
Submitted:
27 June 2023
Posted:
28 June 2023
You are already at the latest version
Abstract
Centropomus undecimalis (common snook, sergeant fish or robalo) and Centropomus parallelus (fat snook, smallscale fat snook, little snook or chucumite) have tropical and subtropical wide distribution from southern Florida (Gulf of Mexico) to southern Brazil (Rio Grande do Sul) waters, and due the value of their flesh, these species have been explored with predatory fishing, generating a deficiency for their conservation. Adequate conservation of this species depends on population genetic parameters. Thus, in this study, we used microsatellite markers to investigate the genetic variability and the genetic characterization of C. undecimalis and C. parallelus populations, identifying if the citations species are geographically connected, using 10 loci of microsatellites, bringing strategies of management, sustainability, and conservation for species citations, and providing genetic subsidies needed to be used in breeding programs for snooks. Microsatellite molecular genetic markers were efficient in the study of the genetic variability of the samples of C. parallelus and C. undecimalis, resulting in data on the genetic structuring of these populations, thus aiding in their management and conservation. The data presented may provide subsidies for future use in programs aimed at animal breeding of sea bass, establishing tools for use in fish farming. In addition, despite being two species that are not on the endangered list, we can observe that there are genetic differences between the studied populations of the same species.
Keywords:
Microsatellites
; Centropomus parallelus
; Centropomus undecimalis
; Centropomidae
; genetic polymorphism
1. Introduction
The Brazil has a coastal area with more than 8,000 kilometers, with a large volume of fresh water, is one of the largest hydrographic basins in the world, with a great marine biodiversity, due this scenario the Brazilian marine fish farming is in constant expansion and economic growth [1,2]. In 2012, the world production of fish was 158 million tons, moving US $ 600 billion / year - US $ 136 billion/year in exports. While one hectare of land can produce 0.12 tons/year of meat, the same area of water can produce 100 to 320 tons/year of fish. Making fish farming a more sustainable activity. With this potential, Brazil intends to increase aquaculture production [2].
Among the various marine species with commercial interest and potential to be exploited, the species of the genus Centropomus spp stand out. Centropomus spp is the sole genus of the predominantly marine fish family Centropomidae (Teleostei) and have 12 species, commonly known as snooks (http://researcharchive.calacademy.org/research/ichthyology/catalog/SpeciesByFamily.asp). Species of this genus can be found in both the eastern Pacific and western Atlantic [3,4], which was separated around 1.6 to 3 Mya after the Panama Isthmus closure [5]. Among them, 6 of them inhabit the Brazilian coasts and we highlighted Centropomus undecimalis (common snook, sergeant fish or robalo) and Centropomus parallelus (fat snook, smallscale fat snook, little snook or chucumite) as the of highest interest for economic and sport fishing, which have wide distribution from southern Florida (Gulf of Mexico) to southern Brazil (Rio Grande do Sul).
These species are carnivores feeding mainly on fish, shrimp and crustaceans, however, there is a certain seasonal variation of feeding because they seem to be quite opportunistic predating depending on the availability of the environment in which they are [6,7], in view of the nobility of its meat, with excellent organoleptic quality, reaching high prices in the domestic and foreign markets, these attributes make these species to be exploited with sportive, artisanal and professional fishing and lack of data for catch reports generate a deficiency for their conservation [8-12] . C. undecimalis is one of the largest members of the genus (140 cm and 22 kg) [13] and has been included in several aquaculture programs in coastal areas of the Atlantic Ocean [4, 14, 15]. C. parallelus may reach up to 75 cm and weigh up to 4 kg with females being larger than males of the same age [16]. Snook species are normally commercialized with size is from 0.5 to 1.0 kg [16].
For the advancement of efficiency in the production and profitability of aquaculture, a diversified range of genetic resources is necessary. Research on genomics, genetics and breeding in aquaculture has the ultimate objective of improving production efficiency, sustainability, product quality, food safety, consumer protection and improving profitability, all with the goal of benefiting consumers [17]. Thus, knowledge of genetic diversity of species is a tool to ensure the sustainability of future populations within their geographic range, despite habitat alterations, as well as captive breeding [18-20].
