
Submitted:
27 June 2023
Posted:
28 June 2023
You are already at the latest version
Abstract
The most recent and non-invasive approach for studying early-stage biomarkers is liquid biopsy. It implies the extraction and analysis of non-solid biological tissues (serum, plasma, saliva, urine and cerebrospinal fluid) without undergoing invasive procedures to determine disease prognosis. Liquid biopsy can be used for the screening of several components such as extracellular vesicles, microRNAs, cell-free DNA, cell-free mitochondrial and nuclear DNA, circulating tumor cells, circulating tumor DNA, transfer RNA and circular DNA or RNA derived from body fluids. Its application includes early disease diagnosis, surveillance of disease activity, and treatment response monitoring, with growing evidence for validating this methodology in cancer, liver disease, and central nervous system (CNS) disorders. This review will provide an overview of mentioned liquid biopsy components, which could serve as valuable biomarkers for the evaluation of complex neurological conditions, including Alzheimer’s disease, Parkinson's disease, amyotrophic lateral sclerosis, multiple sclerosis, epilepsy, stroke, traumatic brain injury, CNS tumors, and neuroinfectious diseases. Furthermore, this review highlights the future directions and potential limitations associated with liquid biopsy.
Keywords:
microRNA
; liquid biopsy
; neurological diseases
1. Introduction
Neurological diseases present a formidable challenge in terms of accurate and timely diagnosis. Traditional diagnostic methods often require invasive procedures that take time and may lack the sensitivity and specificity for early-stage detection. In this regard, a recent non-invasive approach has emerged as a promising avenue for studying early-stage biomarkers: liquid biopsy. This innovative methodology involves extracting and analysing non-solid biological tissues using less-invasive approaches, providing valuable insights into disease diagnosis and limiting potential harm to the patient. Further, it offers a versatile screening platform capable of rapidly assessing various components with potential as specific liquid biomarkers for diagnosing and monitoring complex neurological conditions. However, the challenges and limitations of liquid biopsy are the need for more participants in current studies, variations in laboratory protocols, and the fact that many diseases share common features.
2. Liquid biopsy: definition and applications
Liquid biopsy involves extracting and analysing non-solid biological tissues, such as serum, plasma, saliva, urine, and cerebrospinal fluid (CSF), without requiring invasive procedures. Liquid biopsy is considered the most recent, advanced, and non-invasive approach for discovering biomarkers related to disease prognosis and treatment response status. This technique consists of evaluating several components in the fluid compartments that include extracellular vesicles (EVs) (exosomes, microvesicles, and apoptotic bodies), microRNA (miRNA), cell-free DNA (cfDNA), cell-free mtDNA (cf-mtDNA), cell-free nuclear DNA (cf-nuDNA), circulating tumor cells (CTCs), circulating tumor DNA (ct-DNA), transfer RNA (t-RNA) and circular DNA or RNA (circDNA/RNA). Figure 1, summarise the key components of liquid biopsy in neurological diseases.
EVs are single-lipid membrane vesicles secreted by cells into all body fluids, which can be classified based on their size and origin into three major types. The most studied EVs are exosomes, whose average diameter ranges between 30 to 100 nm, and are the smallest vesicles. Exosomes are produced by the endosomal pathway and release extracellular components, which then participate in cell-to-cell communication and transport of proteins, nucleic acid and metabolites across cells. The second EV type is micro-vesicles (MVs), whose average size ranges between 100 and 1000 nm and are produced by blebbing of the plasma membrane. Lastly, apoptotic bodies, with an average size of 50 to 5000 nm, are produced by dying cells. Cells from the CNS, such as neurons and glial cells, are known to release EVs to the extracellular space, where they can be identified by various methods, including specific cell surface markers or epigenetic information. Epigenetically, the methylation patterns of DNA within EVs can be studied in serum, plasma, or CSF samples, and their comparison with the already available atlas may lead to the identification of the cell types that produce EVs [1]. EVs have several functions in the body; in the CNS, they have been involved in various processes, including neurogenesis, neuron plasticity, and immune responses. Interestingly, it has been suggested that some EV cargoes modulate these critical CNS processes, which may lead to pathogenic degeneration when altered. As an example, co-culture of adipose-derived mesenchymal stem cells (ADMSCs) or ADMSC-secreted EVs with neurons pre-exposed to oxygen-glucose-deprivation (OGD) was associated with a significant reduction in neuronal cell death [2]. However, effects of ADMSCs-derived EVs were reversed if they were pre-treated with an exosomal secretion inhibitor. This neuroprotective effect of ADMSCs-EVs was suggested to be mediated by miR-25-3p, insomuch as miR-25-3p oligonucleotide mimics reduced cell death [2].
Exosomes primarily contain miRNA and small RNA, with minimal linear RNA, which is the most common form of RNA in the cell nuclei. Exosomes act as bidirectional cargo in brain-peripheral communication and within the brain environment [3]. Additionally, exosome cargoes have been linked to the epigenetic regulation of neuroglial communication within the CNS and the brain-body axis [4]. The miRNA received by the recipient is called “exosome shuttle RNA,” which could potentially influence the latter’s production. One study suggested that tumor cells release proteins and exosomes under hypoxic conditions, promoting the tumoral microenvironment for angiogenesis and subsequent metastasis [5]. Likewise, microglia-derived exosomes have been shown to induce inflammatory responses in traumatic brain injury models [6,7].
miRNAs can also freely circulate in body fluids without being enclosed in exosomal cargoes. These have been studied in various neurological diseases, including stroke, where circulating serum miRNA-221-3p and miRNA-382-5p were significantly lower in patients with ischemic stroke than in healthy controls, highlighting the potential role of freely circulating miRNAs in diagnosing this condition [8].
Another component of liquid biopsy is cell-free DNA (cfDNA), which refers to the double-stranded DNA fragments released from nucleated cells that circulate freely in the bloodstream. Elevated levels of cfDNA have been associated with stroke severity and poor outcome during the first three months of recovery. Moreover, tumor-related cfDNA increases in plasma and serum, offering potential therapeutic targets for treating tumors based on their mutational profile and monitoring treatment response [9]. cfDNA can be divided into two parts, namely cell-free mitochondrial DNA (cf-mtDNA) and cell-free nuclear DNA (cf-nu-DNA). cf-mtDNA consists of fragments from dysfunctional mitochondria that can be detected in the body’s fluid components. These fragments act as damage-associated molecular patterns (DAMPs), further triggering innate inflammatory responses. In the case of acute brain injury, elevated levels of cf-mtDNA were found in CSF and serum samples, and their levels were correlated with clinical severity and IL-6 cytokine response [10]. ct-DNA refers to DNA released by tumor cells. In some cancers, higher levels of ct-DNA have been linked to disease progression with specific epigenetic profiles showing certain gene mutations [11].
Circulating tumor cells (CTCs) are the cells that get discarded from the primary site of tumors into the blood circulation. Understanding the metastasis cascade of CTCs is essential, which helps identifying therapeutic targets against cancer metastasis. The low half-life of these cells makes them suitable to predict the real-time situation of cancer in the body [12].
In recent years, tRNA-derived RNA fragments (tRFs) have emerged as essential components of liquid biopsy due to their significant role in human diseases. Numerous studies have demonstrated that tRFs function as homeostatic regulators of the post-stroke immune response, potentially serving as biomarkers for increased infection risk in stroke patients. The overexpression of tRF-22-WE8SPOX52 has been shown to participate in the posttranscriptional regulation of genes by downregulating the Zbp1 protein, which acts as a DAMP sensor and induces NLRP3 inflammasome formation. Additionally, tRFs mediate in post-damage communication between the central nervous system (CNS) and immune cells [13]. Furthermore, a recent study involving transfection of rat neuronal cultures with various tRFs demonstrated neuronal swelling and subsequent cell death. This finding suggests that tRFs mediate the molecular mechanism leading to neuronal cell death via glutamate-induced neuronal necrosis [14].
Circular RNAs (circRNAs) are RNA molecules that circulate freely in body fluids. Most circRNAs function as miRNA sponges or RNA-binding proteins [15]. circRNAs have been implicated in various neurological diseases. Studies have indicated that circRS-7, acts as a miR-7 sponge in Alzheimer’s disease [16] and Parkinson’s disease [17], thereby modulating the expression of downstream genes such as cyclin B1 (G2/mitotic-specific) and CdK1 (Cyclin-dependent kinase 1), which suppresses cell proliferation and cell migration [18].
