
Submitted:
27 June 2023
Posted:
28 June 2023
You are already at the latest version
Abstract
Fish tissue samples from 203 adult individuals were collected in the main ports and markets of the Pacific coast of Panama. Molecular identification based on cytochrome oxidase I gene segment of all species was verified GENBANK reference sequences. 34 species from 14 families (Ariidae, Lutjanidae, Caranjidae, Scianidae, Centropomidae, Serranidae, Scombridae, Sphyraenidae, Haemulidae, Gerreidae, Stromotidae, Lobotidae, Malacanthidae, Mugilidae) were identified at species molecular level from 164 sequences. Also, three Caribbean species were molecularly identified among the analyzed samples (Mycteroperca xenarcha, Paralonchurus brasilensis and Lobotes surinamensis). Species diversity was slightly higher in the Gulf of Panama than in the Gulf of Chiriquí. For species with 5 or more individual sequences, genetic diversity and genetic connectivity parameters such as: total number of haplotypes (H), haplotype diversity (Hd), and nucleotide diversity (π) were calculated. Overall, pelagic-migratory species showed higher values of genetic diversity than coastal and estuarine species with some exceptions. Connectivity between Gulf areas was compared using values of genetic distances and genetic differentiation (Fst). High level of connectivity observed between Gulf of Chiriqui and Gulf of Montijo indicates the existence of a single stock in that area for the following species: Scomberomorus sierra, Caranx caninus and Lutjanus guttatus. Demographic history of the most common species was examined using Tajima’s D values suggesting population expansion for two snapper species, L. peru and L. argentiventris, having significant and higher values. Another important contribution from this research was the production of primers and dual labeled probes for environmental DNA detection using qPCR for the five most abundant species (spotted rose snapper, yellow snapper, green jack, Pacific crevalle jack and the Pacific sierra fish). These markers represent a new set of tools for environmental DNA (eDNA) detection and molecular traceability of three commercially important fish species along the supply chain including landing sites and markets of the main fishery areas.
Keywords:
COI
; teleostei
; nucleotide diversity
; haplotypic diversity
; qPCR
; demographic history
; environmental DNA
1. Introduction
The advent of genetic barcode has improved the capacity to resolve the miscalling of species; detect seafood and product fraud; minimize trafficking of endangered and CITES (International Convention for Endangered Species Trade) species; and reduce illegal, unregulated and unreported fishing (IUU) [1,2]. In addition, barcode has allowed the identification and discovery of new species, separation of cryptic species, species complexes, stock characterization, detection of molecular hybrids and understanding migrations and connectivity patterns[3,4]. In fishes, the use of barcode began in 2003 [5] and by 2009 more than 5,000 fish species were characterized by this method [6]. Considering that Osteichthyes (boney fishes) are the most diverse vertebrate taxa with more than 28,000 species reported [7,8], it is evident that scientists still have some important work to do.
Panamanian fish fauna is highly diverse and includes 1,412 species, of which 1,240 are marine species [9,10]. Approximately, 814 of these species have been reported in the Pacific coast of Panama and represents 60% of the fish diversity for the country, making it the most diverse vertebrate taxa [7]. In a recent review, a total of 223 commercial fish species were reported for the Pacific of Panama, corresponding to 183 bony and 70 cartilaginous fish [10]. These species are locally commercialized or exported as fish products (fillets, fins, and whole fish). However, the final destiny of some of the most commercially important fish species (i.e., snappers, drummers, pacific sierra, groupers, queen corvina, dolphin fish, tunas, jacks and sharks) and their derived products is currently unknown or partially understood due to taxonomic identification mistakes and the lack of species-specific tariff codes that should be based on a binomial nomenclature species (BNS) system that is the key for the establishment of species specific traceability program. In some cases, lack of this BNS is usually magnified by clustering three or more species under the same tariff code. For example, Pacific Panama has 10 species of lutjanids that are clustered under snappers as unique tariff code whereas 37 species of shark and 22 species of rays are clustered under the designation “shark” as tariff code [11,12]. These taxonomic identification mistakes lead to an improbable determination of which species are certainly exploited, overexploited, and exported. Even in food products like “ceviche” (a mix of raw fish cooked with lemon) which is popular among local consumers; contains species that are currently unlisted, invasive, CITES or under any of the 12 IUCN categories like sharks, rays and billfishes[13].
Traceability is a records-based system defined as the ability to trace the history, origin location and application of a traceable resource unit (TRU) which is a species or fishery product along the food supply chain using physical (electronic tags), chemical (isotopes) or molecular methods (i.e., DNA barcode or DNA fingerprint) [14,15]. In the 21st Century food safety, transparency and product quality control is a competitive advantage in a globalized market that relies on traceability [16,17]. In addition, transparency in fisheries is considered one of the main challenges that is currently facing the fishery industry [18].
The top 21 countries for food production and consumption participating in the Organization for Economic Co-Operation and Development (OECD) were ranked based on the nature or scope of their mandatory traceability regulations. Results showed that mandatory legislation for food and feed is only established in EU and Pan EU countries. Australia, New Zealand, Canada, Japan, Brazil and the United States have an overall world score ranking of “average”. For Russia insufficient data were received and China was ranked as poor. Interestingly, Costa Rica was recently included in the OECD without having a national traceability system for fish and fish products. However, no information or the species included in this traceability system is available.
The implementation of a traceability system in Latin America for fish and fishery products is currently in an early development phase or yet to be implemented [19]. Only countries like Peru, Chile and Ecuador have implemented a traceability model for some fishery products [20]. Nonetheless, these traceability programs are partially applied to some species or taxa, or they are only utilized for importation but not for domestic commercialization [21]. When traceability is applied, only physical traceability methods (labeling the products) are employed, and no chemical or molecular traceability methods are currently in use for the public sector [22,23,24]. In fact, for the most common commercial species listed in Central America and the Eastern Pacific from Mexico to Ecuador, stock composition and traditional population dynamics studies have been uniquely conducted in a handful of species e.g., red snapper, Lutjanus peru [25] ; spotted rose snapper, Lutjanus guttatus [26] , green jack, Caranx caballus [27]., Pacific sierra, Scomberomorus sierra [28]., dolphin fish, Coryphaena hippurus [29]., Wahoo, Acanthocybium solandri and the yellowfin Tuna, Thunnus albacares [30]. Likewise, genetic data for the species listed just before are only available for the Mexican populations of spotted rose snapper and red snapper [31,32], Pacific sierra[33] Wahoo [34] and the main tuna species [30]. On the other hand, molecular identification methods have been employed for traceability along the commercial chain and for mislabeling purposes in Caribbean populations of Lutjanus campechanus and Lutjanus purpureus [35,36].
Previous populational and taxonomic studies focused on the Pacific Panama fish fauna have been conducted in specific areas of Panama such as Gulf of Montijo[37,38] ., Coiba National Park[39]., Bahía Honda, Gulf of Chiriquí [40] and Las Perlas Archipelago, Gulf of Panama [41] focusing mainly in taxonomic species diversity. Recently, the first taxonomic list of commercial species for the Pacific reported a total of 183 teleost species [10]. This work from Garces 2021 listed the four most diverse commercial fish families that are consistent with our results: Carangidae, Scianidae, Lutjanidae, and Haemulidae. Other Pacific side fish studies that have used also traditional taxonomic methods reported 28 commercial species belonging to the following families: Scianidae, Lutjanidae, Mugilidae, Scombridae and Serranidae[42]. In addition, fish community descriptions performed with multiple traditional collection techniques have been conducted for Coiba National Park, showing 27 commercial species within the families: Carangidae, Lutjanidae and Haemulidae out of 156 collected fish species including reef and non-commercial species [39].
