
Submitted:
29 June 2023
Posted:
29 June 2023
You are already at the latest version
Abstract
Target pests of genetically engineered crops producing both defensive allelochemicals and Bacillus thuringiensis (Bt) toxins often sequentially or simultaneously uptake allelochemicals, Bt toxins, and/or insecticides. How the three types of toxins interact to kill pests remains underexplored. Here we investigated the interactions of Bt toxin Vip3A, plant allelochemical flavone, and insecticide emamectin benzoate in Spodoptera frugiperda. Simultaneous administration of flavone LC25 + Vip3A LC25, emamectin benzoate LC25 + Vip3A LC25, and flavone LC15 + emamectin benzoate LC15 + Vip3A LC15 but not flavone LC25 + emamectin LC25 yielded a mortality significantly higher than their expected additive mortality (EAM). One-day preexposure to one toxin at LC5 followed by 6-day exposure to the same toxin at LC5 plus another toxin at LC50 showed that the mortality of flavone LC5 + Vip3A LC50, emamectin benzoate LC5 + Vip3A LC50, and Vip3A LC5 + emamectin benzoate LC50, were significantly higher than their EAM, while that of flavone LC5 + emamectin benzoate LC50 was significantly lower than their EAM. No significant difference existed among the mortalities of Vip3A LC5 + flavone LC50, emamectin benzoate LC5 + flavone LC50, and their EAMs. The results suggest that the interactions of the three toxins are largely synergistic (inductive) or additive, depending on their combinations and doses.
Keywords:
Allelochemical
; Bt toxin
; chemical insecticide
; combined toxicity
; induced toxicity
; synergistic interactions
1. Introduction
Plants and herbivorous insects have antagonistically co-evolved for over 400 million years, whereby plants acquire various morphological and chemical defenses to protect themselves from herbivores, and insects disarm plant defenses for food, survival, and reproduction [1,2,3,4,5]. Among plant chemical defenses are various toxic plant secondary metabolites (i.e., allelochemicals, such as tannins, cyanide, glycosides, alkaloids, terpenoids, saponins, flavonoids, furanocoumarins, indoles, and phytoecdysteroids), nonprotein or unusual amino acids (e.g. canavanine, 3-hydroxyproline), plant defense proteins / enzymes [e.g. lectins, proteinase inhibitors (PIs), peroxidases (POD), polyphenol oxidases (PPO)], and volatile organic compounds (VOCs) [1,4,5,6,7,8,9,10]. Direct use and/or augmentation of the natural defenses of crops, especially breeding and planting insect-resistant varieties that produce more anti-herbivore allelochemicals, are one of the major tactics for integrated pest management [7].
The advancement in biotechnology from 1980s-1990s has enabled introduction of insecticidal toxin or protein genes from other organisms into the genomes of crops, making in-planta production of pest-resistant toxins of foreign source possible [11,12,13,14,15,16]. Commercialization of genetically engineered crops expressing insecticidal proteins from Bacillus thuringiesis (Bt) in 1996 has marked the beginning of a new paradigm in pest management—from labor-intensive and pollution-inevitable toxin sprays to in-planta self-production of toxins [17]. Transgenic Bt crops can provide a safe and highly effective control of major insect pests such as the European corn borer, southwestern corn borer, tobacco budworm, cotton bollworm, pink bollworm, and fall armyworm while decreasing use of conventional insecticides, boosting biological control, and enhancing yields [18,19,20,21,22]. These economic, health and environmental benefits have led to rapid global adoption of transgenic Bt crops. The area planted globally to transgenic Bt crops increased from 1 million hectares (ha) in 1996 to 109 million ha in 2019, which accounted for >53% of the global cultivated area of genetically modified crops [23]. The Bt crops planted by millions of farmers in 27 nations in 2019 include corn, cotton, soybean, sugarcane, and eggplant [23].
Other than expressing the introduced Bt toxins such as Cry1Ac, Cry2Ab and Vip3A, Bt crops, like all plants, also produce their own insect-resistant allelochemicals. This makes target pests more hardly to survive on Bt crops because they have a double “green prevention and control” shield for these pests. Nonetheless, supplementary insecticide sprays may be necessary to address the surviving insects when high dose of Bt toxins cannot be reached [24,25], Bt toxin concentrations decline in late season [26], or practical resistance of pests to Bt toxins occurs [27]. Consequently, pests feeding on Bt crops are inevitable to ingest low or high doses of Bt toxin proteins sequentially or simultaneously, naturally occurring plant defense allelochemicals / proteins, and/or insecticides, depending on the growth stage of the plants being attacked [26].
Whether the three types of toxins synergistically or antagonistically interact with each other when simultaneously ingested and induce or inhibit each other’s toxicity under sequential ingestion remains underexplored. Sachs et al. [28] showed that pyramiding Bt toxin Cry1Ab and terpenoid in cotton provided a higher yet additive level of resistance to Heliothis virescens larvae than Cry1Ab or terpenoid alone. Other reported additive cases include tannic acid + Cry1Ac against Helicoverpa armigera [29], leptin glycoalkaloid + Cry3A against Leptinotarsa decemlineata [30], and gossypol plus Cry1Ac + Cry2Ab against Spodoptera littoralis [31]. Among the reported synergistic cases are azadirachtin + Bacillus thuringiensis Berliner sub sp . kurstaki against H. armigera [32], maize insect resistance cysteine protease (Mir1-CP) + Cry2A against Helicoverpa zea, H. virescens, Spodoptera frugiperda and Diatraea grandiosella [33], gossypol + Cry1Ac against a resistant strain of H. zea [34], jasmonic acid-induced resistance plus Cry1Ac or Cry1Ac + Cry2Ab against S. frugiperda [35], and flavone + Cry1Ac against H. armigera [36]. The antagonistic cases are tannis + Cry1Ac against H. armigera [37], quercetin + Cry1Ac against H. armigera [38], and Bt + trichlorfon against Plutella xylostella [39]. So far, there has been only one sequential ingestion case study, which showed that pre-exposure to Cry1Ac significantly induced flavone’s toxicity against H. armigera, whereas pre-exposure to flavone didn’t induce or inhibit Cry1A’s toxicity against the same pest species [36].
In this study, we used S. frugiperda, an invasive polyphagous target pest of Bt crops, to address the questions of whether sequential or simultaneous ingestion of flavone, a representative of the ubiquitous flavonoids [40,41,42], Vip3A, one of the most widely deployed Bt toxins in Bt transgenic crops [23,43], and emamectin benzoate, an effective insecticide for controlling this pest [44,45,46,47], induce and / or synergize each other’s toxicity. Our results reveal that simultaneous ingestion of lethal doses of the three toxins and any two of the three toxins caused a synergistic interaction for flavone + Vip3A, emamectin benzoate + Vip3A, and flavone + emamectin benzoate + Vip3A, but an additive interaction for flavone + emamectin benzoate. When any two of the three toxins were sequentially ingested, emamectin benzoate induced Vip3A’s toxicity, flavone induced Vip3A’s toxicity but inhibited emamectin benzoate’s toxicity, neither of Vip3A and emamectin benzoate affected flavone’s toxicity.
2. Materials and Methods
2.1. Insect Sources
A laboratory strain of S. frugiperda was used in the current study. The strain was received as a gift from the China Agricultural University in September 2020 and thus named CAU strain hereafter. Pupae used in the experiment were soaked in 5% formaldehyde for 5 min, washed with water and placed into a clean cage to wait for adult emergence. Inside moth rearing cages, a 10% sugar water solution was provided for adult supplementary feeding, as well as a piece of cheesecloth for female oviposition. Pieces of cheesecloth used in the experiment had been sterilized by soaking for 30 s in 5% formaldehyde solution, cleaning with tap water, and drying in the ambient room temperature. The egg masses on the collected pieces of cheesecloth were put in plastic cups till they brooded. Within 6 h, the newly brooded neonate larvae were kept at 26±1 °C and 60±10% relative humidity (RH) with a 16: 8 h (L/D) photoperiod on a semisynthetic diet containing wheat germ [48].
2.2. Preparation of Toxin-Containing Diet
Vip3A protoxin, flavone (reagent grade) and emamectin benzoate (abbreviated as emamectin hereafter) were procured from Beijing Honoster Biotechnology Company (Beijing, China), Shanghai Aladdin Biochemical Technology Company (Shanghai, China) and China Agricultural University, respectively. Acetone, pH 7.4 phosphate (PBS) buffer and triton X-100 were obtained from Luoyang Haohua Chemical Reagent Co., LTD (Luoyang, Henan, China), Sangon Biotech Company (Shanghai, China) and Solarbio Company (Beijing, China), respectively.
The toxin diets were prepared by diet incorporation for flavone, diet overlay for Vip3A, emamectin, or their mixture, and combo of diet incorporation and overlay for mixtures involving flavone plus Vip3A, emamectin or both Vip3A and emamectin. As the diets cooled to approximately 47 °C, 300 µL of flavone solution or acetone (solvent control for flavone) was added to 30 mL diets, vortexed vigorously, and apportioned 0.75 mL to each well of 128-well bioassay trays (C-D International, Pitman, NJ, USA). The diets with overlayed toxins were prepared by dispensing 0.75 mL diets in each well of bioassay trays first. After the diets coagulated, we covered the diets of each well with 60 µL of a given concentration of Vip3A protoxin, emamectin, 0.25 × pH 7.4 PBS buffer (Vip3A protoxin solvent) or acetone (emamectin solvent). We then put the bioassay trays on an orbital shaker rotating at 70 r/m for 30 min to entail formation of a uniform layer of Vip3A, emamectin or the two solvents on the surface of the diets. Preparation of the diets with both incorportated and overlayed toxins began with vigorously vortexing 30 mL diets incorporated with 300 µL of a given concentration of flavone or acetone, followeded by dispensing the diet mixtures to wells (0.75 mL diets/well) of bioassay trays, overlaying 60 µL of a given concentration of Vip3A protein, emamectin, 0.25 × pH 7.4 PBS buffer (Vip3A protoxin solvent) or acetone (emamectin solvent) on the surface of the solidified flavone or acetone-incorporated diets, and finally rotating the bioassay trays on an orbital shaker at 70 r/m for 30 min. The double overlay (overlay+ overlay) diets were made by alloting 0.75 mL diet in each well of bioassay trays, followed by coating the coagulated diets with 60 µL of a given concentration of Vip3A protoxin or 0.25 × pH 7.4 PBS buffer (Vip3A protoxin solvent), overlaying 60 µL of a given concentration of emamectin or acetone (emamectin solvent), and orbiting the trays on an orbital shaker at 70 r/m for 30 min.
