
Submitted:
28 June 2023
Posted:
29 June 2023
You are already at the latest version
Abstract
The Oxford English Dictionary includes 17 definitions for the word “model” as a noun and other 11 as a verb. Therefore, context is necessary to understand the meaning of the word model. For in-stance, “model railways” refer to replicas of railways and trains at a smaller scale and a “model student” refers to an exemplar individual. In some cases, a specific context, like cancer research, may not be sufficient to provide one specific meaning of a model. Even if the context is narrowed, specifically to the research related to the tumour microenvironment, a “model” can be understood in a wide variety of ways, from an animal model, to a mathematical expression. This paper presents a review of the different “models” of the tumour microenvironment grouped by the different def-initions of the word into four categories: model organisms, in vitro models, mathematical models, and computational models. Then, the frequency of different meanings of the word “model” related to the tumour microenvironment is measured from the number of entries in the MEDLINE database of the United States National Library of Medicine at the National Institutes of Health. The frequency of the main components of the microenvironment and the organ-related cancers modelled are also assessed quantitatively with specific keywords. Whilst animal models, in particular xenografts and mouse models are the most common used “models”, the number of entries have been slowly de-creasing. Mathematical models, prognostic and risk models follow in frequency and these have been growing in use.
Keywords:
Tumour microenvironment
; In vivo model
; In vitro model
; Mathematical model
; Computational model
1. Introduction
It is now widely accepted that Cancer research cannot solely rely on the study of individual cancer cells, or the tumour in isolation [1] but rather on the collection of many different cells and their interactions in what is known as the tumour microenvironment [2]. The complex relationship of cancerous cells with healthy cells, immune cells, vasculature, extracellular matrix, molecules, and other elements that surround and interact with the cancer cells is crucial in the development of a tumour and the response to treatments [3,4]. The idea of the interaction has been traced back to Stephen Paget who in 1889 proposed a “seed and soil” theory [5], where the cancer cells were the seeds that interacted with the organism, the soil, in the spread of tumour cells [6]. Research into the tumour microenvironment thus tries to elucidate the mechanisms by which elements such as infiltrating cells [7,8,9], soluble factors [10,11,12], extracellular matrix [13,14,15] or the tumour vasculature [16,17] interact with one another [18]. At the time of writing (April 2023), PubMed, the search engine of the United States National Library of Medicine (NLM) database MEDLINE, returned more than 81,000 entries for the keywords “tumour microenvironment” (https://pubmed.ncbi.nlm.nih.gov/?term=tumour+microenvironment) and slightly fewer at 78,714 for the same query with tumor instead of tumour (https://pubmed.ncbi.nlm.nih.gov/?term=tumor+microenvironment). Most of these entries have been indexed after 2000.
The direct study of the tumour microenvironment in patients is restricted by the number of patients that are willing to participate in trials, repeated observations are limited, and the use of new drugs is constrained and would have to have gone through a series of pre-clinical studies. Histopathology is a useful tool of study of the microenvironment, but still has some technical limitations, i.e., it is not possible to visualise blood flow. Therefore, a plethora of indirect methods that overcome these limitations have been developed.
These methods rely on a “model” that on one hand simplifies the study of the microenvironment, and on the other hand, resembles it closely, so that any findings can be successfully translated to a patient in a clinical environment. Unfortunately, the concept of “model” is not universally understood. Different disciplines refer to a model in distinct ways, which are related to the many definitions of the word itself. The Oxford English Dictionary [19] (https://www.oed.com/search?searchType=dictionary&q=model) includes 17 definitions for the word “model” as a noun and other 11 as a verb varying from “Something which accurately resembles or represents something else” to “A three-dimensional representation”.
It is therefore not surprising that there are many different “models” of the tumour microenvironment. The rest of this work will explore some of these definitions as they are understood and used by those who research the tumour microenvironment from different perspectives and will assess the frequency of use of several keywords related to the tumour microenvironment.
2. Different concepts of Model
2.1. Model:
“An animal or plant to which another bears a mimetic resemblance”
Perhaps the most widely used concept of a “model” is that related to a model organism; a non-human species used to perform experiments that can reveal some understanding of a biological phenomenon [19]. From simple organisms like the bacteria Escherichia Coli [20] or yeast like Saccharomyces cerevisiae [21] to zebrafish [22], rodents [23], or drosophila [24] model organisms have been extensively used to elucidate anything from aging [25] to Zika [26]. Part of the success of model organisms has been the fact that the operating principles of some cellular processes like the cell cycle or signalling pathways are similar in humans and other species that branched out from earlier common ancestors [27]. Rodents have taken a predominant place as a model organism in cancer and other conditions due to several factors: ease of maintenance and transport, high fertility rates, relative low costs, and ease of genetic modifications [19]. Specific mouse models can now be used to study perimenopausal depression [28], tuberculosis [29], myocardial infarction[30] and the genetically engineered mouse is considered by some to be the preferred organism used in cancer studies [31,32]. Cancer can be induced in these models by the administration of a carcinogen [33,34], diet-induced [35,36], by transplantation of tissue or cells from patients or cell lines into the model, i.e., xenografts [37,38]. Alternatively, in transgenic animals that have been genetically modified, cancer can occur spontaneously [23,39]. As this type of model is a whole living organism, it is expected that they will intrinsically “capture the intricacies of the tumor immune response and microenvironment” [40]. This on its own is one the most important advantages of model organisms, which do not need the design of an environment to model the tumour microenvironment. The organism itself provide the microenvironment from which aspects like therapeutic implication or side effects can be observed [41]. However, there are important shortcomings as the host organism is a different species than the donor, and there may be a species-mismatch between the tumour and the host microenvironments [32,42]. The reliability of the translation from animal models to human diseases therefore remains controversial [43,44]. The model then bears a resemblance to the microenvironment of a human cancer, but it is not exactly the same.
The tumour microenvironment of the models can be observed through histopathology [45,46,47] and immunohistochemistry [45,48,49], in which tissue is extracted, thinly sliced, and stained with different techniques highlighting important components of the tumour microenvironment such as macrophages and lymphocytes. An important limitation of histopathology is that there is only one time point of observation. When techniques such as dorsal skin fold window chambers [50] are used, the development of the tumour and its microenvironment can be directly observed through intravital imaging techniques [32,51], which allow the repeated observation, and the possible effect of treatments [52,53], for a period of time. Alternatively, tissue can be observed using magnetic resonance imaging [54,55] or positron emission tomography [56,57], which is less invasive, but has much lower resolution than microscopical techniques.
2.2. Model:
To serve or behave as the analogue of (a phenomenon, system, etc.). Or
A three-dimensional representation… esp. one showing the component parts in accurate proportion and relative disposition. Or
To produce (a figure, likeness, etc.) by moulding, carving, etc., esp. in clay, wax, or some other malleable material
Another popular concept of model related to cancer is that of “in vitro” or “in glass” experiments. These models refer to investigations performed with cells, organisms, or parts of organisms in Petri dishes or similar equipment and have been used for a long time in cancer-related experiments such as cell growth [58] and screening of antitumour substances [59]. These experiments imply artificial conditions and a significant simplification of the microenvironment of a tumour. Conversely, these models offer a number of advantages over in vivo experiments with model organisms, not least the avoidance of animal testing. Advantages of in vitro experiments include lower costs, higher throughput, can be considered more amenable to mechanistic analysis [40]. Also, despite the considerable simplification of the environment, the models can have higher human relevance since cancer cells derived from a primary patient material can be directly used [60,61]. In vitro models have been considered to have fewer problems of how valid the result on one species is applicable to another species [62]. On the other hand, the in vitro models are limited as compared with the animal models in the complexity they can offer. There is no physiological response, and it is more difficult to observe side effects.
A simple setting to mimic the tumour microenvironment is to co-culture cancer cells with cells of the tumour microenvironment like myofibroblasts [63], cancer associated fibroblasts [64], endothelial cells [65] or stromal cell types and/or extracellular matrix [66]. The co-cultures can then be used to perform a wide variety of experiments related to cell proliferation [67], migration [68,69], invasion [70], or treatment and drug combinations [71,72]. Despite the simplicity of these experiments, the inherent 2D nature of the cultures is a major limitation as the interactions between cells and the environment do not resemble the 3D nature of tumour and its microenvironment [73,74]. Accordingly, 3D in vitro models of the tumour microenvironment have evolved significantly, for instance in breast cancer [75], and now include multicellular aggregates like spheroids [76,77] or organoids [78,79], which are maintained in different settings such as purified extracellular matrix gels, hanging drop cultures, 3D Gels or 3D scaffolds [80] of meshes or sponges, which offer great number of conditions such as porosity, biodegradability, chemical composition, transparency, etc. [74]. A further complexity can be introduced to in vitro models by allowing external interaction, thus simulating metabolic processes [81] or providing complex geometries such as branching structures that mimic the vasculature of a tumour [82]. These models are known by different names: 3D bioprinted, microfluidic, tumour-on-a-chip or organ-on-a-chip [83,84,85,86,87,88]. One of the major advantages of these models over animal models is the observation, as it the settings themselves are easy to examine in microscopes or other settings.
2.3 Model:
A simplified or idealized description or conception of a particular system, situation, or process, often in mathematical terms.
A mathematical model can be understood as the simplification and abstraction of a complex phenomenon and its subsequent description in mathematical equations. A model should tackle one or more biological or clinical hypotheses and analyse experimental data together with the formulation of the mathematical description, i.e., the model itself, and will go through a cycle of refinements until it can be validated [89,90].
A classic example of a mathematical model is the Malthusian growth model [91] that assumes that a population (P0) will grow in time (t) in an exponential way depending on the growth rate (r) following the equation P(t) = P0 ert. This model is similar to the cancer initiation model proposed by Armitage and Doll [92] describing the incidence rate (I) of a cancer at age t, as I(t) = k tn where k is a constant and n is the number of stages (or mutations) that must be passed for a cell to become malignant. These two models are descriptive models, i.e., they describe the broad characteristics of the phenomenon or could be used to predict or prognosticate a future state. When the description refers to the time of occurrence of an event being modelled, the process is sometimes called a survival analysis [93]. If the model takes into account one factor (say time), but ignores other factors (say ethnic group, age, lifestyle) the model is considered univariate [94]. Multivariate statistical models [95] on the other hand, consider several variables at the same time. For instance, the correlation between overall survival of patients with non-small cell lung cancer with the concentration of amino acids and metabolites measured from blood samples [96].
Alternative to descriptive models are those considered mechanistic or conceptual [97], which attempt to explain the processes that drive the phenomena [98], and from which it is possible to derive biologically important characteristics of a tumour. For instance, that distal recurrence of glioblastoma depends on hypoxic microenvironment and migration and proliferation rates of the tumour cells [99].
Models that provide the same results every time are considered deterministic, and those which include a certain randomness in the process are considered stochastic [97]. Stochastic models of the tumour microenvironment [100,101,102] are more common than deterministic ones [103] by an approximate ratio of 10 to 1, which is probably a reflection that many factors related to cancer, like somatic evolution, are not deterministic [104].
The scale, or point of view of the model, provides different resolutions at which the models operate: at organ scale, they are considered macroscale models [105], at tissue scale they are considered mesoscale models [106], at cell level they are considered microscale models [107]. Some authors stress the importance of incorporating cellular models into whole-organ models [108]. The nature of the tumour microenvironment can be studied at different scales, thus many models are considered “multiscale” [109,110,111,112,113,114] as they consider from molecules to cells to tissue-level phenomena [115,116], how the extracellular matrix is altered [117,118] or an avascular tumour growth and a cell model [119]. It is important to consider that any model should be able to reproduce data that has been observed through experiments [120] and as such, models at different scales, will require validation at different scales as well [121].
An interesting perspective to formulate models is to consider the cell as a basic unit, i.e., a virtual cell [122,123] with a set of rules for behaviour. The unit is sometimes called an “agent”, with rules to proliferate, reproduce or transform depending on interactions with its external microenvironment [110] and probabilistic rules [124]. Different types of cells; tumour, immune, dendritic constitute different agents [125]. Since these approaches build the study from single-cells, they are considered "bottom-up” [126]. “Top-down” approaches on the other hand, zoom out and focus on whole organs, or consider cells as a group or population. The behaviour is considered as a mean of all the cells and not as individuals [121]. It is possible of course, to start not at the top or the bottom, but rather somewhere in between with “middle-out” models [127,128,129]. A middle-out model is useful in cases where there is a rich level of biological data than can be used as a starting point from which to reach up and down [108], or when the phenomena to be modelled is itself in the mesoscale, like the microcirculation [130].