The main objective of this research is to provide the first DNA barcode list of Pacific Panama commercial fish. Also, to report the first haplotypes as well as the first values of genetic diversity, genetic connectivity and demographic history for species collected in the largest fishery areas of Pacific Panama (Gulf of Panama and Gulf of Chiriqui) and finally to provide potential markers for the most common species. Thus, this work represents the first attempt at providing a molecular data base (GeneBank accession numbers) of Panamanian commercial fisheries, i.e., fish species and fishery products detected molecularly. This information is the first step to for the establishment of a future fish traceability program in the country based on a BNS and supported by molecular data. These data will be key not only for traceability of commercial species but also for future genetic stock assessment, the establishment of conservation units and sustainable fishery programs. In addition, having a strong traceability system will strengthen the Hazard Analysis and Critical Control Point (HACCP) system of the country and minimize the impact of (IUU) fishing in the transparency of our fisheries [43]. Furthermore, this research contributes with the first species specific primers and probes designed in silico for environmental DNA (eDNA) detection and traceability of some of the most common commercial fish species reported in Panama waters. The production of molecular markers for eDNA detection of commercial species will be the base for a future molecular traceability pilot program in Panama with the potential to become a model for Latin-America and the Greater Caribbean regions, contributing to conservation, sustainable fisheries and minimizing mislabeling fraud through the supply chain and the importation and exportation process.
2. Materials and methods
2.1. Sample collection on local markets and fish gathering centers.
Main fishery landing ports of the Pacific Panama (Coquira, Pedregal and Mutis) and the biggest fish gathering centers of the Pacific side of the Country (Mercado del Marisco-San Felipe Market, Mercado de Anton, Mercado de Santiago and Isla Verde-Golfo de Montijo) were visited (Figure 1). Additionally, red snapper samples from Pixvae area, located within the buffer zone of Coiba Island National Park (largest marine park in Central America and UNESCO Natural Heritage Site), were also analyzed. At each collection site, fin clips and muscle tissue of the main fish families reported as commercial for the Pacific Panama were collected and storage in vials containing ethanol at 95%. Each sample was labeled and recorded using the species common name, possible fishery zone (Gulf) and landing site name.
2.2. DNA extraction and pcr amplification
A total of 203 fish tissue samples were collected. DNA was extracted from all these samples by different methods including the use of DNA easy kit for blood and tissue (QIAGEN, Inc., Valencia, California) as well as phenol-chlorophorm method. DNA was quantified using a nanodrop (Thermo Scientific, https://www.thermofisher.com/pa/en/home.html). PCR reactions were conducted with 1µl of each primer using fish universal primers C_FishF1 and C_FishR1[44]., 8.5 µl of molecular grade water, 2µl of template DNA and 12.5 µl of 2X master mix reagent (QIAGEN, Inc., Valencia, California). All PCR reactions were conducted for 35 cycles at an annealing temperature (Ta) of 55 ⁰C [44] .
2.3. Sequence analysis
PCR products were visualized in 2% agarose gels. Positive products were cleaned and sequenced by Macrogen Corp (Maryland, USA). Sequences were verified for size and checked for quality using Geneious Prime [45]. High quality sequences were trimmed, edited to a size of 614 bp and aligned for final analysis in Geneious Prime and DnaSP version 6.1. Edited sequences were imported to other programs for further genetic diversity and connectivity analysis as Fasta files. GenBank accession numbers for all obtained sequences were generated through Sequin software version 15.50. Sequence identity was verified to species level using BOLD taxonomic and reference sequence search [46].
2.4. Species and genetic diversity in Panama pacific fisheries
2.4.1. Species and family diversity
Total number of families and species richness (S = total number of species per collection site) was quantified overall (all samples considered) and by site and by Gulf (Gulf of Panama, Gulf of Chiriqui and Gulf of Montijo) (Table 1). Total number of species and families by Gulf was compared using a non-parametric test (Kruskall-Wallis).
2.4.2. Genetic diversity
Genetic diversity of the most common species was determined overall and by site. Genetic diversity was estimated using the following parameters: total number of haplotypes (H), total number of polymorphic sites (S), haplotype diversity (Hd) and nucleotide diversity (π). These values were determined in species with more than five sample sequences available by sampling site using DnaSP v. 6.1 [47] .
2.4.3. Genetic connectivity and demographic history
Genetic differentiation (Fst) was calculated overall by species and by site to examine the existence of genetic stocks and understand patterns of connectivity and differentiation among sites (Table 2 and Table 3). Hudson 2000 statistical (snn) significance p<0.0001 was also compared among species to infer genetic structure and connectivity. The total number of haplotypes (H) and haplotype diversity (Hd) were used as a measure of population identification, stock fitness and sustainability. Connectivity between sites was estimated indirectly using values of pairwise Fst and genetic distances (species with representative sequences in all sites) were calculated using DNAsp 6.0.
Demographic history was estimated using values of Tajimas D and Fu´s. These parameters were calculated in species with more than 10 individual sequences. Population expansion and bigger population size was evidenced with negative and significant values of Tajima´s D while positive and non-significant values of this parameter showed smaller population size and population contraction.
2.4.4. Developing tools for molecular taxonomy and traceability
Primers and dual labeled probes for taqman qPCR environmental DNA detection and quantification were developed in silico using two programs Genescript and Primer Express 3.0. Ten sequences from each species (Caranx caballus, Caranx caninus, Lutjanus guttatus, Lutjanus argentiventris, and Scomberomorus sierra) including sequences from this study and GenBank reference sequences were aligned and compared to find a conserved specific region for eDNA target species detection. Amplicon segments were tested through BLAST to corroborate species specificity as well as primer and probe set.
3. Results
3.1. Sequence analysis
We obtained 164 partial sequences of 614 base pair of the mitochondrial gene cytochrome oxidase I out of 203 tissue samples. For each identified haplotype of the analyzed species, a GenBank accession number was generated for all sequences (Table 1). All analyzed sequences showed between 98 to 100% of query call and a 100% of similarity with GenBank reference sequences. Some sequences reported here represent the first sequences of the species for Panamanian waters (Table 1).
3.2. Species, family and genetic diversity in Panama pacific fisheries
A total of 34 species belonging to 14 families were identified out of 164 sequences obtained from 223 fish tissue samples. The most diverse families were Carangidae (Selene peruviana, Caranx caballus, Caranx caninus, Carangoides vinctus and Caranx sexfasciatus) and Scianidae (Nebris occidentalis, Larimus pacificus, Cynoscion phoxocephalus, Cynoscion leiarchus and Paralunchurus brasiliensis) with five species each. The second most diverse family was Lutjanidae (Lutjanus argentiventris, Lutjanuns guttatus, Lutjanus peru and Lutjanus colorado); and Centropomidae (Centropomus robalito, and Centropomus viridis and Centropomus unionensis and Centropomus medius) with 4 species each. Haemulidae family had 3 species (Pomadasys panamensis, Haemulopsis leuciscus and Orthoprisitis chalceus). Other observed families in this study such as Lobotidae (Lobotes surinamensis and Lobotes pacificus), Scombridae (Scomberomorus sierra and Thunnus albacares) and Serranidae (Myctoperca xenarcha and Hyporthodus acanthistius) had two species each. Finally, families: Sphyraenidae (Sphyraena ensis), Malacanthidae (Caulolatilus affinis), Stromatidae (Peprilus medius), Ariidae (Bagre marinus), Gerridae (Guerres cinereus) and Mugilidae (Mugil curema); were represented by one species.
Among areas, family and species diversity showed higher numbers in the Gulf of Panama (14 families and 34 species) while 7 families and 10 species were registered in the Gulf of Chiriquí. Among landing sites, higher diversity values were found in San Felipe market with 18 species, Coquira Port with 17 species and Anton fishery market with 11 species. All these landing sites are in the Gulf of Panama. In contrast, landing sites located in the Gulf of Chiriquí showed 5 and 10 species in Port of Montijo and Port of Pedregal respectively (Table 1). An interesting finding was the existence of species from the Atlantic side of the country in Pacific Landing sites, i.e., Mycroperca xenarcha, Paralonchurus brasiliensis and Lobotes surinamensis.
3.3. Genetic diversity
COI haplotypes were identified for all species. Nonetheless, genetic diversity values such as nucleotide diversity (π) and haplotype diversity (Hd) were quantified in species with five or more individual sequences. Genetic diversity comparisons between sampling sites were only conducted in species with representative sequences in different Gulf areas (i.e. Lutjanus guttatus 10 sequence in Gulf of Panama, 10 sequences in the Gulf of Chiriquí and five in the Gulf of Montijo and 11 from Coiba (Table 2). Overall genetic diversity parameters were in a range between 0.54-1.00 for haplotypic diversity and between 0.00113 to 0.01656 for nucleotide diversity.