Population genetics are linked to allelic distribution and change in natural selection, mutation, migration, gene flow, geographic distribution, and genetic drift [21]. Loss of genetic variability leads to problems in fish adaptation and survival, genetic knowledge of the population structure of common and fat snook leads to gains in fish production and conservation. The genetic diversity between the natural reserves and/or between the different fish farming systems is fundamental for the correct management of the species [22]. There are several tools for genetic analysis today, mainly the analysis of the genome or regions in the genome and among them, microsatellites stand out. Microsatellite markers, also known as Short Tandem Repeats (STR), are 90–350 bp long non-transcriptional variable number of tandem repeat sequences at certain points of the genome. They are co-dominant and generally high polymorphic and are widely used for genetic diversity analysis and quantitative trait locus [23- 26].
Thus, in this study, we used microsatellite markers to investigate the genetic variability and the genetic characterization of C. undecimalis and C. parallelus populations, identifying if the citations species are geographically connected, using 10 loci of microsatellites, bringing strategies of management, sustainability and conservation for species citations, and providing genetic subsidies needed to be used in breeding programs for snooks.
2. Materials and Methods
2.1. Sample Collection and DNA Extraction
Samples of the C. undecimalis and C. parallelus specimens were collected through artisanal fisheries in the region of Santa Catarina to São Paulo Brazilian federated states (Table 1); 42 C. parallelus samples were collected, 29 from Balneário de Penha – SC (pSC) and 13 from Guarujá – SP (pSP). Sixty six C. undecimalis samples were collected, 18 from Balneário de Penha – SC (uSC), 8 from Guarujá – SP (uSP), 40 captive from CERES (College of Southern Region)/UDESC (Santa Catarina State University), and UFSC (Federal University of Santa Catarina) - LAPMAR (Marine Fish Laboratory) (uCap).
Biological tissues of the animals were collected from the dorsal or caudal fin or from muscle tissue to further DNA extraction and stored in 1.5 ml microtubes with ethanol (100 %) and kept in a freezer at -20˚C in the Laboratory of Genetics and Molecular Biology of CERES- UDESC- Laguna, SC, Brazil.
2.2. DNA Extraction
DNA extraction was performed according to the phenol/chloroform protocol described by Sambrook et al. [27]. Approximately 30 mg of tissue was transferred to 1.5 ml eppendorf tubes containing 500 μl of digestion solution (50 mM Tris-HCl, 50 mM EDTA, 100 mM NaCl, 1% SDS pH 8.0). Then 10μl of SDS and 20μl of Proteinase K (20mg/ml) were added to the tube. These tubes were shaken for 15 seconds, and the solution was then brought to the thermo block at 55 °C for 2 hours. After this time a volume of 500μl of phenol/chloroform/isoamyl alcohol was placed. Thereafter the material was centrifuged at 12,000 rpm for 5 minutes. The supernatant was transferred to a new tube and then added with 500 μl of ice-cold absolute ethanol and incubated overnight at -20 °C for precipitation of the DNA. The sample was centrifuged again for 5 minutes at 12,000 rpm, discarded the supernatant, and the DNA pellet was washed with 70% ethanol and then centrifuged again at 12,000 rpm for 15 minutes. The supernatant was discarded and the DNA was allowed to dry in the oven at 50 °C for 15 minutes. At the end the DNA was resuspended in 60 μl autoclaved distilled water, then stored in the freezer at -20 ° C. Quantification and verification of the amount of DNA were done on 1% agarose gel electrophoresis with quantification marker.
2.3. DNA Amplification by PCR
PCR reactions were performed to amplify the locus using 10 STR Cun01, Cun05B, Cun08, Cun09, Cun10A, Cun11, Cun14, Cun16, Cun21B, Cun22 as described by Seyoum et al. [28]. The PCR reaction mixtures were made with the following reagent and concentrations to a final volume of 25 μl: 50 ng DNA, 2.5 nl 10X buffer (100mM Tris-HCl, pH 8.3, 500 mM KCl ), 1 μl (15 mM) MgCl 2, 2 μl of the dNTPs mixture (0.2 mM dATP, dCTP, dGTP and dTTP) 1 μM of each primer (20 μg / μl), 1.25 μl of the enzyme Taq Polymerase (Invitrogen) and H2O to complete the final volume. The reactions were programmed in a thermal cycler with initial denaturation at 94 °C for 5 minutes, followed by 35 cycles with denaturation of 94 °C for 45 seconds, annealing with temperature 58 - 60 °C (according to the indicator pair) by 90 seconds at 72 °C. The final extension occurred for 7 minutes at 72 °C.
