
Submitted:
28 June 2023
Posted:
28 June 2023
You are already at the latest version
Abstract
The climate and its changes are major risk factors for agricultural activities because agricultural systems are highly affected by climatic factors such as temperature, rain, humidity, wind, and solar radiation. While some agricultural areas of the world are irrigated, the vast majority rely on natural rainfall patterns. Farmers will consume more water as a result of climate change and experience less water availability, resulting in a decrease in production capacity. As one of the most important natural phenomena affecting agricultural productivity, drought is a limiting environment for plant growth. The response of plants to water stress is complex and involves coordination between gene expression and its integration with hormones. It has been shown that bacteria have mechanisms to reduce water stress in these plant species and promote greater growth. The underlined mechanism involves root-to-shoot phenotypic changes in growth rate, architecture, hydraulic conductivity, water conservation, plant cell protection, and damage restoration through integrating phytohormones modulation, stress-induced enzymatic apparatus, and metabolites. This review demonstrates how plant growth-promoting bacteria can mitigate negative effects associated with water stress in plants and provide examples of technology conversion in agroecosystems.
Keywords:
sustainable agriculture
; biofertilizers
; PGPB
; endophytic bacteria
; abiotic stress
; inoculant
1. Introduction
Agricultural systems are directly affected by climate changes and global demands for food, fiber, and energy, which compromise food security and agroecosystem sustainability [1]. Due to fossil fuel demands associated with industrialization and urbanization, deforestation and intensive land use, which have increased greenhouse gas emissions, food chain production activities contribute significantly to global climate change trends. In this century, changes in temperature and precipitation regimes will be a significant constraint on food production. Abiotic factors account for about half of agricultural productivity losses, while biotic factors account for about 30% [1,2]. Agriculture faces a number of challenges under global climate change scenarios and economic constraints, including increasing crop yields or mitigating crop losses. [3]. As a result of an external factor that alters plant physiology, stress can be defined as any adverse change that occurs. In addition to temperature, drought, salinity, flooding, and heavy metals, abiotic stressors reduce plant productivity. An integrative approach to developing drought-tolerant varieties has been used in several studies by combining conventional breeding tools with modern breeding techniques. [4]. The use of plant breeding has been widely applied to select genotypes that are tolerant to diverse environmental stresses. Furthermore, several studies have been conducted to unravel the molecular bases and morpho-physiological traits associated with drought tolerance [5,6]. Environmental stresses can be mitigated by plant growth-promoting bacteria (PGPB) through two primary mechanisms, including water conservation mechanisms and protection-recovery mechanisms [7,8,9]. Drought-mitigating bacteria are being converted into a sustainable solution for agroecosystems, resulting in increased interest [10,11]. The most feasible biological technology for biotic and abiotic crop protection is microbial inoculants [12,13]. Using complex molecular machinery that is mediated by phytohormonal signaling, an induced enzymatic pool, and metabolites, this review examines how bacterial inoculants alleviate plant water stress to increase soil water accessibility and reduce plant water loss. It will also demonstrate how catalytic proteins and metabolites are coordinated to prevent damage to plant cells and trigger repair mechanisms that increase plant tolerance for water scarcity. Additionally, past and current drought stress mitigation strategies, technological applications, and future prospects for bacterial inoculants will be discussed.
2. Adaptation mechanisms of plants to drought
The greatest threat to global agricultural quality and productivity is drought, which limits plant growth and development. It is defined as a meteorological phenomenon characterized by prolonged periods of sub-normal rainfall that compromises soil moisture for a given crop [14]. Consequently, a decrease in water availability impacts plant growth and development negatively, affecting plant life cycles [15]. The water potential of the soil decreases during drought periods, thereby affecting the plant's water potential as well. In order to maintain the balance between soil, plants, and the atmosphere, plants' response to drought is a complex phenomenon involving a variety of molecular, biochemical, and physiological changes. Researchers conducted studies on grape leaves and demonstrated that the stomata completely closed before cavitation occurred, thus preventing cavitation and embolism [16,17]. CO2 influx decreases simultaneously, directly affecting photosynthetic capacity, while photorespiration increases simultaneously. Reduced carbon incorporation into plant biomass under water scarcity influences plant drought responses. To increase plant resilience to water stress, two main approaches must be interconnected. A combination of morpho-physiological mechanisms are employed to increase plant water status, primarily through hormonal signaling mechanisms (i.e. auxin, cytokinin, gibberellin) [18,19,20]. As well as signaling stress, they are essential for growth and development. In drought conditions, abscisic acid (ABA), a stress hormone, is present in significant amounts. In addition to regulating several genes, it promotes stomatal closure and dehydration tolerance [21,22]. A process known as osmotic adjustment can also be used to maintain cell turgor by modulating a cell's osmolarity by increasing solute concentrations within the cell (soluble sugars and glycine) [23]. These solutes prevent plant cells from becoming toxic during droughts in addition to maintaining turgor pressure [23]. The osmotic adjustment of pea roots [24] and sorghum [25] has been reported. In subsequent studies [26,27,28,29,30], osmotic adjustment mechanisms were demonstrated to maintain plant turgor [26,27,28,29,30]. It is important to note, however, that plant species/cultivars and the duration of stress events determine the degree of response to osmotic adjustment. As a result of the application of organic compounds as foliar sprays, plants are able to tolerate stress conditions better, which results in an increase in osmolytes accumulation, such as when sugar beets (Beta vulgaris var. saccharifera L.) [31], Catharanthus roseus [32], Brassica spp. [33], and Raphanus sativus L. [34]. Protecting and repairing machinery are associated with water conservation [10]. Similarly to other abiotic stresses, water deficit causes a decrease in electron transport chain activity, causing plant tissue to photooxidize [35]. As a result, the Rubisco enzyme activity declines and the PSII membrane complex is damaged, repressing photosynthetic activity [36]. An important photoprotective response to drought occurs in plants, as well as a number of molecular and biochemical mechanisms that lead to non-photochemical quenching (NPQ) of chlorophyll fluorescence. Induction of reactive oxygen species (ROS) is one of these well-studied mechanisms that can lead to membrane peroxidation and oxidative damage, impairing cellular functions [37]. Various enzymes are involved in plants' antioxidant defense system in order to reduce oxidative damage (superoxide dismutase (SOD), peroxidase (POX), catalase (CAT), and glutathione reductase (GR). As a result, drought tolerance correlates positively with antioxidant response. Plant species that are more tolerant display better antioxidant responses, resulting in higher levels of antioxidative enzyme activity, which protects the plant from oxidative damage [38]. Conversely, species whose enzyme activity machinery is less sensitive to drought show no such changes. Drought increases the activity of antioxidant enzymes, as shown in several studies. These antioxidant responses can differ between cultivars, as found in rice Oryza sativa L. [39], Triticum aestivum L. [40], and Hordeum vulgare L. [41]. Researchers have shown that drought-tolerant cultivars produce less ROS and oxidative stress than drought-sensitive cultivars as a result of their research. Through breeding and genetic engineering, these differences between drought-sensitive and drought-tolerant genotypes can be used to develop more resistant crops [42,43]. ROS play a critical role in maintaining cellular processes, but when they exceed normal levels, they have a toxic effect. It is therefore important to maintain a balance between cytostatic and cytotoxic ROS levels [35,36]. Plant fitness under environmental stress can be enhanced by microbial communities at the rhizosphere, root-shoot surfaces, and inner tissues (endophytes), in addition to the molecular, biochemical, and physiological responses triggered by drought [44,45]. In natural conditions, scientists are able to understand and enhance drought tolerance through the study of plant-bacteria interactions. In order to reduce the impact of drought on food security and agricultural production, researchers can gain insight into plant-microbe interactions that can lead to the development of drought-tolerant crops.
3. Growth-Promoting Bacteria in Plants
As a group of organisms, soil bacteria communities have the greatest diversity, abundance, and physiological activity, with bacterial phylotypes ranging from 102 to 106 per gram [46]. A new ecological niche is created when seeds or other reproductive plant structures are sown through the soil as the soil microbes act as a "seed bank" of species richness [47]. As described by the authors, rhizospheres are soil perimeters enriched with carbon exudates (including organic acids, amino acids, sugars, flavonoids), mucilaginous matrixes, and detached root cap cells. This environment is highly active and conducive to plant recruitment of taxa capable of colonizing the rhizosphere [48,49,50]. It is important to note that in the rhizosphere, beneficial and pathogenic microorganisms compete for colonization of plant tissues, modulating nutrient flux through the soil-plant system and affecting the growth of plants [51]. Bacteria have played a crucial role as the most common and active fraction interacting with plant hosts among such microorganisms [52]. It has been reported that biofertilizers, biostimulants, and bioprotections enhance plant growth, prevent pathogens, and mitigate environmental stresses, according to several reports. Klebsiella, Paraburkholderia, Azospirillum, Herbaspirillum, Gluconacetobacter, Serratia, Azotobacter, and many others belong to the PGPB family in addition to Pseudomonas and Enterobacter. Plant growth is promoted by these microorganisms in two ways: (1) biological nitrogen fixation; (2) indirect mechanisms: reducing damage caused by pathogens and/or environmental stresses by producing hormones (indole-3-acetic acid, gibberellic acid, and cytokinin, such as zeatin) and acquiring essential nutrients; and (3) indirect mechanisms: relating to biocontrol, which involves mitigating damage caused by pathogens and environmental stresses. [13,44,51,53,54,55,56,57] (Figure 1).