2.3. Bioassay
To determine the baseline toxicity of Vip3A protoxin and emamectin, a total of 48 neonate larvae [3 repeats of 16 insects per treatment (different concentrations and control) of S. frugiperda hatched within 6 h were transferred onto the diet surface of each well (1 neonate per well) containing 0.75 mL diet that overlaid with 0.25 × pH 7.4 PBS buffer (0.05 % tritonx-100), acetone (with 0.05 % tritonx-100), different concentrations of Vip3A protoxin, or different concentrations of emamectin. A similar procedure was followed to mesure the baseline toxicity of flavone except that flavone- and acetone-incorporated (control) diets were used. The bioassay trays with neonate larvae were maintained at 26 ± 1 °C, 60 ± 10% R.H., and a photoperiod of 16: 8 h (L/D). The numbers of dead individuals and live larvae in each of the 1st to the 4th instars were recorded after 7 d. The sum of dead and retarded (1st instar) larvae was used to calculate larval mortalities for each control and concentration of the three toxins.
The yielded toxicity baselines of the three toxins were used to calculate the LC5, LC15, LC25, and LC50 doses of each toxin. To assess the combined toxicities of two-toxin (flavone LC25 + Vip3A LC25, Vip3A LC25 + emamectin LC25, flavone LC25 + emamectin LC25) and three-toxin mixtures (flavone LC15 + Vip3A LC15 + emamectin LC15), 48 newly hatched neonate larvae (3 repeats of 16 insects each) were transferred to wells (1 larva /well) of the bioassay trays containing the corresponding solvent control diets, single toxin diets (flavone: LC15 = 197 µg/mL, LC25=300.08 µg/mL; Vip3A: LC15 = 0.03 µg/cm, LC25 = 0.06 µg/cm; emamectin: LC15 = 0.06 ng/cm, LC25 =0.09 ng/cm), two-toxin mixture diets, and three-toxin mixture diets, respectively.
To examine if one day pre-administration of a sublethal dose (≤ LC10) of flavone induces or inhibits the toxicity of Vip3A or emamectin, a total of 192 neonates were exposed to the acetone- or flavone (LC5 = 89.5 µg/ml diet)-incorporated diets, respectively. After 24 h, the 192 larvae on the diets incorporated with LC5 dose of flavone were divided into 4 quarters of 48 larvae each (3 replicates of 16 larvae each) and then transferred the 4 quarters to the diets incorporated with flavone LC5 and coverred with 0.25 × pH 7.4 PBS buffer (flavone only treatment for flavone LC5 + Vip3A LC50), the diets mixed with flavone LC5 and coated with 0.01% acetone (flavone only treatment for flavone LC5 + emamectin LC50), the diets contained flavone LC5 and covered with Vip3A LC50 (0.1µg/cm) (flavone LC5 + Vip3A LC50) and the diets contained flavone LC5 and covered with emamectin LC50 (0.12 ng/cm) (flavone LC5 + emamectin LC50 ), respectively. Likewise, we transferred 4 quarters of the neonate larvae from the the acetone-incorporated diet after 24 h to the diets contained acetone and covered with pH 7.4 0.25 × PBS (Vip3A control), the diets supplemented with acetone and coated with acetone (emamectin control), the diets mixed with acetone and covered with Vip3A LC50 (Vip3A only treatment), and the diets contained acetone and coated with emamectin LC50 (emamectin only treatment), respectively. In the same manner, bioassays of S. frugiperda neonates with Vip3A LC5 (0.008µg/cm), emamectin LC5 (0.039 ng/cm), flavone LC50 (689.5µg/mL), Vip3A LC50 (0.1µg/cm), emamectin LC50 (0.12 ng/cm), Vip3A LC5 + flavone LC50, Vip3A LC5 + emamectin LC50, emamectin LC5 + Vip3A LC50, emamectin LC5 + flavone LC50, and the corresponding solvent controls were carried out to uncover whether one day earlier exposure to Vip3A LC5 induces / inhibits the toxicity of flavone / emamectin as well as whether one day earlier exposure to emamectin LC5 induces / inhibits the toxicity of Vip3A / flavone.
All the aforementioned combined and induced toxicity bioassay treatments and controls were maintained at 26 ± 1 °C, 60 ± 10% R.H. with a 16: 8 h (L/D) photoperiod. The numbers of dead individuals and live larvae in each instar (1st to the 4th) were recorded after 7 d. We used the sum of the dead and retarded (1st instar) larvae for calculation of larval mortalities.
2.4. Data Analysis
The adjusted mortalities of all concentrations and treatments were calculated with Abbott’s formula [49]. Probit analysis was performed to estimate the dose-response baselines, LC50, LC25, and LC5 doses of flavone, Vip3A and emamectin against S. frugiperda larvae using the SPSS software (SPSS, 1998). The differences among the adjusted mortalities of the single toxin alone treatments and sequential or simultaneous combination treatments were compared by the Tukey’s honestly significant difference (HSD) test at P < 0.05 in GraphPad Prism (GraphPad Software Inc., USA). We used Chi-squared test and co-toxicity factor [50] assay to ascertain the nature of interaction [(i.e., synergistic (inductive), additive, or antagonistic (inhibitive)] among flavone, Vip3A and emamectin. If Chi-squared test revealed that the observed adjusted mortality of a consecutive or concomitant mixture was <, =, or > the expected additive mortality (EAM = sum of adjusted mortalities of the corresponding single toxins) of two or three toxins, then a conclusion of antagonistic (or inhibitive), additive, or synergistic (inductive) interaction between the toxins could be drawn. The co-toxicity factor was computed by the following equation:
The co-toxicity factor within the range of > +20, −20 to +20, and < −20 represented antagonistic (inhibitive), additive, and synergistic (inductive) interaction, respectively [50].
3. Results
3.1. Baseline toxicity of Vip3A, flavone and emamectin against S. frugiperda larvae
The bioassays using the neonate larvae of the CAU strain of S. frugiperda were performed to establish the lethal dose-probit regression lines (LD-P line) of flavone, Vip3A, and emamectin and compute three lethal (LC15, LC25, LC50) and one sublethal (LC5) doses of the three toxins (Table 1) for assessing their sequential and simultaneous interactions. Based on their LC50 values, emamectin was about 1150-fold more toxic than Vip3A, and Vip3A was over 3000-fold more potent than flavone (Table 1). Given that the LC50 of the CAU strain for Vip3A [161 ng/cm2 (95% FL: 90-370)] was about the same with that of a Vip3A-susceptible susceptible strain [156.496 ng/cm2 (95%FL: 87.15-217.96)] [51], the CAU strain measured here is a Vip3A-susceptible strain.
3.2. Toxic synergy among Vip3A, emamectin benzoate and flavone against S. frugiperda larvae
Simultaneous exposure of S. frugiperda neonates to the calculated LC25 doses of either two of the three toxins (Vip3A, emamectin and flavone) (Table 1) were conducted to determine the types of their interactions (additive, synergistic or antagonistic). The corrected mortalities for exposure to the calculated LC25 doses of Vip3A (0.06 µg/cm) and flavone (300.08 µg/mL) were 35.42% and 25%, respectively (Table 2, Figure 1A). When S. frugiperda neonates were simultaneously treated with LC25 doses of Vip3A and flavone, the corrected mortality was 81.25%, yielding a co-toxicity factor of > 20 (Table 1), the cutoff value for a significant synergistic interaction between toxins. Consistent with the co-toxicity factor value (34.48 > 20), the observed corrected mortality of the flavone-Vip3A combination was significantly higher than that of the two single toxin treatments and the expected additive mortality (EAM = 35.42% + 25% = 60.42%) of the two toxins (Figure 1A) (Tukey’s HSD and Chi-Square tests).
When S. frugiperda neonates concurrently ingested the calculated LC25 doses of Vip3A (0.06 µg/cm) and emamectin (0.09 ng/cm), we observed an adjusted mortality of 95.83%, which was significantly higher than that of each toxin and their EAM (43.61% + 31.65% = 75.26%) (Tukey’s HSD and Chi-Square tests) and resulted in a co-toxicity factor of 27.34 (>20, the criterium for synergistic interaction) (Table 2, Figure 1B). By contrast, simultaneous exposure to the LC25 doses of flavone (300.08 µg/mL) and emamectin (0.09 ng/cm) caused an adjusted mortality of 70.83% and a co-toxicity factor of 6.5, belonging to the scope of additive interactions (Table 2). Tukey’s HSD and Chi-Squared tests uncovered that the adjusted mortality of the flavone/emamectin mixture was significantly higher than that of each toxin alone but was not different from their EAM (25.0% + 41.67% = 66.67%) (Table 2, Figure 1C).
We also tested the combined toxicity of Vip3A, flavone and emamectin to S. frugiperda neonates, using a ratio of LC15 : LC15 : LC15. The three-toxin combination produced an adjusted mortality of 87.5% and a co-toxicity factor of 40.0 (>20), suggesting a syne rgistic toxicological interplay among the three toxins (Table 2). The corrected mortality of the three-toxin mixture was significantly higher than that of flavone (197.0 µg/g diets, 14.58%),Vip3A (0,03 µg/cm2, 29.17%), emamectin (0.06 ng/cm2, 19.17%) and their EAM ( 14.58% + 29.17% + 19.17% = 62.92%) (Table 2, Figure 1D) (Tukey’s HSD and Chi-Squared tests).
