
Submitted:
29 June 2023
Posted:
29 June 2023
You are already at the latest version
Abstract
In order to ensure food security worldwide in the face of current climatic changes, a higher quality and quantity of crops is necessary to sustain the growing human population. By developing a sustainable circular economy and biorefinery approaches, we can move from a petroleum-based to a bio-based economy. Plant biostimulants have long been considered an important source of plant growth stimulants in agronomy and agro-industries with both macroalgae (seaweeds) and microalgae (microalgae). There has been extensive exploration of macroalgae biostimulants. A lack of research and high production costs have constrained the commercial implementation of microalgal biostimulants, despite their positive impact on crop growth, development and yields. The current knowledge on potential biostimulatory compounds, key sources and their quantitative information from algae is summarized in the present review. Our goal is to provide a brief overview of the potential for microalgal biostimulants to improve crop production and quality. A number of key aspects are discussed, including the biostimulant effects caused by microalgae extracts, as well as the feasibility and potential for cocultures and coapplication with other biostimulants and biofertilizers. This article also discusses the current knowledge, recent developments and achievements in extraction techniques, types of applications, timings of applications. Ultimately, this review highlights the potential for microalgal biostimulants for sustainable agricultural practices, the algal biochemical components contributing to these traits, and finally bottlenecks and involved prospects in commercializing microalgal biostimulants.
Keywords:
algae biostimulants
; sustainable agriculture
; microalgae
; crop nutrition
; biofertiliser
; crop protection products
1. Introduction
Agro-industry practices and future perspectives are of current relevance, but they face two challenges: improving crop quality and yield due to a growing world population, and minimising impacts on the environment and human health. Besides the growing demand for organic food and growing environmental awareness, the forthcoming regulatory framework will restrict the use of chemical inputs [1,2,3]. In addition, urbanisation, erosion, and adverse effects of climate change further complicate the situation by making farmers worldwide produce more with less due to reaching the genetic potential of stable crops and decreasing fertile land areas [4,5]. It is possible to mitigate these issues with biofertilizers and biostimulants, and they offer a renewable option for improving crop quality and yield. The purpose of biofertilizers is to promote plant growth and development by colonizing the rhizosphere of the plant with microorganisms, such as bacteria, fungi or microalgae, and allowing the plant to absorb nitrogen, phosphorus, potassium, minerals and apply large quantities [6,7]. In contrast, biostimulants are resources that improve crop nutrition, stress tolerance, yield or quality when applied in small quantities without damaging or even improving the surrounding environment when they are applied in small amounts [8,9,10]. Biostimulants increase nutrient efficiency and help plants withstand abiotic stresses, thus enhancing both crop quality and yield. A biostimulant is not a biofertilizer since it does not directly provide nutrients to plants. As a consequence of modifying rhizosphere and plant metabolism, they facilitate nutrient uptake, improving nutrient efficiency, tolerance to abiotic stresses, and crop quality [11]. Humic substances (humic acid, fulvic acid, and humins); algae extracts; protein hydrolysates (signaling peptides and free amino acids); and microorganisms (bacteria, yeast, filamentous fungi, and microalgae) are some of the main plant biostimulants [12]. Macroalgae extracts contain a wide variety of biostimulatory compounds, including amino acids, polysaccharides, vitamins, fatty acids, minerals, phenolics, and phytohormone traces. In the organic plant biostimulant market, macroalgae have been heavily exploited since the early 1980s for their biostimulant potential. In comparison to microalgae, macroalgae have a longer history of biostimulants. The specific mode of biostimulant action of individual bioactives is often unclear due to their diverse compositions and physicochemical properties. Increasing biotechnological advances, including high-throughput phenotyping and -omic platforms, can be used to illustrate the underlying mechanisms of biostimulant action and to develop novel products [12]. As a mainly single-celled photosynthetic organism, microalgae synthesize a wide variety of metabolites using sunlight and carbon dioxide. Currently, microalgae are being explored for use in biofuels, aquaculture and animal feed, bioremediation of waste, nutraceuticals, pharmaceuticals and cosmetics [13]. In the case of agricultural applications, microalgae have not been explored as much as macroalgae. According to traditional research, cyanobacteria (blue-green algae) fix nitrogen in paddy fields and are beneficial for a variety of other crops [14]. Agricultural microalgal biomass is known to act as a biofertilizer and soil conditioner [15,16]. In contrast, living cyanobacteria are known to act as a potential biocontrol agent against plant pathogens, through the activation of plant defence enzymes and the production of hydrolytic enzymes and antimicrobial compounds. In addition, by integrating waste nutrients such as wastewater [17] and anaerobic digestion waste (digestive waste) [18] into microalgal biostimulants, unique opportunities are provided for shaping circular economy platforms. The major emerging concepts identified in a recent literature survey on scientific trends and market opportunities are microalgal bioplastics and biostimulants [19]. Despite this, determining the mechanism of biostimulant action of specific bioactives is still a major challenge due to the variability of algae and crops and the interaction between them based on abiotic factors [12]. It is clear, however, that microalgae have a huge potential, when viewed alongside the promising capabilities of microalgal biostimulants, as well as the increasing cost of chemical fertilizers, pesticide resistance, and climate change. Microalgae can help to make agriculture more sustainable and resilient by providing an immense scope. In this review, we discuss the current state of research on algal biostimulants, their sources, and quantitative data [13]. Phytohormones, proteins, amino acids, humic acids, fulvic acids, polysaccharides, antioxidants, vitamins, and enzymes are among the biostimulants covered. A comparison is also made between the physiological effects and quantitative information on established macroalgal biostimulants to determine what is the best way forward for the microalgal biostimulant industry [14]. It discusses the potential of microalgal biostimulants in improving crop production and quality, including specific biostimulant effects caused by extracts of microalgae, microalgal-algae consortia, microalgal-bacterial consortia, extraction techniques, application types, timing, and current regulatory perspectives.