In an alternative approach, these cells, whether cancer or healthy, can be considered as species that strive for survival, and treat cancer as a problem of ecology and evolution [131,132,133] and consider sub-populations within a single cancer [134]. An example of this approach is a branching process [135,136], in which as time passes, a cell may divide, die, or mutate at certain rates. After a number of cycles, mutations may accumulate in the population of cells. From a simple formulation like this one, it is possible then to significantly increase complexity by adding different types of cells, i.e., cells of the immune system [137]. As such, models have now been proposed for migration [138], tumour growth [139], invasion [140], angiogenesis [141,142], treatment and recurrence [143], cancer cell intravasation [144], fluid transport in vascularised tumours [145], macrophage infiltration [146], response to radiotherapy [147], optimisation of chemotherapy [148]. For reviews into mathematical modelling of cancer, the reader is referred to [89,90,98].
As many of the previously mentioned approaches require computer simulations, these models are sometimes called in silico models or computational models. Some mathematical models are purely mathematical, like the Armitage and Doll, which does not require simulations or computations but merely applies an equation. However, many mathematical models apply numerical methods and are intrinsically computational [149]. Some authors [150] distinguish mathematical models when they use a continuous model using mathematical equations from computational models which are discrete based on a series of steps or instructions. Still, in many cases, distinctions between mathematical and computational are not considered and some authors use the terms “mathematical model” and “computational model” interchangeably [151] and others consider a model itself to be both mathematical and computational [97,152,153,154,155,156]. For more information about mathematical and computational models of tumour microenvironment and cancer, the reader is referred to [97,111,121,157,158].
Mathematical and computational models include numerous advantages: no need of animals or tissues, lower costs, rapidity at which simulations can be generated, However, the limitations are numerous, not least the inherent simplicity of any mathematical model as compared with a living organism, a complex disease like cancer and a complex setting like the tumour microenvironment.
2.4 Model:
To devise a (usually mathematical) model or simplified description of (a phenomenon, system, etc.)
Despite the close relationship between the mathematical and computational approaches, there are different methodologies that are fundamentally computational. In these cases, computational methods are applied to process, analyse and extract information from datasets. As opposed to a “model” that describes the growth of a tumour, these methods could for instance count something [159] or measure colour [160]. What is modelled is not the cells or the cancer itself, but rather derived features, like the shape of a cell or a vessel [72] or movement of cells or its fluorescent intensity. There does not need to exist an underlying mathematical abstraction of cancer or a biological process in the methodologies, but the information extracted will relate to conditions of the cancer, like the cellularity [161].
Computational methods that belong to areas of computer vision, image processing, machine learning or more recently deep learning, can be applied. Features related to important characteristics, like number of nuclei identification [162] or microvessel density [163] can be extracted. Surely, these computational methods can extract features or quantities that can be then used to inform mathematical models. For instance, to estimate the vascular permeability [53] in tumours, the fluorescence intensity can be acquired, then, through image processing techniques, the vasculature can be segmented, the intensity inside and outside the vessels calculated and these quantities fed to the Patlak Model [164] to model the blood extravasation. The effect of vascular disrupting agents on tumours can be assessed from the velocity of red blood cells travelling inside the tumour and a model of movement can be applied to measure the velocity of the cells [165]. The spatial heterogeneity in the tumour microenvironment [166] can be assessed by identifying and mapping cells from histological samples, and then ecological models can be used with the information extracted.
To complicate matters, another quite different computational type of model has been gaining popularity. Namely, those models associated with the areas of artificial intelligence, artificial neural networks, and deep learning. These models have been inspired by neurobiology and the simplification of a neurone as a unit with many input signals, which are weighted, i.e., multiplied by individual values, and then combined (i.e., summed) to produce a single or multiple output value. This model is known as the McCulloch–Pitts model of a neuron [167,168]. Many neurones, sometimes also called nodes or units, with this and many other functions, are then combined into layers with a specific structure, sometimes called an architecture. With time and the increase in computer power, these models of artificial neural networks have been increasing in complexity adding more and more layers with millions of neurones to the architecture, thus gaining the name “deep”. One key difference is that, unlike other mathematical or computational models, in which fine-tuning of the parameters is performed manually by a person (hand-crafted), these have a huge number of parameters that self-tune when presented with a large amount of training data, i.e., raw data, like an image, coupled with class labels that indicate what is where. This process through which the parameters of the architecture adapt is called “learning” and the area in general is known as machine learning and in particular deep learning for the larger architectures. Thus, a specific model can be equally used to analyse images of cats and dogs or images of the tumour microenvironment depending on the training data that has been provided. Sometimes the arrangement of the basic blocks or structure is called an architecture and once it has been specifically trained for a task it is called a model, but as in other cases, architecture and model are used interchangeably. The models are normally known by short acronyms like CNN (for convolutional neural network), VGG [169] (after the Visual Geometry Group at Oxford University) sometimes followed by numbers associated to the number of layers of the architecture like VGG16, AlexNet [170] (after the name of the designer of the architecture Alex Krizhevsky), U-Net [171] (after the shape of the architecture like a letter U), or GoogLeNet [172] (after the affiliation of some of the authors where the architecture was introduced). For introductory reviews to deep learning, the reader is referred to [173] and neural networks and deep learning for biologists to [174]. For more specific reviews on deep learning applied to cancer and histopathology, the reader is referred to [175,176,177,178,179,180]. The following paragraph illustrate with a few examples how deep learning models are applied.
The differences between breast stromal microenvironment and benign biopsies in haematoxylin and eosin (H&E) slides were distinguished by a VGG model [181]. The model was then used in a different dataset to detect a higher amount of tumour associated stroma in ductal carcinoma in situ in grade 3 compared with grade 1. Cancer grading was calculated from prostate cancer H&E slides with a combination of several CNNs that perform detection and classification and the tissue with a posterior slide-level analysis which provided a Gleason grade [182]. Patient survival was predicted from colorectal histology slides [183] by applying a VGG19 model for the classification of the slides into a series of classes (adipose, background, debris, lymphocytes, mucus, smooth, etc.), from which a combination of values was used to create a “deep stromal score” with considerable prognostic power, especially for advanced cancer stages. In another study [184], patient survival is predicted from a score (tumour associated stroma infiltrating lymphocytes TASIL-Score) which is calculated from the spatial co-occurrence statistics (stroma-stroma, stroma-lymphocyte, etc.) that are extracted using a DenseNet model [185] to segment each class in head and neck squamous cell carcinoma H&E slides.
3. Quantitative evaluation of the presence of different models in Medline
To assess the distribution of the different definitions of the word model as related to the tumour microenvironment, a quantitative and unbiased analysis was performed. The analysis mined the MEDLINE database of the United States National Library of Medicine at the National Institutes of Health. Mining was performed using the search engine PubMed through a series of queries with combinations of keywords and basic terms as has been previously described [186]. The basic terms were the search URL of PubMed ('https://www.ncbi.nlm.nih.gov/pubmed/?term=') and tumour microenvironment in British and American spelling (("tumor microenvironment") OR ("tumour microenvironment")) and “cancer microenvironment” was also included with an OR. Dates were restricted to 2000-2023 (2000:2023[dp]). The keywords were manually curated based on the previously described definitions of the word model and are shown in Table 1. The concatenation was performed sequentially with one keyword at a time.
The following caveats should be considered when observing the results. A single entry could be retrieved more than once, e.g., “Imaging interactions between macrophages and tumour cells that are involved in metastasis in vivo and in vitro” will be counted for both in vivo and in vitro. Similarly, the same type of model could be referred to with two different keywords like mouse and mice. The entries are mined if the keyword appears in the PubMed record, which includes title, abstract and Mesh terms. That is, if the keywords only appear in the main text of a paper, it will not be retrieved. Furthermore, it is very important to note that the term tumour/tumor could include benign tumours and thus the results are not restricted to cancer.
The total number of entries in PubMed for each of the keywords is shown in Figure 1 as a bar chart. Colours have been used to group each keyword according to the definitions of model. In Figure 2, the entries have been aggregated into four groups and are shown per year as ribbons with the same colours as in Figure 1.
The first observation is that the most frequent entries for the tumour microenvironment are those related to animal models, far more than the in vitro models. Since the scale of the vertical axis is logarithmic, xenograft and mouse are an order of magnitude above most other keywords. These are followed by mathematical keywords prognostic and risk. Despite the simplicity of in vitro models and the perceived lack of human relevance of animal models, these latter ones dominate the research on tumour microenvironment. However, the temporal trends shown in Figure 2 show that there is a slight decrease of the number of entries related to animal models in the last 3-4 years. Furthermore, whilst the term mathematical model appears much more recently than the model organism or in vitro model, the growth is faster and is overtaking both, especially in the past 5 years. The term computational model appear later but also shows an increasing trend, although not as high as for mathematical model. It will be interesting to observe these trends in future years.
Next, to identify important components of the microenvironment and their frequency of appearance in PubMed, 39 keywords related to the microenvironment (e.g., T cells, endothelial cells, B cells, invasion, metastasis, inflammation, cytokine, pathways, etc.) were added to the queries (e.g., ("cytokine") AND (("tumor microenvironment") OR ("tumour microenvironment") OR (“cancer microenvironment”) AND (model) AND (2000:2023[dp])). Figure 3 shows frequency of appearance of the keywords in decreasing order, starting with Metabolism, Therapeutic, Survival and decreasing towards Adipocytes, Pre-metastatic niche and Extracellular vesicle. Again, besides the caveats previously mentioned, it should be taken into consideration that this figure indicates only how frequently the terms appear in the query. For instance, the frequency of the term neutrophils is one order of magnitude lower than the term macrophages. Still, the most common term and possibly the related research question is to investigate the metabolism of the tumour microenvironment.
Organ-specific keywords were used to investigate what are the most frequently modelled microenvironments and the results are shown in Figure 4. Breast and lung were the most common terms, followed by liver, melanoma and bone with very similar number of entries. These numbers partially correlate with the incidence and mortality rates of cancer. Worldwide, the most common cancers by incidence are breast, lung, colorectal, prostate and stomach, and by mortality are lung, colorectal, liver, stomach and breast [187]. Proportionally, melanoma, bone and brain are more common in research entries in PubMed than their corresponding incidence and mortality rates. At the bottom of the list are bowel, leukaemia, pituitary, testicular and uterus. It is interesting that some related terms like colorectal/colon can have similar number of entries whilst others like uterine/uterus can be orders of magnitude different.
A combination of keywords for the models and the organs as pairs (e.g., “(Xenograft) AND (brain)” is added to the query) is displayed in Figure 5 and a zoom into the results with most results in Figure 6. These figures show that the most common combination is Xenograft with Breast with 479 entries, followed closely by Mouse Model-Breast (398), Mouse Model-Lung (392) and Xenograft-Lung (388). The most frequent entries when using Prognostic or Risk models are Lung, Liver, Breast, Cervical and Colorectal. For in vitro, the most frequent entries are Breast, Brain, Lung and Bone.
4. Conclusions
Whilst the most common setting to investigate the tumour microenvironment are model organisms, recent years have shown a slight decrease in the number of entries in PubMed. In vitro models also show growth with a slowdown in the last 2 years of the analysis here presented. On the other hand, the number of entries using mathematical models has been growing steadily and are now as common as the number of entries for in vivo models. The use of computational models has also been growing, especially agent-based models and convolutional neural networks. It will be interesting to see how these trends continue in the near future.
The basic idea behind the models here described is these constitute a simplified, idealised and more accessible representation of something more complex and hard to observe, in this case the tumour microenvironment. Whilst it should always be well understood that no model is a perfect representation of reality, a good model should capture some essential characteristics of the microenvironment and permit successful experimentation from which observations can be translated to patient treatment or care. It should always be considered that not everyone understands models in the same way, thus it is important to make an effort to use these terms in ways that avoid confusion if possible. For instance, when talking about deep learning, the term “architecture” could be used instead of models. Adding the word “organism” in cases of animal models could also help, e.g., “the mouse has become the favorite mammalian model organism”. Similarly, in mathematical cases, the specification of a risk-model or a mechanistic-model and not just a model would improve clarity. Biologically and clinically, there are still many unanswered questions related to the tumour microenvironment and all its components. Interdisciplinary research related to the microenvironment is growing and as such, a single study may include, say, in vitro models that are then processed with deep learning models, or histopathology slides that are analysed with machine learning models that and then feed a prognostic model of survival. Therefore, a clear understanding of what is meant each time that the word “model” appears in a paper is necessary and researchers from all sides of the spectrum should bear in mind that not everyone understands the same by “model”.