A non-coding RNA (ncRNA) is an RNA molecule not translated into a protein. Long non-coding RNA (lncRNA) includes intergenic long intervening non-coding (lincRNA), intronic ncRNAs, and sense and antisense lncRNAs. lncRNAs have been implicated in several neurological diseases, including ischemic stroke. Few studies have shown that in this entity, increased levels of non-coding nuclear abundant transcript 2 (NEAT2) were found in cultured brain microvascular endothelial cells (BMECs) after oxygen-glucose deprivation (OGD) and in isolated cerebral mice microvessels after middle cerebral artery occlusion (MCAO). Furthermore, silencing of NEAT2 significantly worsens the OGD-induced expression of proapoptotic components, such as Bcl-2 interacting mediator of cell death (Bim). This effect was studied in a NEAT2 knockout which presented a larger brain infarct size [19]. In a separate study, MCAO rats and OGD/R-treated neurocytes were investigated, and up-regulated expression level of lncRNA maternally expressed 3 (MEG3) were observed. MEG3 and absent in melanoma 2 (AIM2) functioned as molecular sponges to suppress miR-485. AIM2 was further validated as a target of miR-485 [20]. In a study focused on patients with acute mountain sickness, the expression levels of ncRNAs were analysed. The results showed significant up-regulation of lnc-CRKL-2 and lnc-NTRK3-4, and down-regulation of RPS6KA2-AS1 and lnc-CALM1-7 [21].
3. Liquid biopsy in neurological diseases
A wide range of techniques have been employed to study liquid biopsy in neurological diseases. These techniques include miRNA arrays for miRNA profiling, highly sensitive next-generation sequencing (NGS) for RNA sequencing, reduced representation bisulfite sequencing (RRBS), and Infinium MethylationEPIC BeadChip (EPIC) for analysing methylation patterns. Additionally, a highly sensitive Proximity Extension Assay (PEA) has been utilized, which combines immunoassay for antibody recognition with highly sensitive qPCR. Another notable technique is Small Input Liquid Volume Extracellular RNA (exRNA) Sequencing (SILVER-seq), which enables sequencing of this molecule in small liquid volumes. This section will provide the main findings of recent liquid biopsy studies in patients with neurological conditions. In addition, Table 1 summarizes the main findings associated with selected biomarkers identified in liquid biopsy studies.
3.1. Alzheimer’s disease
Alzheimer’s disease (AD) is the most common neurodegenerative disorder characterized by progressive cognitive decline and behavioural changes [41]. Key pathological hallmarks of AD include defects in amyloid precursor protein (APP) cleavage and beta-amyloid (Aβ) production, along with hyperphosphorylated tau protein aggregation, leading to an impaired synaptic function, conducing to neurodegeneration [42].
One approach utilizes a highly prevalent DNA modification in the CNS, mainly to derive 5hmC signatures from plasma cfDNA to segregate the multiple AD pathogenesis-related pathways in late-onset AD, compared to healthy controls. Several genes were identified, such as RABEP1, CPNE4, DNAJC15, REEP3, ROR1, CAMK1D, and RBFOX1, showing a strong correlation with disease scores [43].
Components previously considered cellular debris, such as EVs, circRNA, exRNA, and tRFs, have been explored for their potential to serve as biomarkers in AD. For example, neuron-derived extracellular vesicles (NDEVs) were immunoprecipitated by targeting neuronal markers such as L1CAM from AD patients and healthy controls. Higher NDEV Aβ42 levels were associated with better scores in patients’ memory and cognitive status [22]. Another study using SILVER-seq technology observed differentially expressed candidate genes between AD patients and healthy controls, finding phosphoglycerate dehydrogenase (PHGDH) up-regulated in the AD brain. Increased plasma levels of PHGDH’s exRNA were associated with disease converters, reflecting the transition from normal cognition to impairment [23]. Additionally, a separate study reported increased cirHomer1 and hsa_circ_0131235 expressions associated with AD pathology [44,45]. Upregulation of tRFs has also been reported in the hippocampus of AD patients, but the function of these small RNAs remains unclear [46,47]. Overall, these findings suggest that fluid biomarkers can be detected in the body fluids of AD patients using non-invasive techniques, serving as biomarkers for the early detection or progression of the disease.
3.2. Parkinson’s disease
Parkinson’s disease (PD), the second-most common neurodegenerative disease, is characterized by the loss of dopaminergic neurons in the substantia nigra and the widespread accumulation of Lewy bodies, mainly composed of fibrillar aggregates of the presynaptic protein α-synuclein [48]. Despite recent advances in diagnosing PD, including advanced imaging techniques, definite diagnosis remains limited to clinical observations, warranting a high need for reliable biomarkers to identify the early stages of the disease [49]. In this sense, the isolation of EVs from blood or other body fluids has emerged as a promising approach for diagnosing and monitoring PD progression. Studies have shown that levels of α-synuclein inside plasma EVs are significantly greater in patients at early stages of PD than in healthy controls, indicating their potential as a diagnostic and prognostic biomarker [50]. Moreover, the identification and profiling of CSF and blood-derived miRNAs can aid in differentiating PD cases from healthy individuals. Several differentially expressed miRNAs have been identified, including miR-132-5p, miR-34c-3p, miR-132-3p, miR-19b-3p, miR-29c, miR-133b, and miR-331-5p [24,51]. Furthermore, EVs derived from plasma samples of PD patients showed decreased levels of hsa-miR-15b-5p, hsa-miR-138-5p, hsa-miR-338-3p, hsa-miR-106b-3p, and hsa-miR-431-5p, while levels of hsa-miR-30c-2-3p were increased. The expression of the genes hsa-miR-15b-5p, hsa-miR-30c-2-3p, hsa-miR-138-5p, and hsa-miR-338-3p was increased in the dopaminergic synapse and the PD pathway, indicating their potential role in modulating dopamine expression and their use as potential diagnostic biomarkers [25].
Saliva is another potential source of biomarkers for early screening of PD. Commonly detected biomarkers in saliva include α-synuclein, protein deglycase (DJ-1), and oxidative stress markers such as hemeoxygenase-1 (HO-1), acetylcholinesterase (AChE), and total protein [52].
Although urine samples are highly diluted, they can also serve as a source of biomarkers for PD. Higher urine kynurenine (KYN) levels have been associated with early stages of PD and correlated with mild cognitive impairment and disease severity [53]. Additionally, cf-mtDNA has been identified as a potential biomarker, as its levels were significantly reduced in the CSF of PD patients compared to control diseases [54].
In summary, several approaches have been tested in various body fluids to diagnose PD earlier. These studies resulted in the preliminary identification of biomarkers for the disease, although further validation in larger cohorts is needed.
3.3. Amyotrophic lateral sclerosis
Amyotrophic lateral sclerosis (ALS) is a fatal adult-onset neurodegenerative disorder characterized by the progressive loss of upper and lower motor neurons [55]. Although most ALS cases are sporadic (sALS), about 5-10% correspond to familial forms with mutations in diverse genes, including Cu2+/Zn2+ superoxide dismutase (SOD1) and TAR DNA-binding protein 43 (TDP-43), among others [56]. Neuroinflammatory responses associated with the activation of microglia and astrocytes are prominent features of ALS and are believed to contribute to disease progression [57]. Recently, liquid biopsy approaches using CSF have been employed to identify potential biomarkers of ALS. The ultrasensitive protein detection method, PEA, was applied to CSF-EV samples, and it was found that the junctional adhesion molecule A (JAM-A) protein, chitinase-1, and tumor necrosis factor receptor-2 (TNF-R2) were significantly up-regulated in ALS cases. At the same time, myoglobin was down-regulated [58]. NGS was performed on exosome messenger RNA (mRNA) isolated from CSF. This methodology identified CUE Domain Containing 2 (CUEDC2) as the top candidate biomarker for this disease, as it was highly increased in CSF from ALS cases [26]. Similarly, differential methylation markers were obtained from cfDNA isolation from CSF and plasma of ALS patients for biomarker discovery. This study identified a novel differentially methylated region in the RHBDF2 gene in ALS patients compared to controls, highlighting its potential as an epigenetic biomarker of neurodegeneration [59].
A recent study identified a biomarker signature of ncRNA in serum samples of ALS patient including hsa-miR-16-5p, hsa-miR-21-5p, hsa-miR-92a-3p, hsa-piR-33151, TRV-AAC4-1.1, and TRA-AGC6-1.1. This combination accurately differentiated ALS from non-ALS cases [60]. Furthermore, other studies have identified hsa-miR-4299 and hsa-miR-4649-5p as down-regulated and up-regulated miRNAs, respectively in the plasma of sALS patients. Additionally, hsa-miR-663b and hsa-miR-4258 were significantly down-regulated in the CSF of patients, while no miRNAs were found to be up-regulated [28].
In summary, these liquid biopsy approaches offer promising insights into identifying potential biomarkers for ALS. However, further research is needed to validate and translate these findings into clinical practice.
3.4. Multiple Sclerosis
Multiple Sclerosis (MS) is a disease that presents with neuroinflammation and neurodegeneration [61]. Several studies have searched for biomarkers in different clinical phenotypes of MS. Firstly, the presence of anti-myelin lipid-specific oligoclonal IgM bands (LS-OCMBs) in CSF is considered a predictor of the aggressive evolution of the disease. However, obtaining CSF through a lumbar puncture is an invasive approach. To overcome this issue, Iparraguirre et al. (2020) used less invasive approaches, i.e., peripheral blood mononuclear cells (PBMCs). Transcriptomic analysis was performed on these samples, and further bioinformatics analysis suggested that several free circular and linear RNA biomarkers were differentially expressed. After performing validation experiments by qPCR, only two circRNAs (hsa_circ_0000478 and hsa_circ_0116639) and two linear RNAs (IRF5 and MTRNR2L8) were found to be significantly differentially expressed between LS-OCMBS positive and negative patients [62].