To reduce damage caused by environmental stress on plants, several mechanisms have already been identified in the reactions and interactions between PGPB and plants. [58,59]. In addition to producing hormones (auxin, cytokinin, abscisic acid), exopolysaccharides and beneficial enzymes, including 1-aminocyclopropane-1-carboxylate deaminase (ACC deaminase), trehalose and volatile organic compounds, as well as osmoregulation, there are also responses related to osmoregulation. PGPB resists drought by a variety of mechanisms as shown in Table 1.
4. Plant Growth-Promoting Bacteria (PGPB) and stress tolerance
4.1. Plant morpho-physiological traits are modulated by bacterial phytohormones
A complex network of plant hormones is necessary for beneficial microorganisms to promote plant growth. PGPB's interaction with plants results in a number of changes, including changes in hormonal homeostasis, as well as responses to biotic and abiotic stimuli [66,67,68]. Some microorganisms can affect hormone concentrations, locations, and signaling in plants [68,69,70]. Plant hormones play an important role in promoting plant growth through beneficial microorganisms (Figure 2).
Plant hormone auxin has a number of functions including cell division, expansion, and differentiation [71]. Seeds and tubers are stimulated by auxinic activity, xylem and roots are developed at a faster rate, vegetative growth processes are controlled, lateral and adventitious roots are formed, light, gravity, and flowering are mediated, and synthesis of photosynthesis, pigments, metabolites, and resistance to stressful conditions are all affected [72]. A number of PGPBs regulate auxin balance, which affects the growth rate and architecture of roots [69,72,73,74]. Microbes inoculate crops in such a way as to modify root anatomy and biochemistry through the modulation of phytohormones [51,53,55,69,73]. As a matter of interest, these plant-growth promotion effects are crucial to increasing water availability in environmental scarcity situations with a low soil matrix water potential. By increasing root formation (root ramification), root hair density, and root hair length (specific surface enhancement), absorptive root structures increase water uptake. This results in an increase in root surface, volume, and biomass. Additionally, the root system's hydraulic conductivity is modulated by changes in the organization pattern of epidermal, cortical, and vascular root tissue systems [75]. Variables include the number and arrangement of cell layers, water flux resistance, metaxylem size, diameter, and distribution, and water channel transmembrane transporters (such as aquaporin). Additionally, auxin signaling activates electrogenic transmembrane pumps (P-type H+ ATPase at the plasma membrane and V-type H+ ATPase at the vacuolar membrane) which create electrochemical gradients to facilitate secondary nutrient transport. Furthermore, P-type pumps acidify the apoplast microenvironment near the meristematic tip of recently divided cells, which promotes growth of the root axes and tissue proliferation [76,77]. The inoculation of bacteria facilitates plant growth under appropriate water availability or alleviates water scarcity negative effects by modulating auxin signaling and balance. When drought occurs, abscisic acid (ABA), a stress hormone, is detected significantly. ABA promotes stomatal closure and regulates genes involved in dehydration tolerance [22]. The ability of some plants to cope with abiotic stresses is attributed to exogenous ABA provided by bacteria of a particular genus, according to Cohen and collaborators [78]. When maize plants were inoculated with Azospirillum brasilense strains Ab-V5 and Ab-V6, they recovered better from a prolonged drought [79]. Microorganisms and plants interact to control the hormonal balance of ABA in plants, thus promoting plant growth even under stressful conditions. Curá and collaborators [67], demonstrated that inoculating maize plants with Azospirillum and Herbaspirillum directly alters molecular, biochemical, and physiological processes. PGPB inoculation also induces the accumulation of ABA in Vitis vinifera plants, according to Salomon and collaborators [80]. Under water scarcity, bacterial-inoculated plants generate convergent action mechanisms to improve water use efficiency as a result of complex crosstalk between auxin and ABA. In plants, auxiliary signals contribute to water uptake and transport, while ABA signals reduce transpiration losses. Dual-mode actions increase plant tissue water content under stressful conditions. When plants are exposed to severe drought, they display a survival phenotype, and when they are exposed to mild drought, microbial inoculation promotes their growth and development [81]. A number of experiments have shown significant increases in fresh biomass under greenhouse and field conditions despite a non-significant accumulation of dry biomass due to microbial inoculation. Due to the increased water content in plants' bodies, inoculating them with bacteria improved their fitness in water-scarce environments. As a result of its integrative role in water conservation in plant cells and tissues, neither the auxin–ABA signaling network nor the osmoregulation mechanism can be considered separately. However, changes in plant microstructure would increase plant water storage and circulation if they were based on plant phenotypic plasticity [82]. Plant cells vacuolize more, specialized cells for water storage increase, and apoplastic and symplastic compartment volumes change [75,83]. This integrated mechanism of plant water conservation also stimulates plant response enzymatic-metabolic machinery that protects cells and restores damage (i.e., ROS produced as a consequence of damage to biological membranes and biomolecules). When atmospheric temperatures rise and soil water availability declines, leaf water potential decreases, reducing stomatic conductance and transpiration and decreasing photosynthetic rates until the stomata are completely closed, preventing water loss in plant tissue and reducing photosynthetic activity [84,85,86,87]. The inoculation of bacteria can increase net photosynthetic activity to some extent compared with non-inoculated plants with similar stomatic conductance values. Inoculated bacteria are believed to increase water use efficiency by increasing carbon dioxide influx or reducing respiration rates at a similar rate to water loss from leaf blade substomatic chambers, which leads to an additional acquisition of carbon to meet the energetic requirements needed to restore cell homeostasis [88,89]. Plant growth and development are influenced by ethylene gas, another phytohormone, in several ways, but the underlying mechanism remains unclear. In addition to initiating roots, inhibiting root elongation, promoting fruit ripening, causing flower wilting, stimulating seed germination, promoting leaf abscission, and responding to biotic and abiotic stresses, it also stimulates flower wilting, stimulates seed germination, stimulates leaf abscission, and activates plant hormone synthesis. In response to stressful conditions, plants can increase ethylene synthesis. In order to synthesize ethylene, 1-aminocyclopropane-1-carboxylate is required. The first component of this hormone is methionine, which is converted by SAM synthase (SAM synthase) into S-adenosylmethionine (SAM) and ACC synthase into ACC. In response to this, ACC concentrations increase, and so do ethylene levels. However, at high concentrations, ACC inhibits crop growth and yield. Honma and Shimomura first characterized 1-aminocyclopropane-1-carboxylate deaminase (ACC deaminase) in PGPB [90], under stressful conditions, it promotes plant growth. According to Glick et al. [91], PGPB synthesizes and secretes auxin, which promotes plant growth when tryptophan is present. Also, auxin induces S-adenosylmethionine to be converted into ACC by 1-aminocyclopropane-1-carboxylate synthase (ACC synthase). A plant exudes ACC to maintain a balance between the internal and external concentration of ACC, which in turn decreases the outside concentration of ACC. Due to ACC's role as a precursor to ethylene, a reduction in ACC directly leads to a decrease in plant ethylene levels, which promotes plant growth even under limited conditions [92]. As a result of bacteria containing this enzyme, plants can withstand both biotic and abiotic stresses [92]. Inoculation of bacteria that synthesize ACC deaminase under abiotic stress is an excellent growth promoter for plants [60,64,91,92].
4.2. Biosynthetic Bacteria and Induced Accumulation in Plants
Osmolytes are synthesized by plant growth-promoting bacteria and secreted together with other exuded compounds [93]. In addition to osmolytes produced by plants (such as glycine-betaine, soluble sugars, trehalose, and proline), plant-produced osmolytes serve as osmoprotectants. Plants are more likely to tolerate water stress when these microbes accumulate osmolytes [59,61,62,66,67]. Under water deficit conditions, plants inoculated with PGPB may be able to tolerate stress, maintain cell turgor and membrane stability, and prevent electrolyte leakage due to increased proline levels. Plants produce proline as a result of Arthrobacter sp. and Bacillus spp. inoculated [94]. The drought-tolerant strains of Bacillus spp. There was an increase in proline, sugars, and free amino acids in sunflowers (Helianthus annuus L.) and maize (Zea mays L.) when antioxidant enzyme activity was reduced. When Aridopsis thaliana was inoculated with Azospirillum baldaniorum strain Sp 245 under water deficit conditions, proline levels and relative water content were increased, improving the plants' performance under drought conditions [78]. A plant cell's cytoplasmatic compartment has an increased osmolyte content as a result of bacterial inoculants (secreted by microbes or induced by microbes), resulting in a decrease in osmotic pressure, preventing water efflux. To balance water and flux within the plant body, phytohormonal imbalances serve as an orchestrated connection mechanism. The improved hydrated microenvironment promotes photosynthesis recovery, thus protecting subcellular compartments from damage [95].
