Submitted:
23 June 2023
Posted:
27 June 2023
You are already at the latest version
Abstract
(1) Background: Formation of biofilms on food-contact surfaces by Shiga-toxigenic Escherichia coli (STEC) can pose a significant challenge to the food industry, making conventional control-methods insufficient. Targeted use of bacteriophages to disrupt these biofilms could reduce this problem. Previously isolated and characterized bacteriophages (n=52) were evaluated against STEC biofilms in-vitro and on food-contact surfaces. (2) Methods: Phage-treatments (9 logs PFU/ml), in phosphate-buffered-saline, were used individually or as cocktails. Biofilms of STEC (O157, O26, O45, O103, O111, O121, and O145) were formed in 96-well micro-titer plates (7 logs CFU/ml; 24 h) or on stainless-steel (SS) and high-density-polyethylene (HDPE) coupons (9 logs CFU/cm2; 7 h), followed by phage-treatment. Biofilm-disruption was measured in-vitro at 0, 3, and 6 h as a change in optical-density (A595). Coupons were treated with STEC-serotype-specific phage-cocktails or a 21-phage cocktail (3 phages/serotype) for 0, 3, 6 and 16 h and surviving STEC populations enumerated. (3) Results: Of the 52 pages, 77% showed STEC-biofilm disruption in-vitro. Serotype-specific phage-treatments reduced pathogen population within the biofilms by 1.9-4.1 and 2.3-5.6 logs CFU/cm2, while the 21-pahge cocktail reduced it by 4.0 and 4.8 logs CFU/cm2 on SS and HDPE, respectively. (4) Conclusions: Bacteriophages can be used to reduce STEC and their biofilms.
Keywords:
Bacteriophages
; Shiga-toxigenic E. coli
; biocontrol
; biofilms
; food-contact surfaces
1. Introduction
Shiga-toxigenic Escherichia coli (STEC) are human pathogens responsible for multiple foodborne disease outbreaks. Their infections can range from mild to severe bloody diarrhea and hemorrhagic colitis (HC), leading to life-threatening complications, such as hemolytic-uremic syndrome (HUS) and thrombotic thrombocytopenic purpura [1]. While the STEC serotype, O157:H7, is frequently associated with HC and HUS, other non-O157 serotypes, reported to cause similar infections, have also emerged. These include the top 6 serotypes, O26, O45, O103, O111, O121, and O145, that have been responsible for multiple outbreaks and infections in the last two decades [2,3,4]. According to the Centers for Disease Control and Prevention (CDC) estimates, STEC O157 and non-O157 together cause more than 265,000 illnesses each year in the United States [5]. These infections could be due to the consumption of contaminated fresh produce, undercooked meat, unpasteurized milk, or drinking water [6]. Other factors include, person to person contact, or livestock handling and contact with their environment on the farm, livestock events or through petting zoos [7].
These pathogens can also adapt to adverse environmental conditions by forming biofilms on a wide variety of food-contact surfaces, as well as on fresh produce and meat products [8,9,10]. Biofilms consist of a network of adhesive carbohydrates, exopolysaccharides (EPS), that are difficult to penetrate and can protect the bacteria from stressful conditions and antimicrobials [11,12]. It has been reported that STEC biofilms have an increased tolerance to common sanitizers such as chlorine and quaternary ammonium compounds [10,13,14,15]. This could be due to a combination of bacterial resistance mechanisms such as, diffusional resistance of the EPS matrix, chemical and enzymatic inactivation of sanitizers and disinfectants, physiological changes in the cell, and induction of stress responses in the cell [16]. The ability of these pathogens to form strong biofilms poses a significant threat of cross-contamination to the food-processing industry. It is therefore critical to develop effective strategies to prevent, remove or control biofilms in the food industry for improved food safety.
Bacteriophages are viruses that infect and kill bacteria and have garnered significant attention as antibacterial agents, primarily due to their target-specificity towards the host bacteria [17,18,19]. They are present as commensal microflora in the gastrointestinal tract of animals and have also been isolated from various food and water sources [20,21]. The host-specificity and ubiquitous nature of lytic bacteriophages makes them highly desirable as antibacterial agents [18]. Under in-vitro conditions, virulent bacteriophages have shown the potential for selective elimination of foodborne pathogens [22,23]. Attention has also been given to the use of bacteriophages for removal of biofilms formed by foodborne pathogens on various food-contact surfaces [24,25,26,27]. Viazis et al. [27] tested a phage cocktail against E. coli O157:H7 biofilms on stainless steel (SS), ceramic tile, and high-density polyethylene (HDPE) coupons. The phage cocktail effectively reduced pathogen populations in biofilms on all three surfaces, within an hour of the treatment. In another study, phage treatment on spinach-harvesting blade reduced E. coli O157:H7 in biofilms by 4.5 logs CFU/blade [25]. The majority of these studies have been limited to evaluating bacteriophage potential against E. coli O157:H7 and its biofilms. Very little is known about the effectiveness and applicability of phages against non-O157 STEC and their biofilms. The objective of the present study was to evaluate the efficacy of previously isolated and characterized bacteriophages [28,29] against O157 and non-O157 STEC biofilms in-vitro and on food-contact surfaces.
2. Materials and Methods
2.1. Bacterial cultures
The STEC isolates used in the study included, E. coli O157:H7 (ATCC 43895, wild type (WT): LF4, KF10); O26 (CDC 2003-3014, WT: QF6, BF8); O45 (CDC 2000-3039, WT: EF2, AF1); O103 (CDC 2006-3008, WT: GF6, AF10); O111 (CDC 2010C-3114, ATCC: 2440, 2180), O121 (CDC 2002-3211, ATCC: 2219, 2203); and O145 (CDC 99-3311, ATCC: 2208, 1652). The WT isolates were retrieved from the Jaroni laboratory culture collection, originally isolated from bovine feces or cattle farm environment [30]. Biofilm forming capability of these isolates was previously tested in-vitro [31].
For in-vitro studies, overnight cultures of STEC isolates were prepared in Luria Broth (LB; MP Biomedicals, CA) and incubated while shaking (180 rpm; V.390W, Fisher Scientific, NJ) at 37°C for 16 h. For phage propagation and food-contact surface studies, overnight cultures were prepared in tryptic soy broth (TSB; Bacto™, Difco, BD, Sparks, MD) and incubated statically at 37°C for 18 h. Bacterial cocktail suspensions were prepared by mixing (1:1) isolates of STEC serotypes, at a population of 109 CFU/ml (Table 1,Table 2,Table 3).
2.2. Phage preparation
Phages were propagated by suspending 100 µl overnight culture of host bacterium in molten (0.75%) NZCYM agar (RPI Corp, IL; Fisher Scientific, NJ) and plating via double layer agar method [28]. Phage working-stock solutions were prepared as previously described [28] and stored at 4°C, until further use. Prior to an experiment, phage titers were determined as plaque forming units per ml (PFU/ml), by serially diluting the phage working-stock in phosphate buffered saline (PBS: pH 7.4: NaCl, KCl, NaH2PO4 and KH2PO4, Sigma-Aldrich, MO, USA) and performing a plaque assay [28]. All phage treatments were prepared at a population of 109 PFU/ml.
2.3. STEC biofilm disruption
2.3.1. In vitro biofilm disruption
In-vitro STEC biofilm disruption was determined as previously described [32], in the following three experiments (I, II, III). In Experiment-I, previously isolated and characterized phages (n=52) were used individually to test their efficacy against their host STEC biofilms. Based on the results from Experiment-I, phages were selected to prepare host-specific cocktails (Table 1), which were then tested in Experiment-II. In Experiment-III, a multi-phage cocktail, containing 21-phages (3 phages per STEC serotype), was tested against biofilms of multi-serotype STEC cocktail (Table 2).
Overnight pathogen cultures were diluted (1:100) in M9 medium (MP Biomedicals, CA), supplemented with 0.4% (wt/vol) glucose (Fisher Scientific, NJ) and minerals (1.16 mM MgSO4, 2 µM FeCl3, 8 µM CaCl2, and 16 µM MnCl2; Fisher Scientific, NJ), and incubated with shaking (180 rpm) for 24 h at 37°C. Following incubation, bacterial cultures were further diluted (1:100) in M9 medium (containing glucose and minerals) and allowed to form biofilms in 96-well microtiter plates (Thermo Scientific, Denmark) by aliquoting 150 µl in each well (in triplicates) and incubating the plates at 37°C for 24 h. Wells filled with sterile M9 were used as the negative control. After incubation, the liquid culture was carefully removed using a micropipette and the wells washed three times with PBS (150 µl), without disturbing any biofilm formed at the bottom of the wells. Plates were dried at 37°C for 15 min and 150 µl of the respective bacteriophage treatment (individual or cocktail) or PBS (positive control) was added to the wells. Plates were incubated further at 37°C for 0, 3, and 6 h. After each incubation period, the treatment solution was removed and the wells washed with PBS, as described above. After drying at 37°C for 15 min, biofilms were stained by placing crystal violet (CV; Fisher Scientific, NJ) solution (0.1% in distilled water; wt/vol) in the wells for 2 min, washing thrice with PBS, and drying (37°C for 15 min). The stain was released with 150 µl of ethanol:acetone solution (80:20; vol/vol; Fisher Scientific, NJ) and biofilm disruption quantified by measuring the optical density (OD) at 595 nm (SpectroMax M3; Molecular Devices LLC., CA). Wells filled with ethanol:acetone solution were used as the blank.
2.3.2. STEC biofilm disruption on food-contact surfaces
The efficacy of bacteriophages to disrupt STEC biofilms on food-contact surfaces was studied in two experiments (I and II). In Experiment-I, a cocktail of each STEC serotype (described in 2.1) was used to form biofilms on stainless steel (SS) and high-density polyethylene (HDPE) coupons and treated with their respective phage cocktail for 16 h (Table 3). In Experiment-II, a 21-phage cocktail was evaluated against biofilms formed by a cocktail of 14 STEC isolates (2 isolates per serotype), on SS and HDPE coupons, for 0, 3, 6 and 16 h (Table 2).
2.3.2.1. Preparation of coupons
Stainless steel (304 finish, type 4; Stillwater Steels, OK) and HDPE (1/8”x 24”x 48”; Polymersan, Hialeah, FL) coupons (2 x 5 cm2) were used for the study and prepared as described by Hood and Zottola [33]. Coupons were cleaned by soaking in acetone (30 min), followed by distilled water rinse (5 min), soaking (1 h) in 1 N NaOH (Fisher Scientific, NJ) and sonicating (40KHz; Branson, CT) in distilled water (1 h). Following sonication, coupons were rinsed in distilled water, air-dried and sterilized prior to use.
2.3.2.2. Phage treatment of STEC biofilms
For each STEC serotype, 3 coupons were inoculated, where one coupon was designated as the inoculated, untreated control, one was treated with the respective phage cocktail, and one treated with PBS (control). In each experiment (I and II), coupons were first immersed in 30 ml suspension of bacterial cocktail (109 CFU/ml) in a 50 ml centrifuge tube (Fisher Scientific, GA) and incubated at 25°C for 2.5 h, to facilitate bacterial attachment to the surface. Coupons were gently removed from the suspension using sterile forceps and placed in a sterile 50 ml tube for 5 h in a biosafety hood, allowing further bacterial attachment and biofilm formation. After incubation, coupons were rinsed in 30 ml sterile distilled water to remove unattached bacterial cells from the surface. One coupon of each surface material (SS or HDPE) was sampled to determine the initial pathogen population within the biofilm (inoculated, untreated control). The remaining inoculated coupons were then suspended in 30 ml bacteriophage (109 PFU/ml) treatment (Table 3) or PBS at 37°C for 16 h in Experiment-I, and 0, 3, 6 and 16 h in Experiment-II. After incubation, coupons were sonicated for 5 min at 40 KHz to dislodge bacterial cells from the coupon surface. Immediately following, 3 g of glass beads (Genlantis, CA) were added to the tube and agitated for 1 min, using a vortex (Fisher Scientific, NJ), to remove any remaining attached cells from the coupon [26]. Surviving pathogen populations in the solution were enumerated (CFU/cm2) on TSA or STEC CHROM agar (CHROMagar, Paris, France). Any injured cells were recovered by 24 h enrichment in TSB at 37°C.
