
Submitted:
28 June 2023
Posted:
30 June 2023
You are already at the latest version
Abstract
The genus Eucalyptus is a globally captivated source of hardwood and is well known for its medicinal uses. The hybrid and wild species of Eucalyptus are widely used as exotic plantations due to their renowned potential of adapting to various systems and sites, furthermore, making the species the most extensively propagated. Despite unquestionable progression in biotechnological and tissue culture approaches, the productivity of plantations is still limited often due to the low efficiency of clonal propagation from cuttings. The obtained F1 hybrids yield high biomass and High-quality low-cost raw material for large-scale production, however, the development of hybrid, clonal multiplication, proliferation, and post-developmental studies are still major concerns. This riveting review describes the problems concerned with in-vitro and clonal propagation of Eucalypt plantation; recent advances in biotechnological and tissue culture practices for massive and rapid micro-propagation of Eucalyptus; as well, highlights the Eucalypt germplasm preservation techniques.
Keywords:
Eucalyptus
; tissue culture
; F1 hybrids
; clonal propagation
; micro-propagation
; germplasm.
1. Introduction
Eucalyptus (Family: - Myrtaceae) is a genus including rapidly growing evergreen ornamental trees and shrubs indigenous to Australia. Over 900 different species native to Australia, Argentina, Brazil, Chile, France, India, Indonesia, Portugal, Morocco, Spain, South Africa, and USA have been planted, covering more than 20 million hectares of land [1,2]. The augmented urge of its pulp and hardwood as well as adaptivity to various biotic stressors and high productivity have hiked its economic value [3]. Besides these, the essential oils concentrated in different organs of the plant possess biological, toxicological, and pharmacological applications thus making it a promising prospect for ethnomedicine[4]. Owing to the increased future demands, rapidly propagating Eucalyptus plantations are desired [5,6].
The conventional breeding programs involve the cultivation of Eucalyptus from seeds; however, challenges of long generation time and genetic load persists [7]. These challenges have paved the way for modern biotechnological and plant tissue culture approaches allowing highly efficient rapid multiplication of plants in in-vitro environments as well as germplasm storage and preservation [8]. Biotechnological tools along with traditional tree improvement techniques can be considered as a solution for meeting growing human demands for forest produce as compared to conventional tree improvement programs alone, also biotechnology opens doors for understanding a wide range of complex biological problems related to the forest tree species. Forest biotechnology covers a wide array of modern techniques covering aspects of genetic engineering, in vitro regeneration techniques, genomics, metabolomics, proteomics, molecular markers, and marker-assisted breeding. These advanced biotechnological tools have opened new doors for understanding the genetic structure, identifying factors of biotic and abiotic interactions and stress tolerance, and linking some of the important genetic traits making possible accelerated selection and further breeding programs. One such efficient technique is micropropagation. The promising approach of micropropagation is achieved via organogenesis and somatic embryogenesis [9]. This paper attempts to discuss the conventional breeding approach as well as the methods of clonal propagation in upcoming sections. This study also focuses on the factors that influence the clonal propagation of Eucalyptus and further discusses the advances in germplasm preservation techniques.
Previous research concerning Traditional and Modern breeding programs for improving Eucalyptus plantations in the last 25 years is included. Even though successful attempts in micropropagation have been made, however some challenges like hyperhydricity, phenolic oxidation, explant contamination, root and ex vitro survival of the established clone still persists. Addressing these problems requires more efforts and refinement of tissue culture protocols, including the optimization of growth media, hormone formulations, and culture conditions. Additionally, strict aseptic techniques and effective sterilization methods are crucial to minimize endogenous contamination. This comprehensive review aims to cover all gaps for large-scale propagation of the eucalyptus plant henceforth helping the scientific community with further research in a useful direction. Overall, overcoming these challenges will contribute to the successful large-scale propagation of forest species through in vitro techniques, benefiting reforestation efforts and conservation initiatives.
1.1. A Crop with potential biomass production
As reported, nine dominating species of Eucalyptus have been cultivated for hardwood including E. camaldulensis, E. pellita, E. dunnii, E. grandis, E. tereticornis, E. globulus, E. sargila, E. uropylla, E. nitens [10]. The F1 hybrids achieved from the breeding of these species show seed vigor and yield high biomass. Moreover, the F1 hybrids from interspecific breeding show heterosis. Lucrative attempts to achieve commercial-scale plantations have been made for some species (E. deglupta, E. grandis, E. camaldulensis) agile to Tropical and Subtropical regions with high rainfall via macro-cutting and micro-cuttings [11,12]. For those species impotent of rapidly propagating by these methods, a tissue culture is a propitious approach [13]. The events of natural hybridization between Eucalyptus species have been reported [14]. The spontaneous hybrid formation between E. benthamii and E. dunnii at Embrapa forests in Colombo, Paraná State, presented frost-tolerant varieties. Likewise, the artificially pollinated hybrids serve as a viable alternative for future forest plantations while combining the useful traits from different parental combinations [15,16]. Initially thirteen mature F1 eucalyptus hybrid pairings were reported in India. Studies were started to create controlled and natural hybrids using half-sibling progeny generated from seeds gathered from stands of two intercross able species growing close to one another based on cross-ability pattern. The hybrids were created mechanically or naturally through selection in the Dehradun campus of the Forest Research Institute’s New Forest area. Researchers had worked on two important interspecific F1 hybrids, FRI-5 (E. camaldulensis Dehn × E. tereticornis Sm) & FRI-14 (E. torelliana F.V. Muell × E. citriodora Hook) among several interspecific hybrids that were created [17]. The superiority of Eucalyptus hybrids FRI-5 and FRI-14 to their parentage has been established. Additionally, high levels of hybrid vigor have been shown by these plants and they outperformed the parent combinations by three to five times in terms of growth characteristics. The average yield per unit area per unit of time was lower in the F2 population produced from seeds because of the significant segregation that was seen [17] (Table 1, Table 2).