4.3. A self-protective and water-retaining property of bacterial exopolysaccharides
As well as being highly heterogeneous, exopolysaccharides (EPSs) have a high molecular weight. It has been demonstrated that microorganisms in extreme environments secrete a variety of monosaccharides, soluble in water and composed of sugar residues [96], as a means of protecting themselves. Thus, EPS synthesis is one of the most common mechanisms for bacterial self-defense. To grow, adhere to solid surfaces, and survive harsh conditions, bacteria synthesize this compound, which accounts for 40% to 95% of the bacteria's weight. Furthermore, they are essential for forming and maintaining biofilm architecture, retaining water, absorbing nutrients, and ensuring survival in harsh environments [97]. In stressful conditions, EPS is beneficial for both bacteria and plants [96]. Due to their ability to increase soil water retention capacity, bacteria that synthesize exopolysaccharides are essential for promoting plant growth under stressful conditions, such as drought. It is noteworthy that these bacteria are more advantageous and have gained prominence in being used as bio-inoculants for drought tolerance in plants [98]. EPSs have been found to be secreted by bacteria from the genera Bacillus, Pseudomonas, and Azospirillum as well as other microorganisms under water stress [96,98]. Moreover, these compounds can also act as emulsifiers and mitigate ROS-induced effects, as well as alter root structure. Additionally, plants inoculated with bacteria capable of synthesizing EPS accumulate more proline, sugars, and free amino acids, as well as more biomass, leaf area, and protein content [99].
4.4. The role of volatile organic compounds in drought bioprotection by bacteria
Many publications have discussed the potential of PGPB to increase crop yields. As plants develop, they release low molecular weight lipophilic compounds (300 moles. L–1) as a response to biotic and abiotic stress and as a result of gaseous organic compounds (VOCs) [100]. Plants release volatile organic compounds during stressful periods, reducing ROS effects and increasing membrane protection. As a signal molecule, VOCs emitted by microorganisms can be found in the rhizosphere [101]. The first demonstration of this mechanism was made when Bacillus subtilis was inoculated into Arabidopsis thaliana [102]. The compounds are synthesized by many bacteria, including Burkholderia, Pantoea, Serratia, Chromobacterium, Arthrobacter, Proteus, Bacillus, Fusarium, Pseudomonas, Alternaria, and Laccaria. As a result of modulating essential nutrients, hormonal balance, metabolism, and sugar concentrations [100], these organisms release a variety of compounds that are specific to different metabolic pathways, signaling a variety of plant physiological processes and promoting growth. These studies [101,102,103,104] demonstrated that VOCs can be effective for promoting plant growth by using A. thaliana. It has already been demonstrated by some researchers [105,106,107] that these compounds promote plant growth and reduce stress, but most of these studies have been conducted in controlled laboratory settings. Because these compounds are highly biodegradable and reactive, agriculture does not commonly use them. Further research is needed into these compounds, their perception mechanisms in plant tissues, and their application techniques [108].
4.5. Mechanisms of bacterial protection and repair in drought-stressed plant tissue
It has been shown that drought stress leads to the generation of reactive oxygen species (ROS) within subcellular compartments, causing free radicals and redox imbalances (oxidative damage) that damage macromolecules (such as biological membranes) and compromise plant health. There is a differential tolerance to drought stress in plant tissues that is influenced by antioxidant enzymes [37,40,41]. Plant antioxidant enzymes are modulated by bacterial inoculants, which decrease ROS levels, thereby enhancing crop protection. Researchers have found that under stressful conditions, enzymatic antioxidants (such as ascorbate peroxidase, catalases, peroxidases, glutathione reductase, superoxide dismutase) and nonenzymatic antioxidants (such as ascorbic acid, flavonoids, and phenolic compounds) are more effective. ROS accumulation impairs photosynthetic activity, compromising antenna harvesting, electron transport, enzyme activity, and chloroplast membrane function [109]. Inoculating Bacillus pumilus improved the photosynthetic efficiency of Glycyrrhiza uralensis under drought stress [110,111,112]. Bacillus inoculation maintained chloroplasts and mitochondrial integrity. Plants can benefit from a combination of beneficial bacteria and bioactive products to boost their antioxidant response. A combination of humic acids and diazotrophic bacteria enhanced drought stress recovery in sugarcane [89]. Inoculating leaves with bacteria preserved their water potential and increased relative water content, while humic acids activated antioxidant enzymes to mitigate water stress.
5. Drought Stress Mitigation Using Microbial Inoculants in Agroecosystems
In current times, industrial fertilizers, mainly derived from nonrenewable resources, pose a significant environmental problem. They contribute significantly to the degradation of the ozone layer, greenhouse gas emissions, low-efficiency recovery by plants, as well as the high cost of production. In order to reduce agriculture's environmental impact on local and global levels, bioinoculants and other biological products have become increasingly important as sustainable technologies [113]. Bioinputs include formulations containing live microorganisms (fungi, bacteria, and algae) and/or their metabolites that are used to fertilize, stimulate, and protect agroecosystems [12,73,113]. It is possible to inoculate plant surfaces, soil, or seeds with bioinoculants (delivering niches) in various physicochemical formulations (microbiological composition, carriers, and additives), as well as to apply them at the right time, depending on the crop's physiological state. By colonizing plant surfaces, interiors, or rhizospheres, microorganisms boost plant growth: (a) enhancing nutrient availability in the plant-soil system (i.e., biological nitrogen fixation, mineral solubilization, organic compound mineralization); and (b) enhancing nutrient absorption through hormonal action (auxin, cytokinin, gibberellin, abscisic acid) that results in changes in plant morphology that improves nutrient utilization [12,53,73]. The first microbial inoculants used in agriculture were introduced by Nobbe and Hiltner (1895), who introduced "Nitragin," which contains rhizobia. In addition, numerous studies have examined the use of microorganisms as bioinoculants for the promotion of plant growth in sugarcane, rice, soybeans, beans, chickpeas, tomatoes, maize, tropical fruits, and wheat, among others [12,79,114,115,116,117,118]. A rapid increase in farmers' adoption of bioinoculants has been observed [12], leveraging innovation and technologies to fulfill the growing demand for bioproducts. It is estimated that Bradyrhizobium spp. and Azospirillum brasilense are used mainly as biofertilizers in global agriculture [12]. Plant growth is primarily promoted by nitrogen fixation and increased nutrient uptake by roots (Figure 3). There are, however, a variety of side effects associated with commercial inoculants, such as an "increase in water absorption and saline stress" and "produces phytohormones that promote root development, which increases water and nutrient absorption and improves resistance to drought and salinity." However, some mechanisms do not directly promote plant growth promotion, despite the fact that microbial inoculations provide complementary effects on soil nutrient availability and root uptake. The plants, however, rely heavily on them to protect themselves from adverse environmental conditions, mitigating biotic and abiotic stresses. These mechanisms include ACC deaminase activity, ROS-enzyme synthesis, EPSs, volatile organic compounds, and osmolytes, as well as induced systemic resistance [36,45,61,80,94,99,119]. In previous technological attempts, bacteria were collected under non-selective pressure conditions that were used to screen candidate strains. A better-performing isolate was screened and further evaluated in a greenhouse and an open field using assays involving water activity reduction (i.e. osmotic active molecules) [96,108], or studies involving progressive cell-bacterium dissection [120,121]. In bacterial selection programs, plant-growth-enhancing traits and water deficit tolerance traits have also been regarded as addictive traits. Plants adapted to harsh environments have emerged with a new generation of microbial products based on microbe-driven selection for rhizospheres, rhizoplanes, and inner tissues. As abiotic stress affects bacteria cell phenology, an increasing number of studies have been conducted [122]. The secondary metabolites of a halotolerant PGPR could induce tomato salinity stress tolerance by producing auxin and ACC-deaminase [123,124]. By analyzing the entire genomes of bacterial candidates for inoculant formulations, it is possible to conduct phenotypic screening. In Bacillus altitudinis (strain FD48), previously shown to induce antioxidant stress in rice under drought, genes related to distinct mechanisms of water stress evaluation were discovered. Jochum and colleagues [125] proposed a bacterial bioprospecting screen that combines effective root colonization and drought stress mediation to design microbial products for drought stress. PGPBs were selected from perennial grass rhizospheres in a semi-arid environment, prescreened in the laboratory for desired plant phenotypes (delayed symptoms of water scarcity), and (c) bacteria isolates were selected to be formulated and delivered as soon as water stress was detected (rapid colonizers and adequate crop protection). As a result of bioprospecting, another sudamerican commercial bacterial inoculant that relieves plant water stress was developed. Using rhizobacteria for drought-tolerant plant growth, researchers led by Melo [122] examined cacti-associated bacteria from semi-arid environments.