Exosomes derived from body fluids (serum, plasma, or CSF) of MS patients and healthy controls have also been exploited in various liquid biopsy approaches. One group reported increased expression of Epstein-Barr virus (EBV) nuclear antigen EBNA1, and latent membrane proteins LMP1 and 2A on serum exosomes derived from patients with active relapsing-remitting MS (RRMS) compared to healthy controls or stable RRMS [63]. Another study demonstrated diminished expression of hsa-miR-122-5p, hsa-miR-196b-5p, hsa-miR-301a-3p, and hsa-miR-532-5p in serum samples of RRMS patients as compared to healthy controls. Interestingly, these miRNAs have been involved in direct cell-to-cell communication [30]. Lastly, exosomes were isolated from urine, plasma, and CSF samples from MS patients and healthy controls and the authors reported increased levels of miR-155-5p on day 6 before disease onset [31].
Two viruses are related to MS and demyelinating events pathogenesis. EBV is considered a causative event that can lead to MS development and could also be identified by detecting EBV-derived cell-free DNA in CSF or plasma samples [29]. Another viral pathogen related to demyelinating diseases is the human John Cunningham virus (JCV), causing progressive multifocal leukoencephalopathy (PML) [64], which mainly affects MS patients due to the reactivation of a latent JCV infection in the brain as a severe adverse effect originating from the immunosuppressive treatment [65]. The viral DNA can be detected in CSF by PCR assays, helping to arrive to an accurate diagnosis in combination with typical brain MRI findings [66].
These results suggest that MS patients showed differential expression of several components in various body fluids, such as miRNA isolated from exosomes and several free-circulating linear and circular RNA biomarkers.
3.5. Epilepsy
Epilepsy is one of the most common neurological diseases [67], affecting over 70 million people worldwide [68]. It is a chronic disease that primarily affects the brain cortex as a consequence (or not) of CNS insults, predisposing patients to suffer seizures [67]. Therefore, patients with this condition may experience severe long-term consequences, including increased comorbidities, disability, and mortality [69].
Regarding disease epidemiology and pathogenesis, quantifying lncRNA in the peripheral blood of epileptic patients showed that molecules such as HOXA-AS2 and SPRY4-IT1 have a significantly increased expression in male epilepsy patients, suggesting that these two lncRNAs could potentially play a role in the pathogenesis of the disease [70]. In line, studying cfDNA methylation in serum or plasma samples could provide new biomarkers for identifying various stages of epileptogenesis and high-risk patients. This is supported by the fact that some neurodegenerative disorders involve methylation remodelling in the DNA of genes related to inflammation and disease duration, as observed in the anterior temporal neocortex and hippocampus of mesial temporal lobe epilepsy with hippocampal sclerosis (mTLE-HS) patients. Some of these methylation patterns may also be present in the circulation and derive from damaged neurons or glia [71]. For example, a study suggested that the mean baseline concentration of cfDNA was lower in patients with extratemporal lobe epilepsy (XTLE) than in control subjects. Furthermore, the maximum concentration of cfDNA after baseline measurement was significantly lower in patients with a disease duration of more than 18 years than in those with a disease duration of less than 18 years [72].
The most extensively studied liquid biopsy markers for epilepsy are free-circulating or EV-derived miRNAs originating from neurons or glia that circulate in the blood or CSF. In patients with temporal lobe epilepsy (TLE) -which comprises 30% of all epilepsy cases- and status epilepticus (SE), the EVs derived from CSF have been found to contain numerous differentially expressed miRNAs. TLE-derived EVs showed increased levels of miR-19b-3p and decreased levels of seven different miRNAs, while SE-derived EVs had increased levels of miR-21-5p, miR-451a, and six other miRNAs, along with decreased expression of miR-204-5p [73]. Further, serum samples of TLE patients showed elevated levels of the lncRNA ILF3-AS1 compared to controls. This molecule is closely associated with epilepsy due to its role in promoting the expression of metalloproteinases (MMPs) and other pro-inflammatory cytokines commonly found in this disease [74]. Additionally, in mTLE-HS, six miRNAs were found to be differentially expressed compared to healthy controls. Only miR-8071 exhibited high sensitivity and specificity, and was additionally associated with seizure severity [32].
In other focal epilepsies, plasma samples are a good source of tRFs. Specifically, 5′GluCTC-tRF, 5′GlyGCC-tRF, and 5′AlaTGC-tRF are increased in samples prior to seizure compared to post-seizure samples, reflecting the potential role of tRFs as non-invasive predictors of epilepsy risk [33]. Further, differences in the expression levels of microRNAs were observed between epileptogenic and non-epileptogenic tuberous sclerosis complex (TSC) tubers. Epileptogenic tubers showed a significant increase in miR-142-3p, miR-223-3p, and miR-21-5p compared to non-epileptogenic TSC tubers. These miRNAs are known to activate toll-like receptors 7 and 8 (TLR-7/8), resulting in the propagation of neuroinflammatory responses [75]. Moreover, neuronal cfDNA found in plasma could help identify somatic epilepsy mutations, as this cfDNA could have crossed the BBB due to its increased permeability derived from brain damage [76].
Regarding treatment, a few biomarkers have been linked to drug resistance or favourable surgical outcomes. miR-301a-3p was significantly reduced in the serum of patients who developed drug resistance in cases of refractory epilepsy (RE). Researchers concluded that miR-301a-3p could serve as a potential biomarker not only for mesial temporal lobe epilepsy, but also for predicting sudden death. Likewise, miR-654-3p may help predicting favourable surgical outcomes in mTLE-HS patients [33].
Altogether, several studies have outlined the role of exosome-derived miRNA, cfRNA, cfDNA, and tRNA in the early detection of epilepsy, seizure severity, and drug resistance. These studies are in the exploratory stages; until now, no biomarkers have been used in the clinical setting.
3.6. Stroke
Stroke is the second leading cause of death and a significant cause of disability worldwide [77]. There are two major types of strokes: acute ischemic stroke (AIS), which accounts for about 80% of cases and is caused by the interruption of cerebral blood flow, and haemorrhagic stroke, which represents the remaining 20% and is the consequence of a ruptured blood vessel [78].
Several studies have investigated the role of circRNAs in this entity. A study showed that circSCMH1 levels are diminished in AIS. Additionally, functional recovery was promoted in rodent and non-human primates by using rabies virus glycoprotein-circSCMH1-extracellular vesicles [79]. A separate study found that the expression levels of circTLK1 were significantly increased in brain tissue and plasma isolated from animal models of AIS. Moreover, it was observed that it improved long-term neurological deficits and significant reductions in infarct volumes and neuronal injury after knocking down circTLK1 in an animal model of AIS [80]. Furthermore, it was found that circCDC14A was significantly increased in lymphocytes and granulocytes of AIS patients [81]. In another study, the expression levels of circRNA_0001599 were found to be positively correlated with the National Institute of Health´s Stroke Scale Score (NIHSS) and infarct volumes [82].
These studies suggest that circRNAs can be used as biomarkers and therapeutic targets in AIS.
3.7. Traumatic Brain Injury
Traumatic Brain Injury (TBI) is one of the leading causes of long-term disability. It is characterized by an immediate primary mechanical injury followed by a secondary injury that can persist over time due to pathophysiological changes [83,84]. TBI can be classified into mild (mTBI) or severe (sTBI), urging molecular approaches to evaluate this entity in the emergency setting.
A recent study investigated cfDNA levels in patients with TBI. The total amount of cf-mtDNA (mtCOXIII, mtNADI) and cf-nuDNA (nuACTB and nuSIRT1) were analyzed using qPCR from serum samples of healthy controls and patients with mTBI and sTBI. Significantly higher levels of all four cfDNA markers were observed in patients with sTBI compared to healthy controls [34]. In another study, attenuated total reflectance (ATR)-Fourier transform infrared (FTIR) spectroscopy was used to analyse serum samples from mTBI patients and healthy controls. Machine learning algorithms were then developed using the data obtained to identify which patients with mTBI were most likely to present a positive CT scan. This approach presented very high sensitivity and specificity, distinguishing patients with mTBI from controls [85].
As aforementioned, most of the work has been focused on searching biomarkers for differentiating mTBI from sTBI by using several components of liquid biopsy, particularly cfDNA and cf-mtDNAs. The applicability of these selected biomarkers could be used in clinical settings as they present high sensitivity and specificity, although further validations are needed.
