/MAVS, cGAS/STING, TBK1/IRF3, and TRIF (Figure 1b), suggesting its involvement in these signal pathways and role as a component of innate immune signaling pathway. Additionally, we explored the impact of OTUD6A on the innate immune responses mediated by the DNA virus HSV-1 and RNA viruses VSV and SeV. As hypothesized, overexpression of OTUD6A led to a significant inhibition of Ifnβ expression following infection with any of these viruses (Figure 1c), thus confirming its negative regulatory role in the expression of Ifnβ.
3.2. OTUD6A deficiency enhances antiviral innate immunity in Vitro
To gain further insights into the function of OTUD6A in the innate immune responses, we generated Otud6a-/- mice and infected peritoneal macrophages (Figure 2a, Figure S1) and BMDMs (Figure 2b) isolated from both wild-type and Otud6a-/- mice with Sev. We then assessed the mRNA levels of type I interferon using RT-qPCR. Our results showed that the expression of Ifna1, Ifna2, Ifna4, and Ifnβ was significantly increased in Otud6a-/- peritoneal macrophages and BMDMs, confirming that OTUD6A played a critical role as a negative regulator of the innate immune response. We also performed whole-transcriptome sequencing on wild-type and Otud6a-/- iBMDM cells infected with HSV-1 and analyzed the top antiviral response-related terms from GO biological processes, which indicated a strong correlation between innate immunity signal pathways and OTUD6A (Figure 2c). Moreover, the expression of antiviral-related genes was significantly higher in cells from Otud6a-/- mice after HSV-1 infection (Figure 2d). These findings suggested that OTUD6A was an important negative regulator of innate immunity.
3.3. OTUD6A deficiency enhances antiviral innate immunity in Vivo
In vitro experiments demonstrated that the absence of OTUD6A enhanced innate immunity. To investigate the function of OTUD6A in vivo, we established an acute infection model by injecting HSV-1 into mice. Our results showed that Otud6a-/- mice had increased virus resistance and a higher survival rate than wild-type mice (Figure 3a). We also extracted mouse brain tissue on the fifth day after virus infection and detected the virus load. The plaque results indicated that the virus titer in the brain of Otud6a-/- mice was lower than that in wild-type mice (Figure 3b). Furthermore, we collected blood from mice infected with the virus for different days and found that Otud6a-/- mice produced more type I interferon and stronger antiviral-related factors (Ifnβ, Ifit1, Ifit2, and Cxcl10) compared to wild-type mice (Figure 3c). We next investigated if OTUD6A had a similar effect on RNA viruses. To this end, we injected WT and Otud6a-/- mice with vesicular stomatitis virus (VSV) and observed that Otud6a-/- mice had a higher survival rate and lower viral RNA levels (Figure 3d,e, FigureS2). Additionally, the expression of antiviral-related factors, such as Ifna2, Ifna4, Ifna5, and Ifnβ, was significantly higher in the blood of Otud6a-/- mice than in wild-type mice (Figure 3f). These findings demonstrated that OTUD6A played a critical role in facilitating innate immune responses to viral infections.
3.4. OTUD6A deficiency increases HSV-1 induced inflammatory responses
A recent study showed that OTUD6A can promote inflammation response during tumorigenesis[42]. We isolated primary peritoneal macrophages and bone marrow-derived macrophages from WT and Otud6a-/- mice and stimulated them with LPS. RT-qPCR showed that compared with wild-type cells, the expression of inflammatory factors, such as Il1b, Il6, and Tnf-α, decreased in both Otud6a-/- PMC and Otud6a-/- BMDM (Figure 4a,b and Figure S3a,b). Our results also showed that OTUD6A facilitated inflammation response, which is consistent with previous work.
To explore the role of OTUD6A in vivo, we established an acute infection model by infecting mice with LPS. The results showed that compared with WT mice, Otud6a-/- mice had higher survival rates (Figure 4c) and less body weight loss (Figure 4d). Moreover, Otud6a-/- mice had higher serum Ifnβ and Isg15 levels than WT mice (Figure 4E). Consistently, Otud6a-/- mice has much higher survival rates compared with WT mice after 2% DSS treatment (Figure S3c). Otud6a-/- mice also has higher survival rates and less body weight loss after listeria infection (Figure S3d,e). These results showed that OTUD6A promoted induction of proinflammatory cytokines after LPS treatment and plays a critical role in regulating inflammation in vivo.
3.5. OTUD6A participates in the regulation of NF-κB mediated inflammation signaling pathway via UBC13.
OTUD6A is a critical component of antiviral innate immunity and inflammatory signaling pathways, but its specific mechanism remains unclear. To further investigate the mechanism, we induced mouse peritoneal macrophages and infected them with LPS. Western blotting results showed that Otud6a-/- macrophages exhibited reduced levels of phosphorylated p65 compared to WT cells, while the phosphorylated and total IκBα levels increased (Figure 5a). This suggested that the deletion of OTUD6A inhibited the degradation process of phosphorylated IκBα, indicating that NF-κB pathway isn’t efficiently activated. Recent research has shown that OTUD6A directly binds to the NACHT domain of the NLRP3 inflammasome and selectively cleaves K48-linked polyubiquitin chains to enhance the stability of NLRP3. In conclusion, OTUD6A could promote inflammation.
To identify potential target proteins interacting with OTUD6A, we overexpressed Flag-OTUD6A or empty vector in HEK 293 and Hela cells and enriched them with Flag antibody for mass spectrometry analysis. Our results showed that the ubiquitin conjugating enzyme UBE2N (Ubc13) was a major interacting protein of OTUD6A (Figure 5b). Ubc13 is a critical E2-conjugating enzyme in regulation of immunity and inflammation, which can prevent the conversion of regulatory T cells into effector-like T cells[43], promote K63-Linked polyubiquitination of NLRP3[44] and confer processivity to TRAF6 ubiquitin ligase activity[45]. Immunoprecipitation analysis confirmed that the interaction between OTUD6A and Ubc13 was present in the resting state and significantly enhanced after HSV-1 stimulation (Figure 5c). We also investigated the role of Cys87, a key enzyme activity site of Ubc13, on their interaction and found that the loss of this site did not affect their interaction (Figure 5d). Furthermore, truncation of the first 50 or 100 amino acids at the N-terminal of OTUD6A significantly reduced its interaction with Ubc13, suggesting that OTUD6A interacted with Ubc13 through its N-terminal rather than its OTU structural domain (Figure 5e,f). These findings suggested that OTUD6A may rely on Ubc13 to participate in the regulation of NF-κB-mediated inflammation signaling pathways.
4. Discussion
The balance between immunity and inflammation responses is dynamic under steady-state conditions, but can quickly shift in response to pathogen invasion[46,47]. The family of deubiquitinases plays an essential role in regulating this balance, as different deubiquitinases may either up- or down-regulate innate immunity or inflammation response[48,49,50]. In this study, we have demonstrated that OTUD6A can attenuate the anti-virus innate immunity response and promote inflammation response.