According to [18,19], these hybrids produced more biomass than their parents' offspring and the Mysore Gum. They reported a micropropagation strategy for cloning and large-scale production as novel clonal propagation techniques for the commercial cultivation of these priceless Eucalyptus hybrids. The hybrid was observed to be intermediate to its parental species in more than half of the contrasting characteristics tested, it is noteworthy because of the two parent species involved i.e. E. grandis and E. tereticornis. A faster growth rate and good stem form were exhibited by the former and provides the best pulp quality, prefers high rainfall areas while the other was drought tolerant, showing suitability of hybrids towards intermediary environments (hybrid habitat). This reported for the first time in India. the outcomes of hybrid micro propagation. The species and hybrids that are reproduced via traditional methods (such as the many millions of E. grandis × E. urophylla and E. grandis plants produced per year) are often adapted to heavy rainfall areas in the tropics or subtropics. On drier, colder sites where land is more easily accessible and less expensive, hardwood plantations are being planted more frequently. Other eucalypt species are needed for these locations, such as the harder to propagate from cuttings C. citriodora, E. globulus, E. cloeziana F. Muell.), or E. nitens [20,21]. The development of effective techniques for eucalyptus clonal propagation, particularly for species that are difficult to propagate from cuttings, has remained a major issue in hardwood forestry. Tissue culture being among the most promising methods to rapidly propagate desirable genotypes and maintaining germ plasm in vitro has been used sustainably for these species and many others by various workers [22].
2. Micropropagation and its applications
Achieving true-to-type plants with desirable traits using in-vitro protocols is termed micropropagation (Figure 1). Five critical stages need to be accomplished for the successful establishment of micropropagation [23]. These Five stages include (i) Stage 0 described as the preparative stage for developing efficient and reproducible protocol; (ii) Stage 1 aiming to establish an Aseptic and viable culture; (iii) Stage 2 intending propagation without immolating objective to be achieved; (iv) stage 3 involving large scale propagation and (v) stage 4 hardening of the established plant material. Each of the stages involved is discussed comprehensively in the impending section.
2.1. Establishing Axenic Culture
For ensuring the success of micropropagation, maintaining uncontaminated cultures are necessary throughout the protocol. The foremost important step in the in-vitro propagation of plants is to establish a microbial contamination-free culture because the primary explants employed are nonsterile and thus the major source of microbial contamination in the culture [24]. The nodal segments bearing axillary buds, shoot tips, lead discs, and seeds could be used as explants to initiate tissue culture. The seeds as explants do not anticipate true-to-type propagation however the ease of decontamination and juvenility of young seedlings make them superior for rapid clonal propagation [25,26]. Moreover, Dormant Axillary buds from nodes are widely used for maintaining clonal fidelity providing thousands of plantlets by rapid propagation with high multiplication rates. The propagation from nodal segments is also reported in the micro-propagation of woody plants like bamboo [27]. Nonetheless, the explants from leaf discs and shoot tips provide true-to-type propagation, though, they are least preowned due to onerous sterilization [25,27]. The initial step involves surface sterilization of the explant which is usually achieved by first rinsing/washing the explant with non-sterilized water, immersing it in ethyl alcohol (70%), or treating it with chlorine-based sterilant such as Calcium Hypochlorite (Ca (OCl)2, Mercuric chloride (HgCl2), and Sodium Hypochlorite (NaOCl), followed by rinsing with sterile distilled water. To prevent drying and increase the interaction between the explant surface and sterilant, a drop of wetting agents such as Tween 20 is often added. Though Chlorine based sterilant efficiently can sterilize the explants, however their use is concerned with toxic effects on the living cells. Mercuric chloride has been reported to cause mammalian toxicity. So, the use of either Calcium hypochlorite or Sodium hypochlorite is recommended [28,29]. The basic explant sterilization Eucalypt is achieved by treating with Sodium hypochlorite at 67-1340 mM Concentrations. However, the optimal concentrations of Sodium hypochlorite must be optimized before experimentation as it can result in affecting the overall success of micropropagation. For instance, as reported, seed germination of C. citridora and C. torelliana hybrid was decreased when the concentration of NaOCl was increased from 134 to 670 mM [30]. Optimization of sterilization protocol of five different clones of eucalypt was performed using 1% sodium hypochlorite, 0.1% Mercuric chloride, 1mg/mL rifampicin and 70% ethanol as sterilant, study suggested that, along with 1% sodium hypochlorite and 70% ethanol. 0.1 mercuric chloride for 3 minutes was optimum and 1mg/mL rifampicin for 5 minutes was optimum for effective sterilization [31].
Leaching out of phenolics from the cultured explants is also a main concern that prevents establishment of in vitro cultures to a large extent. Reports suggest that Nodal segments cultured on MS medium exudated phenolics. The quantity of phenolics exudated by nodal segments and bud break response varied with the month of the collection of explants. Nodal segments have been reported to be collected every month starting from January to December. It was reported that explants collected during April and July to September showed the least phenolic exudation and better bud break response comparatively and were best for in vitro studies. From October to January and May to June the phenolic exudation was high with poor bud break responses [32].
2.2. In-vitro proliferation of shoot
In plant tissue culture, two main pathways are involved in achieving shoot regeneration: embryogenesis and organogenesis. Both processes play crucial roles in the development of new shoots, but they differ in the cellular mechanisms and morphological changes they undergo. Past literature has only focused on “shoot multiplication” however for a better understanding here we provide a concise explanation for organogenesis and embryogenesis.
2.2.1. Organogenesis
Organogenesis refers to the process by which shoot regeneration occurs through the differentiation and development of new shoots from pre-existing meristematic cells or tissues. In this pathway, the initial step involves the induction of adventitious shoots from explants such as leaf, stem, or root segments. The explants are cultured on a nutrient medium supplemented with specific plant growth regulators, particularly cytokinin, which stimulate the formation of shoot primordia. These primordia then undergo further growth and differentiation to develop into shoots with organized structures, including leaves and stems. Organogenesis is often characterized by the initiation of multiple shoots, referred to as shoot proliferation or multiplication [33]. The formation of new organs directly from explants is termed direct organogenesis while the formation of new organs from cell cultures or suspension, tissues, or callus is termed indirect organogenesis [33,34,35]. The development of new organs depends on a variety of factors such as hormones and type of media [36]. The differential media and nutrients employed in Eucalypt micropropagation are discussed in the upcoming section. Mostly basal MS salts and Murashige Skoog media of different strengths are used for establishing organogenesis in Eucalypt. Besides these, Woody Plant Media, Driver and Kuniyaki Walnut Media, Juan Antonio Diva and Salvin Media, Schenk, Hildebrandt Media, and B5 media have also been extensively used [17,33,37] [Table 1].
