Submitted:
28 June 2023
Posted:
28 June 2023
You are already at the latest version
Abstract
Crimean-Congo hemorrhagic fever virus (CCHFV), a member of the Nairoviridae family and Bunyavirales order, is transmitted to humans via tick bites or contact with the blood of infected animals. It can cause severe symptoms, including hemorrhagic fever, with a mortality rate between 5 to 30%. CCHFV is classified as a high-priority pathogen by the World Health Organization (WHO) due to its high fatality rate and the absence of effective medical countermeasures. CCHFV is endemic in several regions across the world, including Africa, Europe, the Middle East, and Asia, and has the potential for global spread. The emergence of the disease in new areas, as well as the presence of the tick vector in countries without reported cases, emphasizes the need for preventive measures should be taken. In the past, the lack of a suitable animal model susceptible to CCHFV infection has been a major obstacle in the development of vaccines and treatments. However, recent advances in biotechnology and the availability of suitable animal models have significantly expedited the development of vaccines against CCHF. These advancements have not only contributed to an enhanced understanding of the pathogenesis of CCHF but have also facilitated the evaluation of potential vaccine candidates. This review outlines of the immune response to CCHFV and animal models utilized for the study of CCHFV and highlights the progress made in CCHFV vaccine studies. Despite remarkable advancements in vaccine development for CCHFV, it remains crucial to prioritize continued research, collaboration, and investment in this field.
Keywords:
Crimean–Congo hemorrhagic fever virus
; hemorrhagic fever
; immune response
; animal models
; vaccine development.
1. Introduction
Crimean-Congo hemorrhagic fever virus (CCHFV) is a member of the genus Orthonairovirus in the family Nairoviridae and the order Bunyavirales [1,2]. The viral genome is composed of three negative-sense RNA segments: small (S), medium (M), and large (L). The S segment encodes the nucleoprotein (NP), while the M segment encodes the glycoprotein precursor (GPC), which later forms mature Gn, Gc, and several nonstructural proteins such as mucin, GP38, and NSm [3,4,5,6]. The L segment encodes the L protein responsible for viral RNA synthesis, which includes the RNA-dependent RNA polymerase (RdRp) and an ovarian tumor (OTU) protease domain that the L protein is crucial for viral RNA synthesis and includes the RNA-dependent RNA polymerase (RdRp) and an OTU protease domain that may aid in evading the host's innate immunity (Figure 1a,b) [7,8,9].
It was initially detected in Soviet soldiers in Crimea during the 1940s. In the 1960s, a virus with similar symptoms to the Crimean virus was discovered in the Belgian Congo (currently known as the Democratic Republic of the Congo). Further studies revealed that both viruses were antigenically identical, leading to the virus being named CCHFV [10,11,12,13]. CCHFV circulates within an enzootic cycle that involves ticks and vertebrates [14,15,16]. Although CCHFV has been isolated in multiple tick species, Hyolamma ticks serve as the host and biological vector for CCHFV due to its extensive geographical range, which closely correlates with the distribution of CCHF cases [17,18]. Ticks can transmit CCHFV vertically from one generation to the next, transovarially from one developmental stage to another, sexually from males to females during copulation, or through cofeeding from one tick to other ticks feeding on the same non-viremic host [19,20,21,22]. CCHFV infects a wide range of both wild and domestic animals. However, infections in these animals are typically asymptomatic, but they exhibit viremia for more than five days, which helps the maintenance of CCHFV in nature [14,21,25,26].
CCHFV is found in a vast geographic region, from western China to Africa, the Middle East, Spain, and the Balkans [25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41]. Turkey has been experiencing CCHF epidemics since 2002, with the number of cases increasing significantly in recent years [42,43,44,45,46]. Similarly, some Balkan countries have reported regular cases of the disease [47]. CCHF was first reported in Spain in 2016, although tick surveillance studies had already shown the presence of CCHFV in the area [37]. This suggests a shift in the main vector's geographic distribution, as there were no previous reports of autochthonous human cases in Spain [48,49,50]. Ticks primarily disperse over long distances only through their hosts [51]. Therefore, changes in tick populations are primarily associated with bird migrations or expansions of host populations. The geographical spread of tick populations concerning as infected ticks transported to non-endemic areas can spread the disease to humans [49,52,53,54]. Furthermore, uninfected ticks introduced to a new area may establish populations that can sustain the virus after its introduction. The expansion of CCHF's geographic distribution is also driven by several factors, including global warming, increasing of human mobility, and human activities such as deforestation and agricultural growth, leading to more people coming into contact with infected ticks and animals [55,56,57,58]. Transporting livestock is a widespread practice across the world, and its contribution to the spread of CCHF cannot be overlooked. The movement of animals across borders or the transportation of infected ticks from endemic regions can initiate new CCHF outbreaks in non-endemic regions. [59,60,61,62,63].
Human beings are regarded as accidental hosts of the CCHFV. Humans become infected through tick bites or exposure to crushed infected ticks during agricultural activities. Another significant source of infection is the blood of infected agricultural animals, which can be viremic but not display any symptoms of disease [64,65]. Nosocomial transmission contributes to the spread of CCHFV, leading to higher fatality rates compared to those resulting from tick bites. Several cases of nosocomial outbreaks have been linked to infected blood or needle-stick injuries during patient care [65,66,67,68,69]. CCHFV infection in humans can cause mild to severe symptoms, including high fever, malaise, myalgia, and gastrointestinal distress, typically after a short incubation period of about a week. Severe cases can result in hemorrhagic disease with a fatality rate ranging from 5 to 30%, often due to disseminated intravascular coagulopathy, shock, and/or multi-organ failure [64,65,66]. Due to its high fatality rate, widespread vector, and the absence of effective medical countermeasures for prevention and treatment, CCHFV is classified as a high-priority pathogen by the World Health Organization (WHO) [71].
2. Immune Response to CCHF Infection
Effective control of CCHFV infection in the host relies on immune responses from both the innate and adaptive systems. The innate immune system acts as the first line of defense against viruses, by limiting viral entry, translation, replication, and assembly. Additionally, it facilitates the identification and elimination of infected cells, as well as the development of adaptive immunity through coordination and acceleration. Type 1 interferons (IFN-α/β) are produced by the host's innate immune response against viruses. These responses are rapid and efficient, and can be generated and secreted by all mammalian cells. These immunomodulators facilitate the expression of antiviral proteins, inhibit cell proliferation, and help regulate apoptosis [72,73,74].
The innate immune response is stimulated by CCHFV, leading to the production of IFNs and interferon-stimulated genes (ISGs). Andersson et al. conducted in vitro studies that confirmed the antiviral effect of IFN on CCHFV. The studies highlighted a substantial reduction of vRNA levels in cells treate with IFN, emphasizing the crucial role of IFN in controlling CCHFV replication [75]. Hawman et al. recently created a novel model using type I interferon-deficient mice, whereby infection with the human clinical isolate strain Hoti resulted in progressive illness characterized by several days of overt clinical signs [76]. This model also demonstrated the induction and release of IFNs, subsequent upregulation of ISGs, and the involvement of the host's innate immune response to CCHFV [77]. Bente et al. confirmed the crucial role of IFN in combating CCHFV using a STAT-1 KO mouse model, where STAT1 is a central component of IFN signaling pathways [78]. In 2012, CCHFV-infected IFN1 deficient mice (IFNAR-/-) exhibited clinical symptoms resembling CCHF, whereas wild-type mice remained asymptomatic, highlighting the importance of IFN1 in preventing CCHFV infection [79]. IFN is also crucial in controlling infections and preventing diseases in animals even those with compromised adaptive immune systems.
Lindquist et al. demonstrated the temporary suppression of the immune response in various mouse strains (including wild-type and those with impaired adaptive immunity such as NOD/SCID, Prf1− /−, and Rag2− /−) by utilizing an anti-IFNAR1 monoclonal antibody (mAb) [80]. IFN responses play a significant role in determining disease severity. Studies indicated that polymorphisms in toll-like receptor genes (TLR7, 8, 9, and 10) are associated with increased illness severity in Turkish CCHF patients, thereby emphasizing the crucial role of TLRs as an immune-sensing pathway in controlling the virus. [81,82]. Despite IFNs being crucial in the host's immune response, CCHFV utilizes various strategies to evade and counteract the innate immune response. These include removing the 5' triphosphate group from the viral genome to avoid RIG-I recognition, delaying IRF-3 activation through particle recognition pathway, and downregulating NF-kappaB activation [83,84,85]. Additionally, studies have shown that the CCHFV can suppress the body's innate immune system by encoding an ovarian tumor-related deubiquitinase (OTU) domain. This domain deubiquitinates proteins involved in the body's signaling pathways thereby inhibiting innate immune responses including the antiviral response mediated by ISG15 modifications. In addition, the OTU domain has both de-ISGylation and deubiquitinase activity, which are important for viral pathogenesis [8,86].
Apoptosis can serve as a significant innate response to viral infections. It is a critical process involved in viral infections, as its suppression or induction determines the level of infection spread. Viruses can either inhibit host cell apoptosis, which is a defense mechanism against infections, to ensure their survival or on the contrary, promote apoptosis to eliminate uninfected immune cells and facilitate viral spread [87,88]. As with many other viruses, CCHFV also has the ability to regulate apoptosis. CCHF infection induces TNF-α and FasL-mediated apoptosis in cell culture [89]. CCHFV NP inhibits caspase 3 and caspase 9 activation, prevents apoptosis initiated by BAX, and curtails the release of cytochrome c from mitochondria [90]. However, the specific point in the intrinsic apoptosis pathway where NP disrupts activation is yet to be determined. Despite the CCHFV NP inhibiting activation in the intrinsic pathway of apoptosis, the CCHFV NSs, a cryptic ambisense product of the NP have been found to disrupt mitochondrial membrane potential, induce apoptosis by activating caspase 3/7 and cleaving poly ADP-ribose polymerase. Furthermore, the presence of a conserved DEVD motif in the virus' NP that can be cleaved by host caspase 3 implies a regulatory role in the virus's life cycle [91].
Pro-inflammatory cytokines and chemokines, which are produced by various immune cells in response to viral infections, play a crucial role in the innate immune response against viral pathogens [92]. When a virus enters the body, immune cells such as macrophages and dendritic cells recognize it through pattern recognition receptors (PRRs) on their surface. These PRRs identify pathogen-associated molecular patterns (PAMPs) that are unique to the invading virus. Upon recognition of viral PAMPs, immune cells release pro-inflammatory cytokines [92,93]. While pro-inflammatory cytokines and chemokines are critical for the innate immune response to viral infections, excessive or uncontrolled release of these molecules can lead to tissue damage and inflammatory disease [94,95]. CCHFV initially targets immune cells such as dendritic cells, macrophages, and monocytes [96]. This results in the production of pro-inflammatory cytokines, including TNF-α, IL1, IL6, IL8, IL-12, IFN-γ, MCP-1, and MIP-1b [97]. Pro-inflammatory response in severe or fatal diseases can lead to vascular dysfunction, disseminated intravascular coagulation (DIC), organ failure, and shock [98]. Increased levels of TNF-α, IL-8, IL-9, IL-15, IP-10, and MCP-1 are associated with disease severity and negative outcomes in patients from Turkey, Albania, and Kosovo [97,98,99,100,101]. Researchers have also found that the secretion of sTREM-1 by myeloid cells enhances inflammatory responses during CCHF virus infection, though it remains to be empirically demonstrated how excessive levels of these inflammatory agents may drive pathogenic processes [102,103].
While innate immunity serves as the initial protective response against CCHFV infection, a potent adaptive immune response is also essential for effective control of the infection. Anti-CCHFV IgM and IgG antibodies are detectable within 7-9 days from symptoms onset [104,105,106]. IgG antibodies are typically peaking during the second to third week and can remain for up to 3 years. In contrast, IgM titers decline within 3 weeks and become undetectable between 3-5 months after disease onset. The absence of serum antibodies has been linked to higher mortality rates in CCHF patients, suggesting that antibodies may provide protection against fatal CCHFV infection [101,104]. Kaya et al. assessed serial antibody responses on 31 patients with CCHF, 11 of which were fatal cases. The study revealed that all surviving patients had a positive IgG titer within 9 days of onset, whereas none of the fatal cases showed such a response at the same timepoint [107]. In a study conducted on 24 patients with 43 samples, it was observed that quantitative IgG levels and viral loads had a correlation, and none of the fetal patients developed positive IgG titers. Only one sample from nine survivors taken less than nine days after the onset of the disease showed positive IgG titers. There was no correlation between death or viral load and IgM positivity [108]. In a study involving 46 confirmed cases of acute CCHFV infection in Kosovo, it was discovered that there was no correlation between the presence of IgM antibodies and clinical classification. Furthermore, only five out of the 34 patients who survived the disease exhibited IgG antibodies. [98].
Neutralizing antibodies (NAbs) usually appear by day 10 of the illness. are usually found at low levels in CCHF survivors, but are undetectable in fatal cases of CCHF [101,104]. This suggests that antibodies may play a crucial role in protecting individuals from lethal CCHFV infections. As of now, the study has revealed that mAbs and neutralizing mAbs specific to CCHFV have exclusively been derived from mice. [109]. Among the isolated antibodies, three Gc-specific NAbs have demonstrated the ability to neutralize multiple strains of the virus [109,110]. However, despite their cross-neutralizing activity, these NAbs have not been effective in providing protection against CCHF in experiments conducted on mouse models. These epitopes are not associated with the production of NAbs that contribute to the immune response against CCHF. Three Gc-specific NAbs cross-neutralize various strains but are ineffective in protecting mouse models of CCHF [109,111]. Studies have shown that although mAbs targeting pre-Gn and/or GP38 lack neutralizing activity, they can still provide pre-exposure protection in mice [109,110,111,112]. These findings suggest that non-NAbs may also have the ability to protect against CCHF through other mechanisms besides neutralization, as evidenced by their ability to confer protection in fatal CCHF challenges. The GP38-targeting mAbs effectiveness in providing protection depends on complement activity. This finding suggests that the antibody's effector functions, such as complement-mediated lysis and phagocytosis, play a crucial role in protecting against CCHFV [111]. However, a recent study illustrated the efficacy of bispecific antibodies (bsAbs) by incorporating variable domains from widely NAbs to boost their antiviral efficacy. The study found one bsAb to be particularly effective as it provided therapeutic protection against CCHFV with a single dose [113]. Thus, it is currently unclear whether there is any relationship between the neutralization antibody responses and positive disease outcomes.
Several studies have found a strong relationship between a high viral load and fatality rate, with some identifying a viral load of ≥ 10 8 copies/ml as a significant predictor of fatality [66,98,108,114]. While a reduction in CCHF viral load has been connected with the presence of antibodies in clinical infections, the production of antibodies is not always linked to the clearance of the virus. During the first week of infection, viral loads generally decrease irrespective of IgG levels, indicating the critical role of cellular immunity [98,108]. A study conducted with STAT1-deficient mice showed that CD4+ and CD8+ T cells were activated early against CCHFV infection [78]. Studies on infected mice have demonstrated that T-cells play a critical role in controlling CCHFV infection. They limit the virus's spread and prevent further infection by producing antiviral cytokines upon rapid activation [76,77].