Author Contributions
Conceptualization, C.C.R.-A.; methodology, C.C.R.-A.; software, C.C.R.-A.; investigation, C.C.R.-A.; data curation, C.C.R.-A.; writing—original draft preparation, C.C.R.-A.; writing—review and editing, C.C.R.-A. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data was mined from the PubMed with scripts written in Matlab® (The MathworksTM, Natick, USA), and available in https://github.com/reyesaldasoro/TumourMicroenvironmentModels.
Conflicts of Interest
The authors declare no conflict of interest.
References
- L. Laplane, D. Duluc, A. Bikfalvi, N. Larmonier, and T. Pradeu, “Beyond the tumour microenvironment,” International Journal of Cancer, vol. 145, no. 10, pp. 2611–2618, 2019. [CrossRef]
- F. R. Balkwill, M. Capasso, and T. Hagemann, “The tumor microenvironment at a glance,” Journal of Cell Science, vol. 125, no. 23, pp. 5591–5596, Dec. 2012. [CrossRef]
- N. M. Anderson and M. C. Simon, “The tumor microenvironment,” Current Biology, vol. 30, no. 16, pp. R921–R925, Aug. 2020. [CrossRef]
- Y. Xiao and D. Yu, “Tumor microenvironment as a therapeutic target in cancer,” Pharmacol Ther, vol. 221, p. 107753, May 2021. [CrossRef]
- S. Paget, “THE DISTRIBUTION OF SECONDARY GROWTHS IN CANCER OF THE BREAST.,” The Lancet, vol. 133, no. 3421, pp. 571–573, Mar. 1889. [CrossRef]
- M. Akhtar, A. Haider, S. Rashid, and A. D. M. H. Al-Nabet, “Paget’s ‘Seed and Soil’ Theory of Cancer Metastasis: An Idea Whose Time has Come,” Advances in Anatomic Pathology, vol. 26, no. 1, p. 69, Jan. 2019. [CrossRef]
- X. Li et al., “Targeting of tumour-infiltrating macrophages via CCL2/CCR2 signalling as a therapeutic strategy against hepatocellular carcinoma,” Gut, vol. 66, no. 1, pp. 157–167, Jan. 2017. [CrossRef]
- R. García-Marín et al., “CD8+ Tumour-Infiltrating Lymphocytes and Tumour Microenvironment Immune Types as Biomarkers for Immunotherapy in Sinonasal Intestinal-Type Adenocarcinoma,” Vaccines (Basel), vol. 8, no. 2, p. 202, Apr. 2020. [CrossRef]
- M. a. C. Versluis et al., “The prognostic benefit of tumour-infiltrating Natural Killer cells in endometrial cancer is dependent on concurrent overexpression of Human Leucocyte Antigen-E in the tumour microenvironment,” Eur J Cancer, vol. 86, pp. 285–295, Nov. 2017. [CrossRef]
- H. Ahmed, A. Ghoshal, S. Jones, I. Ellis, and M. Islam, “Head and Neck Cancer Metastasis and the Effect of the Local Soluble Factors, from the Microenvironment, on Signalling Pathways: Is It All about the Akt?,” Cancers (Basel), vol. 12, no. 8, p. 2093, Jul. 2020. [CrossRef]
- M. Akimoto, R. Maruyama, H. Takamaru, T. Ochiya, and K. Takenaga, “Soluble IL-33 receptor sST2 inhibits colorectal cancer malignant growth by modifying the tumour microenvironment,” Nat Commun, vol. 7, p. 13589, Nov. 2016. [CrossRef]
- T. Kupsa, J. Vanek, P. Zak, L. Jebavy, and J. M. Horacek, “Serum levels of selected cytokines and soluble adhesion molecules in acute myeloid leukemia: Soluble receptor for interleukin-2 predicts overall survival,” Cytokine, vol. 128, p. 155005, Apr. 2020. [CrossRef]
- C. Walker, E. Mojares, and A. Del Río Hernández, “Role of Extracellular Matrix in Development and Cancer Progression,” Int J Mol Sci, vol. 19, no. 10, p. 3028, Oct. 2018. [CrossRef]
- N. Kolesnikoff, C.-H. Chen, and M. S. Samuel, “Interrelationships between the extracellular matrix and the immune microenvironment that govern epithelial tumour progression,” Clin Sci (Lond), vol. 136, no. 5, pp. 361–377, Mar. 2022. [CrossRef]
- S. Karlsson and H. Nyström, “The extracellular matrix in colorectal cancer and its metastatic settling - Alterations and biological implications,” Crit Rev Oncol Hematol, vol. 175, p. 103712, Jul. 2022. [CrossRef]
- J. K. Tee et al., “Nanoparticles’ interactions with vasculature in diseases,” Chem Soc Rev, vol. 48, no. 21, pp. 5381–5407, Oct. 2019. [CrossRef]
- J. H. E. Baker et al., “Targeting the tumour vasculature: exploitation of low oxygenation and sensitivity to NOS inhibition by treatment with a hypoxic cytotoxin,” PLoS One, vol. 8, no. 10, p. e76832, 2013. [CrossRef]
- S. Maman and I. P. Witz, “A history of exploring cancer in context,” Nat Rev Cancer, vol. 18, no. 6, Art. no. 6, Jun. 2018. [CrossRef]
- S. Leonelli and R. A. Ankeny, “What makes a model organism?,” Endeavour, vol. 37, no. 4, pp. 209–212, Dec. 2013. [CrossRef]
- Z. D. Blount, “The unexhausted potential of E. coli,” Elife, vol. 4, p. e05826, Mar. 2015. [CrossRef]
- J. Nielsen, “Yeast Systems Biology: Model Organism and Cell Factory,” Biotechnol J, vol. 14, no. 9, p. e1800421, Sep. 2019. [CrossRef]
- S. A. Renshaw and N. S. Trede, “A model 450 million years in the making: zebrafish and vertebrate immunity,” Dis Model Mech, vol. 5, no. 1, pp. 38–47, Jan. 2012. [CrossRef]
- V. Paschall and K. Liu, “An Orthotopic Mouse Model of Spontaneous Breast Cancer Metastasis,” J Vis Exp, no. 114, p. 54040, Aug. 2016. [CrossRef]
- W. Lodge et al., “Tumor-derived MMPs regulate cachexia in a Drosophila cancer model,” Dev Cell, vol. 56, no. 18, pp. 2664-2680.e6, Sep. 2021. [CrossRef]
- V. Vanhooren and C. Libert, “The mouse as a model organism in aging research: usefulness, pitfalls and possibilities,” Ageing Res Rev, vol. 12, no. 1, pp. 8–21, Jan. 2013. [CrossRef]
- C. E. Trammell and A. G. Goodman, “Emerging Mechanisms of Insulin-Mediated Antiviral Immunity in Drosophila melanogaster,” Front Immunol, vol. 10, p. 2973, 2019. [CrossRef]
- S. Fields and M. Johnston, “Whither Model Organism Research?,” Science, vol. 307, no. 5717, pp. 1885–1886, Mar. 2005. [CrossRef]
- G. Yao et al., “Astragalin attenuates depression-like behaviors and memory deficits and promotes M2 microglia polarization by regulating IL-4R/JAK1/STAT6 signaling pathway in a murine model of perimenopausal depression,” Psychopharmacology (Berl), vol. 239, no. 8, pp. 2421–2443, Aug. 2022. [CrossRef]
- C. A. Foss et al., “PET/CT imaging of CSF1R in a mouse model of tuberculosis,” Eur J Nucl Med Mol Imaging, vol. 49, no. 12, pp. 4088–4096, Oct. 2022. [CrossRef]
- Y. Nong et al., “Single dose of synthetic microRNA-199a or microRNA-149 mimic does not improve cardiac function in a murine model of myocardial infarction,” Mol Cell Biochem, vol. 476, no. 11, pp. 4093–4106, Nov. 2021. [CrossRef]
- P. Stiedl, B. Grabner, K. Zboray, E. Bogner, and E. Casanova, “Modeling cancer using genetically engineered mice,” Methods Mol Biol, vol. 1267, pp. 3–18, 2015. [CrossRef]
- D. Entenberg, M. H. Oktay, and J. S. Condeelis, “Intravital imaging to study cancer progression and metastasis,” Nat Rev Cancer, vol. 23, no. 1, pp. 25–42, Jan. 2023. [CrossRef]
- W. Thamavit, N. Bhamarapravati, S. Sahaphong, S. Vajrasthira, and S. Angsubhakorn, “Effects of Dimethylnitrosamine on Induction of Cholagiocarcinoma in Opisthorchis viverrini-infected Syrian Golden Hamsters1,” Cancer Research, vol. 38, no. 12, pp. 4634–4639, Dec. 1978.
- R. A. Crallan, N. T. Georgopoulos, and J. Southgate, “Experimental models of human bladder carcinogenesis,” Carcinogenesis, vol. 27, no. 3, pp. 374–381, Mar. 2006. [CrossRef]
- M.-B. Hu et al., “Differential expressions of integrin-linked kinase, β-parvin and cofilin 1 in high-fat diet induced prostate cancer progression in a transgenic mouse model,” Oncol Lett, vol. 16, no. 4, pp. 4945–4952, Oct. 2018. [CrossRef]
- A. Asgharpour et al., “A diet-induced animal model of non-alcoholic fatty liver disease and hepatocellular cancer,” J Hepatol, vol. 65, no. 3, pp. 579–588, Sep. 2016. [CrossRef]
- Y. Chen, X. Liu, L. Gao, and Y. Liu, “Xenograft Mouse Model of Human Uveal Melanoma,” Bio Protoc, vol. 7, no. 21, p. e2594, Nov. 2017. [CrossRef]
- 3A. Fantozzi and G. Christofori, “Mouse models of breast cancer metastasis,” Breast Cancer Research, vol. 8, no. 4, p. 212, 2006. [CrossRef]
- S. Yang, J. J. Zhang, and X.-Y. Huang, “Mouse models for tumor metastasis,” Methods Mol Biol, vol. 928, pp. 221–228, 2012. [CrossRef]
- E. E. Patton et al., “Melanoma models for the next generation of therapies,” Cancer Cell, vol. 39, no. 5, pp. 610–631, May 2021. [CrossRef]
- V. Piskovatska, O. Strilbytska, A. Koliada, A. Vaiserman, and O. Lushchak, “Health Benefits of Anti-aging Drugs,” Subcell Biochem, vol. 91, pp. 339–392, 2019. [CrossRef]
- E. Loeuillard, S. R. Fischbach, G. J. Gores, and S. Rizvi, “Animal models of cholangiocarcinoma,” Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease, vol. 1865, no. 5, pp. 982–992, May 2019. [CrossRef]
- J. Seok et al., “Genomic responses in mouse models poorly mimic human inflammatory diseases,” Proceedings of the National Academy of Sciences, vol. 110, no. 9, pp. 3507–3512, Feb. 2013. [CrossRef]
- H. B. van der Worp et al., “Can Animal Models of Disease Reliably Inform Human Studies?,” PLOS Medicine, vol. 7, no. 3, p. e1000245, Mar. 2010. [CrossRef]
- A. Chung, D. Nasralla, and A. Quaglia, “Understanding the Immunoenvironment of Primary Liver Cancer: A Histopathology Perspective,” J Hepatocell Carcinoma, vol. 9, pp. 1149–1169, Nov. 2022. [CrossRef]
- G. Lendvai et al., “Cholangiocarcinoma: Classification, Histopathology and Molecular Carcinogenesis,” Pathol Oncol Res, vol. 26, no. 1, pp. 3–15, Jan. 2020. [CrossRef]
- F. Mungenast et al., “Next-Generation Digital Histopathology of the Tumor Microenvironment,” Genes, vol. 12, no. 4, Art. no. 4, Apr. 2021. [CrossRef]
- K. Jia et al., “Multiplex immunohistochemistry defines the tumor immune microenvironment and immunotherapeutic outcome in CLDN18.2-positive gastric cancer,” BMC Med, vol. 20, no. 1, p. 223, Jul. 2022. [CrossRef]
- J. S. Ahn et al., “Plasmablastic Lymphomas: Characterization of Tumor Microenvironment Using CD163 and PD-1 Immunohistochemistry,” Ann Clin Lab Sci, vol. 50, no. 2, pp. 213–218, Mar. 2020.