2. Crop production and quality can be improved by using algal-bacteria and algae consortiums
Research has highlighted the potential of bacterial plant biostimulants, including non-pathogenic bacteria such as Pseudomonas and Bacillus, Azotobacter, Serratia, and Azospirillum. Upon seed or root inoculation, Azospirillum brasilense promotes the growth of many terrestrial plants and improves the yield of a variety of crops [20]. Consequently, it has been proposed that co-immobilized microalgae and PGPB are effective means to increase microalgal populations within confined environments, as well as to improve the biostimulant potential of algal-bacterial consortia. In the spring and summer, such consortia resulted in 18.9% and 12.9% weight increases in romaine lettuce, respectively, and 16.5% and 22.7% increases in leaf lettuce. In contrast to control romaine lettuces, treated romaine lettuces increased their carotenoid content 26.7% during the summer, suggesting that the algal-bacterial consortia reduced the negative effects caused by excess light and heat, resulting in better light-dependent metabolite development. Compared to the control, romaine lettuce had a 2.5-times higher antioxidant capacity for algal-bacterial treatment in the summer [10]. Additionally, it has been shown that salt stress and temperature stress increase antioxidant activity, where elevated concentrations of guaiacol-specific peroxidase were observed in French beans [21]. Despite this implying the plants were stressed due to high summer temperatures, the addition of the biostimulant consortia improved biomass compared to the control plants. Plant yields can also be improved by consortia of algal strains without bacteria [22]. A coculture of Anabaena cylindrica and Nostoc sp. produced the highest levels of exopolysaccharide (EPS). As compared to monocultures, this consortia application achieved the highest plant yield in lettuce crops, indicating the EPS was beneficial [23]. Chrorophyta improve growth and nutritional quality, while cyanobacteria aid in N-fixation, soil quality, and biocontrol. Accordingly, combining the two strains or more is likely to improve agricultural aspects more than combining just one strain, and specific combinations of synergistic traits can be selected, depending on the desired outcome [22]. In order to fully exploit the potential of natural co-culture, it is important to select the right consortia partners strategically [24]. Therefore, further research into different combinations of algae should be conducted to confirm whether they should be cultivated together or whether mutual benefits will still occur if they are cultured separately and applied simultaneously.
3. Biomass production of microalgae
Microalgae can be produced using wastewater, recovering nutrients and preserving water for further use [25,26], making it one of the most rapidly growing activities in the world [27]. As a result, microalgae reduce greenhouse gas emissions by sequestering CO2 and nitrous oxide (N2O) from industrial byproducts. A variety of systems for producing microalgal biomass have been proposed and are being used both in laboratories and in industries [28,29]. Raceway ponds are among the most commonly used methods. In raceway ponds, the water depth is between 10 and 50 cm, which allows for adequate illumination, and the paddle wheel mixes and circulates the gas/medium. As a result of direct exposure to the air, the growing medium evaporates, regulating the temperature of the culture medium. Arthrospira spp., Dunaliella spp., Anabaena spp., Phaeodactylum spp., Pleurochrysis spp., Chlorella spp. and Nannochloropsis spp. are common microalgae and cyanobacteria grown in this system [30]. Due to the better capture of light and the optimal use of cultivated space, photobioreactors typically have higher volumetric productivity than open ponds. Raceway ponds, on the other hand, require less energy to mix culture medium because they are made with less expensive materials [31]. Photobioreactors have better radiant energy utilisation, reduced gas/liquid mass transfer, temperature control, and lower microalgae productivity than open ponds. As a strategy to increase the cost-effectiveness of microalgal production, optimisation of culture medium has been suggested [32]. Microalgal production can be made more cost-effective by employing low-cost resources, such as nutrient-rich wastewater, agricultural by-products, and inexpensive fertilisers [25,33]. In hydroponics, microalgae usually grow spontaneously, but they are considered a critical point because they can cause nutritional competence and pipeline clogs. In contrast, microalgae, through the biochemical process of photosynthesis, can produce oxygen (O2) for crop roots to respire and grow through nutrient solutions. Microalgae could be co-cultivated without any additional inputs, according to the same authors [34]. In addition, Barone et al. [35] suggested the co-production of tomato plants and microalgae (Scenedesmus quadricauda or Chlorella vulgaris). Microalgae growth and chemical composition could be affected by a number of factors, including nutrients, light intensity, pH, and electroconductivity. Moreover, the different characteristics of the growing medium, particularly the source and concentration of nitrogen (N), can affect the growth and biochemical composition of microalgal species [36,37,38]. A variation of the N source influences Arthrospira species' biomass production [39,40]. Meanwhile, the growth phase of the marine microalgae Isochrysis spp. had a greater impact on gross biochemical composition than the source of N [38]. Microalgae productivity might be enhanced by biochemical stimulants such as phytohormones and polyamines [41,42]. For the green alga Chlorella spp., biochemical stimulants were tested for their influence on growth and chlorophyll concentration. This microalga grew significantly faster and produced a greater amount of proteins, saccharides and chlorophylls using natural and synthetic auxins [43].
4. Microalgae as a new source of biostimulants
Agricultural applications of microalgae have long focused on their use as biofertilisers and soil conditioners, whose effects on crops are mainly attributable to the improvement of physical, chemical and biological soil fertility [44]. However, in recent years, numerous studies have shown that the variety of physiological responses in plants following the application of these microbial biomasses cannot be attributed solely to the increase in the nutrients available to the plant, but derives from the action of a wide range of bioactive molecules (e.g. phytohormones, amino acids, vitamins, polysaccharides, carbohydrates, polyamines, polyphenols) that are effective on the plant at concentrations considerably lower than those of the macroelements (such as nitrogen, phosphorous and potassium) contained in biofertilisers [45,46]. The ability of microalgae to produce these bioactive molecules, which plants can absorb and metabolise both foliar and root, and the possibility of improving crop productivity using very small quantities of the product compared to biofertilisers, has led the scientific community and companies to take an interest in studying the biostimulant properties of these microalgae [47]. Considering that most of the results in this field have been published in recent years and that very few microalgae and cyanobacteria-based biostimulant products are available on the market today, we can say that research in this area, although very promising, is still in its infancy [1,45]. Due to the enormous biodiversity of these microorganisms (it is estimated that only half of the approximately 55,000 existing species of microalgae and cyanobacteria have been described to date) [48], only a small number of strains belonging to a few genera have been investigated for their biostimulant properties to date (Figure 1). Most of the products currently on the market are obtained from the cyanobacterium Arthrospira platensis and the green microalgae Chlorella spp. Arthrospira and Chlorella spp. are the two genera most extensively cultivated worldwide for various commercial applications (mainly for the nutraceutical market) and most frequently studied for their biostimulant activities on different plant species, appearing in 49% and 56% of the scientific publications in the field related to cyanobacteria and microalgae.