2.4. Statistical analysis
All the experiments were repeated 3 times. Surviving STEC populations, recovered after treatments, were converted to log10 CFU/cm2 and the mean values of the three replicates obtained. Data were analyzed using General Linear Model (SAS v.9.3 software; SAS Inst., Cary, NC, USA) to determine analysis of variance (ANOVA) for phage treatment effects. Significant differences between the treatment means were separated by the least significant difference (LSD) at P<0.10 for in-vitro studies and P<0.05 for food-contact surface studies.
3. Results and Discussion
3.1. STEC biofilm disruption
Effectiveness of individual and cocktails of phages to disrupt biofilms of 7 STEC serotypes, in-vitro and on food-contact surfaces (SS and HDPE), was determined. Results demonstrated that the phages were more effective in reducing biofilms on food-contact surfaces than in-vitro.
3.1.1. In vitro STEC biofilm disruption
Biofilm disruption by individual, serotype-specific bacteriophages, was determined in-vitro by measuring OD (A595) at 0, 3 and 6 h. STEC-biofilm disruption was observed with a reduction in OD from 2.262 nm (0 h) to 0.808 nm (6 h) in phage-treated wells. All phages (n=52) reduced biofilm formation by their respective host bacteria, with 77% showing significant (P<0.10) reductions at 3 and 6 h (Figure 1A-G).
Figure 1.
(A). In-vitro biofilm disruption by individual bacteriophages specific to E. coli O157:H7 after treatment for 0, 3, and 6 h.
Figure 1.
(A). In-vitro biofilm disruption by individual bacteriophages specific to E. coli O157:H7 after treatment for 0, 3, and 6 h.
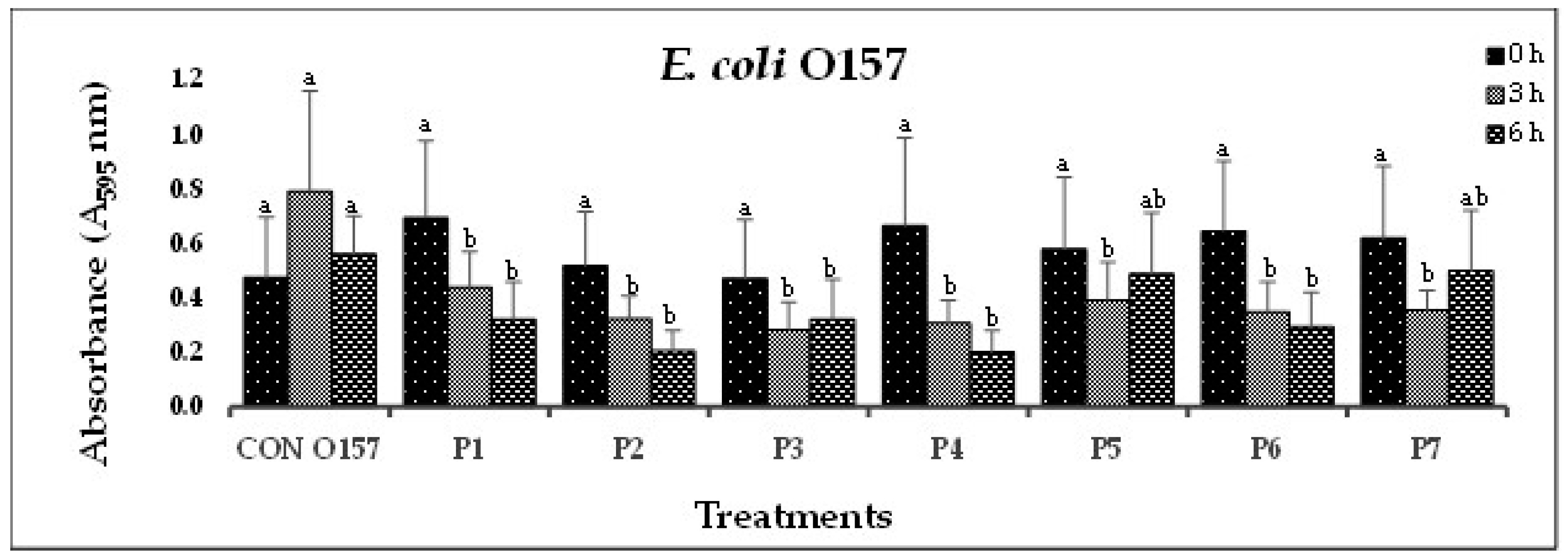
Figure 1.
(B). In-vitro biofilm disruption by individual bacteriophages specific to E. coli O26 after treatment for 0, 3, and 6 h.
Figure 1.
(B). In-vitro biofilm disruption by individual bacteriophages specific to E. coli O26 after treatment for 0, 3, and 6 h.
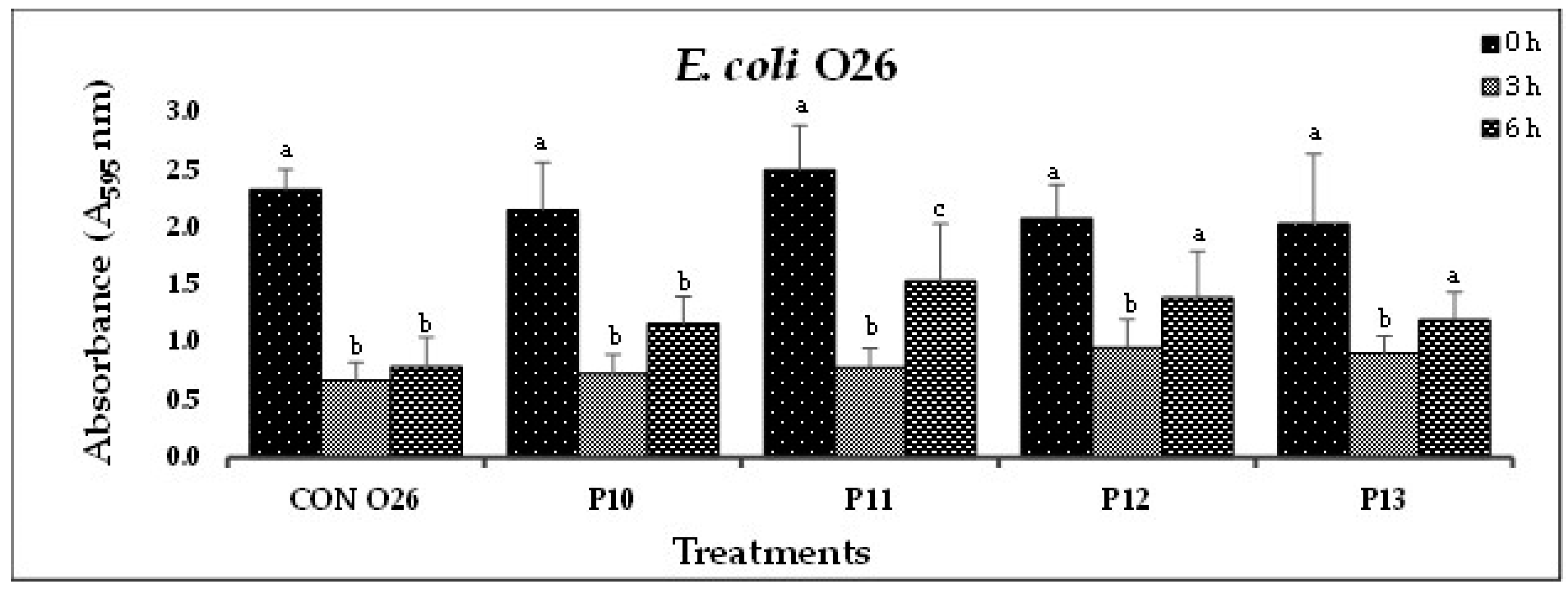
Figure 1.
(C). In-vitro biofilm disruption by individual bacteriophages specific to E. coli O111 after treatment for 0, 3, and 6 h. .
Figure 1.
(C). In-vitro biofilm disruption by individual bacteriophages specific to E. coli O111 after treatment for 0, 3, and 6 h. .
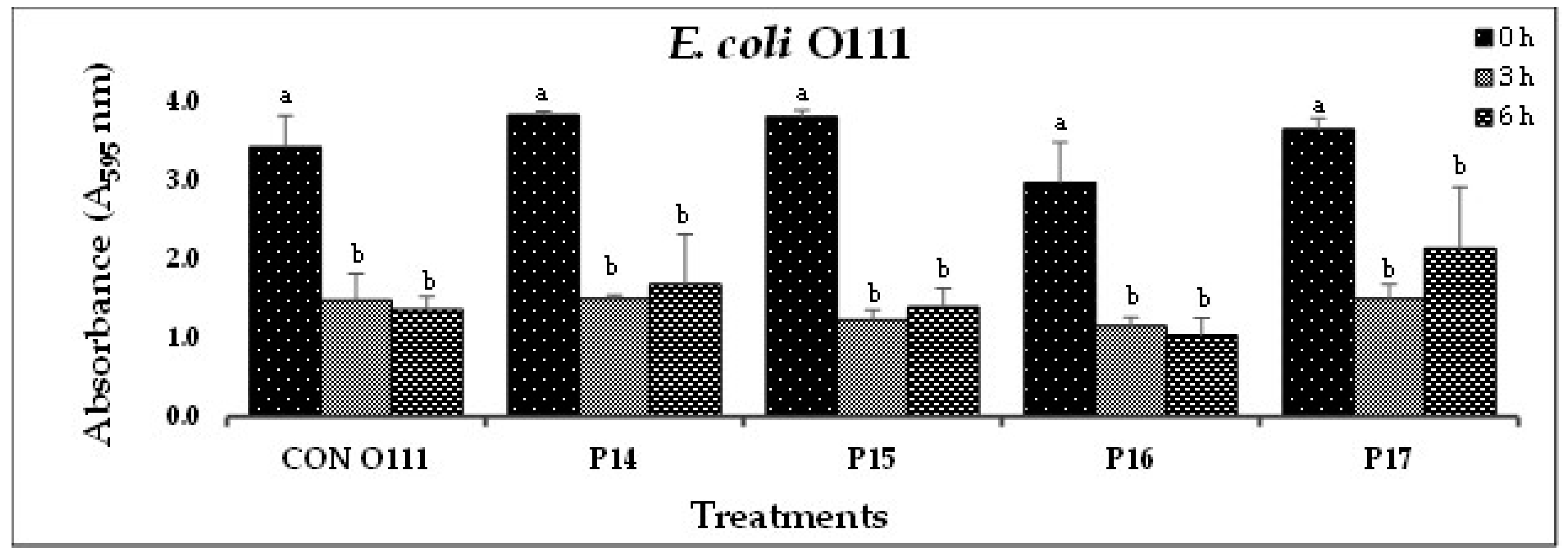
Figure 1.
(D). In-vitro biofilm disruption by individual bacteriophages specific to E. coli O103 after treatment for 0, 3, and 6 h.
Figure 1.
(D). In-vitro biofilm disruption by individual bacteriophages specific to E. coli O103 after treatment for 0, 3, and 6 h.