- H. D. Papenfuss, J. F. Gross, M. Intaglietta, and F. A. Treese, “A transparent access chamber for the rat dorsal skin fold,” Microvasc Res, vol. 18, no. 3, pp. 311–318, Nov. 1979. [CrossRef]
- S. J. Lunt, C. Gray, C. C. Reyes-Aldasoro, S. J. Matcher, and G. M. Tozer, “Application of intravital microscopy in studies of tumor microcirculation,” J Biomed Opt, vol. 15, no. 1, p. 011113, Feb. 2010. [CrossRef]
- S. Akerman et al., “Influence of soluble or matrix-bound isoforms of vascular endothelial growth factor-A on tumor response to vascular-targeted strategies,” Int J Cancer, vol. 133, no. 11, pp. 2563–2576, Dec. 2013. [CrossRef]
- C. C. Reyes-Aldasoro et al., “Estimation of apparent tumor vascular permeability from multiphoton fluorescence microscopic images of P22 rat sarcomas in vivo,” Microcirculation, vol. 15, no. 1, pp. 65–79, Jan. 2008. [CrossRef]
- S. Prasad et al., “Optical and magnetic resonance imaging approaches for investigating the tumour microenvironment: state-of-the-art review and future trends,” Nanotechnology, vol. 32, no. 6, p. 062001, Nov. 2020. [CrossRef]
- M. Matsuo, S. Matsumoto, J. B. Mitchell, M. C. Krishna, and K. Camphausen, “Magnetic resonance imaging of the tumor microenvironment in radiotherapy: perfusion, hypoxia, and metabolism,” Semin Radiat Oncol, vol. 24, no. 3, pp. 210–217, Jul. 2014. [CrossRef]
- B. Zinnhardt et al., “Imaging of the glioma microenvironment by TSPO PET,” Eur J Nucl Med Mol Imaging, vol. 49, no. 1, pp. 174–185, Dec. 2021. [CrossRef]
- D. M. L. Lilburn and A. M. Groves, “The role of PET in imaging of the tumour microenvironment and response to immunotherapy,” Clin Radiol, vol. 76, no. 10, p. 784.e1-784.e15, Oct. 2021. [CrossRef]
- R. A. Lambert, “COMPARATIVE STUDIES UPON CANCER CELLS AND NORMAL CELLS : II. THE CHARACTER OF GROWTH IN VITRO WITH SPECIAL REFERENCE TO CELL DIVISION,” J Exp Med, vol. 17, no. 5, pp. 499–510, May 1913. [CrossRef]
- P. A. Eichorn, K. V. Huffman, J. J. Oleson, S. L. Halliday, and J. H. Williams, “A Comparison of in Vivo and in Vitro Tests for Antineoplastic Activity of Eight Compounds,” Annals of the New York Academy of Sciences, vol. 58, no. 7, pp. 1172–1182, 1954. [CrossRef]
- D. Tuveson and H. Clevers, “Cancer modeling meets human organoid technology,” Science, vol. 364, no. 6444, pp. 952–955, Jun. 2019. [CrossRef]
- S. C. Baker, S. Shabir, and J. Southgate, “Biomimetic Urothelial Tissue Models for the in Vitro Evaluation of Barrier Physiology and Bladder Drug Efficacy,” Mol. Pharmaceutics, vol. 11, no. 7, pp. 1964–1970, Jul. 2014. [CrossRef]
- P. Pound and M. Ritskes-Hoitinga, “Is it possible to overcome issues of external validity in preclinical animal research? Why most animal models are bound to fail,” Journal of Translational Medicine, vol. 16, no. 1, p. 304, Nov. 2018. [CrossRef]
- M. Musa, D. Ouaret, and W. F. Bodmer, “In Vitro Analyses of Interactions Between Colonic Myofibroblasts and Colorectal Cancer Cells for Anticancer Study,” Anticancer Res, vol. 40, no. 11, pp. 6063–6073, Nov. 2020. [CrossRef]
- A. Alhussan et al., “Potential of Gold Nanoparticle in Current Radiotherapy Using a Co-Culture Model of Cancer Cells and Cancer Associated Fibroblast Cells,” Cancers (Basel), vol. 14, no. 15, p. 3586, Jul. 2022. [CrossRef]
- E. Hamilton et al., “Mimicking the tumour microenvironment: three different co-culture systems induce a similar phenotype but distinct proliferative signals in primary chronic lymphocytic leukaemia cells,” Br J Haematol, vol. 158, no. 5, pp. 589–599, Sep. 2012. [CrossRef]
- R. Xu and F. M. Richards, “Development of In Vitro Co-Culture Model in Anti-Cancer Drug Development Cascade,” Comb Chem High Throughput Screen, vol. 20, no. 5, pp. 451–457, 2017. [CrossRef]
- M. Curtis et al., “Fibroblasts Mobilize Tumor Cell Glycogen to Promote Proliferation and Metastasis,” Cell Metab, vol. 29, no. 1, pp. 141-155.e9, Jan. 2019. [CrossRef]
- B. Erdogan et al., “Cancer-associated fibroblasts promote directional cancer cell migration by aligning fibronectin,” J Cell Biol, vol. 216, no. 11, pp. 3799–3816, Nov. 2017. [CrossRef]
- C. Kanthou et al., “Tumour cells expressing single VEGF isoforms display distinct growth, survival and migration characteristics,” PLoS One, vol. 9, no. 8, p. e104015, 2014. [CrossRef]
- S. Wen et al., “Cancer-associated fibroblast (CAF)-derived IL32 promotes breast cancer cell invasion and metastasis via integrin β3-p38 MAPK signalling,” Cancer Lett, vol. 442, pp. 320–332, Feb. 2019. [CrossRef]
- J. Kikuchi, D. Koyama, H. Y. Mukai, and Y. Furukawa, “Suitable drug combination with bortezomib for multiple myeloma under stroma-free conditions and in contact with fibronectin or bone marrow stromal cells,” Int J Hematol, vol. 99, no. 6, pp. 726–736, Jun. 2014. [CrossRef]
- S. J. Lunt et al., “Vascular effects dominate solid tumor response to treatment with combretastatin A-4-phosphate,” Int J Cancer, vol. 129, no. 8, pp. 1979–1989, Oct. 2011. [CrossRef]
- M. Kapałczyńska et al., “2D and 3D cell cultures – a comparison of different types of cancer cell cultures,” Arch Med Sci, vol. 14, no. 4, pp. 910–919, Jun. 2018. [CrossRef]
- S. J. Ellem, E. M. De-Juan-Pardo, and G. P. Risbridger, “In vitro modeling of the prostate cancer microenvironment,” Advanced Drug Delivery Reviews, vol. 79–80, pp. 214–221, Dec. 2014. [CrossRef]
- M. Huerta-Reyes and A. Aguilar-Rojas, “Three-dimensional models to study breast cancer (Review),” Int J Oncol, vol. 58, no. 3, pp. 331–343, Mar. 2021. [CrossRef]
- S. I. Bär, B. Biersack, and R. Schobert, “3D cell cultures, as a surrogate for animal models, enhance the diagnostic value of preclinical in vitro investigations by adding information on the tumour microenvironment: a comparative study of new dual-mode HDAC inhibitors,” Invest New Drugs, vol. 40, no. 5, pp. 953–961, Oct. 2022. [CrossRef]
- L. A. Kunz-Schughart, M. Kreutz, and R. Knuechel, “Multicellular spheroids: a three-dimensional in vitro culture system to study tumour biology,” Int J Exp Pathol, vol. 79, no. 1, pp. 1–23, Feb. 1998. [CrossRef]
- S. Gunti, A. T. K. Hoke, K. P. Vu, and N. R. London, “Organoid and Spheroid Tumor Models: Techniques and Applications,” Cancers (Basel), vol. 13, no. 4, p. 874, Feb. 2021. [CrossRef]
- T. Xia, W.-L. Du, X.-Y. Chen, and Y.-N. Zhang, “Organoid models of the tumor microenvironment and their applications,” J Cell Mol Med, vol. 25, no. 13, pp. 5829–5841, May 2021. [CrossRef]
- R. Rizzo et al., “A pH-sensor scaffold for mapping spatiotemporal gradients in three-dimensional in vitro tumour models,” Biosens Bioelectron, vol. 212, p. 114401, Sep. 2022. [CrossRef]
- G. Mazzoleni, D. Di Lorenzo, and N. Steimberg, “Modelling tissues in 3D: the next future of pharmaco-toxicology and food research?,” Genes Nutr, vol. 4, no. 1, pp. 13–22, Mar. 2009. [CrossRef]
- L. Neufeld, E. Yeini, S. Pozzi, and R. Satchi-Fainaro, “3D bioprinted cancer models: from basic biology to drug development,” Nat Rev Cancer, vol. 22, no. 12, Art. no. 12, Dec. 2022. [CrossRef]
- R. Leek, D. R. Grimes, A. L. Harris, and A. McIntyre, “Methods: Using Three-Dimensional Culture (Spheroids) as an In Vitro Model of Tumour Hypoxia,” Adv Exp Med Biol, vol. 899, pp. 167–196, 2016. [CrossRef]
- I. Manini et al., “Role of Microenvironment in Glioma Invasion: What We Learned from In Vitro Models,” Int J Mol Sci, vol. 19, no. 1, p. 147, Jan. 2018. [CrossRef]
- H.-F. Tsai, A. Trubelja, A. Q. Shen, and G. Bao, “Tumour-on-a-chip: microfluidic models of tumour morphology, growth and microenvironment,” J R Soc Interface, vol. 14, no. 131, p. 20170137, Jun. 2017. [CrossRef]
- J. Nolan, O. M. T. Pearce, H. R. C. Screen, M. M. Knight, and S. W. Verbruggen, “Organ-on-a-Chip and Microfluidic Platforms for Oncology in the UK,” Cancers (Basel), vol. 15, no. 3, p. 635, Jan. 2023. [CrossRef]
- A. Ozcelikkale, H.-R. Moon, M. Linnes, and B. Han, “In vitro microfluidic models of tumor microenvironment to screen transport of drugs and nanoparticles,” Wiley Interdiscip Rev Nanomed Nanobiotechnol, vol. 9, no. 5, Sep. 2017. [CrossRef]
- B. Kundu, D. Caballero, C. M. Abreu, R. L. Reis, and S. C. Kundu, “The Tumor Microenvironment: An Introduction to the Development of Microfluidic Devices,” Adv Exp Med Biol, vol. 1379, pp. 115–138, 2022. [CrossRef]
- H. M. Byrne, “Dissecting cancer through mathematics: from the cell to the animal model,” Nat Rev Cancer, vol. 10, no. 3, pp. 221–230, Mar. 2010. [CrossRef]
- P. M. Altrock, L. L. Liu, and F. Michor, “The mathematics of cancer: integrating quantitative models,” Nat Rev Cancer, vol. 15, no. 12, pp. 730–745, Dec. 2015. [CrossRef]
- T. R. Malthus, An Essay on the Principle of Population: Or, A View of Its Past and Present Effects on Human Happiness. J. Johnson, 1807.
- P. Armitage and R. Doll, “The Age Distribution of Cancer and a Multi-stage Theory of Carcinogenesis,” Br J Cancer, vol. 8, no. 1, pp. 1–12, Mar. 1954.