3.8. CNS tumors
Primary CNS tumors are highly heterogeneous and can be benign or malignant. Gliomas are the most frequently diagnosed among the latter, and glioblastoma multiforme (GBM) is the most aggressive form, particularly prevalent in adults [86]. These tumors are challenging to treat, with an overall 5-year survival rate of 35% [87], dropping to 5% for GBM [36]. Accordingly, a significant amount of research in liquid biopsy has been focused on GBM, including EVs, cell-free DNA, RNA, and CTCs.
EVs isolated from blood and CSF samples revealed that IDH1-R132H is a diagnostic biomarker of GBM. Further analysis revealed that higher concentrations of EVs containing this molecule are related to tumor recurrence in patients who underwent resection [35].
Liquid biopsy from CSF samples revealed that combinations of miRNAs, such as miR-2 and miR-15b, could distinguish GBM from primary CNS lymphoma, with high sensitivity and specificity. Furthermore, a combination of a signature of nine miRNAs was found to correlate with higher sensitivity and specificity to tumor volume [36]. Additionally, a panel of exosomal miRNAs could serve as predictive biomarkers for GBM patients, aiding in monitoring treatment response to chemotherapy and drug resistance [35].
ctDNA of some tumor suppressors, such as P53 and PTEN, can be detected in all recurrent gliomas, making ctDNA a valuable biomarker for predicting response to adjuvant therapies, providing real-time information about tumor progression and recurrence. In the same study, the authors found that mutations in ctDNA of RB1 and EGFR could be used as biomarkers for predicting the IDH-wildtype subtype of GBM and, consequently, the malignancy of the tumor [37].
Overall, several approaches have suggested a promising role of EVs, CTCs, and ctDNA in determining tumor progression, treatment effect, early diagnosis, and tumor recurrence.
3.9. Neuroinfectious diseases
Neuroinfectious diseases are pathological conditions that produce brain or spinal cord inflammation triggered by pathogenic microorganisms such as bacteria, viruses, parasites, and fungi. Current diagnostic methods rely on microbial examination of biological samples, particularly CSF. Examples include microscopic observation, Gram staining, cultures, pathogen-specific antibody reactions, and detection of pathogen nucleic acids by PCR, among others. However, these diagnostic procedures have some limitations, as some lack sensitivity and require time to arrive at a final diagnosis. As a result, there is an urgent need to find more accurate, fast, and comprehensive techniques to accurately diagnose such diseases [88].
For bacterial pathogens, such as M. tuberculosis, amplification of cfDNA IS6110 sequence in CSF of tuberculosis meningitis patients is more sensitive than some traditional diagnostic protocols. NGS cfDNA can be applied to other infections, such as C. canimorsus meningitis. Moreover, antibiotic exposure does not affect this diagnostic method [38].
Regarding viral CNS infections, determining cf-mtDNA levels in HIV-infected patients, with or without highly active antiretroviral therapy (HAART), may reflect the current status of CNS damage. In this sense, an inverse correlation between the levels of cf-mtDNA in CSF samples and proteins necessary for angiogenesis and iron metabolism was observed, suggesting that these molecular events are related to CNS damage [29].
Parasitic infections caused by T. solium, a causative agent of neurocysticercosis, can be detected in CSF and urine samples via cfDNA determination by amplifying the pTsol9-gene through PCR primers [39]. The same approach can be applied to P. falciparum, which causes cerebral malaria, by measuring total plasmatic cfDNA, both host cfDNA in response to the infection and parasite cfDNA [40].
Finally, NGS can be useful for pathogenic agent detection when performed on CSF samples in presumed or definitive cases of neuroinfectious diseases. Bacterial capture sequencing (BacCapSeq) and all vertebrate virus sequencing (VirCapSeq-VERT) show promising capacities for detecting various pathogens. However, the clinical application of these techniques still needs to be explored [89].
4. Conclusions and future directions
Liquid biopsy assesses components that are shed from cells into body fluids, and over the last decade, several sensitive technologies have emerged for detecting them. Previously considered as insignificant or waste material, these technologies have elucidated their functional roles in disease progression or as prognostic biomarkers. Liquid biopsy represents a sensitive and non-invasive approach for early diagnosis, identification of therapeutic targets, real-time disease surveillance and monitoring of treatment effects. Furthermore, this technique has already been employed as outcome measure in clinical trials, mostly in CNS tumors. However, further investigation is needed for other CNS conditions.
Despite the promising potential of non-invasive approaches in diagnosing, prognosis, and monitoring neurodegenerative diseases, the clinical application of these biomarkers still needs to be improved when compared to traditional diagnostic techniques, being essential to acknowledge the several inconveniences it faces. The number of participants in liquid biopsy studies is often insufficient, there is heterogeneity among participants, and many neurological diseases share common features often accompanied by neurodegeneration/neuronal damage. Consequently, many biomarkers encountered in liquid samples result in overlapping conclusions.
The future of liquid biopsy will depend on the identification and validation of genetic, epigenetic, and protein components through the standardization of protocols in larger multicentric cohorts. Additionally, it will be crucial to develop simplified tests that facilitate a seamless transition from the laboratory to clinical settings, ensuring cost-effectiveness and reducing the expertise required for implementation. This approach will facilitate the integration of liquid biopsies into the existing extensive clinical and radiological evaluations, making the process more accessible and efficient.
Author Contributions
All authors have contributed to the conception and design of the review. S.M., M.C.M.M., A.P., X.M. and M.C have contributed to the preparing the figure and table. All authors revised the manuscript for intellectual content.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
S.M was supported by the grant from Instituto Salud Carlos III (PI20/01697). The authors would like to express their gratitude to Dr. Josep Villena and Dr. Sumedha Sachar for their valuable scientific feedback to improve this review.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Loyfer, N.; Magenheim, J.; Peretz, A.; Cann, G.; Bredno, J.; Klochendler, A.; Fox-Fisher, I.; Shabi-Porat, S.; Hecht, M.; Pelet, T.; Moss, J.; et al. A DNA methylation atlas of normal human cell types. Nature 2023, 613, 355–364. [Google Scholar] [CrossRef]