5. Processes and applications of biostimulating algal biomass
The production of biostimulants from algal biomass and cyanobacteria may involve the use of various techniques mainly aimed at breaking down cells by making bioactive molecules contained in or bound to cell walls available to the plant. This disruption can be achieved through physical/mechanical, chemical or enzymatic methods [47]. The choice of extraction method is mainly dictated by the type of biomass used and the target molecules. For example, the physical/mechanical methods, most commonly used for research purposes today, which involve mechanical cell wall disruption or the use of high pressure, high temperature, ultrasound or combinations thereof, cannot guarantee high extraction yields for micro- and macroalgae, which may have a thicker cell wall than cyanobacteria [45]. Cell disruption may be followed by a phase of separation from the extract of cell residues by centrifugation or filtration, or by an extraction phase using solvents in order to obtain specific fractions of the crude extract [1]. For instance, in the production of biostimulant polysaccharide extracts, polysaccharides are usually precipitated with ethanol following the physical breakdown of cells. A rather recent technique is the extraction using supercritical CO2 as a solvent, i.e. with chemical-physical properties intermediate between a liquid and a gas, obtained at low temperatures (50°C) and under high pressure (200-500 bar), ensuring the preservation of the thermolabile bioactive compounds in the biomass [47]. In the preparation of microalgal and cyanobacterial hydrolysates, the use of chemical agents, mainly acids or bases such as sulphuric acid, hydrochloric acid and sodium hydroxide, generally results in the breakdown of the macromolecules contained in the cells. However, these methods are less and less used as they may lead to the degradation and inactivation of some bioactive molecules contained in the biomass and require the subsequent disposal of large quantities of chemical compounds [49]. Enzymatic methods use single enzymes capable of breaking cell walls and/or proteolytic enzymes that cleave peptide bonds to produce protein hydrolysates, i.e. products rich in free amino acids and soluble peptides. Extracts and hydrolysates can be applied directly to the foliar apparatus by spraying or nebulisation, to the growing medium by fertigation where the active molecules are absorbed by the root system, or they can be used for pre-sowing treatments [44,47]. Foliar application is generally preferred to soil application as it allows the use of lower doses of product, limits losses due to leaching and prevents degradation by soil microorganisms. To avoid the costs associated with extraction/hydrolysis processes, the live cells can be applied directly into the growth medium or onto the plant leaves or be used for seed treatment. Alternatively, the culture medium separated by filtration or centrifugation from the microalgal biomass can be used directly for biostimulant treatment, exploiting the action of the compounds released by the microbial cultures [45].
6. Main biostimulating effects of microalgae on the plant
The application of microalgae and cyanobacteria or formulations derived from them (biomass, extracts, hydrolysates) on plants has been shown to produce a wide range of often interconnected beneficial effects (Table 1). These responses vary according to the microalgal species used to produce the biostimulant, but also in relation to the plant species treated and the growing conditions (Figure 2) [50,51]. Among the most common effects observed is an increase in vegetative growth and, consequently, yield in leafy vegetables (lettuce, spinach, rocket) and aromatic herbs (mint, basil) [45,50]. The increase in plant growth and fresh weight was associated with a stimulation of nitrogen and carbon metabolism in plants treated with microalgal extracts, whereby an increase in leaf content, protein, carbohydrate and photosynthetic pigments (chlorophylls and carotenoids) was observed [52]. The stimulation effects of primary metabolism can be attributed to an increase in nutrient uptake by the plants subjected to the biostimulant treatment. In this sense, biostimulants can act either directly, by improving soil structure and nutrient availability in the soil if applied basally, or by directly influencing plant physiology if applied basally or by foliar application [53]. Indeed, it is known that the inoculation of cyanobacteria in the soil can promote the uptake of zinc and iron by plants through the production of siderophores [54]. Moreover, the extracellular polysaccharides produced by many cyanobacterial species can stimulate the microbiota of the rhizosphere by providing organic carbon for microbial growth and can improve soil aggregation and water retention capacity, increasing the volume of soil that can be explored by roots and indirectly promoting root growth [55]. Stimulation of growth and root development has also been observed in several studies following treatment with microalgal and cyanobacterial extracts and hydrolysates [1,45]. For example, the use of Chlorella vulgaris and Scenedesmus quadricauda on beetroot produced positive effects on root architecture including an increase in root length, but also in the number of lateral roots and thus in the root surface area for nutrient uptake [35]. These stimulation effects occur both when the biostimulant is applied to the basal part of the plant and is absorbed directly by the roots, and when it is applied to the leaves and induces a concomitant increase in the macro- and micro-nutrient content of the plant tissue. In addition to leaf or soil application, seedling growth can also be stimulated following seed treatment in the pre-sowing phase. Seed treatment can also have the effect of increasing the germination percentage [50]. Due to their ability to accelerate germination and seedling development, stimulate early flowering and increase the number of flowers, microalgal and cyanobacterial biostimulants can also find interesting applications in floriculture [50,56]. For instance, aqueous extracts and lyophilised biomass of Desmodesmus subspicatus increased germination in vitro and accelerated development in the subsequent transplanting and acclimatisation phase in the greenhouse of the orchid Cattleya warneri [57]. The effect of biostimulants on the plant does not only result in improved vegetative growth. The effect of biostimulants on the plant does not only result in improved vegetative growth. For example, treatments on wheat and vines have led to a significant increase in grape yield [50]. Furthermore, the application of biostimulants can trigger biochemical processes that lead to the accumulation of important metabolites resulting in improved quality traits and shelf life of the final product [58,59]. Among these, we may mention the increase in leaf content in essential oils in peppermint treated with extracts of Anabaena vaginicola and Cylindrospermum michailovskoense [60], and the increase in total soluble solids content and reduction in weight loss during storage of onions treated with extracts of Arthrospira platensis [61]. Although the incidence of abiotic stresses, such as drought, salinity and extreme temperatures, is set to increase in the years to come as climate change phenomena intensify, few strategies are available to date to mitigate the negative effects of such stresses [62]. Many abiotic factors manifest themselves in plants as osmotic stresses, leading to the accumulation of reactive oxygen species (ROS) that can cause severe oxidative damage to DNA, lipids, carbohydrates and proteins [63]. The application of microalgae and cyanobacteria in formulations derived from them has been shown to promote the growth and yield of certain plant species such as rice, wheat and tomato under abiotic stress conditions, inducing an enhancement of antioxidant defences in plant tissues [45,50]. In rice plants inoculated with cyanobacteria, an accumulation of phenolic acids and flavonoids was observed in leaf tissue [46]. Furthermore, according to recent studies, polysaccharide extracts obtained from different microalgal strains including Chlamydomonas reinhardtii, Chlorella rokiniana, Porphirydium spp. and Dunaliella salina, can increase the activity of antioxidant enzymes such as catalase, peroxidase and superoxide dismutase in tomato plants subjected to salt stress [58]. Another important function of exopolysaccharides released into the soil is to sequester metal ions and sodium ions, reducing their uptake by plants and stimulating their growth in saline or polluted soils [64,65]. For example, seed coating on maize with Arthrospira platensis led to a reduction of more than 90% of cadmium absorbed by the roots 12 days after sowing [66]. Finally, it is important to remember that the concentration of the biostimulant and the number of applications are a determining factor in the success of the treatment and that an increase in the dose does not always correspond to an increase in the positive effects on the plant [67]. In fact, it has been found in some studies that intermediate dilutions of biostimulant can be more effective in promoting growth and flowering, while the application of high doses usually reduces or even neutralises the effect. Effective doses may vary considerably depending on the plant species treated and the method of application. Generally, foliar application is effective at lower concentrations than seed or soil application [45].