- T. G. Clark, M. J. Bradburn, S. B. Love, and D. G. Altman, “Survival Analysis Part I: Basic concepts and first analyses,” Br J Cancer, vol. 89, no. 2, pp. 232–238, Jul. 2003. [CrossRef]
- J. Fan and Z. Yu, “A univariate model of calcium release in the dyadic cleft of cardiac myocytes,” Annu Int Conf IEEE Eng Med Biol Soc, vol. 2009, pp. 4499–4503, 2009. [CrossRef]
- J. Bradburn, T. G. Clark, S. B. Love, and D. G. Altman, “Survival Analysis Part II: Multivariate data analysis – an introduction to concepts and methods,” Br J Cancer, vol. 89, no. 3, pp. 431–436, Aug. 2003. [CrossRef]
- K. Azuma et al., “Clinical significance of plasma-free amino acids and tryptophan metabolites in patients with non-small cell lung cancer receiving PD-1 inhibitor: a pilot cohort study for developing a prognostic multivariate model,” J Immunother Cancer, vol. 10, no. 5, p. e004420, May 2022. [CrossRef]
- R. A. Beckman, I. Kareva, and F. R. Adler, “How Should Cancer Models Be Constructed?,” Cancer Control, vol. 27, no. 1, p. 1073274820962008, Sep. 2020. [CrossRef]
- R. A. Anderson and V. Quaranta, “Integrative mathematical oncology,” Nat Rev Cancer, vol. 8, no. 3, Art. no. 3, Mar. 2008. [CrossRef]
- L. Curtin, A. Hawkins-Daarud, A. B. Porter, K. G. van der Zee, M. R. Owen, and K. R. Swanson, “A Mechanistic Investigation into Ischemia-Driven Distal Recurrence of Glioblastoma,” Bull Math Biol, vol. 82, no. 11, p. 143, Nov. 2020. [CrossRef]
- D. R. Menon and M. Fujita, “A state of stochastic cancer stemness through the CDK1-SOX2 axis,” Oncotarget, vol. 10, no. 27, pp. 2583–2585, Apr. 2019. [CrossRef]
- N. Kumar, G. M. Cramer, S. A. Z. Dahaj, B. Sundaram, J. P. Celli, and R. V. Kulkarni, “Stochastic modeling of phenotypic switching and chemoresistance in cancer cell populations,” Sci Rep, vol. 9, no. 1, p. 10845, Jul. 2019. [CrossRef]
- C. J. Gommes, T. Louis, I. Bourgot, A. Noël, S. Blacher, and E. Maquoi, “Remodelling of the fibre-aggregate structure of collagen gels by cancer-associated fibroblasts: A time-resolved grey-tone image analysis based on stochastic modelling,” Front Immunol, vol. 13, p. 988502, 2022. [CrossRef]
- V. Morales and L. Soto-Ortiz, “Modeling Macrophage Polarization and Its Effect on Cancer Treatment Success,” Open J Immunol, vol. 8, no. 2, pp. 36–80, Jun. 2018. [CrossRef]
- W. Blaszczak and P. Swietach, “What do cellular responses to acidity tell us about cancer?,” Cancer Metastasis Rev, vol. 40, no. 4, pp. 1159–1176, Dec. 2021. [CrossRef]
- A. Belfatto et al., “Comparison between model-predicted tumor oxygenation dynamics and vascular-/flow-related Doppler indices,” Med Phys, vol. 44, no. 5, pp. 2011–2019, May 2017. [CrossRef]
- L. Possenti et al., “A Mesoscale Computational Model for Microvascular Oxygen Transfer,” Ann Biomed Eng, vol. 49, no. 12, pp. 3356–3373, Dec. 2021. [CrossRef]
- A. Zhang, L. X. Xu, G. A. Sandison, and J. Zhang, “A microscale model for prediction of breast cancer cell damage during cryosurgery,” Cryobiology, vol. 47, no. 2, pp. 143–154, Oct. 2003. [CrossRef]
- D. Noble, “Modeling the heart--from genes to cells to the whole organ,” Science, vol. 295, no. 5560, pp. 1678–1682, Mar. 2002. [CrossRef]
- H. Chen, Y. Cai, Q. Chen, and Z. Li, “Multiscale modeling of solid stress and tumor cell invasion in response to dynamic mechanical microenvironment,” Biomech Model Mechanobiol, vol. 19, no. 2, pp. 577–590, Apr. 2020. [CrossRef]
- S. Sadhukhan, P. K. Mishra, S. K. Basu, and J. K. Mandal, “A multi-scale agent-based model for avascular tumour growth,” Biosystems, vol. 206, p. 104450, Aug. 2021. [CrossRef]
- Z. Wang, J. D. Butner, R. Kerketta, V. Cristini, and T. S. Deisboeck, “Simulating cancer growth with multiscale agent-based modeling,” Seminars in Cancer Biology, vol. 30, pp. 70–78, Feb. 2015. [CrossRef]
- P. Gerlee, E. Kim, and A. R. A. Anderson, “Bridging scales in cancer progression: Mapping genotype to phenotype using neural networks,” Seminars in Cancer Biology, vol. 30, pp. 30–41, Feb. 2015. [CrossRef]
- A. Wijeratne et al., “Multiscale modelling of solid tumour growth: the effect of collagen micromechanics,” Biomech Model Mechanobiol, vol. 15, no. 5, pp. 1079–1090, Oct. 2016. [CrossRef]
- P. Kumar, J. Li, and C. Surulescu, “Multiscale modeling of glioma pseudopalisades: contributions from the tumor microenvironment,” J Math Biol, vol. 82, no. 6, p. 49, Apr. 2021. [CrossRef]
- G. G. Powathil, M. Swat, and M. A. J. Chaplain, “Systems oncology: towards patient-specific treatment regimes informed by multiscale mathematical modelling,” Semin. Cancer Biol., vol. 30, pp. 13–20, Feb. 2015. [CrossRef]
- M. R. Nikmaneshi and B. Firoozabadi, “Investigation of cancer response to chemotherapy: a hybrid multi-scale mathematical and computational model of the tumor microenvironment,” Biomech Model Mechanobiol, vol. 21, no. 4, pp. 1233–1249, Aug. 2022. [CrossRef]
- L. Peng, D. Trucu, P. Lin, A. Thompson, and M. A. J. Chaplain, “A Multiscale Mathematical Model of Tumour Invasive Growth,” Bull Math Biol, vol. 79, no. 3, pp. 389–429, Mar. 2017. [CrossRef]
- M. Chowkwale, G. J. Mahler, P. Huang, and B. T. Murray, “A multiscale in silico model of endothelial to mesenchymal transformation in a tumor microenvironment,” J Theor Biol, vol. 480, pp. 229–240, Nov. 2019. [CrossRef]
- F. Pourhasanzade and S. H. Sabzpoushan, “A New Mathematical Model for Controlling Tumor Growth Based on Microenvironment Acidity and Oxygen Concentration,” Biomed Res Int, vol. 2021, p. 8886050, 2021. [CrossRef]
- K. H. W. J. ten Tusscher, D. Noble, P. J. Noble, and A. V. Panfilov, “A model for human ventricular tissue,” Am J Physiol Heart Circ Physiol, vol. 286, no. 4, pp. H1573-1589, Apr. 2004. [CrossRef]
- K.-A. Norton, C. Gong, S. Jamalian, and A. S. Popel, “Multiscale Agent-Based and Hybrid Modeling of the Tumor Immune Microenvironment,” Processes (Basel), vol. 7, no. 1, p. 37, Jan. 2019. [CrossRef]
- D. C. Walker and J. Southgate, “The virtual cell--a candidate co-ordinator for ‘middle-out’ modelling of biological systems,” Brief Bioinform, vol. 10, no. 4, pp. 450–461, Jul. 2009. [CrossRef]
- D. Walker, S. Wood, J. Southgate, M. Holcombe, and R. Smallwood, “An integrated agent-mathematical model of the effect of intercellular signalling via the epidermal growth factor receptor on cell proliferation,” J Theor Biol, vol. 242, no. 3, pp. 774–789, Oct. 2006. [CrossRef]
- A. Rojas-Domínguez, R. Arroyo-Duarte, F. Rincón-Vieyra, and M. Alvarado-Mentado, “Modeling cancer immunoediting in tumor microenvironment with system characterization through the ising-model Hamiltonian,” BMC Bioinformatics, vol. 23, p. 200, May 2022. [CrossRef]
- S. Rahbar et al., “Agent-based Modeling of Tumor and Immune System Interactions in Combinational Therapy with Low-dose 5-fluorouracil and Dendritic Cell Vaccine in Melanoma B16F10,” Iran J Allergy Asthma Immunol, vol. 21, no. 2, pp. 151–166, Apr. 2022. [CrossRef]
- G. Cesaro et al., “MAST: a hybrid Multi-Agent Spatio-Temporal model of tumor microenvironment informed using a data-driven approach,” Bioinform Adv, vol. 2, no. 1, p. vbac092, Dec. 2022. [CrossRef]
- Z. Tylutki, S. Polak, and B. Wiśniowska, “Top-down, Bottom-up and Middle-out Strategies for Drug Cardiac Safety Assessment via Modeling and Simulations,” Curr Pharmacol Rep, vol. 2, no. 4, pp. 171–177, 2016. [CrossRef]
- E. Tsirvouli, V. Touré, B. Niederdorfer, M. Vázquez, Å. Flobak, and M. Kuiper, “A Middle-Out Modeling Strategy to Extend a Colon Cancer Logical Model Improves Drug Synergy Predictions in Epithelial-Derived Cancer Cell Lines,” Front Mol Biosci, vol. 7, p. 502573, 2020. [CrossRef]
- K. Sugano, “Lost in modelling and simulation?,” ADMET DMPK, vol. 9, no. 2, pp. 75–109, Mar. 2021. [CrossRef]
- W. Secomb and A. R. Pries, “The microcirculation: physiology at the mesoscale,” J Physiol, vol. 589, no. Pt 5, pp. 1047–1052, Mar. 2011. [CrossRef]
- K. S. Korolev, J. B. Xavier, and J. Gore, “Turning ecology and evolution against cancer,” Nat Rev Cancer, vol. 14, no. 5, pp. 371–380, May 2014. [CrossRef]
- M. Dujon et al., “Identifying key questions in the ecology and evolution of cancer,” Evol Appl, vol. 14, no. 4, pp. 877–892, Feb. 2021. [CrossRef]
- A. Bukkuri et al., “Modeling cancer’s ecological and evolutionary dynamics,” Med Oncol, vol. 40, no. 4, p. 109, Feb. 2023. [CrossRef]
- B. Morris et al., “Identifying the spatial and temporal dynamics of molecularly-distinct glioblastoma sub-populations,” Math Biosci Eng, vol. 17, no. 5, pp. 4905–4941, Jul. 2020. [CrossRef]
- T. Antal and P. L. Krapivsky, “Exact solution of a two-type branching process: models of tumor progression,” J. Stat. Mech., vol. 2011, no. 08, p. P08018, Aug. 2011. [CrossRef]
- I. Bozic et al., “Accumulation of driver and passenger mutations during tumor progression,” Proceedings of the National Academy of Sciences, vol. 107, no. 43, pp. 18545–18550, Oct. 2010. [CrossRef]
- D. Lewin, B. Avignon, A. Tovaglieri, L. Cabon, N. Gjorevski, and L. G. Hutchinson, “An in silico Model of T Cell Infiltration Dynamics Based on an Advanced in vitro System to Enhance Preclinical Decision Making in Cancer Immunotherapy,” Front Pharmacol, vol. 13, p. 837261, 2022. [CrossRef]
- L. Curtin, A. Hawkins-Daarud, K. G. van der Zee, K. R. Swanson, and M. R. Owen, “Speed Switch in Glioblastoma Growth Rate due to Enhanced Hypoxia-Induced Migration,” Bull Math Biol, vol. 82, no. 3, p. 43, Mar. 2020. [CrossRef]
- B. de Melo Quintela, S. Hervas-Raluy, J. M. Garcia-Aznar, D. Walker, K. Y. Wertheim, and M. Viceconti, “A theoretical analysis of the scale separation in a model to predict solid tumour growth,” J Theor Biol, vol. 547, p. 111173, Aug. 2022. [CrossRef]
- R. A. Anderson, “A Hybrid Multiscale Model of Solid Tumour Growth and Invasion: Evolution and the Microenvironment,” in Single-Cell-Based Models in Biology and Medicine, A. R. A. Anderson, M. A. J. Chaplain, and K. A. Rejniak, Eds., in Mathematics and Biosciences in Interaction. Basel: Birkhäuser, 2007, pp. 3–28. [CrossRef]
- M. a. J. Chaplain, S. R. McDougall, and A. R. A. Anderson, “Mathematical modeling of tumor-induced angiogenesis,” Annu Rev Biomed Eng, vol. 8, pp. 233–257, 2006. [CrossRef]
- M. A. Chaplain, S. M. Giles, B. D. Sleeman, and R. J. Jarvis, “A mathematical analysis of a model for tumour angiogenesis,” J Math Biol, vol. 33, no. 7, pp. 744–770, 1995. [CrossRef]
- H. Enderling, M. A. J. Chaplain, A. R. A. Anderson, and J. S. Vaidya, “A mathematical model of breast cancer development, local treatment and recurrence,” J Theor Biol, vol. 246, no. 2, pp. 245–259, May 2007. [CrossRef]
- I. Ramis-Conde, M. A. J. Chaplain, A. R. A. Anderson, and D. Drasdo, “Multi-scale modelling of cancer cell intravasation: the role of cadherins in metastasis,” Phys Biol, vol. 6, no. 1, p. 016008, Mar. 2009. [CrossRef]
- B. D. Sleeman and H. R. Nimmo, “Fluid transport in vascularized tumours and metastasis,” IMA J Math Appl Med Biol, vol. 15, no. 1, pp. 53–63, Mar. 1998.