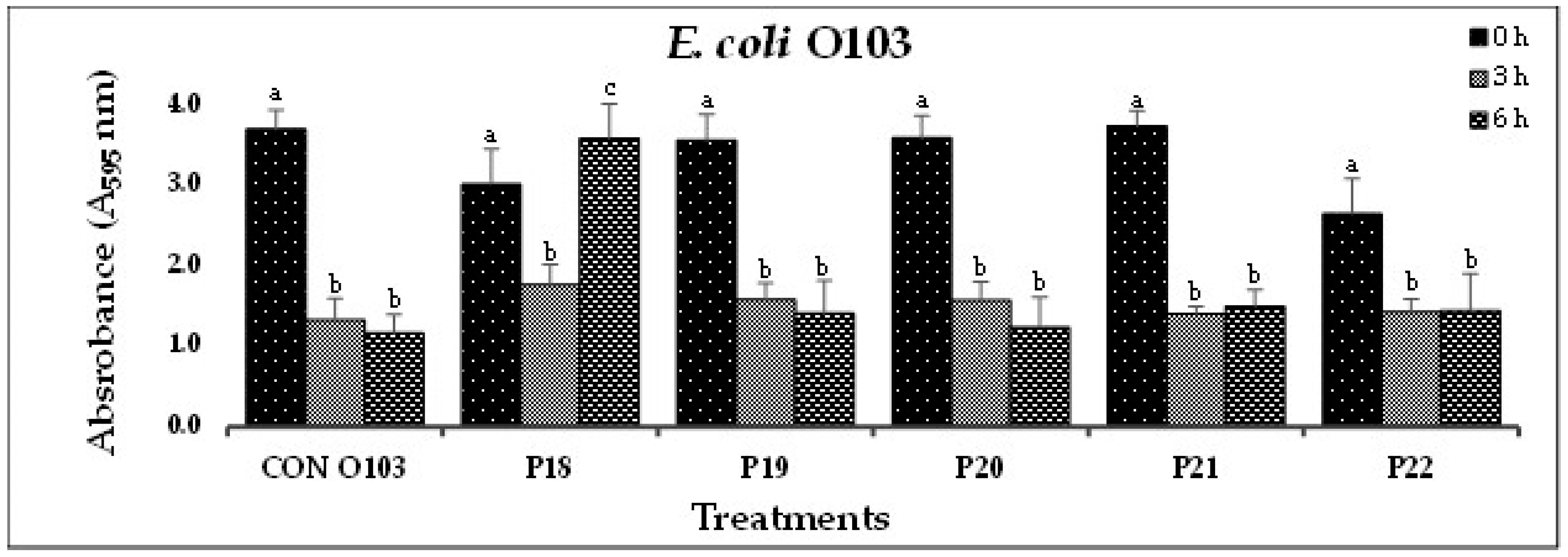
Figure 1.
(E). In-vitro biofilm disruption by individual bacteriophages specific to E. coli O121 after treatment for 0, 3, and 6 h.
Figure 1.
(E). In-vitro biofilm disruption by individual bacteriophages specific to E. coli O121 after treatment for 0, 3, and 6 h.
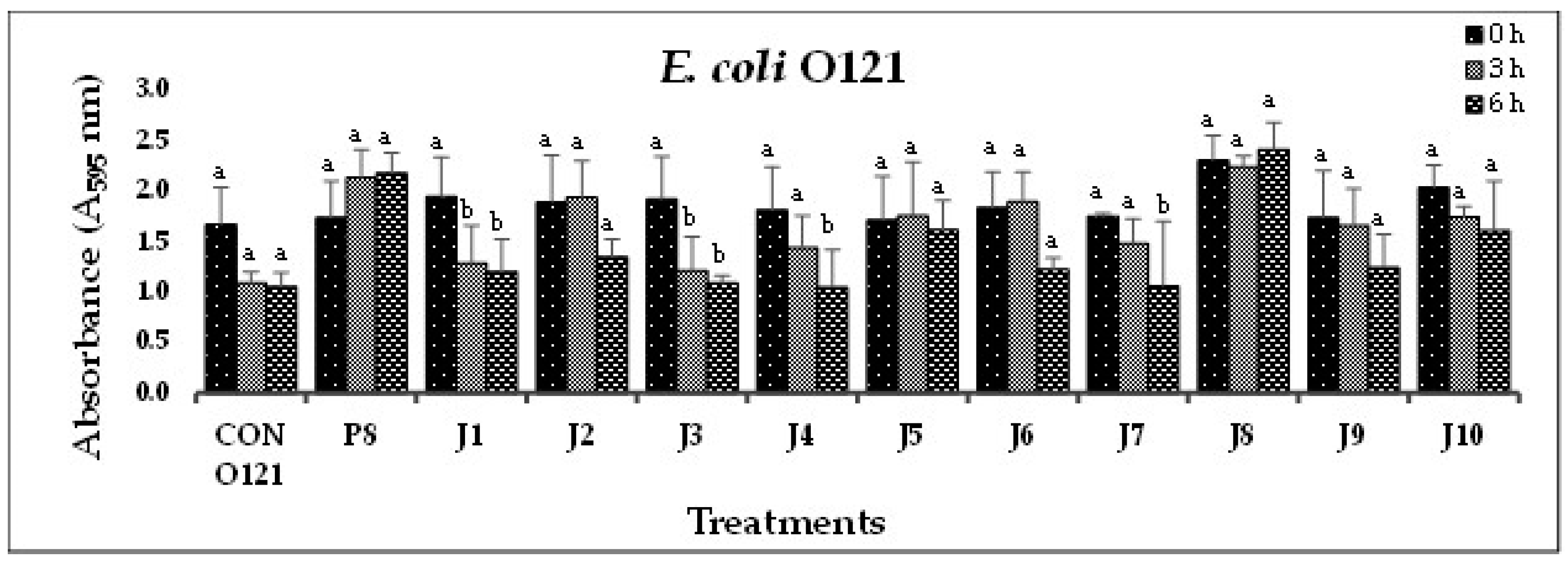
Figure 1.
(F). In-vitro biofilm disruption by individual bacteriophages specific to E. coli O45 after treatment for 0, 3, and 6 h. .
Figure 1.
(F). In-vitro biofilm disruption by individual bacteriophages specific to E. coli O45 after treatment for 0, 3, and 6 h. .
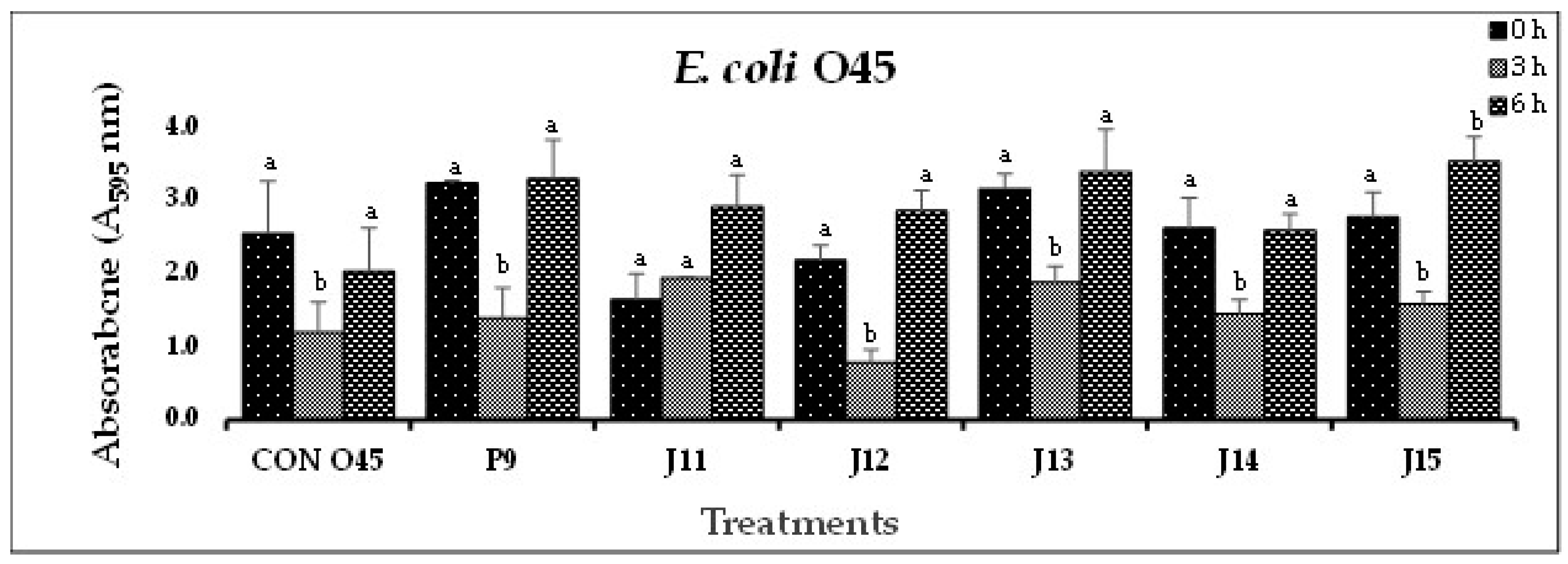
Figure 1.
(G). In-vitro biofilm disruption by individual bacteriophages specific to E. coli O145 after treatment for 0, 3, and 6 h. .
Figure 1.
(G). In-vitro biofilm disruption by individual bacteriophages specific to E. coli O145 after treatment for 0, 3, and 6 h. .
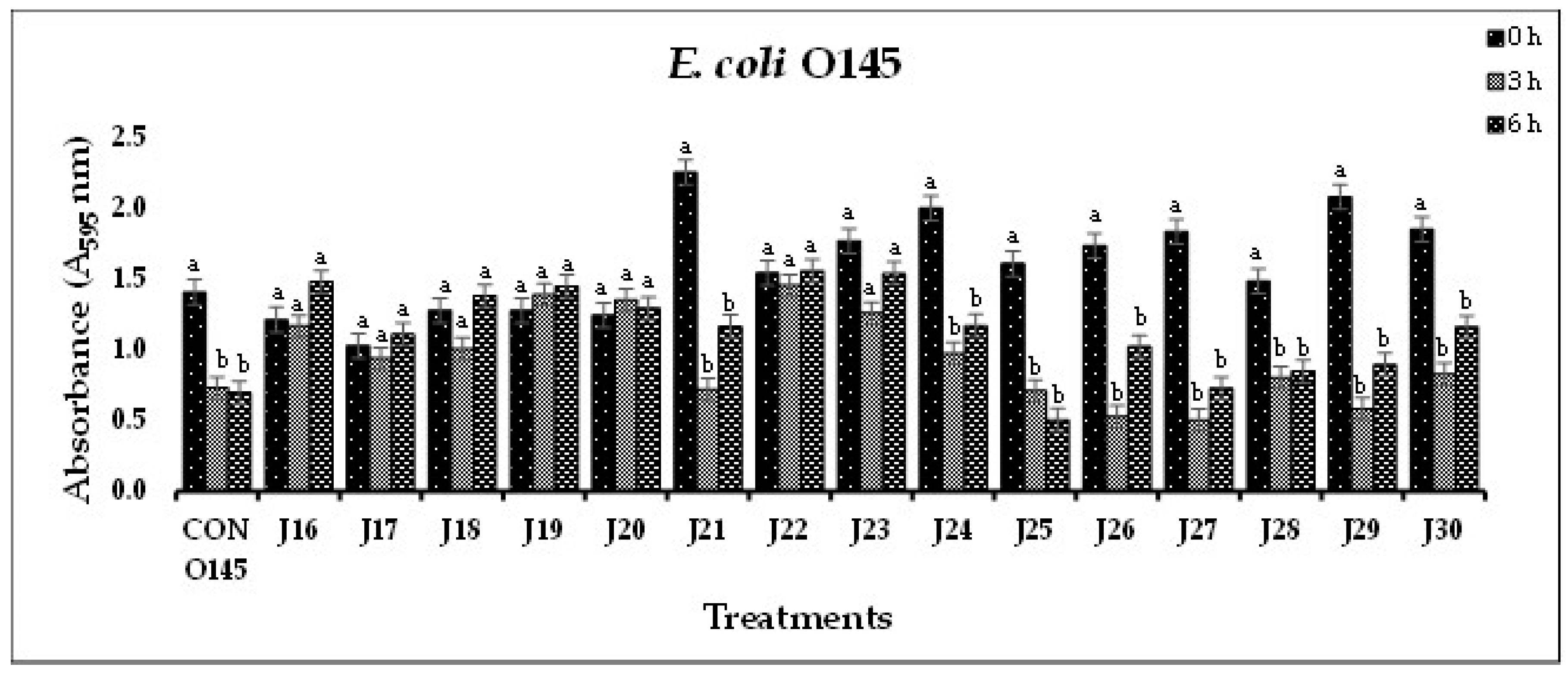
In Experiment-I, where individual phages were tested against their host pathogens, varying results were obtained. All the O157-phages (P1-O157 to P7-O157) showed a reduction in pathogen biofilms at 0 h (A595 = 0.472 – 0.696), 3 h (A595 = 0.284 – 0.441) and 6 h (A595 = 0.202 – 0.321), except phages P3, P5 and P7, where an increase in absorbance was observed at 6 h (Figure 1A). At 3 and 6 h, phage P4-O157 showed the highest reduction compared to the rest of the phages. Non-O157 STEC biofilms, treated with phages, also showed significant (P<0.10) reductions at 0 h (A595 =1.034 – 3.853), 3 h (A595 = 0.506 – 2.631) and 6 h (A595 = 0.202 – 3.577) (Figure 1B-G). However, STEC O45-specific phages were more effective after 3 h than after 6 h treatment. A list of the most effective serotype-specific phages is provided in Table 4.