6. Conclusions
Plant survival and resilience under stress is influenced by a series of physiological, biochemical, and molecular complex network responses. It is well known that there are two core mechanisms: one that conserves plant water and one that protects and restores plant damage. In addition to reducing drought stress in plants, bacteria that promote plant growth have also been shown to be effective. In order to increase crop yields by using strategies and techniques that support water deficits, it is still necessary to have a comprehensive understanding of these response mechanisms triggered by microorganisms. However, with current technological advancements, we are able to formulate, apply, and design bacterial inoculants that can help plants cope with droughts. In our knowledge, microbial bacteria can mitigate the adverse effects and reduce agricultural productivity. Inoculants for drought stress mitigation should be designed according to the main mechanisms that enhance plant–microbe interactions under drought tolerance. The survival and efficacy of bacterial inoculum are affected by field conditions. A bacterial cell maintains its viability by accumulating osmoprotectants, antioxidants, genes related to stress, and essential proteins. Microorganisms can produce and/or modulate several hormonal classes, which are regulated by changes in concentrations, locations, and signaling of hormones, which affect plant concentrations and balances, making it important to characterize drought-induced responses molecularly, and identify hormonal homeostasis. By altering plant structural changes triggered by ABA signaling, it is also possible to increase plant water content during water shortages in inoculated plants. Inoculating plants with BPCV could increase plant response under drought conditions or increase agricultural productivity by altering the hormonal balance of ABA in plants, since abscisic acid is a sign of plant stress. Using this information, we can select stress-tolerant microorganisms and improve the use of BPCVs to reduce the damage observed in agricultural production systems. New generations of bacterial inoculants aimed at mitigating water stress in plants could benefit from recent initiatives involving bacterial bioprospection under appropriate selective pressure (arid environments). This involves the distinct soil-plant compartments (rhizosphere, rhizoplane, and inner tissue) under intense selective pressure and constant water deficits. We also selected bacteria strains that produced exopolysaccharides (EPSs) under osmotic stress as well as batch reactor growth media and inoculant formulations that stimulated EPS secretion. In EPS systems that are rich in microenvironments, water is trapped and desiccation is reduced, allowing bacteria to thrive. A formulation containing synthetic microbial communities was developed using metataxonomic and metagenomic data gathered from plant microbiomes cultivated in drought-stressed environments. Furthermore, proper formulations contain additives or carriers that promote bacteria's survival or protect plants from abiotic stressors (humic substances).
Author Contributions
Conceptualization, D.P.; methodology, writing—original draft preparation D.P. and R.F.; software and investigation, D.P.; writing—review and editing, D.P. and R.F.; funding acquisition, D.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the CREA Research Centre for Vegetable and Ornamental Crops.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data, tables, and figures in this manuscript are original.
Acknowledgments
The authors would like to express his heartfelt gratitude to colleagues and providers of plants and to all other sources for their cooperation and guidance in writing this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Malhi, G.S.; Kaur, M.; Kaushik, P. Impact of Climate Change on Agriculture and Its Mitigation Strategies: A Review. Sustainability 2021, 13, 1318. [Google Scholar] [CrossRef]
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Alam, S.; Khalil, S.; Ayub, N.; Rashid, M. In vitro solubilization of inorganic phosphate by phosphate solubilizing microorganism (PSM) from maize rhizosphere. Intl. J. Agric. Biol. 2002, 4, 454–458. [Google Scholar]
- Ashraf, M. Inducing drought tolerance in plants: Recent advances. Biotechnol. Adv. 2010, 28, 169–183. [Google Scholar] [CrossRef]
- Akhtar, M.S.; Siddiqui, Z.A. Effect of phosphate solubilizing microorganisms and Rizobium sp. on the growth, nodulation, yield and root-rot disease complex of chickpea under field condition. Afr. J. Biotech. 2009, 8, 3489–3496. [Google Scholar]
- Rao, D.E.; Chaitanya, K.V. Photosynthesis and antioxidative defense mechanisms in deciphering drought stress tolerance of crop plants. Biol. Plant. 2016, 60, 201–218. [Google Scholar]
- Berg, G. Plant-microbe interactions promoting plant growth and health: perspectives for controlled use of microorganisms in agriculture. Appl. Microbiol Biotech. 2009, 84, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Broeckling, C.D. 2008. Root exudates regulate soil fungal community composition and diversity. Appl. Environ Microbiol. 2008, 74, 738–744. [Google Scholar] [CrossRef]
- Li, C.; Li, L.; Reynolds, M.P.; Wang, J.; Chang, X.; Mao, X.; Jing, R. Recognizing the hidden half in wheat: Root system attributes associated with drought tolerance. J. Exp. Bot. 2021, 72, 5117–5133. [Google Scholar] [CrossRef]
- Deubel, A.; Gransee, G.; Merbach, W. Transformation of organic rhizodeposits by rhizoplane bacteria and its influence on the availability of tertiary calcium phosphate. J. Plant Nutr. Soil Sci. 2000, 163, 387–392. [Google Scholar] [CrossRef]
- Rasheed, A.; Mahmood, A.; Maqbool, R.; Albaqami, M.; Sher, A.; Sattar, A.; Khosa, G.B.; Nawaz, M.; Hassan, M.U.; Al-Yahyai, R. Key insights to develop drought-resilient soybean: A review. J. King Saud Univ.-Sci. 2022, 34, 102089. [Google Scholar] [CrossRef]
- Gull, M.; Hafeez, F.E.; Saleem, M.; Malik, K.A. Phosphorus uptake and growth promotion of chickpea by co-inoculation of mineral phosphate solubilizing bacteria and a mixed rhizobial culture. Aust J. Exp Agric. 2004, 44, 623–628. [Google Scholar] [CrossRef]
- Poudel, M.; Mendes, R.; Costa, L.A.S.; Bueno, C.G.; Meng, Y.; Folimonova, S.Y.; Garrett, K.A.; Martins, S.J. The role of plant- associated bacteria, fungi, and viruses in drought stress mitigation. Front. Microbiol. 2021, 12, 3058. [Google Scholar] [CrossRef] [PubMed]
- Duponnois, R.; Kisa, M.; Plenchette, C. Phosphate solubilizing potential of the nemato fungus Arthrobotrys oligospora. J. Plant Nutr. Soil Sci. 2006, 169, 280–282. [Google Scholar] [CrossRef]
- Dutton, V.M.; Evans, C.S. Oxalate production by fungi: its role in pathogenicity and ecology in the soil environment. Can J. Microbiol. 1996, 42, 881–895. [Google Scholar] [CrossRef]
- Cochard, H.; Coll, L.; Le Roux, X.; Améglio, T. Unraveling the effects of plant hydraulics on stomatal closure during water stress in walnut. Plant Physiol. 2002, 128, 282–290. [Google Scholar] [CrossRef]
- Hochberg, U.; Windt, C.W.; Ponomarenko, A.; Zhang, Y.-J.; Gersony, J.; Rockwell, F.E.; Holbrook, N.M. Stomatal closure, basal leaf embolism, and shedding protect the hydraulic integrity of grape stems. Plant Physiol. 2017, 174, 764–775. [Google Scholar] [CrossRef]
- Habibi, A.; Heidari, G.; Sohrabi, Y.; Badakhshan, H.; Mohammadi, K. 2011. Influence of bio, organic and chemical fertilizers on medicinal pumpkin traits. J. Med. Plants Res. 2011, 5, 5590–5597. [Google Scholar]
- Iqbal, S.; Wang, X.; Mubeen, I.; Kamran, M.; Kanwal, I.; Díaz, G.A.; Abbas, A.; Parveen, A.; Atiq, M.N.; Alshaya, H.; et al. Phytohormones Trigger Drought Tolerance in Crop Plants: Outlook and Future Perspectives. Front. Plant Sci. 2022, 12, 3378. [Google Scholar] [CrossRef]
- Bandurska, H. Drought Stress Responses: Coping Strategy and Resistance. Plants 2022, 11, 922. [Google Scholar] [CrossRef]