3.3. Asymmetrical toxicity induction between flavone and Vip3A against S. frugiperda larvae
One day earlier exposure of S. frugiperda neonates to the LC5 sublethal dose of flavone (89.5 µg/g diets) followed by 6-day exposure to the LC5 sublethal dose of flavone and the LC50 dose of Vip3A (0.1 µg/cm) (flavone LC5 + Vip3A LC50) effected a corrected mortality of 81.25% and a co-toxicity factor of 85.71 (Table 3). Tukey’s HSD and Chi-Squared tests uncovered that the corrected mortality caused by flavone LC5 + Vip3A LC50 was significantly greater than that of the larvae feeding on the diets with the LC5 sublethal dose of flavone for 7 d (flavone LC5) and of the larvae feeding for 1 d on the diets with acetone (flavone solvent) and then for 6 d on the diets with LC50 dose of Vip3A (Vip3A LC50) and the EAM of flavone LC5 and Vip3A LC50 (6.25% + 37.5% = 43.75%) (Table 3, Figure 2A).
By contrast, one day earlier exposure of S. frugiperda neonates to the LC5 sublethal concentration of Vip3A (0.008 µg/cm2) and then 6-day exposure to the LC5 sublethal concentration of Vip3A plus the LC50 dosage of flavone (689.6 µg/g diets) (Vip3A LC5 + flavone LC50) provoked an adjusted mortality of 62.08% and a co-toxicity factor of 6.43 (Table 3), which fell within the range (-20 to 20) of additive interactions [50]. Consistent with this inference, the correct mortality elicited by Vip3A LC5 + flavone LC50 was not greater than that of the larvae exposed to 0.25 × PBS buffer (Vip3A solvent) for 1 d than then to the LC50 dose of flavone for 6 d (flavone LC50) and the EAM of Vip3A LC5 + flavone LC50 (Tukey’s HSD and Chi-Square tests) (Table 3 and Figure 2B).
3.4. Asymmetrical toxicity induction between Vip3a and emamectin benzoate against S. frugiperda larvae
The corrected mortality for S. frugiperda neonates exposed to LC5 sublethal dose of Vip3A (0.008 µg/cm2) for 1 d followed by 6 d of LC5 sublethal dose of Vip3A plus LC50 dose of emamectin (0.12 ng/cm2) (Vip3A LC5 + emamectin LC50) was 49.4%, yielding a co-toxicity factor of 5.86 (Table 3). This mortality was significantly higher than that of the larvae fed for 1 d on the diets overlaid with 0.25 × PBS buffer (Vip3A solvent) and then for 6 d on the diets covered with LC50 dosage of emamectin (emamectin LC50), but not different from the expected additive mortality (EAM= 46.67%) of Vip3A LC5 (4.17%) and emamectin LC50 (42.5%) (Tukey’s HSD and Chi-Square tests) (Table 3, Figure 3A).
When S. frugiperda neonates were first exposed to the LC5 sublethal concentration of emamectin (0.039 ng/cm2) and then to the LC5 sublethal concentration of emamectin plus LC50 dose of Vip3A (0.1ug/cm) (emamectin LC5 + Vip3A LC50), we observed a corrected mortality of 52.08.% and a co-toxicity factor of 47.06 (Table 3), belonging to the range of induction. Tukey’s HSD and Chi-Squared tests revealed that the adjusted mortality caused by emamectin LC5 + Vip3A LC50 was notably higher than that of Vip3A LC50 (33.33%) and the EAM (35.42%) of Vip3A LC50 and emamectin LC5 (2.08%) (Table 3, Figure 3B).
3.5. Asymmetrical toxicity inhibition between flavone and emamectin benzoate against S. frugiperda larvae
Seven-day exposure to LC5 sublethal dose (89.5 µg/g diets) of flavone (flavone LC5) and 1-d exposure to acetone (flavone solvent) plus subsequent 6-d exposure to LC50 dose (0.12 ng/cm2) of emamectin (emamectin LC50) resulted in an adjusted death rate of 12.5% and 33.33% (Table 3), respectively. Sequential exposure to the two toxins, i.e., 1-d exposure to flavone LC5 followed by 6-d exposure to flavone LC5 plus emamectin LC50 (flavone LC5 + emamectin LC50), yielded a corrected mortality of 25.0% and co-toxicity factor of -45.45, far below the critical value (-20) for inhibitive interaction (Table 3). In agreement with this inference, the corrected mortality of flavone LC5 + emamectin LC50 was significantly lower than that of the EAM (45.83%) of flavone LC5 and emamectin LC50 (Table 3, Figure 4A) (Tukey’s HSD and Chi-Squared tests).
When the order of sequential exposure to flavone and emamectin was reversed, the corrected mortality (56.25%) of emamectin LC5 + flavone LC50 was not significantly different from that (50.0%) of flavone LC50 (689.6 µg/g diets) and the EAM (0% + 50% = 50%) of flavone LC50 and emamectin LC5 (0.039 ng/cm2) (Table 3, Figure 4B) (Tukey’s HSD and Chi-Squared tests). The co-toxicity factor was 12.5, which fell within the range of additive interaction (-20 to <20) (Table 3, Figure 4B).
4. Discussion
S. frugiperda and other target pests of Bt-transgenic crops may concurrently or sequentially encounter Bt toxins, ant-herbivore allelochemicals and/or insecticides, depending on the spatiotemporal expression patterns of Bt toxins [52,53] and plant defense allelochemicals [54,55,56] as well as the timing of insecticide sprays. Binary and ternary mixtures of flavone, Vip3A and emamectin devised to imitate concurrent consuming of the three types of poisons exhibited a toxicological synergy for flavone + Vip3A, emamectin + Vip3A, and flavone + emamectin + Vip3A, but an additive interaction for flavone + emamectin (Table 2 and Figure 1). In theory, synergy can arise only if the mixed poisons reciprocally elevate each other’s toxicity or at least one of the mixed poisons boosts the potency of the other one or two poisons. Our induced toxicity experiments simulated the sequential intake of any two of the three toxins detected a significant induction effect of one-day earlier feeding of a sublethal concentration (LC5) of flavone and emamectin on the toxicity of Vip3A LC50 but did not find an induction effect of one-day earlier ingestion of Vip3A LC5 on the toxicity of flavone LC50 and emamectin LC50 (Table 3 and Figure 2 and Figure 3). On the contrary, consuming flavone LC5 one day in advance inhibited the toxicity of emamectin LC50, but consuming emamectin LC5 one day in advance didn’t affect the toxicity of flavone LC50 (Table 3 and Figure 4). The asymmetrical induction of the poisonousness hints at that the synergy found in the mixtures of flavone + Vip3A, emamectin + Vip3A, and Flavone + emammectin +Vip3A (Table 2 and Figure 1) were likely resulted from elevation of Vip3A toxicity by flavone, emamectin, or both, rather than they reciprocally strengthened each other’s toxicity. The fact that the co-toxicity factor values of the binary mixtures of flavone + Vip3A and emamectin + Vip3A were about half of those of the sequential mixtures of flavone LC5 + Vip3A LC50 and emamectin LC5 + Vip3A LC50 (Table 3) supports this speculation.
There are three possible routes by which flavone and/or emamectin can enhance the toxicity of Vip3A. Flavone and/or emamectin may synergistically upregulate the expression of the receptors of Vip3A, such as the putative Vip3Aa-binding ribosomal protein S2 [57], and/or the Vip3A-activated apoptosis pathway genes [58]. They may also upregulate the protease genes responsible for activation of Vip3A, such as trypsin [59]. The third approach is to inprove the accessibility of Vip3A to its receptor protein(s) by promoting Vip3A’s entrance into its receptor sites and/or decreasing its degradation, accumulation, and/or evacuation. In agreement with the three possible routes, flavone is known to up- and down-regulate 295 and 125 genes, respectively, in Spodopera litura [60], a sister species of S. frugiperda, and 38 and 10 genes, respectively, in H. armigera [61]. By the same token, emamectin can up- and down-regulate 599 and 1658 genes, respectively, in the predatory beetle Paederus fuscipes [62], and 1495-2784 and 1622-2351 genes, respectively, in Spodoptera exigua, depending on the dose of emamectin [63]. Additional experiments are required to illuminate how flavone and emamectin induce and elevate the pontency of Vip3A to S. frugiperda.
On the other hand, the sequential flavone inhibition of emamectin toxicity failed to decide the type or nature of concurrent interactions between emamectin and flavone since an additive but not antagonistic interaction was observed for the binary mixture of flavone + emamectin (Table 2 and Figure 1). This suggests that the nature of simultaneous interactions of two or more toxins may not be always explained by their sequential interactions, especially when asymmetrical inhibition occurs. Our finding of asymmetrical inhibition of emamectin toxicity by flavone is consistent with the lack of cross-resistance to the plant allelochemical 2-tridecanone in the insecticide (fenvalerate)-resistant Colorado potato beetle [64] as well as the unidirectional (asymmetrical) cross resistance to several organophosphate pesticides of the triterpenoid cucurbitacin-C-selected twospotted spider mite [65] and to the pyrethroid insecticide α-cypermethrin of the plant allelochemical xanthotoxin-exposed H. zea survivors and their offspring [66]. Along the same line, pre-exposure to flavone effectively enhanced detoxification enzyme activities and larval tolerance to multiple synthetic insecticides in Spodoptera litura by turning on ROS-CncC-mediating xenobiotic metabolism pathway [60]. Pre-ingestion of the flavonoids catechin, myricetin kaempferol, quercetin, and rutin markedly enhanced P450 activity and resistance to flupyradifurone and thiamethoxam in Bemisia tabaci [67]. And preexposure to visnagin, DIMBOA (2,4-Dihydroxy-7-methoxy-1,4-benzoxazin-3-one), coumarin and flavone significantly decreased larval susceptibility of H. armigera to methomyl via upregulation of P450s including CYP6B2, CYP6B6, and CYP6B7 [68]. Such unidirectional sequential inhibition and unidirectional cross resistance between insecticides and anti-herbivore allelochemicals are probably prompted by the absence of behavioral adjustment to the repellent and/or antifeedant influences of plant allelochemicals in insecticide-preexposed individuals or survivors, rather than by the lack of metabolic adaptation [66]. After all, insecticides are also capable of inducing metabolic enzymes [63,66], and at least some of the induced or selected detoxification enzymes are capable of metabolizing both insecticides and allelochemicals [66].