Our findings demonstrate that overexpression of OTUD6A led to the downregulation of Ifnβ expression induced by RIG-I/MAVS, cGAS/STING, TBK1/IRF3, or TRIF overexpression. These results indicate that OTUD6A had a broad inhibitory effect on the production pathway of Ifnβ. It is worth noting that OTUB1 and OTUB2 negatively regulate the expression of Ifnβ induced by VISA-mediated pathway, but not TBK1-mediated pathway[51]. The downregulation of Ifnβ expression during VSV, SeV, or HSV-1 infection was also observed upon overexpression of OTUD6A. In vitro experiments demonstrated that OTUD6A-deficient BMDMs and PMCs expressed more interferons when infected with SeV compared to WT. Furthermore, in vivo experiments revealed that Otud6a-/- mice expressed more interferons when infected with VSV or HSV-1 and were more resistant to death than WT mice. NGS analysis showed that Otud6a-/- iBMDM expressed more interferons when infected with HSV-1 compared to WT.
In vitro LPS treatment in Otud6a-/- BMDMs and PMCs showed that Otud6a-/- mice were more resistant to death induced by LPS or DSS compared to WT. These findings are consistent with the work of Xin Liu et al[42], which showed that OTUD6A directly bound to the NACHT domain of the NLRP3 inflammasome and selectively cleaved K48-linked polyubiquitin chains from NLRP3 at K430 and K689 to enhance the stability of NLRP3, leading to increased Il1β levels and inflammation[42]. In vivo DSS treatment in Otud6a-/- mice also confirmed that OTUD6A promoted the induction of proinflammatory cytokines.
To elucidate the mechanism by which OTUD6A participates in the immune response, we employed immunoprecipitation (IP) and mass spectrometry (MS) to enrich and identify potential interacting proteins of OTUD6A. Our results revealed that OTUD6A could interact with Ubc13, and further verification experiments demonstrated that this interaction was dependent on the N-terminal of Ubc13. Ubc13 is a well-known E2 ubiquitin-conjugating enzyme and immune receptor, which has an important role in the process of tagging target proteins with lysine 63-linked polyubiquitin chains that are essential for the activation of NF-κB[52,53]. Recent research has shown that dendritic cell-specific deletion of OTUB1 impaired cytokine production after T. gondii infection by increasing the stability of Ubc13. Conversely, OTUD1 physically interacted with receptor-interacting serine/threonine-protein kinase 1 (RIPK1) and selectively cleaved K63-linked polyubiquitin chains from RIPK1 to inhibit the recruitment of NF-κB essential modulator (NEMO), thereby inhibiting the inflammatory response[53]. This controversy suggests the diverse roles of deubiquitinases in regulating immune and inflammatory responses, indicating that they have redundant and opposite effects on inflammation response pathways. Furthermore, we found that Otud6a-/- PMC exhibited a reduced level of phosphorylated p65, but increased levels of phosphorylated and total IκBα, indicating that OTUD6A promotes the degradation process of pIκBα.
In summary, our findings demonstrated that OTUD6A plays a critical role in regulating both innate immunity and inflammatory response. Our data reveal that OTUD6A can both inhibit innate immunity and promote inflammatory response, highlighting its importance in accelerating the cellular response to deadly virus or bacterial infections. These results provided new insights for the treatment of infectious and inflammatory diseases, emphasizing the potential therapeutic value of targeting OTUD6A to modulate immune and inflammatory responses.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, F.Y, C.W and S.L; methodology, S.L, Z.L and G.L.; software, G.L.; validation, Z.L and G.L; formal analysis, G.L; investigation, Z.L, G.L and S.L; resources, F.Y and C.W; data curation, Z.L and G.L; writing—original draft preparation, Z.L,G.L, Y.J and Z.Z; writing—review and editing, C.W and F.Y; visualization, Z.L and G.L; supervision, F.Y; project administration, F.Y; funding acquisition, F.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Beijing Natural Science Foundation (Z210014), the National Key Research and Development Program of China (2021YFC2302602, 2020YFA0707800), The National Natural Science Foundation of China (31570891; 31872736; 32022028; 81991505; 82201928), Peking University Clinical + X (PKU2020LCXQ009), the Peking University Medicine Fund (PKU2020LCXQ009), Zhuhai Science and Technology Innovation Bureau (ZH22036302200063PWC to Z.Y.), China Postdoctoral Science Foundation (2022M710265 to H.Y.), and a grant from Tianjin Natural Science Foundation of China (No. 21JCQNJC01870 to D.W.) and Incubation fund of Tianjin Third Central Hospital (No. 2019YNR6 to D.W.).
Institutional Review Board Statement
The study was conducted according to the Peking University Animal Care and Use Committee. The project license number is LA2016240.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ebstein, F.; Keller, M.; Paschen, A.; Walden, P.; Seeger, M.; Burger, E.; Kruger, E.; Schadendorf, D.; Kloetzel, P. M.; Seifert, U., Exposure to Melan-A/MART-126-35 tumor epitope specific CD8(+)T cells reveals immune escape by affecting the ubiquitin-proteasome system (UPS). Sci Rep 2016, 6, 25208. [CrossRef]