- M. R. Owen, H. M. Byrne, and C. E. Lewis, “Mathematical modelling of the use of macrophages as vehicles for drug delivery to hypoxic tumour sites,” J Theor Biol, vol. 226, no. 4, pp. 377–391, Feb. 2004. [CrossRef]
- T. D. Lewin, H. M. Byrne, P. K. Maini, J. J. Caudell, E. G. Moros, and H. Enderling, “The importance of dead material within a tumour on the dynamics in response to radiotherapy,” Phys Med Biol, vol. 65, no. 1, p. 015007, Jan. 2020. [CrossRef]
- M. Italia, K. Y. Wertheim, S. Taschner-Mandl, D. Walker, and F. Dercole, “Mathematical Model of Clonal Evolution Proposes a Personalised Multi-Modal Therapy for High-Risk Neuroblastoma,” Cancers (Basel), vol. 15, no. 7, p. 1986, Mar. 2023. [CrossRef]
- A. Araujo, L. M. Cook, C. C. Lynch, and D. Basanta, “An integrated computational model of the bone microenvironment in bone-metastatic prostate cancer,” Cancer Res, vol. 74, no. 9, pp. 2391–2401, May 2014. [CrossRef]
- M. A. Clarke and J. Fisher, “Executable cancer models: successes and challenges,” Nat Rev Cancer, vol. 20, no. 6, pp. 343–354, Jun. 2020. [CrossRef]
- I. Myung, Y. Tang, and M. A. Pitt, “Chapter 11 Evaluation and Comparison of Computational Models,” in Methods in Enzymology, in Computer Methods, Part A, vol. 454. Academic Press, 2009, pp. 287–304. [CrossRef]
- B. Goldstein, J. R. Faeder, and W. S. Hlavacek, “Mathematical and computational models of immune-receptor signalling,” Nat Rev Immunol, vol. 4, no. 6, Art. no. 6, Jun. 2004. [CrossRef]
- Z. Ji, K. Yan, W. Li, H. Hu, and X. Zhu, “Mathematical and Computational Modeling in Complex Biological Systems,” BioMed Research International, vol. 2017, p. e5958321, Mar. 2017. [CrossRef]
- A. Konstorum, A. T. Vella, A. J. Adler, and R. C. Laubenbacher, “Addressing current challenges in cancer immunotherapy with mathematical and computational modelling,” Journal of The Royal Society Interface, vol. 14, no. 131, p. 20170150, Jun. 2017. [CrossRef]
- V. Garcia, S. Bonhoeffer, and F. Fu, “Cancer-induced immunosuppression can enable effectiveness of immunotherapy through bistability generation: A mathematical and computational examination,” J Theor Biol, vol. 492, p. 110185, May 2020. [CrossRef]
- R. Vega, M. Carretero, R. D. M. Travasso, and L. L. Bonilla, “Notch signaling and taxis mechanisms regulate early stage angiogenesis: A mathematical and computational model,” PLoS Comput Biol, vol. 16, no. 1, p. e1006919, Jan. 2020. [CrossRef]
- J. West, M. Robertson-Tessi, and A. R. A. Anderson, “Agent-based methods facilitate integrative science in cancer,” Trends Cell Biol, vol. 33, no. 4, pp. 300–311, Apr. 2023. [CrossRef]
- J. Metzcar, Y. Wang, R. Heiland, and P. Macklin, “A Review of Cell-Based Computational Modeling in Cancer Biology,” JCO Clin Cancer Inform, vol. 3, pp. 1–13, Feb. 2019. [CrossRef]
- A. Homeyer et al., “Automated quantification of steatosis: agreement with stereological point counting,” Diagnostic Pathology, vol. 12, no. 1, p. 80, Nov. 2017. [CrossRef]
- M. Dawood, K. Branson, N. M. Rajpoot, and F. ul A. A. Minhas, “All You Need is Color: Image Based Spatial Gene Expression Prediction Using Neural Stain Learning,” in Machine Learning and Principles and Practice of Knowledge Discovery in Databases, M. Kamp, I. Koprinska, A. Bibal, T. Bouadi, B. Frénay, L. Galárraga, J. Oramas, L. Adilova, G. Graça, et al., Eds., in Communications in Computer and Information Science. Cham: Springer International Publishing, 2021, pp. 437–450. [CrossRef]
- A. Ortega-Ruiz, C. Karabağ, V. G. Garduño, and C. C. Reyes-Aldasoro, “Morphological Estimation of Cellularity on Neo-Adjuvant Treated Breast Cancer Histological Images,” Journal of Imaging, vol. 6, no. 10, p. 101, Oct. 2020. [CrossRef]
- F. Serin, M. Erturkler, and M. Gul, “A novel overlapped nuclei splitting algorithm for histopathological images,” Computer Methods and Programs in Biomedicine, vol. 151, pp. 57–70, Nov. 2017. [CrossRef]
- A. W. Sullivan, S. Ghosh, I. T. Ocal, R. L. Camp, D. L. Rimm, and G. G. Chung, “Microvessel area using automated image analysis is reproducible and is associated with prognosis in breast cancer,” Human Pathology, vol. 40, no. 2, pp. 156–165, Feb. 2009. [CrossRef]
- C. S. Patlak, R. G. Blasberg, and J. D. Fenstermacher, “Graphical evaluation of blood-to-brain transfer constants from multiple-time uptake data,” J Cereb Blood Flow Metab, vol. 3, no. 1, pp. 1–7, Mar. 1983. [CrossRef]
- C. C. Reyes-Aldasoro, S. Akerman, and G. M. Tozer, “Measuring the velocity of fluorescently labelled red blood cells with a keyhole tracking algorithm,” J Microsc, vol. 229, no. Pt 1, pp. 162–173, Jan. 2008. [CrossRef]
- Y. Yuan, “Spatial Heterogeneity in the Tumor Microenvironment,” Cold Spring Harb Perspect Med, vol. 6, no. 8, p. a026583, Aug. 2016. [CrossRef]
- S. McCULLOCH and W. Pitts, “The statistical organization of nervous activity,” Biometrics, vol. 4, no. 2, pp. 91–99, Jun. 1948.
- P. beim Graben and J. Wright, “From McCulloch-Pitts neurons toward biology,” Bull Math Biol, vol. 73, no. 2, pp. 261–265, Feb. 2011. [CrossRef]
- K. Simonyan and A. Zisserman, “Very Deep Convolutional Networks for Large-Scale Image Recognition.” arXiv, Apr. 10, 2015. [CrossRef]
- A. Krizhevsky, I. Sutskever, and G. E. Hinton, “ImageNet classification with deep convolutional neural networks,” Commun. ACM, vol. 60, no. 6, pp. 84–90, May 2017. [CrossRef]
- O. Ronneberger, P. Fischer, and T. Brox, “U-Net: Convolutional Networks for Biomedical Image Segmentation,” arXiv:1505.04597 [cs], May 2015, Accessed: Mar. 25, 2021. [Online]. Available: http://arxiv.org/abs/1505.04597.
- C. Szegedy et al., “Going deeper with convolutions,” in 2015 IEEE Conference on Computer Vision and Pattern Recognition (CVPR), Jun. 2015, pp. 1–9. [CrossRef]
- Y. LeCun, Y. Bengio, and G. Hinton, “Deep learning,” Nature, vol. 521, no. 7553, Art. no. 7553, May 2015. [CrossRef]
- N. Kriegeskorte and T. Golan, “Neural network models and deep learning,” Current Biology, vol. 29, no. 7, pp. R231–R236, Apr. 2019. [CrossRef]
- S. Kuntz et al., “Gastrointestinal cancer classification and prognostication from histology using deep learning: Systematic review,” Eur J Cancer, vol. 155, pp. 200–215, Sep. 2021. [CrossRef]
- A. Davri et al., “Deep Learning on Histopathological Images for Colorectal Cancer Diagnosis: A Systematic Review,” Diagnostics, vol. 12, no. 4, Art. no. 4, Apr. 2022. [CrossRef]
- A. Tran, O. Kondrashova, A. Bradley, E. D. Williams, J. V. Pearson, and N. Waddell, “Deep learning in cancer diagnosis, prognosis and treatment selection,” Genome Med, vol. 13, no. 1, p. 152, Sep. 2021. [CrossRef]
- B. Bhinder, C. Gilvary, N. S. Madhukar, and O. Elemento, “Artificial Intelligence in Cancer Research and Precision Medicine,” Cancer Discov, vol. 11, no. 4, pp. 900–915, Apr. 2021. [CrossRef]
- J. Wu and D. Lin, “A Review of Artificial Intelligence in Precise Assessment of Programmed Cell Death-ligand 1 and Tumor-infiltrating Lymphocytes in Non-Small Cell Lung Cancer,” Adv Anat Pathol, vol. 28, no. 6, pp. 439–445, Nov. 2021. [CrossRef]
- N. Thakur, H. Yoon, and Y. Chong, “Current Trends of Artificial Intelligence for Colorectal Cancer Pathology Image Analysis: A Systematic Review,” Cancers (Basel), vol. 12, no. 7, p. 1884, Jul. 2020. [CrossRef]
- B. E. Bejnordi et al., “Using deep convolutional neural networks to identify and classify tumor-associated stroma in diagnostic breast biopsies,” Modern Pathology, vol. 31, no. 10, pp. 1502–1512, Jan. 2018. [CrossRef]
- L. Pantanowitz et al., “An artificial intelligence algorithm for prostate cancer diagnosis in whole slide images of core needle biopsies: a blinded clinical validation and deployment study,” The Lancet Digital Health, vol. 2, no. 8, pp. e407–e416, Aug. 2020. [CrossRef]
- J. N. Kather et al., “Predicting survival from colorectal cancer histology slides using deep learning: A retrospective multicenter study,” PLOS Medicine, vol. 16, no. 1, p. e1002730, Jan. 2019. [CrossRef]
- M. Shaban et al., “A digital score of tumour-associated stroma infiltrating lymphocytes predicts survival in head and neck squamous cell carcinoma,” J Pathol, vol. 256, no. 2, pp. 174–185, Feb. 2022. [CrossRef]
- G. Huang, Z. Liu, L. van der Maaten, and K. Q. Weinberger, “Densely Connected Convolutional Networks.” arXiv, Jan. 28, 2018. [CrossRef]
- C. C. Reyes-Aldasoro, “The proportion of cancer-related entries in PubMed has increased considerably; is cancer truly ‘The Emperor of All Maladies’?,” PLoS One, vol. 12, no. 3, p. e0173671, 2017. [CrossRef]
- H. Sung et al., “Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries,” CA: A Cancer Journal for Clinicians, vol. 71, no. 3, pp. 209–249, 2021. [CrossRef]
Figure 1.
Number of entries indexed in PubMed for individual queries. Each query concatenated the individual keyword with the date range (2000:2023[dp]), and the restrictions corresponding to tumour microenvironment ((("tumor microenvironment") OR ("tumour microenvironment")) OR (“cancer microenvironment”)). Colours have been allocated for model organism (red), mathematical (blue), in vitro (green) and computational (brown) for visualisation purposes. The legend in the top right indicates the aggregates per group.
Figure 1.
Number of entries indexed in PubMed for individual queries. Each query concatenated the individual keyword with the date range (2000:2023[dp]), and the restrictions corresponding to tumour microenvironment ((("tumor microenvironment") OR ("tumour microenvironment")) OR (“cancer microenvironment”)). Colours have been allocated for model organism (red), mathematical (blue), in vitro (green) and computational (brown) for visualisation purposes. The legend in the top right indicates the aggregates per group.