The maintenance of CCHFV-specific T-cells for an extended period after infection suggests that memory T-cells may offer long-term immunity against the virus, responding rapidly to future exposure and serving as a lasting defense [115]. The study on a DNA vaccine highlights the significance of a TH1 response for effective protection [116]. It was shown that CD8+ T-cell responses in human survivors lasted for 13 years after acute infection. Most T-cell epitopes were found on the NP, but there were two instances of reactivity to GC-derived peptides. None of the epitopes were considered immunodominant. [117]. Lindquist et al. found that an IFN blockade antibody treatment in mice effectively controlled CCHFV through adaptive immune responses, specifically cytolytic T-cell activity, while avoiding liver damage, which is a common issue caused directly by CCHFV [80]. The removal of CD4+ or CD8+ T-cells in mice infected with the virus resulted in a higher mortality rate, highlighting the indispensability of these cells in survival [77]. Hawman et al. also found that the absence of CD4 T-cells eliminated the host's IFN-γ response and blocking IFN-γ signaling led to lethality in IFNAR-/- mice, suggesting that cellular immunity and type II IFN may control the CCHFV infection. Despite the existing research on the role of the adaptive immune response in CCHFV infection, further studies are needed to determine the immune responses and their effector functions essential for protection.
3. Animal Models for CCHF
CCHFV infections are asymptomatic in animals despite infecting many species and causing detectable viremia. With the exception of humans, only newborn mice and rats were suseptible to CCHFV among vertebrates. Disease signs and lethality are observed in human and newborn mice and rats through intracranial or intraperitoneal inoculation, making newborn rodents the first animal models for CCHF research [104,118]. However, they could not be used as models owing to their immature immune systems. Adult mice genetically deficient in type I IFN due to IFNα receptor or signal transducer and activator of transcription 1 (Stat1-/-) have been used as lethal models of CCHF disease [78,79]. Interferon-deficient mice typically develop a rapid onset of severe illness resulting in death within four days after postinoculation. These mice exhibit elevated levels of inflammatory cytokines, liver enzymes, liver lesions, and spleen lymphocyte depletion which is comparable to the symptoms observed in humans infected with CCHFV. However, immunocompromised mice may not be suitable for vaccine testing or fully understanding immune protection against CCHFV, as IFN-I signaling is critical in generating mature antigen-presenting cells, T and B cell responses, and memory T and B cell pools [78,79].
Recently, it has been created a novel murine system that utilizes the MAR1-5A3 antibody targeting IFN-I receptor A [80]. This method has been previously used to generate severe disease models for different viruses, and has proved to be effective in inducing lethal or severe CCHFV infection in mice by producing a temporary blockade of IFN-I [119,120]. The MAR1-5A3 antibody induces a temporary IFN-I blockade in mice, leading to a predictable and severe CCHFV infection [80,121,122]. A major benefit of this model is that it can mimic the same phenotype as an IFN-I receptor knockout animal in almost any wild-type or transgenic mouse, without requiring cross-breeding. This system enables vaccination of an animal with an intact immune system prior to virus exposure, reducing the potential impact of IFN-I disruption on secondary immune responses upon challenge. The disease resulting from the antibody-mediated IFN-I blockade model is almost identical to the disease observed in genetic knockout animals, both displaying equivalent mean times to death.
It was recently reported that a humanized mouse model has been developed for CCHFV [123]. They are excellent small-animal models that have been transplanted with human cells or tissues, additionally equipped with human transgenes. These animal models are particularly useful in analyzing human hematopoiesis and studying pathogens with a special affinity towards humans, especially those that have been conditioned to support the engraftment of human immune cells [124,125]. Spengler and colleagues created a new type of humanized mouse model by introducing human CD34+ stem cells into NOD-SCID-gamma Hu-NSG-SGM3 mice, which have severe immunodeficiency and impaired cytokine signaling resulting from the absence of mature T-cells, B-cells, and natural killer cells as well as deficiency in the common gamma chain. While exposure to CCHFV strains from Oman and Turkey showed diverse disease patterns, it was only the Turkish strain that resulted in fatal outcomes. The humanized mice, Hu-NSGTM-SGM3, exhibited severe histopathological changes in the brain and are a promising model for investigating the cellular and molecular factors contributing to CCHF-related neurological disease [123].
A mouse-adapted strain of CCHFV is capable of infecting immunocompetent wild type lab mice, causing significant pathology in the liver and spleen, high viral loads in multiple tissues, and inducing inflammatory cytokine production. Upon convalescence, robust humoral and cellular immunity was observed. Five coding mutations were detected in this virus through sequencing: two were found in the viral NP along with one mutation that also affected the viral NSs, one in the NSm, and two in the L protein (115). Although sex-linked differences have occasionally been reported for the CCHF patients [126,127,128], female mice showed greater resistance to severe disease than male mice, which exhibited a sex-linked bias in disease severity. The severe disease observed in male mice shared similar characteristics with poor outcomes in human CCHF cases, such as increased production of inflammatory cytokines, prolonged viraemia, and greater tissue pathology [115].
Previous studies have shown that the infection of African green monkeys, baboons, and patas monkeys with CCHFV was not successful [129,130,131]. However, Haddock et. al. have developed a cynomolgus macaque model of CCHF that was infected with a human clinical isolate of CCHFV (the Hoti strain) administering a high dose (5 log10 TCID50) through intravenous (IV) or combined IV and subcutaneous (SC) exposure. The infected animals showed viremia and developed a severe, occasionally lethal disease, with symptoms such as inflammatory immune responses, heightened liver enzymes, extended clotting times, thrombocytopenia, leukopenia, and fever. Four out of eight animals were humanely euthanized by day seven post-infection for meeting humane endpoint criteria. Animals that experienced severe disease had liver pathology, inflammatory cytokines, high viral loads, and coagulation disorders, which are similar hallmarks found in severe human disease [132]. In a study assessing the antiviral drug favipiravir's effectiveness for CCHFV-infected macaques, only one of eight animals in the placebo group met euthanasia criteria by day eight, while the other animals had moderate disease outcomes. [115]. Although this model has already been utilized to evaluate antivirals and vaccines, the disease outcomes in the model were variable.
Through the use of the cynomolgus macaque model, Cross et al. enhanced the understanding of CCHFV pathogenesis by comparing two distinct strains, Afg09-2990 and Hoti [133]. Two separate groups of cynomolgus macaques were intravenously injected with each strain, and their disease courses were closely monitored. All animals exhibited clinical illness, viremia, significant changes in clinical chemistry, hematology values, and serum cytokine profiles consistent with CCHF disease in humans. However, in contrast to the earlier study, all NHPs recovered fully, and none of the animals met the euthanasia criteria [133]. In a separate study conducted by Smith et al, it is worth mentioning that the persistence of CCHFV has been observed in the testes and latent tuberculosis granulomas of macaques infected with the virus. This observation suggests that the virus may have the ability to persist in immune-privileged sites [134]. Although the factors that contribute to disease variability are not yet fully understood, the use of cynomolgus macaques as a model has proven useful in preclinical evaluation of anti-CCHFV therapeutics and vaccines.
4. Vaccine Targets of CCHF
4.1. The nucleoprotein (NP)
The NP is an essential component of CCHFV, as it plays a crucial role in the viral life cycle. The primary function of the NP is to encapsidate the viral RNA and form the ribonucleoprotein (RNP) complex (Figure 1a). The RNP complex is the basic unit of the virus and is required for viral replication and transcription. CCHFV NP are involved in a range of important functions, many of which relate to interactions with components of the host cell. These interactions may facilitate the transport of viral RNPs, the evasion of the host's immune system, and the regulation of apoptosis [3,135]. The NP is a promising vaccine target for CCHF due to its abundance and high immunogenicity [136]. The NP contains in both B and T cell epitopes, making it capable of stimulating both the humoral and cellular immune responses [117]. The genetic diversity and multiple distinct lineages of CCHFV existing in different global regions are crucial factors to consider for the development of vaccines. The NP of CCHFV exhibits a remarkable degree of conservation across various strains of the virus, suggesting NP could provide protection against multiple strains of the virus [137,138].
4.2. The Glycoproteins (GPC)
The CCHFV M segment encodes a polyprotein that undergoes post-translational processing to form intermediate glycoprotein precursors, PreGn (140 kDa) and PreGc (85 kDa) (Figure 1b). These intermediate glycoproteins are subsequently processed to yield envelope glycoproteins (Gn and Gc), nonstructural M protein (NSM), as well as secreted non-structural proteins (GP160, GP85, and GP38) and mucin-like domain (MLD) [5,6,109,139] (Figure 1b). Gn and Gc glycoproteins are essential for virus attachment and envelope fusion with host cells [140,141]. One of the most promising approaches to developing a CCHFV vaccine is the use of glycoproteins Gn and Gc as antigens. These glycoproteins are located on the surface of the virus and are responsible for facilitating viral entry into host cells (Figure 1a). They are also the primary targets of the immune response during CCHFV infection, making them ideal candidates for vaccine development. Early studies indicates that antibodies that target Gc glycoprotein in CCHF virus have neutralizing capabilities, while no such neutralizing effect has been observed for Gn-targeting antibodies [109]. It has been observed that neonatal mice can be protected from lethal CCHFV infection by both neutralizing and non-neutralizing mAbs antibodies, but these findings are limited due to the fact that neonatal mice do not accurately represent CCHF disease, thereby making it difficult to interpret the results. However, a recent study examined the effectiveness of murine mAbs in protecting adult mice from CCHFV infection. The findings showed that non- NAbs targeting the GP38 protein provided protection against lethal CCHFV infection in mature animals [111]. The study highlights the potential of developing antibody-based CCHFV countermeasures.
The diversity of the M segment of CCHFV, particularly the region encoding the nonstructural proteins, has long been suspected to impact crossreactivity and ultimately, the neutralization ability against different strains [72]. Understanding this diversity is crucial for developing vaccines, antiviral therapies, and understanding the virus's evolution and pathogenesis. Further studies will be needed to fully elucidate the mechanisms underlying this diversity and its impact on viral pathogenesis.
5. Platforms for CCHF vaccine candidates
Animal models for CCHFV have historically been limited by the lack of suitable animal hosts. Prior to the discovery of CCHFV animal models, there were limited attempts to develop a vaccine for CCHF, and efficacy studies were not possible. However, in recent years, researchers have developed new animal models that closely mimic the disease in humans, providing a more accurate representation of the virus. Also, recent advances in biochemical and molecular techniques have enabled researchers to employ different vaccine platforms for developing CCHFV vaccine candidates (Figure 1c and Table 1). Below are summarized the various approaches and platforms used in vaccine development.
5.1. Inactivated Vaccines
In the 1960s, Soviet scientists developed an experimental CCHF vaccine using brain tissue from infected newborn laboratory mice and rats. The vaccine was developed by cultivating CCHFV in suckling mouse brain and subsequently inactivating the virus through chloroform treatment and heat exposure at 58°C. The inactivated virus was subsequently absorbed onto aluminum hydroxide (Al(OH)3) [142]. It was approved and licensed in 1974 in Bulgaria and have been used in military and medical personnel and people living in endemic regions. To date, the only tested CCHFV vaccine for humans is the suckling mouse brain-derived vaccine, utilized exclusively in Bulgaria. Between 1953-1996, the Bulgarian Ministry of Health observed a reduction in CCHF cases from 1105 to 279 [143], however, vaccination may not solely account for this reduction as factors such as changes to ecology and epidemiology may have also contributed. While data on the immunogenicity of the mouse brain-derived vaccine is limited. In 2012, a study showed that repeated vaccinations among healthy volunteers were associated with high levels of CCHFV antibodies, anti-CCHFV specific T-cell activity, and low levels of neutralization activity (Table 1) [144]. However, this vaccine is only authorized for use in Bulgaria and not in other countries with at-risk populations due to potential allergic and autoimmune reactions and is unlikely to receive international approval due to safety issues and scalability limitations.
Our group developed a purified and formalin-inactivated CCHF vaccine candidate derived from cell culture in 2015. The inactivated vaccine was prepared by growing the CCHFV Turkey-Kelkit06 strain in cell culture, harvesting, and inactivating it with formaldehyde. Alum adjuvant was added, and administered through three inoculations of 5, 20, and 40 μg dosages to IFNAR-/-. Two weeks after the last immunization, the mice were challenged with high lethal dose (1000 PPFU) of CCHFV Turkey-Kelkit06 strain to test the vaccine's efficacy. Immunization with the cell culture-based CCHF vaccine at doses of 5, 20, and 40 μg provided partial protection ranging from 60% to 80%, with a significant delay in time to death (Table 1) [145]. Until recently, there was no data available on the efficacy of the mouse brain-derived vaccine. Our recent study using the immune-suppressed (IS) mouse model investigated the potential efficacy of mouse brain-derived vaccine. The challenge studies showed that mouse brain-derived vaccine provided complete protection but the cell culture-based vaccine more effectively stimulated CCHFV-specific antibody and T cell responses (Table 1) [121]. In a recent study, Engin et al investigated the IgG and neutralizing antibody titers over a duration of one year in BALB/c mice following vaccination with the the cell culture-based and mouse brain-derived vaccines. Consistent with previous finding, the cell-culture-based vaccine resulted in higher IgG and neutralizing antibody titers than the mouse-brain-based vaccine at all measured time points [146].
5.2. Subunit Vaccines
Subunit vaccines are a safe and effective vaccine preparation strategy that relies on viral proteins to generate an immune response without eliciting antibodies against unrelated antigens or viral particles. The CCHFV envelope glycoproteins Gn and Gc were expressed using insect expression technology in Drosophila Schneider 2 (S2) cells. Adjuvanted Gn or Gc ectodomains were able to generate NAbs, but they did not provide protection to STAT-1 knockout mice after challenge (Table 1) [147]. Xia et al used an affinity purification method based on a gram-positive enhancer matrix-protein anchor (GEM-PA) surface display system to exhibit eGN, eGC, and NAb epitopes (NAb, aa1443 and 1566 of the M gene in IbAr10200 strain) of glycoprotein. The immunogenicity of these epitopes was evaluated in BALB/c mice [148], but the efficacy of the vaccine candidates was not assessed. In the study conducted in 2023, three vaccine candidates (rvAc-Gn, rvAc-Np, and rvAc-Gn-Np) expressing CCHFV's glycoprotein Gn and NP on a baculovirus using the insect baculovirus vector expression system were evaluated for immunogenicity in mice. Results indicated limited immunogenicity for rvAc-Gn-NP, while rvAc-Gn elicited cellular and humoral immunity [149]. Once again, no vaccine effectiveness study has been conducted.
Plant-based vaccines offer scalable and cost-effective approaches for foreign gene expression in transgenic plants compared to conventional methods. In recent years, there has been considerable growth in producing human vaccine candidates in plants that address different targets [150]. Using plant cloning vectors, researchers introduced the Gn and Gc coding regions of CCHFV into transgenic tobacco plants for expression. Feeding the roots and leaves from these plants to mice resulted in oral/mucosal immunization that induced CCHFV-specific anti-Gn/Gc IgG and IgA antibodies in their serum and fecal material [151]. Nevertheless, the neutralizing ability of these antibodies was not evaluated, and there were no challenge studies conducted.
5.3. DNA Vaccines
DNA vaccines are a safe, efficient, and cost-effective means of inducing an immune response. Recombinant DNA is used to express antigens in antigen presenting cells, inducing an immune response similar to viral infections. The delivered DNA can be translated into a desired protein in the cell cytoplasm, and the resulting peptide fragments can be presented by MHC class I and/or II molecules, promoting innate and adaptive immune responses. DNA vaccines have several advantages over other vaccines, including ease of manufacturing, improved safety, and simplified production. While there are some potential drawbacks to this approach, such as autoimmune responses and low immunogenicity, these can be addressed through innovative design [152,153]. Overall, DNA-based vaccines have significant potential for the development of vaccines against CCHFV. The first DNA vaccine development study against CCHFV date back to 2006. A DNA-based vector was created to deliver the GPC of IbAr 10200 CCHFV. Vaccination alone or combined with other DNA vaccines against Rift Valley fever virus, tick-borne encephalitis virus, and Hantaan virus showed NAbs detection in only 50% of vaccinated mice. Cell-mediated immune responses were not assessed and challenge studies were not conducted due to the unavailability of an appropriate animal model during that time [154].