- Kuang, Y.; Zheng, X.; Zhang, L.; Ai, X.; Venkataramani, V.; Kilic, E.; Hermann, D.M.; Majid, A.; Bähr, M.; Doeppner, T.R. Adipose-derived mesenchymal stem cells reduce autophagy in stroke mice by extracellular vesicle transfer of miR-25. J Extracell vesicles 2020, 10, e12024. [Google Scholar] [CrossRef]
- Kanninen, K.M.; Bister, N.; Koistinaho, J.; Malm, T. Exosomes as new diagnostic tools in CNS diseases. Biochim Biophys Acta 2016, 1862, 403–410. [Google Scholar] [CrossRef]
- Lai, C.P.; Breakefield, X.O. Role of exosomes/microvesicles in the nervous system and use in emerging therapies. Front Physiol. 2012, 3, 228. [Google Scholar] [CrossRef]
- Park, J.E.; Tan, H.S.; Datta, A.; Lai, R.C.; Zhang, H.; Meng, W.; Lim, S.K.; Sze, S.K. Hypoxic tumor cell modulates its microenvironment to enhance angiogenic and metastatic potential by secretion of proteins and exosomes. Mol Cell Proteomics 2010, 9, 1085–1099. [Google Scholar] [CrossRef]
- Verderio, C.; Cagnoli, C.; Bergami, M.; Francolini, M.; Schenk, U.; Colombo, A.; Riganti, L.; Frassoni, C.; Zuccaro, E.; Danglot, L.; et al. TI-VAMP/VAMP7 is the SNARE of secretory lysosomes contributing to ATP secretion from astrocytes. Biol Cell 2012, 104, 213–228. [Google Scholar] [CrossRef]
- Kumar, A.; Stoica, B.A.; Loane, D.J.; Yang, M.; Abulwerdi, G.; Khan, N.; Kumar, A.; Thom, S.R.; Faden, A.I. Microglial-derived microparticles mediate neuroinflammation after traumatic brain injury. J Neuroinflammation 2017, 14, 47. [Google Scholar] [CrossRef]
- Wang, Y.; Ma, Z.; Kan, P.; Zhang, B. The Diagnostic Value of Serum miRNA-221-3p, miRNA-382-5p, and miRNA-4271 in Ischemic Stroke. J Stroke Cerebrovasc Dis. 2017, 26, 1055–1060. [Google Scholar] [CrossRef]
- Vajpeyee, A.; Wijatmiko, T.; Vajpeyee, M.; Taywade, O. Cell free DNA: A Novel Predictor of Neurological Outcome after Intravenous Thrombolysis and/or Mechanical Thrombectomy in Acute Ischemic Stroke Patients. Neurointervention 2018, 13, 13–19. [Google Scholar] [CrossRef]
- Kayhanian, S.; Young, A.M.H.; Ewen, R.L.; Piper, R.J.; Guilfoyle, M.R.; Donnelly, J.; Fernandes, H.M.; Garnett, M.; Smielewski, P.; Czosnyka, M.; et al. Thresholds for identifying pathological intracranial pressure in paediatric traumatic brain injury. Sci Rep. 2019, 9, 3537. [Google Scholar] [CrossRef]
- Pessoa, L.S.; Heringer, M.; Ferrer, V.P. ctDNA as a cancer biomarker: A broad overview. Crit Rev Oncol Hematol. 2020, 155, 103109. [Google Scholar] [CrossRef]
- Lin, D.; Shen, L.; Luo, M.; Zhang, K.; Li, J.; Yang, Q.; Zhu, F.; Zhou, D.; Zheng, S.; Chen, Y.; et al. Circulating tumor cells: biology and clinical significance. Signal Transduct Target Ther. 2021, 6, 404. [Google Scholar] [CrossRef]
- Winek, K.; Lobentanzer, S.; Nadorp, B.; Dubnov, S.; Dames, C.; Jagdmann, S.; Moshitzky, G.; Hotter, B.; Meisel, C.; Greenberg, D.S.; et al. Transfer RNA fragments replace microRNA regulators of the cholinergic poststroke immune blockade. Proc Natl Acad Sci. 2020, 117, 32606–32616. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Liu, K.; Xiong, Y.; Zhao, C.; Liu, L. Increased expression of fragmented tRNA promoted neuronal necrosis. Cell Death Dis. 2021, 12, 823. [Google Scholar] [CrossRef]
- Li, M.L.; Wang, W.; Jin, Z.B. Circular RNAs in the Central Nervous System. Front Mol Biosci. 2021, 8, 629593. [Google Scholar] [CrossRef]
- Zhao, Y.; Alexandrov, P.N.; Jaber, V.; Lukiw, W.J. Deficiency in the Ubiquitin Conjugating Enzyme UBE2A in Alzheimer’s Disease (AD) is Linked to Deficits in a Natural Circular miRNA-7 Sponge (circRNA; ciRS-7). Genes (Basel) 2016, 7, 116. [Google Scholar] [CrossRef]
- Ghosal, S.; Das, S.; Sen, R.; Basak, P.; Chakrabarti, J. Circ2Traits: a comprehensive database for circular RNA potentially associated with disease and traits. Front Genet. 2013, 4, 283. [Google Scholar] [CrossRef] [PubMed]
- Hua, K.; Jin, J.; Zhang, H.; Zhao, B.; Wu, C.; Xu, H.; Fang, L. MicroRNA-7 inhibits proliferation, migration and invasion of thyroid papillary cancer cells via targeting CKS2. Int J Oncol. 2016, 49, 1531–1540. [Google Scholar] [CrossRef]
- Zhang, X.; Tang, X.; Liu, K.; Hamblin, M.H.; Yin, K.J. Long Noncoding RNA Malat1 Regulates Cerebrovascular Pathologies in Ischemic Stroke. J Neurosci Off J Soc Neurosci. 2017, 37, 1797–1806. [Google Scholar] [CrossRef]
- Liang, J.; Wang, Q.; Li, J.Q. , Guo, T.; Yu, D. Long non-coding RNA MEG3 promotes cerebral ischemia-reperfusion injury through increasing pyroptosis by targeting miR-485/AIM2 axis. Exp Neurol. 2020, 325, 113139. [Google Scholar] [CrossRef]
- Xu, X.; Zhuang, C.; Chen, L. Exosomal Long Non-Coding RNA Expression from Serum of Patients with Acute Minor Stroke. Neuropsychiatr Dis Treat. 2020, 16, 153–160. [Google Scholar] [CrossRef]
- Eren, E.; Leoutsakos, J.M.; Troncoso, J.; Lyketsos, C.G.; Oh, E.S.; Kapogiannis, D. Neuronal-Derived EV Biomarkers Track Cognitive Decline in Alzheimer’s Disease. Cells 2022, 11, 436. [Google Scholar] [CrossRef]
- Yan, Z.; Zhou, Z.; Wu, Q.; Chen, Z.B.; Koo, E.H.; Zhong, S. Presymptomatic Increase of an Extracellular RNA in Blood Plasma Associates with the Development of Alzheimer’s Disease. Curr Biol. 2020, 30, 1771–1782. [Google Scholar] [CrossRef]
- Ramaswamy, P.; Yadav, R.; Pal, P.K.; Christopher, R. Clinical Application of Circulating MicroRNAs in Parkinson’s Disease: The Challenges and Opportunities as Diagnostic Biomarker. Ann Indian Acad Neurol. 2020, 23, 84–97. [Google Scholar]
- Xie, S.; Niu, W.; Xu, F.; Wang, Y.; Hu, S.; Niu, C. Differential expression and significance of miRNAs in plasma extracellular vesicles of patients with Parkinson’s disease. Int J Neurosci. 2022, 132, 673–688. [Google Scholar] [CrossRef]
- Otake, K.; Kamiguchi, H.; Hirozane, Y. Identification of biomarkers for amyotrophic lateral sclerosis by comprehensive analysis of exosomal mRNAs in human cerebrospinal fluid. BMC Med Genomics 2019, 12, 7. [Google Scholar] [CrossRef]
- Joilin, G.; Gray, E.; Thompson, A.G.; Bobeva, Y.; Talbot, K.; Weishaupt, J.; Ludolph, A.; Malaspina, A.; Leigh, P.N.; Newbury, S.F.; et al. Identification of a potential non-coding RNA biomarker signature for amyotrophic lateral sclerosis. Brain Commun. 2020, 2, fcaa053. [Google Scholar] [CrossRef]
- Takahashi, I.; Yabe, I.; Hama, Y.; Uwatoko, H.; Shirai, S.; Matsushima, M.; Utsumi, J.; Sasaki, H.; et al. Evaluation of microRNAs in patients with sporadic amyotrophic lateral sclerosis by liquid biopsy as a disease biomarker (P1.9-027). Neurology 2019, 92, P1.9-027. [Google Scholar]
- Gaitsch, H.; Franklin, R.J.M.; Reich, D.S. Cell-free DNA-based liquid biopsies in neurology. Brain 2023, 146, 1758–1774. [Google Scholar] [CrossRef]
- Selmaj, I.; Cichalewska, M.; Namiecinska, M.; Galazka, G.; Horzelski, W.; Selmaj, K.W.; Mycko, M.P. Global exosome transcriptome profiling reveals biomarkers for multiple sclerosis. Ann Neurol. 2017, 81, 703–717. [Google Scholar] [CrossRef]
- Singh, J.; Deshpande, M.; Suhail, H.; Rattan, R.; Giri, S. Targeted Stage-Specific Inflammatory microRNA Profiling in Urine During Disease Progression in Experimental Autoimmune Encephalomyelitis: Markers of Disease Progression and Drug Response. J neuroimmune Pharmacol Off J Soc NeuroImmune Pharmacol. 2016, 11, 84–97. [Google Scholar] [CrossRef]