7. Advantages and critical issues in the use of microalgae for biostimulants
Although there is increasing scientific evidence on the beneficial effects of using microalgae (Figure 3), their application in agriculture is still very limited and very few products are currently on the market, especially when compared to the large number of macroalgae products [1]. One of the main obstacles to be overcome in the commercial exploitation of microalgae relates to the cost of producing biomass, which is generally higher than for macroalgae [45]. Indeed, microalgae are cultivated in controlled and confined systems (photobioreactors and tanks) that require significant amounts of electricity, fertilisers, water and materials for construction and operation. On the other hand, cultivation in a controlled environment is also one of the main advantages of using microalgal biomass for biostimulant production, as it allows for greater standardisation of production processes [76]. Macroalgal biomasses, on the other hand, have biochemical and functional characteristics that can vary considerably depending on the phenological stage, environmental conditions and nutrient availability at the time of harvest, and are therefore more difficult to standardise. In order to make microalgae biostimulants more competitive with other products on the market, it will be necessary to reduce biomass production costs, e.g. by supplementing cultivation with wastewater treatment, using waste CO2 or cultivating thermotolerant strains that do not require cooling of the crop [77,78]. Ideally, the production of biostimulants from microalgae can be integrated with the production of other products. For example, the residual pellets from extraction could be used as a biofertiliser, or the remaining lipid fraction could be used for the production of biofuels, to obtain polyunsaturated fatty acids with various cosmetic, medical and nutraceutical applications, or polyhydroxyalkanoates used for the production of bioplastics [45]. Residual proteins could be used to formulate food or feed for animal husbandry and aquaculture. However, the design of an efficient biorefinery system requires that the fractions that contribute most to the biostimulating action be clearly identified, in order to assess the possible reuses of the remaining fractions [45].
8. Conclusions
Microalgae are to date considered one of the most promising sources for the development of biostimulant products as their wide genetic and metabolic biodiversity is still far from being adequately explored. In recent years, scientific evidence has accumulated to support the biostimulant action of these microorganisms and their derived formulations, which have been shown to increase yields while improving the efficiency of fertilisation treatments by stimulating nutrient uptake by the plant. Furthermore, the application of microalgal biostimulants has been shown to reduce the negative effects on vegetative growth caused by abiotic stresses, increasing the resilience of crops to climate change. The success of a biostimulant treatment depends on many factors including the microalgal species used, the method of biomass processing, the plant species on which it is applied, and the concentration and method of application. It follows that the development of a new biostimulant must involve several stages, ranging from in vitro screening on model species to identify potentially bioactive strains, to extensive agronomic testing on different plant species to determine the most convenient and effective doses, timing and mode of application depending on the species and crop conditions. Although the development and commercialisation of a biostimulant does not currently require a clear demonstration of the mechanisms of action, a greater understanding of how different bioactive molecules affect plant physiology would make it possible to accelerate the selection of new biostimulant strains and to plan strategies aimed at increasing the concentration of the bioactive compounds of interest in microalgal biomass, paving the way for a new category of biostimulants characterised by greater standardisation and reliability and greater efficacy on crops.
Author Contributions
Conceptualization, D.P.; methodology, writing—original draft preparation D.P. and D.S.; software and investigation, D.P.; writing—review and editing, D.P.; funding acquisition, D.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the CREA Research Centre for Vegetable and Ornamental Crops.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data, tables, and figures in this manuscript are original.
Acknowledgments
The author would like to express his heartfelt gratitude to his colleagues at CREA Research Centre for Vegetable and Ornamental Crops in Pescia and to all other sources for their cooperation and guidance in writing this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chiaiese, P.; Corrado, G.; Colla, G.; Kyriacou, M.C.; Rouphael, Y. Renewable sources of plant biostimulation: microalgae as a sustainable means to improve crop performance. Front. Plant Sci. 2018, 9, 1782. [Google Scholar] [CrossRef] [PubMed]
- Dmytryk, A.; Chojnacka, K. Algae as fertilizers, biostimulants, and regulators of plant growth. In: Algae Biomass: Characteristics and Applications. Springer International Publishing. 2018, 115–122.
- Mzibra, A.; Aasfar, A.; Benhima, R.; Khouloud, M.; Boulif, R.; Douira, A.; Bamouh, A.; Meftah Kadmiri, I. Biostimulants derived from moroccan seaweeds: seed germination metabolomics and growth promotion of tomato plant. J. Plant Growth Regul. 2020, 1–18. [Google Scholar] [CrossRef]
- Povero, G.; Mejia, J.F.; Di Tommaso, D.; Piaggesi, A.; Warrior, P. A systematic approach to discover and characterize natural plant biostimulants. Front. Plant Sci. 2016, 7, 435. [Google Scholar] [CrossRef] [PubMed]
- Shukla, P.S.; Borza, T.; Critchley, A.T.; Prithiviraj, B. Carrageenans from red seaweeds as promoters of growth and elicitors of defense response in plants. Front. Mar. Sci. 2016, 3, 81. [Google Scholar] [CrossRef]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed extracts as biostimulants in horticulture. Sci. Hortic. (Amsterdam). 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Win, T.T.; Barone, G.D.; Secundo, F.; Fu, P. Algal biofertilizers and plant growth stimulants for sustainable agriculture. Ind. Biotechnol. 2018, 14, 203–211. [Google Scholar] [CrossRef]
- Arnau, L.; Richard, H. Techno-Economic Feasibility Study for the Production of Microalgae Based Plant Biostimulant. KTH Royal Institute of Technology. 2016; pp. 300–365.
- Barone, V.; Baglieri, A.; Stevanato, P.; Broccanello, C.; Bertoldo, G.; Bertaggia, M.; Cagnin, M.; Pizzeghello, D.; Moliterni, V.M.C.; Mandolino, G.; Fornasier, F.; Squartini, A.; Nardi, S.; Concheri, G. Root morphological and molecular responses induced by microalgae extracts in sugar beet (Beta vulgaris L. ). J. Appl. Phycol. 2018, 30, 1061–1071. [Google Scholar] [CrossRef]
- Kopta, T.; Pavlíkova, M.; Se¸ kara, A.; Pokluda, R.; Marˇsa´lek, B. Effect of bacterial- algal biostimulant on the yield and internal quality of Lettuce (Lactuca sativa L. ) produced for spring and summer crop. Not. Bot. Horti Agrobot. Cluj-Napoca. 2018, 46, 615–621. [Google Scholar] [CrossRef]
- Drobek, M.; Frąc, M.; Cybulska, J. Plant biostimulants: importance of the quality and yield of horticultural crops and the improvement of plant tolerance to abiotic stress—A review. Agronomy. 2019, 9, 335. [Google Scholar] [CrossRef]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants application in horticultural crops under abiotic stress conditions. Agronomy. 2019, 9, 306. [Google Scholar] [CrossRef]
- Chanda, M. J.; Merghoub, N.; EL Arroussi, H. Microalgae polysaccharides: the new sustainable bioactive products for the development of plant bio-stimulants? World J. Microbiol. Biotechnol. 2019; pp. 411–442.
- Chakdar, H.; Jadhav, S.D.; Dhar, D.W.; Pabbi, S. Potential applications of blue green algae. J. Sci. Ind. Res. 2012, 71, 13–20. [Google Scholar]
- Dasgan, H.Y.; Aydoner, G.; Akyol, M. Use of some microorganisms as bio- fertilizers in soilless grown squash for saving chemical nutrients. In: Acta Horticulturae. International Society for Horticultural Science. 2012, 155–162.