The resistance of pests to Bt-transgenic crops and insecticides is the major challenge for pest management. Developing new Bt crops that can dramatically enhance the manufacture of anti-herbivore allelochemicals / proteins is one possible strategy to address this challenge [31,69], exposing target pests to the redundant and synergistic killing effect of the two or even three (when spraying insecticides) types of toxins. The basic requirements for anti-herbivore allelochemicals / proteins to be co-expressed with Bt insecticidal proteins in new Bt crops include 1) additive or synergistic interaction with the corresponding Bt toxins and even with the commonly used insecticides if possible; 2) negative or no cross resistance with the corresponding Bt toxins and even with insecticides if possible; 3). natural presence in major crops. The allelochemical flavone apparently meets the third requirement as it is naturally present in a wide range of plants including corn, cotton, and soybean [40,41,42]. The data reported here demonstrate that flavone also meets the first requirement as it synergistically interacted with Vip3a and additively with emamectin against S. frigiperda. Moreover, flavone is known to toxicologically synergize with Cry1Ac [36] and Cry2Ab (unpublish data) against H. armigera and with Cry1Ab against S. frugiperda (unpublish data) (He H., personal communication). Given the popularity of Cry1Ab, Cry1Ac, Cry105, Cry2Ab, and Vip3a in Bt crops (https://www.texasinsects.org/bt-corn-trait-table.html), flavone would be a suitable allelochemical to be stacked with these Bt toxins to manage pests if further cross-resistance studies confirm that it also meets the second requirement.
Supplementary Materials
Not applicable.
Author Contributions
X.L. and Z.D. conceived and designed the experiments. K.H., S.W. and H.H. performed the experiments. X.L., Z.D., H.H. and K.H. analyzed the data. X.L., K.H., M.Z., X.C. and X.N. wrote the manuscript. X.L., Z.D. and K.H. shared in the scoping and writing responsibilities. All authors have read and approved the manuscript for publication.
Funding
This research was funded by the National Science Foundation of China (NSFC)-Henan Joint major grant (No. U2004206), Key Scientific Research Projects of Colleges and Universities in Henan Province (No. 21A210027), State Key Laboratory of Cotton Biology (No. CB2020A06) and the USDA National Institute of Food and Agriculture [hatch grant ARZT-1360890-H31-164 and multi state grant ARZT-1370680-R31-172 (NC246)].
Data Availability Statement
Not applicable.
Acknowledgments
We appreciate Dr. Shaohua Gu for providing the CAU strain of S. frugiperda and MS. Lihong Yang for her help in rearing S. frugiperda.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Li, X. and Ni, X. Deciphering the plant-insect phenotypic arms race. Recent Adv Entomol Res 2011, 3, 3–33. [CrossRef]
- Schuman, M. C.; Baldwin, I. T. , The layers of plant responses to insect herbivores. Annu Rev Entomol 2016, 61, 373–94. [Google Scholar] [CrossRef]
- Erb, M. , Plant defenses against herbivory: closing the fitness gap. Trends in plant science 2018, 23(3), 187–194. [Google Scholar] [CrossRef]
- Tanda, A.S. Mutualistic plant associations related to insect resistance. In Molecular Advances in Insect Resistance of Field Crops: Modern and Applied Approaches, Tanda, A.S., Ed. Springer International Publishing: Cham, 2022; pp. 1-42.
- War, A.R.; Buhroo, A.A.; Hussain, B.; Ahmad, T.; Nair, R.M.; Sharma, H.C. Plant defense and insect adaptation with reference to secondary metabolites. In Co-Evolution of Secondary Metabolites, Mérillon, J.-M., Ramawat, K.G., Eds. Springer International Publishing: Cham, 2020; pp. 795-822.
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.; Brennan, R.M.; Graham, J.; Karley, A.J. Plant Defense against herbivorous pests: exploiting resistance and tolerance traits for sustainable crop protection. Front. Plant Sci. 2016, 7, 1132. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Malook, S.U.; Shen, G.; Gao, L.; Zhang, C.; Li, J.; Zhang, J.; Wang, L.; Wu, J. Current understanding of maize and rice defense against insect herbivores. Plant Divers. 2018, 40, 189–195. [Google Scholar] [CrossRef]
- Karban, R. The ecology and evolution of induced responses to herbivory and how plants perceive risk. Ecol. Entomol. 2020, 45, 1–9. [Google Scholar] [CrossRef]
- Brooker, N.L.; Long, J.H.; Stephan, S.M. Field assessment of plant derivative compounds for managing fungal soybean diseases. Biochem. Soc. Trans. 2000, 28, 917–920. [Google Scholar] [CrossRef]
- Hilder, V.A.; Gatehouse, A.M.R.; Sheerman, S.E.; Barker, R.F.; Boulter, D. A novel mechanism of insect resistance engineered into tobacco. Nature 1987, 330, 160–163. [Google Scholar] [CrossRef]
- Adang, M.J.; Brody, M.S.; Cardineau, G.; Eagan, N.; Roush, R.T.; Shewmaker, C.K.; Jones, A.; Oakes, J.V.; McBride, K.E. The reconstruction and expression of a Bacillus thuringiensis cryIIIA gene in protoplasts and potato plants. Plant Mol. Biol. 1993, 21, 1131–1145. [Google Scholar] [CrossRef] [PubMed]
- Peferoen, M. Engineering of insect-resistant plants with Bacillus thuringiensis crystal protein genes. In Plant genetic manipulation for crop protection., Gatehouse, A., Hilder, V., Boulter, D., Eds. CAB International: Wallingford, UK, 1992; pp. 135-153.
- Koziel, M.G.; Carozzi, N.B.; Currier, T.C.; Currier, T.C.; Warren, G.W.; Evola, S.V. The insecticidal crystal proteins of Bacillus thuringiensis: past, present and future uses. Biotechnol. Genet. Eng. Rev. 1993, 11, 171–228. [Google Scholar] [CrossRef]
- Vaeck, M.; Reynaerts, A.; Höfte, H.; Jansens, S.; De Beuckeleer, M.; Dean, C.; Zabeau, M.; Montagu, M.V.; Leemans, J. Transgenic plants protected from insect attack. Nature 1987, 328, 33–37. [Google Scholar] [CrossRef]
- Potrykus, I. Gene transfer to plants: assessment of published approaches and results. Ann. Rev. Plant Biol. 1991, 42, 205–225. [Google Scholar] [CrossRef]
- Estruch, J.J.; Carozzi, N.B.; Desai, N.; Duck, N.B.; Warren, G.W.; Koziel, M.G. Transgenic plants: an emerging approach to pest control. Nat. Biotechnol. 1997, 15, 137–141. [Google Scholar] [CrossRef]
- Betz, F.S.; Hammond, B.G.; Fuchs, R.L. Safety and advantages of Bacillus thuringiensis-protected plants to control insect pests. Regul. Toxicol. Pharmacol. 2000, 32, 156–173. [Google Scholar] [CrossRef] [PubMed]
- Hutchison, W.D.; Burkness, E.C.; Mitchell, P.D.; Moon, R.D.; Leslie, T.W.; Fleischer, S.J.; Abrahamson, M.; Hamilton, K.L.; Steffey, K.L.; Gray, M.E. , et al. Areawide suppression of European corn borer with Bt maize reaps savings to non-Bt maize growers. Science 2010, 330, 222–225. [Google Scholar] [CrossRef] [PubMed]
- Dively, G.P.; Venugopal, P.D.; Bean, D.; Whalen, J.; Holmstrom, K.; Kuhar, T.P.; Doughty, H.B.; Patton, T.; Cissel, W.; Hutchison, W.D. Regional pest suppression associated with widespread Bt maize adoption benefits vegetable growers. Proc. Natl. Acad. Sci. U. S. A. 2018, 115, 3320–3325. [Google Scholar] [CrossRef] [PubMed]
- Romeis, J.; Naranjo, S.E.; Meissle, M.; Shelton, A.M. Genetically engineered crops help support conservation biological control. Biol. Control 2019, 130, 136–154. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Liesner, L.R.; Ellsworth, P.C.; Unnithan, G.C.; Fabrick, J.A.; Naranjo, S.E.; Li, X.; Dennehy, T.J.; Antilla, L.; Staten, R.T. , et al. Transgenic cotton and sterile insect releases synergize eradication of pink bollworm a century after it invaded the United States. Proc. Natl. Acad. Sci. U. S. A. 2021, 118, e2019115118. [Google Scholar] [CrossRef]
- ISAAA. Global status of commercialized biotech/GM crops in 2019: Biotech crops drive socio-economic development and sustainable environment in the new frontier; Cornell University: Ithaca, NY, USA, 2019. [Google Scholar]
- IRAC. Industry perspectives on insect resistance monitoring for transgenic insect-protected crops. Availabe online: https://irac-online.org/documents/industry-perspectives-on-ir-monitoring/?ext=pdf (accessed on 06 June 2023).