- Henri, F.; Laurette, N.N.; Annette, A.; John, Q.; Wolfgang, M.; François-Xavier, E.; Dieudonné, E. Solubilization of inorganic phosphates and plant growth promotion by strains of Pseudomonas fluorescens isolated from acidic soils of Cameroon. Afri J. Microbiol Res. 2008, 2, 171–178. [Google Scholar]
- Hilda, R.; Fraga, R. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotech Adv. 2000, 17, 319–359. [Google Scholar]
- Turner, N.C. Turgor maintenance by osmotic adjustment, an adaptive mechanism for coping with plant water deficits. Plant Cell Environ. 2017, 40, 1–3. [Google Scholar] [CrossRef]
- Greacen, E.L.; Oh, J.S. Physics of Root Growth. Nat. New Biol. 1972, 235, 24–25. [Google Scholar] [CrossRef]
- Constable, G.A.; Hearn, A.B. Agronomic and physiological responses of soybean and sorghum crops to water deficits I. Growth, development and yield. Funct. Plant Biol. 1978, 5, 159–167. [Google Scholar] [CrossRef]
- Hinsinger, P. Bioavailability of soil inorganic P in the rhizosphere as affected by root induced chemical changes: a review. Plant Soil. 2001, 237, 173–195. [Google Scholar] [CrossRef]
- Igual, J.M.; Valverde, A.; Cervantes, E.; Velázquez, E. Phosphate-solubilizing bacteria asinoculants for agriculture: use of updated molecular techniques in their study. Agronomie. 2001, 21, 561–568. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef]
- Chen, H.; Jiang, J.-G. Osmotic adjustment and plant adaptation to environmental changes related to drought and salinity. Environ. Rev. 2010, 18, 309–319. [Google Scholar] [CrossRef]
- Kathiresan, G.; Manickam, G.; Parameswaran, P. 1995. Efficiency of phosphobacteria addition on cane yield and quality. Cooperative Sugar. 1995, 26, 629–631. [Google Scholar]
- Hussein, H.-A.A.; Mekki, B.B.; Abd El-Sadek, M.E.; El Lateef, E.E. Effect of L-Ornithine application on improving drought tolerance in sugar beet plants. Heliyon 2019, 5, e02631. [Google Scholar] [CrossRef] [PubMed]
- Chang, B.; Yang, L.; Cong, W.; Zu, Y.; Tang, Z. The improved resistance to high salinity induced by trehalose is associated with ionic regulation and osmotic adjustment in Catharanthus roseus. Plant Physiol. Biochem. 2014, 77, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.M.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Trehalose-induced drought stress tolerance: A comparative study among different Brassica species. Plant Omics 2014, 7, 271–283. [Google Scholar]
- Akram, N.A.; Waseem, M.; Ameen, R.; Ashraf, M. Trehalose pretreatment induces drought tolerance in radish (Raphanus sativus L.) plants: Some key physio-biochemical traits. Acta Physiol. Plant. 2016, 38, 3. [Google Scholar] [CrossRef]
- Khan, M.S.; Zaidi, A.; Wani, P.A. Role of phosphate-solubilizing microorganisms in sustainable agriculture - A review. Agron Sustain Dev. 2007, 27, 29–43. [Google Scholar] [CrossRef]
- Kim, K.Y.; Jordan, D.; Mc Donald, G.A. Effect of phosphate-solubilizing bacteria and vesicular-arbuscular mycorrhizae on tomato growth and soil microbial activity. Biol. Fertil Soils. 1998, 26, 79–87. [Google Scholar] [CrossRef]
- Sohbat, Z.I. Non-photochemical quenching of chlorophyll fluorescence and its components–recent advances. J. Life Sci. Biomed. 2022, 4, 76–86. [Google Scholar]
- Hernández, I.; Cela, J.; Alegre, L.; Munné-Bosch, S. Antioxidant Defenses Against Drought Stress. In Plant Responses to Drought Stress: From Morphological to Molecular Features; Aroca, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 231–258. [Google Scholar]
- Pyngrope, S.; Bhoomika, K.; Dubey, R.S. Oxidative stress, protein carbonylation, proteolysis and antioxidative defense system as a model for depicting water deficit tolerance in Indica rice seedlings. Plant Growth Regul. 2013, 69, 149–165. [Google Scholar] [CrossRef]
- Sheoran, S.; Thakur, V.; Narwal, S.; Turan, R.; Mamrutha, H.M.; Singh, V.; Tiwari, V.; Sharma, I. Differential activity and expression profile of antioxidant enzymes and physiological changes in wheat (Triticum aestivum L.) under drought. Appl. Biochem. Biotechnol. 2015, 177, 1282–1298. [Google Scholar] [CrossRef]
- Harb, A.; Awad, D.; Samarah, N. Gene expression and activity of antioxidant enzymes in barley (Hordeum vulgare L. ) under controlled severe drought. J. Plant Interact. 2015, 10, 109–116. [Google Scholar]
- Kpomblekou, K.; Tabatabai, M.A. Effect of organic acids on release of phosphorus from phosphate rocks. Soil Sci. 1994, 158, 442–453. [Google Scholar] [CrossRef]
- Lee, K.E.; Pankhurst, C.E. Soil organisms and sustainable productivity. Australian J. Soil Res. 1992, 30, 855–892. [Google Scholar] [CrossRef]
- Nahas, E. Factors determining rock phosphate solubilization by microorganism isolated from soil. World J. Microb Biotech. 1996, 12, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Nihorimbere, V.; Ongena, M.; Smargiassi, M.; Thonart, P. Beneficial effect of the rhizosphere microbial community for plant growth and health. Biotechnol Agron Soc Environ. 2011, 15, 327–337. [Google Scholar]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef]
- Ponmurugan, P.; Gopi, G. Distribution pattern and screening of phosphate solubilizing bacteria isolated from different food and forage crops. J. Agron. 2006, 5, 600–604. [Google Scholar]
- Rodrguez, H.; Fraga, R. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotech Adv. 1999, 17, 319–339. [Google Scholar] [CrossRef]
- Rokhzadi, A.; Asgharzadeh, A.; Darvish, F.; Nourmohammadi, G.; Majidi, E. Influence of plant growth-promoting rhizobacteria on dry matter accumulation and yield of chickpea (Cicer arietinum L. ) under field condition. Am-Euras. J. Agric. Environ. Sci. 2008, 3, 253–257. [Google Scholar]
- Jjemba, P.K.; Alexander, M. Possible determinants of rhizosphere competence of bacteria. Soil Biol. Biochem. 1999, 31, 623–632. [Google Scholar] [CrossRef]
- Rokhzadi, A.; Toashih, V. Nutrient uptake and yield of chickpea (Cicer arietinum L.) inoculated with plant growth promoting rhizobacteria. Aust J. Crop Sci. 2011, 5, 44–48. [Google Scholar]
- Rosas, S.B. , Andre´ s J.A., Rovera M. and Correa N.S. Phosphate-solubilizing Pseudomonas putida can influence the rhizobia-legume symbiosis. Soil BiolBiochem. 2006, 38, 3502–3505. [Google Scholar] [CrossRef]
- Olivares, F.L.; Busato, J.G.; de Paula, A.M.; da Silva Lima, L.; Aguiar, N.O.; Canellas, L.P. Plant growth promoting bacteria and humic substances: Crop promotion and mechanisms of action. Chem. Biol. Technol. Agric. 2017, 4, 30. [Google Scholar] [CrossRef]
- Kumar, A.; Verma, J.P. Does plant—Microbe interaction confer stress tolerance in plants: A review? Microbiol. Res. 2018, 207, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Etesami, H.; Maheshwari, D.K. Use of plant growth promoting rhizobacteria (PGPRs) with multiple plant growth promoting traits in stress agriculture: Action mechanisms and future prospects. Ecotoxicol. Environ. Saf. 2018, 156, 225–246. [Google Scholar] [CrossRef] [PubMed]
- Saikia, S.P.; Jain, V. Biological nitrogen fixation with non- legumes:an achievable able target or a dogma. Curr. Sci. 2007, 92, 317–322. [Google Scholar]
- Schroth, M.N.; Hancock, J.G. Selected topics in biological control. Ann Rev Microbiol. 1981, 35, 453––476. [Google Scholar] [CrossRef]
- Shanmugam, P.M.; Veeraputhran, R. Effect of organic manure, biofertilizers, inorganic nitrogen and zinc on growth and yield of rabi rice. Madras Agric J. 2000, 2, 87–90. [Google Scholar]
- Shehata, M.M.; El-khawas, S.A. Effect of biofertilizers on growth parameters, yield characters, nitrogenous components, nucleic acids content, minerals, oil content, protein profiles and DNA banding pattern of sunflower (Helianthus annus L. cv. Vedock) yield. Pak. J. Biol. Sci. 2003, 6, 1257–1268. [Google Scholar] [CrossRef]
- Arzanesh, M.H.; Alikhani, H.A.; Khavazi, K.; Rahimian, H.A.; Miransari, M. Wheat (Triticum aestivum L.) growth enhancement by Azospirillum sp. under drought stress. World J. Microbiol. Biotechnol. 2011, 27, 197–205. [Google Scholar] [CrossRef]
- Moreno-Galván, A.E.; Cortés-Patiño, S.; Romero-Perdomo, F.; Uribe-Vélez, D.; Bashan, Y.; Bonilla, R.R. Proline accumulation and glutathione reductase activity induced by drought-tolerant rhizobacteria as potential mechanisms to alleviate drought stress in Guinea grass. Appl. Soil Ecol. 2020, 147, 103367. [Google Scholar] [CrossRef]