2.4. Data Analysis
Sequences were submitted to the Methods and Statistics program in GenAlEx 6.5 [29], where parameters of allele frequency, observed and expected heterozygosity, Chi-square and Hardy-Weinberg equilibrium were calculated.
R environment (R Development Core Team 2018) packages adegenet [30] and ggplot2 [31] was used to have a visual assessment of between-population differentiation, we performed a principal component analysis (PCA)
Heterozygosity observed (HO) and expected (HE), as well allelic richness (Ar), were used to estimate the diversity of each species in each sampling locality. We also estimated inbreeding coefficients (FIS) for the populations within each sampling locality using the diveRsity [32] and PopGenKit [33] packages. Confidence intervals were obtained with 10,000 bootstrap replicates.
3. Results and Discussion
The analyzed microsatellites showed high polymorphism of the ten loci used to verify the genetic variability of C. udecimalis and C. paralellus populations. Microsatellite loci have been evaluated in many species of fish, studies with these markers are being carried out, since the number of alleles per loco is quite variable. Microsatellites are abundant in the genomes of higher organisms [34]. Thousands of such microsatellite markers have been identified in humans [34, 35]. For C. undecimalis, microsatellites were described by Seyoum et al. [28] and a study was performed by Hernández-Vidal et al. [36]. No study with microsatellite markers has been performed in C. parallelus so far. The alleles length used in this study from C. undecimalis vary from 80 bp for locus Cun21B to 293 bp for locus Cun09. In the study by Seyoum et al. [28] the variation was 100 bp for locus Cun21B to 243 bp for locus Cun14.
Numbers of alleles vary in the 3 populations of C. udecimalis. In in uSC, the locus Cun08 and Cun11 showed the highest number of all alleles (22), while loco Cun05B showed the lowest number of alleles (7). In uSP, the locus Cun12 showed the highest number of all alleles (12), while loco Cun05B showed the lowest number of alleles (3). In uCap, the locus Cun01 showed the highest number of all alleles (32), while loco Cun05B showed the lowest number of alleles (13). In C. paralellus in pSC, the locus Cun08 showed the highest number of all alleles (24), while loco Cun05B showed the lowest number of alleles (4). In pSP, the locus Cun14 showed the highest number of all alleles (15), while loco Cun16 showed the lowest number of alleles (6).
Although the loci with the highest number of alleles varied, the locus that showed the lowest number of alleles was Cun05B in all populations of C. paralellus and C. udecimalis ranging from 3 to 13.
A high number of different alleles per locus is a good indicator of high polymorphism. However, this is often strongly related to the sample size of each group studied [37, 38], for example, in the case of uCap, which showed the largest number of alleles at all loci. Thus, there are other parameters to be considered in a population genetics analysis. Ho and He are parameters that are quite reliable, and the locus Cun05B also showed the lowest Ho in general. In the uSP population, this same locus showed a Ho of 0.5, while all other loci showed a Ho of 1.
In genetics of populations the presence of null alleles has been commonly reported and may interfere with estimates of genetic variability and genetic distances [39]. Due to the high He index observed for all loci, it was estimated there is no null allele in the locus analyzed in this study of the populations of C. undecimalis and C. paralellus. However, null allele was observed in C. undecimalis in the locus Cun5B, Cun16 and Cun21B in uSP; in the locus Cun14, Cun21B and Cun22 in uSC; and in Cun10A, Cun11, Cun14 and Cun22 in uCap; while in C. paralellus in the locus Cun10A, Cun11 and Cun16 in pSC; and in 14, 21B and 22 in pSP.
In the present study, genotypic and genotypic differentiation values, based on allele and genotype frequencies observed, indicated a possible genetic differentiation in the analyzed populations. Population differentiation studies in fish have been carried out to identify and manage different genetic pools. Population differentiation can occur in different forms (from genetically distinct populations to panmitic populations) and in this way, the basic conservation unit would be represented by a group genetically homogeneous individuals, such as the existence of different local populations [40].
Regarding the populations of C. undecimalis, in fSP population the heterozygosity observed (HO) ranged from 1 to 0.5 with a mean of 0.95 whereas the heterozygosity expected (HE) ranged from 0.8 to 0.91 with an average of 0.82. In fSC population the HO ranged from 1 to 0.39 with an average of 0.89 whereas the HE ranged from 0.9 to 0.94 with an average of 0.89. In fCap population the HO ranged from 1 to 0.68 with an average of 0.93 whereas the HE ranged from 0.95 to 0.85 with an average of 0.92.