- Yates-Stewart, A.D.; Yorke, B.T.; Willse, A.; Fridley, J.; Head, G.P. Using sentinel plots to monitor for changes in thrips susceptibility to MON 88702 cotton containing the Cry51Aa2. 834_16 Bt Protein. Insects 2023, 14, 497. [Google Scholar] [CrossRef] [PubMed]
- Carrière, Y.; DeGain, B.; Unnithan, G.C.; Harpold, V.S.; Li, X.; Tabashnik, B.E. Seasonal declines in Cry1Ac and Cry2Ab concentration in maturing cotton favor faster evolution of resistance to pyramided Bt cotton in Helicoverpa zea (Lepidoptera: Noctuidae). J. Econ. Entomol. 2019, 112, 2907–2914. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Fabrick, J.A.; Carrière, Y. Global patterns of insect resistance to transgenic Bt crops: The first 25 years. J. Econ. Entomol. 2023, 116, 297–309. [Google Scholar] [CrossRef]
- Sachs, E.S.; Benedict, J.H.; Taylor, J.F.; Stelly, D.M.; Davis, S.K.; Altman, D.W. Pyramiding CryIA (b) insecticidal protein and terpenoids in cotton to resist tobacco budworm (Lepidoptera: Noctuidae). Environ. Entomol. 1996, 25, 1257–1266. [Google Scholar] [CrossRef]
- Guan, X.; Liu, X.; Lu, Z.; Zhao, Z.; Zhang, Q.; Xia, J. Influence of tannic acid and Cry1Ac toxin of Bacillus thuringiensis on larval survival, growth, and development of Helicoverpa armigera. Entomol. Exp. Appl. 2009, 132, 50–58. [Google Scholar] [CrossRef]
- Coombs, J.J.; Douches, D.S.; Li, W.; Grafius, E.J.; Pett, W.L. Combining engineered (Bt-cry3A) and natural resistance mechanisms in potato for control of Colorado potato beetle. J. Am. Soc. Hortic. Sci. 2002, 127, 62–68. [Google Scholar] [CrossRef]
- Hagenbucher, S.; Eisenring, M.; Meissle, M.; Romeis, J. Interaction of transgenic and natural insect resistance mechanisms against Spodoptera littoralis in cotton. Pest Manag. Sci. 2017, 73, 1670–1678. [Google Scholar] [PubMed]
- Abedi, Z.; Saber, M.; Vojoudi, S.; Mahdavi, V.; Parsaeyan, E. Acute, sublethal, and combination effects of azadirachtin and Bacillus thuringiensis on the cotton bollworm, Helicoverpa armigera. J. Insect Sci. 2014, 14, 30. [Google Scholar] [CrossRef]
- Mohan, S.; Ma, P.W.; Williams, W.P.; Luthe, D.S. A naturally occurring plant cysteine protease possesses remarkable toxicity against insect pests and synergizes Bacillus thuringiensis toxin. PLoS One 2008, 3, e1786. [Google Scholar] [CrossRef]
- Anilkumar, K.J.; Sivasupramaniam, S.; Head, G.; Orth, R.; Van Santen, E.; Moar, W.J. Synergistic interactions between Cry1Ac and natural cotton defenses limit survival of Cry1Ac-resistant Helicoverpa zea (Lepidoptera: Noctuidae) on Bt cotton. J. Chem. Ecol. 2009, 35, 785–795. [Google Scholar] [CrossRef]
- Mészáros, A.; Beuzelin, J.M.; Stout, M.J.; Bommireddy, P.L.; Rita Riggio, M.; Rogers Leonard, B. Jasmonic acid-induced resistance to the fall armyworm, Spodoptera frugiperda, in conventional and transgenic cottons expressing Bacillus thuringiensis insecticidal proteins. Entomol. Exp. Appl. 2011, 140, 226–237. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, M.; Huang, J.; Li, L.; Huang, K.; Zhang, Y.; Li, Y.; Deng, Z.; Ni, X.; Li, X. Inductive and synergistic interactions between plant allelochemical flavone and Bt toxin Cry1Ac in Helicoverpa armigera. Insect Sci. 2021, 28, 1756–1765. [Google Scholar] [PubMed]
- Olsen, K.M.; Daly, J.C. Plant-toxin interactions in transgenic Bt cotton and their effect on mortality of Helicoverpa armigera (Lepidoptera: Noctuidae). J. Econ. Entomol. 2000, 93, 1293–1299. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Guan, X.; Michaud, J.P.; Zhang, Q.; Liu, X. Quercetin interacts with Cry1Ac protein to affect larval growth and survival of Helicoverpa armigera. Pest Manag. Sci. 2016, 72, 1359–1365. [Google Scholar] [CrossRef]
- Xu, J.; Cai, Y.S.; Zhu, J.L. The study on join-toxicity of Bacillus thuringiensis with insecticides. Jiangsu Agricultural Sciences 2001, 010, 45–46. [Google Scholar]
- Nix, A.; Paull, C.; Colgrave, M. Flavonoid profile of the cotton plant, Gossypium hirsutum: a review. Plants (Basel) 2017, 6, 43. [Google Scholar] [CrossRef]
- Mathesius, U. Flavonoid functions in plants and their interactions with other organisms. Plants (Basel) 2018, 7, 30. [Google Scholar] [CrossRef]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [PubMed]
- Xiao, Y.; Wu, K. Recent progress on the interaction between insects and Bacillus thuringiensis crops. Philos. Trans. R Soc. Lond B Biol. Sci. 2019, 374, 20180316. [Google Scholar]
- Koffi, D.; Kyerematen, R.; Osae, M.; Amouzou, K.; Eziah, V.Y. Assessment of Bacillus thuringiensis and emamectin benzoate on the fall armyworm Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) severity on maize under farmers’ fields in Ghana. Int. J. Trop. Insect Sci. 2022, 42, 1619–1626. [Google Scholar] [CrossRef]
- Zhang, J.; Jiang, J.; Wang, K.; Zhang, Y.; Liu, Z.; Yu, N. A Binary mixture of emamectin benzoate and chlorantraniliprole supplemented with an adjuvant effectively controls Spodoptera frugiperda. Insects 2022, 13, 1157. [Google Scholar] [PubMed]
- Bird, L.; Miles, M.; Quade, A.; Spafford, H. Insecticide resistance in Australian Spodoptera frugiperda (J.E. Smith) and development of testing procedures for resistance surveillance. PLoS One 2022, 17, e0263677. [Google Scholar]
- Zhang, D.-d.; Xiao, Y.-t.; Xu, P.-j.; Yang, X.-m.; Wu, Q.-l.; Wu, K.-m. Insecticide resistance monitoring for the invasive populations of fall armyworm, Spodoptera frugiperda in China. J. Integr. Agr. 2021, 20, 783–791. [Google Scholar] [CrossRef]
- Waldbauer, G.P.; Friedman, S. Self-selection of optimal diets by insects. Ann. Rev. Entomol. 1991, 36, 43–63. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. 1925. J. Am. Mosq. Control Assoc. 1987, 3, 302–303. [Google Scholar] [PubMed]
- Mansour, N.A.; Eldefrawi, M.E.; Toppozada, A.; Zeid, M. Toxicological studies on the egyptian cotton leaf worm, Prodenia litura. VI. Potentiation and antagonism of organophosphorus and carbamate insecticides. J. Econ. Entomol. 1966, 59, 307–311. [Google Scholar] [CrossRef]
- Bergamasco, V.B.; Mendes, D.R.; Fernandes, O.A.; Desidério, J.A.; Lemos, M.V. Bacillus thuringiensis Cry1Ia10 and Vip3Aa protein interactions and their toxicity in Spodoptera spp. (Lepidoptera). J. Invertebr. Pathol. 2013, 112, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Zhang, D.; Zhang, R.; Wang, Y.; Meng, Z.; Zhou, T.; Liang, C.; Zhu, T.; Guo, S. Bt protein expression in the transgenic insect-resistant cotton in China. Sci. Bull. 2016, 61, 1555–1557. [Google Scholar] [CrossRef]