- Garcia-Gonzalez, J.; Sommerfeld, M. Biofertilizer and biostimulant properties of the microalga Acutodesmus dimorphus. J. Appl. Phycol. 2016, 28, 1051–1061. [Google Scholar] [CrossRef]
- Renuka, N.; Prasanna, R.; Sood, A.; Bansal, R.; Bidyarani, N.; Singh, R.; Shivay, Y.S.; Nain, L.; Ahluwalia, A.S. Wastewater grown microalgal biomass as inoculants for improving micronutrient availability in wheat. Rhizosphere. 2017, 3, 150–159. [Google Scholar] [CrossRef]
- Stiles, W.A.V.; Styles, D.; Chapman, S.P.; Esteves, S.; Bywater, A.; Melville, L.; Silkina, A.; Lupatsch, I.; Grünewald, C.F.; Lovitt, R. Using microalgae in the circular economy to valorise anaerobic digestate: challenges and opportunities. Bioresour. Technol. 2018, 267, 732–742. [Google Scholar] [CrossRef] [PubMed]
- Rumin, J.; Nicolau, E.; Gonçalves de Oliveira Junior, R.; Fuentes-Grünewald, C.; Flynn, K.J.; Picot, L. European commission. Mar. Drugs. 2020, 18, 79. [Google Scholar] [CrossRef]
- Gonzalez, L.E.; Bashan, Y. Increased growth of the microalga chlorella vulgariswhen coimmobilized and cocultured in alginate beads with the plant-growth-promoting bacterium Azospirillum brasilense. Appl. Environ. Microbiol. 2000, 66, 1527–1531. [Google Scholar] [CrossRef]
- Babu, N.R.; Devaraj, V.R. High temperature and salt stress response in French bean (Phaseolus vulgaris). Aust. J. Crop. Sci. 2008, 2, 40–48. [Google Scholar]
- Renuka, N.; Guldhe, A.; Prasanna, R.; Singh, P.; BuX, F. Microalgae as multi- functional options in modern agriculture: current trends, prospects and challenges. Biotechnol. Adv. 2018, 36, 1255–1273. [Google Scholar] [CrossRef]
- Xue, C.; Wang, L.; Wu, T.; Zhang, S.; Tang, T.; Wang, L.; Zhao, Q.; Sun, Y. Characterization of co-cultivation of cyanobacteria on growth, productions of polysaccharides and extracellular proteins, nitrogenase activity, and photosynthetic activity. Appl. Biochem. Biotechnol. 2017, 181, 340–349. [Google Scholar] [CrossRef]
- Padmaperuma, G.; Kapoore, R.V.; Gilmour, D.J.; Vaidyanathan, S. Microbial consortia: a critical look at microalgae co-cultures for enhanced biomanufacturing. Crit. Rev. Biotechnol. 2018, 38, 690–703. [Google Scholar] [CrossRef]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for biodiesel production and other applications: A review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef]
- Hultberg, M.; Carlsson, A.S.; Gustafsson, S. Treatment of drainage solution from hydroponic greenhouse production with microalgae. Bioresour. Technol. 2013, 136, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Brennan, L.; Owende, P. Biofuels from microalgae—A review of technologies for production, processing, and extractions of biofuels and co-products. Renew. Sustain. Energy Rev. 2010, 14, 557–577. [Google Scholar] [CrossRef]
- Carvalho, A.P.; Meireles, L.A.; Malcata, X. Microalgal reactors: A review of enclosed system designs and performances. Biotechnol. Prog. 2006, 22, 1490–1506. [Google Scholar] [CrossRef] [PubMed]
- Ugwu, C.U.; Aoyagi, H.; Uchiyama, H. Photobioreactors for mass cultivation of algae. Bioresour. Technol. 2008, 99, 4021–4028. [Google Scholar] [CrossRef]
- Moheimani, N.R.; Borowitzka, M.A. The long-term culture of the coccolithophore Pleurochrysis carterae (Haptophyta) in outdoor raceway ponds. J. Appl. Phycol. 2006, 18, 703–712. [Google Scholar] [CrossRef]
- Pushparaj, B.; Pelosi, E.; Tredici, M.R.; Pinzani, E.; Materassi, R. As integrated culture system for outdoor production of microalgae and cyanobacteria. J. Appl. Phycol. 1997, 9, 113–119. [Google Scholar] [CrossRef]
- Lee, Y.K. Microalgal mass culture systems and methods: Their limitation and potential. J. Appl. Phycol. 2001, 13, 307–315. [Google Scholar] [CrossRef]
- Gong, Y.; Jiang, M. Biodiesel production with microalgae as feedstock: From strains to biodiesel. Biotechnol. Lett. 2011, 33, 1269–1284. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.; Zhou, Q. Co-cultivation of Chlorella spp. and tomato in a hydroponic system. Biomass Bioenerg. 2017, 97, 132–138. [Google Scholar] [CrossRef]
- Barone, V.; Puglisi, I.; Fragalà, F.; Piero, A.R.L.; Giuffrida, F.; Baglieri, A. Novel bioprocess for the cultivation of microalgae in hydroponic growing system of tomato plants. J. Appl. Phycol. 2019, 31, 465–470. [Google Scholar] [CrossRef]
- Brown, M.R.; Jeffrey, S.W.; Volkman, J.K.; Dunstan, G.A. Nutritional properties of microalgae for mariculture. Aquaculture 1997, 151, 315–331. [Google Scholar] [CrossRef]
- Kaplan, D.; Richmond, A.E.; Dubinsky, Z.; Aaronson, S. Algal nutrition. In Handbook of Microalgal Mass Culture; Richmond, A., Ed.; CRC Press: Boca Raton, FL, USA, 1986; pp. 147–198. [Google Scholar]
- Levasseur, M.; Thompson, P.A.; Harrison, P.J. Physiological acclimation of marine phytoplankton to different nitrogen sources 1. J. Phycol. 1993, 29, 587–595. [Google Scholar] [CrossRef]