Regarding the populations of C. paralellus, in pSC population the HO ranged from 1 to 0.55 with an average of 0.86 while the HE ranged from 0.9 to 0.94 with an average of 0.89. In pSP population the HO ranged from 1 to 0.77 with an average of 0.95 whereas the HE ranged from 0, 9 to 0.85 with a mean of 0.84.
FIS (coefficient of inbreeding) for those who did not present negative values indicating that there is no excess of heterozygotes in these populations. FIS, varied among all locus of the populations and the two species (Table 2 and Table 3). The distance and the genetic identity among the C. undecimalis populations was evidenced by the FST (fixation index) value of 0.0376 with an IC of (0.0250-0.0532), which according to Wright [41] are indications of low genetic differentiation. PCA data showed that there is a genetic distinction between populations, especially between pSC and pSP populations of C. paralellus, and in uSC and uSP of C. undecimalis, however uCap is more like the uSP population than to uSC (Figure).
The locus (Cun01, Cun05B, Cun08, Cun09, Cun10A, Cun11, Cun14, Cun16, Cun21B, and Cun22) did not exhibit Hardy-Weinberg equilibrium according to populations and species [42]. There was no linkage disequilibrium between the locus, suggest that all are genetically independent.
Its capture is associated with migratory movements to freshwater ecosystems and, also, to spawning on the coast. The environmental differences found along the coast play an ecological and evolutionary role, favoring genetic differentiation. So far, there has been no study comparing the species and populations of C. undecimalis and C. parallelus between the coast of the Brazilians federated states of São Paulo and Santa Catarina [43- 45].
Figure 1.
and second (PC2) principal components based on the analysis of the individual C. undecimalis (A) and C. parallelus (B) using 10 microsatellites loci. The ellipses, shown in different colours, represent the analysed individuals in two different analyses.
Figure 1.
and second (PC2) principal components based on the analysis of the individual C. undecimalis (A) and C. parallelus (B) using 10 microsatellites loci. The ellipses, shown in different colours, represent the analysed individuals in two different analyses.
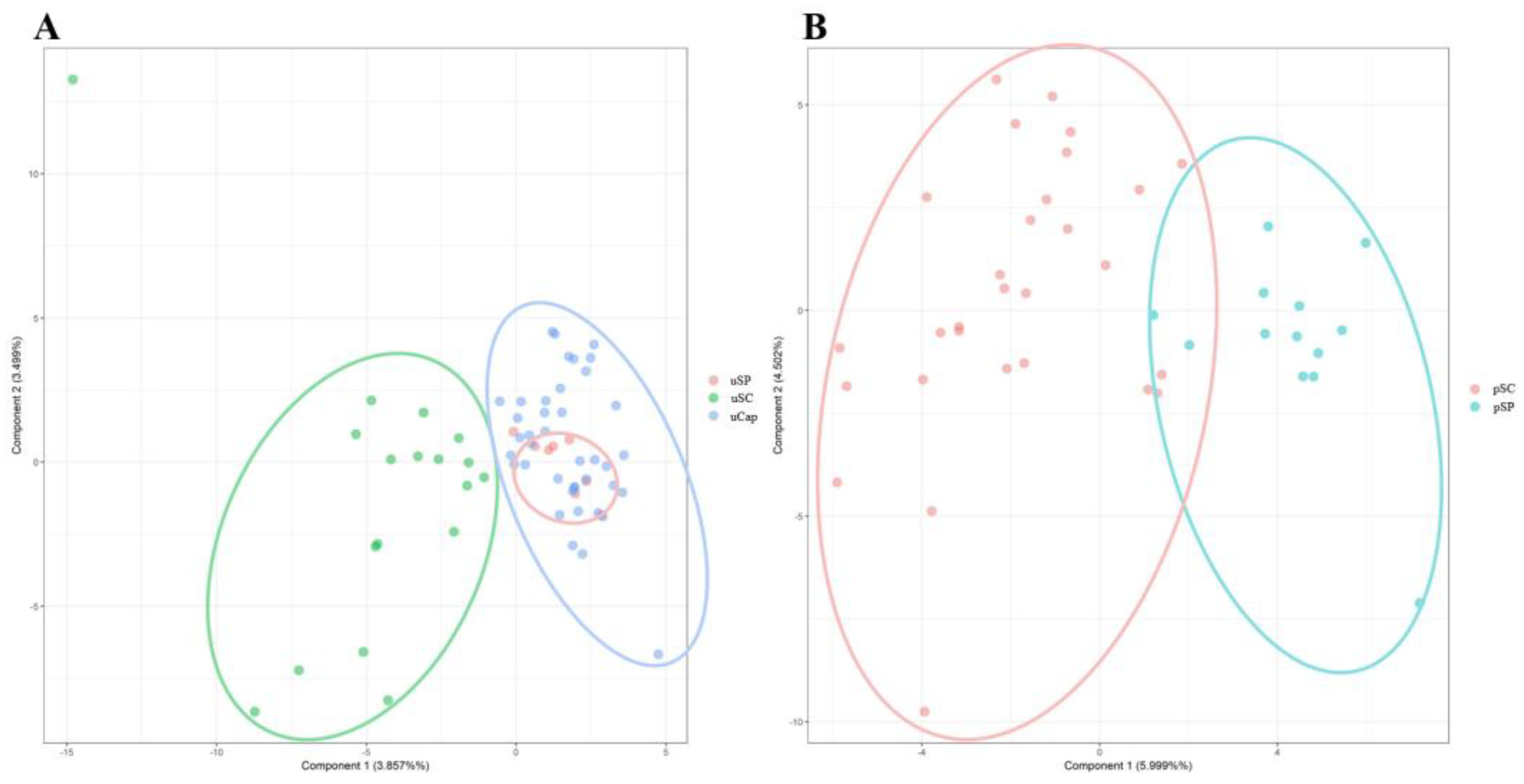
Although the scientific literature lacks sources about these two species, especially regarding their genetics, some considerations can be made. In summary, it was demonstrated by our data that these populations are found with high genetic variability, including the recently domesticated population of C. parallelus. The populations showed distinct allelic frequencies of some alleles, which demonstrates that they are consolidated populations. The Figure demonstrates that there are some points of convergence between populations of C. parallelus, which may indicate some reproductive contact. In addition, the same figure demonstrates that the domesticated population of C. undecimalis is more related to the population of the state of São Paulo than to Santa Catarina.