A DNA vector encoding mature CCHFV envelope glycoproteins (Gn and Gc) as well as the NP of the IbAr 10200 strain was intradermally immunized three times. IFNAR-/- mice were successfully protected from lethal CCHFV challenge through the successful elicitation of both antibody and T cell immune responses [116]. In the same study, mice that received a VLP construct showed higher in vitro NAbs compared to the CCHFV DNA vaccine, although their protection was only partial. These results suggest that NAbs alone may not be sufficient to provide protection against CCHFV.
Garrison et al. evaluated the immunogenicity and protective efficacy of a DNA vaccine that expresses the M-segment glycoprotein precursor gene of CCHFV in two lethal mouse models of disease: IFNAR-/- mice, and a transiently immune suppressed (IS) mouse model [155]. In the study, the vaccine was administered via muscle electroporation at a dose of 25 μg, which stimulated a robust humoral immune response. After three vaccinations, neutralizing titers were detected in both mouse models. Both mouse models exhibited predominantly Th1 antibody responses, but the IS model had a significantly lower Th1/Th2 ratio, indicating a more balanced antibody response in immunocompetent mice. Although complete protection was not achieved in either mouse model, the survival rate was higher in the IFNAR-/- model (71.4%) compared to the IS model (60%) (Table 1).
The same group conducted a recent study in 2021 to evaluate the effectiveness of two CCHFV DNA vaccines, namely CCHFV-M10200 and CCHFV-MAfg09, in mice [156]. The vaccine dose for CCHFV-M10200 was increased to 50 μg, administered three times, three weeks apart, resulting in 100% protection after a final vaccination and challenge with CCHFV-IbAr 10200. Challenge studies also showed that CCHFV-MAfg09 provided complete protection against homologous CCHFV-Afg09-2990 challenge, while CCHFV-M10200 provided 80% protection against heterologous CCHFV-Afg09-2990 challenge (Table 1). They also found that a DNA vaccine expressing the GP38 region of CCHFV-IbAr 10200 partially protected against homologous challenge, and high levels of anti-GP38 antibodies provided protection from CCHFV challenge, suggesting genetic diversity in the GP38 region is responsible for the diminished protection provided by the vaccines against heterologous challenge.
The study assessed the protection potential of a DNA vector expressing CCHFV's NP and CD24 in mice, discovering that CD24 led to induce immune response by regulating B and T cell proliferation [157]. CCHFV's NP is a promising candidate for vaccination due to its ability to stimulate a balanced immune response. A study identified a complete protection in IFNAR-/- mice with a DNA vaccine based on the nucleocapsid (Table 1). These results showed that the introduction of appropriate adjuvants for DNA vaccine immunization is a promising approach to enhance the immune response and efficacy of DNA vaccines.
Hu et al. recently developed three DNA vaccines that encode the NP, glycoprotein N-terminal (Gn) and C-terminal (Gc) of CCHFV [158]. These vaccines are fused with lysosome-associated membrane protein 1 (LAMP1) and have been tested for their immunogenicity and protective efficacy in a human MHC (HLA-A11/DR1) transgenic mouse model. Vaccination of mice with pVAX-LAMP1-CCHFV-NP was the most effective vaccine, inducing balanced Th1 and Th2 responses and providing effective protection against CCHFV transcription and tecVLPs infection. In contrast, pVAX-LAMP1-CCHFV-Gc elicited mainly specific anti-Gc and NAbs, while pVAX-LAMP1-CCHFV-Gn provided inadequate protection against CCHFV tecVLPs infection.
A DNA-based vaccine was tested in cynomolgus macaque model for CCHFV [159]. The vaccine contains plasmid-expressed CCHFV strain Hoti NP and GPC, delivered through intramuscular injections with in vivo electroporation. The vaccine was well-tolerated and induced CCHFV-specific antibody and T-cell responses. However, the vaccined macaques produced low levels of NAbs against CCHFV (Table 1). They also showed reduced viremia, clinical signs, and pathology following CCHFV challenge compared to unvaccinated controls. The DNA vaccine is the first to show efficacy in a non-human primate model of CCHF and supports the vaccine's advancement into human clinical trials.
5.4. Virus Like Replicon Vaccines
Virus like replicon vaccines (VRP) are virus-like particles that enter cells, undergo limited transcription and translation to synthesize proteins, but don't produce infectious progeny. They are engineered virus genomes to express multiple proteins and lead to a strong immune response as it produces a high level of antigen expression in a single round of infection. The VRP vaccine lacks the M segment, which limits replication to one cycle and includes S and L genome segments from the IbAr10200 strain. However, co-transfection with a plasmid containing the optimized GPC of the Oman-98 strain enhances VRP generation and amplification for optimized cell entry. When tested in IFNAR-/- mouse model, the VRP vaccine, based on IbAr 10200 strain with the GPC sequence from the Oman-1998 strain, provides complete protection against lethal challenge following a single high dose (105 TCID50 of VRP) subcutaneous vaccination. However, when a low dose (103 TCID50 of VRP) of the vaccine was given, it was able to protect seven out of the nine mice (Table 1) [160]. This demonstrates that even a lower dose of the vaccine can still provide significant protection. Based on a related study, it has been discovered that the VRP candidate vaccine offers heterologous protection against CCHF disease. This protection was observed in IFNAR-/- mice that were vaccinated with a single dose of VRP and subsequently challenged with CCHFV-Turkey and Oman-97 strains. (Table 1) [161]. In a recent study, the efficacy of vaccinating IFNAR-/- mice at different time intervals before exposure to the CCHFV was evaluated. It was observed that all non-vaccinated mice succumbed to the infection within 8 days, whereas mice vaccinated 14 or 7 days before the virus challenge were fully protected, while those vaccinated 3 days before showed symptoms but later recovered (Table 1). This findings suggest that the VRP vaccine could be used in shorter vaccination protocols to protect against severe disease outcomes [162].
Flavivirus replicons, which are viral self-replicating sub-genomic replicons, are a potent tool for studying viral genome replication, antiviral screening, and creating chimeric vaccines [163]. Recently, the Kunjin strain of West Nile Virus (WNV) was used to create flavivirus-based replicon virus-like particles for CCHF vaccine development. The C-prM-E genes in the WNVKUN replicon replaced with the genes encoding the Gn and Gc glycoproteins of CCHFV to generate a replicon capable of expressing CCHFV proteins. The CCHFV Gn and Gc glycoproteins were expressed in RVP vaccine platform however, it induced a weak antibody response against them. T cell responses were not measured, and its protective efficacy in vivo was not assessed (Table 1) [164].
Alpha vector systems have emerged as a promising platform for developing prophylactic and therapeutic vaccines against infectious diseases [165]. A vaccine for CCHFV has been developed using the DNA-based Sindbis replicon platform. The vaccine, pSinCCHF-52S, replaces the structural proteins of Sindbis virus with the NP gene of CCHFV, inducing NP-specific antibody and T cell responses with a Th1 skew [166]. However, the vaccine has not yet been tested for efficacy against CCHFV in a challenge model. The Venezuelan Equine Encephalitis Virus (VEEV) RNA replicon is another Alpha vector vaccine platform that has been used to produce potential CCHFV vaccines for NP (repNP), GPC (repGPC), and a combination of both (repNP + repGPC) [167]. The repNP vaccine elicited a strong antibody response but a weak T-cell response, whereas the repGPC vaccine stimulated a weak antibody response but a strong T-cell response. The repNP and repNP + repGPC vaccines provided the complete protection after challenge with a heterologous strain of CCHFV in mice treated with an anti-IFNAR antibody blockade, whereas only 40% of mice immunized with repGPC were protected (Table 1).
5.5. mRNA Vaccines
The progress made in mRNA design, nucleic acid delivery technology, and the identification of new antigen targets has elevated mRNA vaccines to an exceptional tool for fighting emerging infectious diseases [168]. During the Covid-19 pandemic, mRNA-based vaccines have proven highly effective against the virus and are considered a robust alternative to traditional vaccines due to their potency, safety, and efficiency [168,169]. The study evaluated the efficacy of a conventional mRNA vaccine expressing NP from the non-optimized S segment of the Ank-2 strain of CCHFV. Single and booster doses were given, and challenge assays showed a 100% protection rate in the booster group and 50% in the single dose group, indicating lower effectiveness with a single dose (Table 1) [170].
In a recent study, Appelberg et al. designed two nucleoside-modified mRNA-lipid nanoparticle (LNP) vaccines that encoded either the CCHFV IbAr10200 glycoproteins (GcGn) or NP and tested in both immunocompromised and immunocompetent mice. Challenge studies demonstrated complete protection for IFNAR-/- mice vaccinated with either GnGc mRNA-LNP or NP mRNA-LNP (Table 1) [171]. The study does not provide conclusive evidence that a cellular immune response alone can protect against CCHFV, and it remains uncertain if antibodies are always necessary. However, the inclusion of NP in the vaccine can improve the protection against different strains of CCHFV. Further studies are necessary to determine if cellular immunity through NP mRNA-LNP is enough to prevent CCHFV infection.
5.6. Viral Vector Vaccines
Recombinant viral vectors have been extensively studied as a promising vaccine platform due to their ability to express the antigens, stimulate antigen-specific immune responses, and generate potent antibody titers, all without the need for external adjuvants [172,173]. Multiple vaccine candidates have been developed for CCHF using various viral vectors, including the modified Vaccinia Ankara virus (MVA) [174,175,176], recombinant adenovirus type 5 (AdHu5) [177,178], recombinant chimpanzee adenovirus (ChAdOx2) [179], recombinant vesicular stomatitis virus (rVSV) [180], and recombinant bovine herpesvirus type 4 (BoHV-4) [178].
The MVA vector was used to create CCHF MVA-GP vaccine candidate, encodes the entire M segment open reading frame (ORF) of the IbAr 10200 CCHFV strain. Following intramuscular administration twice, the MVA-GP vaccine has shown the capability to induce NAbs and T cell responses leading to complete protection against intradermal lethal challenge in IFNAR-/- mice (Table 1) [174]. Dowall et al. demonstrated that MVA-based vaccines effectively stimulate both immune system arms, essential in eliciting protective effects against lethal CCHFV challenge [176].
A recent study found that the ChAdOx2 CCHF vaccine which employs a recombinant chimpanzee adenovirus to express the entire M segment of CCHFV, produced comparable results to a study using the MVA-GP vaccine. This study has been conducted to examine the immunogenicity and protection provided by ChAdOx2 CCHF vaccine either alone or combined with the MVA-GP CCHF vaccine. They found strong antibody responses and IFN-γ-mediated cellular immunity upon administration of various vaccine combinations in both immunocompetent BALB/c and immune-deficient A129 mice. The study demonstrated that a single dose of ChAdOx2 CCHF vaccine or homologous/heterologous prime-boost vaccination regimens resulted in full protection against CCHFV-induced disease in the A129 lethal mouse model (Table 1) [179].
The recombinant vesicular stomatitis virus vector vaccine (rVSV) for CCHF is another example that utilizes the GPC antigen. In a STAT-1/mouse model, a replication-competent recombinant rVSV that encoded the CCHFV GPC gene of the IbAr 10200 CCHFV strain provided complete protection following a single intraperitoneal immunization. Conversely, a replication-deficient rVSV construct failed to provide protection against a lethal virus challenge administered intraperitoneally [180].
The MVA-NP vaccine candidate, which encodes the S segment ORF of the IbAr 10200 CCHFV strain, failed to provide protection after a lethal challenge, even though it was capable of activating both arms of the immune system against CCHFV [175]. On the contrast to the the MVA-NP vaccine candidate, Zivcec et al. developed a promising NP-based candidate vaccine (Ad-N) that uses human adenovirus 5 to encode the NP of the CCHFV strain IbAr 10200 [177]. The Ad-NP was able to provide partial protection in IFNAR-/- mice against virus challenge. Additionally, a prime-boost strategy was used which resulted in enhanced protection against the virus and reduced clinical symptoms compared to single-dose vaccination methods (Table 1). Another study confirms that using recombinant AdV-5 encoding NP from CCHFV can protect IFNα/β/γR- mice after CCHFV challenge (Table 1). Furthermore, the antibody passive transfer and T cell adoptive transfer experiments demonstrated a 50% survival rate of mice after a lethal CCHFV challenge [178].
BoHV-4 is another viral vector that has been used in the development of a vaccine for CCHFV. BoHV-4 possesses several characteristics that make it an ideal vaccine vector, such as its ability to stably express foreign genes, replication capability in foreign hosts, and minimal pathogenicity in various hosts. When used in combination with a prime and boost strategy, the BoHV-4 vector encoding the full-length NP of CCHFV provided complete protection after lethal challenge (Table 1). Additionaly, partial protection was observed in experiments involving antibody passive transfer and T cell adoptive transfer [178].

Table 1.
This is a concise summary of different vaccine design platforms for the Crimean-Congo hemorrhagic fever virus (CCHFV).
Table 1.
This is a concise summary of different vaccine design platforms for the Crimean-Congo hemorrhagic fever virus (CCHFV).