3.3.1. Genetic connectivity and demographic history.
Overall values of Fst (genetic differentiation) by species, Fst per pairs (genetic differentiation between pairs of populations) and genetic distances (Da) were used as indirect values of genetic connectivity by species and among study areas. Fst per pairs estimates were only conducted on species with representative number of sequences in all areas (Gulf) (Table 3).
Demographic history of the most common species was determined using Tajima’s D and Fu´s values. Results from these parameters are presented in Table 2. Tajima’s D values were negative in five of the analyzed species. However, was only negative and significant in two snapper species: Lutjanus peru and Lutjanus argentiventris.
3.3.2. Developing tools for molecular taxonomy and traceability:
Five primer sets (forward and reverse) and five double dye species specific FAM-TAMRA probes were designed and tested in silico for five species: Lutjanus guttatus, Lutjanus peru, Scomberomorus sierra, Caranx caballus, Caranx caninus (Table 4). Average amplicon size for these species was 119bp with a range between 97 and 168bp and a probe size that varied between 20 to 35bp (Table 4).
4. Discussion
4.1. Species diversity
In this study, 34 fish species were identified out of 164 sequences. This is the first molecular identification study and largest number of DNA sequences deposited in Genebank for Panama Pacific marine commercial teleost species in a single publication including the main fishery areas of Panama Pacific. No similar data set have been reported for the Eastern Tropical Pacific. However. There is a similar study focused on Caribbean reef fishes; however, that study does not include Panama samples [42]. This research is also pioneer because we provide the first molecular list of commercial bony fishes collected and commercialized along the main fishery areas of the country including different points of the commercial chain i.e., landing sites and fish markets. The total number of species and families obtained in this study represent a high percentage of the total commercial species already reported for Pacific side using traditional taxonomic methods.
Regarding feeding habits, most samples corresponded to carnivores and piscivores species, with coastal and demersal habits (Lutjanidae, Scombridae, Scianidae, Serranidae, Haemulidae and Centropomidae). Nonetheless, large pelagic carnivores with more oceanic and epipelagic habits were also molecularly identified i.e., Jacks and Pompanos (Carangidae) barracudas (Sphyraenidae) and tunas (Scombridae).
Among the registered families of carnivore fish, the most diverse was Carangidae with four species: Caranx caninus (Pacific crevalle jack), Caranx vinctus (Cocinero jack), Caranx caballus (Green jack) and Selene peruviana (Pacific moonfish). For corvinas and croakers (Scianidae) five species were identified Cynoscion othonopterus (Gulf weakfish); Nebris occidentalis (Pacific smalleye croaker); Cynoscyon phoxocephalus (Cachema weakfish), Cynoscion leiarchus (Smooth weakfish) and Paralochurus dumerilli (Suco croaker). For snappers, 4 species were collected and molecularly identified. One was an exclusively estuarine species Lutjanus argentiventris (yellow snapper) [48].; one coastal species, Lutjanus guttatus (spotted rose snapper) [49] and two species with oceanic habits Lutjanus peru (Pacific red snapper) and Lutjanus colorado (colorado snapper) were also identified [50].
Other important families were Scombridae, Haemulidae and Serranidae. Among these families the most common species was Scomberomorus sierra. As a juvenile, Scomberomorus sierra is common in lower estuary areas. However, as it becomes an adult, it starts migration to open coastal and oceanic areas [38]. Haemulidae (grunts) family was represented by three species mainly located in the Gulf of Panama.
Other families with different feeding habits than midsize carnivores were collected in small numbers like: Stromatidae, Lobotidae, Sphyraenidae, Ariidae, Mugilidae, Gerreidae, and Malacanthidae for a total of 14 families and 34 species. Ecologically, these feeding habits results show that fishing pressure is mainly centered on secondary consumers, large pelagics, demersal carnivores and planktivores as an evidence of a reduced selection pressure for herbivores and omnivores fish in Pacific Panamanian waters. Other non-traditionally commercial families or less popular among consumers were also molecularly identified. That was the case of Ariidae, Malacanthidae and Guerridae. Species composition of these families were very similar among Gulfs (No significant differences were observed P>0.05). Nonetheless, Gulf of Panama was slightly more diverse. Different factors could be generating a greater number of fish species in the Gulf of Panama landings.
Some other species listed in previous taxonomic and fishery studies for Panama Pacific were not collected in the present study, e.g., Pomadasys macracanthus, Cynoscion squamipinnis, Cynoscion albus, Coryphaena hippurus, Hyporthodus acanthistius, Hyporthodus niphobles, Scomber japonicus, Scomberomorus maculatus, Sarda orientalis, Katsuwonus pelamis, Thunnus alalunga, Thunnus obesus and Thunus albacares. One possible explanation to that is that most all of our samples were collected during visits to landing sites, markets and gathering sites with the exception of red snapper samples from Coiba National Park, collected from artisanal fishermen. The absence of large pelagic species like tunas, dolphin fish and bonito in our analyzed landing sites and fishery markets obeyed to a direct commercialization of these species from the industrial fishery sector to international boats and external processing plants.
Another finding was the existence species reported in the Caribbean side of the country like Mycroperca xenarcha, Paralonchurus dumerilli and Lobotes surinamensis in gathering sites and public markets of the Pacific side of the country as evidence for local commercialization and traceability of Caribbean species into bigger Pacific markets with a higher consumer demand.
Genetic diversity: understanding genetic diversity patterns can help us to understand sustainability of fisheries and fitness. Higher values of this parameter indicate which species are likely to adapt to future disturbance. Genetic diversity is also a measure of reproduction strategy, fecundity, effective population size [51,52] and life history variation base on habitat type and period of larval development [53]. Genetic diversity values make it possible to establish comparisons of these parameters for species occupying different habitats or having different life histories [53,54,55]. Despite of its importance genetic diversity data are limited and are regionally oriented mainly epipelagic species i.e., tunas, jacks [30,56].
For coastal and demersal species studies are limited to the molecular identification of species of the genus Cynoscion at Panama Bay[57] as well in some important commercial species such as corvinas (Cynoscion albus) and snappers (Lutjanus peru and Lutjanus guttatus) from Pacific Costa Rica [58].; demersal shark species like Mustelus lunulatus and Mustelus henlei [59] and deep-sea water species [60]. Therefore, values of genetic diversity for commercial species in the Eastern Tropical Pacific are reduced and for Panama Pacific are completely absent. Thus, collected information from this research for commercial species will be valuable for the establishment of a better species-specific tariff code base on a binomial nomenclature species system supported by molecular data of the country (haplotypes reported in Panama)[13].
In this study genetic diversity parameters were in a range between 0.54-1.00 for haplotypic diversity and between 0.00113 to 0.01656 for nucleotide diversity. Overall species with less retention and higher migratory habits showed higher values of genetic diversity and elevated connectivity as an evidence of a bigger population size [53,54,61] (Table 2). Among the five analyzed species for genetic diversity, three species from Lutjanidae family (Snappers) with different life histories showed contrast in genetic diversity values (Table 2). Genetic studies for this family in the Pacific are mainly focused in Mexico and Baja California [62]. In previous studies Lutjanus peru (red snapper) showed elevated genetic diversity in Mexican populations suggesting elevated population size and lack of genetic structure in the analyzed populations [63]. Regionally, neither population boundaries nor genetic stocks have been established along the Eastern Tropical Pacific for Lutjanus peru or Lutjanus guttatus (rose spotted snapper)[64].
In contrast, species with reduced migration (estuarine habits) reported lower values of genetic diversity and connectivity i.e., Lutjanus argentriventris among snappers in this study and Lutjanus guttatus in a previous study[65] . These results are the first genetic data for the species and no genetic information is available for this species in GenBank, only a few unpublished sequences have been uploaded to the data base.