5. Conclusions
Microsatellite molecular genetic markers were efficient in the study of the genetic variability of the samples of C. parallelus and C. undecimalis, resulting in data on the genetic structuring of these populations, thus aiding in their management and conservation. In this way, ten polymorphic microsatellite loci obtained in C. parallelus and C. undecimalis, with the possibility of being used in the heterologous amplification of the other. Despite being two species that are not on the endangered list, we can observe that there are genetic differences between the studied populations of the same species. Centropomidae species, all important both from the economic point of view and from the sport fishing. The data presented may provide subsidies for future use in programs aimed at animal breeding of sea bass, establishing tools for use in fish farming. As far as we know, this is the first study genetically characterizing populations of these two species in Brazil, which is their main habitat.
6. Patents
Author Contributions
Conceptualization, M.E.H and C.A.V.L.R.C.; methodology, M.E.H.; validation, M.E.H.; formal analysis, M.E.H. and M.M.B.; investigation, X.X.; resources, C.A.V.L.R.C.; data curation, M.M.B.; writing—original draft preparation, M.E.H.; writing—review and editing, M.E.H., M.M.B. and C.A.V.L.R.C.; supervision, C.A.V.L.R.C.; project administration, M.E.H.;. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), grant number #2014/03062-0, and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), grant number #158284/2013-5.
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Ethics Committee of Comissão de Ética no Uso de Animais (CETEA) of Santa Catarina State University (protocol code 1.10.12 and date of approval at October 1st, 2012)
Acknowledgments
We would like to thank Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), Santa Catarina State University and University of São Paulo for the support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- FAO, Food and Agriculture Organization. The State of World Fisheries and Aquaculture; FAO: Rome, 2009. [Google Scholar]
- Brazil, Ministry of Agriculture. Aquicultura tem potencial para crescer mais de 80% em cinco anos. 2020. Available online: https://www.gov.br/casacivil/pt-br/assuntos/noticias/2015/junho/aquicultura-tem-potencial-para-crescer-mais-de-80-em-cinco-anos (accessed on 27 April 2020).