- Carrière, Y.; Degain, B.A.; Unnithan, G.C.; Harpold, V.S.; Heuberger, S.; Li, X.; Tabashnik, B.E. Effects of seasonal changes in cotton plants on the evolution of resistance to pyramided cotton producing the Bt toxins Cry1Ac and Cry1F in Helicoverpa zea. Pest Manag. Sci. 2018, 74, 627–637. [Google Scholar] [CrossRef] [PubMed]
- Justesen, H.; Andersen, A.S.; Brandt, K. Accumulation of anthocyanins and flavones during bud and flower development in Campanula isophylla Moretti. Ann. Bot. 1997, 79, 355–360. [Google Scholar] [CrossRef]
- Sunila, A.V.; Murugan, K. Variation in phenolics, flavonoids at different stages of fruit development of Pouteria campechiana (Kunth) baehni and its antioxidant activity. Int. J. Pharm. Pharm. Sci. 2017, 9, 70–75. [Google Scholar] [CrossRef]
- Chang, X.; Lu, Y.; Lin, Z.; Qiu, J.; Guo, X.; Pan, J.; Abbasi, A.M. Impact of leaf development stages on polyphenolics profile and antioxidant activity in Clausena lansium (Lour.) Skeels. Biomed. Res. Int. 2018, 2018, 7093691. [Google Scholar] [CrossRef]
- Ayra-Pardo, C.; Ochagavía, M.E.; Raymond, B.; Gulzar, A.; Rodríguez-Cabrera, L.; Rodríguez de la Noval, C.; Morán Bertot, I.; Terauchi, R.; Yoshida, K.; Matsumura, H. , et al. HT-SuperSAGE of the gut tissue of a Vip3Aa-resistant Heliothis virescens (Lepidoptera: Noctuidae) strain provides insights into the basis of resistance. Insect Sci. 2019, 26, 479–498. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Shan, Y.; Peng, Y.; Wang, P.; Li, Q.; Yu, S.; Zhang, L.; Xiao, Y. An Integrative Analysis of Transcriptomics and Proteomics Reveals novel insights into the response in the midgut of Spodoptera frugiperda Larvae to Vip3Aa. Toxins (Basel) 2022, 14, 55. [Google Scholar] [CrossRef] [PubMed]
- Song, F.; Chen, C.; Wu, S.; Shao, E.; Li, M.; Guan, X.; Huang, Z. Transcriptional profiling analysis of Spodoptera litura larvae challenged with Vip3Aa toxin and possible involvement of trypsin in the toxin activation. Sci. Rep. 2016, 6, 23861. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Cheng, Y.; Li, Y.; Li, W.; Zeng, R.; Song, Y. Phytochemical flavone confers broad-spectrum tolerance to insecticides in Spodoptera litura by activating ROS/CncC-mediated xenobiotic detoxification pathways. J. Agric. Food Chem. 2021, 69, 7429–7445. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Zhang, Y.; Fang, L.; Zhang, M.; Wang, L.; Ni, X.; Li, X. Identification of the flavone-inducible counter-defense genes and their cis-elements in Helicoverpa armigera. Toxins 2023, 15, 365. [Google Scholar] [CrossRef]
- Khan, M.M.; Khan, A.H.; Ali, M.W.; Hafeez, M.; Ali, S.; Du, C.; Fan, Z.; Sattar, M.; Hua, H. Emamectin benzoate induced enzymatic and transcriptional alternation in detoxification mechanism of predatory beetle Paederus fuscipes (Coleoptera: Staphylinidae) at the sublethal concentration. Ecotoxicology 2021, 30, 1227–1241. [Google Scholar] [CrossRef]
- Sun, Y.; Tao, S.; Zhang, W.; Jiang, B.; Dai, H.-Y.; Liu, B.-S.; Zhang, Y.-J.; Kong, X.-d.; Zhao, J.; Bai, L.-X. Transcriptome profile analysis reveals the emamectin benzoate-induced genes associated with olfaction and metabolic detoxification in Spodoptera exigua Hübner (Lepidoptera: noctuidae). All Life 2022, 15, 340–357. [Google Scholar] [CrossRef]
- Kennedy, G.G.; Farrar Jr, R.R. Response of insecticide-resistant and susceptible Colorado potato beetles, Leptinotarsa decemlineata to 2-tridecanone and resistant tomato foliage: the absence of cross resistance. Entomol. Exp. Appl. 1987, 45, 187–192. [Google Scholar] [CrossRef]
- Gould, F.; Carroll, C.R.; Futuyma, D.J. Cross-resistance to pesticides and plant defenses: A study of the two-spotted spider mite. Entomol. Exp. Appl. 1982, 31, 175–180. [Google Scholar] [CrossRef]
- Li, X.; Zangerl, A.R.; Schuler, M.A.; Berenbaum, M.R. Cross-resistance to alpha-cypermethrin after xanthotoxin ingestion in Helicoverpa zea (Lepidoptera: Noctuidae). J. Econ. Entomol. 2000, 93, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yang, F.; Tong, H.; Hu, Y.; Zhang, X.; Tian, T.; Zhang, Y.; Su, Q. Plant flavonoids enhance the tolerance to thiamethoxam and flupyradifurone in whitefly Bemisia tabaci (Hemiptera: Aleyrodidae). Pestic. Biochem. Physiol. 2021, 171, 104744. [Google Scholar] [CrossRef]
- Chen, S.; Elzaki, M.E.A.; Ding, C.; Li, Z.F.; Wang, J.; Zeng, R.S.; Song, Y.Y. Plant allelochemicals affect tolerance of polyphagous lepidopteran pest Helicoverpa armigera (Hübner) against insecticides. Pestic. Biochem. Physiol. 2019, 154, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Trapero, C.; Wilson, I.W.; Stiller, W.N.; Wilson, L.J. Enhancing integrated pest management in GM cotton systems using host plant resistance. Front. Plant Sci. 2016, 7, 500. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Combined toxicity of Vip3A + flavone (A), Vip3A + emamectin (B), flavone + emamectin (C), and Vip3A + flavone + emamectin (D) against S. frugiperda neonates. Unconverted average mortalities ± SE (standard error) are presented. Mortality data were arcsine converted prior to statistical analysis. Within each small graph, mean mortalities followed by the same letter are not significantly different at P > 0.05 (Tukey’s HSD test). Significant difference is depicted by an asterisk (*) between the observed mortality and expected additive mortality (EAM) of each mixture at P < 0.05 (Chi-square test). Fla = flavone, EMB = emamectin benzoate, NS = No significance.
Figure 1.
Combined toxicity of Vip3A + flavone (A), Vip3A + emamectin (B), flavone + emamectin (C), and Vip3A + flavone + emamectin (D) against S. frugiperda neonates. Unconverted average mortalities ± SE (standard error) are presented. Mortality data were arcsine converted prior to statistical analysis. Within each small graph, mean mortalities followed by the same letter are not significantly different at P > 0.05 (Tukey’s HSD test). Significant difference is depicted by an asterisk (*) between the observed mortality and expected additive mortality (EAM) of each mixture at P < 0.05 (Chi-square test). Fla = flavone, EMB = emamectin benzoate, NS = No significance.
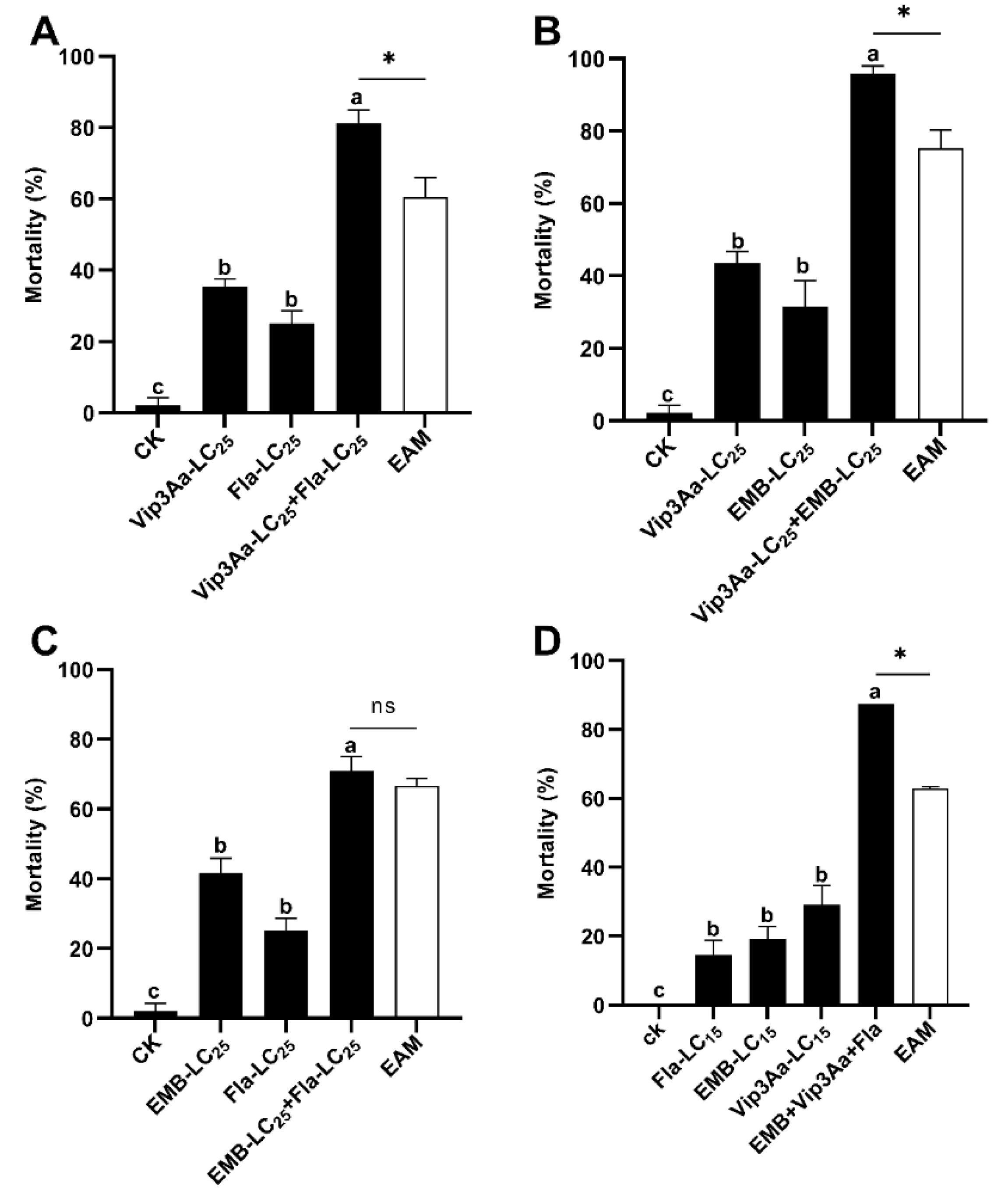
Figure 2.
Induced toxicity of flavone LC5 + Vip3A LC50 (A) and Vip3A LC5 + flavone LC50 (B) against S. frugiperda neonates. Untransformed average mortality ± SE are presented. Mortality data were arcsine converted prior to statistical analysis. Within each small graph, mean mortalities followed by the same letter are not significantly different at P > 0.05 (Tukey’s HSD tests). Significant difference is depicited by an asterisk (*) between the observed mortality and the expected additive mortality (EAM) of each mixture at P < 0.05 (Chi-squared test). Fla = flavone, NS = No significance.
Figure 2.