- Katayama, S.; Kukita, T.; Ishikawa, E.; Nakashima, S.; Masuda, S.; Kanda, T.; Akiyama, H.; Teshima, R.; Nakamura, S., Apple polyphenols suppress antigen presentation of ovalbumin by THP-1-derived dendritic cells. Food Chem 2013, 138, (2-3), 757-61. [CrossRef]
- Liu, J.; Guan, D.; Dong, M.; Yang, J.; Wei, H.; Liang, Q.; Song, L.; Xu, L.; Bai, J.; Liu, C.; Mao, J.; Zhang, Q.; Zhou, J.; Wu, X.; Wang, M.; Cong, Y. S., UFMylation maintains tumour suppressor p53 stability by antagonizing its ubiquitination. Nat Cell Biol 2020, 22, (9), 1056-1063. [CrossRef]
- Wang, B.; Jie, Z. L.; Joo, D. H.; Ordureau, A.; Liu, P.; Gan, W. J.; Guo, J. P.; Zhang, J. F.; North, B. J.; Dai, X. P.; Cheng, X. H.; Bian, X. W.; Zhang, L. Q.; Harper, J. W.; Sun, S. C.; Wei, W. Y., TRAF2 and OTUD7B govern a ubiquitin-dependent switch that regulates mTORC2 signalling. Nature 2017, 545, (7654), 365-+. [CrossRef]
- Liu, J.; Zou, X.; Gotoh, T.; Brown, A. M.; Jiang, L.; Wisdom, E. L.; Kim, J. K.; Finkielstein, C. V., Distinct control of PERIOD2 degradation and circadian rhythms by the oncoprotein and ubiquitin ligase MDM2. Sci Signal 2018, 11, (556). [CrossRef]
- Chen, S.; Yang, J.; Yang, L.; Zhang, Y.; Zhou, L.; Liu, Q.; Duan, C.; Mieres, C. A.; Zhou, G.; Xu, G., Ubiquitin ligase TRAF2 attenuates the transcriptional activity of the core clock protein BMAL1 and affects the maximal Per1 mRNA level of the circadian clock in cells. FEBS J 2018, 285, (16), 2987-3001. [CrossRef]
- Jia, J. Y.; Bissa, B.; Brecht, L.; Allers, L.; Choi, S. W.; Gu, Y. X.; Zbinden, M.; Burge, M. R.; Timmins, G.; Hallows, K.; Behrends, C.; Deretic, V., AMPK is activated during lysosomal damage via a galectin-ubiquitin signal transduction system. Autophagy 2020, 16, (8), 1550-1552. [CrossRef]
- Hansen, F. M.; Tanzer, M. C.; Bruning, F.; Bludau, I.; Stafford, C.; Schulman, B. A.; Robles, M. S.; Karayel, O.; Mann, M., Data-independent acquisition method for ubiquitinome analysis reveals regulation of circadian biology. Nat Commun 2021, 12, (1), 254. [CrossRef]
- Murray, S. S.; Wong, A. W.; Yang, J.; Li, Y.; Putz, U.; Tan, S. S.; Howitt, J., Ubiquitin Regulation of Trk Receptor Trafficking and Degradation. Mol Neurobiol 2019, 56, (3), 1628-1636. [CrossRef]
- Murakami, T.; Felinski, E. A.; Antonetti, D. A., Occludin phosphorylation and ubiquitination regulate tight junction trafficking and vascular endothelial growth factor-induced permeability. J Biol Chem 2009, 284, (31), 21036-46. [CrossRef]
- Teh, C. E.; Lalaoui, N.; Jain, R.; Policheni, A. N.; Heinlein, M.; Alvarez-Diaz, S.; Sheridan, J. M.; Rieser, E.; Deuser, S.; Darding, M.; Koay, H. F.; Hu, Y.; Kupresanin, F.; O'Reilly, L. A.; Godfrey, D. I.; Smyth, G. K.; Bouillet, P.; Strasser, A.; Walczak, H.; Silke, J.; Gray, D. H., Linear ubiquitin chain assembly complex coordinates late thymic T-cell differentiation and regulatory T-cell homeostasis. Nat Commun 2016, 7, 13353. [CrossRef]
- Verheul, T. C. J.; Philipsen, S., A ubiquitin ligase toggles red cell differentiation. Blood 2021, 137, (2), 143-144. [CrossRef]
- Sun, H.; Zhang, Q.; Jing, Y. Y.; Zhang, M.; Wang, H. Y.; Cai, Z.; Liuyu, T.; Zhang, Z. D.; Xiong, T. C.; Wu, Y.; Zhu, Q. Y.; Yao, J.; Shu, H. B.; Lin, D.; Zhong, B., USP13 negatively regulates antiviral responses by deubiquitinating STING. Nat Commun 2017, 8, 15534. [CrossRef]
- Zhang, Z.; Fang, X.; Wu, X.; Ling, L.; Chu, F.; Li, J.; Wang, S.; Zang, J.; Zhang, B.; Ye, S.; Zhang, L.; Yang, B.; Lin, S.; Huang, H.; Wang, A.; Zhou, F., Acetylation-Dependent Deubiquitinase OTUD3 Controls MAVS Activation in Innate Antiviral Immunity. Mol Cell 2020, 79, (2), 304-319 e7. [CrossRef]
- Jiang, X.; Chen, Z. J., The role of ubiquitylation in immune defence and pathogen evasion. Nat Rev Immunol 2011, 12, (1), 35-48. [CrossRef]
- Mukhopadhyay, D.; Riezman, H., Proteasome-independent functions of ubiquitin in endocytosis and signaling. Science 2007, 315, (5809), 201-5. [CrossRef]
- Li, Y.; Xie, P.; Lu, L.; Wang, J.; Diao, L.; Liu, Z.; Guo, F.; He, Y.; Liu, Y.; Huang, Q.; Liang, H.; Li, D.; He, F., An integrated bioinformatics platform for investigating the human E3 ubiquitin ligase-substrate interaction network. Nat Commun 2017, 8, (1), 347. [CrossRef]
- Lv, Z.; Rickman, K. A.; Yuan, L.; Williams, K.; Selvam, S. P.; Woosley, A. N.; Howe, P. H.; Ogretmen, B.; Smogorzewska, A.; Olsen, S. K., S. pombe Uba1-Ubc15 Structure Reveals a Novel Regulatory Mechanism of Ubiquitin E2 Activity. Mol Cell 2017, 65, (4), 699-714 e6. [CrossRef]
- Hjerpe, R.; Bett, J. S.; Keuss, M. J.; Solovyova, A.; McWilliams, T. G.; Johnson, C.; Sahu, I.; Varghese, J.; Wood, N.; Wightman, M.; Osborne, G.; Bates, G. P.; Glickman, M. H.; Trost, M.; Knebel, A.; Marchesi, F.; Kurz, T., UBQLN2 Mediates Autophagy-Independent Protein Aggregate Clearance by the Proteasome. Cell 2016, 166, (4), 935-949. [CrossRef]
- Li, F.; Sun, Q.; Liu, K.; Zhang, L.; Lin, N.; You, K.; Liu, M.; Kon, N.; Tian, F.; Mao, Z.; Li, T.; Tong, T.; Qin, J.; Gu, W.; Li, D.; Zhao, W., OTUD5 cooperates with TRIM25 in transcriptional regulation and tumor progression via deubiquitination activity. Nat Commun 2020, 11, (1), 4184. [CrossRef]