- Rivas, L.R. ; Systematic review of the perceived fishes of the Centropomus genus. Copeia 1986, 1, 579–611. [Google Scholar] [CrossRef]
- Taylor, R.G.; Whittington, J.A.; Grier, H.J.; Crabtree, R.E. Age, growth, maturation and protandric sex reversal in common snook, Centropomus undecimalis, from the east and west coasts of South Florida. Fish Bull 2000, 98, 612–624. [Google Scholar]
- Tringali, M.D., Bert. Molecular phylogenetics and ecological diversification of the transisthmian fish genus Centropomus (Perciformes:Centropomidae). Molecular Phylogenetics and Evolution 1999, 16, 193–207. [Google Scholar] [CrossRef] [PubMed]
- Patrona, L.D. Contribution to the biology of "sea bass" Centropomus parallelus (Pisces Centropomidae) of Sud-Est du Brésil: possibilités quacoles. 175 f. Thèse (Doctorat decème Cycle, Sciences et Techniques in Productions Animales) - Institut National Polytechnique de Toulouse, France, 1984.
- Mendonça, M.C.F.B. Autoecology of camorim, Centrpomus undecimalis (Bloch, 1792), (Peciformes: Centropomidadae) in hypersaline environment in Galinhos, RN. Brazil. São Carlos. UFSCar. PhD thesis, 145p. 2004.
- Aoki, P. C. M. Thermal tolerance of juveniles of sea bass Centropomus parallelus poey, 1860 (pisces: centropomidae) in freshwater. Dissertation (Master in Biological Sciences) Postgraduate course in Animal Biology, Federal University of Espírito Santo, 1999.
- Barbuio, M. A. T. Comparison of growth and body composition of the seabream Centropomus parallelus (poey, 1860) fed on a commercial diet and dry and semi-moist experimental diets. 57f. Dissertation (Master in Aquaculture) Postgraduate Course in Aquaculture, Federal University of Santa Catarina, Florianópolis. 1999.
- Cerqueira, V.R. Robalo culture: aspects of reproduction, larviculture and fattening. Florianópolis. Ed of the author 2002, 94p.
- Cerqueira, V.R. Cultivation of sea bass, Centropomus parallelus. In Native species for fish farming in Brazil. UFSM: Santa Maria, Brazil, 2005, p.403-431.
- Soligo, T.A. First experiments with breeding, larvae and weaning of sea bass, Centropomus undecimalis in. Brazil. Dissertation, (master's degree) - Federal University of Santa Catarina. Postgraduate Program in Aquaculture, 2007.
- Alvarez-Lajonchère, L.; Tsuzuki, M.Y. A review of methods for Centropomus spp. (snooks) aquaculture and recommendations for the establishment of their culture in Latin America. Aquaculture Research. [CrossRef]
- Adams, A.J.; Hill, J.E.; Kurth, B.N.; Barbour, A.B. Effects of a severe cold event on the subtropical, estuarine-dependent common snook, Centropomus undecimalis. Gulf and Caribbean Research 2010, 24, 13–21. [Google Scholar] [CrossRef]
- Mendonça, J.T.; Chao, L.; Albieri, R.J.; Giarrizzo, T.; da Silva, F.M.S.; Castro, M.G.; Brick Peres, M.; Villwock de Miranda, L.; Vieira, J.P. Centropomus undecimalis. The IUCN Red List of Threatened Species, 1918. [Google Scholar] [CrossRef]
- Cerqueira, V.R.; Tsuzuki, M.Y. A review of spawning induction, larviculture, and juvenile rearing of the fat snook, Centropomus parallelus. Fish Physiology and Biochemistry 2009, 35, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Abdelrahman, H.; El Hady, M.; Alcivar-Warren, A.; Allen, S.; Al-Tobasei, R.; Bao, L., et al..; et al. Aquaculture genomics, genetics and breeding in the United States: current status, challenges, and priorities for future research. BMC Genomics 2007, 18, 191. [Google Scholar] [CrossRef]
- Pritchard, V.L.; Jones, K.; Cowley, D.E. Genetic Diversity within Fragmented Cutthroat Trout Populations. Transactions of the American Fisheries Society 2007, 136, 606–623. [Google Scholar] [CrossRef]
- Mehner, T.; Pohlmann, K.; Elkin, C.; Monaghan, M.T.; Freyhof, J. Genetic mixing from enhancement stocking in commercially exploited vendace populations. Journal of Applied Ecology 2009, 46, 1340–1349. [Google Scholar] [CrossRef]