Induced toxicity of flavone LC5 + Vip3A LC50 (A) and Vip3A LC5 + flavone LC50 (B) against S. frugiperda neonates. Untransformed average mortality ± SE are presented. Mortality data were arcsine converted prior to statistical analysis. Within each small graph, mean mortalities followed by the same letter are not significantly different at P > 0.05 (Tukey’s HSD tests). Significant difference is depicited by an asterisk (*) between the observed mortality and the expected additive mortality (EAM) of each mixture at P < 0.05 (Chi-squared test). Fla = flavone, NS = No significance.
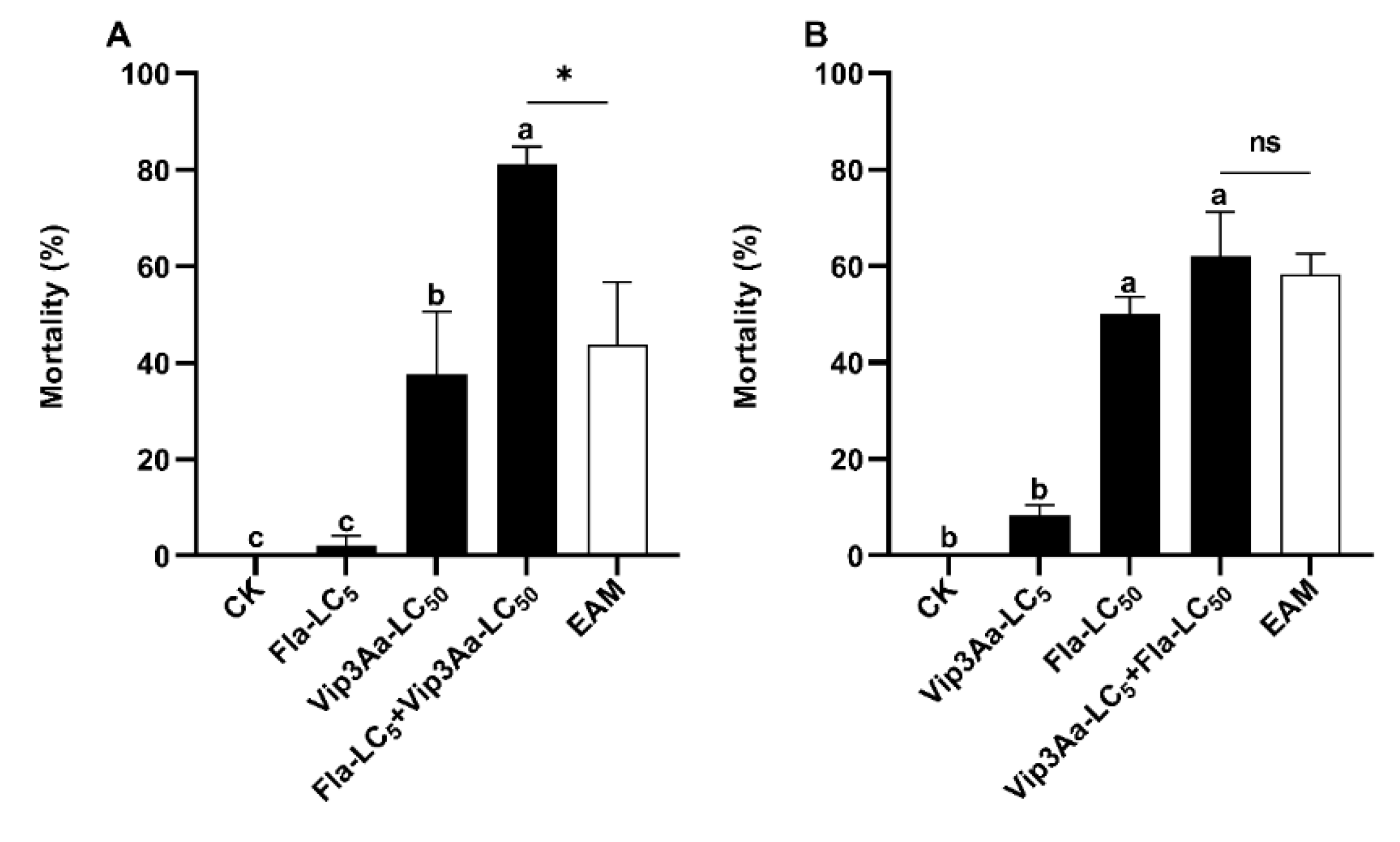
Figure 3.
Induced toxicity of Vip3A LC5 + emamectin LC50 (A) and emamectin LC5 +Vip3A LC50 (B) against S. frugiperda neonates. Untransformed average mortality ± SE are presentated. Mortality values were arcsine converted prior to analysis. Within each small graph, mean mortalities followed by the same letter are not significantly different at P > 0.05 (Tukey’s HSD tests). Extremely significant difference is indicated by two asterisks (**) between the observed mortality and the expected additive mortality (EAM) of each mixture at P < 0.05 (Chi-squared test). EMB = emamectin benzoate, NS = No Significance.
Figure 3.
Induced toxicity of Vip3A LC5 + emamectin LC50 (A) and emamectin LC5 +Vip3A LC50 (B) against S. frugiperda neonates. Untransformed average mortality ± SE are presentated. Mortality values were arcsine converted prior to analysis. Within each small graph, mean mortalities followed by the same letter are not significantly different at P > 0.05 (Tukey’s HSD tests). Extremely significant difference is indicated by two asterisks (**) between the observed mortality and the expected additive mortality (EAM) of each mixture at P < 0.05 (Chi-squared test). EMB = emamectin benzoate, NS = No Significance.
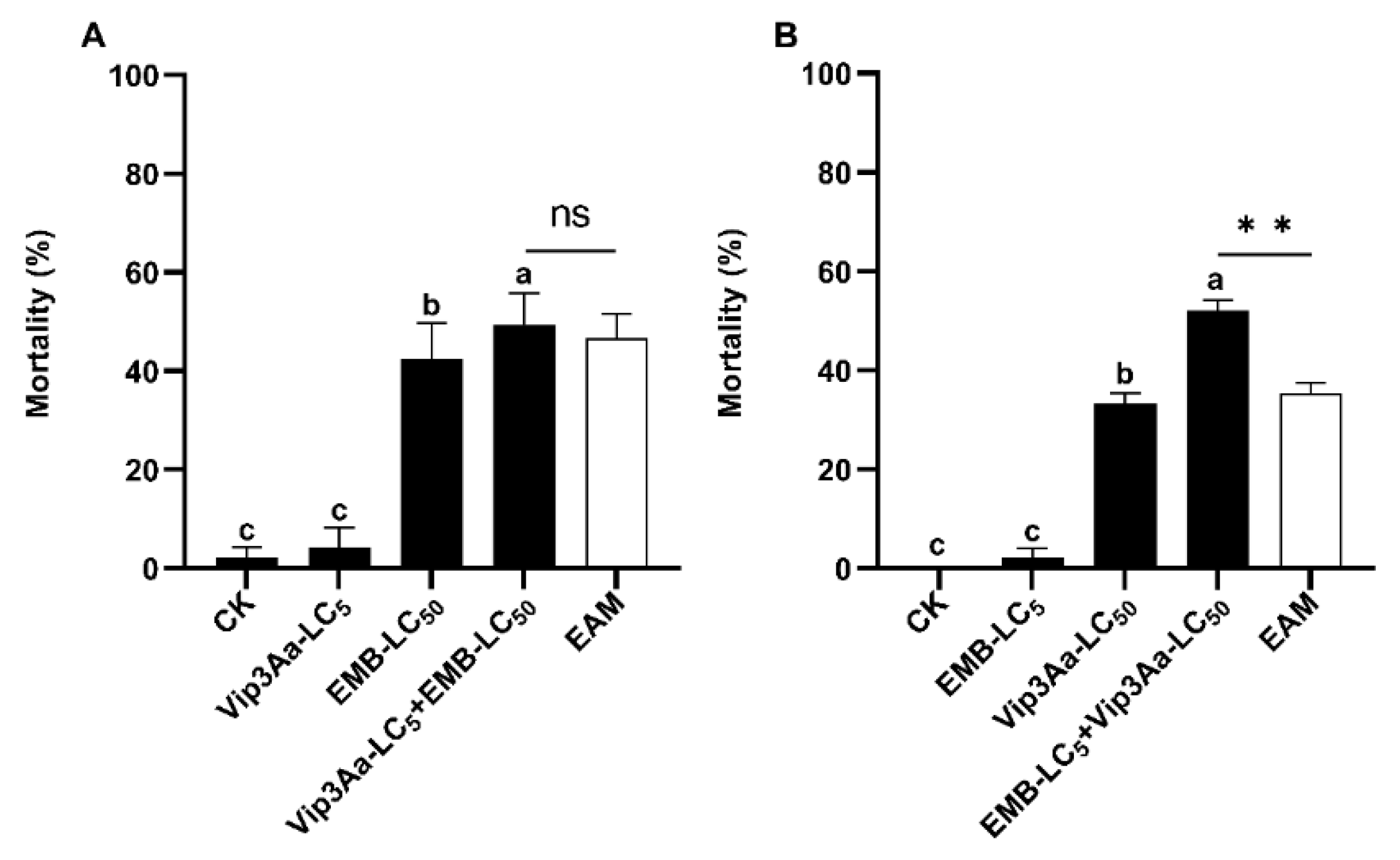
Figure 4.
Induced toxicity of flavone LC5 + emamectin LC50 (A) and emamectin LC5 + flavone LC50 (B) against S. frugiperda neonates. Untransformed average mortality ± SE are presentated. Mortality data were arcsine converted prior to analysis. Within each small graph, mean mortalities followed by the same letter are not significantly different at P > 0.05 (Tukey’s HSD tests). Significant difference is depicted by one asterisk (*) between the observed mortality and the expected additive mortality (EAM) of each mixture at P < 0.05 (Chi-squared test). Fla = flavone, EMB = emamectin benzoate, NS = No Significance.