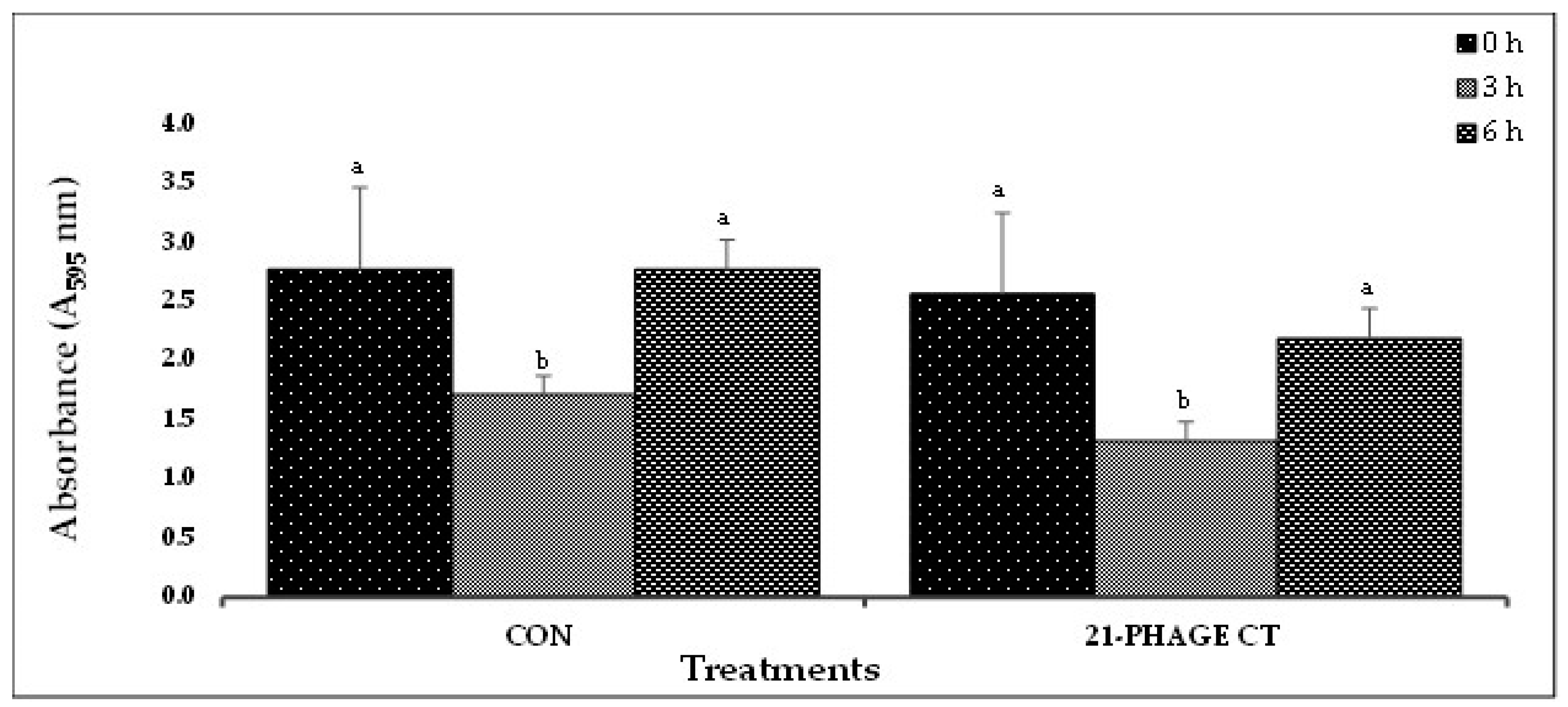
Based on the results from Experiment-I, selected phage cocktails were tested against their respective STEC-serotype biofilms (Experiment-II). A list of the most effective serotype-specific phage cocktails is provided in Table 4. Results revealed that all O157-phage cocktails (CT-1 to CT-4) were able to disrupt E. coli O157:H7 biofilms, reducing OD from 1.990 – 2.329 at 0 h to 0.576 – 0.636 at 6 h (Figure 2A). The CT-3 cocktail showed the highest reduction from 2.250 at 0 h to 0.748 at 3 h and CT-2 showed the highest reduction at 6 h from 2.319 to 0.576. Among the STEC nonO157 phage cocktails, CT5-O26, CT7-O103 and CT12-O145 performed the best by continuing to reduce biofilms at 6 h (Figure 2B,C). Phages from Experiment-II were selected to make a 21-phage cocktail (3 phages per serotype) to treat multi-serotype STEC biofilms (2 bacterial isolates per serotype). Results showed that phage cocktail reduced biofilm after 3 h of treatment, where OD decreased from 2.561 at 0 h to 1.321 at 3 h (Figure 2D). However, a slight increase in absorbance (2.186) was recorded after 6 h of phage-cocktail treatment (Figure 2D).
Figure 2.
(A). In-vitro biofilm disruption by bacteriophage cocktails specific to E. coli O157 after treatment for 0, 3, and 6 h.
Figure 2.
(A). In-vitro biofilm disruption by bacteriophage cocktails specific to E. coli O157 after treatment for 0, 3, and 6 h.
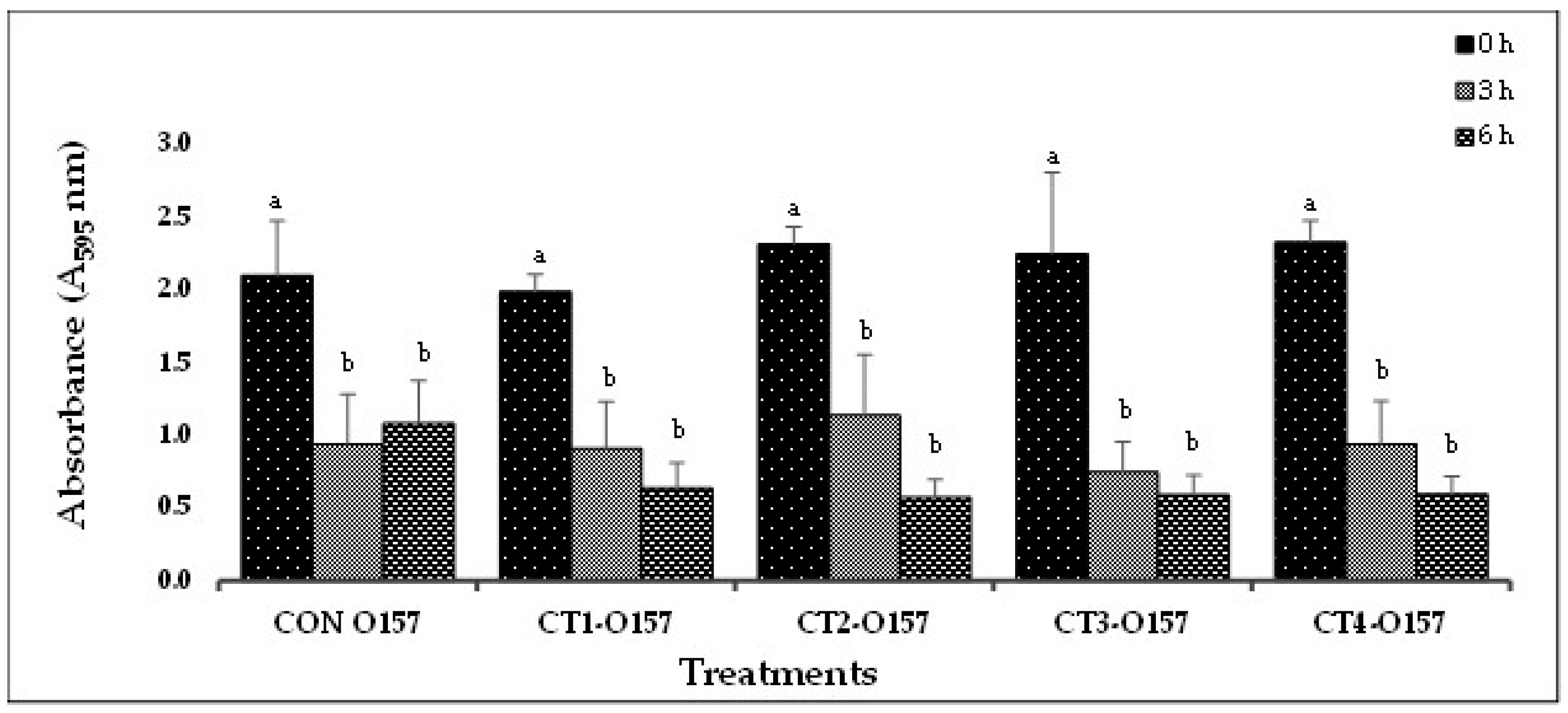
Figure 2.
(B). In-vitro biofilm disruption by bacteriophage cocktails specific to E. coli O121 and O145 after treatment for 0, 3, and 6 h.
Figure 2.
(B). In-vitro biofilm disruption by bacteriophage cocktails specific to E. coli O121 and O145 after treatment for 0, 3, and 6 h.