- Wertz, I. E.; Newton, K.; Seshasayee, D.; Kusam, S.; Lam, C.; Zhang, J.; Popovych, N.; Helgason, E.; Schoeffler, A.; Jeet, S.; Ramamoorthi, N.; Kategaya, L.; Newman, R. J.; Horikawa, K.; Dugger, D.; Sandoval, W.; Mukund, S.; Zindal, A.; Martin, F.; Quan, C.; Tom, J.; Fairbrother, W. J.; Townsend, M.; Warming, S.; DeVoss, J.; Liu, J.; Dueber, E.; Caplazi, P.; Lee, W. P.; Goodnow, C. C.; Balazs, M.; Yu, K.; Kolumam, G.; Dixit, V. M., Phosphorylation and linear ubiquitin direct A20 inhibition of inflammation. Nature 2015, 528, (7582), 370-5. [CrossRef]
- Wertz, I. E.; O'Rourke, K. M.; Zhou, H.; Eby, M.; Aravind, L.; Seshagiri, S.; Wu, P.; Wiesmann, C.; Baker, R.; Boone, D. L.; Ma, A.; Koonin, E. V.; Dixit, V. M., De-ubiquitination and ubiquitin ligase domains of A20 downregulate NF-kappaB signalling. Nature 2004, 430, (7000), 694-9. [CrossRef]
- Pickart, C. M.; Eddins, M. J., Ubiquitin: structures, functions, mechanisms. Biochim Biophys Acta 2004, 1695, (1-3), 55-72. [CrossRef]
- Myeku, N.; Clelland, C. L.; Emrani, S.; Kukushkin, N. V.; Yu, W. H.; Goldberg, A. L.; Duff, K. E., Tau-driven 26S proteasome impairment and cognitive dysfunction can be prevented early in disease by activating cAMP-PKA signaling. Nat Med 2016, 22, (1), 46-53. [CrossRef]
- Collins, G. A.; Goldberg, A. L., The Logic of the 26S Proteasome. Cell 2017, 169, (5), 792-806. [CrossRef]
- Bhoj, V. G.; Chen, Z. J., Ubiquitylation in innate and adaptive immunity. Nature 2009, 458, (7237), 430-7. [CrossRef]
- 27. Yoshida, Y.; Saeki, Y.; Murakami, A.; Kawawaki, J.; Tsuchiya, H.; Yoshihara, H.; Shindo, M.; Tanaka, K., A comprehensive method for detecting ubiquitinated substrates using TR-TUBE. Proc Natl Acad Sci U S A 2015, 112, (15), 4630-5. [CrossRef]
- Cheng, J.; Guo, J.; North, B. J.; Wang, B.; Cui, C. P.; Li, H.; Tao, K.; Zhang, L.; Wei, W., Functional analysis of deubiquitylating enzymes in tumorigenesis and development. Biochim Biophys Acta Rev Cancer 2019, 1872, (2), 188312. [CrossRef]
- Mevissen, T. E.; Hospenthal, M. K.; Geurink, P. P.; Elliott, P. R.; Akutsu, M.; Arnaudo, N.; Ekkebus, R.; Kulathu, Y.; Wauer, T.; El Oualid, F.; Freund, S. M.; Ovaa, H.; Komander, D., OTU deubiquitinases reveal mechanisms of linkage specificity and enable ubiquitin chain restriction analysis. Cell 2013, 154, (1), 169-84. [CrossRef]
- Yao, F.; Zhou, Z.; Kim, J.; Hang, Q.; Xiao, Z.; Ton, B. N.; Chang, L.; Liu, N.; Zeng, L.; Wang, W.; Wang, Y.; Zhang, P.; Hu, X.; Su, X.; Liang, H.; Sun, Y.; Ma, L., SKP2- and OTUD1-regulated non-proteolytic ubiquitination of YAP promotes YAP nuclear localization and activity. Nat Commun 2018, 9, (1), 2269. [CrossRef]
- Zhang, Z.; Wang, D.; Wang, P.; Zhao, Y.; You, F., OTUD1 Negatively Regulates Type I IFN Induction by Disrupting Noncanonical Ubiquitination of IRF3. J Immunol 2020, 204, (7), 1904-1918. [CrossRef]
- Wiener, R.; DiBello, A. T.; Lombardi, P. M.; Guzzo, C. M.; Zhang, X.; Matunis, M. J.; Wolberger, C., E2 ubiquitin-conjugating enzymes regulate the deubiquitinating activity of OTUB1. Nat Struct Mol Biol 2013, 20, (9), 1033-9. [CrossRef]
- Wiener, R.; Zhang, X.; Wang, T.; Wolberger, C., The mechanism of OTUB1-mediated inhibition of ubiquitination. Nature 2012, 483, (7391), 618-22. [CrossRef]
- Nakada, S.; Tai, I.; Panier, S.; Al-Hakim, A.; Iemura, S.; Juang, Y. C.; O'Donnell, L.; Kumakubo, A.; Munro, M.; Sicheri, F.; Gingras, A. C.; Natsume, T.; Suda, T.; Durocher, D., Non-canonical inhibition of DNA damage-dependent ubiquitination by OTUB1. Nature 2010, 466, (7309), 941-6. [CrossRef]
- Shi, L.; Liu, J.; Peng, Y.; Zhang, J.; Dai, X.; Zhang, S.; Wang, Y.; Liu, J.; Long, J., Deubiquitinase OTUD6A promotes proliferation of cancer cells via regulating Drp1 stability and mitochondrial fission. Mol Oncol 2020, 14, (12), 3169-3183. [CrossRef]
- Zhao, Y.; Huang, X.; Zhu, D.; Wei, M.; Luo, J.; Yu, S.; Tian, Y.; Zheng, X., Deubiquitinase OTUD6A promotes breast cancer progression by increasing TopBP1 stability and rendering tumor cells resistant to DNA-damaging therapy. Cell Death Differ 2022, 29, (12), 2531-2544. [CrossRef]
- Fu, X.; Zhao, J.; Yu, G.; Zhang, X.; Sun, J.; Li, L.; Yin, J.; Niu, Y.; Ren, S.; Zhu, Y.; Xu, B.; Huang, L., OTUD6A promotes prostate tumorigenesis via deubiquitinating Brg1 and AR. Commun Biol 2022, 5, (1), 182. [CrossRef]
- Peng, Y.; Liu, J.; Wang, Z.; Cui, C.; Zhang, T.; Zhang, S.; Gao, P.; Hou, Z.; Liu, H.; Guo, J.; Zhang, J.; Wen, Y.; Wei, W.; Zhang, L.; Liu, J.; Long, J., Prostate-specific oncogene OTUD6A promotes prostatic tumorigenesis via deubiquitinating and stabilizing c-Myc. Cell Death Differ 2022, 29, (9), 1730-1743. [CrossRef]
- Park, S. H.; Jung, E. H.; Kim, G. Y.; Kim, B. C.; Lim, J. H.; Woo, C. H., Itch E3 ubiquitin ligase positively regulates TGF-beta signaling to EMT via Smad7 ubiquitination. Mol Cells 2015, 38, (1), 20-5. [CrossRef]
- Smith, K.; Gunaratnam, L.; Morley, M.; Franovic, A.; Mekhail, K.; Lee, S., Silencing of epidermal growth factor receptor suppresses hypoxia-inducible factor-2-driven VHL-/- renal cancer. Cancer Res 2005, 65, (12), 5221-30. [CrossRef]
- You, F.; Wang, P.; Yang, L.; Yang, G.; Zhao, Y. O.; Qian, F.; Walker, W.; Sutton, R.; Montgomery, R.; Lin, R.; Iwasaki, A.; Fikrig, E., ELF4 is critical for induction of type I interferon and the host antiviral response. Nat Immunol 2013, 14, (12), 1237-46. [CrossRef]
- Liu, X.; Fang, Y.; Lv, X.; Hu, C.; Chen, G.; Zhang, L.; Jin, B.; Huang, L.; Luo, W.; Liang, G.; Wang, Y., Deubiquitinase OTUD6A in macrophages promotes intestinal inflammation and colitis via deubiquitination of NLRP3. Cell Death Differ 2023. [CrossRef]