- Ellegren, H.; Galtier, N. Determinants of genetic diversity. Nature Review Genetics 2016, 17, 422–433. [Google Scholar] [CrossRef]
- Futuyma, D.J.; Kirkpatrick, M. Evolution, 4th edition; Sinauer Associates is an imprint of Oxford University Press: Oxford, UK, 2017. [Google Scholar]
- Sekino, M.; Sugaya, T.; Hara, M.; Taniguchi, N. Relatedness inferred from microsatellite genotypes as a tool for broodstock management of Japanese flounder Paralichthys olivaceus. Aquaculture 2004, 233, 163–172. [Google Scholar] [CrossRef]
- Qiu, D.; Morgan, C.; Shi, J.; Long, Y.; Liu, J.; Li, R.; et al. A comparative linkage map of oilseed rape and its use for QTL analysis of seed oil and erucic acid content. Theor. Appl. Genet 2006, 114, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Chen, L.; Zou, J.; Tian, E.; Xia, W.; Meng, J. Development of a population for substantial new type Brasscia napus diversified at both A/C genomes. Theor. Appl. Genet 2010, 121, 1141–1150. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Ma, B.; Li, H.; Chang, Y.; Han, Y.; Li, J., et al.; et al. Identification, characterization, and utilization of genome-wide simple sequence repeats to identify a QTL of acidity in apple. BMC Genomics 2012, 13, 537. [Google Scholar] [CrossRef]
- Xia, W.; Xiao, Y.; Liu, Z.; Luo, Y.; Mason, A. S.; Fan, H. , et al. Development of gene-based simple sequence repeat markers for association analysis in Cocos nucifera. Mol. Breed.
- Sambrook, J.; Fritschi, E.F.; Maniatis, T. Molecular cloning: a laboratory manual, Cold Spring Harbor Laboratory Press: New York, 1989.
- Seyoum, S.; Tringali, M. D.; Sullivan, J. Isolation and characterization of 27 polymorphic microsatellite loci for the common snook Centropomus undecimalis. Molecular Ecology Notes 2005, 5, 192–194. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research--an update. Bioinformatics, 2537. [Google Scholar]
- Jombart, T. Adegenet: a R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis, 2nd Edition; Springer, New York, 2009.
- Keenan, K.; McGinnity, P.; Cross, T.F.; Crozier, W.W.; Prodöhl, P.A. diveRsity: An R package for the estimation and exploration of population genetics parameters and their associated errors. Methods in Ecology and Evolution 2013, 4, 782–788. [Google Scholar] [CrossRef]
- Paquette, S.R. PopGenKit: Useful Functions for (Batch) File Conversion and Data Resampling in Microsattelite Datasets. R Package Version 1.0. 2012.
- Ellegren, H. Microsatellites: simple sequences with complex evolution. Nat Rev Genet 2004, 5, 435–45. [Google Scholar] [CrossRef]
- International Human Genome Sequencing Consortium. Initial sequencing and analysis of the human genome. Nature 2001, 409, 860–921. [Google Scholar] [CrossRef]
- Hernández-Vidal, U.; Lesher-Gordillo, J.; Contreras-Sánchez, W.M.; Chiappa-Carrara, X. Genetic variability of the common Snook Centropomus undecimalis (Perciformes: Centropomidae) in connected marine and riverine environments. Revista de biologia tropical 2014, 62, 627–636. [Google Scholar] [CrossRef]
- Forbes, S.H.; Hogg, J.T.; Buchanan, F.C.; Crawford, A.M.; Allendorf, F.W. Microsatellite evolution in congeneric mammals: domestic and bighorn sheep. Molecular Biology and Evolution 1995, 12, 1106–1113. [Google Scholar] [PubMed]
- Oliveira, D.C.; Milheiriço, C.; Vinga, S.; de Lencastre, H. Assessment of allelic variation in the ccrAB locus in methicillin-resistant Staphylococcus aureus clones. Journal of Antimicrobial Chemotherapy 2006, 58, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Frankham, R.; Ballou, J.; Briscoe, D. Introduction to Conservation Genetics, 2nd edition; Cambridge University Press: New York, 2010. [Google Scholar]