2.2.2. Somatic embryogenesis
The development of bipolar embryos from somatic cells/ tissues asexually is termed as somatic embryogenesis [38]. It is a pathway for shoot regeneration that mimics the process of embryo development in plants that involves the dedifferentiation of somatic cells to a totipotent state, where they regain the ability to form an embryonic structure. Somatic embryos can be formed via two pathways, direct embryogenesis in which the embryo develops from pre-embryonic cells, or via indirect embryogenesis in which the embryo develops from a callus raised on culture media [33,38]. A protocol for somatic embryogenesis has been reported by [39], suggesting the use of leaves from adult trees to be better for inducing somatic embryogenesis than floral tissues. The dedifferentiated cells progress through various stages of embryo development, including the formation of a proembryo, globular embryo, heart-shaped embryo, and a mature embryo. Shoots arise from these embryogenic structures, and they can be further multiplied through subsequent subculturing. However, the inability of somatic embryos to reach the maturation stage is an adverse limitation of clonal propagation, the success of which is directly dependent upon the optimization of PGRs used, the age of tissue used, and the type of media used for establishment [Table 1]. Typically semi-solid MS media supplemented with sucrose is used to initiate Eucalypt somatic embryogenesis, however, B5 media supplemented with sucrose have also been reported to induce somatic embryogenesis in C. citridora. Moreover, various PGRs have also been reported to induce embryogenesis in Eucalypt, for instance a hormone-free media for inducing somatic embryogenesis was also suggested [40,41,42,43]
2.3. Adventitious root formation and root hardening
The bipolar structure formed after somatic embryogenesis can germinate directly by using nutrients from basal media for the shoot and root proliferation. Contrastingly, the unipolar structure needs the proliferation of adventitious roots at the base of their shoots for the development of plantlets. This is usually achieved via Semisolid media, often activated charcoal is also introduced in the media as it regulates the pH of the media, and as well reported to adsorb inhibitory compounds from the media, moreover, reducing irradiance at the base of the shoots [44,45,46]. Improved in vitro rooting in E. grandis × E. nitens was reported that, by reducing the strength of MS media from full to half strength and decreasing the concentration of sucrose in shooting media from 20 to 15 gL-1. Additionally, it was observed that increasing the concentration of NAA from 0.1 mgL-1 to 0.5mgL-1 increased the average percentage of adventitious roots [29]. Also, increasing the IBA concentration from 0.1 mgL-1 to 0.5 mgL-1 increased root hair formation. Similar studies on E. erythronema × E. stricklandii suggested 8 weeks of continuous exposure to IBA on roots resulted in the longest root length [47]. Light studies on E. grandis × E. urophylla suggested the use of Red/ blue light to be superior for rooting and showed the highest mean number of roots [48].
Before adventitious root formation, the acclimatization of shoots was necessary for their future success in nursery conditions. Micro propagated plantlets were hardened using liquid MS media (1/4 strength) possessing 2% sucrose in it. Further for supporting roots, adsorbent cotton was soaked in this liquid media. After maintaining for 2 weeks, the plantlets were transferred to mist bags containing a 1:1:1 ratio of soil, manure, and sand and then transferred to a net house. Finally, the plantlets were transferred to field conditions and showed 85-95% success rates in field conditions [17]. However in another study, 58% of success rate of plantlets in field conditions due to the loss of some planets during handling [49]. Another innovative approach for acclimatization was reported where the shoots were maintained in photoautotrophic culture at high carbon dioxide and low sugar concentrations. These conditions promote carbon fixation and transpiration. 86 – 96% of success rate for E.camaldulensis and 100% success rate for E. grandis × E. urophylla have been achieved via this method [50,51,52]. Also, a similar study suggested that increasing the temperature from 18-13°C to 33-28°C increased number of root cuttings per stock plant [53]. Moreover, improvement of rooting efficiency in clones of E. urophylla was achieved by in vitro rejuvenation/ reinvigoration [54].
3. Factors affecting the efficiency of micropropagation
The efficiency of micropropagation, can be influenced by various factors (Figure 2). Growth regulators play a crucial role in promoting shoot proliferation and rooting in tissue culture. The type and concentration of cytokinins and auxins in the culture medium can significantly impact the rate and quality of shoot formation. Additionally, the composition of the culture media, including the types and concentrations of nutrients, vitamins, and carbon sources, affects the growth and development of explants. Organic elements such as amino acids and complex organic compounds can enhance shoot regeneration and plantlet growth, while inorganic elements, including macronutrients and micronutrients, are vital for overall plant health and development. Moreover, light is an essential environmental factor that affects micropropagation efficiency. Proper light intensity and photoperiod can influence shoot initiation, elongation, and rooting. The spectrum of light, particularly the ratio of red to far-red light, also plays a role in regulating plant growth and differentiation. Optimizing these factors in micropropagation protocols is crucial for maximizing the efficiency of shoot proliferation and the production of healthy plantlets. Here we elaborate these all factors and their effect on Eucalypt micropropagation.
3.1. Role of Plant Growth Regulators:
Indole-3-Acetic acid (IAA) is the natural auxin in plants and Indole-3-butyric Acid (IBA) is the analog of auxin found in plants, both IAA and IBA are syntheised via Tryptophan dependent or independent pathways [55,56]. Both IAA and IBA can be quickly metabolised in Tissues of Eucalyptus. Since auxins plays regulatory role in regulating cell divison , elongation of plant and many other phases of their development, so these are stored by plants as IAA or in the form of IBA which is converted into IAA whenever required [57,58].
Plant growth regulators have been reported to directly as well indirectly influence the proliferation of plantlets including the differentiation of embryos and different organs. For instance, optimal concentrations of cytokinin/ auxin necessary for inducing organogenesis in E. cloeziana micropropagation was suggested by [59]. Moreover, the highest rate of somatic embryogenesis by introducing 0.1 mgL-1 NAA and 0.5mgL-1 BA was also reproted [60]. Increased shoot multiplication by the addition of 0.5mgL-1 BAP in WPM and ½ MS media was also reported [61]. It is important to consider that optimization of PGRs in culture media is necessary as it could negatively affect the growth of plantlets [61,62]. Similar studies on Auxin type on E. salgina, and E. globulus were reported, suggesting the best rooting obtained with IBA than IAA. Best rooting was achieved in both species when treated with IBA, the possible explanation supporting this was IAA is highly susceptible to enzymatic degradation as well is 5 times more susceptible to photo-oxidation than IBA. In support of this, a similar study on E. sideroxylon micorcuttings was reported on Comparing IBA and NAA using different concentrations of both auxins from 0-10μM. High frequency of callus indusction was reproted by culturing cotyledon explants on MS media supplemented 1mgL-1 NAA + 0.5mgL-1 BA, also the same study suggested MS media supplemented with 0.5mgL-1 NAA + 1mgL-1 BA + 1mgL-1 GA3 , as well ½ MS media supplemeted with 0.5 – 1mgL-1 resulted in high frequency adventitious root formation in E. bosistoana. Furthermore, 100% survival of preacclimatized plantlets was obtained [63]. Also, similar study revealed best shoot elongation by supplementing the media with 0.05mgL-1 BAP + 1mgL-1 NAA and 0.05mgL-1 BAP + 1mgL-1 NAA + 1mgL-1 IBA-1 [64]. The results showed that increased callus induction in micorcuttings exposed to IBA was observed than the micorcuttings exposed to NAA . However, the responses to auxins may vary from species to species based on their differential affinities, uptake, and metabolization of auxins [65,66].