- Chang, J. H.; Xiao, Y.; Hu, H.; Jin, J.; Yu, J.; Zhou, X.; Wu, X.; Johnson, H. M.; Akira, S.; Pasparakis, M.; Cheng, X.; Sun, S. C., Ubc13 maintains the suppressive function of regulatory T cells and prevents their conversion into effector-like T cells. Nat Immunol 2012, 13, (5), 481-90. [CrossRef]
- Ni, J.; Guan, C.; Liu, H.; Huang, X.; Yue, J.; Xiang, H.; Jiang, Z.; Tao, Y.; Cao, W.; Liu, J.; Wang, Z.; Wang, Y.; Wu, X., Ubc13 Promotes K63-Linked Polyubiquitination of NLRP3 to Activate Inflammasome. J Immunol 2021, 206, (10), 2376-2385. [CrossRef]
- Hu, L.; Xu, J.; Xie, X.; Zhou, Y.; Tao, P.; Li, H.; Han, X.; Wang, C.; Liu, J.; Xu, P.; Neculai, D.; Xia, Z., Oligomerization-primed coiled-coil domain interaction with Ubc13 confers processivity to TRAF6 ubiquitin ligase activity. Nat Commun 2017, 8, (1), 814. [CrossRef]
- McNab, F.; Mayer-Barber, K.; Sher, A.; Wack, A.; O'Garra, A., Type I interferons in infectious disease. Nat Rev Immunol 2015, 15, (2), 87-103. [CrossRef]
- Medzhitov, R., Origin and physiological roles of inflammation. Nature 2008, 454, (7203), 428-35. [CrossRef]
- Damgaard, R. B.; Walker, J. A.; Marco-Casanova, P.; Morgan, N. V.; Titheradge, H. L.; Elliott, P. R.; McHale, D.; Maher, E. R.; McKenzie, A. N. J.; Komander, D., The Deubiquitinase OTULIN Is an Essential Negative Regulator of Inflammation and Autoimmunity. Cell 2016, 166, (5), 1215-1230 e20. [CrossRef]
- Panda, S.; Nilsson, J. A.; Gekara, N. O., Deubiquitinase MYSM1 Regulates Innate Immunity through Inactivation of TRAF3 and TRAF6 Complexes. Immunity 2015, 43, (4), 647-59. [CrossRef]
- Wang, X. M.; Yang, C.; Zhao, Y.; Xu, Z. G.; Yang, W.; Wang, P.; Lin, D.; Xiong, B.; Fang, J. Y.; Dong, C.; Zhong, B., The deubiquitinase USP25 supports colonic inflammation and bacterial infection and promotes colorectal cancer. Nat Cancer 2020, 1, (8), 811-825. [CrossRef]
- Li, S.; Zheng, H.; Mao, A. P.; Zhong, B.; Li, Y.; Liu, Y.; Gao, Y.; Ran, Y.; Tien, P.; Shu, H. B., Regulation of virus-triggered signaling by OTUB1- and OTUB2-mediated deubiquitination of TRAF3 and TRAF6. J Biol Chem 2010, 285, (7), 4291-7. [CrossRef]
- Yamamoto, M.; Okamoto, T.; Takeda, K.; Sato, S.; Sanjo, H.; Uematsu, S.; Saitoh, T.; Yamamoto, N.; Sakurai, H.; Ishii, K. J.; Yamaoka, S.; Kawai, T.; Matsuura, Y.; Takeuchi, O.; Akira, S., Key function for the Ubc13 E2 ubiquitin-conjugating enzyme in immune receptor signaling. Nat Immunol 2006, 7, (9), 962-70. [CrossRef]
- Wu, B.; Qiang, L.; Zhang, Y.; Fu, Y.; Zhao, M.; Lei, Z.; Lu, Z.; Wei, Y. G.; Dai, H.; Ge, Y.; Liu, M.; Zhou, X.; Peng, Z.; Li, H.; Cui, C. P.; Wang, J.; Zheng, H.; Liu, C. H.; Zhang, L., The deubiquitinase OTUD1 inhibits colonic inflammation by suppressing RIPK1-mediated NF-kappaB signaling. Cell Mol Immunol 2022, 19, (2), 276-289. [CrossRef]
Figure 1.
Overexpression of OTUD6A restricts innate immune signaling. (a) HEK293 cells were seeded onto 24-well cell culture plates and transfected with 50 ng of Ifnβ-Luci and 20 ng of Actin-Luci reporter gene plasmids, along with 100 ng of Flag-cGAS and 100 ng of Flag-STING expression plasmids in each well. After 12 hours, the cells were co-transfected with 42 DUBs expression plasmids. Twenty-four hours later, the activation of the reporter gene was measured using a single-well microplate reader. (b) HEK293 cells were transfected with IFNβ-Luci and Actin-Luci reporter gene plasmids, along with Flag-OTUD6A expression plasmids. The cells were then co-transfected with STING, MAVS, TBK1, IRF3, TRIF, and RIG-1 expression plasmids, respectively. The control group was transfected with an empty vector plasmid. After 24 hours, the biological activity of dual luciferase was assessed using a single-well microplate reader. (c) HEK293 cells were transfected with Flag-OTUD6A expression plasmid or an empty vector plasmid, and then infected with SeV, VSV, and HSV-1 viruses after 24 hours. After 24 hours of infection, the biological activity of dual luciferase was measured using a single-well microplate reader.
Figure 1.
Overexpression of OTUD6A restricts innate immune signaling. (a) HEK293 cells were seeded onto 24-well cell culture plates and transfected with 50 ng of Ifnβ-Luci and 20 ng of Actin-Luci reporter gene plasmids, along with 100 ng of Flag-cGAS and 100 ng of Flag-STING expression plasmids in each well. After 12 hours, the cells were co-transfected with 42 DUBs expression plasmids. Twenty-four hours later, the activation of the reporter gene was measured using a single-well microplate reader. (b) HEK293 cells were transfected with IFNβ-Luci and Actin-Luci reporter gene plasmids, along with Flag-OTUD6A expression plasmids. The cells were then co-transfected with STING, MAVS, TBK1, IRF3, TRIF, and RIG-1 expression plasmids, respectively. The control group was transfected with an empty vector plasmid. After 24 hours, the biological activity of dual luciferase was assessed using a single-well microplate reader. (c) HEK293 cells were transfected with Flag-OTUD6A expression plasmid or an empty vector plasmid, and then infected with SeV, VSV, and HSV-1 viruses after 24 hours. After 24 hours of infection, the biological activity of dual luciferase was measured using a single-well microplate reader.