4.2. Genetic connectivity and demographic history
The study of population genetics, demographic history and genetic connectivity have proved to be an important tool for fishery management and understanding population structure, reproduction patterns, migration, passive larval dispersal and determine selection pressures that allow scientists and decision makers to generate conservation strategies in fish exploited populations[51,66]. Connectivity patterns of three species: L. guttatus, Caranx caballus and Scomberomorus sierra were examined. Only L. guttatus showed significant differences among the three fishery areas (Gulf of Montijo, Gulf of Panama and Gulf of Chiriquí) (Table 3). However, elevated connectivity or lack of genetic differentiation among Gulf areas was evidenced among samples of the following species: S. sierra and C. caballus (Table 3) specially between Gulf of Chiriquí and Gulf of Panama. In contrast, other studies for this area also suggest elevated connectivity among Gulf of Chiriquí and Gulf of Montijo[59].
In terms of demographic history, despite L. guttatus showed negative values of Tajimas D (Table 2). These values were non-significant and close to 0 indicating balancing selection, possibly associated to an intermediate population size, characteristic of a coastal demersal species that is under exploitation from both industrial and artisanal fisheries. In contrast, other snapper species like L. peru considered an offshore species and L. argentiventris a coastal estuarine species exhibited negative and significant values of Tajima´s D (Table 2) indicating population expansion and sustainability of their fisheries in the Pacific Panama.
4.3. Developing tools for molecular taxonomy and traceability:
Developing Environmental DNA molecular markers (primers and probes) for molecular taxonomy and traceability
Molecular data from Panama commercial fish species is not available for most of the listed and reported fish species. Recently, some species have been characterized in ceviche samples in an effort of conservation agencies and regional NGO´s, like MarViva and COIBA AIP, to avoid the consumption of CITES and endangered shark and ray species in ceviche (Díaz-Ferguson et al. in preparation). Molecular information of invasive cichlid species including the Nile tilapia reported in Panamanian waters have been recently published [65]. Also, the genetic connectivity patterns and local movements of the largest fish of the world (Rhincodon typus) including transient individuals collected in Panama waters has been reported [67]. Environmental DNA (eDNA) or short size DNA fragments that an organism leaves behind in nonliving components of the ecosystems (i.e., water, sediments, or any other surface) has proved to be effective in detecting and tracking fish and fish products as well as other vertebrate species without the necessity of sample collection or even species observation [68,69,70]. Through this research we provide primers, probes and target amplicon regions for qPCR eDNA detection of five important commercial fish species (Lutjanus guttatus, Lutjanus argentiventris, Caranx caballus, Caranx caninus, Scomberomorus sierra) (Suplementary material). All designed amplicons had an average size of 119 base pairs (Table 4). These primers and probes could be used as a methodology to avoid exportation fraud and the use of false tariff codes for exportation. Detection of amplicons this size for eDNA is documented to be more efficient than bigger fragments. However, small sequences might not be fully sequenced for confirmation, and ambiguities within the sequences are common to the degraded nature of these products especially when samples are coming from processed foods such as liver, oil pills, shark fin soup, skin care products [71] and ceviche[13]. In addition, of the new developed markers, new obtained haplotypes were deposited on Genebank for all analyzed species. This information will become a reference for fishery studies providing the origin and fishing area of these species. Also, fish mitochondrial haplotypes have been used for determining and track farm fish population origin as well as to introduce new variants into hatcheries to improve genetic variability, fitness and improve breeding programs [72].
5. Conclusions
Conservation and management of genetic diversity is key for sustainable fisheries. This research provides important information and molecular tools for accurate identification of species, its traceability along the supply chain by providing species specific tariff codes using a binomial species-specific nomenclature system, conduct species specific biodiversity monitoring and fishery management. In addition, this information contributes to the future generation of a molecular data base for accurate stock identification, genetic health programs, the establishment of population boundaries, connectivity patterns, understanding population patterns and effective management of population size (Ne). Better understanding of these features will provide accurate information for decision makers and policy planners that contribute, and support national and regional policies based on scientific data and creates a balance between the power of the industrial fishery sector, local governments, and fishery managers. Thus, by providing the first haplotypes and values of genetic diversity for some species like L. guttatus, S. sierra and C. caballus collected in Panamanian waters we initiate the molecular database that will be a step forward for the future establishment of marine species and fishery product traceability program that will be crucial to achieve an ecosystem management approach of our fisheries, recover overexploited stocks and reduce illegal, unreported and unregulated fisheries (IUU) [23,73]. In the future, similar efforts should be considered for sharks, rays and billfishes since several populations are currently endangered, banned or has CITES status.
Author Contributions
Conceptualization of the paper, E. Díaz-Ferguson and C. Ramos; sequence analysis and sequences editing, E. Díaz-Ferguson and C. Ramos; primer and probes design for eDNA qPCR detection, E. Díaz-Ferguson; original draft preparation, writing, review and editing, E. Díaz-Ferguson & C. Ramos; sample collection, C. Ramos, M. Chial, O. Chen, M. González, O. Durán, A. Vega; DNA extraction and PCR, E. Díaz-Ferguson, S. Justo, C. Ramos; generation of accession numbers through Genebank, S. Justo; final review of the manuscript and response to reviewers E. Díaz-Ferguson.
Funding
This research was fund by Coiba Scientific Station and VIP of the University of Panama.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
We want to thank VIP of the University of Panama and Coiba Scientific Station (COIBA AIP) for funding this project. To the Panamanian Association for the Advance of Science (APANAC) for the award received as best biodiversity research poster during their last international congress. To our Lab Tech, Sara Justo, for helping in the process of editing sequences and generating accession numbers to Genebank and in the collection site map design. We also want to thank the Direction of Research and Development of the National Authority of Aquatic Resources of the Republic of Panama (ARAP) for the logistic support provided during the collection of fish samples from Pedregal and Remedios ports in Chiriqui Province and from Port Mutis, in Veraguas Province.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Helyar, S.J.; Lloyd, H.A.D.; De Bruyn, M.; Leake, J.; Bennett, N.; Carvalho, G.R. Fish Product Mislabelling: Failings of Traceability in the Production Chain and Implications for Illegal, Unreported and Unregulated (IUU) Fishing. PLoS One 2014, 9. [Google Scholar] [CrossRef]
- Costa Leal, M.; Pimentel, T.; Ricardo, F.; Rosa, R.; Calado, R. Seafood Traceability: Current Needs, Available Tools, and Biotechnological Challenges for Origin Certification. Trends Biotechnol 2015, 33, 331–336. [Google Scholar] [CrossRef]
- Chambers, E.A.; Hebert, P.D.N. Assessing Dna Barcodes for Species Identification in North American Reptiles and Amphibians in Natural History Collections. PLoS One 2016, 11. [Google Scholar] [CrossRef]
- Radulovici, A.E.; Archambault, P.; Dufresne, F. DNA Barcodes for Marine Biodiversity: Moving Fast Forward? Diversity (Basel) 2010, 2, 450–472. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; DeWaard, J.R. Biological Identifications through DNA Barcodes. Proceedings of the Royal Society B: Biological Sciences 2003, 270, 313–321. [Google Scholar] [CrossRef]
- Ward, R.D.; Hanner, R.; Hebert, P.D.N. The Campaign to DNA Barcode All Fishes, FISH-BOL. J Fish Biol 2009, 74, 329–356. [Google Scholar] [CrossRef]
- Allen G & Robertson R Fishes of the Tropical Eastern Pacific; Univ. Hawaii Press.; Honolulu, Hawaii., 1994.
- Paul Humann; Ned DeLoach Reef Fish Identification : Baja to Panama; 1st ed.; Jacksonville, Fla. : New World, 2004.
- Froese, R. and D.P. FishBase. http://www.fishbase.org/search.php 2000, 344.