3.2. Effect of culture media
The initiation and shoot proliferation is usually achieved by culturing explant on semi-media that is made up of Gelling agents such as 1.5-4.0g/ L gelrite, 4-8g/L agar, or 1.5 -4.5 g/L phytagel, and the pH is adjusted between 5-6. Also, use of liquid media has been reported for the initiation and proliferation of nodes and shoots [67,68]. The culture of the shoot depends on the ability to encourage the development of axillary and accessory buds that are present at the base of each leaf axil. In previous research attempts at Eukalypt micropropagation, the basal media used includes Murashige and Skoog media ( Different strengths ½, ¼ ); JADS medium (Juan, Antonio, Diva, and Silvian), WPM (Woody Plant medium) and DKW media (Driver and Kuniyaki Walnut) [69]. Additionally, the form of medium, whether liquid suspension medium or semisolid medium, affects the growth of plantlets in vitro. Reportedly, better shoot multiplication of Eucalypts has been observed in liquid media as compared to semi-solid media [70]. WPM as the optimal media for micropropagation of E. benthamii was reported by [71]. Similarly, JADS media was observed optimal for trunk base shoot elongation of E. grandis. DKW media was used as an alternative for micropropagation of E. nitens [72]. Adjustment of pH of the medium to 5.8 prior to autoclaving at 121° C for 15 min has been recommended. Temperature incubation at 25 ± 2°C and 16 h photoperiod with the photon flux density of 2500 lux, from white fluorescent tubes is the recommended cultured conditions for eucalyptus spp. For improving the survival of explant, polyvinyl pyrrolidone, activated charcoal, and ascorbate have also been added to the culture media. However, the type of media and the response of plantlets varies among species of Eucalypts and their hybrids, besides these some drawbacks such as chlorosis, tissue browning, and oxidation have been reported in almost all types of media used for Eucalypt micropropagation.
3.3. Importance of organic and inorganic elements
Organic and inorganic elements play a crucial role in plant micropropagation media, which are used for the propagation and growth of plants under sterile conditions. Elements like Calcium (Ca), Nitrogen (N), Phosphorous (P) and Boron (B) serve as macro and micronutrients essential for nourishing plant growth. These have been introduced in in vitro cultures to promote the proliferation, and differentiation of organs from the shoot. An appropriate balance and concentration of these organic and inorganic elements are crucial to ensure successful propagation and growth of plants in vitro [73]. The media composition can be adjusted based on the specific requirements of different plant species and their growth stages. The elements required by plants lower than 0.5mM/L concentration as referred to as micronutrients and more than this concentration are referred to as macronutrients [74,75,76]. Magnesium (Mg), Calcium (Ca), Hydrogen (H), Sulphur (S), Potassium (K), Nitrogen (N), Phosphorous (P), and Oxygen (O) serve as macronutrients. Calcium act as an important cofactor and cellular messenger involved in various signal transduction pathways as well known to play various important roles in plant stress [77]. For instance, in E. urophylla and E. grandis calcium has been reported to trigger organogenesis [78]. Manganese (Mn), Chlorine (Cl), Iron (Fe), Zinc (Zn), Boron (B), Sodium (Na), Iodine (I), and Copper (Cu) serve as the microelements from which Iron is the most critical element. Also, it was reported that the deficiency of Boron in media led to necrosis and callus accumulation, additionally seedling inhibition in E. grandis [79]. Furthmeore a study suggests that calcium, in the form of calcium chloride in agitated liquid media decreases the hyperhydricity in E. saligna , However due to toxicity caused by chlorine, it had been not effective in completely eliminating hyperhydricity [80]. Also, calcium chloride dihydrate was reproted to induce shoot elongation as well decrease vitrification [81]. Nonethless efforts in improving the optimization and choice of organic / inorganic elements have been increased, thpugh is insuuficient to cover the individual role of vast available macro and micronutreints.
3.4. Role of carbohydrates
Carbohydrates are important biomolecules that provide biofuel and a carbon source to the growing cells. Different reducing and non-reducing sugars are available and have been employed in micropropagation like Glucose, fructose, Sucrose, and Galactose, of which sucrose still being the most preferable in Eucalypt micropropagation due to its ease of translocation in plant tissues. Sucrose is a non-reducing sugar, specifically a disaccharide comoposed of fructose and glucose. Some of the reports suggested that increased sucrose concentrations can hinder the water and nutrient uptake in plants as well as hinders photosynthesis by influencing photosynthetic enzymes. However, contrastingly some studies suggest that plants remain uninfluenced by high sucrose concentrations. Furthermore, in E. cloiziana, high glucose, and sucrose concentrations were reported to decrease the shoot length (conc.> 15g/mL in media) [82]. For instance, studies reported different concentrations of sucrose (1%-6%) in MS media for in vitro shoot proliferation. Best results were reported using 3% sucrose in MS media with a 6-7 fold increased shoot multiplication. Similar findings reported that media devoid of sucrose results in inhibition of shoot multiplication as well the leaves and shoots turned to a pale green color [83,84]. The results of this study are in line with those of several studies that used 3 percent sucrose as a carbohydrate source to promote. The growth of shoots in a variety of Eucalyptus species. However, similar findings on many other woody plants are also being reported for instance, in bamboo shoots successful multiplication was observed when media was supplemented with 2% sucrose [85,86]. Also, it has been reported that increases in sucrose levels at 3 to 4% do not cause any effect on shoots but result in albinism. Similarly, at 1% sucrose concentration, thin shoots and leaves were developed that were inappropriate for subculturing. Investigation on Myo-inositol has been conducted to determine its role in in vitro shoot multiplication. MS media supplemented with 100mgL-1 of myo-inositol yielded the best shoot multiplication rates, while MS media devoid of Myo-inositol showed decreased shoot multiplication. Moreover, MS media supplemented with excess myo-inositol (more than 150 mgL-1) not only decreased shoot multiplication but alongwith resulted indetrimentak effect on shoots. For maximizing shoot multiplication rate and growth, 100mgL-1 myo-imositol was supplemnted in culture media for all trials [6,83].