- Laikre, L.; Palm, S.; Ryman, N. Genetic Population Structure of Fishes: Implications for Coastal Zone Management. Ambio 2005, 34, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Wright, S. Evolution and the Genetics of Populations, vol. 4. Variability Within and Among Natural Populations; University of Chicago Press: Chicago, 1978. [Google Scholar]
- Rice, W.R. Analyzing tables of statistical tests. Evolution 1989, 43, 223–225. [Google Scholar] [CrossRef] [PubMed]
- Able, K.W. A re-examination of fish estuarine dependence: Evidence for connectivity between estuarine and ocean habitats. Estuarine, Coastal and Shelf Science 2005, 64, 5–17. [Google Scholar] [CrossRef]
- Silvano, R. A. M.; MacCord, P. F. L.; Lima, R.V.; Begossi, A. When does this fish Spawn? Fishermen‟s local knowledge of migration and reproduction of Brazilian coastal fishes. Environmental Biology of Fishes 2006, 76, 371–386. [Google Scholar] [CrossRef]
- D’Aanatro, A.; Pereira, A.N.; Lessa, E. P. Genetic structure of the white croaker, Micropogonias furnieri Desmarest 1823 (Perciformes: Sciaenidae) along Uruguayan coasts: contrasting marine, estuarine, and lacustrine populations. Environmental Biology of Fishes 2011, 91, 407–420. [Google Scholar] [CrossRef]

Table 1.
This is a table. Tables should be placed in the main text near to the first time they are cited.
Table 1.
This is a table. Tables should be placed in the main text near to the first time they are cited.
| Specie | Sample code | Number (n) of samples collected | Collection point | |
|---|---|---|---|---|
| Federal State | County or region | |||
| C. parallelus | pSC | 29 | Santa Catarina | Balneário de Penha |
| pSP | 13 | São Paulo | Guarujá | |
| C. undecimalis | uCap | 40 | Santa Catarina | Captive* |
| uSC | 18 | Santa Catarina | Balneário de Penha | |
| uSP | 8 | São Paulo | Guarujá | |
* Captive from CERES/UDESC and UFSC - LAPMAR (Marine Fish Laboratory).
Table 2.
The measure of genetic structure of Centropomus udecimalis and Centropomus paralellus F-statistics. Fst is the proportion of the total genetic variance contained in a subpopulation and Fis (inbreeding coefficient) is the proportion of the variance in the subpopulation contained in an individual.
Table 2.
The measure of genetic structure of Centropomus udecimalis and Centropomus paralellus F-statistics. Fst is the proportion of the total genetic variance contained in a subpopulation and Fis (inbreeding coefficient) is the proportion of the variance in the subpopulation contained in an individual.
| Specie | Parameters | Mean IC | Lower IC | Higher IC |
|---|---|---|---|---|
| C. udecimalis | Fst | 0.0376 | 0.0250 | 0.0532 |
| Fis | -0.0021 | -0.0262 | 0.0202 | |
| C. paralellus | Fst | 0.0592 | 0.0368 | 0.0832 |
| Fis | 0.0020 | -0.0263 | 0.0308 |
Table 3.
The measure of genetic structure of the populations of Centropomus udecimalis and Centropomus paralellus. Fis (inbreeding coefficient) is the proportion of the variance in the subpopulation contained in an individual and AR allele designations represent the number of repeats.
Table 3.
The measure of genetic structure of the populations of Centropomus udecimalis and Centropomus paralellus. Fis (inbreeding coefficient) is the proportion of the variance in the subpopulation contained in an individual and AR allele designations represent the number of repeats.
| Polpulations | Mean_AR | Lower Bound_AR | Higher Bound_AR | Mean_Fis | Lower Bound_Fis | Higher Bound_Fis |
|---|---|---|---|---|---|---|
| uSP | 6.58 | 5.2 | 7.9 | -0.1537 | -0.2618 | -0.0992 |
| uSC | 8.9 | 6.7 | 10.6 | -0.0049 | -0.0655 | 0.0455 |
| uCap | 9.9 | 8 | 11.3 | -0.0113 | -0.0388 | 0.0142 |
| pSC | 10.87 | 9.1 | 12.4 | 0.0275 | -0.0086 | 0.0649 |
| pSP | 8.17 | 6.9 | 9.3 | -0.1404 | -0.1949 | -0.1016 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