- Abbasi, S.; Sadeghi, A.; Safaie, N. Streptomyces alleviate drought stress in tomato plants and modulate the expression of transcription factors ERF1 and WRKY70 genes. Sci. Hortic. 2020, 265, 109206. [Google Scholar] [CrossRef]
- Yasmin, H.; Bano, A.; Wilson, N.L.; Nosheen, A.; Naz, R.; Hassan, M.N.; Ilyas, N.; Saleem, M.H.; Noureldeen, A.; Ahmad, P. Drought-tolerant Pseudomonas sp. showed differential expression of stress-responsive genes and induced drought tolerance in Arabidopsis thaliana. Physiol. Plant. 2022, 174, e13497. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, B.; Shahid, M.; Syed, A.; Rajput, V.D.; Elgorban, A.M.; Minkina, T.; Bahkali, A.H.; Lee, J. Drought tolerant Enterobactersp./Leclercia adecarboxylata secretes indole-3-acetic acid and other biomolecules and enhances the biological attributes of Vigna radiata (L.) R. Wilczek in water deficit conditions. Biology 2021, 10, 1149. [Google Scholar] [CrossRef] [PubMed]
- Kasim, W.A.; Osman, M.E.H.; Omar, M.N.; Salama, S. Enhancement of drought tolerance in Triticum aestivum L. seedlings using Azospirillum brasilense NO40 and Stenotrophomonas maltophilia B11. Bull. Natl. Res. Cent. 2021, 45, 95. [Google Scholar] [CrossRef]
- Sundara, B.; Natarajan, V.; Hari, K. Influence of phosphorus solubilizing bacteria on the changes in soil available phosphorus and sugarcane yields. Field Crops Res. 2002, 77, 43–49. [Google Scholar] [CrossRef]
- SundraRao, W.V.B; Sinha, M.K. Phosphate dissolving organisms in soil and rhizosphere. Indian J. Agric Sci. 1963, 33, 272–278. [Google Scholar]
- Tambekar, D.H.; Gulhane, S.R.; Somkuwar, D.O.; Ingle, K.B.; Kanchalwar, S.P. Potential Rhizobium and phosphate solubilizers as a biofertilizers from saline belt of Akola and Buldhana district, India. Res. J. Agric. Biol. Sci. 2009, 5, 578–582. [Google Scholar]
- Pantoja-Guerra, M.; Valero-Valero, N.; Ramírez, C.A. Total auxin level in the soil–plant system as a modulating factor for the effectiveness of PGPR inocula: A review. Chem. Biol. Technol. Agric. 2023, 10, 6. [Google Scholar] [CrossRef]
- Sgroy, V.; Cassán, F.; Masciarelli, O.; Del Papa, M.F.; Lagares, A.; Luna, V. Isolation and characterization of endophytic plant growth-promoting (PGPB) or stress homeostasis-regulating (PSHB) bacteria associated to the halophyte Prosopis strombulifera. Appl. Microbiol. Biotechnol. 2009, 85, 371–381. [Google Scholar] [CrossRef]
- Tao, G.; Tian, S.; Cai, M.; Xie, G. Phosphate solubilizing and mineralizing abilities of bacteria isolated from soils. Pedosphere. 2008, 18, 515–523. [Google Scholar] [CrossRef]
- Vazquez, P.; Holguin, G.; Puente, M.; Cortes, AE.; Bashan, Y. Phosphate solubilizing microorganisms associated with the rhizosphere of mangroves in a semi arid coastal lagoon. Biol Fertil Soils. 2000, 30, 460–468. [Google Scholar] [CrossRef]
- Valente Lima, J.; Tinôco, R.S.; Olivares, F.L.; Moraes, A.J.G.d.; Chia, G.S.; Silva, G.B.d. Hormonal imbalance triggered by rhizobacteria enhance nutrient use efficiency and biomass in oil palm. Sci. Hortic. 2020, 264, 109161. [Google Scholar] [CrossRef]
- Ahmed, A.; Hasnain, S. Auxin-producing Bacillus sp. : Auxin quantification and effect on the growth of Solanum tuberosum. Pure Appl. Chem. 2010, 82, 313–319. [Google Scholar]
- Mohammadi Alagoz, S.; Zahra, N.; Hajiaghaei Kamrani, M.; Asgari Lajayer, B.; Nobaharan, K.; Astatkie, T.; Siddique, K.H.M.; Farooq, M. Role of Root Hydraulics in Plant Drought Tolerance. J. Plant Growth Regul. 2022, 1–16. [Google Scholar] [CrossRef]
- Whitelaw, M.A. Growth promotion of plants inoculated with phosphate solubilizing fungi. Adv Agron. 2000, 69, 99–151. [Google Scholar]
- Xiao, C.Q.; Chi, R.A.; Huang, X.H.; Zhang, W.X. Optimization for rock phosphate solubilization by phosphate-solubilizing fungi isolated from phosphate mines. Ecol. Eng. 2008, 33, 187–193. [Google Scholar] [CrossRef]
- Cohen, A.C.; Bottini, R.; Piccoli, P.N. Azospirillum brasilense Sp 245 produces ABA in chemically-defined culture medium and increases ABA content in arabidopsis plants. Plant Growth Regulation 2008, 54, 97–103. [Google Scholar] [CrossRef]
- Hungria, M.; Campo, R.J.; Souza, E.M.; Pedrosa, F.O. Inoculation with selected strains of Azospirillum brasilense and A. lipoferum improves yields of maize and wheat in Brazil. Plant Soil 2010, 331, 413–425. [Google Scholar] [CrossRef]
- Salomon, M.V.; Bottini, R.; de Souza Filho, G.A.; Cohen, A.C.; Moreno, D.; Gil, M.; Piccoli, P. Bacteria isolated from roots and rhizosphere of Vitis vinifera retard water losses, induce abscisic acid accumulation and synthesis of defense-related terpenes in in vitro cultured grapevine. Physiol. Plant. 2014, 151, 359–374. [Google Scholar] [CrossRef]
- Yahya, A.; Azawi, S.K.A. Occurrence of phosphate solubilizing bacteria in some Iranian soils. Plant Soil. 1998, 117, 135–141. [Google Scholar] [CrossRef]
- Zaddy, E.; Perevolosky, A. Enhancement of growth and establishment of oak seedling by inoculation with Azospirillum brasilense. Forest Eco Manage. 1995, 72, 81–83. [Google Scholar] [CrossRef]
- Zaddy, E.; Perevolosky, A.; Okon, Y. Promotion of plant growth by inoculation with aggregated and single cell suspension by Azospirillum brasilense. Soil Biol. Biochem. 1993, 25, 819–823. [Google Scholar] [CrossRef]
- Bhat, M.I.; Rashid, A.; Faisul-ur-Rasool; Mahdi, S.S.; Haq, S.A.; Raies, A.B. Effect of Rhizobium and Vesicular arbuscular mycorrhizae fungi on green Gram (Vigna radiata L.Wilczek) under temperate conditions. Res. J. Agric. Sci. 2010, 1, 113–118. [Google Scholar]
- Bisleski, R.L. Phosphate transport and phosphate availability, Ann. Rev.Pl.Physiol. 1973, 24, 225–252. [Google Scholar] [CrossRef]
- Brundrett, M.C.; Abbott, L.K. Mycorrhizal fungus propagules in the jarrh forest. I. Spatial variability in inoculam levels. New Phytol. 1995, 131, 461–469. [Google Scholar] [CrossRef]
- Champawat, R.S. ; Pathak,V. N. Effect of Vesicular-arbuscular mycorrhizal fungi on growth and nutrition uptake of pearl millet, Indian J.Mycol.Pl.Pathol. 1993, 23, 30–34. [Google Scholar]
- Naveed, M.; Mitter, B.; Reichenauer, T.G.; Wieczorek, K.; Sessitsch, A. Increased drought stress resilience of maize through endophytic colonization by Burkholderia phytofirmans PsJN and Enterobacter sp. FD17. Environ. Exp. Bot. 2014, 97, 30–39. [Google Scholar] [CrossRef]
- Aguiar, N.O.; Medici, L.O.; Olivares, F.L.; Dobbss, L.B.; Torres-Netto, A.; Silva, S.F.; Novotny, E.H.; Canellas, L.P. Metabolic profile and antioxidant responses during drought stress recovery in sugarcane treated with humic acids and endophytic diazotrophic bacteria. Ann. Appl. Biol. 2016, 168, 203–213. [Google Scholar] [CrossRef]
- Honma, M.; Shimomura, T. Metabolism of 1-aminocyclopropane-1-carboxylic acid. Agric. Biol. Chem. 1978, 42, 1825–1831. [Google Scholar]
- Glick, B.R.; Penrose, D.M.; Li, J. A model for the lowering of plant ethylene concentrations by plant growth-promoting bacteria. J. Theor. Biol. 1998, 190, 63–68. [Google Scholar] [CrossRef]
- Duchense, L.C.; Peterson, R.L.; Ellis, B.E. The future of ectomycorrhizal fungi as biological control agents. Phytoprotection 1989, 70, 51–57. [Google Scholar]
- Dumas, G.E.; Guillaume, P.; Tahiri, A.A.; Gianinazzi-Pearson, V.; Gianinazzi, S. Changes in polypeptide patterns in tobacco roots by Glomus species. Mycorrhiza 1994, 4, 215–221. [Google Scholar]
- Sziderics, A.H.; Rasche, F.; Trognitz, F.; Sessitsch, A.; Wilhelm, E. Bacterial endophytes contribute to abiotic stress adaptation in pepper plants (Capsicum annuum L. ). Can. J. Microbiol. 2007, 53, 1195–1202. [Google Scholar] [CrossRef]
- Vardharajula, S.; Zulfikar Ali, S.; Grover, M.; Reddy, G.; Bandi, V. Drought-tolerant plant growth promoting Bacillus spp.: Effect on growth, osmolytes, and antioxidant status of maize under drought stress. J. Plant Interact. 2011, 6, 1–14. [Google Scholar] [CrossRef]
- Ghosh, D.; Gupta, A.; Mohapatra, S. A comparative analysis of exopolysaccharide and phytohormone secretions by four drought- tolerant rhizobacterial strains and their impact on osmotic-stress mitigation in Arabidopsis thaliana. World J. Microbiol. Biotechnol. 2019, 35, 90. [Google Scholar] [CrossRef] [PubMed]