3.5. Effects of radiation and light exposure
Light is a critical external aspects influencing the different phases of plant growth. Light hour durations and intensity is directly linked to the photosynthetic rates in plants [87]. Many hybrids have been studied that suggest the effects of light and radiation on the success of micropropagation. The effects of five sources of lights, fluorescent lamps, white LEDs, red LEDs, blue LEDs, and red/blue LEDs on E. grandis × E. urophylla hybrid was studied and reported the Use of Red/ Blue LEDs and Florescent lights to be superior for E. grandis × E. urophylla micropropagation. Response of Eucalypt to micropropagation varies among genotypes. Moreover, a low level of irradiations triggered rooting in E. globulus, contrastingly, some studies proved low-level irradiations hindered root proliferation in E. globulus [48,88,89]. Also, studies on E. salgina and E. globulus were conducted for the effect of light on rooting capacity using a White fluorescent lamp. E. globulus did show any effect on the exposure and was observed to be dependent only on exogenous auxin concentration for rooting, while E. salgina cuttings were observed to show increased root density per rooted cutting upon exposure to light in conjugation with exogenous auxin application [90]. Also, a similar study suggested that preservation under low light intensity effectively presrved cultures for 3 months [91]. Besides these, increasing light intensity alongwith increased Carbondioxide content have shown to increase the growth of explants photoautotrophically [92]. In a similar study, the effect of light quality on the clone of E. urophylla in photoautotrophic system was assessed focusing on the stomatal density, carotenoid content, chlorophyll content, number of shoots and longest shoot. The results indicated that blue LED resulted in fewer shoots, while, high production of carotenoids was observed under white light [93]. In another study on Eucalyptus dunnii and Eucalyptus grandis × E. urophylla, it was observed that white light was realted to increased buds per plants, decreased tissue oxidation and longer shoot length. In E. dunnii , blue, red and yellow light resulted in increased chlorophyll a & b content. Also, blue, white, puple and red light increased stomatal densities. Moreover, the study suggests that irrespective of light spectra, E. dunnii showed decreased adventitious rooting [94]. An another similar study on E. grandis × E. urophylla clone was carried out to assess the impact of five different light sources including fluorescent lamp, blue, green, red and yellow cellophane light in bioreactor system. This study suggests yellow and blue light to be more suitable for the clone as less hyperhydricity was observed alongwith spongy parenchymatic tissue, thicker mesophyll, increased shoot length and more shoots per explant [95]. Besides, an other study suggests that for in vitro multiplication of E. pilularis , white light was more suitable, and for E. urograndis clone, blue light was more suitable because it increased number of buds, shoots length and fresh weight of per explant [96].
4. In-vitro germplasm preservation
In vitro germplasm preservation plays a vital role in conserving plant biodiversity, protecting endangered species, and safeguarding important genetic resources for future research, breeding programs, and restoration efforts. It helps to maintain a diverse and resilient gene pool, ensuring the availability of plant materials for sustainable agriculture, forestry, and environmental conservation. A brief insights to recent preservation techniques in Eucalypt spp. Have been discussed below.
4.1. Cryopreservation and cold storage
Preservation of germplasm is peremptory for breeding programs. Cryopreservation is a protocol holding germplasm storage at ultra lower temperatures (-135°C to -196°C) in liquid nitrogen. At this temperature the cell viability and genetic stability are preserved however, the aging of the cell is hindered due to the halting of all biochemical and physiological pathways of the cell [97]. Various reporting on successful cryopreservation of Eucalypt spp. Have been reported including shoot tips of E. grandis × E. camaldulensis, E. urophylla × E. grandis, E. grandis, E. grandis × E. urophylla, and E. camaldulensis by droplet vitrification method, firstly by preculturing shoot tips on MS medium for 24 hours containing 0.25M sucrose concentration and then again at 24 hours on MS media containing 0.625M sucrose concentration followed by subjecting the individual shoot tips to Plant Vitrification Solution (PVS) and Preservation of Axillary buds of E. grandis × E. camaldulensis have also been reported by placing the axillary buds in semi-solid MS media on significantly increasing the concentration of sucrose and glycerol at 0.4, 0.7 and 1.0 M, this type of method showed 49% of regrowth [97,98,99].
Cold storage involves the preservation of plants at lower temperatures. Some attempts have been made to preserve shoots of E.grandis in cold storage on Half strength and full-strength media. However, half-strength MS media allowed the preservation of shoots at 24°C to 28°C for 10 months, contrastingly full strength media could preserve the same shoots at 10°C only for 6 months. Similar studies were conducted on C. toleriana and C. citridora showed preservation of shoots on Full strength MS media at 14°C for 12 months [100].
5. Conclusions
Micropropagation has allowed for achieving rapid clonal plantations, however, the efficiency and success of micropropagation depends upon the rate of shoot multiplication. Eucalypt spp and Its hybrids are among the commercially important tree spps and have been extensively propagated by micropropagation. These are continuously studied for their genetic improvement and establishment of superior hybrid species. The micropropagation of Eucalypts can be achieved in 4 main steps (i) suitable explant collection, (ii) establishment of aseptic culture, (iii) shoot and root proliferation, (iv) hardening of the root. A wide range of media including, semi solid and liquid cultire media have been described for efficient clonal propagation of Eucalypt. Currently, many studies are still being conducted for the development of improved tissue culture protocols as well as improving the wood quality of Eucalypts. For achieving success in callogenesis and histogenesis of Eucalypt spp, complete knowledge of all optimizations including the type of media, choice of the hormone, their concentration, and ratios must be known. Apart from that, the age and ttype of tissues, as well as the seasons during the collection of explants, are necessary to consider for ensuring the success of tissue proliferation. Moreover, field success of plantlets is necessary for which proper conditions and protocols for acclimatization of plantlets must be known All of these important factors have been compiled in the present study. However, more studies are required to close the gap between in vitro raising of plantlets of Eucalypt to their growth in field conditions which is important for its industrial success.
Author Contributions
“Conceptualization, V.S. and I.S.; methodology, AnK. and V.S.J; writing—original draft preparation, V.S. and AnK, B.K.; writing—review and editing, S.S., A.K., V.S.J.; visualization, V.S.J. and S.G.; supervision, V.S. and I.S..; project administration, I.S.; funding acquisition, I.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data are contained within this article.