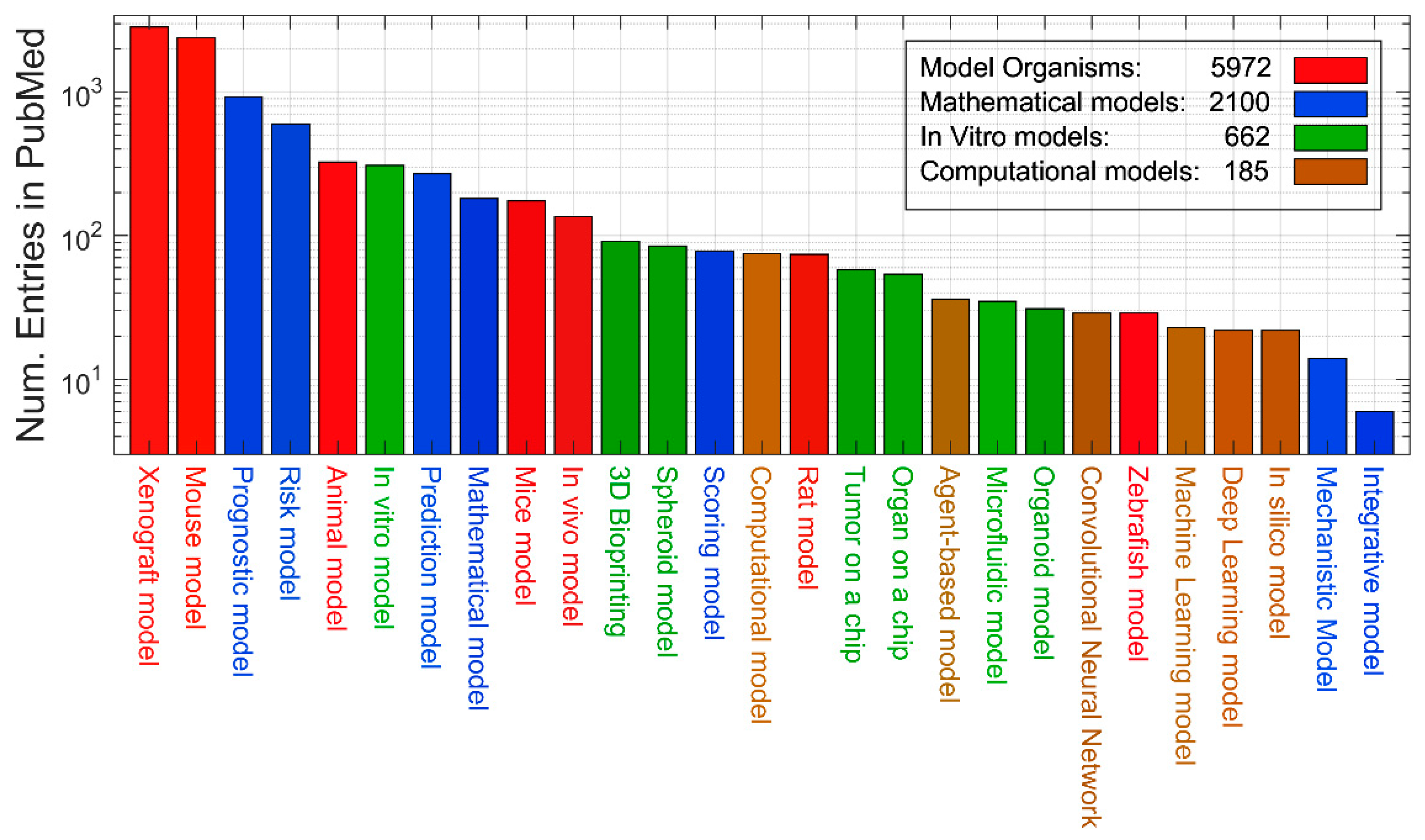
Figure 2.
Number of entries indexed in PubMed for individual queries per year of publication aggregated by the four groups. Each coloured ribbon corresponds to a sum of the keywords of each group and the year increases as indicated by the axes on the right. It should be noticed how some techniques are more established (i.e., Model organism) whilst others are more recent (Computational model).
Figure 2.
Number of entries indexed in PubMed for individual queries per year of publication aggregated by the four groups. Each coloured ribbon corresponds to a sum of the keywords of each group and the year increases as indicated by the axes on the right. It should be noticed how some techniques are more established (i.e., Model organism) whilst others are more recent (Computational model).
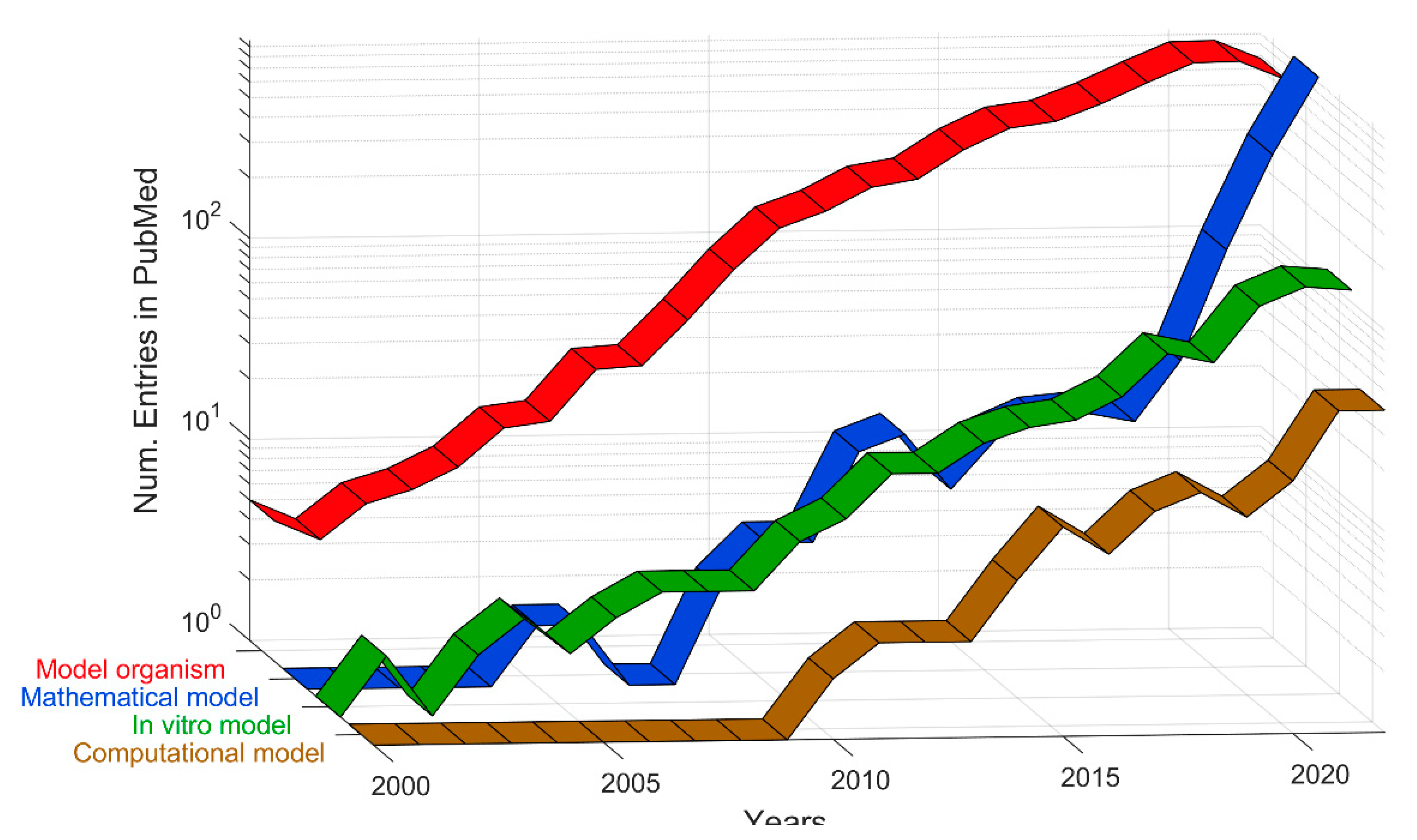
Figure 3.
Number of entries indexed in PubMed for individual queries. In this case, the query included a keyword, e.g., angiogenesis, and the word “model”. It should be noted that the vertical axis is logarithmic.
Figure 3.
Number of entries indexed in PubMed for individual queries. In this case, the query included a keyword, e.g., angiogenesis, and the word “model”. It should be noted that the vertical axis is logarithmic.
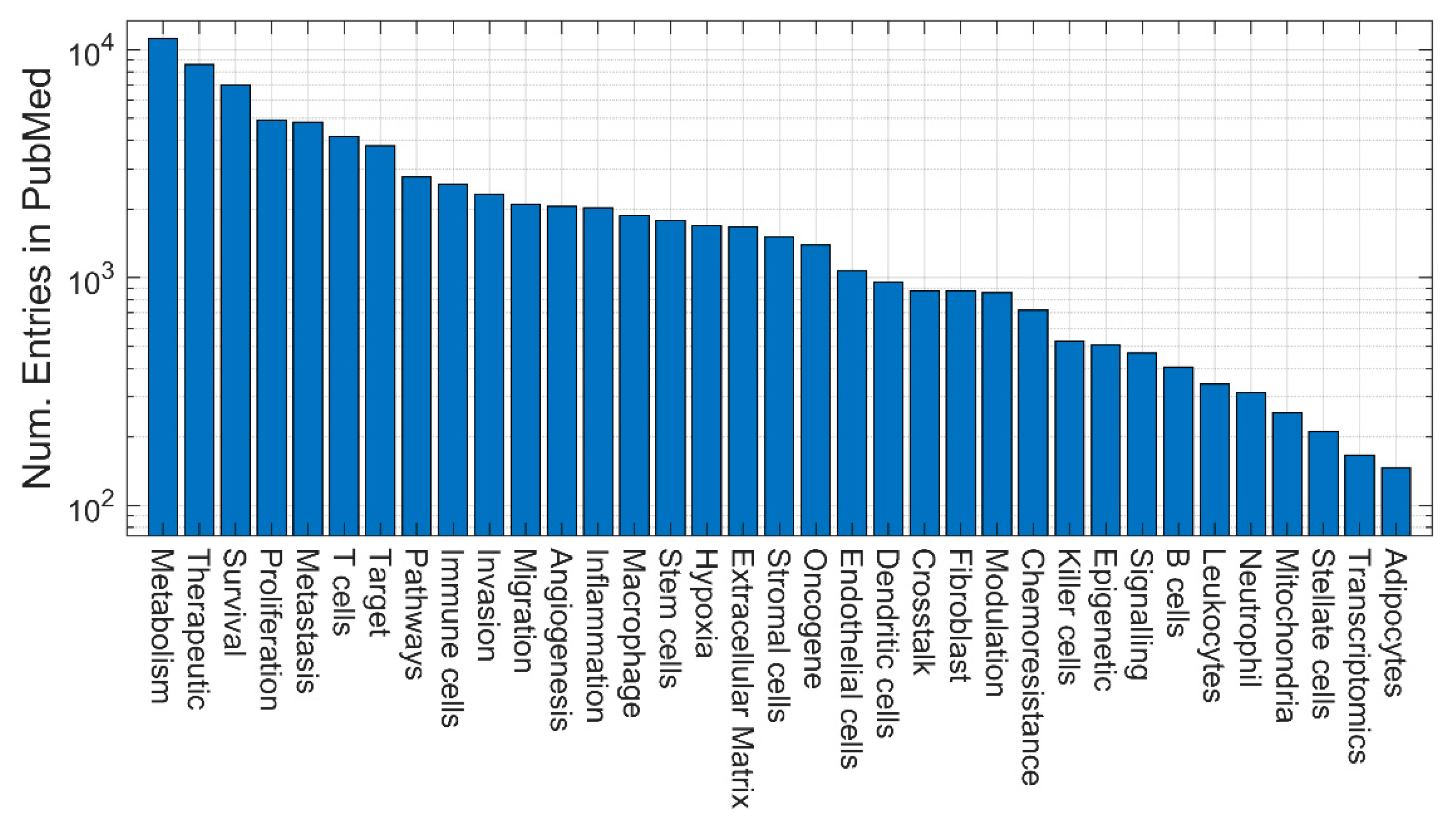
Figure 4.
Number of entries indexed in PubMed for individual queries. In this case, the query included organ-specific keywords.
Figure 4.
Number of entries indexed in PubMed for individual queries. In this case, the query included organ-specific keywords.