- Fa Yuan, W.; Zhao Yong, S. Biodiversity of Arbuscular Mycorrhizal Fungi in China: a Review. Advances in Environmental Biology 2008, 2, 31–39. [Google Scholar]
- Ojuederie, O.B.; Olanrewaju, O.S.; Babalola, O.O. Plant growth promoting rhizobacterial mitigation of drought stress in crop plants: Implications for sustainable agriculture. Agronomy 2019, 9, 712. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A. Exopolysaccharide producing rhizobacteria and their impact on growth and drought tolerance of wheat grown under rainfed conditions. PLoS ONE 2019, 14, e0222302. [Google Scholar] [CrossRef]
- Hart, M.M.; Reader, R.J. Host plant benefit from association with arbuscular mycorrhizal fungi: variation due to differences in size of mycelium, Biol. Ferti.Soils 2002, 36, 357–366. [Google Scholar] [CrossRef]
- Brilli, F.; Loreto, F.; Baccelli, I. Exploiting plant volatile organic compounds (VOCs) in agriculture to improve sustainable defense strategies and productivity of crops. Front. Plant Sci. 2019, 10, 264. [Google Scholar] [CrossRef]
- Ryu, C.-M.; Farag, M.A.; Hu, C.-H.; Reddy, M.S.; Wei, H.-X.; Paré, P.W.; Kloepper, J.W. Bacterial volatiles promote growth in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 4927–4932. [Google Scholar] [CrossRef] [PubMed]
- Blom, D.; Fabbri, C.; Connor, E.C.; Schiestl, F.P.; Klauser, D.R.; Boller, T.; Eberl, L.; Weisskopf, L. Production of plant growth modulating volatiles is widespread among rhizosphere bacteria and strongly depends on culture conditions. Environ. Microbiol. 2011, 13, 3047–3058. [Google Scholar] [CrossRef] [PubMed]
- Subrahmanyam, G.; Kumar, A.; Sandilya, S.P.; Chutia, M.; Yadav, A.N. Diversity, plant growth promoting attributes, and agricultural applications of rhizospheric microbes. Plant Microbiomes Sustain. Agric. 2020, 25, 1–52. [Google Scholar]
- Jeffries, A. Use of mycorrhiza in agriculture. Cri. Rev. Biotech. 1987, 5, 319–357. [Google Scholar] [CrossRef]
- Khan, I.A.; Ayub, N.; Mirza, S.N.; Nizami, S.M.; Azam, M. Synergistic effect of dual inoculation (Vesicualr- arbuscular mycorrhizae) on the growth and nutrients uptake of Medicago sativa, Pak. J.Bot. 2008, 40, 939–945. [Google Scholar]
- Cho, S.M.; Kang, B.R.; Han, S.H.; Anderson, A.J.; Park, J.-Y.; Lee, Y.-H.; Cho, B.H.; Yang, K.-Y.; Ryu, C.-M.; Kim, Y.C. 2R, 3R-butanediol, a bacterial volatile produced by Pseudomonas chlororaphis O6, is involved in induction of systemic tolerance to drought in Arabidopsis thaliana. Mol. Plant-Microbe Interact. 2008, 21, 1067–1075. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.; Wang, X.; Saleem, M.H.; Sumaira; Hafeez, A.; Afridi, M.S.; Khan, S.; Zaib Un, N.; Ullah, I.; Amaral Júnior, A.T.D.; et al. PGPR-Mediated Salt Tolerance in Maize by Modulating Plant Physiology, Antioxidant Defense, Compatible Solutes Accumulation and Bio-Surfactant Producing Genes. Plants 2022, 11, 345. [Google Scholar] [CrossRef]
- Nivetha, N.; Lavanya, A.K.; Vikram, K.V.; Asha, A.D.; Sruthi, K.S.; Bandeppa, S.; Annapurna, K.; Paul, S. PGPR-Mediated Regulation of Antioxidants: Prospects for Abiotic Stress Management in Plants. In Antioxidants in Plant-Microbe Interaction; Singh, H.B., Vaishnav, A., Sayyed, R.Z., Eds.; Springer: Singapore, 2021; pp. 471–497. [Google Scholar]
- El-Komy, M.; Hesham, A. Coimmobilization of Azospirillum lipoferum and Bacillus megaterium for successful phosphorus and nitrogen nutrition of wheat plants. Food Technol. Biotechnol. 2004, 43, 19–27. [Google Scholar]
- El-Komy, H.M.; Hamdia, M.A.; El-Baki, G.K.A. 2003. Nitrate reductase in wheat plants grown under water stress and inoculated with Azospirillum spp. Biol. Plant. 2003, 46, 281–287. [Google Scholar] [CrossRef]
- Zhang, W.; Xie, Z.; Zhang, X.; Lang, D.; Zhang, X. Growth-promoting bacteria alleviates drought stress of G. uralensis through improving photosynthesis characteristics and water status. J. Plant Interact. 2019, 14, 580–589. [Google Scholar] [CrossRef]
- Fulchieri, M.; Frioni, L. Azospirillum inoculation on maize (Zea mays): Effect of yield in a field experiment in Central Argentina. Soil Biol. Biochem. 1994, 26, 921–924. [Google Scholar] [CrossRef]
- Patil, P.L.; Medhane, N.S. Seed inoculation studies in gram (Cicer arietinum L.) with different strains of Rhizobium sp. Plant Soil 1974, 40, 221–223. [Google Scholar] [CrossRef]
- Khan, A.L.; Halo, B.A.; Elyassi, A.; Ali, S.; Al-Hosni, K.; Hussain, J.; Al-Harrasi, A.; Lee, I.-J. Indole acetic acid and ACC deaminase from endophytic bacteria improves the growth of Solanum lycopersicum. Electron. J. Biotechnol. 2016, 21, 58–64. [Google Scholar] [CrossRef]
- Mc A Fee, B.J.; Fortin, J.A. Comparative effects of the soil microflora on ectomycorrhizal inoculation of conifer seedling. New Phytol. 1986, 108, 108–443. [Google Scholar]
- de Souza, R.; Schoenfeld, R.; Passaglia, L.M.P. Bacterial inoculants for rice: Effects on nutrient uptake and growth promotion. Arch. Agron. Soil Sci. 2016, 62, 561–569. [Google Scholar] [CrossRef]
- Baldotto, L.E.B.; Baldotto, M.A.; Canellas, L.P.; Bressan-Smith, R.; Olivares, F.L. Growth promotion of pineapple’Vitória’by humic acids and Burkholderia spp. during acclimatization. Revista Brasileira de Ciência do Solo 2010, 34, 1593–1600. [Google Scholar] [CrossRef]
- Garg, S.; Bhatnagar, A.; Kalla, A.; Narula, N. In vitro nitrogen fixation, phosphate solubilization, survival and nutrient release by Azotobacter strains in an aquatic system. Biores. Technol. 2001, 80, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Geurts, R.; Bisseling, T. Rhizobium nod factor perception and signalling. Plant Cell 2002, 14, S239–S249. [Google Scholar] [CrossRef]
- Khosro, M.; Yousef, S. Bacterial biofertilizers for sustainable crop production: A review. ARPN J. Agric. Biol. Sci. 2012, 7, 307–316. [Google Scholar]
- Kavamura, V.N.; Santos, S.N.; Silva, J.L.D.; Parma, M.M.; Ávila, L.A.; Visconti, A.; Zucchi, T.D.; Taketani, R.G.; Andreote, F.D.; Melo, I.S.d. Screening of Brazilian cacti rhizobacteria for plant growth promotion under drought. Microbiol. Res. 2013, 168, 183–191. [Google Scholar] [CrossRef]
- Kang, S.-M.; Shahzad, R.; Bilal, S.; Khan, A.L.; Park, Y.-G.; Lee, K.-E.; Asaf, S.; Khan, M.A.; Lee, I.-J. Indole-3-acetic-acid and ACC deaminase producing Leclercia adecarboxylata MO1 improves Solanum lycopersicum L. growth and salinity stress tolerance by endogenous secondary metabolites regulation. BMC Microbiol. 2019, 19, 80. [Google Scholar] [CrossRef] [PubMed]
- Narayanasamy, S.; Thankappan, S.; Kumaravel, S.; Ragupathi, S.; Uthandi, S. Complete genome sequence analysis of a plant growth-promoting phylloplane Bacillus altitudinis FD48 offers mechanistic insights into priming drought stress tolerance in rice. Genomics 2023, 115, 110550. [Google Scholar] [CrossRef] [PubMed]
- West, T.P.; Strohfus, B. Short Communication: Effect of carbon source on exopolysaccharide production by Sphingomonas paucimobilis ATCC 31461. Microbiol. Res. 1999, 153, 327–329. [Google Scholar] [CrossRef]
Figure 1.
Plant growth is facilitated by PGPB, which produces siderophores, increases iron availability, and produces hormones like auxins, gibberellins, and cytokinin which influence plant hormone balance. By utilizing the nitrogenase enzyme complex and solubilizing inorganic phosphate in the soil, biological nitrogen fixation (BNF) occurs, as well as siderophores are produced. Plant pathogens and nematodes are repelled by PGPB by occupying niches and producing substances that repel them.
Figure 1.
Plant growth is facilitated by PGPB, which produces siderophores, increases iron availability, and produces hormones like auxins, gibberellins, and cytokinin which influence plant hormone balance. By utilizing the nitrogenase enzyme complex and solubilizing inorganic phosphate in the soil, biological nitrogen fixation (BNF) occurs, as well as siderophores are produced. Plant pathogens and nematodes are repelled by PGPB by occupying niches and producing substances that repel them.