Figure 4.
Induced toxicity of flavone LC5 + emamectin LC50 (A) and emamectin LC5 + flavone LC50 (B) against S. frugiperda neonates. Untransformed average mortality ± SE are presentated. Mortality data were arcsine converted prior to analysis. Within each small graph, mean mortalities followed by the same letter are not significantly different at P > 0.05 (Tukey’s HSD tests). Significant difference is depicted by one asterisk (*) between the observed mortality and the expected additive mortality (EAM) of each mixture at P < 0.05 (Chi-squared test). Fla = flavone, EMB = emamectin benzoate, NS = No Significance.
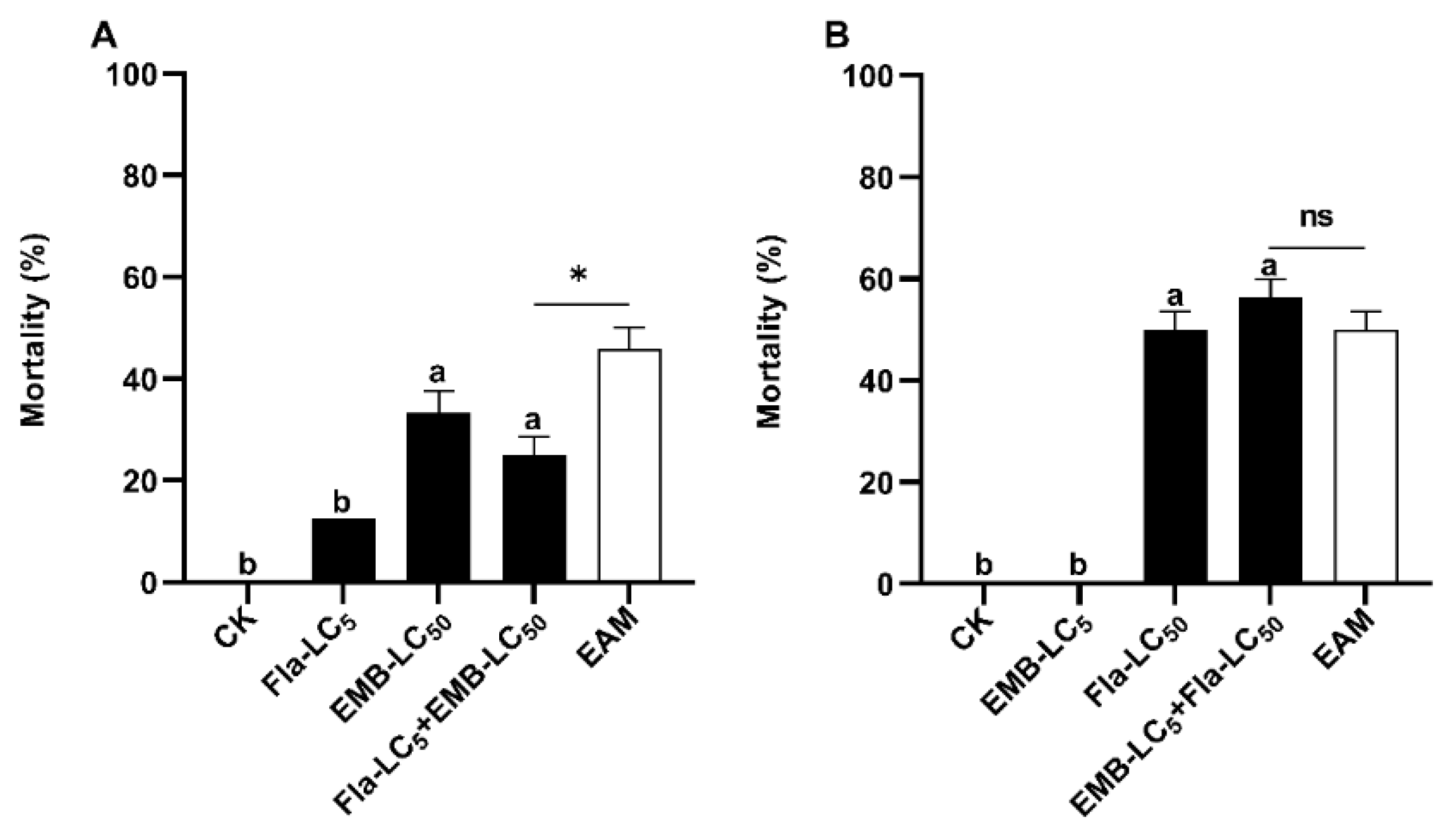

Table 1.
Baseline toxicity of Vip3Aa, flavone, and emamectin benzoate against S. frugiperda neonate larvae.
Table 1.
Baseline toxicity of Vip3Aa, flavone, and emamectin benzoate against S. frugiperda neonate larvae.
| Toxins | Na | LD-P line | LC5 (95% CLb) | LC25 (95% CLb) | LC50 (95% CLb) | R2c |
|---|---|---|---|---|---|---|
| Vipa3A | 285 | Y=1.97X + 1.56 | 24d (1-50) | 73 (19-130) | 161 (90-370) | 0.89 |
| Flavone | 275 | Y=3.05X 8.27 | 147.95e (27.0-244.6) | 307.5 (142.1-348.1) | 511.1 (354.6-801.8) | 0.91 |
| Emamectin benzoate | 285 | Y=3.50X + 2.99 | 0.047d (0.03-0.06) | 0.09 (0.074-0.1) | 0.14 (0.12-0.16) | 0.96 |
a-Number of neonates tested. b-95% confidence limits. c-Correlation coefficient. d-ng/cm2 for Vip3Aa and Emamectin benzoate. e-ug/g diets for flavone.
Table 2.
Co-toxicity factors of concurrent exposure of S. frugiperda neonates to emamectin benzoate, Vip3Aa, and flavone.
Table 2.
Co-toxicity factors of concurrent exposure of S. frugiperda neonates to emamectin benzoate, Vip3Aa, and flavone.
| Exposure to | Observed mortality (%) | EAM* (%) | Co-toxicity factor | Interaction | |
|---|---|---|---|---|---|
| Concurrent Vip3Aa_LC25 + Fla_LC25 |
Vip3Aa | 35.42 ± 2.08a | |||
| Flab | 25.00 ± 3.61 | ||||
| Vip3Aa + Fla | 81.25 ± 3.61 | 60.42 | +20 < 34.48 | Synergism | |
| Concurrent Vip3Aa_LC25 + EMB_LC25 |
Vip3Aa | 43.61 ± 3.06 | |||
| EMBc | 31.65 ± 7.01 | ||||
| Vip3Aa + EMB | 95.83 ± 2.08 | 75.26 | +20 < 27.34 | Synergism | |
| Concurrent Fla_LC25 + EMB_LC25 |
Fla | 25.00 ± 3.61 | |||
| EMB | 41.67 ± 4.17 | ||||
| Vip3Aa + EMB | 70.83 ± 4.17 | 66.67 | +20 > 6.25 | Addition | |
| Concurrent Fla_LC15 + EMB_LC15 + Vip3Aa_LC15 |
Fla | 14.58 ± 4.17 | |||
| EMB | 19.17 ± 3.63 | ||||
| Vip3Aa | 29.17 ± 5.51 | ||||
| Fla + EMB + Vip3Aa | 87.50 ± 0.00 | 62.92 | +20 < 40.00 | Synergism | |
a-Standard error of the means. b-Flavone. c-Emamectin benzoate. *EAM= expected additive mortality.
Table 3.
Co-toxicity factors of in sequence exposure of S. frugiperda neonates to emamectin benzoate, Vip3Aa and flavone.
Table 3.
Co-toxicity factors of in sequence exposure of S. frugiperda neonates to emamectin benzoate, Vip3Aa and flavone.
| Exposure to | Observed mortality (%) | EAM* (%) | Co-toxicity factor | Interaction | |
|---|---|---|---|---|---|
| Sequential Vip3Aa_LC5 + Fla_LC50 |
Vip3Aa | 8.33 ± 2.08a | |||
| Fla | 50.00 ± 3.61 | ||||
| Vip3Aa+Flab | 62.08 ± 9.14 | 58.33 | -20 < 6.43 < +20 | Addition | |
| Sequential Fla_LC5 + Vip3Aa_LC50 | Fla | 6.25 ± 0.00 | |||
| Vip3Aa | 37.50 ± 13.01 | ||||
| Fla + Vip3Aa | 81.25 ± 3.61 | 43.75 | +20 < 85.71 | Induction | |
| Sequential Fla_LC5+EMB_LC50 |
Fla | 12.5 ± 0.00 | |||
| EMBc | 33.33 ± 4.17 | ||||
| Fla + EMB | 25.00 ± 3.61 | 45.83 | -45.45 < -20 | Inhibition | |
| Sequential EMB_LC5+Fla_LC50 |
EMB | 0.00 ± 0.00 | |||
| Fla | 50.00 ± 3.61 | ||||
| EMB + Fla | 56.25 ± 3.61 | 50.00 | -20 < 12.50 < +20 | Addition | |
| Sequential Vip3Aa_LC5+EMB_LC50 |
Vip3Aa | 4.17 ± 4.17 | |||
| EMB | 42.50 ± 7.32 | ||||
| Vip3Aa + EMB | 49.40 ± 6.37 | 46.67 | -20 < 5.86 < +20 | Addition | |
| Sequential EMB_LC5+Vip3Aa_LC50 |
EMB | 2.08 ± 2.08 | |||
| Vip3Aa | 33.33 ± 2.08 | ||||
| EMB+Vip3Aa | 52.08 ± 2.08 | 35.42 | +20 < 47.06 | Induction | |
a-Standard error of the means; b-Flavone; c-Emamectin benzoate; *EAM= expected additive mortality.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