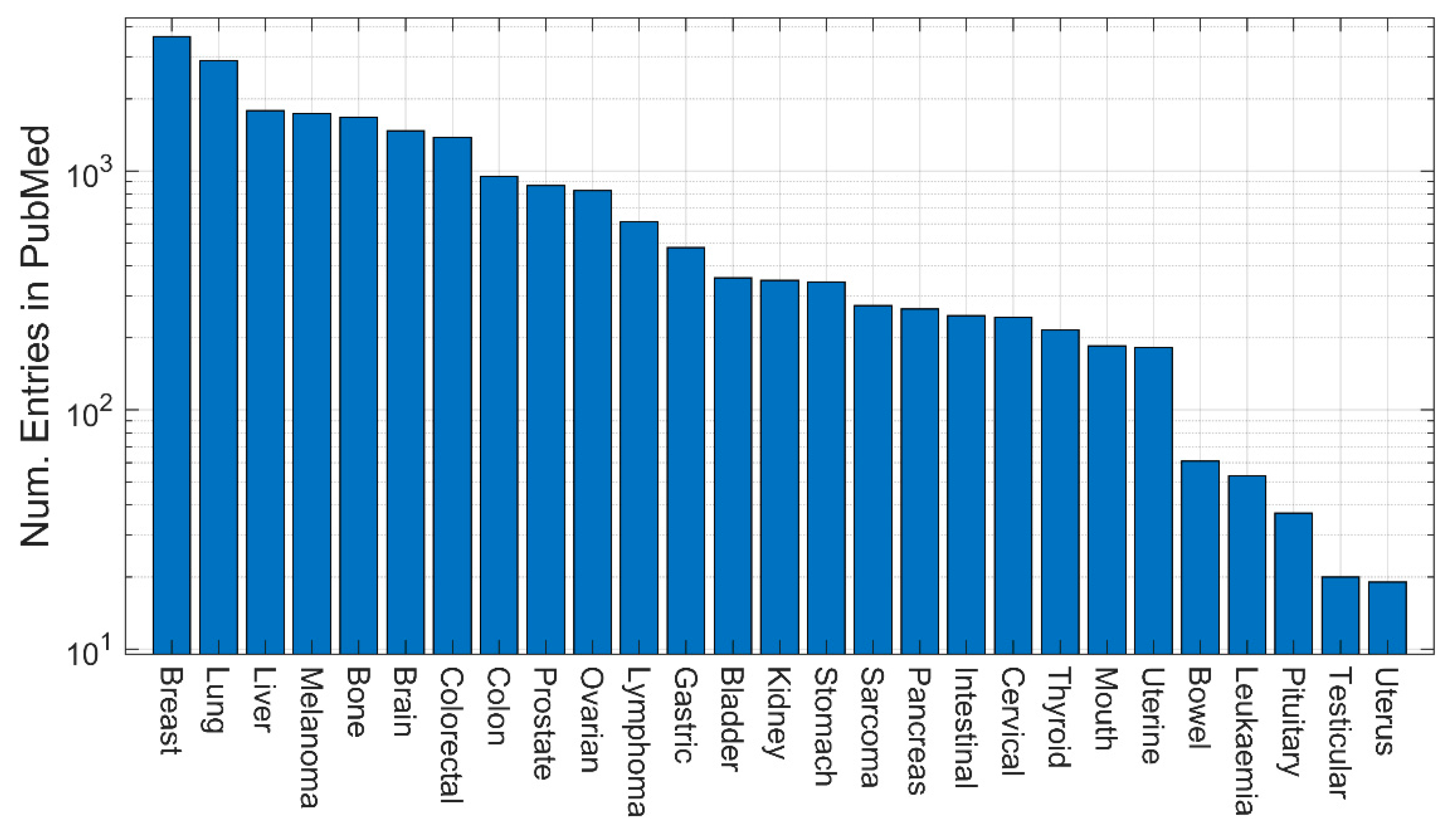
Figure 5.
Number of entries indexed in PubMed for queries with pairs of keywords for models and organs. Each 3D bar corresponds to the number of entries in PubMed for a specific pair of keywords. The colour correspond to those if Figure 1.
Figure 5.
Number of entries indexed in PubMed for queries with pairs of keywords for models and organs. Each 3D bar corresponds to the number of entries in PubMed for a specific pair of keywords. The colour correspond to those if Figure 1.
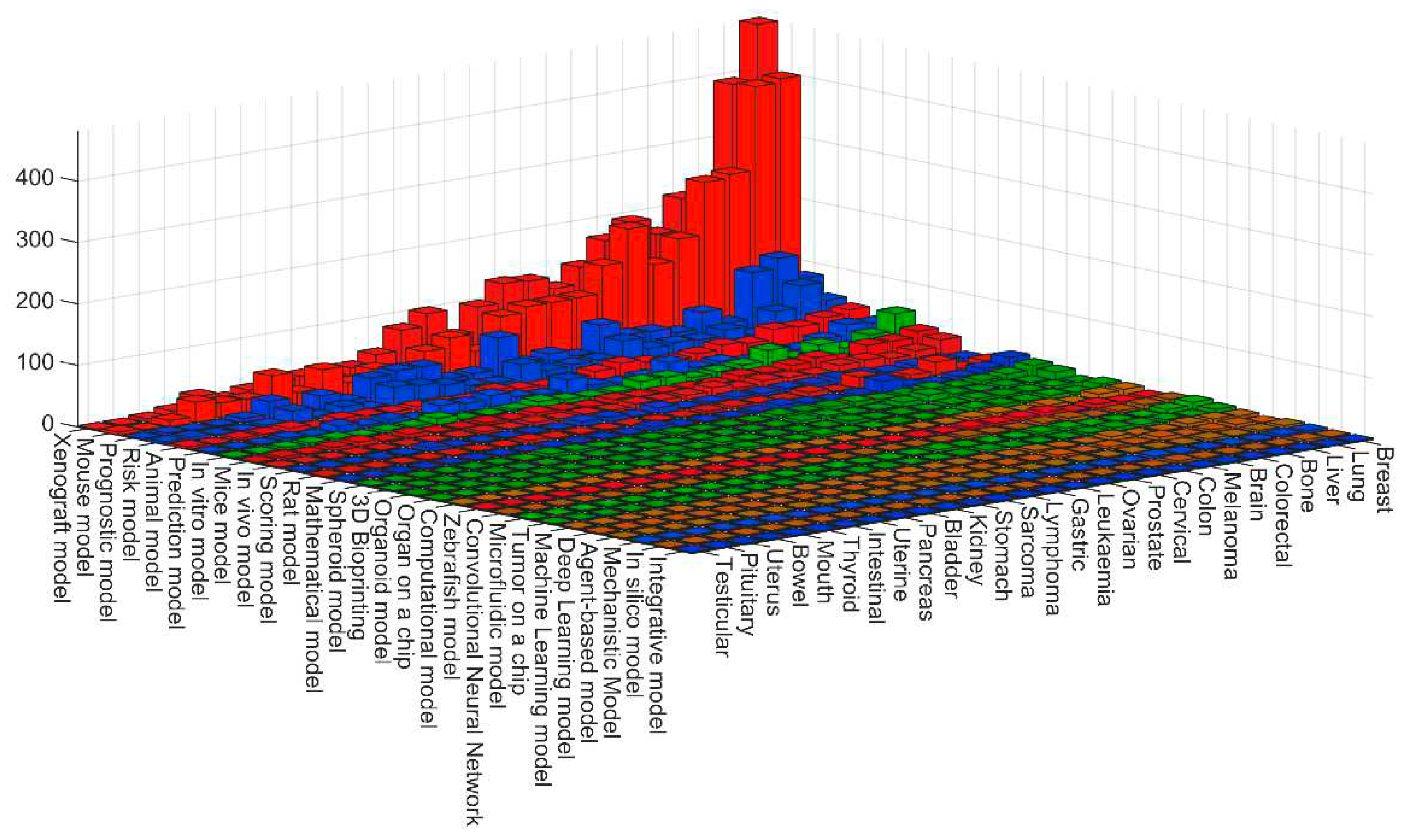
Figure 6.
Number of entries indexed in PubMed for queries with pairs of keywords for models and organs for the cases with highest results from Figure 5.
Figure 6.
Number of entries indexed in PubMed for queries with pairs of keywords for models and organs for the cases with highest results from Figure 5.
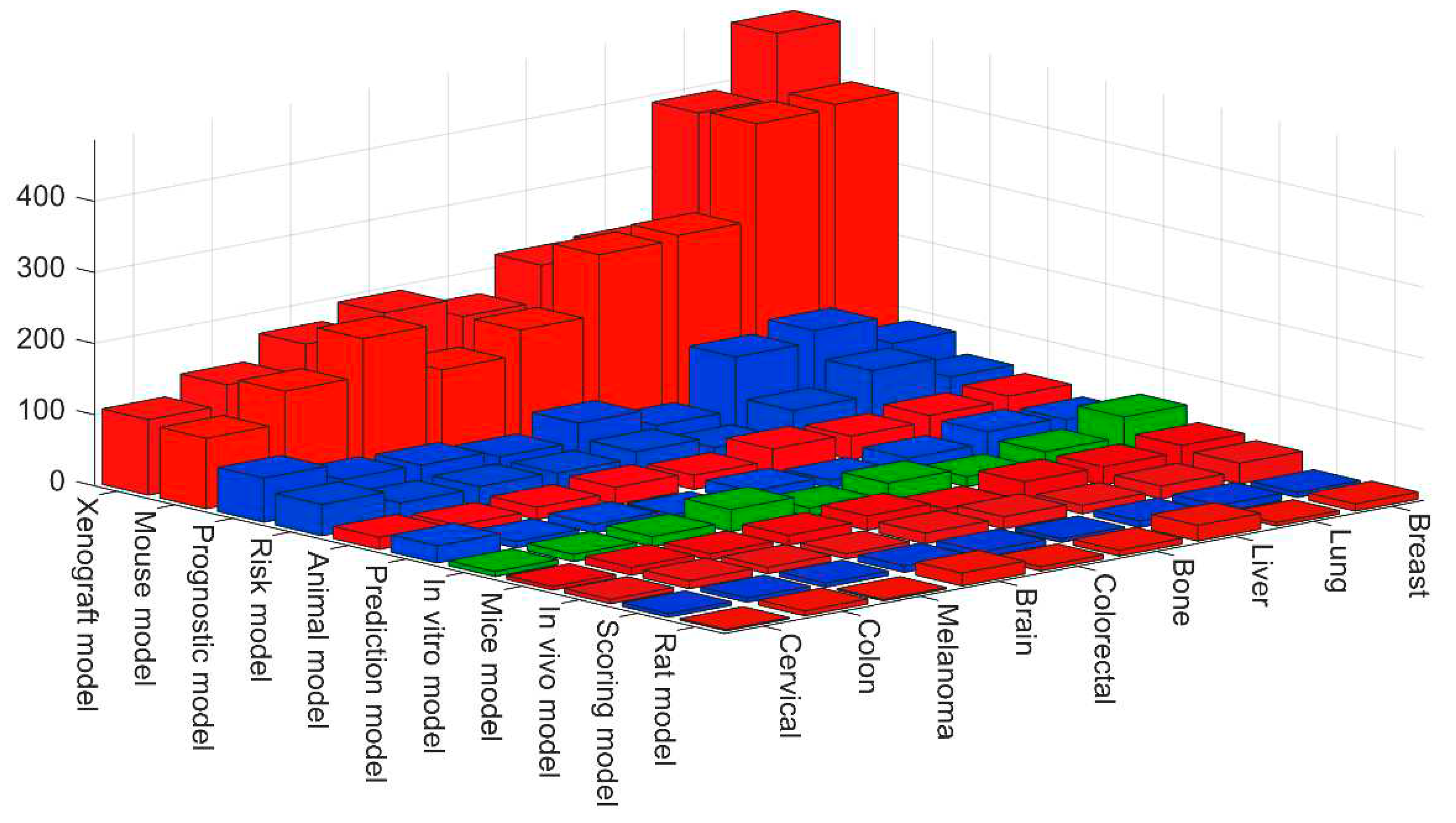

Table 1.
Keywords used to create the queries to mine PubMed grouped according to the four definitions of model.
Table 1.
Keywords used to create the queries to mine PubMed grouped according to the four definitions of model.
| Definition | Keywords |
|---|---|
| Model organism | Animal model. Mouse model. Mice model. Rat model. Zebrafish model. Xenograft model. In vivo model |
| In vitro model | In vitro model. Tumor on a chip. Microfluidic model. 3D Bioprinting. 3D model. Organoid model. Spheroid model. Organ on a chip. |
| Mathematical model | Mechanistic Model. Scoring model. Prediction model. Risk model. Integrative model. Mathematical model. Prognostic model. |
| Computational model | In silico model. Computational model. Deep Learning model. Machine Learning model. Convolutional Neural Network. Agent-based model |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