- Danesi, E.D.G.; Rangel-Yagui, C.O.; de Carvalho, J.C.M.; Sato, S. An investigation of effect of replacing nitrate by urea in the growth and production of chlorophyll by Spirulina platensis. Biomass Bioenerg. 2002, 23, 261–269. [Google Scholar] [CrossRef]
- Colla, L.M.; Reinehr, C.O.; Reichert, C.; Costa, J.A.V. Production of biomass and nutraceutical compounds by Spirulina platensis under different temperature and nitrogen regimes. Bioresour. Technol. 2007, 98, 1489–1493. [Google Scholar] [CrossRef]
- Czerpak, R.; Bajguz, A.; Piotrowska, A.; Dobrogowska, R.; Matejczyk, W.; Wieslawski, W. Biochemical activity of di- and polyamines in the green alga Chlorella vulgaris Beijerinck (Chlorophyceae). Acta Soc. Bot. Pol. 2003, 72, 19–24. [Google Scholar] [CrossRef]
- Hourmant, A.; Mereau, N.; Penot, M.; Cann, C.; Caroff, J. Influence of polyamines on growth and metabolism of Dunaliella primolecta. Acta Bot. Neerl. 1994, 43, 129–136. [Google Scholar] [CrossRef]
- Czerpak, R.; Krotke, A.; Mical, A. Comparison of stimulatory effect of auxins and cytokinins on protein, saccharides and chlorophylls content in Chlorella pyrenoidosa Chick. Pol. Arch. Hydrobiol. 1999, 46, 71–82. [Google Scholar]
- Carillo, P.; Ciarmiello, L.F.; Woodrow, P. , Corrado, G., Chiaiese, P., Rouphael, Y. Enhancing sustainability by improving plant salt tolerance through macro- and microalgal biostimulants. Biology. 2020, 9, 253. [Google Scholar] [CrossRef]
- Santini, G.; Biondi, N.; Rodolfi, L.; Tredici, M.R. Plant biostimulants from cyanobacteria: an emerging strategy to improve yields and sustainability in agriculture. Plants. 2021, 10, 643. [Google Scholar] [CrossRef]
- Singh, J.S.; Kumar, A.; Rai, A.N.; Singh, D.P. Cyanobacteria: a precious bio-resource in agriculture, ecosystem, and environmental sustainability. Front. Microbiol. 2016, 7, 529. [Google Scholar] [CrossRef] [PubMed]
- Ronga, D.; Biazzi, E.; Parati, K. , Carminati, D., Carminati, E., Tava, A. Microalgal biostimulants and biofertilisers in crop productions. Agronomy. 2019, 9, 192. [Google Scholar] [CrossRef]
- Guiry, M.D. How many species of algae are there? J.Phycol. 2012, 48, 1057–1063. [Google Scholar] [CrossRef]
- Rachidi, F.; Benhima, R. , Sbabou, L.; El Arroussi, H. Microalgae polysaccharides biostiluating effect on tomato plants: growth and metabolic distribution. Biotechnol. Rep. 2020, 25, e00426. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, A.L.; Weyers, S.L.; Goemann, H.M.; Peyton, B.M.; Gardner, R.D. Microalgae, soil and plants: a critical review of microalgae as renewable resources for agriculture. Algal Research. 2021, 54, 102200. [Google Scholar] [CrossRef]
- Toscano, S.; Romano, D.; Massa, D.; Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulant applications in low input horticultural cultivation systemes. Italus hortus. 2018, 25, 27–36. [Google Scholar] [CrossRef]
- Puglisi, I.; La Bella, E.; Rovetto, E.I.; Lo Piero, A.R.; Baglieri, A. Biostimulant effect and biochemical response in lettuce seedlings treated with a Scenedesmus quadricauda extract. Plants. 2020, 9, 123. [Google Scholar] [CrossRef]
- Halpern, M.; Bar-Tal, A.; Ofek, M.; Minz, D.; Muller, T.; Yermiyahu, U. The use of biostimulants for enhancing nutrient uptake. Adv. Agron. 2015, 130, 141–174. [Google Scholar]
- Manjunath, M.; Kanchan, A.; Rajan, K.; Venkatachalam, S.; Prasanna, R.; Ramakrishnan, B.; Hossain, F.; Nain, L.; Shivay, Y.S.; Rai, A.B. Beneficial cyanobacteria and eubacteria synergistically enhance bioavailability of soil nutrients and yield of okra. Heliyon. 2016, 2, e00066. [Google Scholar] [CrossRef]
- Adessi, A.; de Carvalho, R.C.; De Philippis, R.; Branquinho, C.; da Silva, J.M. Microbial extracellular polymeric substances improve water retention in dryland biological soil crusts. Soil Biol. Biochem. 2018, 116, 67–69. [Google Scholar]
- Plaza, B.M.; Gomez-Serrano, C.; Acien-Fernandez, F.G.; Jimenez-Becker, S. Effetc of microalgae hydrolysate foliar application (Arthrospira platensis and Scenedesmus sp. ) on Petunia x hybrida growth. Environ. Boil. Fishes. 2018, 30, 2359–2365. [Google Scholar]
- Navarro, Q.R.; de Oliveira Correa, D.; Behling, A.; Noseda, M.D.; Amano, E.; Mamoru Suzuki, R.; Lopes Fortes Ribas, L. Efficient use of biomass and extract of the microalga Desmodesmus subspicatus (Scenedesmaceae) in asymbotic seed germination and seedling development of the orchid Cattleya warneri. J. Appl. Phycol. 2021, 2, 134–141. [Google Scholar]
- Mutale-Joan, C.; Redouane, B.; Najib, E.; Yassine, K.; Lyamlouli, K.; Laila, S.; Zeroual, Y.; El Arroussi, H. Screening of microalgae liquid extracts for their biostimulant properties on plant growth, nutrient uptake and metabolite profile of Solanum lycopersicum L. Sci. Rep. 2020, 10, 2820. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.; Baptistella, J.L.C.; Horz, D.C.; Bortolato, L.M.; Mazzafera, P. Organic plant biostimulants and fruit quality- A review. Agronomy. 2020, 10, 98. [Google Scholar] [CrossRef]