| Vaccine design platforms | Strain name and types of antigen | Animal Models |
Doses and vaccination strategies | Spesific antibody response | Neutralizing antibody response | T cell immune response | CCHFV challenge |
Survival rate% | References |
|---|---|---|---|---|---|---|---|---|---|
| Inactivated vaccines | Whole CCHFV from mouse-brain | NE | Several thousand people took repeated vaccination | NE | Yes | NE | NE | NE | [142] |
| Bulgarian V42/81 strain; CCHFV whole antigen from mouse brain | NE | One group received a single dose, while the second group received four doses. | Yes | Yes | Yes | NE | NE | [143,144] | |
| Turkey-Kelkit06 strain; CCHFV whole antigen from cell culture (Vero-E6) | IFNAR-/- mice and BALB/c mice with transiently immune suppressed by mAb-5A3 | IFNAR-/- mice: Administered (IP), at doses 5 μg, 20 μg and 40 μg of inactivated vaccine, on days 0, 21 and 42. BALB/c mice: Administered (IP), at doses 5 μg, 10 μg and 20 μg of inactivated vaccine, on days 0, 14 and 27. |
Yes | Yes | Yes | Turkey-Kelkit06 strain; 1000FFU (IFNAR-/-) and 100FFU (Balb/C) |
80% protected (IFNAR-/-), 100% protected (Balb/C) | [121,145,146] | |
| Subunit vaccines | IbAr10200 strain; CCHFV Gn and Gc ectodomain | STAT1 knockout mice | Administered (IP), at doses 1.4 μg, 7.5 μg and 15 μg of Gn and Gc, on days 0 and 21 |
NE | Yes | NE | IbAr10200 strain; 100PFU | Not protected | [147] |
| IbAr10200 strain; extracellular region of Gn (eGn), extracellular region of Gc with truncation of C terminal (eGc), neutralizing antibody region of Gc (NAB) | BALB/c mice | Administered (SC), at doses1 μg, 5 μg and 20 μg of surface display protein of G-GP with eGn, eGc, at weeks 0, 3, 6 and 9 |
Yes | Yes | Yes | NE | NE | [148] | |
| Chinese Xinjiang strain HANM18; Gn and NP from CCHFV expressed in baculovirus expression system as rvAc-Gn, rvAc-NP and rvAc-Gn-NP | BALB/c mice | 107 PFU, on days 0, 14 and 28 |
Yes | NE | Yes | NE | NE | [149] | |
| CCHFV Iranian strain; Gn and Gc expressed in transgenic tobacco leaves | BALB/c mice | Feeding (leaves), at dose 10 μg of Gc/Gn, at weeks 0, 1, 2 and 3 Feeding (roots), at dose 10 μg of Gc/Gn, at weeks 0, 1, 2, 3 |
Yes | NE | NE | NE | NE | [151] | |
| DNA vaccines | IbAr10200 strain; GPC | BALB/c mice | Administered (gene gun), at dose 2.5 μg of the vaccine from each CCHFV+ RVFV + HTNV+ TBEV as total 10 μg, alone or combined, at weeks 0, 4 and 8 |
NE | Yes | NE | NE | NE | [154] |
| IbAr10200 strain; Ubiquitin linked version of Gn, Gc and NP | IFNAR-/- mice | Administered (ID), at dose 15 μg of the vaccine three times with 4 weeks interval between 1st and 2nd dose and three weeks interval between 2nd and 3rd dose | Yes | Yes | Yes | IbAr10200 strain; 400FFU | 100% protected | [116] | |
| IbAr10200 strain; GPC | IFNAR-/- mice and C57BL/6 mice with transiently immune suppressed by mAb-5A3 |
Administered (IM), at dose 25 μg of GPC, at weeks 0, 3 and 6 |
Yes | Yes | NE | IbAr10200 strain; 100PFU | Protective efficacy 71,4% for IFNAR-/- mice and 60% for transiently immune suppressed mice | [155] | |
| IbAr 10200 strain; GPC from | C57BL/6 mice with transiently immune suppressed by mAb-5A3 | Administered (IM), at dose 50 μg of GPC, at weeks 0, 3 and 6 |
Yes | NE | Yes | IbAr10200 and Afg09-2990 strains; 100PFU |
100% protected against IbAr10200, 80% protected against Afg09-2990 |
[156] | |
| Ank-2 strain; NP with CD24 |
BALB/c for immunological responses and IFNAR-/- mice for challenge studies | Administered (IM), at dose 50 μg of pV-N13 and 40 μg of pV-N13 with 10 μg of CD24, on days 0 and 14 |
Yes | NE | Yes | Ank-2 strain; 1000 TCID50 |
100% protected | [157] | |
| IbAr10200 strain; NP, N terminal Gn and C terminal Gc fused with LAMP1 to generate three candidate vaccines | Human MHC (HLA-A11/DR1) | Administered (IM), at dose 70 μg pVAX-LAMP1-NP, 70 μg pVAX-LAMP1-Gn, 70 μg pVAX-LAMP1-Gc, at weeks 0, 3 and 6 |
Yes | Yes | Yes | IbAr10200 strain; 100 TCID50 CCHFV tecVLPS |
Instead of measuring survival percentages, NanoLuc activities measured, NP had the lowest levels of NanoLuc activities in their liver, spleen, and kidney | [158] | |
| Hoti strain; Ubiquitin fused with GPC and NP |
Cynomolgus macaque | Administered (IM), at dose1 mg of pNP + 1 mg of pGPC, on days 0, 21 and 42 |
Yes | Poor neutralization acitivity | Yes | Hoti strain; 1X105 TCID50 |
Survival percentage was not assessed due to the non-uniform lethality of CCHFV in this animal model | [159] | |
| Viral like replicon particles (VRP) vaccines | IbAr10200 strain; S and L segment, Oman 98 strain; M segment |
IFNAR-/- mice | Administered (SC), at dose 105 TCID50 or 103 TCID50 of VRPs, single vaccination |
Yes | NE | NE | IbAr1020 strain; 100 TCID50 |
Low dose showed 77% protection and high dose showed 100% protection | [160] |
| IbAr10200 strain; S and L segment, Oman 98 strain; M segment |
IFNAR-/- mice | Administered (SC), at dose 1 × 105 TCID50 VRPs, single vaccination |
Yes | NE | NE | Oman 97 strain and CCHFV-Turkey strain; 100 TCID50 |
100% protected | [161] | |
| IbAr10200 strain; S and L segment, Oman 98 strain; M segment |
IFNAR-/- mice | Administered (SC), at dose 1 × 105 TCID50 VRPs, single vaccination |
NE | NE | NE | CCHFV-Turkey strain; 100 TCID50 |
After 14,7, and 3days following vaccination; 100% protected |
[162] | |
| Hoti strain; Gn and Gc |
BALB/c mice | Administered (SC), at dose 106 particles, at weeks 0, 2 and 5 |
Yes | Yes | NE | NE | NE | [164] | |
| SPU 187/90 strain; NP | NIH-III Heterozygous mice strain | Administered (IM), at dose 100 μg Sindbis replicon (expressing NP), on days 0, 21 and 42 Administered (IM), at dose 100 μg of Sindbis replicon (expressing NP) + 50 μg Poly (I:C), on days 0, 21 and 42 |
Yes | NE | Yes | NE | NE | [166] | |
| Hoti strain; NP and GPC |
C57BL/6 mice with transiently immune suppressed by mAb-5A3 | Administered (IM), at dose 2.5 μg of NP, 2.5 μ GPC and 5 μg of NP+GPC, on days 0 and 28 |
Yes | Poor neutralization activity | Yes | UG3010; 100TCID50 | 100% protected for NP and NP+GPC, 40% protected for GPC |
[167] | |
| mRNA vaccines | Ank-2 strain; NP |
C57BL/6 mice for immunogenicity and IFNAR-/- mice for challenge studies |
Administered (IM), at dose 25 μg of NP, at weeks 0 and 2 |
Yes | No | Yes | Ank-2 strain; 100LD50 |
Double dose immunized group showed 100% protection | [170] |
| IbAr10200 strain; NP, Gn and Gc | IFNAR-/- mice | Administered (ID), at doses 10 μg of NP, 10 μg of Gn, 10 μg of Gc and 20 μg of NP+ Gn+ Gc, at weeks 0 and 3 |
Yes | Yes | Yes | IbAr10200 strain; 400FFU |
100% protected | [171] | |
| Viral vector-based vaccines | IbAr10200 strain; GPC expressed in modified vaccinia virus Ankara | IFNAR-/- mice | Administered (IM), at dose 107 PFU, at weeks 0 and 2 |
Yes | NE | Yes | IbAr10200 strain; 200TCID50 |
100% protected | [174] |
| IbAr10200 strain; NP expressed in modified vaccinia virus Ankara | IFNAR-/- mice | Administered (IM), at dose 107 PFU, at weeks 0 and 2 |
Yes | NE% | Yes | IbAr10200 strain; 200TCID50 | Not protected | [175] | |
| IbAr10200 strain; NP expressed in Adenovirus type 5 | IFNAR-/- mice | Administered (IM), 1.25x107 PFU for first dose at day 0, Administered (IN), 1x108 PFU for second dose at day 28 |
Yes | NE | NE | IbAr10200 strain; 1000LD50 | 78% protected | [177] | |
| Ank-2 strain; NP expressed in Bovine Herpesvirus Type 4 (BoHV-4) |
BALB/c mice for serological assay and IFNAR-/- mice for challenge studies | Administered (IP), at dose 100 TCID50, at weeks 0 and 2 |
Yes | No | Yes | Ank-2 strain 100LD50 | 100% protected | [178] | |
| IbAr10200 strain; GPC expressed in ChAdOx2 (Chimpanzee Adenovirus) | BALB/c mice for immunogenicity and IFNAR-/- mice for challenge studies | Administered (IM), at dose 5X107 infectious unit (IU), on days 0 and 14 |
Yes | Yes | Yes | IbAr10200 strain; 200FFU |
100% protected | [179] | |
| IbAr10200 strain; GPC was expressed in Vesicular Stomatitis Virus expression system | STAT1 knock out mice | Administered (IP), at dose 107 PFU on days 0 and 21 |
Yes | Yes | NE | Turkey2004 strain; 50PFU |
100% protected | [180] |
NP-Nucleoprotein; GPC- Glycoprotein Complex; NE-Not evaluated; IM- Intramuscular; SC- Subcutaneous; ID- Intradermal; IP- Intraperitoneal; IN- Intranasal; PFU- Plaque Forming Unit; TCID50- Tissue Culture Infectious Dose 50; FFU- Fluorescence Focus Unit; LD50- Lethal dose 50; IFN-γ- Interferon Gamma; IL- Interleukin, CD24- Cluster of differentiation 24; RVFV- Rift Valley Fiver Virus; HTNV-Hantaan Virus; TBEV- Tick-Borne Encephalitis Virus.
6. Conclusions
CCHF is a severe viral infection that poses a significant threat to public health. The high fatality rate and the absence of a specific treatment or vaccine make it crucial to understand the pathogenesis and immunology of CCHF to develop effective countermeasures. The use of animal models, including interferon-deficient mice and the Cynomolgus macaque, has significantly advanced the study of the disease's mechanisms and potential treatments. Preclinical vaccine studies for various vaccine platforms have shown promising results. However, the genetic variability of CCHFV makes it challenging to develop a vaccine that can provide broad protection against all strains of the virus. Additionally, most CCHF vaccine studies have primarily employed the prototype IbAr 10200 CCHFV strain, isolated in ticks, with uncertain human virulence. To address these challenges, heterologous challenge studies are required to develop more reliable vaccines that can provide broad protection against different strains of the CCHFV. Although preclinical studies show potential, these vaccines have not yet been tested in humans, and it remains to be determined if these findings can be successfully applied in human clinical trials.
Funding
This research received no external funding
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| CCHFV | Crimean-Congo hemorrhagic fever virus |
| S | small |
| M | medium |
| L | large |
| RdRp | RNA-dependent RNA polymerase |
| NP | nucleoprotein |
| GPC | glycoprotein precursor |
| OTU | ovarian tumor |
| IFN | interferon |
| ISGs | interferon-stimulated genes |
| IFNAR-/- | IFN1 deficient mice |
| mAb | monoclonal antibody |
| TLR | toll-like receptor |
| PRRs | pattern recognition receptors |
| PAMPs | pathogen-associated molecular patterns |
| DIC | disseminated intravascular coagulation |
| Nabs | neutralizing antibodies |
| bsAbs | bispecific antibodies |
| Stat1−/− | signal transducer and activator of transcription 1 |
| IV | intravenous |
| SC | subcutaneous |
| NSM | nonstructural M protein |
| MLD | mucin-like domain |
| Al(OH)3 | aluminum hydroxide |
| IS | immune-suppressed |
| S2 | drosophila schneider 2 |
| GEM-PA | enhancer matrix-protein anchor |
| LAMP1 | lysosome-associated membrane protein 1 |
| VRP | Virus like replicon |
| VEEV | Venezuelan equine encephalitis virus |
| WNV | West Nile virus |
| LNP | lipid nanoparticle |
| AdHu5 | adenovirus type 5 |
| ChAdOx2 | chimpanzee adenovirus |
| rVSV | recombinant vesicular stomatitis virus |
| BoHV-4 | bovine herpesvirus type 4 |
| ORF | open reading frame |
References
- Abudurexiti, A.; Adkins, S.; Alioto, D.; Alkhovsky, S.V.; Avšič-Županc, T.; Ballinger, M.J.; Bente, D. A.; Beer, M.; Bergeron, É.; Blair, C.D.; et al. Taxonomy of the order Bunyavirales: update 2019. Arch of Virol 1949–1965 2019, 164. [Google Scholar] [CrossRef] [PubMed]
- Garrison, A.R.; Alkhovsky Альхoвский Сергей Владимирoвич, S.V.; Avšič-Županc, T.; Bente, D. A.; Bergeron, É.; Burt, F.; Di Paola, N.; Ergünay, K.; Hewson, R.; Kuhn, J.H.; et al. ICTV virus taxonomy profile: Nairoviridae. J Gen Virol 2020, 101, 798–799. [Google Scholar] [CrossRef] [PubMed]