- Yan, S.; Zhang, H.; Xie, W.; Meng. ; F Zhang, K.; Jiang, Y.; Zhang, X.; Zhang, J. Altered microRNA profiles in plasma exosomes from mesial temporal lobe epilepsy with hippocampal sclerosis. Oncotarget 2017, 8, 4136–4146. [Google Scholar] [CrossRef]
- Whitlock, J.H.; Soelter, T.M.; Williams, A.S.; Hardigan, A.A.; Lasseigne, B.N. Liquid biopsies in epilepsy: biomarkers for etiology, diagnosis, prognosis, and therapeutics. Hum Cell 2022, 35, 15–22. [Google Scholar] [CrossRef]
- Marcatti, M.; Saada, J.; Okereke, I.; Wade, C.E.; Bossmann, S.H.; Motamedi, M.; Szczesny, B. Quantification of Circulating Cell Free Mitochondrial DNA in Extracellular Vesicles with PicoGreen™ in Liquid Biopsies: Fast Assessment of Disease/Trauma Severity. Cells 2021, 10, 819. [Google Scholar] [CrossRef]
- Sareen, H.; Garrett, C.; Lynch, D.; Powter, B.; Brungs, D.; Cooper, A.; Po, J.; Koh, E.S.; Vessey, J.Y.; McKechnie, S.; et al. The Role of Liquid Biopsies in Detecting Molecular Tumor Biomarkers in Brain Cancer Patients. Cancers (Basel) 2020, 12, 1831. [Google Scholar] [CrossRef]
- Bauman, M.M.J.; Bouchal, S.M.; Monie, D.D.; Aibaidula, A.; Singh, R.; Parney, I.F. Strategies, considerations, and recent advancements in the development of liquid biopsy for glioblastoma: a step towards individualized medicine in glioblastoma. Neurosurg Focus. 2022, 53, E14. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, C.; Li, M.; Shen, Y.; Feng, S.; Liu, J.; Li, F.; Hou, L.; Chen, Z.; Jiang, J.; et al. Applications of cerebrospinal fluid circulating tumor DNA in the diagnosis of gliomas. Jpn J Clin Oncol. 2020, 50, 325–332. [Google Scholar] [CrossRef]
- Asif, A.A.; Roy, M.; Tellier, B.R.; Ahmad, S. Capnocytophaga canimorsus meningitis diagnosed using next- generation sequencing of microbial cell-free DNA. IDCases 2021, 24, e01126. [Google Scholar] [CrossRef]
- Toribio, L.; Romano, M.; Scott, A.L.; Gonzales, I.; Saavedra, H.; Garcia, H.H.; Shiff, C. For The Cysticercosis Working Group In Peru. Detection of Taenia solium DNA in the Urine of Neurocysticercosis Patients. Am J Trop Med Hyg. 2019, 100, 327–329. [Google Scholar] [CrossRef]
- Vera, I.M.; Kessler, A.; Ting, L.M.; Harawa, V.; Keller, T.; Allen, D.; Njie, M.; Moss, M.; Soko, M.; Ahmadu, A.; et al. Plasma cell-free DNA predicts pediatric cerebral malaria severity. JCI Insight. 2020, 5, e136279. [Google Scholar] [CrossRef]
- Scheltens, P.; Blennow, K.; Breteler, M.M.B.; de Strooper, B.; Frisoni, G.B.; Salloway, S.; Van der Flier, W.M. Alzheimer’s disease. Lancet (London, England) 2016, 388, 505–517. [Google Scholar] [CrossRef]
- Soria Lopez, J.A.; González, H.M.; Léger, G.C. Alzheimer’s disease. Handb Clin Neurol. 2019, 167, 231–255. [Google Scholar]
- Chen, L.; Shen, Q.; Xu, S.; Yu, H.; Pei, S.; Zhang, Y.; He, X.; Wang, Q.; Li, D. 5-Hydroxymethylcytosine Signatures in Circulating Cell-Free DNA as Diagnostic Biomarkers for Late-Onset Alzheimer’s Disease. J Alzheimers Dis. 2022, 85, 573–585. [Google Scholar] [CrossRef]
- Dube, U.; Del-Aguila, J.L.; Li, Z.; Budde, J.P.; Jiang, S.; Hsu, S.; Ibanez, L.; Fernandez, M.V.; Farias, F.; Norton, J.; et al. An atlas of cortical circular RNA expression in Alzheimer disease brains demonstrates clinical and pathological associations. Nat Neurosci. 2019, 22, 1903–1912. [Google Scholar] [CrossRef]
- Bigarré, I.M.; Trombetta, B.A.; Guo, Y.J.; Arnold, S.E.; Carlyle, B.C. IGF2R circular RNA hsa_circ_0131235 expressions in the middle temporal cortex is associated with AD pathology. Brain Behav. 2021, 11, e02048. [Google Scholar] [CrossRef]
- Kuscu, C.; Kumar, P.; Kiran, M.; Su, Z.; Malik, A.; Dutta, A. tRNA fragments (tRFs) guide Ago to regulate gene expression post-transcriptionally in a Dicer-independent manner. RNA 2018, 24, 1093–1105. [Google Scholar] [CrossRef]
- Wu, W.; Lee, I.; Spratt, H.; Fang, X.; Bao, X. tRNA-Derived Fragments in Alzheimer’s Disease: Implications for New Disease Biomarkers and Neuropathological Mechanisms. J Alzheimers Dis. 2021, 79, 793–806. [Google Scholar] [CrossRef]
- Poewe, W.; Seppi, K.; Tanner, C.M.; Halliday, G.M.; Brundin, P.; Volkmann, J.; Schrag, A.E.; Lang, A.E. Parkinson disease. Nat Rev Dis Prim. 2017, 3, 17013. [Google Scholar] [CrossRef]
- Tolosa, E.; Garrido, A.; Scholz, S.W.; Poewe, W. Challenges in the diagnosis of Parkinson’s disease. Lancet Neurol. 2021, 20, 385–397. [Google Scholar] [CrossRef]
- Liu, J.; Chen, Y.; Pei, F.; Zeng, C.; Yao, Y.; Liao, W.; Zhao, Z. Extracellular Vesicles in Liquid Biopsies: Potential for Disease Diagnosis. Biomed Res Int. 2021, 2021, 6611244. [Google Scholar] [CrossRef]
- Burgos, K.; Malenica, I.; Metpally, R.; Courtright, A. , Rakela, B.; Beach, T.; Shill, H.; Adler, C.; Sabbagh, M.; Villa, S.; et al. Profiles of extracellular miRNA in cerebrospinal fluid and serum from patients with Alzheimer’s and Parkinson’s diseases correlate with disease status and features of pathology. PLoS One 2014, 9. [Google Scholar] [CrossRef]
- Pawlik, P.; Błochowiak, K. The Role of Salivary Biomarkers in the Early Diagnosis of Alzheimer’s Disease and Parkinson’s Disease. Diagnostics (Basel, Switzerland) 2021, 11. [Google Scholar] [CrossRef]
- Bai, J.; Zheng, Y.; Yu, Y. Urinary kynurenine as a biomarker for Parkinson’s disease. Neurol Sci. 2021, 42, 697–703. [Google Scholar] [CrossRef]
- Pyle, A.; Brennan, R.; Kurzawa-Akanbi, M.; Yarnall, A.; Thouin, A.; Mollenhauer, B.; Burn, D.; Chinnery, P.F.; Hudson, G. Reduced cerebrospinal fluid mitochondrial DNA is a biomarker for early-stage Parkinson’s disease. Ann Neurol. 2015, 78, 1000–1004. [Google Scholar] [CrossRef]
- Hardiman, O.; Al-Chalabi, A.; Chio, A.; Corr, E.M.; Logroscino, G.; Robberecht, W.; Shaw, P.J; Simmons, Z.; van den Berg, L.H.; et al. Amyotrophic lateral sclerosis. Nat Rev Dis Prim. 2017, 3, 17071. [Google Scholar] [CrossRef]
- Ince, P.G.; Highley, J.R.; Kirby, J.; Wharton, S.B.; Takahashi, H.; Strong, M.J.; Shaw, P.J.; et al. Molecular pathology and genetic advances in amyotrophic lateral sclerosis: an emerging molecular pathway and the significance of glial pathology. Acta Neuropathol. 2011, 122, 657–671. [Google Scholar] [CrossRef]
- Philips, T.; Robberecht, W. Neuroinflammation in amyotrophic lateral sclerosis: role of glial activation in motor neuron disease. Lancet Neurol. 2011, 10, 253–263. [Google Scholar] [CrossRef]
- Sjoqvist, S.; Otake, K. A pilot study using proximity extension assay of cerebrospinal fluid and its extracellular vesicles identifies novel amyotrophic lateral sclerosis biomarker candidates. Biochem Biophys Res Commun. 2022, 613, 166–173. [Google Scholar] [CrossRef]
- Mendioroz, M.; Martínez-Merino, L.; Blanco-Luquin, I.; Urdánoz, A.; Roldán, M.; Jericó, I. Liquid biopsy: a new source of candidate biomarkers in amyotrophic lateral sclerosis. Ann Clin Transl Neurol. 2018, 5, 763–768. [Google Scholar] [CrossRef]
- Joilin, G.; Gray, E.; Thompson, A.G.; Bobeva, Y.; Talbot, K.; Weishaupt, J.; Ludolph, A.; Malaspina, A.; Leigh, P.N.; Newbury, S.F.; et al. Identification of a potential non-coding RNA biomarker signature for amyotrophic lateral sclerosis. Brain Commun. 2020, 2, fcaa053. [Google Scholar] [CrossRef]