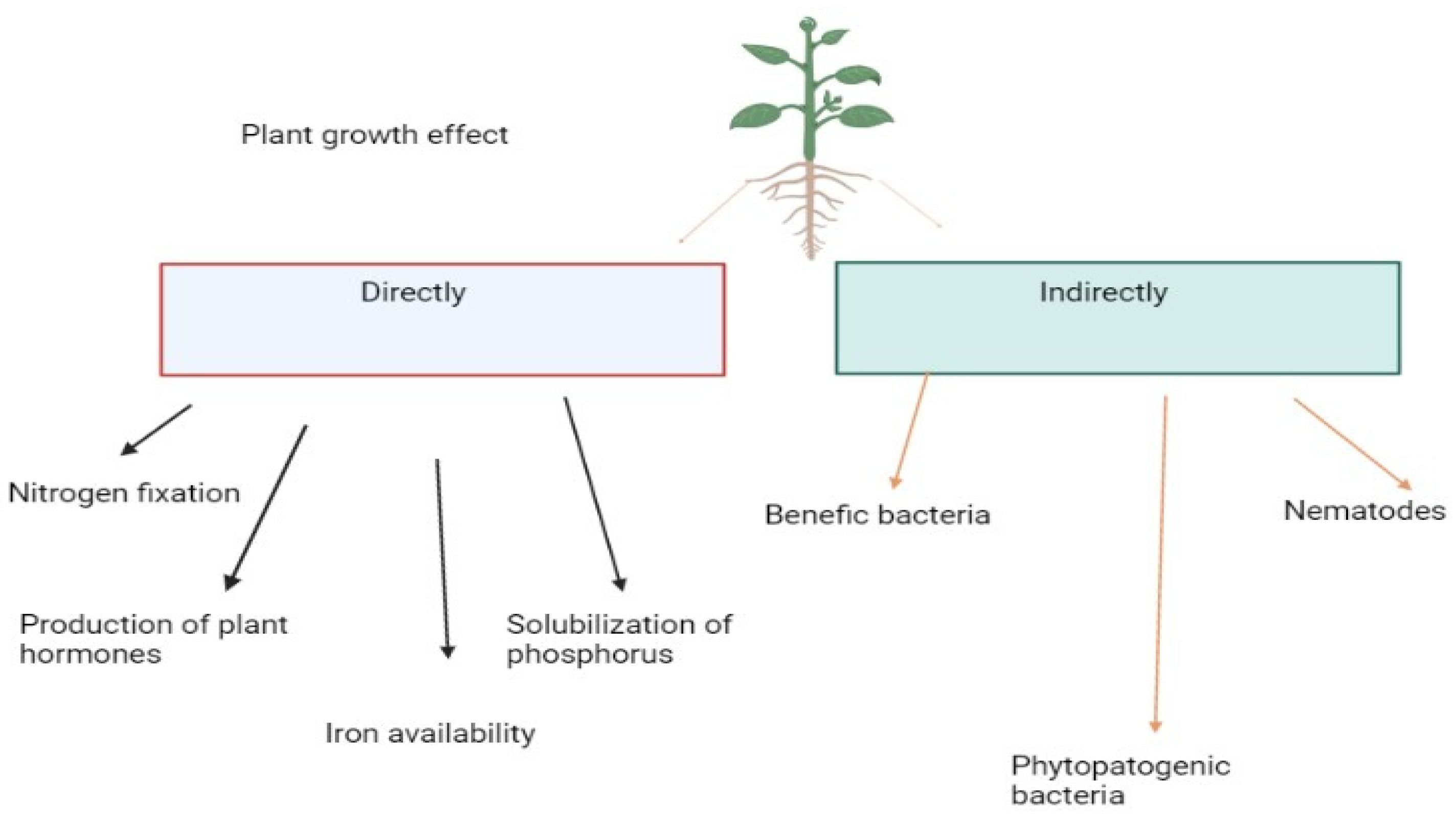
Figure 2.
Microorganisms and plants produce metabolites that act as signaling molecules and substrates when they interact. Besides fixing nitrogen and solubilizing nutrients (phosphate and iron), microorganisms also produce hormones, which stimulate plant growth. A dashed line indicates a positive relationship between plants and bacteria. Abbreviations: 1-aminocyclopropane-1 carboxylate deaminase; BNF, biological nitrogen fixation.
Figure 2.
Microorganisms and plants produce metabolites that act as signaling molecules and substrates when they interact. Besides fixing nitrogen and solubilizing nutrients (phosphate and iron), microorganisms also produce hormones, which stimulate plant growth. A dashed line indicates a positive relationship between plants and bacteria. Abbreviations: 1-aminocyclopropane-1 carboxylate deaminase; BNF, biological nitrogen fixation.
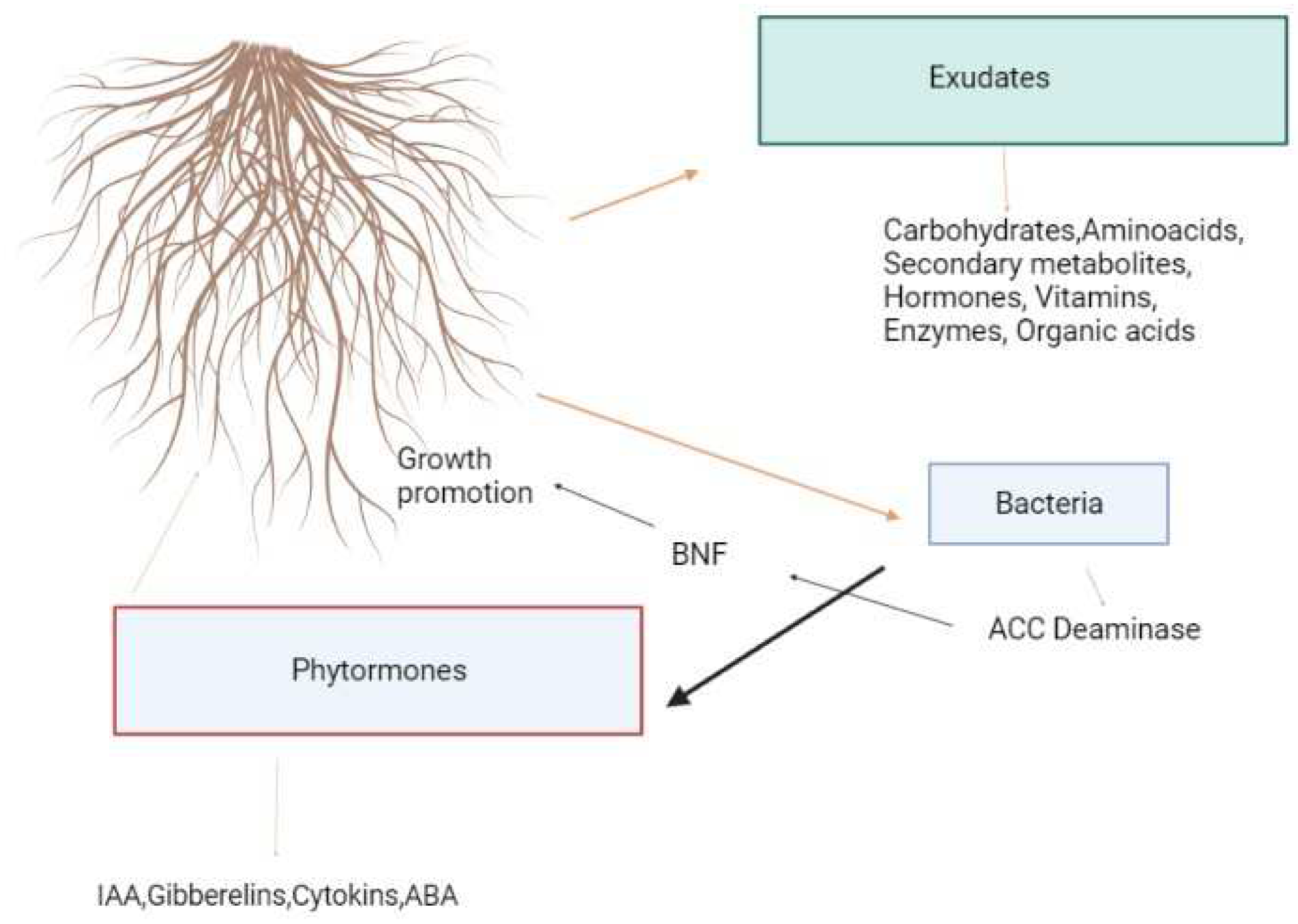
Figure 3.
Inoculation of different strains of Azospirillum sp., Bacillus sp. and Streptomyces sp., in the growth promotion of water-stressed lettuce plants.
Figure 3.
Inoculation of different strains of Azospirillum sp., Bacillus sp. and Streptomyces sp., in the growth promotion of water-stressed lettuce plants.


Table 1.
Bacterial genera and species that are drought resistant.
| Microorganisms | Plant | Method of action | Cit. |
|---|---|---|---|
| Azospirillum sp. | Wheat | Auxin and N concentrations are highest | [60] |
| Bacillus sp. | Grass | Antioxidant system response and early proline accumulation | [61] |
| Streptomyces sp. | Tomato | Increasing the content of different sugars | [62] |
| Pseudomonas sp. | Arabidopsis | Exopolysaccharide, gibberellic acid, abscisic acid, and indole acetic acid deaminase activity are higher | [63] |
| Enterobacter sp. | Bean | Increase the levels of proline, malondialdehyde, and antioxidant enzymes | [64] |
| Azospirillum brasilense | Wheat | A decrease in H2O2 accumulation and a decrease in the production of proline and catalase as well as peroxidase activity | [65] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