- Carter, S.D.; Surtees, R.; Walter, C.T.; Ariza, A.; Bergeron, É.; Nichol, S.T.; Hiscox, J.A.; Edwards, T.A.; Barr, J.N. Structure, function, and evolution of the Crimean-Congo hemorrhagic fever virus nucleocapsid protein. J Virol 2012, 86, 10914–10923. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Wang, W.; Ji, W.; Deng, M.; Sun, Y.; Zhou, H.; Yang, C.; Deng, F.; Wang, H.; Hu, Z.; et al. Crimean–Congo hemorrhagic fever virus nucleoprotein reveals endonuclease activity in bunyaviruses. P Proc Natl Acad Sci USA. 2012, 109, 5046–5051. [Google Scholar] [CrossRef]
- Sanchez, A.J.; Vincent, M.J.; Nichol, S.T. ; Characterization of the glycoproteins of Crimean-Congo hemorrhagic fever virus. J Virol 2002, 76, 7263–7275. [Google Scholar] [CrossRef]
- Sanchez, A.J.; Vincent, M.J.; Erickson, B.R.; Nichol, S.T. Crimean-Congo hemorrhagic fever virus glycoprotein precursor is cleaved by furin-like and SKI-1 proteases to generate a novel 38-kilodalton glycoprotein. J Virol 2006, 80, 514–525. [Google Scholar] [CrossRef]
- Honig, J.E.; Osborne, J.C.; Nichol, S.T. Crimean–Congo hemorrhagic fever virus genome L RNA segment and encoded protein. Virology 2004, 321, 29–35. [Google Scholar] [CrossRef]
- Scholte, F.E.M.; Zivcec, M.; Dzimianski, J.V.; Deaton, M.K.; Spengler, J.R.; Welch, S.R.; Nichol, S.T.; Pegan, S.D.; Spiropoulou, C.F.; Bergeron, É. Crimean-Congo hemorrhagic fever virus suppresses innate immune responses via a ubiquitin and ISG15 specific protease. Cell reports 2017, 20, 2396–2407. [Google Scholar] [CrossRef]
- Perng, Y.C.; Lenschow, D.J. ISG15 in antiviral immunity and beyond. Nat Rev Microbiol 2018, 16, 423–439. [Google Scholar] [CrossRef]
- Chumakov, M. P. 30 Years of investigation of Crimean hemorrhagic fever (Russian). Труды Института пoлиoмиелита и вирусных энцефалитoв Академии медицинских наук CCCР 1974, 22, 5–18. [Google Scholar]
- Butenko, A.M.; Chumakov, M.P.; Rubin, V.N. Isolation and investigation of Astrakhan strain (‘‘Drozdov’’) of Crimean hemorrhagic fever virus and data on serodiagnosis of this infection. Mater. 15 Nauchn. Sess. Inst. Polio Virus Entsefalitov (Moscow) 3, 88–90. Whitehouse C.A. Crimean-Congo hemorrhagic fever. Antiviral Research 2004, 145–160. [Google Scholar]
- Chumakov, M.P.; Butenko, A.M.; Shalunova, N.V.; Mart'ianova, L.I.; Smirnova, S.E.; Bashkirtsev, I.N.; Zavodova, T. I.; Rubin, S.G.; Tkachenko, E.A.; Karmysheva, V.I.a. Novye dannye o viruse-vozbuditele krymskoĭ gemorragicheskoĭ likhoradki [New data on the viral agent of Crimean hemorrhagic fever]. Voprosy virusologii 1968, 13, 377. [Google Scholar] [PubMed]
- Simpson, D.I.; Knight, E.M.; Courtois, G.; Williams, M.C.; Weinbren, M.P.; Kibukamusoke, J.W. Congo virus: a hitherto undescribed occurring In Africa. I. Human isolations—clinical notes. East Afr Med J 1967, 44, 86–92. [Google Scholar]
- Whitehouse, C.A. Crimean-Congo hemorrhagic fever. Antiviral Res 2004, 64, 145–160. [Google Scholar] [CrossRef]
- Vorou, R.; Pierroutsakos, I.N.; Maltezou, H.C. Crimean-Congo hemorrhagic fever. Curr Opin Infect Dis 2007, 20, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Gargili, A.; Estrada-Peña, A.; Spengler, J.R.; Lukashev, A.; Nuttall, P.A.; Bente, D.A. The role of ticks in the maintenance and transmission of Crimean-Congo hemorrhagic fever virus: a review of published field and laboratory studies. Antiviral Res 2017, 144, 93–119. [Google Scholar] [CrossRef] [PubMed]
- Ergönül, Ö. Crimean-Congo haemorrhagic fever. Lancet Infect Dis 2006, 6, 203–214. [Google Scholar] [CrossRef]
- Mourya, D.T.; Yadav, P.D.; Shete, A.; Majumdar, T.D.; Kanani, A.; Kapadia, D.; Chandra, V.; Kachhiapatel, A.J.; Joshi, P.T.; Upadhyay, K.J.; et al. Serosurvey of Crimean-Congo hemorrhagic fever virus in domestic animals, Gujarat, India, 2013. Vector Borne Zoonotic Dis 2014, 14, 690–692. [Google Scholar] [CrossRef]
- Shepherd, A.J.; Swanepoel, R.; Cornel, A.J.; Mathee, O. Experimental studies on the replication and transmission of Crimean-Congo hemorrhagic fever virus in some African tick species. Am J Trop Med Hyg 1989, 40, 326–331. [Google Scholar] [CrossRef]
- Gordon, S.W.; Linthicum, K.J.; Moulton, J.R. Transmission of Crimean-Congo hemorrhagic fever virus in two species of Hyalomma ticks from infected adults to cofeeding immature forms. Am J Trop Med Hyg 1993, 48, 576–580. [Google Scholar] [CrossRef]
- Logan, T.M.; Linthicum, K.J.; Bailey, C.L.; Watts, D.M.; Moulton, J.R. Experimental transmission of Crimean- Congo haemorrhagic fever virus by Hyalomma truncatum Koch. Am J Trop Med Hyg 1989, 40, 207–212. [Google Scholar] [CrossRef]
- Okorie, T.G. Comparative studies on the vector capacity of the different stages of Amblyomma variegatum Fabricius and Hyalomma rufipes Koch for Congo virus, after intracoelomic inoculation. Veterinary Parasitology 1991, 38, 215–223. [Google Scholar] [CrossRef]
- El-Azazy, O.M.; Scrimgeour, E.M. Crimean-Congo haemorrhagic fever virus infection in the western province of Saudi Arabia. Trans R Soc Trop Med Hyg 1997, 91, 275–278. [Google Scholar] [CrossRef] [PubMed]
- Swanepoel, R.; Shepherd, A.J.; Leman, P.A.; Shepherd, S.P.; McGillivray, G.M.; Erasmus, M.J.; Searle, L.A.; Gill, D.E. Epidemiologic and clinical features of Crimean- Congo hemorrhagic fever in southern Africa. Am J Trop Med Hyg 1987, 36, 120–132. [Google Scholar] [CrossRef] [PubMed]
- Yen, Y.C.; Kong, L.X.; Lee, L.; Zhang, Y.Q.; Li, F.; Cai, B.J.; Gao, S.Y. Characteristics of Crimean-Congo hemorrhagic fever virus (Xinjiang strain) in China. Am J Trop Med Hyg 1985, 34, 1179–1182. [Google Scholar]
- Gear, J.H.; Thomson, P.D.; Hopp, M.; Andronikou, S.; Cohn, R.J.; Ledger, J.; Berkowitz, F.E. Congo-Crimean haemorrhagic fever in South Africa. Report of a fatal case in the Transvaal. S Afr Med J 1982, 62, 576–580. [Google Scholar]
- Saluzzo, J.F.; Aubry, P.; McCormick, J.; Digoutte, J.P. Haemorrhagic fever caused by Crimean Congo haemorrhagic fever virus in Mauritania. Trans R Soc Trop Med Hyg 1985, 79, 268. [Google Scholar] [CrossRef]
- Dunster, L.; Dunster, M.; Ofula, V.; Beti, D.; Kazooba-Voskamp, F.; Burt, F.; Swanepoel, R.; DeCock, K.M. First documentation of human Crimean-Congo hemorrhagic fever, Kenya. Emerg Infect Dis 2002, 8, 1005–1006. [Google Scholar] [CrossRef]
- Tantawi, H.H.; Al-Moslih, M.I.; Al-Janabi, N.Y.; Al-Bana, A.S.; Mahmud, M.I.; Jurji, F.; Yonan, M.S.; Al-Ani, F.; Al-Tikriti, S.K. Crimean-Congo haemorrhagic fever virus in Iraq: isolation, identification and electron microscopy. Acta Virol 1980, 24, 464–467. [Google Scholar] [PubMed]
- Schwarz, T.F.; Nitschko, H.; Jäger, G.; Nsanze, H.; Longson, M.; Pugh, R.N.; Abraham, A.K. Crimean-Congo haemorrhagic fever in Oman. Lancet 1995, 346, 1230. [Google Scholar] [CrossRef]
- El Azazy, O.M.; Scrimgeour, E.M. Crimean-Congo haemorrhagic fever virus infection in the western province of Saudi Arabia. Trans R Soc Trop Med Hyg 1997, 91, 275–278. [Google Scholar] [CrossRef] [PubMed]
- Chinikar, S.; Ghiasi, S.M.; Naddaf, S.; Piazak, N.; Moradi, M.; Razavi, M.R.; Afzali, N.; Haeri, A.; Mostafavizadeh, K.; Ataei, B.; et al. A serological survey in suspected human patients of Crimean-Congo hemorrhagic fever in Iran by determination of IgM-specific ELISA method during 2000–2004. Arch Iranian Med 2005, 8, 52–55. [Google Scholar]
- Mustafa, M.L.; Ayazi, E.; Mohareb, E.; Yingst, S.; Zayed, A.; Rossi, C.A.; Schoepp, R.J.; Mofleh, J.; Fiekert, K.; Akhbarian, Z.; et al. Crimean-Congo hemorrhagic fever, Afghanistan, 2009. Emerg Infect Dis 2011, 17, 1940–1941. [Google Scholar] [CrossRef]
- Burney, M.I.; Ghafoor, A.; Saleen, M.; Webb, P.A.; Casals, J. Nosocomial outbreak of viral hemorrhagic fever caused by Crimean hemorrhagic fever-Congo virus in Pakistan, January 1976. Am J Trop Med Hyg 1980, 29, 941–947. [Google Scholar] [CrossRef]
- Mishra, A.C.; Mehta, M.; Mourya, D.T.; Gandhi, S. Crimean-Congo haemorrhagic fever in India. Lancet 2011, 378, 372. [Google Scholar] [CrossRef]
- Yadav, P.D.; Patil, D.Y.; Shete, A.M.; Kokate, P.; Goyal, P.; Jadhav, S.; Sinha, S.; Zawar, D.; Sharma, S. K.; Kapil, A.; et al. Nosocomial infection of CCHF among healthcare workers in Rajasthan, India. BMC Infect Dis 2016, 16, 624. [Google Scholar] [CrossRef]
- Negredo, A.; de la Calle-Prieto, F.; Palencia-Herrejón, E.; Mora-Rillo, M.; Astray-Mochales, J.; Sánchez-Seco, M. P.; Bermejo Lopez, E.; Menárguez, J.; Fernández-Cruz, A.; Sánchez-Artola, B.; et al. Autochthonous Crimean-Congo hemorrhagic fever in Spain. N Engl J Med 2017, 377, 154–161. [Google Scholar] [CrossRef]
- Papa, A.; Bino, S.; Llagami, A.; Brahimaj, B.; Papadimitriou, E.; Pavlidou, V.; Velo, E.; Cahani, G.; Hajdini, M.; Pilaca, A.; et al. Crimean-Congo hemorrhagic fever in Albania, 2001. Eur J Clin Microbiol Infect Dis 2002, 21, 603–606. [Google Scholar] [CrossRef] [PubMed]
- Papa, A.; Christova, I.; Papadimitriou, E.; Antoniadis, A. Crimean-Congo hemorrhagic fever in Bulgaria. EmergInfect Dis 2004, 10, 1465–1467. [Google Scholar] [CrossRef] [PubMed]
- Drosten, C.; Minnak, D.; Emmerich, P.; Schmitz, H.; Reinicke, T. Crimean-Congo hemorrhagic fever in Kosovo. J Clin Microbiol 2002, 40, 1122–1123. [Google Scholar] [CrossRef]
- Ceianu, C.S.; Panculescu-Gatej, R.I.; Coudrier, D.; Bouloy, M. First serologic evidence for the circulation of Crimean-Congo hemorrhagic fever virus in Romania. Vector Borne Zoonotic Dis 2012, 12, 718–721. [Google Scholar] [CrossRef] [PubMed]
- Karti, S.S.; Odabasi, Z.; Korten, V.; Yilmaz, M.; Sonmez, M.; Caylan, R.; Akdogan, E.; Eren, N.; Koksal, I.; Ovali, E.; et al. Crimean-Congo hemorrhagic fever in Turkey. Emerg Infect Dis 2004, 10, 1379–1384. [Google Scholar] [CrossRef] [PubMed]
- Tonbak, S.; Aktas, M.; Altay, K.; Azkur, A.K.; Kalkan, A.; Bolat, Y.; Dumanli, N.; Ozdarendeli, A. Crimean-Congo hemorrhagic fever virus: genetic analysis and tick survey in Turkey. J Clin Microbiol 2006, 44, 4120–4124. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, G.R.; Buzgan, T.; Irmak, H.; Safran, A.; Uzun, R.; Cevik, M.A.; Torunoglu, M.A. The epidemiology of Crimean-Congo hemorrhagic fever in Turkey 2002-2007. Int J Infect Dis 2009, 13, 380–386. [Google Scholar] [CrossRef]
- Midilli, K.; Gargili, A.; Ergonul, O.; Elevli, M.; Ergin, S.; Turan, N.; Sengöz, G.; Ozturk, R.; Bakar, M. The first clinical case due to AP92 like strain of Crimean-Congo Hemorrhagic Fever virus and a field survey. BMC Infect Dis 2009, 9, 90. [Google Scholar] [CrossRef]
- Canakoglu, N.; Berber, E.; Ertek, M.; Yoruk, M.D.; Tonbak, S.; Bolat, Y.; Aktas, M.; Kalkan,A. ; Ozdarendeli, A. Pseudo-Plaque Reduction Neutralization Test (PPRNT) for the Measurement of Neutralizing Antibodies to Crimean-Congo Hemorrhagic Fever Virus. Virol J 2013, 10. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control: Crimean-Congo Haemorrhagic Fever. In: ECDC. Annual epidemiological report for 2019. Available online: https://www.ecdc.europa.eu/en/publications-data/crimean-congo-haemorrhagic-fever-annual-epidemiological-report-2019 (accessed on 3 February 2021).
- Estrada-Pena, A.; Palomar, A.M.; Santibanez, P. Crimean-Congo Hemorrhagic Fever Virus in Ticks, Southwestern Europe, 2010. Emerg Infect Dis 2012, 18, 179–180. [Google Scholar] [CrossRef]
- Spengler, J.R.; Bergeron, E.; Spiropoulou, C.F. Crimean-Congo hemorrhagic fever and expansion from endemic regions. Curr Opin Virol 2019, 34, 70–78. [Google Scholar] [CrossRef]
- World Health Organization. Geographic distribution of Crimean-Congo haemorrhagic fever. Available online: https://www.who.int/ emergencies/diseases/crimean-congo-haemorrhagic-fever/Global_ CCHFRisk_2017.jpg?ua=1 (accessed on 4 November 2019).