Acknowledgments
NA.
Conflicts of Interest
Declare conflicts of interest or state “The authors declare no conflict of interest.”.
References
- Watt M., P. and Blakeway, F. C. and M. M. E. O. and J. S. M. Micropropagation of Eucalyptus. In Micropropagation of Woody Trees and Fruits; Jain S. Mohan and Ishii, K., Ed.; Springer Netherlands: Dordrecht, 2003; pp. 217–244. [Google Scholar] [CrossRef]
- Trueman, S. J.; Hung, C. D.; Wendling, I. Tissue Culture of Corymbia and Eucalyptus. Forests 2018, 9. [Google Scholar] [CrossRef]
- Nogueira, M. C. de J. A.; DE ARAUJO, V. A.; Vasconcelos, J. S.; Christoforo, A. L.; Lahr, F. A. R. Sixteen Properties of Eucalyptus Tereticornis Wood for Structural Uses. Bioscience Journal 2020, 36, 449–457. [Google Scholar] [CrossRef]
- Silva, J.; Abebe, W.; Sousa, S. M.; Duarte, V. G.; Machado, M. I. L.; Matos, F. J. A. Analgesic and Anti-Inflammatory Effects of Essential Oils of Eucalyptus. J Ethnopharmacol 2003, 89, 277–283. [Google Scholar] [CrossRef]
- Kamal, B.; Arya, I.; Sharma, V.; Jadon, V. S. In Vitro Enhanced Multiplication and Molecular Validation of Eucalyptus F1 Hybrids. Plant Cell Biotechnol Mol Biol 2016, 17, 167–175. [Google Scholar]
- Arya, I. D.; Sharma, S.; Chauhan, S.; Arya, S. Micropropagation of Superior Eucalyptus Hybrids FRI-5 (Eucalyptus Camaldulensis Dehn x E. Tereticornis Sm) and FRI-14(Eucalyptus Torelliana F.V. Muell x E. Citriodora Hook): A Commercial Multiplication and Field Evaluation. Afr J Biotechnol 2009, 8, 5718–5726. [Google Scholar] [CrossRef]
- Kendurkar Shuchishweta Vinay and Rangaswamy, M. Genetic Transformation in Eucalyptus. In Biotechnologies of Crop Improvement, Volume 2: Transgenic Approaches; Gosal Satbir Singh and Wani, S. H., Ed.; Springer International Publishing: Cham, 2018; pp. 335–336. [Google Scholar] [CrossRef]
- El-Esawi, M. A. Micropropagation Technology and Its Applications for Crop Improvement. In Plant Tissue Culture: Propagation, Conservation and Crop Improvement; Anis Mohammad and Ahmad, N., Ed.; Springer Singapore: Singapore, 2016; pp. 523–545. [Google Scholar] [CrossRef]
- Trueman, S. J. Clonal Propagation and Storage of Subtropical Pines in Queensland, Australia. Southern African Forestry Journal 2006, 208, 49–52. [Google Scholar] [CrossRef]
- Booth, T. H. Eucalypt Plantations and Climate Change. For Ecol Manage 2013, 301, 28–34. [Google Scholar] [CrossRef]
- Booth, T. H. Eucalypt Plantations and Climate Change. For Ecol Manage 2013, 301, 28–34. [Google Scholar] [CrossRef]
- Shanthi, K.; Bachpai, V. K. W.; Anisha, S.; Ganesan, M.; Anithaa, R. G.; Subashini, V.; Chakravarthi, M.; Sivakumar, V.; Yasodha, R. Micropropagation of Eucalyptuscamaldulensis for the Production of Rejuvenated Stock Plants for Microcuttings Propagation and Genetic Fidelity Assessment. New For (Dordr) 2015, 46, 357–371. [Google Scholar] [CrossRef]
- Kataria, V. and Masih, A. and C. S. and S. S. K. and K. A. and A. I. D. Clonal Propagation, a Tested Technique for Increasing Productivity: A Review of Bamboos, Eucalyptus and Chirpine. In Agricultural Biotechnology: Latest Research and Trends; Kumar Srivastava Dinesh and Kumar Thakur, A. and K. P., Ed.; Springer Nature Singapore: Singapore, 2021; pp. 37–51. [Google Scholar] [CrossRef]
- Butcher, P. A.; Skinner, A. K.; Gardiner, C. A. Increased Inbreeding and Inter-Species Gene Flow in Remnant Populations of the Rare Eucalyptus Benthamii. Conservation Genetics 2005, 6, 213–226. [Google Scholar] [CrossRef]
- Potts, B. M.; Dungey, H. S. Interspecific Hybridization of Eucalyptus: Key Issues for Breeders and Geneticists. New For (Dordr) 2004, 27, 115–138. [Google Scholar] [CrossRef]
- Jovanovic, T.; Arnold, R.; Booth, T. Determining the Climatic Suitability of Eucalyptus Dunnii for Plantations in Australia, China and Central and South America. New For (Dordr) 2000, 19, 215–226. [Google Scholar] [CrossRef]
- Singh, D.; Kaur, S.; Kumar, A. In Vitro Drought Tolerance in Selected Elite Clones of Eucalyptus Tereticornis Sm. Acta Physiol Plant 2020, 42, 17. [Google Scholar] [CrossRef]
- Venkatesh, C. S.; Sharma, V. K. Hybrid Vigour in Controlled Interspecific Crosses of Eucalyptus Tereticornis X E. Camaldulensis. Silvae Genet 1977, 26, 121–124. [Google Scholar]
- Paramathma, M.; Surendran, C.; Rai, R. S. V. STUDIES ON HETEROSIS IN SIX EUCALYPTUS SPECIES. Journal of Tropical Forest Science 1997, 9, 283–293. [Google Scholar]
- de Almeida, M. R.; de Bastiani, D.; Gaeta, M. L.; de Araújo Mariath, J. E.; de Costa, F.; Retallick, J.; Nolan, L.; Tai, H. H.; Strömvik, M. V; Fett-Neto, A. G. Comparative Transcriptional Analysis Provides New Insights into the Molecular Basis of Adventitious Rooting Recalcitrance in Eucalyptus. Plant Science 2015, 239, 155–165. [Google Scholar] [CrossRef]