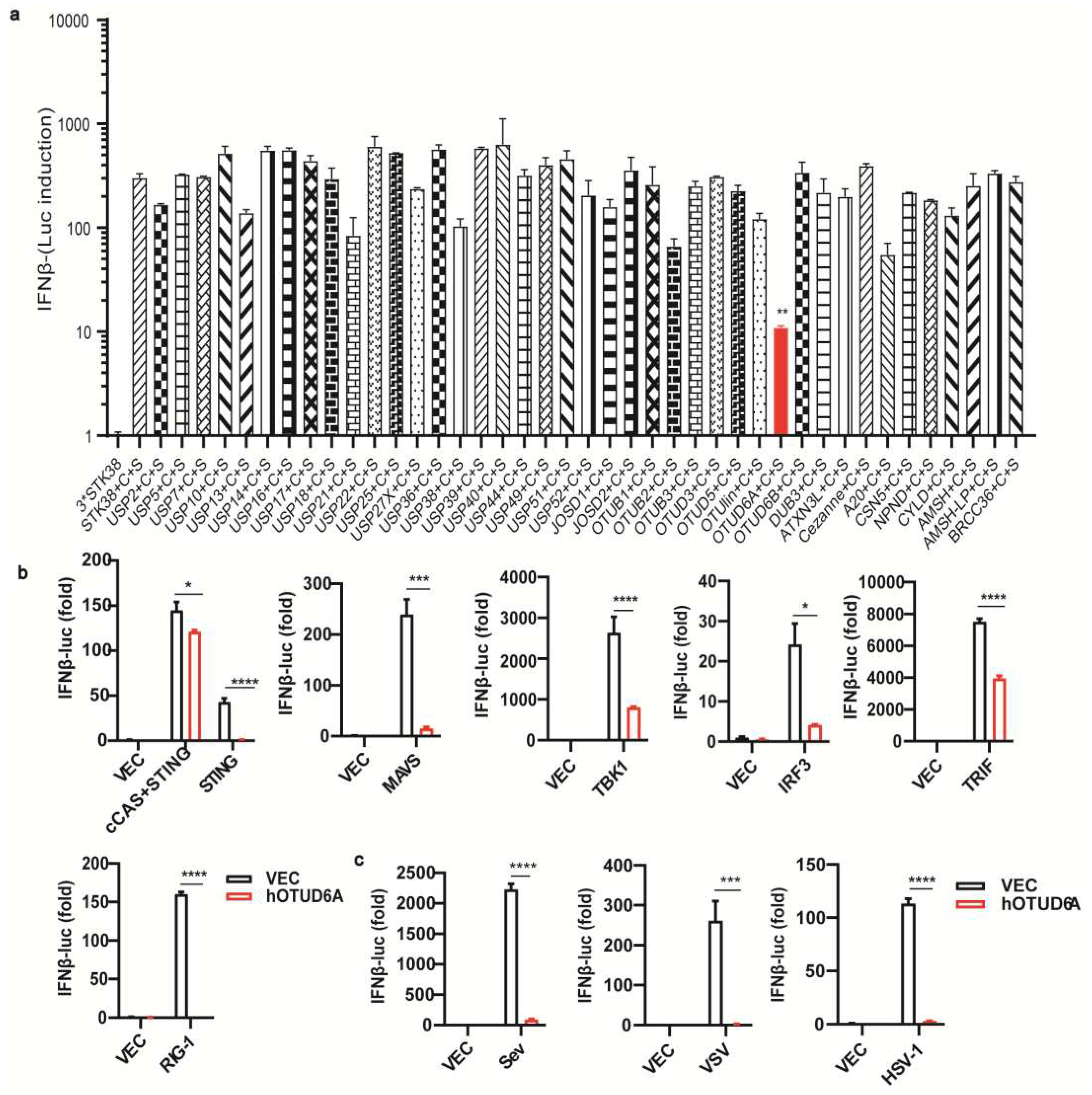
Figure 2.
OTUD6A deficiency enhances antiviral innate immunity in Vitro. (a-b) The mRNA expression levels of Ifna1, Ifna2, Ifna4, and Ifnβ were measured by RT-qPCR in Otud6a-/- mouse-derived peritoneal macrophages (PMs) and bone marrow-derived macrophages (BMDMs) infected with Sev (0.1 MOI) for 8 hours, compared to wild-type (WT) cells (c) RNA extracted from Otud6a-/- and wild-type iBMDM cells, 6 hours after HSV-1 infection, was used for whole transcriptome sequencing. Differentially expressed genes were identified and subjected to gene ontology (GO) analysis. (d) The heatmap was generated to represent the normalized expression levels of viral infection-induced genes, as indicated.
Figure 2.
OTUD6A deficiency enhances antiviral innate immunity in Vitro. (a-b) The mRNA expression levels of Ifna1, Ifna2, Ifna4, and Ifnβ were measured by RT-qPCR in Otud6a-/- mouse-derived peritoneal macrophages (PMs) and bone marrow-derived macrophages (BMDMs) infected with Sev (0.1 MOI) for 8 hours, compared to wild-type (WT) cells (c) RNA extracted from Otud6a-/- and wild-type iBMDM cells, 6 hours after HSV-1 infection, was used for whole transcriptome sequencing. Differentially expressed genes were identified and subjected to gene ontology (GO) analysis. (d) The heatmap was generated to represent the normalized expression levels of viral infection-induced genes, as indicated.

Figure 3.
OTUD6A deficiency enhances antiviral innate immunity in Vivo. (a) Wild-type (WT) mice and Otud6a-/- mice (n=6) were subjected to tail vein injection of HSV-1 (5 x 107 PFU per mouse), and the mice were monitored for survival. (b) On the fifth day after HSV-1 virus infection, brain tissue was extracted from the mice to measure virus titers using plaque assays. (c) One day after HSV-1 infection, total mRNA was extracted from the blood, and the mRNA levels of Ifnβ, Ifit1, Ifit2 and Cxcl10 were measured using RT-qPCR. (d) WT mice and Otud6a-/- mice (n=6) were tail vein injected with VSV (1x108 PFU per mouse), and mouse survival was monitored. (e) On the fifth day after VSV virus infection, brain tissue was extracted from the mice to determine virus titers using plaque assays. (f) One day after VSV infection, total mRNA was extracted from the blood, and the mRNA levels of Ifna2, Ifna4, Ifna5, and Ifnβ were measured using RT-qPCR.
Figure 3.