- Hoogstraal, H. The epidemiology of tick-borne Crimean-Congo hemorrhagic fever in Asia, Europe, and Africa. J Med Entomol 1979, 15, 307–417. [Google Scholar] [CrossRef]
- Jameson, L.J.; Morgan, P.J.; Medlock, J.M.; Watola, G.; Vaux, A.G.C. Importation of Hyalomma marginatum, vector of CrimeanCongo haemorrhagic fever virus, into the United Kingdom by migratory birds. Ticks Tick Borne Dis 2012, 3, 95–99. [Google Scholar] [CrossRef]
- Palomar, A.M.; Portillo, A.; Santiba ́ n ̃ ez, P. Crimean-Congo hemorrhagic fever virus in ticks from migratory birds, Morocco. Emerg Infect Dis 2013, 19, 260–263. [Google Scholar] [CrossRef] [PubMed]
- Capek, M.; Literak, I.; Kocianova, E.; Sychra, O.; Najer, T.; Trnka, A.; Kverek, P. Ticks of the Hyalomma marginatum complex transported by migratory birds into Central Europe. Ticks Tick Borne Dis 2014, 5, 489–493. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Peña, A.; Venzal, J.M. Climate niches of tick species in the Mediterranean region: modeling of occurrence data, distributional constraints, and impact of climate change. J Med Entomol 2007, 44, 1130–1138. [Google Scholar] [CrossRef]
- Vatansever, Z. ; Uzun, R.; Estrada-Peña, A.; Ergonul, O. etc. In Crimean-Congo Hemorrhagic Fever: A Global Perspective, 1st ed.; Ergonul, O., Whitehouse, C.A., Eds.; Dordrecht, Netherlands, 2007; p. 328. ISBN 978-1-4020-6105-9. [Google Scholar]
- Estrada-Peña, A.; Sánchez, N.; Estrada-Sánchez, A. An assessment of the distribution and spread of the tick Hyalomma marginatum in the western Palearctic under different climate scenarios. Vector Borne Zoonotic Dis 2012, 12, 758–768. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Peña, A.; Jameson, L.; Medlock, J.; Vatansever, Z.; Tishkova, F. Unraveling the ecological complexities of tickassociated Crimean-Congo hemorrhagic fever virus transmission: a gap analysis for the western Palearctic. Vector Borne Zoonotic Dis 2012, 12, 743–752. [Google Scholar] [CrossRef]
- Saleem, J.; Usman, M.; Nadeem, A.; Sethi, S.A.; Salman, M. Crimean-Congo hemorrhagic fever: a first case from Abbottabad, Pakistan. Int J Infect Dis 2009, 13, 121–123. [Google Scholar] [CrossRef]
- Jameson, L.J.; Medlock, J.M. Results of HPA tick surveillance in Great Britain. Vet Rec 2009, 165, 154. [Google Scholar] [CrossRef]
- Spengler, J.R.; Estrada-Peña, A.; Garrison, A.R.; Schmaljohn, C.; Spiropoulou, C.F.; Bergeron, E.; Bente, D. A chronological review of experimental infection studies on the role of wild animals and livestock in maintenance and transmission of Crimean-Congo hemorrhagic fever virus. Antiviral Res 2016, 135, 31–47. [Google Scholar] [CrossRef]
- Mallhi, T.H.; Khan, Y.H.; Sarriff. A.; Khan, A.H. Crimean-Congo haemorrhagic fever virus and Eid-Ul- Adha festival in Pakistan. Lancet Infect Dis 2016, 16, 1332–1333. [Google Scholar] [CrossRef]
- Chisholm, K.; Dueger, E.; Fahmy, N.T.; Samaha, H.A.T.; Zayed, A.; Abdel-Dayem, M.; Villinski, J.T. Crimean-congo hemorrhagic fever virus in ticks from imported livestock, Egypt. Emerg Infect Dis 2012, 18, 181–182. [Google Scholar] [CrossRef]
- Whitehouse, C.A. Crimean-Congo hemorrhagic fever. Antivir Res 2004, 64. [Google Scholar] [CrossRef] [PubMed]
- Ergonul, O. Crimean-Congo haemorrhagic fever. Lancet lnfect Dis 2006, 6. [Google Scholar] [CrossRef]
- Çevik, M.A.; Erbay, A.; Bodur, H.; Gülderen, E.; Baştuğ, A.; Kubar, A. Clinical and laboratory features of CrimeanCongo hemorrhagic fever: predictors of fatality. Int J Infect Dis 2008, 12, 374–382. [Google Scholar] [CrossRef]
- Gürbüz, Y.; Sencan, I.; Oztürk, B.; Tütüncü, E. A case of nosocomial transmission of Crimean-Congo hemorrhagic fever from patient to patient. Int J Infect Dis 2009, 13, 105–111. [Google Scholar] [CrossRef]
- Naderi, H.; Sheybani, F.; Bojdi, A.; Khosravi, N.; Mostafavi, I. Fatal nosocomial spread of Crimean-Congo hemorrhagic fever with very short incubation period. Am J Trop Med Hyg 2013, 88, 469–539. [Google Scholar] [CrossRef] [PubMed]
- Yildirmak, T.; Tulek, N.; Bulut, C. Crimean-Congo haemorrhagic fever: transmission to visitors and healthcare workers. Infection 2016, 44, 687–695. [Google Scholar] [CrossRef]
- Leblebicioglu, H.; Sunbul, M.; Guner, R.; Bodur, H.; Bulut, C.; Duygu, F. Healthcare-associated Crimean-Congo haemorrhagic fever in Turkey, 2002–2014: a multicentre retrospective cross-sectional study. Clin Microbiol Infect 2016, 22, 387–390. [Google Scholar] [CrossRef]
- Mehand, M.S.; Al-Shorbaji, F.; Millett, P.; Murgue, B. The WHO R&D Blueprint: 2018 review of emerging infectious diseases requiring urgent research and development efforts. Antivir Res 2018, 159, 63–67. [Google Scholar] [CrossRef]
- Bente, D. A. Crimean-Congo hemorrhagic fever: history, epidemiology, pathogenesis, clinical syndrome and genetic diversity. Antivir. Res 2013, 100, 159–189. [Google Scholar] [CrossRef]
- Rodriguez, S.E.; Hawman, D.W.; Sorvillo, T.E; O'Neal, T.J.; Bird, B.H.; Rodriguez, L.L; Bergeron, É.; Nichol, S.T.; Montgomery, J.M.; Spiropoulou, C.F.; et al. Immunobiology of Crimean-Congo hemorrhagic fever. Antiviral Res 2022, 199, 105244. [Google Scholar] [CrossRef]
- Hawman, D.W; Feldmann, H. Crimean-Congo haemorrhagic fever virus. Nat Rev Microbiol 2023, 14, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Andersson, I.; Lundkvist, A.; Haller, O.; Mirazimi, A. 2006. Type I interferon inhibits Crimean-Congo hemorrhagic fever virus in human target cells. J Med Virol 2006, 78, 216–222. [Google Scholar] [CrossRef]
- Hawman, D.W.; Meade-White, K.; Haddock, E.; Habib, R.; Scott, D.; Thomas, T.; Rosenke, R.; Feldmann, H. Crimean-Congo hemorrhagic fever mouse model recapitulating human convalescence. J Virol 2019, 93. [Google Scholar] [CrossRef] [PubMed]
- Hawman, David. W.; Meade-White, K.; Leventhal, S.; Carmody, A.; Haddock, E.; Hasenkrug, K.; Feldmann, H. T- cells and interferon gamma are necessary for survival following Crimean-Congo hemorrhagic fever virus infection in mice. Microorganisms 2021, 9, 279. [Google Scholar] [CrossRef] [PubMed]
- Bente, D.A.; Alimonti, J.B.; Shieh, W.-J.; Camus, G.; Stro ̈her, U.; Zaki, S.; Jones, S.M. Pathogenesis and immune response of Crimean-Congo hemorrhagic fever virus in a STAT-1 knockout mouse model. J Virol 2010, 84, 11089–11100. [Google Scholar] [CrossRef]
- Bereczky, S.; Lindegren, G.; Karlberg, H. Crimean-Congo hemorrhagic fever virus infection is lethal for adult type I interferon receptor- knockout mice. J Gen Virol 2010, 91, 1473–1477. [Google Scholar] [CrossRef]
- Lindquist, M.E.; Zeng, X.; Altamura, L.A.; Daye, S.P.; Delp, K.L.; Blancett, C.; Coffin, K.M.; Koehler, J.W.; Coyne, S.; Shoemaker, C.J.; et al. Exploring Crimean-Congo hemorrhagic fever virus-induced hepatic injury using antibody-mediated type I interferon blockade in mice. J Virol 2018, 92. [Google Scholar] [CrossRef]
- Arslan, S.; Engin, A.; Ozbilüm, N.; Bakır, M. Toll-like receptor 7 Gln11Leu, c.4- 151A/G, and +1817G/Tpolymorphisms in Crimean-Congo hemorrhagic fever. J Med Virol 2015, 87, 1090–1095. [Google Scholar] [CrossRef]
- Engin, A.; Arslan, S.; O ̈zbilüm, N.; Bakir, M. Is there any relationship between Toll-like receptor 3 c.1377C/Tand -7C/A polymorphisms and susceptibility to Crimean Congo hemorrhagic fever? J Med. Virol 2016, 88, 1690–1696. [Google Scholar] [CrossRef]
- Weber, F.; Mirazimi, A. Interferon and cytokine responses to Crimean Congo hemorrhagic fever virus; an emerging and neglected viral zonoosis. Cytokine Growth Factor Rev 2008, 19, 395–404. [Google Scholar] [CrossRef]
- Habjan, M.; Andersson, I.; Klingström, J.; Schümann, M.; Martin, A.; Zimmermann, P.; Wagner, V.; Pichlmair, A.; Schneider, U.; Mühlberger, E.; et al. Processing of genome 51 termini as a strategy of negative-strand RNA viruses to avoid RIG-I-dependent interferon induction. PLoS One 2008, 3, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Spengler, J.R.; Patel, J.R.; Chakrabarti, A.K.; Zivcec, M.; García-Sastre, A.; Spiropoulou, C.F.; Bergeron, E. RIG-I mediates an antiviral response to Crimean-Congo hemorrhagic fever virus. J Virol 2015, 89, 10119–10229. [Google Scholar] [CrossRef] [PubMed]
- Zivcec, M.; Metcalfe, M.G.; Albariño, C.G.; Guerrero, L.W.; Pegan, S.D.; Spiropoulou, C.F.; Bergeron, É. Assessment of inhibitors of pathogenic Crimean-Congo hemorrhagic fever virus strains using virus-like particles. PLoS Negl Trop Dis 2015, 9. [Google Scholar] [CrossRef]
- Everett, H.; McFadden, G. 1999. Apoptosis: an innate immune response to virus infection. Trends Microbiol 1999, 7, 160–165. [Google Scholar] [CrossRef]
- Barber, G. Host defense, viruses and apoptosis. Cell Death Differ 2001, 8, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Karlberg, H.; Tan, Y.; J. & Mirazimi, A. Crimean–Congo haemorrhagic fever replication interplays with regulation mechanisms of apoptosis. J Gen Virol 2015, 96, 538–546. [CrossRef]
- Rodrigues, R; Paranhos-Baccalà, G; Vernet, G.; Peyrefitte, C. N. Crimean-Congo hemorrhagic fever virus-infected hepatocytes induce ER-stress and apoptosis crosstalk. PLoS ONE 2012, 7, 29712. [CrossRef]
- Barnwal, B.; Karlberg, H.; Mirazimi, A.; Tan, Y.J. The Non-structural Protein of Crimean-Congo Hemorrhagic Fever Virus Disrupts the Mitochondrial Membrane Potential and Induces Apoptosis. J Biol Chem 2016, 291, 582–592. [Google Scholar] [CrossRef]
- Li, D.; Wu, M. Pattern recognition receptors in health and diseases. Signal Transduct Target Ther 2021, 6, 291. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nat Immunol 2010, 11, 373–384. [Google Scholar] [CrossRef]
- Mogensen, T.H. Pathogen recognition and inflammatory signaling in innate immune defenses. Clin Microbiol Rev 2009, 22, 240–273. [Google Scholar] [CrossRef]
- Newton, K.; Dixit, V.M. Signaling in innate immunity and inflammation. Cold Spring Harb Perspect Biol 2012, 4, a006049. [Google Scholar] [CrossRef] [PubMed]
- Connolly-Andersen, A.M.; Douagi, I.; Kraus, A.A.; Mirazimi, A. Crimean Congo hemorrhagic fever virus infects human monocyte-derived dendritic cells. Virology 2009, 390, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Ergönül, Ö.; Şeref, C.; Eren, Ş.; Çelikbaş, A.; Baykam, N.; Dokuzoğuz, B.; Gönen, M.; Can, F. Cytokine response in crimean-congo hemorrhagic fever virus infection. J Med Virol 2017, 89, 1707–1713. [Google Scholar] [CrossRef]
- Saksida, A.; Duh, D.; Wraber, B.; Dedushaj, I.; Ahmeti, S.; Avsic-Zupanc, T. Interacting roles of immune mechanisms and viral load in the pathogenesis of crimean-congo hemorrhagic fever. Clin Vaccine Immunol 2010, 17, 1086–1093. [Google Scholar] [CrossRef]
- Papa, A.; Bino, S.; Velo, E.; Harxhi, A.; Kota, M.; Antoniadis, A. Cytokine levels in Crimean-Congo hemorrhagic fever. J Clin Virol 2006, 36, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Papa, A.; Tsergouli, K.; Çağlayık, D.Y.; Bino, S.; Como, N.; Uyar, Y.; Korukluoglu, G. Cytokines as biomarkers of Crimean-Congo hemorrhagic fever. J Med Virol 2016, 88, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Ergönül, O. Crimean-Congo haemorrhagic fever. Lancet Infect Dis 2006, 6, 203–214. [Google Scholar] [CrossRef]
- Altay, F.A.; Elaldi, N.; Şentürk, G.Ç.; Altin, N.; Gözel, M.G.; Albayrak, Y.; Şencan, İ. Serum sTREM-1 level is quite higher in Crimean Congo Hemorrhagic Fever, a viral infection. J Med Virol 2016, 88, 1473–1478. [Google Scholar] [CrossRef]
- Hamidinejad, M.A.; Esmaeili Gouvarchin Ghaleh, H.; Farzanehpour, M.; Bolandian, M.; Dorostkar, R. Crimean-Congo hemorrhagic fever from the immunopathogenesis, clinical, diagnostic, and therapeutic perspective: A scoping review. Asian Pac J Trop Med 2021, 14, 254–265. [Google Scholar] [CrossRef]
- Shepherd, A.J.; Swanepoel, R.; Leman, P.A. Antibody response in Crimean-Congo hemorrhagic fever. Rev Infect Dis 1989, 11, S801–S806. [Google Scholar] [CrossRef] [PubMed]
- Burt, F.J.; Leman, P.A.; Abbott, J.C.; Swanepoel, R. Serodiagnosis of Crimean-Congo haemorrhagic fever. Epidemiol Infect 1994, 113, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Ergunay, K.; Kocak Tufan, Z.; Bulut, C.; Kinikli, S.; Demiroz, A.P.; Ozkul, A. Antibody responses and viral load in patients with Crimean-Congo hemorrhagic fever: a comprehensive analysis during the early stages of the infection. Diagn Microbiol Infect Dis 2014, 79, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Kaya, S.; Elaldi, N.; Kubar, A.; Gursoy, N.; Yilmaz, M.; Karakus, G.; Gunes, T.; Polat, Z.; Gozel, M.G.; Engin, A.; Dokmetas, I.; Bakir, M.; Yilmaz, N.; Sencan, M. Sequential determination of serum viral titers, virus-specific IgG antibodies, and TNF-α, IL-6, IL-10, and IFN-γ levels in patients with Crimean-Congo hemorrhagic fever. BMC Infect Dis 2014, 14, 416. [Google Scholar] [CrossRef]