- Shariatmadari, Z.; Riahi, H.; Abdi, M.; Hashtroudi, M.S.; Ghassempour, A.R. Impact of cyanobacterial ectracts on the growth and oil content of the medicinal plant Menthas piperita L. Environ. Boil. Fishes. 2015, 27, 2279–2287. [Google Scholar]
- Geries, L.S.M.; Elsadany, A.Y. Maximizing growth and productivity of onion (Allium cepa L.) by Spirulina platensis extract and nitrogen fixing endophyte Pseudomonas stutzeri. Arch. Microbiol. 2021, 203, 169–181. [Google Scholar] [CrossRef]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed extracts as biostimulants in horticulture. Sci hortic. 2015, 196, 38–48. [Google Scholar] [CrossRef]
- Panda, D.; Pramanik, K.; Nayak, B.R. Use of seaweed extracts as plant growth regulators for sustainable agriculture. Int. J. Bioresource Stress Manag. 2012, 3, 404–411. [Google Scholar]
- De Philippis, R.; Colica, G.; Micheletti, E. Exopolysaccharide-producing cyanobacteria in heavy metal removal from water: Molecular basis and practical applicability of the biosorption process. Appl. Microbial. Biotechnol. 2011, 92, 697–708. [Google Scholar] [CrossRef]
- Arora, M.; Kaushik, A.; Rani, N. Kaushik, C.P. Effect of cyanobacterial exopolysaccharides on salt stress alleviation and seed germination J. Environ. Biol. 2010, 31, 701–704. [Google Scholar]
- Seifikalhor, M.; Hassani, S.B.; Aliniaeifard, S. Seed priming by cyanobacteria (Spirulina platensis ) and Salep Gum enhances tolerance of Maize plant against Cadmium toxicity. J. Plant Growth Regul. 2019, 39, 1009–1021. [Google Scholar] [CrossRef]
- Godlewska, K.; Michalak, I.; Pacyga, P.; Basladynska, S. , Chojnacka, K. Potential applications of cyanobacteria: Spirulina platensis filtrates and homogenates in agriculture. World J. Microbiol. Biotechnol. 2019, 35, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Mogor, A.F.; Ordog, V.; Pace Pereira Lima, G.; Molnar, Z.; Mogor, G. Biostimulant properties of cyanobacterial hydrolysate related to polyamines. J. Appl. Phycol. 2018, 30, 453–460. [Google Scholar] [CrossRef]
- Tuhy, L. Samoraj, M.; Witkowska, Z.; Chojnacka, K. Biofortification of maize with micronutrients by Spirulina. OpenChem. 2015, 13, 1119–1126. [Google Scholar]
- Dias, G.A.; Rocha, R.H.C.; Araujo, J.L.; de Lima, J.F.; Guedes, W.A. Growth, yield, and postharvest quality in eggplant produced under different foliar fertilizer (Spirulina platensis) treatments. Semina: Ciencias Agrarias, Londrina. 2016, 6, 3893–3902. [Google Scholar] [CrossRef]
- Coppens, J.; Grunert, O.; Van Den Hende, S.; Vanhoutte, I.; Boon, N.; Haesaert, G.; De Gelder, L. The use of microalgae as a high-value organic slow-release fertiliser results in tomatoes with increased carotenoid and sugar levels. J.Appl. Phycol. 2016, 28, 2367–2377. [Google Scholar] [CrossRef]
- Ertani, A.; Sambo, P.; Nicoletto, C.; Santagata, S.; Schiavon, M.; Nardi, S. The use of organic biostimulants in hot pepper plants to help low input sustainable agriculture. Chem. Biol. Technol. Agric. 2015, 2, 11. [Google Scholar] [CrossRef]
- Osman, M.E.H.; Abo-shady, A.M.; El-Nagar, M.M.F. Cyanobacterial Arthrospira (Spirulina platensis) as safener against harmaful effects of fusilade herbicide on faba bean plant. Rend. Fis. Acc. Lincei. 2016, 27, 455–462. [Google Scholar] [CrossRef]
- Shalaby, T.A.; El-Ramady, H. Effect of foliar application of biostimulants on growth, yield, components, and storability of garlic (Allium sativum L.). Aust. J. Crop. Sci. 2014, 8, 271. [Google Scholar]
- Gemin, L.G.; Mogor, A.F.; De Oliveira Amatussi, J.; Mogor, G. Microalgae associated to humic acid as a novel biostimulant improving onion growth and yield. Sci. Horticult. 2019, 256, 108560. [Google Scholar] [CrossRef]
- Tredici, M.R. Mass production of microalgae: Photobioreactors. In: A. Richmond (Ed.), Handbook of Microalgal Culture: Biotechnology and Applied Phycology. 2004; pp. 178–214.
- Valderrama, D.; Cai, J.; Hishamunda, N.; Ridler, N.; Neish, I.C.; Hurtado, A.Q.; Msuya, F.E.; Krishnan, M.; Narayanakumar, R.; Kronen, M.; Robledo, D.; Gasca-Leyva, E.; Fraga, J. The economics of Kappaphycus seaweed cultivation in developing countries: a comparative analysis of farming systems. Aquac. Econ. Manag. 2015, 19, 251–277. [Google Scholar] [CrossRef]
- Fortser, J.; Radulovich, R. Seaweed and food security. Seaweed sustainability: Food and Non-Food applications, Elsevier. 2015; pp. 289–313.
Figure 1.
Genera of microalgae studied to date for their biostimulant activities with in vivo tests, listed in descending order by number of published papers, and currently on the market for biostimulant products.
Figure 1.
Genera of microalgae studied to date for their biostimulant activities with in vivo tests, listed in descending order by number of published papers, and currently on the market for biostimulant products.
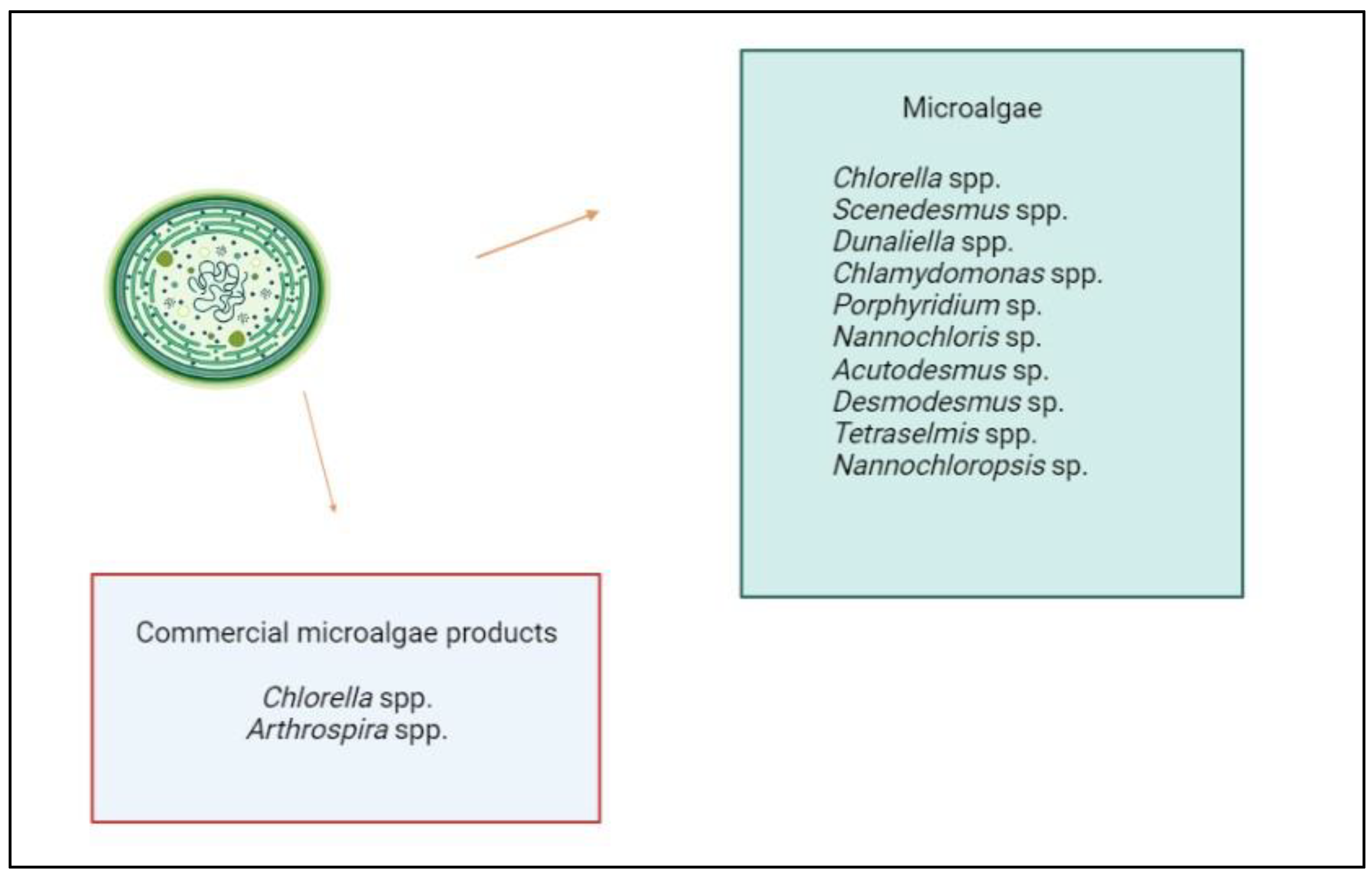
Figure 2.