OTUD6A deficiency enhances antiviral innate immunity in Vivo. (a) Wild-type (WT) mice and Otud6a-/- mice (n=6) were subjected to tail vein injection of HSV-1 (5 x 107 PFU per mouse), and the mice were monitored for survival. (b) On the fifth day after HSV-1 virus infection, brain tissue was extracted from the mice to measure virus titers using plaque assays. (c) One day after HSV-1 infection, total mRNA was extracted from the blood, and the mRNA levels of Ifnβ, Ifit1, Ifit2 and Cxcl10 were measured using RT-qPCR. (d) WT mice and Otud6a-/- mice (n=6) were tail vein injected with VSV (1x108 PFU per mouse), and mouse survival was monitored. (e) On the fifth day after VSV virus infection, brain tissue was extracted from the mice to determine virus titers using plaque assays. (f) One day after VSV infection, total mRNA was extracted from the blood, and the mRNA levels of Ifna2, Ifna4, Ifna5, and Ifnβ were measured using RT-qPCR.
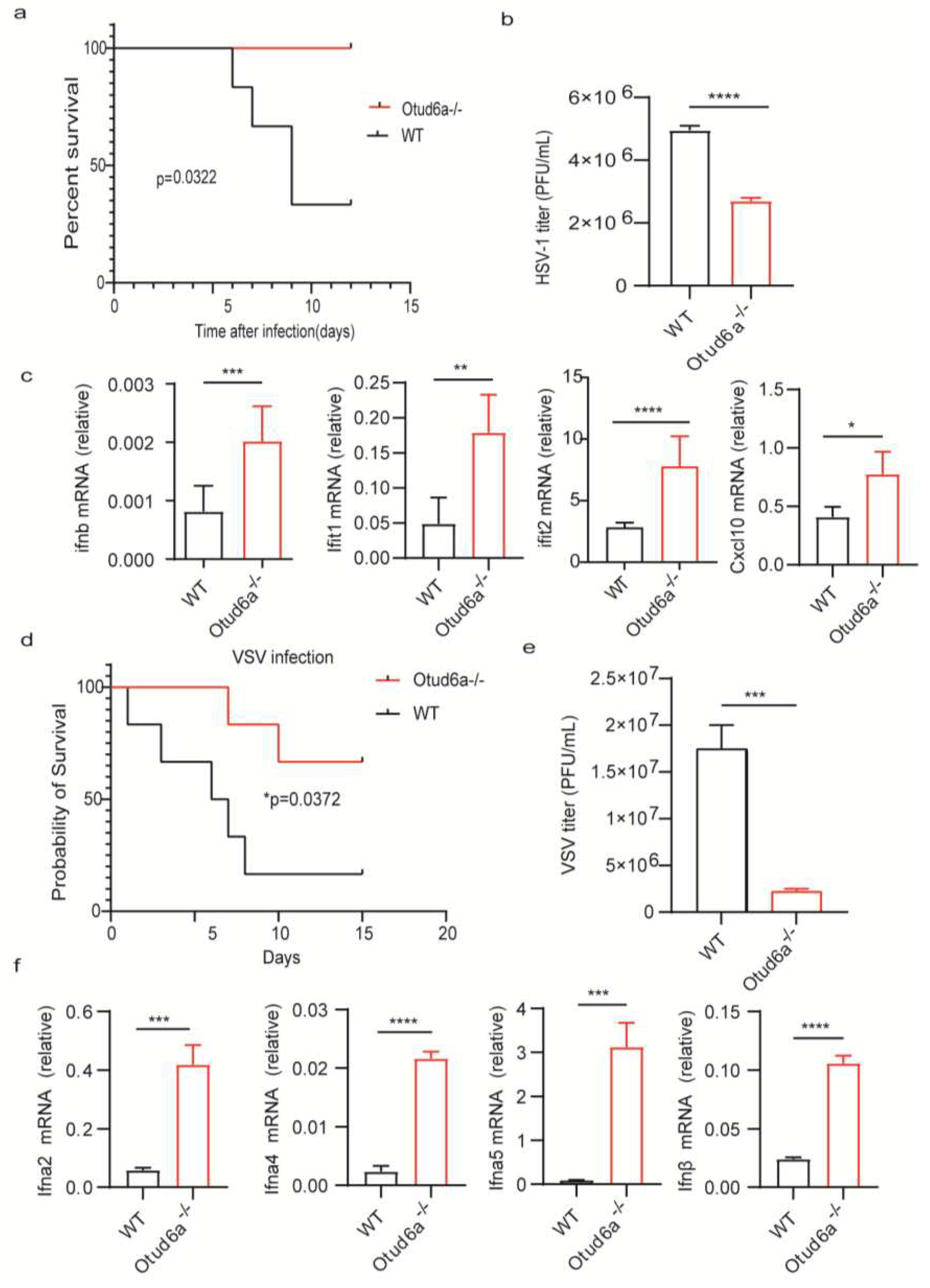
Figure 4.
OTUD6A deficiency attenuates the inflammatory response. (a-b) Peritoneal macrophages and bone marrow-derived macrophages were isolated from wild-type and Otud6a-/- mice and stimulated with 1 μg/mL LPS for 6 hours. Total mRNA was extracted, and the mRNA levels of Il6, Tnf-α, and Il1b were measured using RT-qPCR. (c-d) Wild-type and Otud6a-/- mice were intraperitoneally injected with LPS (8.74 μg/g). The mice were observed daily for mortality and changes in body weight in each group. (e) RT-qPCR analysis was performed to measure the mRNA levels of Ifnβ and Isg15 in the serum (n=6) obtained from wild-type or Otud6a-/- mice intraperitoneally injected with LPS for 3 and 5 days.
Figure 4.
OTUD6A deficiency attenuates the inflammatory response. (a-b) Peritoneal macrophages and bone marrow-derived macrophages were isolated from wild-type and Otud6a-/- mice and stimulated with 1 μg/mL LPS for 6 hours. Total mRNA was extracted, and the mRNA levels of Il6, Tnf-α, and Il1b were measured using RT-qPCR. (c-d) Wild-type and Otud6a-/- mice were intraperitoneally injected with LPS (8.74 μg/g). The mice were observed daily for mortality and changes in body weight in each group. (e) RT-qPCR analysis was performed to measure the mRNA levels of Ifnβ and Isg15 in the serum (n=6) obtained from wild-type or Otud6a-/- mice intraperitoneally injected with LPS for 3 and 5 days.