- Dickinson, G. R.; Wallace, H. M.; Lee, D. J. Reciprocal and Advanced Generation Hybrids between Corymbia Citriodora and C. Torelliana: Forestry Breeding and the Risk of Gene Flow. Ann For Sci 2013, 70, 1–10. [Google Scholar] [CrossRef]
- Wendling, I.; Warburton, P. M.; Trueman, S. J. Maturation in Corymbia Torelliana × C. Citriodora Stock Plants: Effects of Pruning Height on Shoot Production, Adventitious Rooting Capacity, Stem Anatomy, and Auxin and Abscisic Acid Concentrations. Forests 2015, 6, 3763–3778. [Google Scholar] [CrossRef]
- Debergh P., C. and Read, P. E. Micropropagation. In Micropropagation: Technology and Application; Debergh P. C. and Zimmerman, R. H., Ed.; Springer Netherlands: Dordrecht, 1991. [Google Scholar] [CrossRef]
- Cassells, A. C. Pathogen and Biological Contamination Management in Plant Tissue Culture: Phytopathogens, Vitro Pathogens, and Vitro Pests. In Plant Cell Culture Protocols; Loyola-Vargas Víctor M. and Ochoa-Alejo, N., Ed.; Humana Press: Totowa, NJ, 2012; pp. 57–80. [Google Scholar] [CrossRef]
- Wendling, I.; Brooks, P. R.; Trueman, S. J. Topophysis in Corymbia Torelliana × C. Citriodora Seedlings: Adventitious Rooting Capacity, Stem Anatomy, and Auxin and Abscisic Acid Concentrations. New For (Dordr) 2015, 46, 107–120. [Google Scholar] [CrossRef]
- Bag, N.; Chandra, S.; Palni, L. M. S.; Nandi, S. K. Micropropagation of Dev-Ringal [Thamnocalamus Spathiflorus (Trin.) Munro] — a Temperate Bamboo, and Comparison between in Vitro Propagated Plants and Seedlings. Plant Science 2000, 156, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Giri, C. C.; Shyamkumar, B.; Anjaneyulu, C. Progress in Tissue Culture, Genetic Transformation and Applications of Biotechnology to Trees: An Overview. Trees 2004, 18, 115–135. [Google Scholar] [CrossRef]
- Ansar, S.; Iqbal, M. Effect of Dietary Antioxidant on Mercuric Chloride Induced Lung Toxicity and Oxidative Stress. Toxin Rev 2015, 34, 168–172. [Google Scholar] [CrossRef]
- Keret, R.; Nakhooda, M.; Jones, N. B.; Hills, P. N. Optimisation of Micropropagation Protocols for Temperate Eucalypt Hybrids in South Africa, with a Focus on Auxin Transport Proteins. Southern Forests: a Journal of Forest Science 2021, 83, 254–263. [Google Scholar] [CrossRef]
- Trueman, S. J.; Richardson, D. M. In Vitro Propagation of Corymbia Torelliana C. Citriodora (Myrtaceae) via Cytokinin-Free Node Culture. Aust J Bot 2007, 55, 471–481. [Google Scholar] [CrossRef]
- Kuppusamy, S.; Ramanathan, S.; Sengodagounder, S.; Senniappan, C.; Shanmuganathan, R.; Brindhadevi, K.; Kaliannan, T. Optimizing the Sterilization Methods for Initiation of the Five Different Clones of the Eucalyptus Hybrid Species. Biocatal Agric Biotechnol 2019, 22, 101361. [Google Scholar] [CrossRef]
- Kamal, B.; Arya, I. D.; Gupta, S. In-Vitro Regeneration of Interspecific F1 Hybrid (Eucalyptus Citriodora and Eucalyptus Torelliana) of Eucalyptus. Journal of Mountain Research 2022, 17. [Google Scholar] [CrossRef]
- Aggarwal, D.; Kumar, A.; Reddy, M. S. Shoot Organogenesis in Elite Clones of Eucalyptus Tereticornis. Plant Cell, Tissue and Organ Culture (PCTOC) 2010, 102, 45–52. [Google Scholar] [CrossRef]
- Girijashankar, V. In Vitro Regeneration of Eucalyptus Camaldulensis. Physiology and Molecular Biology of Plants 2012, 18, 79–87. [Google Scholar] [CrossRef]
- Fernando, S. C.; Goodger, J. Q. D.; Gutierrez, S. S.; Johnson, A. A. T.; Woodrow, I. E. Plant Regeneration through Indirect Organogenesis and Genetic Transformation of Eucalyptus Polybractea R.T. Baker. Ind Crops Prod 2016, 86, 73–78. [Google Scholar] [CrossRef]
- Sluis, A.; Hake, S. Organogenesis in Plants: Initiation and Elaboration of Leaves. Trends in Genetics 2015, 31, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Oberschelp, G. P. J.; Gonçalves, A. N. Assessing the Effects of Basal Media on the in Vitro Propagation and Nutritional Status of Eucalyptus Dunnii Maiden. In Vitro Cellular & Developmental Biology - Plant 2016, 52, 28–37. [Google Scholar] [CrossRef]
- Méndez-Hernández, H. A.; Ledezma-Rodríguez, M.; Avilez-Montalvo, R. N.; Juárez-Gómez, Y. L.; Skeete, A.; Avilez-Montalvo, J.; De-La-Peña, C.; Loyola-Vargas, V. M. Signaling Overview of Plant Somatic Embryogenesis. Frontiers in Plant Science. Frontiers Media S.A. February 1, 2019. 1 February. [CrossRef]
- Martínez, M. T.; San-José, M. del C.; Arrillaga, I.; Cano, V.; Morcillo, M.; Cernadas, M. J.; Corredoira, E. Holm Oak Somatic Embryogenesis: Current Status and Future Perspectives. Front Plant Sci 2019, 10. [Google Scholar] [CrossRef]
- Pinto, G.; Silva, S.; Park, Y.-S.; Neves, L.; Araújo, C.; Santos, C. Factors Influencing Somatic Embryogenesis Induction in Eucalyptus Globulus Labill.: Basal Medium and Anti-Browning Agents. Plant Cell Tissue Organ Cult 2008, 95, 79–88. [Google Scholar] [CrossRef]
- Pinto, G.; Park, Y.-S.; Neves, L.; Araújo, C.; Santos, C. Genetic Control of Somatic Embryogenesis Induction in Eucalyptus Globulus Labill. Plant Cell Rep 2008, 27, 1093–1101. [Google Scholar] [CrossRef] [PubMed]