- Comerciales, M.; Pacífico De Panamá, D.; Garcés, H. LISTA SISTEMÁTICA PRELIMINAR DE LOS PECESMARINOS COMERCIALES DEL PACÍFICO DE PANAMÁ. Tecnociencia (Panama) 2021, 23, 198–237. [Google Scholar] [CrossRef]
- Agnew, D.J.; Pearce, J.; Pramod, G.; Peatman, T.; Watson, R.; Beddington, J.R.; Pitcher, T.J. Estimating the Worldwide Extent of Illegal Fishing. PLoS One 2009, 4. [Google Scholar] [CrossRef]
- Rodríguez-Arriati Plan de Acción Nacional Para La Conservación y Ordenación de Las Pesquerías de Tiburones; Ciudad de Panamá, Panamá, 2010.
- Díaz-Ferguson E; Justo S; Posada J; Ramos C; Del Cid V Fishery and Conservation Implications of Molecular Characterization and Traceability of Ceviche Samples from Pacific Panama; Panamá, 2023. 2023.
- Olsen, P.; Borit, M. How to Define Traceability. Trends Food Sci Technol 2013, 29, 142–150. [Google Scholar] [CrossRef]
- Amanda Naaum; Robert Hanner Seafood Authenticity and Traceability for Fisheries Compliance 2016, 1–188.
- Poghosyan, A.; Gonzalez-Diaz; F., B.; Y., M.; F., G.; H., H.; V., S.S.; Marino, M.; Mena, S.A.; Ahmedov, Z.; et al. Traceability and Assurance Protocols in the Global Food System. International Food and Agribusiness Management Review 2004, 7, 118–126. [CrossRef]
- Alfaro, J.A.; Rábade, L.A. Traceability as a Strategic Tool to Improve Inventory Management: A Case Study in the Food Industry. Int J Prod Econ 2009, 118, 104–110. [Google Scholar] [CrossRef]
- Ogden, R. Fisheries Forensics: The Use of DNA Tools for Improving Compliance, Traceability and Enforcement in the Fishing Industry. Fish and Fisheries 2008, 9, 462–472. [Google Scholar] [CrossRef]
- Galimberti, A.; De Mattia, F.; Losa, A.; Bruni, I.; Federici, S.; Casiraghi, M.; Martellos, S.; Labra, M. DNA Barcoding as a New Tool for Food Traceability. Food Research International 2013, 50, 55–63. [Google Scholar] [CrossRef]
- Marín, A.; Serna, J.; Robles, C.; Ramírez, B.; Reyes-Flores, L.E.; Zelada-Mázmela, E.; Sotil, G.; Alfaro, R. A Glimpse into the Genetic Diversity of the Peruvian Seafood Sector: Unveiling Species Substitution, Mislabeling and Trade of Threatened Species. PLoS One 2018, 13. [Google Scholar] [CrossRef]
- Charlebois, S.; Sterling, B.; Haratifar, S.; Naing, S.K. Comparison of Global Food Traceability Regulations and Requirements. Compr Rev Food Sci Food Saf 2014, 13, 1104–1123. [Google Scholar] [CrossRef]
- Maralit, B.A.; Aguila, R.D.; Ventolero, M.F.H.; Perez, S.K.L.; Willette, D.A.; Santos, M.D. Detection of Mislabeled Commercial Fishery By-Products in the Philippines Using DNA Barcodes and Its Implications to Food Traceability and Safety. Food Control 2013, 33, 119–125. [Google Scholar] [CrossRef]
- Díaz-Ferguson E; Duran I Evaluación de La Trazabilidad de Los Productos de La Pesca Del Corredor Marino Del Pacífico Este Tropical – Panamá; Panamá, 2012 .
- Díaz-Ferguson, E. Introducción a La Ecología Molecular Marina : Aplicaciones y Perspectivas; Primera Edición.; Panamá, 2012; ISBN 9789962686743 .
- Díaz Uribe; J. Gabriel; Ernesto A.; Garay, E.; F. Juan Assessment of the Pacific Red Snapper (Lutjanus Peru) Fishery in the Southwestern Gulf of California. Cienc Mar 2004, 30, 561–574. [CrossRef]
- Amezcua, F.; Soto-Avila, C.; Green-Ruiz, Y. Age, Growth, and Mortality of the Spotted Rose Snapper Lutjanus Guttatus from the Southeastern Gulf of California. Fish Res 2006, 77, 293–300. [Google Scholar] [CrossRef]
- Mair, J.M.; Cipriani, R.; Guzman, H.M.; Usan, D. Fishery of the Green Jack Caranx Caballus (Osteichytes: Carangidae) in Las Perlas Archipelago, Pacific Panama. Rev. Biol. Trop. (Int. J. Trop. Biol. ISSN 2012, 60, 1271–1288 . [CrossRef]
- Nava-Ortega R; Espino-Barr E; Gallardo-Cabello M; Garcia-Boa A; Puente-Gomez M; Cabral-Solis E Growth Analysis of the Pacific Sierra Scomberomorus Sierra in Colima, México. Revista de Biologia Marina y Oceanografia 2012, 47, 273–281 . [CrossRef]
- Guzman, H.M.; Díaz-Ferguson, E.; Vega, A.J.; Robles, Y.A. Assessment of the Dolphinfish Coryphaena Hippurus (Perciformes: Coryphaenidae) Fishery in Pacific Panama. Rev. Biol. Trop. (Int. J. Trop. Biol. ISSN 2015, 63, 705–716. [Google Scholar]
- Caballero S; Puentes V Genetic Characterization of the Main Tuna Species (Yellow Fin Tuna, Big Eye Tuna and Skipjack Tuna) and Tuna by Catch Species (Sharks, Mahi-Mahi, Wahoo, Sea Turtles, Marine Mammals and Billfishes) Regulated by the Interamerican Tropical Tuna Commission in the Eastern Pacific Ocean. 2011, 0–25.
- Landínez-García, R.M.; Ospina-Guerrero, S.P.; Rodríguez-Castro, D.J.; Arango, R.; Márquez, E. Análisis Genético de Lutjanus Synagris En Poblaciones Del Caribe Colombiano. Cienc Mar 2009, 35, 321–331. [Google Scholar] [CrossRef]
- Bayona-Vásquez, N.J.; Hernández-Álvarez, C.A.; Glenn, T.; Domínguez-Domínguez, O.; Uribe-Alcocer, M.; Díaz-Jaimes, P. Complete Mitogenome Sequences of the Pacific Red Snapper (Lutjanus Peru) and the Spotted Rose Snapper (Lutjanus Gutattus). Mitochondrial DNA A DNA Mapp Seq Anal 2017, 28, 223–224. [Google Scholar] [CrossRef]
- Domínguez López, M.; Alcocer, M.U.; Jaimes, P.D. Phylogeography and Historical Demography of the Pacific Sierra Mackerel (Scomberomorus Sierra) in the Eastern Pacific. BMC Genet 2010, 11. [Google Scholar] [CrossRef]
- Theisen, T.C.; Bowen, B.W.; Lanier, W.; Baldwin, J.D. High Connectivity on a Global Scale in the Pelagic Wahoo, Acanthocybium Solandri (Tuna Family Scombridae). Mol Ecol 2008, 17, 4233–4247. [Google Scholar] [CrossRef]
- Marko PB; Lee SC; Rice AM, G.J.; Fitzhenry TM; Mcallister JS; Harper GR; Moran AL Mislabeling of Depleted Reef Fish. Nature 2004, 430, 309–310. [CrossRef]
- Veneza, I.; Silva, R.; Freitas, L.; Silva, S.; Martins, K.; Sampaio, I.; Schneider, H.; Gomes, G. Molecular Authentication of Pargo Fillets Lutjanus Purpureus (Perciformes: Lutjanidae) by DNA Barcoding Reveals Commercial Fraud. Neotropical Ichthyology 2018, 16. [Google Scholar] [CrossRef]
- Vega A; Robles Y; Boniche S; Rodrígues M ASPECTOS BIOLÓGICO – PESQUEROS DEL GÉNERO (PISCES: SCIAENIDAE) EN EL GOLFO DE MONTIJO, PACÍFICO PANAMEÑO. Tecnociencia (Panama) 2008, 10.