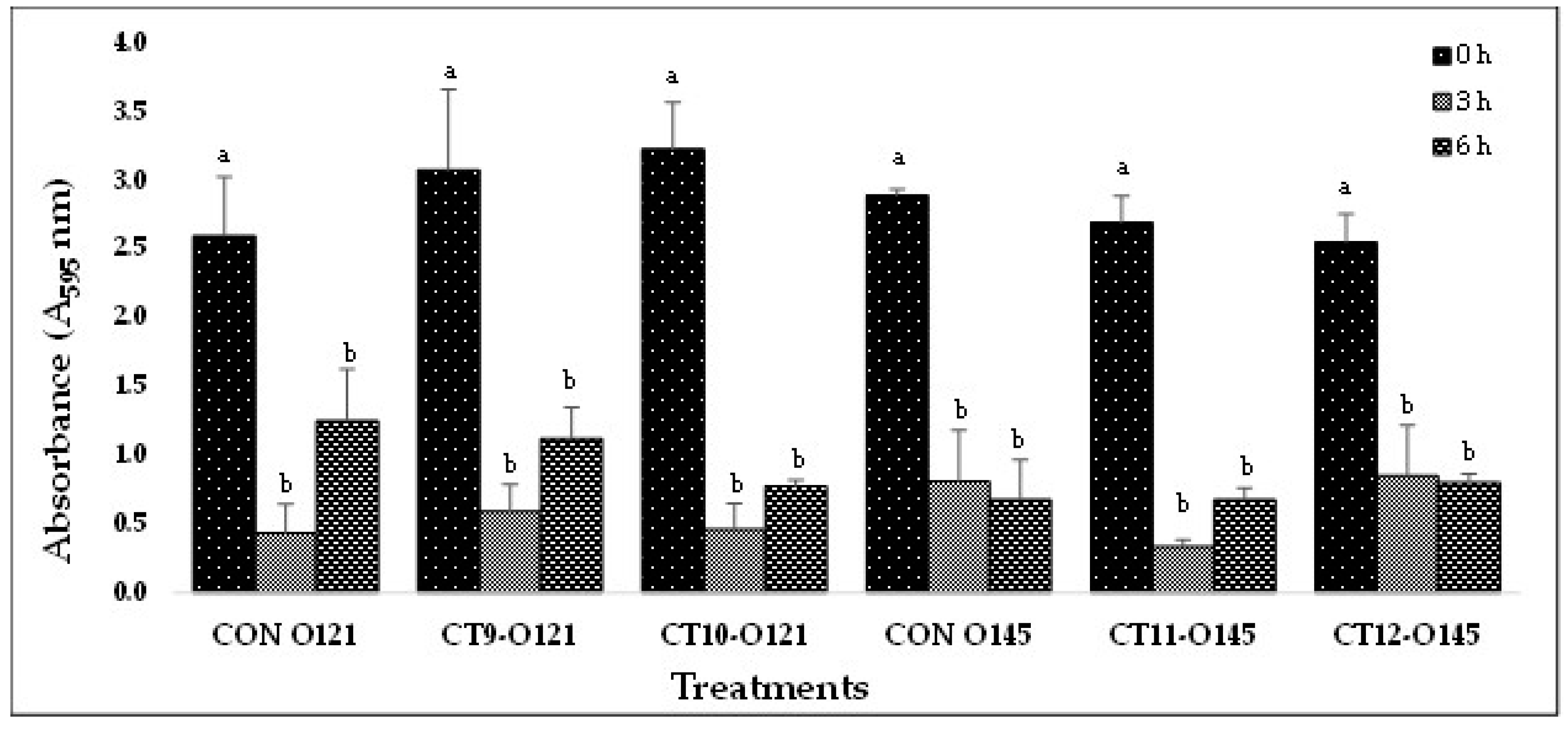
Figure 2.
(C). In-vitro biofilm disruption by bacteriophage cocktails specific to E. coli O26, O45, O103 and O111 after treatment for 0, 3, and 6 h.
Figure 2.
(C). In-vitro biofilm disruption by bacteriophage cocktails specific to E. coli O26, O45, O103 and O111 after treatment for 0, 3, and 6 h.
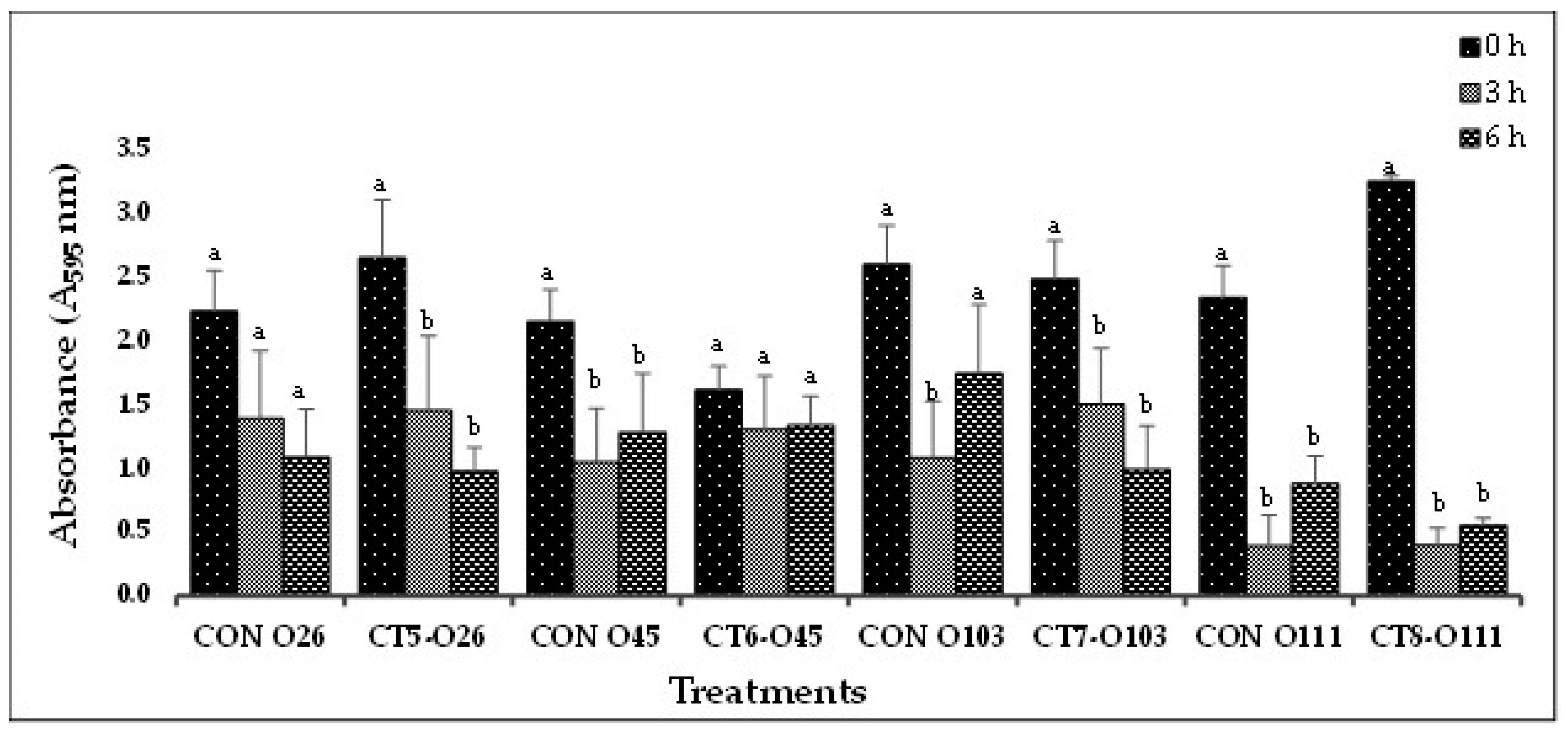
Figure 2.
(D). In-vitro STEC biofilm disruption by 21-phage cocktail after treatment for 0, 3, and 6 h. Values represent the average of three replications. Different letters a, b etc. represent significant difference (P<0.10) between incubation times (0, 3, 6 h) for the control or phage treatment. The bars with the same letters indicate that there is no significance difference (P<0.10) between incubation times (0, 3, 6 h) for the control or phage treatment..
Figure 2.
(D). In-vitro STEC biofilm disruption by 21-phage cocktail after treatment for 0, 3, and 6 h. Values represent the average of three replications. Different letters a, b etc. represent significant difference (P<0.10) between incubation times (0, 3, 6 h) for the control or phage treatment. The bars with the same letters indicate that there is no significance difference (P<0.10) between incubation times (0, 3, 6 h) for the control or phage treatment..
In the current study, a slight increase in OD was observed with some phages and phage cocktails at 6 h treatment. Results of the current study agree with other studies assessing phage treatment effects on bacterial biofilms [34,35]. Chan et al. [34] observed an increase in E. coli biofilms treated with T-4 phage on day 4, after an initial decrease (30%) on day 3. Similarly, Hughes et al. [36] obtained a maximum reduction in biofilm within 2 h or 5 h, depending on the biofilm studied. Factors such as bacterial appendages, used by the bacteria for biofilm attachment, could decrease the efficacy of phages in the biofilm [36,37]. Additionally, changes in bacterial biofilm profile over time, such as inconsistent expression and accumulation of protein through biofilm development stages, could affect phage efficacy [38,39]. Higher protein production at later stages of biofilm formation could impair phage movement within the medium and interfere with phage efficacy against bacterial biofilm [34,40]. Furthermore, static biofilms are known to produce higher polysaccharide and protein content, which could interfere with phage infection efficacies [41]. Vogeleer et al. [32] showed variation in biofilm matrices of individual non-O157 STEC serotypes under static and dynamic conditions. This variation in biofilm matrix could minimize phage effectiveness against biofilm, due to the highly specific nature of its depolymerase enzyme [24,36]. This enzyme is highly specific to EPS produced by host-bacteria, and even a minor change in the EPS composition could prohibit its activity, leading to reduced phage activity against biofilm.
Loss in phage effectiveness after 6 h could also be due to generation of phage resistant mutants [42]. Incubation of biofilms at optimum temperatures (30-37°C) could trigger the rapid growth of bacteria in the biofilm, including phage-resistant bacteria, resulting in decreased efficacy of the phage treatment [37,43]. It has also been shown that STEC serotypes, grown together to form biofilms, could generate an abundance of morphological variants, which could exhibit varying susceptibility to treatments [32,44]. Some newly generated mutants could lose the receptor responsible for bacteriophage susceptibility and become resistant to phages. The use of more diverse phages with the ability to bind to different bacterial receptors could help in controlling the emergence of bacterial mutants by exerting selective pressure on bacterial population in the biofilm and increasing the effectiveness of phage cocktail treatment [45]. At the same time, phage resistance has been shown to be transient in bacterial cells and they could revert to phage susceptible state [46]. Additionally, this decrease in phage effectiveness can be overcome by application of a cocktail of multiple phages [46,47]. Multi-phage cocktails were therefore evaluated in the current study for their effectiveness against STEC biofilms.
3.2. STEC biofilm disruption on food-contact surfaces
Based on the in-vitro experiments, the most effective phage cocktails were selected to conduct application studies on SS and HDPE coupons. In Experiment-I, where STEC serotype-specific cocktails were used to form biofilms on SS and HDPE coupons, phage cocktails significantly (P<0.05) reduced bacterial population in the biofilm when compared to the control (Figure 3A,B). Populations of E. coli O45, O111, O121, and O145 on SS were reduced to undetectable levels after treatment with respective phage cocktails (Figure 3A). A reduction of 3.1 and 3.7 logs was observed in E. coli O26 and O103 populations on SS coupons treated with phages, compared to the control (Figure 4A). On HDPE surface, E. coli O121 population was reduced to undetectable levels after respective phage cocktail treatment (Figure 3B). A reduction between 2.3-5.3 logs was observed in E. coli O26, O45, O103, O111 and O145 populations on HDPE coupons treated with respective phage cocktails, compared to the control (Figure 4B). Phage cocktails for specific STEC serotypes were very effective in reducing pathogen populations on both, SS (1.9-4.1 logs CFU/cm2 reduction) and HDPE (2.3-5.6 logs CFU/cm2 reduction) surfaces. These cocktails were more effective on SS than on HDPE, with the phage cocktails for E. coli O45, O145, O111, and O121 reducing pathogen populations to undetectable levels. These differences could be due to the surface type or difference in the biofilm composition, where different types of EPS were produced, which could provide increased resistance to bacteria against phage [47,48,49].