- Duh, D.; Saksida, A.; Petrovec, M.; Ahmeti, S.; Dedushaj, I.; Panning, M.; Drosten, C.; Avsic-Zupanc, T. Viral load as predictor of Crimean-Congo hemorrhagic fever outcome. Emerg Infect Dis 2007, 13, 1769–1772. [Google Scholar] [CrossRef]
- Bertolotti-Ciarlet, A.; Smith, J.; Strecker, K.; Paragas, J.; Altamura, L.A.; McFalls, J.M.; Frias-Stäheli, N.; García-Sastre, A.; Schmaljohn, C.S.; Doms, R.W. Cellular localization and antigenic characterization of crimean-congo hemorrhagic fever virus glycoproteins. J Virol 2005, 79, 6152–6161. [Google Scholar] [CrossRef]
- Zivcec, M.; Guerrero, L.I.W.; Albariño, C.G.; Bergeron, É.; Nichol, S.T.; Spiropoulou, C.F. Identification of broadly neutralizing monoclonal antibodies against Crimean-Congo hemorrhagic fever virus. Antiviral Res 2017, 146, 112–120. [Google Scholar] [CrossRef]
- Golden, J.W.; Shoemaker, C.J.; Lindquist, M.E.; Zeng, X.; Daye, S.P.; Williams, J.A.; Liu, J.; Coffin, K.M.; Olschner, S.; Flusin, O.; Altamura, L.A.; Kuehl, K.A.; Fitzpatrick, C.J.; Schmaljohn, C.S.; Garrison, A.R. GP38-targeting monoclonal antibodies protect adult mice against lethal Crimean-Congo hemorrhagic fever virus infection. Sci Adv 2019, 5, eaaw9535. [Google Scholar] [CrossRef]
- Mishra, A.K.; Moyer, C.L.; Abelson, D.M.; Deer, D.J.; ElOmari, K.; Duman, R.; Lobel, L.; Lutwama, J.J.; Dye, J.M.; Wagner, A.; Chandran, K.; Cross, R.W.; Geisbert, T.W.; Zeitlin, L.; Bornholdt, Z.A.; McLellan, J.S. Structure and Characterization of Crimean-Congo Hemorrhagic Fever Virus GP38. J Virol 2020, 94, e02005–19. [Google Scholar] [CrossRef]
- Fels, J.M.; Maurer, D.P.; Herbert, A.S.; Wirchnianski, A.S.; Vergnolle, O.; Cross, R.W.; Abelson, D.M.; Moyer, C.L.; Mishra, A.K.; Aguilan, J.T.; et al. Protective neutralizing antibodies from human survivors of Crimean-Congo hemorrhagic fever. Cell 2021, 184, 3486–3501.e21. [Google Scholar] [CrossRef]
- Hasanoglu, I.; Guner, R.; Carhan, A.; Kocak Tufan, Z.; Yagci-Caglayik, D.; Guven, T.; Yilmaz, G.R.; Tasyaran, M.A. Crucial parameter of the outcome in Crimean Congo hemorrhagic fever: Viral load. J Clin Virol 2016, 75, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Hawman, D.W.; Meade-White, K.; Leventhal, S.; Feldmann, F.; Okumura, A.; Smith, B.; Scott, D.; Feldmann, H. Immunocompetent mouse model for Crimean-Congo hemorrhagic fever virus. eLife 2021, 10, e63906. [Google Scholar] [CrossRef] [PubMed]
- Hinkula, J.; Devignot, S.; Åkerström, S.; Karlberg, H.; Wattrang, E.; Bereczky, S.; MousaviJazi, M.; Risinger, C.; Lindegren, G.; Vernersson, C.; et al. Immunization with DNA Plasmids Coding for Crimean-Congo Hemorrhagic Fever Virus Capsid and Envelope Proteins and/or Virus-Like Particles Induces Protection and Survival in Challenged Mice. J Virol 2017, 91, e02076–16. [Google Scholar] [CrossRef] [PubMed]
- Goedhals, D.; Paweska, J.T.; Burt, F.J. Long-lived CD8+ T cell responses following Crimean-Congo haemorrhagic fever virus infection. PLoS Negl Trop Dis 2017, 11, e0006149. [Google Scholar] [CrossRef]
- Smirnova, S.E. A comparative study of the Crimean hemorrhagic fever-Congo group of viruses. Arch Virol 1979, 62, 137–143. [Google Scholar] [CrossRef]
- Teijaro, J.R.; Ng, C.; Lee, A.M.; Sullivan, B.M.; Sheehan, K.C.; Welch, M.; Schreiber, R.D.; de la Torre, J.C.; Oldstone, M.B. Persistent LCMV infection is controlled by blockade of type I interferon signaling. Science 2013, 340, 207–211. [Google Scholar] [CrossRef]
- Smith, D.R.; Hollidge, B.; Daye, S.; Zeng, X.; Blancett, C.; Kuszpit, K.; Bocan, T.; Koehler, J.W.; Coyne, S.; Minogue, T.; et al. Neuropathogenesis of Zika Virus in a Highly Susceptible Immunocompetent Mouse Model after Antibody Blockade of Type I Interferon. PLoS Negl Trop Dis 2017, 11, e0005296. [Google Scholar] [CrossRef]
- Pavel, S.T.I.; Yetiskin, H.; Kalkan, A.; Ozdarendeli, A. Evaluation of the cell culture based and the mouse brain derived inactivated vaccines against Crimean-Congo hemorrhagic fever virus in transiently immune-suppressed (IS) mouse model. PLoS Negl Trop Dis 2020, 14, e0008834. [Google Scholar] [CrossRef]
- Golden, J.W.; Zeng, X.; Cline, C.R.; Smith, J.M.; Daye, S.P.; Carey, B.D.; Blancett, C.D.; Shoemaker, C.J.; Liu, J.; Fitzpatrick, C.J.; et al. The host inflammatory response contributes to disease severity in Crimean-Congo hemorrhagic fever virus infected mice. PLoS Pathog 2022, 18, e1010485. [Google Scholar] [CrossRef]
- Spengler, J.R.; KellyKeating, M.; McElroy, A.K.; Zivcec, M.; ColemanMcCray, J.D.; Harmon, J.R.; Bollweg, B.C.; Goldsmith, C.S.; Bergeron, É.; Keck, J.G.; et al. Crimean-Congo Hemorrhagic Fever in Humanized Mice Reveals Glial Cells as Primary Targets of Neurological Infection. J Infect Dis 2017, 216, 1386–1397. [Google Scholar] [CrossRef]
- Bility, M.T.; Cheng, L.; Zhang, Z.; Luan, Y.; Li, F.; Chi, L.; Zhang, L.; Tu, Z.; Gao, Y.; Fu, Y.; et al. Hepatitis B virus infection and immunopathogenesis in a humanized mouse model: induction of human-specific liver fibrosis and M2-like macrophages. PLoS Pathog 2014, 10, e1004032. [Google Scholar] [CrossRef] [PubMed]
- Lüdtke, A.; Oestereich, L.; Ruibal, P.; Wurr, S.; Pallasch, E.; Bockholt, S.; Ip, W.H.; Rieger, T.; Gómez-Medina, S.; Stocking, C.; et al. Ebola virus disease in mice with transplanted human hematopoietic stem cells. J Virol 2015, 89, 4700–4704. [Google Scholar] [CrossRef] [PubMed]
- Chinikar, S.; Ghiasi, S.M.; Moradi, M.; Goya, M.M.; Shirzadi, M.R.; Zeinali, M.; Meshkat, M.; Bouloy, M. Geographical distribution and surveillance of Crimean-Congo hemorrhagic fever in Iran. Vector Borne Zoonotic Dis 2010, 10, 705–708. [Google Scholar] [CrossRef] [PubMed]
- Yagci-Caglayik, D.; Korukluoglu, G.; Uyar, Y. Seroprevalence and risk factors of Crimean-Congo hemorrhagic fever in selected seven provinces in Turkey. J Med Virol 2014, 86, 306–314. [Google Scholar] [CrossRef]
- Monsalve-Arteaga, L.; Alonso-Sardón, M.; Muñoz Bellido, J.L.; Vicente Santiago, M.B.; Vieira Lista, M.C.; López Abán, J.; Muro, A.; Belhassen-García, M. Seroprevalence of Crimean-Congo hemorrhagic fever in humans in the World Health Organization European region: A systematic review. PLoS Negl Trop Dis 2020, 14, e0008094. [Google Scholar] [CrossRef] [PubMed]
- Butenko, A.M.; Chumakov, M.P.; Smirnova, S.E.; Vasilenko, S.M.; Zavodova, T.I.; Tkachenko, E.A.; et al. Isolation of Crimean hemorrhagic fever virus from blood of patients and corpse material (from 1968–1969 investigation data) in Orstov, Astrakhan Oblast, and Bulgaria. Mater 3 Oblast Nauchn Prakt Konf (Rostovno-Donu, May 1970); 1970. p. 6–25. [Russian, translated NAMRU-3 T522]. 19 May.
- Fagbami, A.H.; Tomori, O.; Fabiyi, A.; Isoun, T.T. Experimantal Congo virus (Ib -AN 7620) infection in primates. Virologie. 1975, 26, 33–7. [Google Scholar]
- Garcia-Sastre, A. Diversity, replication, pathogenicity and cell biology of Crimean Congo hemorrhagic fever virus. Mount Sinai School of Medicine New York, 2010.
- Haddock, E.; Feldmann, F.; Hawman, D.W.; Zivcec, M.; Hanley, P.W.; Saturday, G.; Scott, D.P.; Thomas, T.; Korva, M.; Avšič-Županc, T.; Safronetz, D.; Feldmann, H. A cynomolgus macaque model for Crimean-Congo haemorrhagic fever. Nat Microbiol 2018, 3, 556–562. [Google Scholar] [CrossRef]
- Cross, R.W.; Prasad, A.N.; Borisevich, V.; Geisbert, J.B.; Agans, K.N.; Deer, D.J.; Fenton, K.A.; Geisbert, T.W. Crimean-Congo hemorrhagic fever virus strains Hoti and Afghanistan cause viremia and mild clinical disease in cynomolgus monkeys. PLoS Negl Trop Dis 2020, 14, e0008637. [Google Scholar] [CrossRef]
- Smith, D.R.; Shoemaker, C.J.; Zeng, X.; Garrison, A.R.; Golden, J.W.; Schellhase, C.W.; Pratt, W.; Rossi, F.; Fitzpatrick, C.J.; Shamblin, J.; et al. Persistent Crimean-Congo hemorrhagic fever virus infection in the testes and within granulomas of non-human primates with latent tuberculosis. PLoS Pathog 2019, 15, e1008050. [Google Scholar] [CrossRef]
- Schmaljohn, C.S.; Nichol, S.T. Bunyaviridae. In Fields virology, 5th ed.; Knipe, D.M., Howley, P.M., Griffin, D.E., Lamb, R.A., Martin, M.A., Roizman, B., Straus, S.E., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, 2007; Volume 1, pp. 1741–1789. ISBN 0781760607/9780781760607. [Google Scholar]
- Karaaslan, E.; Çetin, N.S.; Kalkan-Yazıcı, M.; Hasanoğlu, S.; Karakeçili, F.; Özdarendeli, A.; Kalkan, A.; Kılıç, A.O.; Doymaz, M.Z. Immune responses in multiple hosts to Nucleocapsid Protein (NP) of Crimean-Congo Hemorrhagic Fever Virus (CCHFV). PLoS Negl Trop Dis 2021, 15, e0009973. [Google Scholar] [CrossRef]
- Deyde, V.M.; Khristova, M.L.; Rollin, P.E.; Ksiazek, T.G.; Nichol, S.T. Crimean-Congo Hemorrhagic Fever Virus Genomics and Global Diversity. J Virol 2006, 80, 8834–8842. [Google Scholar] [CrossRef] [PubMed]
- Kalkan-Yazıcı, M.; Karaaslan, E.; Çetin, N.S.; Hasanoğlu, S.; Güney, F.; Zeybek, U.; Doymaz, M.Z. Cross-Reactive anti-Nucleocapsid Protein Immunity against Crimean-Congo Hemorrhagic Fever Virus and Hazara Virus in Multiple Species. J Virol 2021, 95, e02156–20. [Google Scholar] [CrossRef] [PubMed]
- Altamura, L.A.; Bertolotti-Ciarlet, A.; Teigler, J.; Paragas, J.; Schmaljohn, C.S.; Doms, R.W. Identification of a novel C-terminal cleavage of Crimean-Congo hemorrhagic fever virus PreGN that leads to generation of an NSM protein. J Virol 2007, 81, 6632–6642. [Google Scholar] [CrossRef] [PubMed]
- Garrison, A.R.; Radoshitzky, S.R.; Kota, K.P.; Pegoraro, G.; Ruthel, G.; Kuhn, J.H.; Altamura, L.A.; Kwilas, S.A.; Bavari, S.; Haucke, V.; Schmaljohn, C.S. Crimean-Congo hemorrhagic fever virus utilizes a clathrin and early endosome-dependent entry pathway. Virology 2013, 444, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Freitas, N.; Enguehard, M.; Denolly, S.; Levy, C.; Neveu, G.; Lerolle, S.; Devignot, S.; Weber, F.; Bergeron, E.; Legros, V.; Cosset, F.L. The interplays between Crimean-Congo hemorrhagic fever virus (CCHFV) M segment-encoded accessory proteins and structural proteins promote virus assembly and infectivity. PLoS Pathog 2020, 16, e1008850. [Google Scholar] [CrossRef]
- Tkachenko, Y.A.; Butenko, A.M.; Butenko, S.A.; Zavodova, T.I.; Chumakov, M.P. Prophylactic characteristics of a protective Crimean hemorrhagic fever vaccine. In: Chumakov, M.P.(Ed.), Crimean Hemorrhagic Fever-Materials of the 3rd Scientific-practical Conference in Rostov-na-Donu Region, Rostav-na-Donu, USSR [Russian], 1970; pp. 136–138.
- Christova, I.; Kovacheva, T.; Georgieva, D. , Ivanova, S.; Argirov, D. Vaccine against Congo- Crimean haemorhagic fever virus—Bulgarian input in fighting the disease. Probl Infect Parasit Dis 2010, 37, 7–8. [Google Scholar]
- Mousavi-Jazi, M.; Karlberg, H.; Papa, A.; Christova, I.; Mirazimi, A. Healthy individuals’ immune response to the Bulgarian Crimean-Congo hemorrhagic fever virus vaccine. Vaccine 2012, 30, 6225–6229. [Google Scholar] [CrossRef]
- Canakoglu, N.; Berber, E.; Tonbak, S.; Ertek, M.; Sozdutmaz, I.; Aktas, M.; Kalkan, A.; Ozdarendeli, A. Immunization of knock-out a/b interferon receptor mice against high lethal dose of Crimean-Congo hemorrhagic fever virus with a cell culture based vaccine. PLoS Negl Trop Dis 2015, 9, e0003579. [Google Scholar] [CrossRef]
- Berber, E.; Canakoglu, N.; Tonbak, S.; Ozdarendeli, A. Development of a protective inactivated vaccine against Crimean-Congo hemorrhagic fever infection. Heliyon 2021, 7, e08161. [Google Scholar] [CrossRef]
- Kortekaas, J.; Vloet, R.P.; McAuley, A.J.; Shen, X.; Bosch, B.J.; de Vries, L.; Moormann, R.J. , Bente, D.A. Crimean-Congo hemorrhagic fever virus subunit vaccines induce high levels of neutralizing antibodies but no protection in STAT1 knockout mice. Vector Borne Zoonotic Dis 2015, 15, 759–764. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, S.; Shi, Z.; Li, Z.; Zhao, Y.; Feng, N.; Bi, J.; Jiao, C.; Li, E.; Wang, T.; Wang, J.; Jin, H.; Huang, P.; Yan, F.; Yang, S.; Xia, X. GEM-PA-Based Subunit Vaccines of Crimean Congo Hemorrhagic Fever Induces Systemic Immune Responses in Mice. Viruses 2022, 14, 1664. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Wang, P.; Jiang, L.; Wang, S.; Zhang, S.; Li, Y. Evaluation of the immunogenicity of vaccine candidates developed using a baculovirus surface display system for Crimean-Congo hemorrhagic fever virus in mice. Front Microbiol. 2023, 14, 1107874. [Google Scholar] [CrossRef] [PubMed]