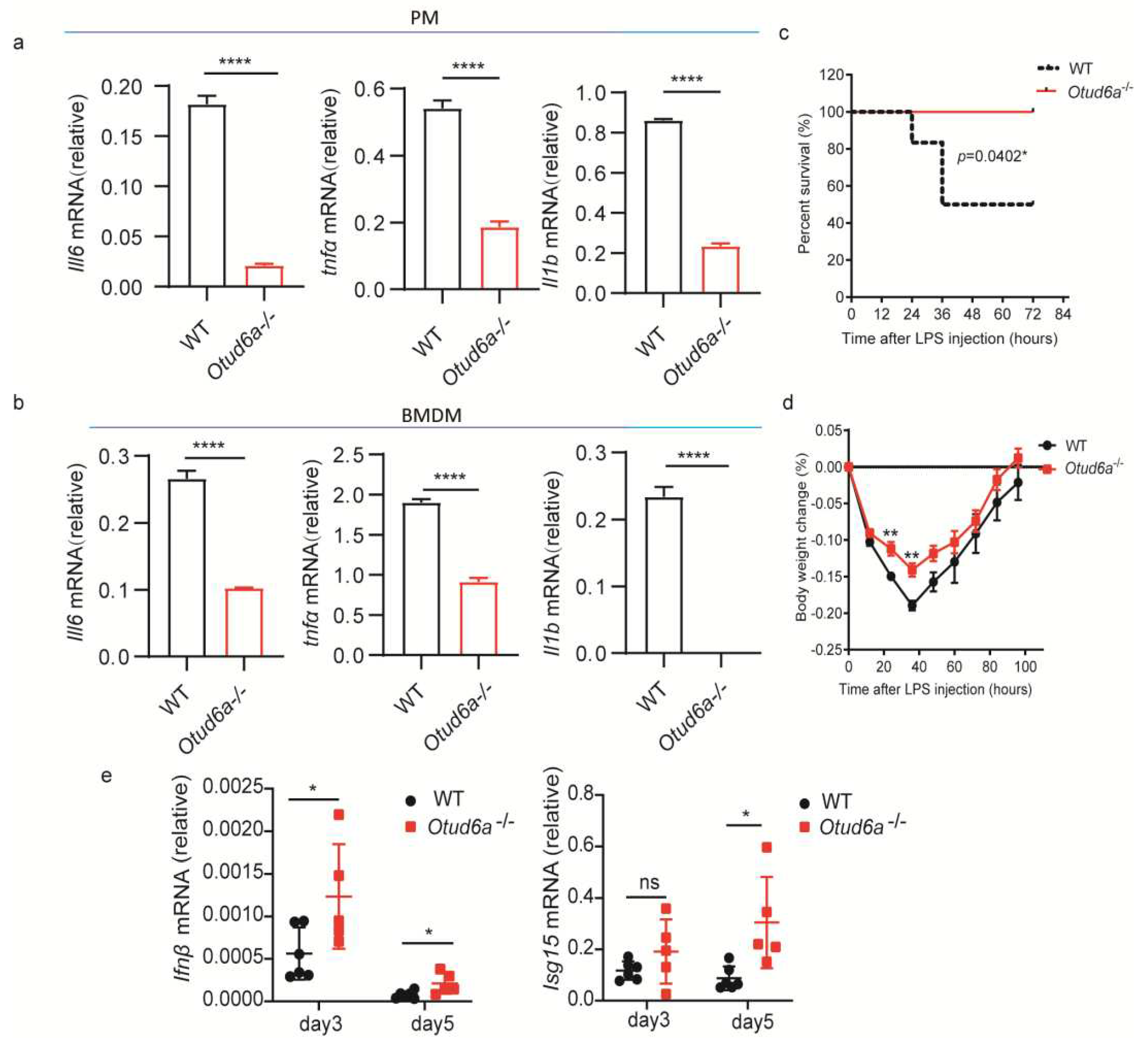
Figure 5.
OTUD6A participates in the regulation of NF-κB mediated inflammation signaling pathway via Ubc13. (a) Macrophages derived from the bone marrow of wild-type and Otud6a-/- mice were induced and stimulated with 1 μg/mL LPS for 30 minutes and 120 minutes. Total proteins were extracted for an immunoblotting experiment to detect the protein levels of endogenous p65, p-p65, IκBα, and p-IκBα. (b) FLAG-M2 beads were used to enrich proteins in HEK293 and Hela cells, respectively. Ubc13 was identified as a potential interaction protein for OTUD6A through mass spectrometry. (c) Flag-Ubc13 and HA-OTUD6A were simultaneously overexpressed in HEK293 cells. After 24 hours, the cells were infected with HSV-1. Co-immunoprecipitation experiments were performed using Flag antibody and IgG antibody after 6 hours, respectively. (d) The cysteine residue at position 87 of Ubc13 was mutated to alanine to obtain the Flag-Ubc13-C87A mutant plasmid. HA-OTUD6A and Flag-Ubc13 or Flag-Ubc13 C87A plasmids were overexpressed in HEK293 cells for 24 hours, followed by HSV infection. Co-immunoprecipitation experiments were performed using Flag antibody and IgG antibody after 6 hours, respectively. (e) The pattern of Ubc13 point mutations and OTUD6A truncation mutations is shown. (f) OTUD6A with 50 or 100 amino acids missing from the N-terminal (HA-OTUD6A d1-50, HA-OTUD6A d1-100) and full-length OTUD6A (HA-OTUD6A fl) were transfected into HEK293 cells with Ubc13 for 24 hours. The cells were then infected with HSV for 6 hours before co-immunoprecipitation (with anti-FLAG) and immunoblotting analysis using the indicated antibodies.
Figure 5.
OTUD6A participates in the regulation of NF-κB mediated inflammation signaling pathway via Ubc13. (a) Macrophages derived from the bone marrow of wild-type and Otud6a-/- mice were induced and stimulated with 1 μg/mL LPS for 30 minutes and 120 minutes. Total proteins were extracted for an immunoblotting experiment to detect the protein levels of endogenous p65, p-p65, IκBα, and p-IκBα. (b) FLAG-M2 beads were used to enrich proteins in HEK293 and Hela cells, respectively. Ubc13 was identified as a potential interaction protein for OTUD6A through mass spectrometry. (c) Flag-Ubc13 and HA-OTUD6A were simultaneously overexpressed in HEK293 cells. After 24 hours, the cells were infected with HSV-1. Co-immunoprecipitation experiments were performed using Flag antibody and IgG antibody after 6 hours, respectively. (d) The cysteine residue at position 87 of Ubc13 was mutated to alanine to obtain the Flag-Ubc13-C87A mutant plasmid. HA-OTUD6A and Flag-Ubc13 or Flag-Ubc13 C87A plasmids were overexpressed in HEK293 cells for 24 hours, followed by HSV infection. Co-immunoprecipitation experiments were performed using Flag antibody and IgG antibody after 6 hours, respectively. (e) The pattern of Ubc13 point mutations and OTUD6A truncation mutations is shown. (f) OTUD6A with 50 or 100 amino acids missing from the N-terminal (HA-OTUD6A d1-50, HA-OTUD6A d1-100) and full-length OTUD6A (HA-OTUD6A fl) were transfected into HEK293 cells with Ubc13 for 24 hours. The cells were then infected with HSV for 6 hours before co-immunoprecipitation (with anti-FLAG) and immunoblotting analysis using the indicated antibodies.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