- Tur, C.; Carbonell-Mirabent, P.; Cobo-Calvo, Á.; Otero-Romero, S.; Arrambide, G.; Midaglia, L.; Castilló, J.; Vidal-Jordana, Á.; Rodríguez-Acevedo, B.; Zabalza, A.; et al. Association of Early Progression Independent of Relapse Activity With Long-term Disability After a First Demyelinating Event in Multiple Sclerosis. JAMA Neurol. 2023, 80, 151–160. [Google Scholar] [CrossRef]
- Iparraguirre, L.; Olaverri, D.; Blasco, T.; Sepúlveda, L.; Castillo-Triviño, T.; Espiño, M.; Costa-Frossard, L.; Prada, Á.; Villar, L.M.; Otaegui, D.; et al. Whole-Transcriptome Analysis in Peripheral Blood Mononuclear Cells from Patients with Lipid-Specific Oligoclonal IgM Band Characterization Reveals Two Circular RNAs and Two Linear RNAs as Biomarkers of Highly Active Disease. Biomedicines 2020, 8, 540. [Google Scholar] [CrossRef]
- Mrad, M.F.; Saba, E.S.; Nakib, L.; Khoury, S.J. Exosomes from Subjects with Multiple Sclerosis Express EBV-Derived Proteins and Activate Monocyte-Derived Macrophages. Neurol Neuroimmunol neuroinflammation 2021, 8, e1004. [Google Scholar] [CrossRef]
- D’Apolito, M.; Rispoli, M.G.; Ajdinaj, P.; Digiovanni, A.; Tomassini, V.; Gentile, L.; De Luca, G. Progressive multifocal leukoencephalopathy or severe multiple sclerosis relapse following COVID-19 vaccine: a diagnostic challenge. Neurol Sci. 2023, 44, 1141–1146. [Google Scholar] [CrossRef]
- Hatchwell, E.; Smith, E.B.; Jalilzadeh, S.; Bruno, C.D.; Taoufik, Y.; Hendel-Chavez, H.; Liblau, R.; Brassat. D.; Martin-Blondel, G.; Wiendl, H.; et al. Progressive multifocal leukoencephalopathy genetic risk variants for pharmacovigilance of immunosuppressant therapies. Front Neurol. 2022, 13, 1016377. [Google Scholar] [CrossRef]
- Ngouth, N.; Monaco, M.C.; Walker, L.; Corey, S.; Ikpeama, I.; Fahle, G.; Cortese, I.; Das, S.; Jacobson, S. Comparison of qPCR with ddPCR for the Quantification of JC Polyomavirus in CSF from Patients with Progressive Multifocal Leukoencephalopathy. Viruses 2022, 14, 1246. [Google Scholar] [CrossRef]
- Beghi, E. The Epidemiology of Epilepsy. Neuroepidemiology 2020, 54, 185–191. [Google Scholar] [CrossRef]
- Thijs, R.D. , Surges, R.; O’Brien, T.J.; Sander, J.W. Epilepsy in adults. Lancet (London, England) 2019, 393, 689–701. [Google Scholar] [CrossRef]
- Moshé, S.L.; Perucca, E.; Ryvlin, P.; Tomson, T. Epilepsy: new advances. Lancet (London, England) 2015, 385, 884–898. [Google Scholar] [CrossRef]
- Hashemian, F.; Ghafouri-Fard, S.; Arsang-Jang, S.; Mirzajani, S.; Fallah, H.; Mehvari Habibabadi, J.; Sayad, A.; Taheri, M. Epilepsy Is Associated with Dysregulation of Long Non-coding RNAs in the Peripheral Blood. Front Mol Biosci. 2019, 6, 113. [Google Scholar] [CrossRef]
- Martins-Ferreira, R.; Leal, B.G.; Costa, P.P. The Potential of Circulating Cell-Free DNA Methylation as an Epilepsy Biomarker. Front Cell Neurosci. 2022, 16, 852151. [Google Scholar] [CrossRef]
- Alapirtti, T.; Jylhävä, J.; Raitanen, J.; Mäkinen, R.; Peltola, J.; Hurme, M.A.; Liimatainen, S. The concentration of cell-free DNA in video-EEG patients is dependent on the epilepsy syndrome and duration of epilepsy. Neurol Res. 2016, 38, 45–50. [Google Scholar] [CrossRef]
- Upadhya, D.; Shetty, A.K. Promise of extracellular vesicles for diagnosis and treatment of epilepsy. Epilepsy Behav. 2021, 121, 106499. [Google Scholar] [CrossRef]
- Cai, X.; Long, L.; Zeng, C.; Ni, G.; Meng, Y.; Guo, Q.; Chen, Z.; Li, Z.; et al. LncRNA ILF3-AS1 mediated the occurrence of epilepsy through suppressing hippocampal miR-212 expression. Aging (Albany NY) 2020, 12, 8413–8422. [Google Scholar] [CrossRef] [PubMed]
- Cukovic, D.; Bagla, S.; Ukasik, D.; Stemmer, P.M.; Jena, B.P.; Naik, A.R.; Sood, S.; Asano, E.; Luat, A.; Chugani, D.C.; et al. Exosomes in Epilepsy of Tuberous Sclerosis Complex: Carriers of Pro-Inflammatory MicroRNAs. Non-coding RNA 2021, 7, 40. [Google Scholar] [CrossRef] [PubMed]
- Mayo, S.; Gómez-Manjón, I.; Fernández-Martínez, F.J.; Camacho, A.; Martínez, F.; Benito-León, J. CfDNA Measurement as a Diagnostic Tool for the Detection of Brain Somatic Mutations in Refractory Epilepsy. Int J Mol Sci. 2022, 23, 4879. [Google Scholar] [CrossRef] [PubMed]
- Katan, M.; Luft, A. Global Burden of Stroke. Semin Neurol. 2018, 38, 208–211. [Google Scholar] [CrossRef]
- Feng, Y.S.; Tan, Z.X.; Wang, M.M.; Xing, Y.; Dong, F.; Zhang, F. Inhibition of NLRP3 Inflammasome: A Prospective Target for the Treatment of Ischemic Stroke. Front Cell Neurosci. 2020, 14, 155. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Han, B.; Zhang, Z.; Wang, S.; Bai, Y.; Zhang, Y.; Tang, Y.; Du, L.; Xu, L.; Wu, F.; et al. Extracellular Vesicle-Mediated Delivery of Circular RNA SCMH1 Promotes Functional Recovery in Rodent and Nonhuman Primate Ischemic Stroke Models. Circulation 2020, 142, 556–574. [Google Scholar] [CrossRef]
- Wu, F.; Han, B.; Wu, S.; Yang, L.; Leng, S.; Li, M.; Liao, J.; Wang, G.; Ye, Q.; Zhang, Y.; et al. Circular RNA TLK1 Aggravates Neuronal Injury and Neurological Deficits after Ischemic Stroke via miR-335-3p/TIPARP. J Neurosci. 2019, 39, 7369–7393. [Google Scholar] [CrossRef]
- Zuo, L.; Zhang, L.; Zu, J.; Wang, Z.; Han, B.; Chen, B.; Cheng, M.; Ju, M.; Li, M.; Shu, G.; et al. Circulating Circular RNAs as Biomarkers for the Diagnosis and Prediction of Outcomes in Acute Ischemic Stroke. Stroke 2020, 51, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Hu, W.; Deng, F.; Chen, S.; Zhu, P.; Wang, M.; Chen, X; Wang, Y. ; Hu, X.; Zhao, B.; et al. Identification of Circular RNA hsa_circ_0001599 as a Novel Biomarker for Large-Artery Atherosclerotic Stroke. DNA Cell Biol. 2021, 40, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.A.; Bell, J.M.; Breiding, M.J.; Xu, L. Traumatic Brain Injury-Related Emergency Department Visits, Hospitalizations, and Deaths—United States, 2007 and 2013. Morb Mortal Wkly report Surveill Summ (Washington, DC 2002) 2017, 66, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.Y.; Lee, A.Y.W. Traumatic Brain Injuries: Pathophysiology and Potential Therapeutic Targets. Front Cell Neurosci. 2019, 13, 528. [Google Scholar] [CrossRef] [PubMed]
- Theakstone, A.G.; Brennan, P.M.; Ashton, K.; Czeiter, E.; Jenkinson, M.D.; Syed, K.; Reed, M.J.; Baker, M.J.; CENTER-TBI Participants and Investigators. Vibrational Spectroscopy for the Triage of Traumatic Brain Injury Computed Tomography Priority and Hospital Admissions. J Neurotrauma. 2022, 39, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Bunda, S.; Zuccato, J.A.; Voisin, M.R.; Wang, J.Z.; Nassiri, F.; Patil, V.; Mansouri, S.; Zadeh, G. Liquid Biomarkers for Improved Diagnosis and Classification of CNS Tumors. Int J Mol Sci. 2021, 22, 4548. [Google Scholar] [CrossRef] [PubMed]
- Lapointe, S.; Perry, A.; Butowski, N.A. Primary brain tumours in adults. Lancet 2018, 392, 432–446. [Google Scholar] [CrossRef]
- Guo, Y.; Yang, Y.; Xu, M.; Shi, G.; Zhou, J.; Zhang, J.; Li, H. Trends and Developments in the Detection of Pathogens in Central Nervous System Infections: A Bibliometric Study. Front Cell Infect Microbiol. 2022, 12, 856845. [Google Scholar] [CrossRef]
- Boruah, A.; Kroopnick, A.; Kim, C.; Thakkar, R.; Dugue, R.; Harrigan, E.; Lipkin, I.; Mishra, N.; Thakur, K. Application of Metagenomic Next-Generation Sequencing in the Diagnosis of Neuroinfectious Diseases (P2-9.004). Neurology 2022, 98, 2866. [Google Scholar]
Figure 1.