- Vega, Á.J.; Quezada, F.; Robles, Y.A. Aspectos Biológicos y Pesqueros de Scomberomorus Sierra (Perciformes:Scombridae) En El Golfo de Montijo Pacífico de Panamá. Tecnociencia (Panama) 2013, 15, 23–70. [Google Scholar]
- Vega, A.J.; Nelva Villarreal, Y. PECES ASOCIADOS A ARRECIFES Y MANGLARES EN EL PARQUE NACIONAL COIBA. Tecnociencia (Panama) 2003, 5, 65–74. [Google Scholar]
- Dominici-Arosemena, A.; Wolff, M. Reef Fish Community Structure in the Tropical Eastern Pacific (Panamá): Living on a Relatively Stable Rocky Reef Environment. Helgol Mar Res 2006, 60, 287–305. [Google Scholar] [CrossRef]
- Benfield, S.; Baxter, L.; Guzman, H.M.; Mair, J.M. A Comparison of Coral Reef and Coral Community Fish Assemblages in Pacific Panama and Environmental Factors Governing Their Structure. Journal of the Marine Biological Association of the United Kingdom 2008, 88, 1331–1341. [Google Scholar] [CrossRef]
- Beleño F; Adames P Estudio de La Abundacia y Determinacion Taxonomica de Las Especies de Peces Comerciales Del Pacifico Veraguense., Universidad de Panamá, 1992.
- Joint FAO/WHO Codex Alimentarius Commission.; World Health Organization.; Food and Agriculture Organization of the United Nations. Code of Practice for Fish and Fishery Products.; World Health Organization, 2012; ISBN 9789251070185.
- Ward, R.D.; Zemlak, T.S.; Innes, B.H.; Last, P.R.; Hebert, P.D.N. DNA Barcoding Australia’s Fish Species. Philosophical Transactions of the Royal Society B: Biological Sciences 2005, 360, 1847–1857. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An Integrated and Extendable Desktop Software Platform for the Organization and Analysis of Sequence Data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The Barcode of Life Data System: Barcoding. Mol Ecol Notes 2007, 7, 355–364. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A Software for Comprehensive Analysis of DNA Polymorphism Data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Bonilla-Gómez, J.L.; Robles, Y.A.; Vega, A.J. Length-Weight Relationship and Biological Information of the Yellow Snapper Lutjanus Argentiventris from a Tropical Estuary: Río Caté, Gulf of Montijo, Panama. Journal of Applied Ichthyology 2014, 30, 227–229. [Google Scholar] [CrossRef]
- Vega Angel; Mate, J. Robles Yolani First Report of Reproductive Aggregations for Pacific Red Snappers Lutjanus Peru (Nicholson y Murphy, 1992) and Spotted Rose Snapper L. Guttatus (Steindachner, 1869) in the Coiba National Park, Pacific of Panama. GCF 2015, 68, 112–117.
- Gomes, G.; Schneider, H.; Vallinoto, M.; Santos, S.; Orti, G.; Sampaio, I. Can Lutjanus Purpureus (South Red Snapper) Be “Legally” Considered a Red Snapper (Lutjanus Campechanus)? Genet Mol Biol 2008, 31, 372–376. [Google Scholar] [CrossRef]
- Hauser, L.; Adcock, G.J.; Smith, P.J.; Bernal Ramírez, J.H.; Carvalho, G.R. Loss of Microsatellite Diversity and Low Effective Population Size in an Overexploited Population of New Zealand Snapper (Pagrus Auratus). Prog Nucl Energy 6 Biol Sci 2002, 99, 11742–11747. [Google Scholar] [CrossRef]
- Waples, R.S.; Luikart, G.; Faulkner, J.R.; Tallmon, D.A. Simple Life-History Traits Explain Key Effective Population Size Ratios across Diverse Taxa. Proceedings of the Royal Society B: Biological Sciences 2013, 280. [Google Scholar] [CrossRef]
- Martinez, A.S.; Willoughby, J.R.; Christie, M.R. Genetic Diversity in Fishes Is Influenced by Habitat Type and Life-History Variation. Ecol Evol 2018, 8, 12022–12031. [Google Scholar] [CrossRef]
- Ward R; Woodwark M; Skibinski O A Comparison of Genetic Diversity Levels in Marine, Freshwater and Anadromous Fishes. Fish Biology 1994, 44, 213–232. [CrossRef]
- DeWoody, J.A.; Avise, J.C. Microsatellite Variation in Marine, Freshwater and Anadromous Fishes Compared with Other Animals. J Fish Biol 2000, 56, 461–473. [Google Scholar] [CrossRef]
- Weiwen, L.; Chen, X.; Xu, Q.; Zhu, J.; Dai, X.; Xu, L. Genetic Population Structure of Thunnus Albacares in the Central Pacific Ocean Based on MtDNA COI Gene Sequences. Biochem Genet 2015, 53, 8–22. [Google Scholar] [CrossRef]
- Vergara-Chen, C.; González-Wangüemert, M.; Bermingham, E.; D’Croz, L. Identification of Weakfish Cynoscion (Gill) in the Bay of Panama with RFLP Markers. J Fish Biol 2009, 75, 1101–1107. [Google Scholar] [CrossRef]
- Edgardo Díaz-Ferguson Diversidad y Conectividad Genética de Tres Especies de Teleósteos de Importancia Pesquera En El Pacífico de Costa Rica. Informe Final de Proyecto. ; 2022.
- Justo, S. Estructura y Variabilidad Genética de Tiburones Del Género Mustelus Spp. En El Golfo de Chiriquí y Montijo Como Áreas de Influencia Del Parque Nacional Coiba. Thesis, International Maritime University of Panama: Panamá, 2022.
- Robertson, D.R.; Angulo, A.; Baldwin, C.C.; Pitassy, D.; Driskell, A.; Weigt, L.; Navarro, I.J.F. Deep-Water Bony Fishes Collected by the B/O Miguel Oliver on the Shelf Edge of Pacific Central America: An Annotated, Illustrated and DNA-Barcoded Checklist. Zootaxa 2017, 4348, 1–125. [Google Scholar] [CrossRef]
- Díaz-Ferguson, E. Desarrollo de Marcadores Moleculares y Determinación de La Estructura Genético Poblacional de Poblaciones Naturales de Solea Senegalensis (Perciformes: Soleidae), En El Golfo de Cádiz. Tesis Doctoral, Universidad de Cádiz: Cádiz, España, 2004.
- Gold, J.R.; Willis, S.C.; Renshaw, M.A.; Buentello, A.; Walker, H.J.; Puritz, J.B.; Hollenbeck, C.M.; Voelker, G. Phylogenetic Relationships of Tropical Eastern Pacific Snappers (Lutjanidae) Inferred from Mitochondrial DNA Sequences. Syst Biodivers 2015, 13, 916–927. [Google Scholar] [CrossRef]
- Rocha-Olivares A; Sandoval Castillo J Diversidad Mitocondrial y Estructura Genética En Poblaciones Alopátricas Delhuachinango Del Pacífico Lutjanus Peru. Cienc Mar 2003, 29, 197–209.