In Experiment-II, biofilms of a cocktail of 14 STEC isolates (two isolates per serotype), on SS and HDPE coupons, were treated with a 21-phage cocktail (three phages per serotype). The cocktail showed an immediate (0 h) reduction in STEC populations on HDPE (1.5 logs CFU/cm2) and SS (1.0 log CFU/cm2) (Figure 4A,B). At 3, 6 and 16 h, the 21-phage cocktail showed significant reductions (P<0.05) in STEC populations on both the surfaces when compared to the control (Figure 4A,B). At 3 h, pathogen populations were reduced by 2.8 and 1.7 logs CFU/cm2 on SS and HDPE, respectively. At 6 h, pathogen populations were reduced to undetectable levels on SS and by 1.7 logs CFU/cm2 on HDPE. At 16 h, STEC populations were reduced to undetectable levels on both surfaces. These results suggest that the tested bacteriophage treatments effectively reduced STEC populations in the biofilms on SS and HDPE surfaces.
Studies have previously shown that bacteriophages can effectively reduce pathogens attached to hard surfaces found in food processing environments [26,50,51]. Reductions between 3.5 to 5.4 logs CFU, in attached Listeria monocytogenes population, were observed on SS after 24 h of phage P100 treatment [52]. In a study by Sharma et al. [26], lytic bacteriophages reduced E. coli O157:H7 populations by 1.2 logs CFU on SS coupon. In the current study, phage treatments reduced E. coli O157:H7 by 1.9 logs CFU/cm2 and non-O157 STEC by 3.1-4.1 logs CFU/cm2 on SS surface. Studies have shown that phages can diffuse through the biofilm formed by bacteria [53] and that the formation of biofilms does not provide additional protection to bacteria against phage attack [26]. Biofilm break-down mechanism of phage is speculated to be associated with enzymatic means. Studies have shown that phages produce enzymes that can degrade the EPS layer, the major component of a biofilm matrix [24,36]. Hughes et al. [36] showed that bacteriophage specific for Enterobacter agglomerans disrupted biofilm through a combination of lytic activity against bacterial cells and degradation of EPS through phage-associated polysaccharide depolymerase enzyme. The presence of phage-depolymerase in an O45-phage (P-9), used in the current study, has been confirmed (data not shown), suggesting that other phages in the cocktail tested may also produce EPS-degrading enzyme, resulting in reduction of bacteria embedded in the biofilm matrix.
Variations in pathogen attachment and biofilm formation were observed on the food- contact surfaces tested in the present study. Higher STEC attachment was observed on HDPE (up to 5.6 logs CFU/cm2) compared to SS (up to 4.3 logs CFU/cm2) surface. A similar trend has been observed in previous studies. Higher bacterial attachment and biofilm formation has been observed on plastic surfaces (polyethylene, polypropylene, polyvinyl chloride) than on stainless steel or glass surfaces [48]. Variations found in polymer surfaces, such as smoothness, charge, Zeta-potential and active chemical groups, could influence bacterial attachment to the surface [54]. Studies have also shown that a variety of active chemical groups released by HDPE surfaces, such as phenols, quinones, aromatic hydrocarbons, aldehydes, and ketones, could be utilized by bacteria as carbon sources and result into higher bacterial attachment [55,56,57].
5. Conclusions
This study has shown the potential for use of lytic bacteriophages, against 7 STEC serotypes as a treatment to control biofilm formation. Future studies need to be conducted to understand phage-bacteria interactions in biofilm using kinetic modeling. The preliminary results from the current study show promise, due to the fact that phages within the biofilms effectively targeted and lysed STEC and were also able to disperse the extracellular matrix forming the biofilm.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, D.J. and P.K.L.; methodology, D.J. and P.K.L.; validation, D.J. and P.K.L.; formal analysis, D.J. and P.K.L.; investigation, D.J. and P.K.L.; resources, D.J.; data curation, D.J., P.K.L., K.R., and P.B; writing—original draft preparation, D.J. and P.K.L.; writing—review and editing, D.J., K.R. and P.B.; visualization, D.J.; supervision, D.J.; project administration, D.J.; funding acquisition, D.J. All authors have read and agreed to the published version of the manuscript.”
Funding
This research received no external funding
Data Availability Statement
Data is contained within the article.
Acknowledgments
This research was supported by the Technology and Business Development Program at the Oklahoma State University, Stillwater, Oklahoma.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.-A.; Roy, S.L.; et al. Foodborne illness acquired in the United States-major pathogens. Emerg. Infect. Dis. 2011, 17. [Google Scholar] [CrossRef] [PubMed]
- Brooks, J.T.; Sowers, E.G.; Wells, J.G.; Greene, K.D.; Griffin, P.M.; Hoekstra, R.M.; et al. Non-O157 Shiga toxin-producing Escherichia coli infections in the United States, 1983-2002. J. Infect. Dis. 2005, 192, 1422–1429. [Google Scholar] [CrossRef] [PubMed]
- Dewey-Mattia, D.; Manikonda, K.; Hall, A.J.; Wise, M.E.; Crowe, S.J. Surveillance for foodborne disease outbreaks-United States, 2009-2015. MMWR Surveill. Summ. 2018, 67, 1. [Google Scholar] [CrossRef]
- Tack, D.M.; Marder, E.P.; Griffin, P.M.; Cieslak, P.R.; Dunn, J.; Hurd, S.; et al. Preliminary incidence and trends of infections with pathogens transmitted commonly through food-Foodborne Diseases Active Surveillance Network, 10 US sites, 2015-2018. Morb. Mortal. Wkly. Rep. 2019, 68, 369. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). National Shiga toxin-producing Escherichia coli (STEC) Surveillance Annual Report, 2020. Available online: https://www.cdc.gov/ecoli/surv2020/ (accessed on May 2023).
- Ferens, W.A.; Hovde, C.J. Escherichia coli O157: H7: animal reservoir and sources of human infection. Foodborne Pathog. and Dis. 2011, 8, 465–487. [Google Scholar] [CrossRef] [PubMed]
- Kintz, E. , Brainard, J.; Hooper, L.; Hunter, P. Transmission pathways for sporadic Shiga-toxin producing E. coli infections: A systematic review and meta-analysis. Int. J. Hyg. and Environ. Health. 2017, 220, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Marouani-Gadri, N.; Augier, G.; Carpentier, B. Characterization of bacterial strains isolated from a beef-processing plant following cleaning and disinfection-influence of isolated strains on biofilm formation by Sakaï and EDL 933 E. coli O157: H7. Int. J. Food Microbiol. 2009, 133, 62–67. [Google Scholar] [CrossRef]
- Silagyi, K.; Kim, S.-H.; Lo, Y.M.; Wei, C.-i. Production of biofilm and quorum sensing by Escherichia coli O157: H7 and its transfer from contact surfaces to meat, poultry, ready-to-eat deli, and produce products. Food Microbiol. 2009, 26, 514–519. [Google Scholar] [CrossRef]
- Wang, R.; Bono, J.L.; Kalchayanand, N.; Shackelford, S.; Harhay, D.M. Biofilm formation by Shiga toxin-producing Escherichia coli O157: H7 and Non-O157 strains and their tolerance to sanitizers commonly used in the food processing environment. J. Food Prot. 2012, 75, 1418–1428. [Google Scholar] [CrossRef]
- Gilbert, P.; McBain, A.J. Potential impact of increased use of biocides in consumer products on prevalence of antibiotic resistance. Clin. Microbiol. Rev. 2003, 16, 189–208. [Google Scholar] [CrossRef]
- Weerarathne, P.; Payne, J.; Saha, J.; Kountoupis, T.; Jadeja, R.; Jaroni, D. Evaluating the efficacy of sodium acid sulfate to reduce Escherichia coli O157:H7 and its biofilms on food-contact surfaces. LWT- Food Sci. Technol. 2021, 139, 110501. [Google Scholar] [CrossRef]
- Fouladkhah, A.; Geornaras, I.; Sofos, J.N. Biofilm Formation of O157 and Non-O157 Shiga toxin-producing Escherichia coli and multidrug-resistant and susceptible Salmonella Typhimurium and Newport and their inactivation by sanitizers. J. Food Sci. 2013, 78. [Google Scholar]
- Houari, A.; Di Martino, P. Effect of chlorhexidine and benzalkonium chloride on bacterial biofilm formation. Lett. Appl. Microbiol. 2007, 45, 652–656. [Google Scholar] [CrossRef] [PubMed]
- Uhlich, G.A.; Rogers, D.P.; Mosier, D.A. Escherichia coli serotype O157: H7 retention on solid surfaces and peroxide resistance is enhanced by dual-strain biofilm formation. Foodborne Pathog. and Dis. 2010, 7, 935–943. [Google Scholar] [CrossRef]
- Gilbert, P.; Allison, D.; McBain, A. Biofilms in vitro and in vivo: do singular mechanisms imply cross-resistance. J. Appl. Microbiol. 2002, 92(s1). [Google Scholar] [CrossRef]
- Coates, A.; Hu, Y. Novel approaches to developing new antibiotics for bacterial infections. Br. J. Pharmacol. 2007, 152, 1147–1154. [Google Scholar] [CrossRef]
- Sulakvelidze, A.; Alavidze, Z.; Morris, J.G. Bacteriophage therapy. Antimicrob. Agents and Chemother. 2001, 45, 649–659. [Google Scholar] [CrossRef]
- Summers, W.C. Bacteriophage therapy. Annu. Rev. Microbiol. 2001, 55, 437–451. [Google Scholar] [CrossRef]
- Ashelford, K.E.; Day, M.J.; Fry, J.C. Elevated abundance of bacteriophage infecting bacteria in soil. Appl. Environ. Microbiol. 2003, 69, 285–289. [Google Scholar] [CrossRef]
- Synnott, A.J.; Kuang, Y.; Kurimoto, M.; Yamamichi, K.; Iwano, H.; Tanji, Y. Isolation from sewage influent and characterization of novel Staphylococcus aureus bacteriophages with wide host ranges and potent lytic capabilities. Appl. Environ. Microbiol. 2009, 75, 4483–4490. [Google Scholar]
- Raya, R.R.; Varey, P.; Oot, R.A.; Dyen, M.R.; Callaway, T.R.; Edrington, T.S.; et al. Isolation and characterization of a new T-even bacteriophage, CEV1, and determination of its potential to reduce Escherichia coli O157: H7 levels in sheep. Appl. Environ. Microbiol. 2006, 72, 6405–6410. [Google Scholar] [CrossRef] [PubMed]
- Tanji, Y.; Shimada, T.; Fukudomi, H.; Miyanaga, K.; Nakai, Y.; Unno, H. Therapeutic use of phage cocktail for controlling Escherichia coli O157: H7 in gastrointestinal tract of mice. J. Biosci. and Bioeng. 2005, 100, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Hughes, K.A.; Sutherland, I.W.; Jones, M.V. Biofilm susceptibility to bacteriophage attack: the role of phage-borne polysaccharide depolymerase. Microbiology 1998, 144, 3039–3047. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.; Sharma, M.; Millner, P.; Calaway, T.; Singh, M. Inactivation of Escherichia coli O157: H7 attached to spinach harvester blade using bacteriophage. Foodborne Pathog. Dis. 2011, 8, 541–546. [Google Scholar] [CrossRef]
- Sharma, M.; Ryu, J.H.; Beuchat, L. Inactivation of Escherichia coli O157: H7 in biofilm on stainless steel by treatment with an alkaline cleaner and a bacteriophage. J. Appl. Microbiol. 2005, 99, 449–459. [Google Scholar] [CrossRef]
- Viazis, S.; Akhtar, M.; Feirtag, J.; Diez-Gonzalez, F. Reduction of Escherichia coli O157: H7 viability on hard surfaces by treatment with a bacteriophage mixture. Int. J. Food Microbiol. 2011, 145, 37–42. [Google Scholar] [CrossRef]
- Litt, P.K.; Jaroni, D. Isolation and physio-morphological characterization of Escherichia coli O157:H7-infecting bacteriophages recovered from beef cattle operations. Int. J. Microbiol. 2017, 7013236. [Google Scholar]
- Litt, P.K.; Saha, J.; and Jaroni, D. Characterization of bacteriophages targeting non-O157 Shiga toxigenic Escherichia coli. J. Food Prot. 2018, 81, 785–794. [Google Scholar] [CrossRef]
- Jayasundera, B.P.K. Identification of contamination sources and occurrence of shigatoxigenic E. coli on small-scale cow/calf operations in Oklahoma and Louisiana. Masters Thesis, Oklahoma State University, 2015. [Google Scholar]
- Kountoupis, T.; Litt, P.K.; Kakani, R.; Jaroni, D. Biofilm forming capabilities of Shiga-toxigenic Escherichia coli recovered from cow calf operations in Oklahoma and Louisiana. Poster session presentation at the annual meeting of International Association for Food Protection, Tampa, FL,USA, 2017.