Overview of liquid biopsy. Figure showing brain-periphery communication. CNS cell populations, including neurons, astrocytes and glial cells, release the principal elements of liquid biopsy components, including extracellular vesicles (micro-vesicles, exosomes and apoptotic bodies), transfer-RNA (t-RNA), circular RNA, and cell-free (cfDNA) within the CNS. The most studied component of liquid biopsy constitutes exosomes. It contains functional miRNA and small RNA. A process of membrane vesicle trafficking performs the transfer of exosomes from the donor cell to the recipient cells. t-RNA is mainly comprised of less than 90 nucleotides. tRNA-derived RNA fragments (tRFs) act as essential mediators of CNS and immune communications via tRFs. Circular RNA (circRNA) is a single-stranded RNA, primarily present in circular form. Another component of liquid biopsy includes cell-free DNA (cfDNA), composed of double helix DNA fragments released from CNS cells/nucleated cells and circulating freely in the bloodstream. Cell-free mitochondrial (cf-mtDNA) is a double stranded fragment produced by dysfunctional mitochondria. Circulating tumor DNA (ctDNA) is produced by tumor cells, because of their half-life, these are real time indicators of disease status in the body.
Figure 1.
Overview of liquid biopsy. Figure showing brain-periphery communication. CNS cell populations, including neurons, astrocytes and glial cells, release the principal elements of liquid biopsy components, including extracellular vesicles (micro-vesicles, exosomes and apoptotic bodies), transfer-RNA (t-RNA), circular RNA, and cell-free (cfDNA) within the CNS. The most studied component of liquid biopsy constitutes exosomes. It contains functional miRNA and small RNA. A process of membrane vesicle trafficking performs the transfer of exosomes from the donor cell to the recipient cells. t-RNA is mainly comprised of less than 90 nucleotides. tRNA-derived RNA fragments (tRFs) act as essential mediators of CNS and immune communications via tRFs. Circular RNA (circRNA) is a single-stranded RNA, primarily present in circular form. Another component of liquid biopsy includes cell-free DNA (cfDNA), composed of double helix DNA fragments released from CNS cells/nucleated cells and circulating freely in the bloodstream. Cell-free mitochondrial (cf-mtDNA) is a double stranded fragment produced by dysfunctional mitochondria. Circulating tumor DNA (ctDNA) is produced by tumor cells, because of their half-life, these are real time indicators of disease status in the body.
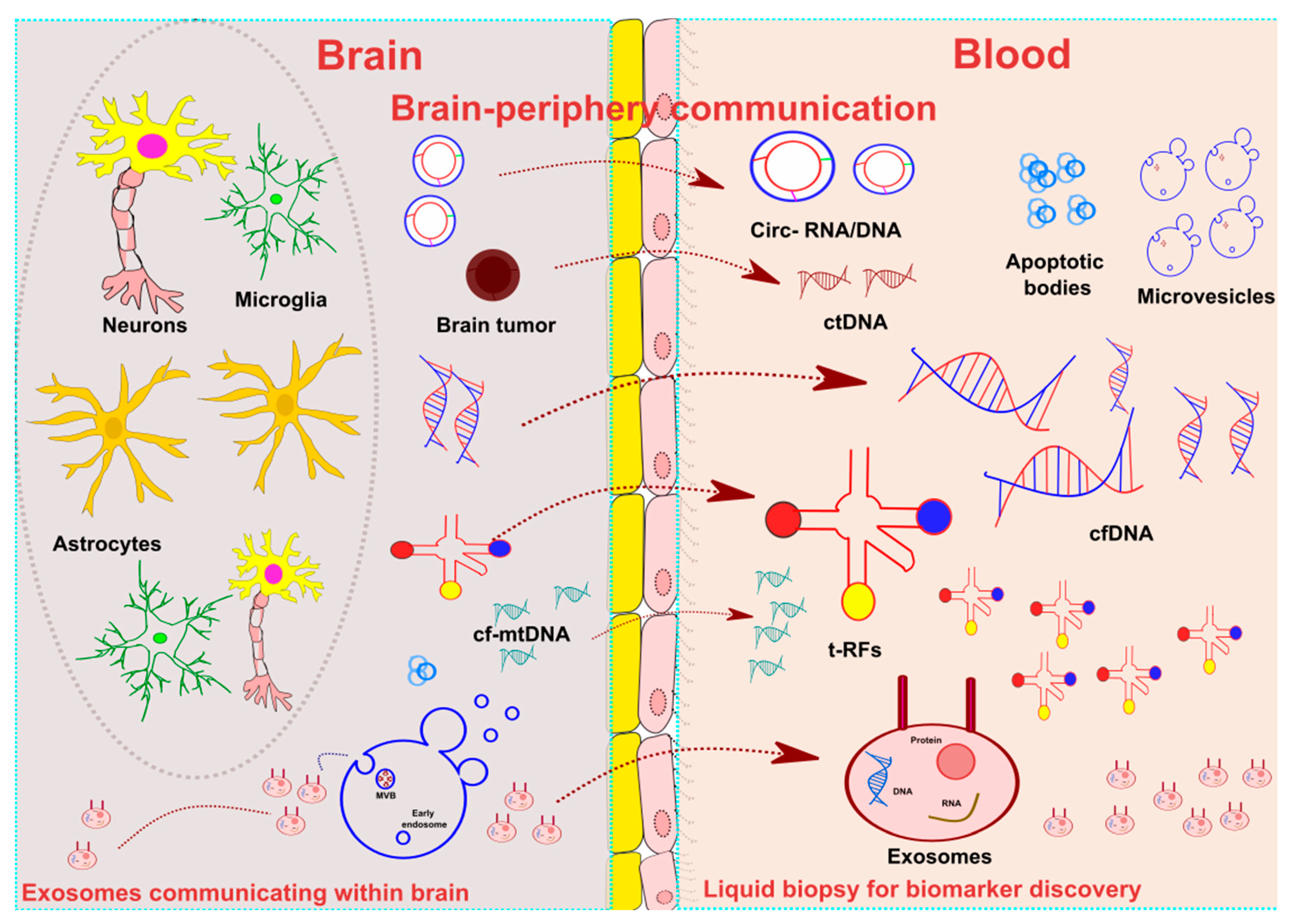

Table 1.
Selected liquid biopsy biomarkers in neurological diseases.
| Biomarker | Source | Main Findings |
| Alzheimer’s Disease | ||
| ↑ Aβ42 | NDEVs |
|
| PHGDH’s exRNA | Plasma |
|
| Parkinson’s Disease | ||
|
↓miR-19b, miR-29c and miR-133b ↑miR-132 and miR-331-5p |
Blood, CSF and exosomes |
|
|
↑ hsa-miR-30c-2-3p ↓hsa-miR-15b-5p, hsa-miR-138-5p, hsa-miR-106b-3p |
Plasma EVs |
|
| Amyotrophic lateral sclerosis | ||
| CUEDC2 exosomal mRNA | CSF EVs |
|
| hsa-miR-16-5p, hsa-piR-33151 and TRV-AAC4-1.1 | Serum |
|
|
↓ hsa-miR-4299 ↑ hsa-miR-4649-5p |
Plasma |
|
| ↓ hsa-miR-663b and has-miR-4258 | CSF |
|
| Biomarker | Source | Main findings |
| Multiple sclerosis | ||
| EBV-derived cfDNA | CSF/ plasma |
|
| ↓ hsa-miR-196b-5p, hsa-miR-532-5p, hsa-miR-122-5p and hsa-miR-301a-3p | Serum |
|
| ↑ miR-155-5p | CSF/plasma/ urine exosomes |
|
| Epilepsy | ||
| miR-8071 | Plasma exosomes |
|
| miR-654-3p | Blood |
|
| Stroke | ||
| circRNA_0001599 | Plasma |
|
| Traumatic brain injury (TBI) | ||
| ↑ ccf-DNA: mtDNA (mtCOXIII, mtNADI) and nuDNA (nuACTB, nuSIRT1) | Serum |
|
| Biomarker | Source | Main findings |
| CNS tumours | ||
| IDH1-R132H | Blood/ CSF EVs |
|
| miR-2 and miR-15b | CSF |
|
| Tumour suppressor genes-derived ctDNA (TP53 and PTEN). | Blood |
|
| Mutation in RB1 and EGFR ctDNA |
|
|
|
Neuroinfectious diseases Bacterial meningitis |
||
| Mycobacterium tuberculosis cfDNA (IS6110 sequence) | CSF |
|
| Capnocytophaga canimorsus meningitis cfDNA | Blood |
|
| Neurocysticercosis (infection by Taenia solium) | ||
| cf-DNA: pTsol19-gene amplification | Urine/CSF |
|
| Cerebral malaria (infection by Plasmodium falciparum) | ||
| Total cf-DNA: host cfDNA (in response to infection) and parasite cf-DNA | Plasma |
|
Aβ42 = amyloid-β 42; AD = Alzheimer disease; ALS = amyotrophic lateral sclerosis; cfDNA = cell-free DNA; CNS = central nervous system; CSF = cerebrospinal fluid; EBV = Epstein-barr virus; EVs = extracellular vesicles; exRNA = extracellular RNA; GBM = glioblastoma; IDH = isocitrate dehydrogenase; miR = microRNA; MS = multiple sclerosis; PD = Parkinson disease; RRMS = relapsing-remitting multiple sclerosis; TBI = traumatic brain injury.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