- Stander, J.; Mbewana, S.; Meyers, A.E. Plant-Derived Human Vaccines: Recent Developments. BioDrugs 2022, 36, 573–589. [Google Scholar] [CrossRef] [PubMed]
- Ghiasi, S.M.; Salmanian, A.H.; Chinikar, S.; Zakeri, S. Mice orally immunized with a transgenic plant expressing the glycoprotein of Crimean- Congo hemorrhagic fever virus. Clin Vaccine Immunol 2011, 18, 2031–2037. [Google Scholar] [CrossRef]
- Robinson, H.L.; Pertmer, T.M. DNA vaccines for viral infections: basic studies and applications. Adv Virus Res 2000, 55, 1–74. [Google Scholar] [CrossRef]
- Qin, F.; Xia, F.; Chen, H.; Cui, B.; Feng, Y.; Zhang, P.; Chen, J.; Luo, M. A Guide to Nucleic Acid Vaccines in the Prevention and Treatment of Infectious Diseases and Cancers: From Basic Principles to Current Applications. Front Cell Dev Biol 2021, 9, 633776. [Google Scholar] [CrossRef]
- Spik, K.; Shurtleff, A.; McElroy, A.K.; Guttieri, M.C.; Hooper, J.W.; SchmalJohn, C. Immunogenicity of combination DNA vaccines for Rift Valley fever virus, tick-borne encephalitis virus, Hantaan virus, and Crimean Congo hemorrhagic fever virus. Vaccine 2006, 24, 4657–4666. [Google Scholar] [CrossRef]
- Garrison, A.R.; Shoemaker, C.J.; Golden, J.W.; Fitzpatrick, C.J.; Suschak, J.J.; Richards, M.J.; Badger, C.V.; Six, C.M.; Martin, J.D.; Hannaman, D.; et al. A DNA vaccine for Crimean-Congo hemorrhagic fever protects against disease and death in two lethal mouse models. PLoS Negl Trop Dis 2017, 11, e0005908. [Google Scholar] [CrossRef]
- Suschak, J.J.; Golden, J.W.; Fitzpatrick, C.J.; Shoemaker, C.J.; Badger, C.V.; Schmaljohn, C.S.; Garrison, A.R. A CCHFV DNA vaccine protects against heterologous challenge and establishes GP38 as immunorelevant in mice. NPJ vaccines 2021, 6, 31. [Google Scholar] [CrossRef]
- Aligholipour Farzani, T.; Hanifehnezhad, A.; Földes, K.; Ergünay, K.; Yilmaz, E.; Hashim Mohamed Ali, H.; Ozkul, A. Co-delivery effect of CD24 on the Immunogenicity and Lethal Challenge Protection of a DNA Vector Expressing Nucleocapsid Protein of Crimean Congo Hemorrhagic Fever Virus. Viruses 2019, 11, 75. [Google Scholar] [CrossRef]
- Hu, Y.L.; Zhang, L.Q.; Liu, X.Q.; Ye, W.; Zhao, Y.X.; Zhang, L.; Qiang, Z.X.; Zhang, L.X.; Lei, Y.F.; Jiang, D.B.; Cheng, L.F.; Zhang, F.L. Construction and evaluation of DNA vaccine encoding Crimean Congo hemorrhagic fever virus nucleocapsid protein, glycoprotein N- terminal and C-terminal fused with LAMP1. Front Cell Infect Microbiol 2023, 13, 1121163. [Google Scholar] [CrossRef] [PubMed]
- Hawman, D.W.; Ahlén, G.; Appelberg, K.S.; Meade-White, K.; Hanley, P.W.; Scott, D.; Monteil, V.; Devignot, S.; Okumura, A.; Weber, F.; Feldmann, H.; Sällberg, M.; Mirazimi, A. A DNA-based vaccine protects against Crimean-Congo haemorrhagic fever virus disease in a cynomolgus macaque model. Nat Microbiol 2021, 6, 187–195. [Google Scholar] [CrossRef]
- Scholte, F.E.M.; Spengler, J.R.; Welch, S.R.; Harmon, J.R.; Coleman-McCray, J.D.; Freitas, B.T.; Kainulainen, M.H.; Pegan, S.D.; Nichol, S.T.; Bergeron, É.; Spiropoulou, C.F. Single-dose replicon particle vaccine provides complete protection against Crimean-Congo hemorrhagic fever virus in mice. Emerg Microbes lnfect 2019, 8, 575–578. [Google Scholar] [CrossRef] [PubMed]
- Spengler, J.R.; Welch, S.R.; Scholte, F.E.M.; Coleman-McCray, J.D.; Harmon, J.R.; Nichol, S.T.; Bergeron, É.; Spiropoulou, C.F. Heterologous protection against Crimean-Congo hemorrhagic fever in mice after a single dose of replicon particle vaccine. Antiviral Res 2019, 170, 104573. [Google Scholar] [CrossRef] [PubMed]
- Spengler, J.R.; Welch, S.R.; Scholte, F.E.M.; Rodriguez, S.E.; Harmon, J.R.; Coleman-McCray, J.D.; Nichol, S.T.; Montgomery, J.M.; Bergeron, É.; Spiropoulou, C.F. Viral replicon particles protect IFNAR-/- mice against lethal Crimean- Congo hemorrhagic fever virus challenge three days after vaccination. Antiviral Res 2021, 191, 105090. [Google Scholar] [CrossRef] [PubMed]
- Kümmerer, B.M. Establishment and Application of Flavivirus Replicons. Adv Exp Med Biol 2018, 1062, 165–173. [Google Scholar] [CrossRef]
- Tran, P.T.; Höglund, U.; Larsson, O.; Appelberg, S.; Mirazimi, A.; Johansson, M.; Melik, W. Enhanced Seroconversion to West Nile Virus Proteins in Mice by West Nile Kunjin Replicon Virus-like Particles Expressing Glycoproteins from Crimean-Congo Hemorrhagic Fever Virus. Pathogens 2022, 11, 233. [Google Scholar] [CrossRef]
- Gorchakov, R.; Volkova, E.; Yun, N.; Petrakova, O.; Linde, N.S.; Paessler, S.; Frolova, E.; Frolov, I. Comparative analysis of the alphavirus-based vectors expressing Rift Valley fever virus glycoproteins. Virology 2007, 366, 212–225. [Google Scholar] [CrossRef]
- Tipih, T.; Heise, M.; Burt, F.J. Immunogenicity of a DNA-Based Sindbis Replicon Expressing Crimean-Congo Hemorrhagic Fever Virus Nucleoprotein. Vaccines 2021, 9, 1491. [Google Scholar] [CrossRef]
- Leventhal, S.S.; Meade-White, K.; Rao, D.; Haddock, E.; Leung, J.; Scott, D.; Archer, J.; Randall, S.; Erasmus, J.H.; Feldmann, H.; Hawman, D.W. Replicating RNA vaccination elicits an unexpected immune response that efficiently protects mice against lethal Crimean-Congo hemorrhagic fever virus challenge. eBioMedicine 2022, 82, 104188. [Google Scholar] [CrossRef]
- Chaudhary, N.; Weissman, D.; Whitehead, K.A. mRNA vaccines for infectious diseases: principles, delivery and clinical translation. Nat Rev Drug Discov 2021, 20, 817–838. [Google Scholar] [CrossRef] [PubMed]
- Gote, V.; Bolla, P.K.; Kommineni, N.; Butreddy, A.; Nukala, P.K.; Palakurthi, S.S.; Khan, W.A. A Comprehensive Review of mRNA Vaccines. Int J Mol Sci 2023, 24, 2700. [Google Scholar] [CrossRef] [PubMed]
- Aligholipour Farzani, T.; Földes, K.; Ergunay, K.; Gurdal, H.; Bastug, A.; Ozkul, A. Immunological Analysis of a CCHFV mRNA Vaccine Candidate in Mouse Models. Vaccines 2019, 7, 115. [Google Scholar] [CrossRef] [PubMed]
- Appelberg, S.; John, L.; Pardi, N.; Végvári, Á.; Bereczky, S.; Ahlén, G.; Monteil, V.; Abdurahman, S. ; Mikaeloff, F.; Beattie, M.; Tam, Y.; Sällberg, M.; Neogi, U.; Weissman, D.; Mirazimi, A. Nucleoside-modified mRNA vaccines protect IFNAR−/− mice against Crimean Congo hemorrhagic fever virus infection. J Virol 2021, 96, e01568–21. [Google Scholar] [CrossRef]
- McCann, N.; O'Connor, D.; Lambe, T.; Pollard, A.J. Viral vector vaccines. Curr Opin Immunol 2022, 77, 102210. [Google Scholar] [CrossRef]
- Wang, S.; Liang, B.; Wang, W.; Li, L.; Feng, N.; Zhao, Y.; Wang, T.; Yan, F.; Yang, S.; Xia, X. Viral vectored vaccines: design, development, preventive and therapeutic applications in human diseases. Sig Transduct Target Ther 2023, 8, 149. [Google Scholar] [CrossRef]
- Buttigieg, K.R.; Dowall, S.D.; Findlay-Wilson, S.; Miloszewska, A.; Rayner, E.; Hewson, R.; Carroll, M.W. A novel vaccine against Crimean-Congo haemorrhagic fever protects 100% of animals against lethal challenge in a mouse model. PLoS One 2014, 9, e91516. [Google Scholar] [CrossRef] [PubMed]
- Dowall, S.D.; Buttigieg, K.R.; Findlay-Wilson, S.J.; Rayner, E.; Pearson, G.; Miloszewska, A.; Graham, V.A.; Carroll, M.W.; Hewson, R.A. Crimean-Congo hemorrhagic fever (CCHF) viral vaccine expressing nucleoprotein is immunogenic but fails to confer protection against lethal disease. Hum Vaccines Immunother 2016, 12, 519–527. [Google Scholar] [CrossRef]
- Zivcec, M.; Safronetz, D.; Scott, D.P.; Robertson, S.; Feldmann, H. Nucleocapsid protein-based vaccine provides protection in mice against lethal Crimean-Congo hemorrhagic fever virus challenge. PLoS Negl Trop Dis 2018, 12, e0006628. [Google Scholar] [CrossRef]
- Aligholipour Farzani, T.; Földes, K.; Hanifehnezhad, A.; Yener Ilce, B.; Bilge Dagalp, S.; Amirzadeh Khiabani, N.; Ergünay, K.; Alkan, F.; Karaoglu, T.; Bodur, H.; Ozkul, A. Bovine herpesvirus type 4 (BoHV-4) vector delivering nucleocapsid protein of Crimean-congo hemorrhagic fever virus induces comparable protective immunity against lethal challenge in IFNα/β/γR−/− mice models. Viruses 2019, 11, 237. [Google Scholar] [CrossRef]
- Saunders, J.E.; Gilbride, C.; Dowall, S.; Morris, S.; Ulaszewska, M.; Spencer, A.J.; Rayner, E.; Graham, V.A.; Kennedy, E.; Thomas, K.; Hewson, R.; Gilbert, S.C.; Belij-Rammerstorfer, S.; Lambe, T. Adenoviral vectored vaccination protects against Crimean-Congo Haemorrhagic Fever disease in a lethal challenge model. eBioMedicine 2023, 90, 104523. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, S.E.; Cross, R.W.; Fenton, K.A.; Bente, D.A.; Mire, C.E.; Geisbert, T.W. Vesicular Stomatitis Virus-Based Vaccine Protects Mice against Crimean-Congo Hemorrhagic Fever. Sci Rep 2019, 9, 7755. [Google Scholar] [CrossRef] [PubMed]
- Dowall, S.D.; Graham, V.A.; Rayner, E.; Hunter, L.; Watson, R.; Taylor, I.; Rule, A.; Carroll, M.W.; Hewson, R. Protective effects of a Modified Vaccinia Ankara-based vaccine candidate against Crimean-Congo Haemorrhagic Fever virus require both cellular and humoral responses. PLoS One 2016, 11, e0156637. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Crimean-Congo hemorrhagic fever virus (CCHFV) virion structure and CCHFV vaccine platforms. (a) CCHFV virion contains three single-stranded RNA segments with a negative-sense orientation. Nucleoprotein (NP) and RNA-dependent RNA polymerase (RdRp; L protein) protect the RNA by enclosing the RNA segments and forming ribonucleoprotein complexes (RNPs). Once the ribonucleoprotein (RNP) complexes are formed, they are surrounded by a protective envelope that originates from the membrane of the host cell. This envelope is coated with specialized glycoproteins known as Gn and Gc. (b) CCHFV is consisting of three genomic segments - small (S), medium (M), and large (L). The S segment is responsible for encoding the NP within one open reading frame, while the small non-structural protein (NSs) is encoded in an opposite-sense open reading frame. The M segment is quite intricate, as it encodes a glycoprotein precursor (GPC) that undergoes processing by host proteases. This processing results in the production of a GP160/85 domain, which is then further processed into a mucin-like domain (MLD) and GP38. Additionally, the M segment encodes the Gn and Gc glycoproteins, as well as the medium non-structural protein (NSm). The L segment of CCHFV, which is distinctively larger than other bunyaviruses, encodes for the viral RNA-dependent RNA polymerase (RdRP) and an ovarian tumor-like protease (OTU) at its N terminus. (c) A diagrammatic representation of different CCHFV vaccine platforms. The diagram was created with Adobe Illustrator.
Figure 1.
Crimean-Congo hemorrhagic fever virus (CCHFV) virion structure and CCHFV vaccine platforms. (a) CCHFV virion contains three single-stranded RNA segments with a negative-sense orientation. Nucleoprotein (NP) and RNA-dependent RNA polymerase (RdRp; L protein) protect the RNA by enclosing the RNA segments and forming ribonucleoprotein complexes (RNPs). Once the ribonucleoprotein (RNP) complexes are formed, they are surrounded by a protective envelope that originates from the membrane of the host cell. This envelope is coated with specialized glycoproteins known as Gn and Gc. (b) CCHFV is consisting of three genomic segments - small (S), medium (M), and large (L). The S segment is responsible for encoding the NP within one open reading frame, while the small non-structural protein (NSs) is encoded in an opposite-sense open reading frame. The M segment is quite intricate, as it encodes a glycoprotein precursor (GPC) that undergoes processing by host proteases. This processing results in the production of a GP160/85 domain, which is then further processed into a mucin-like domain (MLD) and GP38. Additionally, the M segment encodes the Gn and Gc glycoproteins, as well as the medium non-structural protein (NSm). The L segment of CCHFV, which is distinctively larger than other bunyaviruses, encodes for the viral RNA-dependent RNA polymerase (RdRP) and an ovarian tumor-like protease (OTU) at its N terminus. (c) A diagrammatic representation of different CCHFV vaccine platforms. The diagram was created with Adobe Illustrator.
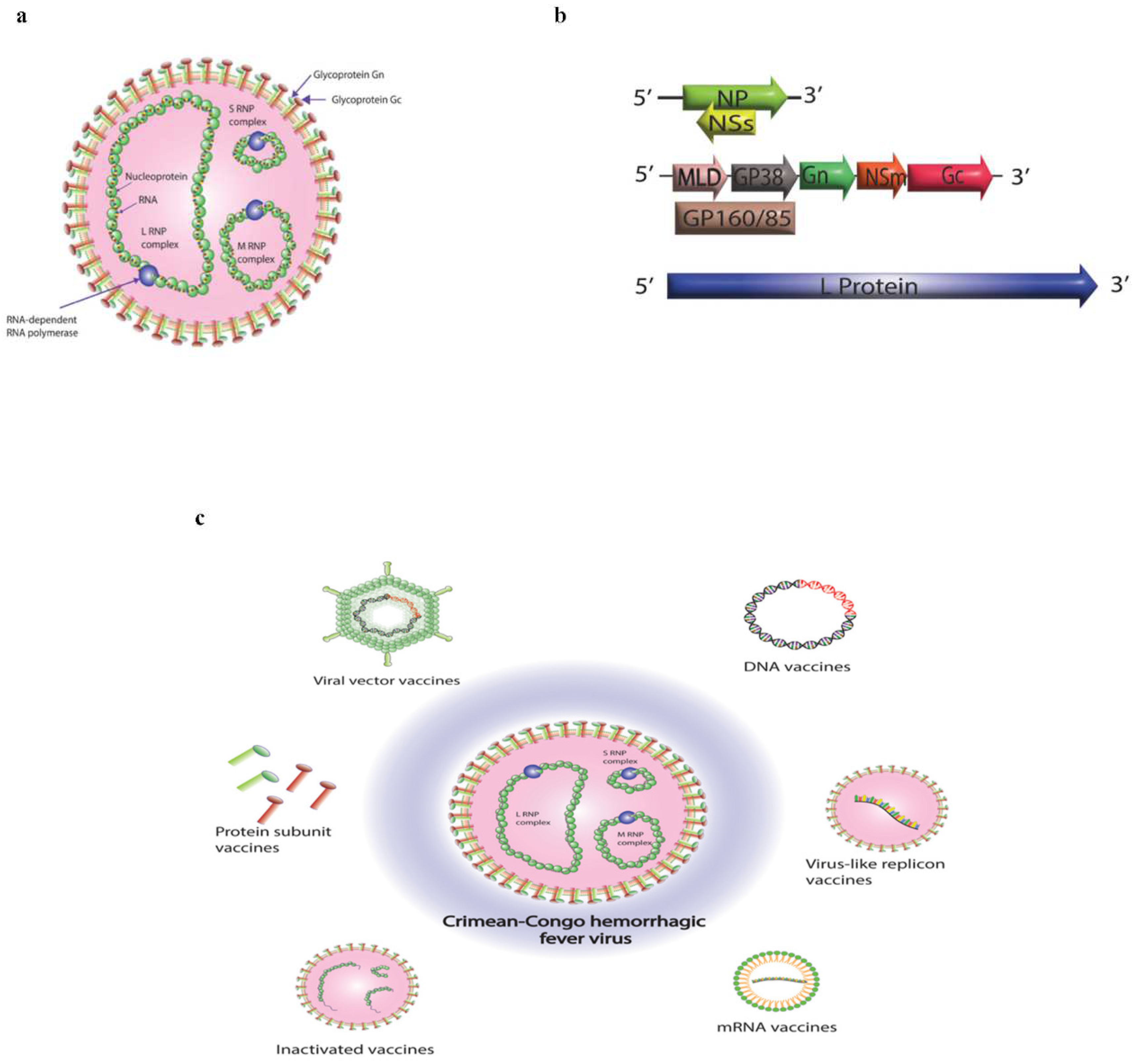
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