- Vogeleer, P.; Tremblay, Y.D.; Jubelin, G.; Jacques, M.; Harel, J. Biofilm-forming abilities of Shiga toxin-producing Escherichia coli isolates associated with human infections. Appl. Environ. Microbiol. 2016, 82, 1448–1458. [Google Scholar] [CrossRef]
- Hood, S.K.; Zottola, E.A. Adherence to stainless steel by foodborne microorganisms during growth in model food systems. Internat. J. Food Microbiol. 1997, 37, 145–153. [Google Scholar] [CrossRef]
- Chan, T.; Lee, K.; Roulin, P. Escherichia coli biofilm development and bacteriophage T4 susceptibility. J. of Expt. Microbiol. Immunol. 2007, 11, 73–80. [Google Scholar]
- Lu, T.K.; Collins, J.J. Dispersing biofilms with engineered enzymatic bacteriophage. Proc. Natl. Acad. Sci. 2007, 104, 11197–11202. [Google Scholar] [CrossRef] [PubMed]
- Hughes, K.A.; Sutherland, I.W.; Clark, J.; Jones, M.V. Bacteriophage and associated polysaccharide depolymerases-novel tools for study of bacterial biofilms. J. Appl. Microbiol. 1998, 85, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Gong, C.; Jiang, X. Application of bacteriophages to reduce biofilms formed by hydrogen sulfide producing bacteria on surfaces in a rendering plant. Can. J. Microbiol. 2015, 61, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Domka, J.; Lee, J.; Bansal, T.; Wood, T.K. Temporal gene-expression in Escherichia coli K-12 biofilms. Environ. Microbiol. 2007, 9, 332–346. [Google Scholar] [CrossRef]
- Schembri, M.A.; Kjaergaard, K.; Klemm, P. Global gene expression in Escherichia coli biofilms. Mol. Microbio. 2003, 48, 253–267. [Google Scholar] [CrossRef]
- Rabinovitch, A.; Aviram, I.; Zaritsky, A. Bacterial debris-an ecological mechanism for coexistence of bacteria and their viruses. J. Theor. Biol. 2003, 224, 377–383. [Google Scholar] [CrossRef]
- Sillankorva, S.; Neubauer, P.; Azeredo, J. Phage control of dual species biofilms of Pseudomonas fluorescens and Staphylococcus lentus. Biofouling 2010, 26, 567–575. [Google Scholar] [CrossRef]
- Meng, X.; Shi, Y.; Ji, W.; Meng, X.; Zhang, J.; Wang, H.; et al. Application of a bacteriophage lysin to disrupt biofilms formed by the animal pathogen Streptococcus suis. Appl. Environ. Microbiol. 2011, 77, 8272–8279. [Google Scholar] [CrossRef]
- Gong, C.; Heringa, S.; Singh, R.; Kim, J.; Jiang, X. Isolation and characterization of bacteriophages specific to hydrogen-sulfide-producing bacteria. Can. J. Microbiol. 2012, 59, 39–45. [Google Scholar] [CrossRef]
- Tremblay, Y.D.; Vogeleer, P.; Jacques, M.; Harel, J. High-throughput microfluidic method to study biofilm formation and host-pathogen interactions in pathogenic Escherichia coli. Appl. Environ. Microbiol. 2015, 81, 2827–2840. [Google Scholar] [CrossRef] [PubMed]
- Goodridge, L.D.; Bisha, B. Phage-based biocontrol strategies to reduce foodborne pathogens in foods. Bacteriophage 2011, 1, 130–137. [Google Scholar] [CrossRef] [PubMed]
- O'Flynn, G.; Ross, R.; Fitzgerald, G.; Coffey, A. Evaluation of a cocktail of three bacteriophages for biocontrol of Escherichia coli O157: H7. Appl.Environmen. Microbiol. 2004, 70, 3417–3424. [Google Scholar] [CrossRef] [PubMed]
- Kudva, I.T.; Hatfield, P.G.; Hovde, C.J. Characterization of Escherichia coli O157: H7 and other Shiga toxin-producing E. coli serotypes isolated from sheep. J. Clin. Microbiol. 1997, 35, 892–899. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.; Dowsett, A.; Dennis, P.; Lee, J.; Keevil, C. Influence of plumbing materials on biofilm formation and growth of Legionella pneumophila in potable water systems. Appl. Environ. Microbiol. 1994, 60, 1842–1851. [Google Scholar] [CrossRef]
- Sutherland, I.W. Biofilm exopolysaccharides: a strong and sticky framework. Microbiology 2001, 147, 3–9. [Google Scholar] [CrossRef]
- Abuladze, T.; Li, M.; Menetrez, M.Y.; Dean, T.; Senecal, A.; Sulakvelidze, A. Bacteriophages reduce experimental contamination of hard surfaces, tomato, spinach, broccoli, and ground beef by Escherichia coli O157: H7. Appl. Environ. Microbiol. 2008, 74, 6230–6238. [Google Scholar] [CrossRef]
- Hibma, A.M.; Jassim, S.A.; Griffiths, M.W. Infection and removal of L-forms of Listeria monocytogenes with bred bacteriophage. Int. J. Food Microbiol. 1997, 34, 197–207. [Google Scholar] [CrossRef]
- Soni, K.A.; Nannapaneni, R. Removal of Listeria monocytogenes biofilms with bacteriophage P100. J. Food Protect. 2010, 73, 1519–1524. [Google Scholar] [CrossRef]
- Briandet, R.; Lacroix-Gueu, P.; Renault, M.; Lecart, S.; Meylheuc, T.; Bidnenko, E.; et al. Fluorescence correlation spectroscopy to study diffusion and reaction of bacteriophages inside biofilms. Appl. Environ. Microbiol. 2008, 74, 2135–2143. [Google Scholar] [CrossRef]
- Triandafillu, K.; Balazs, D.; Aronsson, B.-O.; Descouts, P.; Quoc, P.T.; van Delden, C.; et al. Adhesion of Pseudomonas aeruginosa strains to untreated and oxygen-plasma treated poly (vinyl chloride)(PVC) from endotracheal intubation devices. Biomaterials 2003, 24, 1507–1518. [Google Scholar] [CrossRef] [PubMed]
- Brocca, D.; Arvin, E.; Mosbaek, H. Identification of organic compounds migrating from polyethylene pipelines into drinking water. Water Res. 2002, 36, 3675–3680. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, B.; Kowalski, D.; Rożej, A. Organic compounds migrating from plastic pipes into water. J. Water Supply: Res. Technol.-AQUA 2011, 60, 137–146. [Google Scholar] [CrossRef]
- Skjevrak, I.; Lund, V.; Ormerod, K.; Herikstad, H. Volatile organic compounds in natural biofilm in polyethylene pipes supplied with lake water and treated water from the distribution network. Water Res. 2005, 39, 4133–4141. [Google Scholar] [CrossRef] [PubMed]
Figure 3.
(A). Biofilm disruption on SS surface by bacteriophage cocktails specific to STEC after treatment for 16 h.
Figure 3.
(A). Biofilm disruption on SS surface by bacteriophage cocktails specific to STEC after treatment for 16 h.
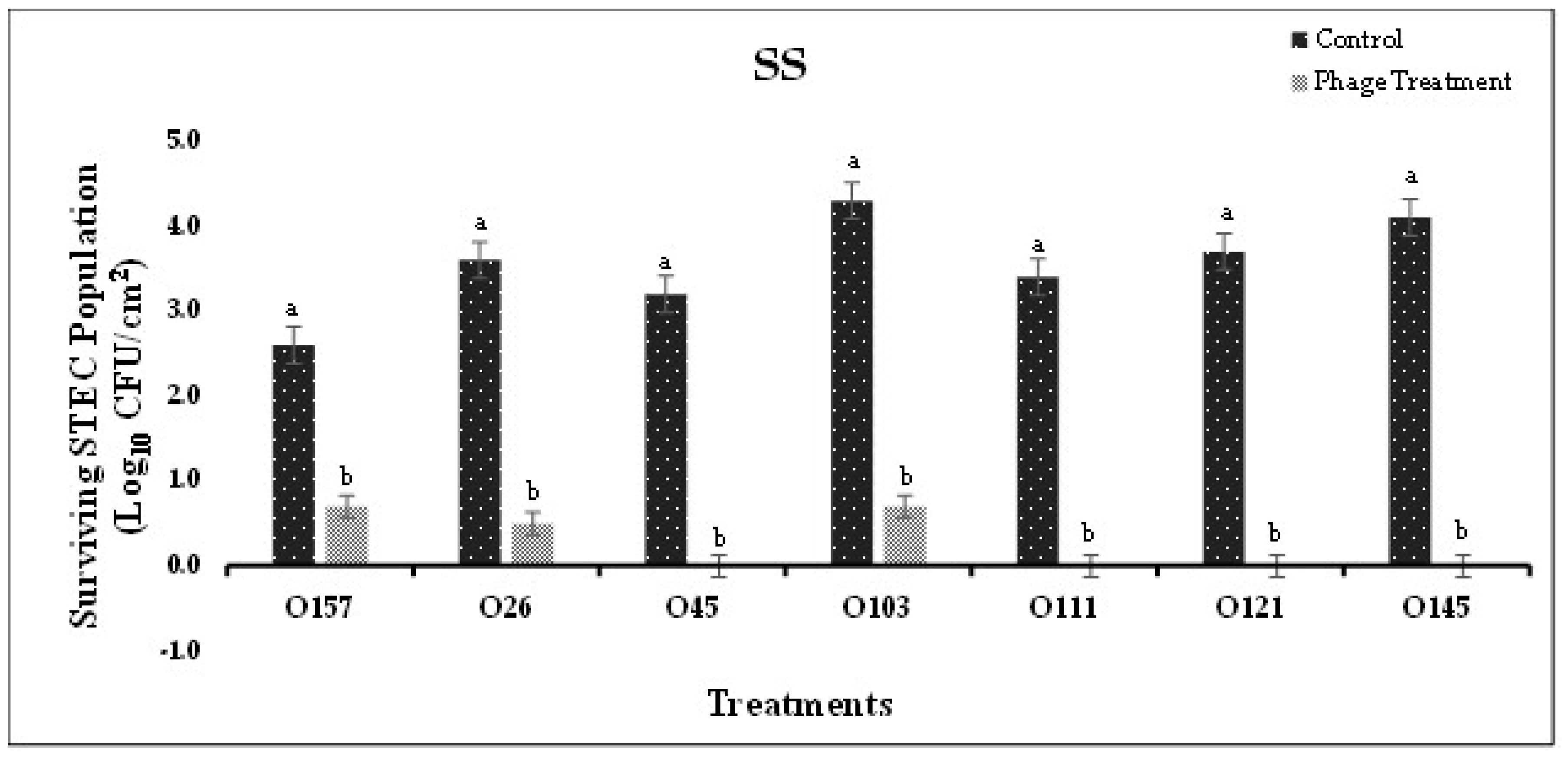
Figure 3.