Positive effects of the application of microalgal biostimulants on plant growth and physiology according to the literature.
Figure 2.
Positive effects of the application of microalgal biostimulants on plant growth and physiology according to the literature.
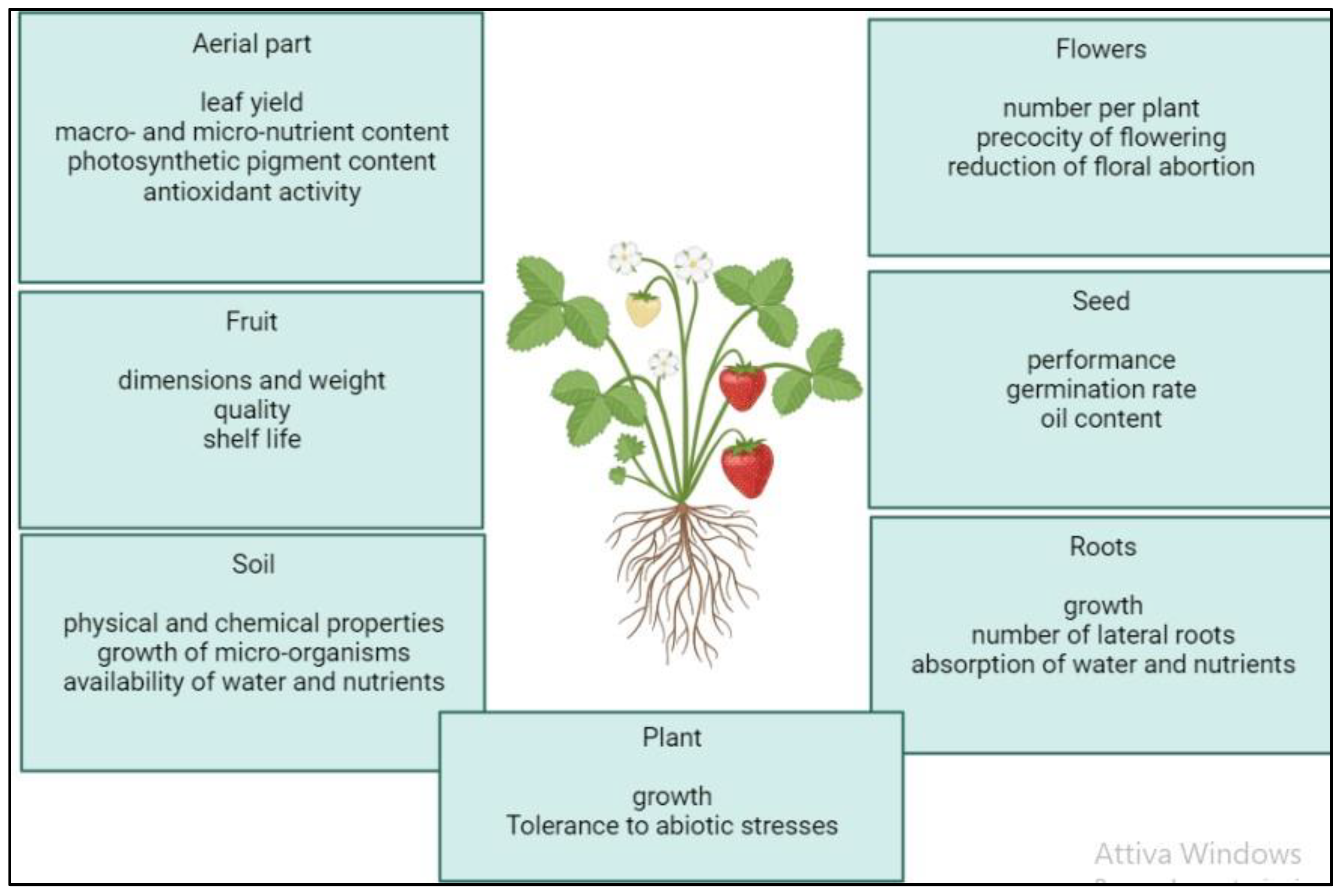
Figure 3.
Positive effect on the vegetative development of lettuce (Lactuca sativa L.) of the microalgae Chlorella spp., Arthrospira spp. and Scenedesmus spp.
Figure 3.
Positive effect on the vegetative development of lettuce (Lactuca sativa L.) of the microalgae Chlorella spp., Arthrospira spp. and Scenedesmus spp.


Table 1.
Effects of microalgae on plant species.
| Species | Genera of microalgae | Effects | Ref. |
|---|---|---|---|
| Lettuce (Lactuca sativa L.) | Chlorella, Scenedesmus quadricauda, Spirulina platensis | Improved productivity, antioxidant capacity, carotenoid content, increased dry matter, chlorophyll and protein in seedlings | [68] |
| Mais (Zea mays L.) | Spirulina platensis | Increased production of caryopses and micro-nutrients | [69] |
| Aubergine (Solanum melongena L.) | Spirulina platensis | Increased vegetative growth and fruit production | [70] |
| Tomato (Solanum lycopersicum L.) | Acutodesmus dimorphus, Chlorella vulgaris, Scenedesmus quadricauda, Nannochloropsis oculata | Increased seed germination, crop biomass, root development and dry matter. Increased sugar and carotenoid content in the fruit | [58,71] |
| Pepper (Capsicum annuum L.) | Spirulina platensis, Dunaliella salina | Plant growth stimulation and salt stress mitigation in seed germination | [72] |
| Cucumber (Cucumis sativus L.) | Spirulina platensis | Improving fresh weight | [68] |
| Fava (Vicia faba L.) | Spirulina platensis | Improved protein and amino acid levels of roots and sprouts | [73] |
| Garlic (Allium sativum L.) | Arthrospira fusiformis | Increasing plant height | [74] |
| Onion (Allium cepa L.) | Spirulina platensis, Scenedesmus subspicatus | Increased production, photosynthetic pigments, root development and sugar and protein content | [75] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