- Lago, F.C.; Herrero, B.; Vieites, J.M.; Espiñeira, M. Genetic Identification of Horse Mackerel and Related Species in Seafood Products by Means of Forensically Informative Nucleotide Sequencing Methodology. J Agric Food Chem 2011, 59, 2223–2228. [Google Scholar] [CrossRef]
- Díaz-Ferguson, E.; Allard, A.; Mendizabal, M.; Ramos, C. Genetic Structure of the Invasive Nile Tilapia Oreochromis Niloticus (Perciformes, Cichlidae) and Molecular Identification of Tilapia Species in Panama Using Barcode. Panam J Aquat Sci 2022, 17, 62–70. [Google Scholar] [CrossRef]
- Hadi, S.; Andayani, N.; Muttaqin, E.; Simeon, B.M.; Ichsan, M.; Subhan, B.; Madduppa, H. Genetic Connectivity of the Scalloped Hammerhead Shark Sphyrna Lewini across Indonesia and the Western Indian Ocean. PLoS One 2020, 15. [Google Scholar] [CrossRef]
- Guzmán, H.M.; Beaver, C.E.; Díaz-Ferguson, E. Novel Insights Into the Genetic Population Connectivity of Transient Whale Sharks (Rhincodon Typus) in Pacific Panama Provide Crucial Data for Conservation Efforts. Front Mar Sci 2021, 8. [Google Scholar] [CrossRef]
- Díaz-Ferguson, E.; Herod, J.; Galvez, J.; Moyer, G. Development of Molecular Markers for EDNA Detection of the Invasive African Jewelfish (Hemichromis Letourneuxi): A New Tool for Monitoring Aquatic Invasive Species in National Wildlife Refuges. Management of Biological Invasions 2014, 5, 121–131. [Google Scholar] [CrossRef]
- Díaz-Ferguson, E.E.; Moyer, G.R. History, Applications, Methodological Issues and Perspectives for the Use of Environmental DNA (EDNA) in Marine and Freshwater Environments. Rev. Biol. Trop. (Int. J. Trop. Biol. ISSN 2014, 62, 1273–1284. [Google Scholar] [CrossRef]
- Edgardo Díaz-Ferguson; Margaret Hunter; Héctor M. Guzmán Genetic Composition and Connectivity of the Antillean Manatee (Trichechus Manatus Manatus) in Panama. Panama Aquatic Mammals 2017, 43, 378–386. [CrossRef]
- Cardeñosa, D. Genetic Identification of Threatened Shark Species in Pet Food and Beauty Care Products. Conservation Genetics 2019, 20, 1383–1387. [Google Scholar] [CrossRef]
- Freitas, M. V.; Ariede, R.B.; Hata, M.E.; Mastrochirico-Filho, V.A.; Pazo, F. Del; Villanova, G. V.; Mendonça, F.F.; Porto-Foresti, F.; Hashimoto, D.T. Haplotypes Traceability and Genetic Variability of the Breeding Population of Pacu (Piaractus Mesopotamicus) Revealed by Mitochondrial Dna. Genet Mol Biol 2021, 44. [Google Scholar] [CrossRef]
- Lewis, S.G.; Boyle, M. The Expanding Role of Traceability in Seafood: Tools and Key Initiatives. J Food Sci 2017, 82, A13–A21. [Google Scholar] [CrossRef]
Figure 1.
Map of the Republic of Panama showing collection sites (ports associated to landing sites in red and gathering centers in blue) per Gulf along the Pacific Panama.
Figure 1.
Map of the Republic of Panama showing collection sites (ports associated to landing sites in red and gathering centers in blue) per Gulf along the Pacific Panama.
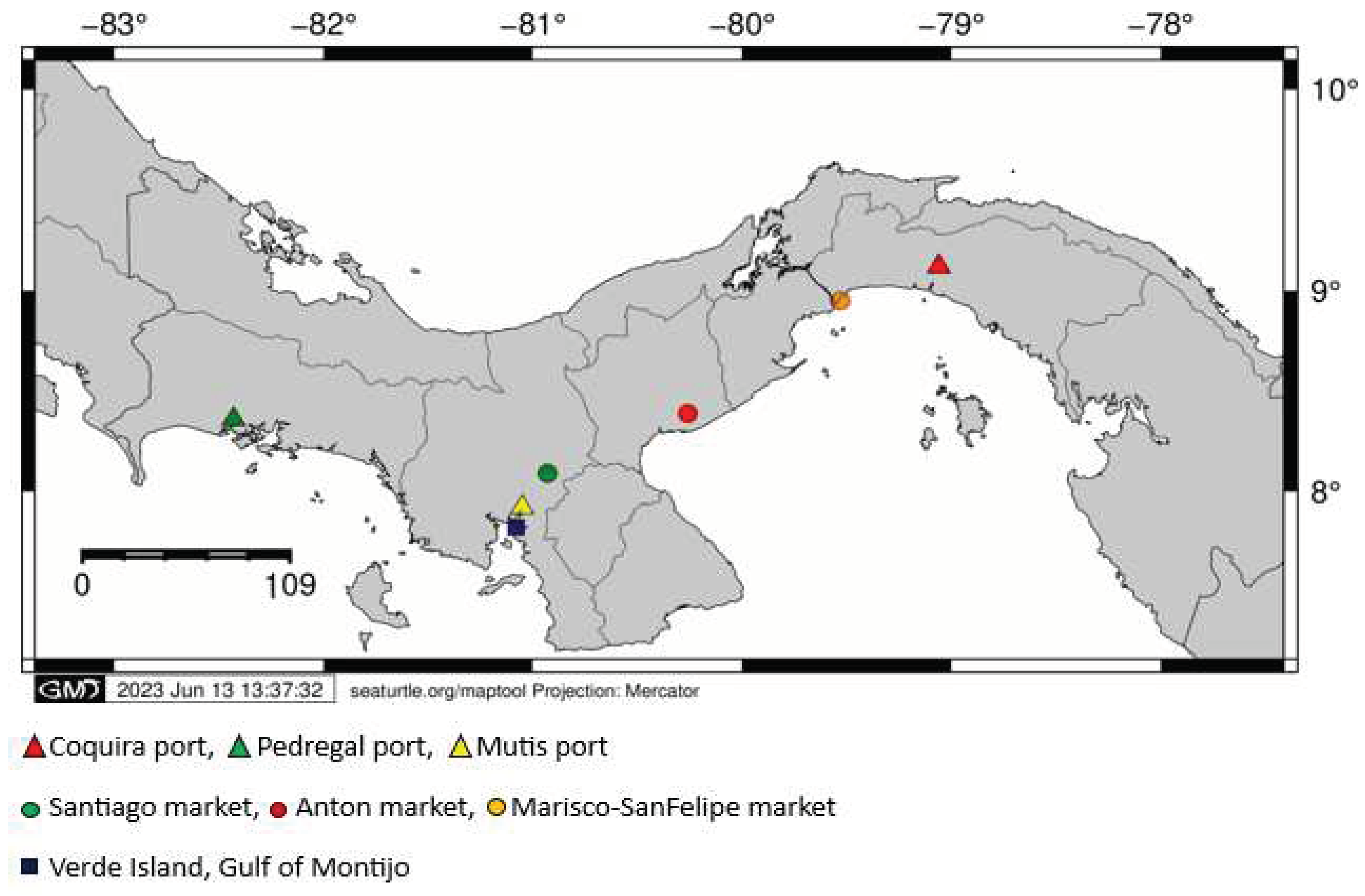

Table 1.
Teleost fish families and species identified per site, common name, and GenBank Accession number.
Table 1.
Teleost fish families and species identified per site, common name, and GenBank Accession number.