(B). Biofilm disruption on HDPE surface by bacteriophage cocktails specific to STEC after treatment for 16 h.
Figure 3.
(B). Biofilm disruption on HDPE surface by bacteriophage cocktails specific to STEC after treatment for 16 h.
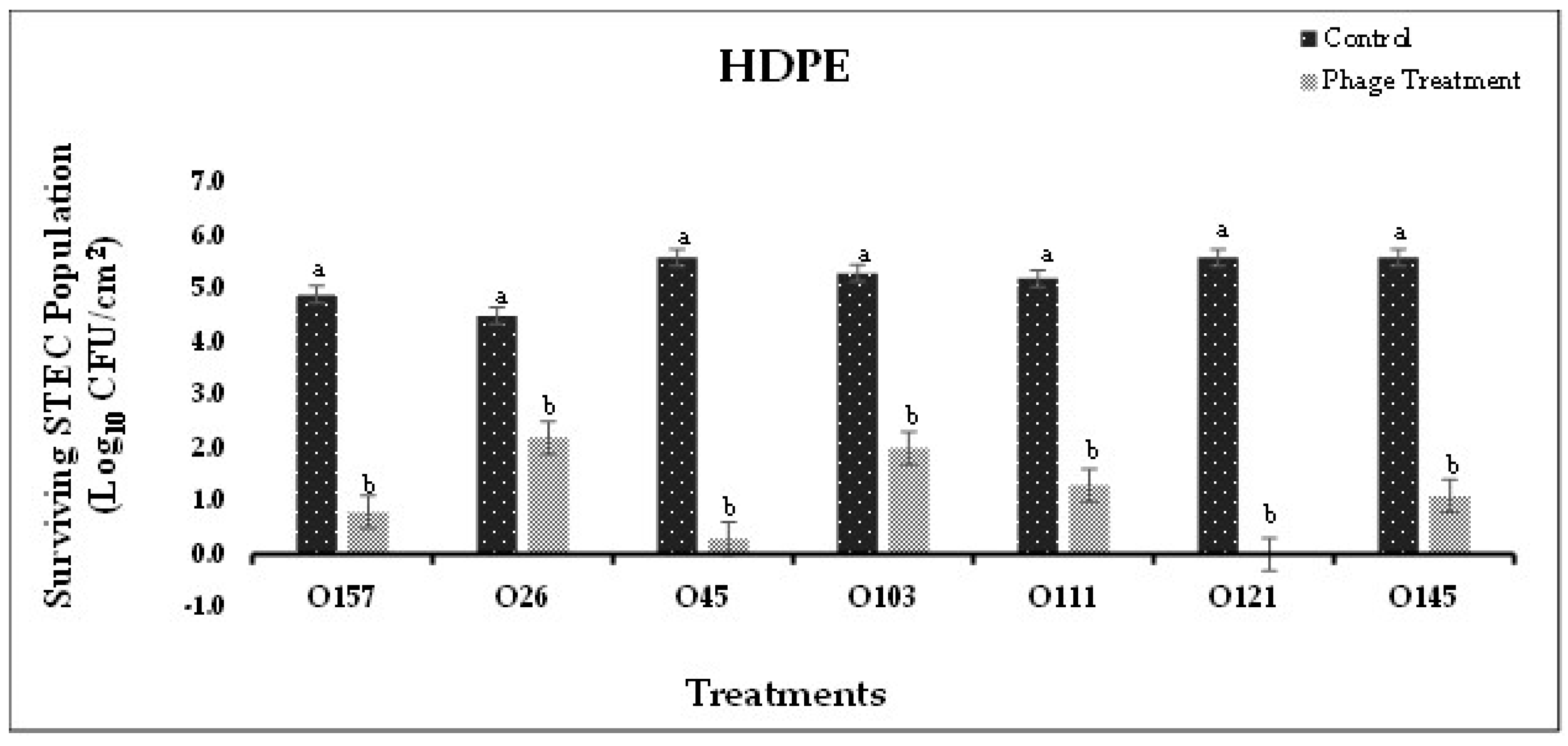
Figure 4.
Biofilm disruption on food contact surfaces by 21-phage cocktail after treatment for 0, 3, 6 and 16 h on (A) SS and (B) HDPE surfaces. Values represent the average of three replications ± standard deviation. Different letters a, b etc. represent significant difference (P < 0.05) between control (0 h and PBS) and treatments. The bars with the same letters indicate that there is no significance difference (P<0.05) between control (0 h and PBS) and treatments..
Figure 4.
Biofilm disruption on food contact surfaces by 21-phage cocktail after treatment for 0, 3, 6 and 16 h on (A) SS and (B) HDPE surfaces. Values represent the average of three replications ± standard deviation. Different letters a, b etc. represent significant difference (P < 0.05) between control (0 h and PBS) and treatments. The bars with the same letters indicate that there is no significance difference (P<0.05) between control (0 h and PBS) and treatments..
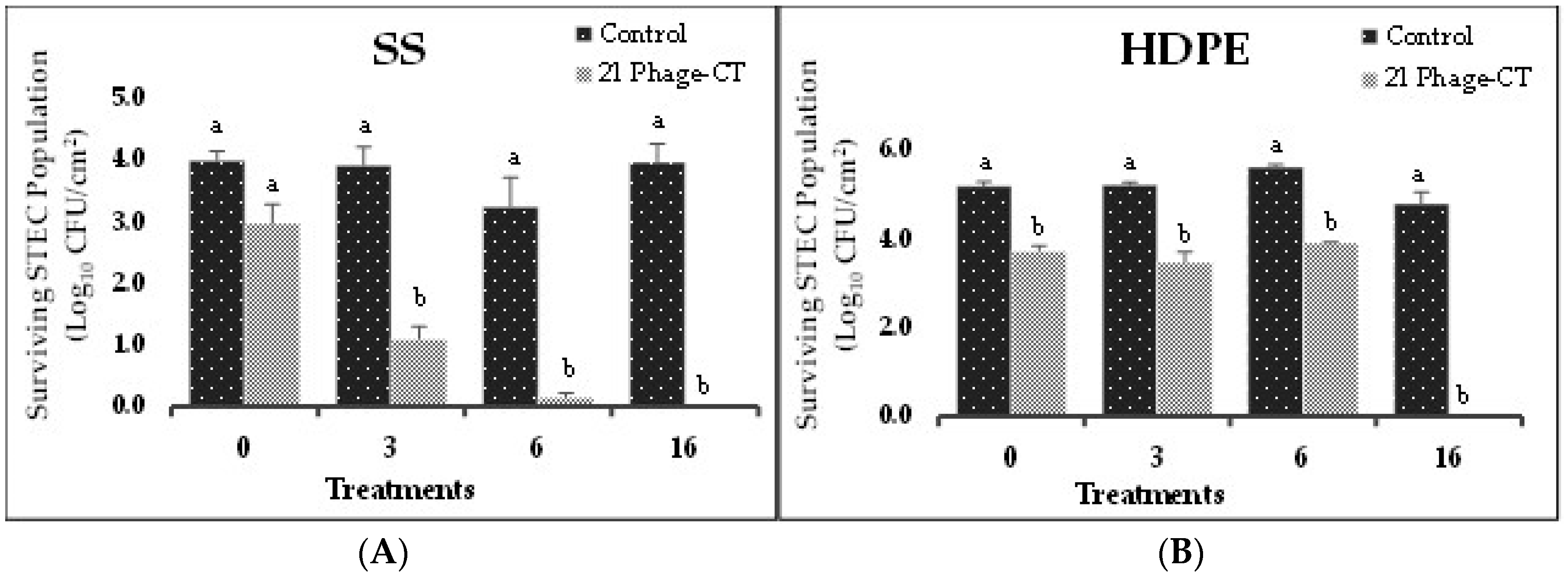

Table 1.
Bacterial and phage cocktails used in in-vitro Experiment-II.
| STEC | Bacterial Cocktail | Phage Cocktail Treatment (CT) | |
|---|---|---|---|
| O157 | ATCC 43895, WT: LF4, KF10 | CT1-O157 | P1, P2, P4, P6 |
| CT2-O157 | P1, P2, P5, P7 | ||
| CT3-O157 | P3, P5, P7 | ||
| CT4-O157 | P2, P3, P4, P7 | ||
| O26 | CDC 2003-3014, WT: BF8, QF6 | CT5-O26 | P10, P11, P12, P13 |
| O45 | CDC 2000-3039, WT: AF1, EF2 | CT6-O45 | P9, J12, J13, J15 |
| O103 | CDC 2006-3008, WT: AF10, GF6 | CT7-O103 | P19, P20, P21 |
| O111 | CDC 2010-3114, ATCC: 2440, 2180 | CT8-O111 | P14, P15, P16, P17 |
| O121 | CDC 2002-3211, ATCC 2219, 2203 | CT9-O121 | P8, J1, J4, J7 |
| CT10-O121 | P8, J3, J6, J9 | ||
| O145 | CDC 99-3311, ATCC 1652, 2208 | CT11-O145 | J21, J24, J26, J27 |
| CT12-O145 | J25, J28, J29, J30 | ||
Table 2.
Multi-serotype bacterial (14 strains) and phage (21 serotype-specific phages) cocktails used in in-vitro Experiment-III and in food-contact surface Experiment-II.
Table 2.
Multi-serotype bacterial (14 strains) and phage (21 serotype-specific phages) cocktails used in in-vitro Experiment-III and in food-contact surface Experiment-II.
| STEC | Bacterial Cocktail | Phage Cocktail |
|---|---|---|
| O157 | ATCC 43895, WT LF4 | P2, P6, P7 |
| O26 | CDC 2003-3014, WT QF6 | P11, P12, P13 |
| O45 | CDC 2000-3039, WT AF1 | P9, J12, J15 |
| O103 | CDC 2006-3008, WT AF10 | P19, P20, P21 |
| O111 | CDC 2010-3114, ATCC 2180 | P14, P15, P17 |
| O121 | CDC 2002-3211, ATCC 2219 | P8, J3, J7 |
| O145 | CDC 99-3311, ATCC 1652 | J18, J21, J29 |
Table 3.
Selected phage cocktails used in food-contact surface study, in Experiment I.
| STEC | Bacterial Cocktail | Phage Cocktail |
|---|---|---|
| O157 | ATCC 43895, WT: LF4, KF10 | P3, P5, P7 |
| O26 | CDC 2003-3014, WT: BF8, QF6 | P10, P11, P12, P13 |
| O45 | CDC 2000-3039, WT: AF1, EF2 | P9, J12, J13, J15 |
| O103 | CDC 2006-3008, WT: AF10, GF6 | P19, P20, P21 |
| O111 | CDC 2010-3114, ATCC: 2440, 2180 | P14, P15, P16, P17 |
| O121 | CDC 2002-3211, ATCC: 2219, 2203 | P8, J3, J6, J9 |
| O145 | CDC 99-3311, ATCC: 2208, 1652 | J21, J24, J26, J27 |
Table 4.
Most effective serotype-specific individual phages and phage cocktail treatments in-vitro.
| STEC | Individual Phage Treatment | Phage Cocktail Treatment (CT) |
|---|---|---|
| O157 | P4 | CT-2, CT-3 |
| O26 | P11 | CT-5 |
| O45 | P9 | CT-6 |
| O103 | P21 | CT-7 |
| O111 | P15 | CT-8 |
| O121 | J3 | CT-10 |
| O145 | J29 | CT-12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