| Site | Family | Species | Common name | GenBank Accession number |
|---|---|---|---|---|
| GP _1_SFelipe | Scombridae | Scomberomorus sierra | Pacific Sierra | OQ790166 -OQ790174 |
| Euthynnus affinis | Tuna | OQ844071 | ||
| Carangidae | Caranx vinctus | Cocinero | OQ790077 | |
| Caranx caninus | Pacific Crevalle Jack | OQ801572 | ||
| Centropomus unionensis | Bigeye trevally | OQ850133 | ||
| Caranx caballus | Green Jack | OQ789895 | ||
| Selene peruviana | Peruvian moonfish | OQ843845 | ||
| Haemulidae | Pomadasys panamensis | Panama grunt | OQ789718 | |
| Lobotidae | Lobotes surinamensis* | Atlantic Triple Tail | OQ844072 | |
| Lobotes pacificus | Pacific Triple Tail | OQ844073 | ||
| Serranidae | Mycteroperca xenarcha* | Caribbean grouper | OQ844067 | |
| Hyporthodus acanthistius | Pacific grouper | OQ844070 | ||
| Sphyraenidae | Sphryraena ensis | Pacific barracuda | OQ850073 | |
| Scianidae | Nebris occidentalis | Guabina | OQ843562 | |
| Cynoscion phoxocephalus | Cachema weakfish | OQ843904 | ||
| Lutjanidae | Lutjanus guttatus | Rose spotted snapper | OQ850300 | |
| Lutjanus argentiventris | Yellow snapper | OQ790134- OQ790136 | ||
| Melacanthidae | Caulolatilus affinis | Bighead tilefish | OQ844074 | |
| GP_2_Anton | Scombridae | Scomberomorus sierra | Sierra fish | OQ790166 -OQ790174 |
| Carangidae | Caranx caballus | Green Jack | OQ789895 | |
| Caranx caninus | Pacific Crevalle Jack | OQ801572 | ||
| Selene peruviana | Peruvian Moonfish | OQ843845 | ||
| Centropomidae | Centropomus robalito | Yellowfin Snook | OQ844075 | |
| Lutjanidae | Lutjanus guttatus | Spotted rose snapper | OQ850300 | |
| Scianidae | Cynoscion othonopterus | Gulf weakfish | OQ850144 | |
| Ophioscion scierus | Tuza croaker | OQ848437 | ||
| Paralonchurus dumerilii | Banded croacker | OQ848738 | ||
| Stromateidae | Peprilus medius | Pacific Harvest Fish | OQ848659 | |
| GP_3Coquira | ||||
| Carangidae | Caranx caninus | Pacific crevalle Jack | OQ801572 | |
| Caranx caballus | Green Jack | OQ789895 | ||
| Centropomidae | Centropomus viridis* | Common Snook | ||
| Centropomus medius | Blackfin snook | OQ848755 | ||
| Lutjanidae | Lutjannus gutattus | Rose spotted snapper | OQ850300 | |
| Haemulidae | Haemulopsis leuciscus | Raucous grunt | OQ849145 | |
| Orthopristis chalceus | Brassy grunt | OQ849164 | ||
| Scianidae | Nebris occidentalis | Pacific smalleye croaker | OQ843562 | |
| Cynoscion phoxocephalus | Cachema weakfish | OQ843904 | ||
| Cynoscion leiarchus* | Smooth weak fish | |||
| Larimus pacificus | Pacific drum | OQ849155 | ||
| Guerridae | Gerres simillimus | Yellow fin mojarra | OQ872776 | |
| Lobotidae | Lobotus pacificus | Pacific Triple Tail | OQ844073 | |
| Ariidae | Bagre marinus* | Gafftopsail catfish | ||
| Mugilidade | Mugil curema* | White mullet | ||
| GCH_1-Pedregal | ||||
| Carangidae | Caranx caballus | Green Jack | OQ789895 | |
| Lutjanidae | Lutjanus argentiventris | Yellow snapper | OQ790134- OQ790136 | |
| Lutjanus guttatus | Rose spotted snapper | OQ850300 | ||
| Lutjanus peru | Red Snapper | OQ791284 | ||
| Scombridae | Scomberomorus sierra | Pacific sierra | OQ790166 - OQ790174 | |
| GMon_1-Mutis | ||||
| Scombridae | Scomberomorus sierra | Pacific sierra | OQ790166 - OQ790174 | |
| Scianidae | Cynoscion phoxocephalus | Cachema weakfish | OQ843904 | |
| Stromatidae | Peprilus medius | Pacific Harvest fish | OQ848659 | |
| Lutjanidae | Lutjanus argentiventris | Yellow snapper | OQ790134- OQ790136 | |
| Lutjanus guttatus | Rose spotted snapper | OQ850300 | ||
| GMon_2-Santiago | ||||
| Carangidae | Caranx vinctus | Jack | OQ790077 | |
| Caranx caninus | Pacific Crevalle Jack | OQ801572 | ||
| Caranx caballus | Green Jack | OQ789895 | ||
| Lutjanidae | Lutjanus Colorado | Colorado snapper | OQ801537-OQ801540 | |
| GCH_PixCOIBA | Lutjanidae | Lutjanus guttatus | Rose spotted snapper | OQ850300 |
| Lutjanus peru | Red Snapper | OQ791284 |
Note: GP (Gulf of Panama); GCH (Gulf of Chiriqui); GMon (Montijo area); GCH-PNC (Coiba); sequences with (*) have being edited and verified to create their accession numbers. However, accession numbers have not being assigned yet.
Table 2.
Overall values of genetic diversity and Tajima’s D of the six most common teleost fish species sampled in ports and markets of the Pacific Panama.
Table 2.
Overall values of genetic diversity and Tajima’s D of the six most common teleost fish species sampled in ports and markets of the Pacific Panama.
| Common name | Species | Familia | Total de secuencias | Haplotype diversity (Hd) | Nucleotide diversity (π) | Tajima’s D |
|---|---|---|---|---|---|---|
| Spotted rose snapper | Lutjanus guttatus | Lutjanidae | 36 | 0.775 | 0.020 | -0.155 |
| Yellow snapper | Lutjanus argentiventris | Lutjanidae | 12 | 0.63 | 0.031 | -2.25** |
| Red snapper |
Lutjanus peru |
Lutjanidae | 17 | 0.86 | 0.010 | -2.17** |
| Pacific Sierra | Scomberomorus sierra | Scombridae | 16 | 0.90 | 0.0045 | -1.09 |
| Green jack | Caranx caballus | Caranjidae | 13 | 0.91 | 0.0039 | -1.26 |
| Pacific crevalle jack | Caranx caninus | Caranjidae | 10 | 0.93 | 0.0038 | -0.86 |
* All species presented in this table showed 10 or more sequences and were represented in more than one Gulf.
Table 3.
Pairwise Fst between sites values for the most common taxa (all sampled individuals by Gulf). Comparisons between samples of Coiba National Park were only conducted in L. guttatus.
Table 3.
Pairwise Fst between sites values for the most common taxa (all sampled individuals by Gulf). Comparisons between samples of Coiba National Park were only conducted in L. guttatus.
| Lutjanus guttatus | Gulf of Panama | Gulf of Montijo | Gulf of Chiriqui | Overall by species Hudson 2000 Snn P<0.0001 |
|---|---|---|---|---|
| Gulf of Panama | 0.00 | |||
| Gulf of Montijo | 0.087 | 0.00 | ||
| Gulf of Chiriqui | 0.41 | 0.45 | 0.00 | L.guttatus** |
| Coiba-Pixvae | 0.052 | 0.00 | 0.44 | |
| Caranx caballus | ||||
| Gulf of Panama | 0.00 | |||
| Gulf of Montijo | 0.424 | 0.00 | C.caballus (P>0.05 ns) | |
| Gulf of Chiriqui | 0.500 | 0.00 | 0.00 | |
| Scomberomorus sierra | ||||
| Gulf of Panama | 0.00 | |||
| Gulf of Montijo | 0.0694 | 0.00 | S.sierra (P>0.05 ns) | |
| Gulf of Chiriqui | 0.0614 | -0.058 | 0.00 |
Table 4.
Developing real time PCR eDNA detection markers for molecular traceability of Pacific Panama commercial fish species.
Table 4.
Developing real time PCR eDNA detection markers for molecular traceability of Pacific Panama commercial fish species.
| Species | Primers sequences | Probe name and sequence | Amplicon size |
|---|---|---|---|
| Lutjanus guttatus | L-GATCGGAGGATTCGGGAACT R-GGGTAGACTGTTCACCCAGT |
5’FAM-TCCAGCACCGGCTTCTACTCCA-3´TAMRA | 168bp |
| Lutjanus argentiventris | L-TACTACTCGCCTCCTCTGGA R-TGGTTAGGTCAACAGACGCT |
5´FAM-TCCTGTTCCGGCACCGGCTT- 3´TAMRA | 108bp |
| Caranx caballus | L-TGGGACTGGCTGAACTGTTT R-CCCTGCTAGGTGAAGGGAAA |
5´FAM-TGCCCACGCGGGAGCATCAGT-TAMRA | 97bp |
| Caranx caninus | L-GCTTCTGACTTCTCCCTCCT R-GGCATGGGCAAGATTACCAG |
5´FAM-AGCCTGTTCCAGCTCCGGCT-TAMRA | 116bp |
| Scomberomorus sierra | L-AGCCCTTCTTGGAGATGACC R-TCAGTTTCCAAACCCTCCGA |
5´FAM-ACAATGTAATCGTTACGGCCCATGCC-TAMRA | 109bp |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