- Pinto, G.; Park, Y.-S.; Silva, S.; Neves, L.; Araújo, C.; Santos, C. Factors Affecting Maintenance, Proliferation, and Germination of Secondary Somatic Embryos of Eucalyptus Globulus Labill. Plant Cell Tissue Organ Cult 2008, 95, 69–78. [Google Scholar] [CrossRef]
- Pinto, G.; Silva, S.; Neves, L.; Araújo, C.; Santos, C. Histocytological Changes and Reserve Accumulation during Somatic Embryogenesis in Eucalyptus Globulus. Trees 2010, 24, 763–769. [Google Scholar] [CrossRef]
- Thomas, T. D. The Role of Activated Charcoal in Plant Tissue Culture. Biotechnol Adv 2008, 26, 618–631. [Google Scholar] [CrossRef]
- Jones, N. B.; van Staden, J. Micropropagation and Establishment of Eucalyptus Grandis Hybrids. South African Journal of Botany 1994, 60, 122–126. [Google Scholar] [CrossRef]
- Sapaeing, A.; Sutthinon, P.; Hilae, A.; Wattanapan, N. Effects of BA, NAA, and Activated Charcoal on Micropropagation of Nepenthes Mirabilis (Lour.) Druce; 2020. [CrossRef]
- Glocke, P.; Delaporte, K.; Collins, G.; Sedgley, M. Micropropagation of Juvenile Tissue of Eucalyptus Erythronema × Eucalyptus Stricklandii Cv. ‘Urrbrae Gem.’ In Vitro Cellular & Developmental Biology - Plant 2006, 42, 139–143. [Google Scholar] [CrossRef]
- Costa Souza, D. M. S.; Fernandes, S. B.; Avelar, M. L. M.; do Prado Frade, S. R.; Molinari, L. V.; Gonçalves, D. S.; Pereira Pinto, J. E. B.; Brondani, G. E. Light Quality in Micropropagation of Eucalyptus Grandis × Eucalyptus Urophylla. Scientia Forestalis/Forest Sciences 2020, 48. [Google Scholar] [CrossRef]
- Keret, R.; Nakhooda, M.; Jones, N.; Hills, P. Optimisation of Micropropagation Protocols for Temperate Eucalypt Hybrids in South Africa, with a Focus on Auxin Transport Proteins. Southern Forests: a Journal of Forest Science 2022, 83, 1–10. [Google Scholar] [CrossRef]
- Agrisexport.
- Zobayed, S. M. A.; Afreen-Zobayed, F.; Kubota, C.; Kozai, T. Mass Propagation of Eucalyptus Camaldulensis in a Scaled-up Vessel Under In Vitro Photoautotrophic Condition. Ann Bot 2000, 85, 587–592. [Google Scholar] [CrossRef]
- Tanaka, M.; Giang, D. T. T.; Murakami, A. Application of a Novel Disposable Film Culture System to Photoautotrophic Micropropagation of Eucalyptus Uro-Grandis (Urophylia x Grandis). In Vitro Cellular & Developmental Biology - Plant 2005, 41, 173–180. [Google Scholar] [CrossRef]
- Trueman, S. J.; McMahon, T. V; Bristow, M. Production of Eucalyptus Cloeziana Cuttings in Response to Stock Plant Temperature. Journal of Tropical Forest Science 2013, 25, 60–69. [Google Scholar]
- Mendonça, E. G.; Batista, T. R.; Stein, V. C.; Balieiro, F. P.; de Abreu, J. R.; Pires, M. F.; de Souza, P. A.; Paiva, L. V. In Vitro Serial Subculture to Improve Rooting of Eucalyptus Urophylla. New For (Dordr) 2020, 51, 801–816. [Google Scholar] [CrossRef]
- Leva, A.; Rinaldi, L. M. R. Recent Advances in Plant in Vitro Culture; IntechOpen: Rijeka, 2012. [Google Scholar] [CrossRef]
- Leva, A.; Rinaldi, L. M. R. Recent Advances in Plant in Vitro Culture; IntechOpen: Rijeka, 2012. [Google Scholar] [CrossRef]
- Frick, E. M.; Strader, L. C. Roles for IBA-Derived Auxin in Plant Development. J Exp Bot 2018, 69, 169–177. [Google Scholar] [CrossRef]
- Strader, L. C.; Bartel, B. Transport and Metabolism of the Endogenous Auxin Precursor Indole-3-Butyric Acid. Molecular Plant. Oxford University Press 2011, pp 477–486. [CrossRef]
- de Oliveira, L. S.; Brondani, G. E.; Molinari, L. V.; Dias, R. Z.; Teixeira, G. L.; Gonçalves, A. N.; de Almeida, M. Optimal Cytokinin/Auxin Balance for Indirect Shoot Organogenesis of Eucalyptus Cloeziana and Production of Ex Vitro Rooted Micro-Cuttings. J For Res (Harbin) 2022, 33, 1573–1584. [Google Scholar] [CrossRef]
- Prakash, M. G.; Gurumurthi, K. Effects of Type of Explant and Age, Plant Growth Regulators and Medium Strength on Somatic Embryogenesis and Plant Regeneration in Eucalyptus Camaldulensis. Plant Cell, Tissue and Organ Culture (PCTOC) 2010, 100, 13–20. [Google Scholar] [CrossRef]
- Brondani, G. E.; Dutra, L. F.; Wendling, I.; Grossi, F.; Hansel, F. A.; Araujo, M. A. <B>Micropropagation of an <Em>Eucalyptus</Em> Hybrid (<Em>Eucalyptus Benthamii x Eucalyptus Dunnii</Em>)</B> - Doi: 10.4025/Actasciagron.V33i4.8317. Acta Sci Agron 2011, 33 (4). [CrossRef]
- Nazirah, A.; Hasnida, N.; Mohd-Saifuldullah, A. W.; Y, M.-F.; Y, A.-Z.; K, R. DEVELOPMENT OF AN EFFICIENT MICROPROPAGATION PROTOCOL FOR EUCALYPTUS HYBRID (E. UROPHYLLA × E. GRANDIS) THROUGH AXILLARY SHOOT PROLIFERATION. JOURNAL OF TROPICAL FOREST SCIENCE 2021, 33, 391–397. [Google Scholar] [CrossRef]
- Shwe, S. S.; Leung, D. W. M. Plant Regeneration from Eucalyptus Bosistoana Callus Culture. In Vitro Cellular & Developmental Biology - Plant 2020, 56, 718–725. [Google Scholar] [CrossRef]
- Faria, T.; Cezar, J.; Cézar, J.; Ribeiro-Kumara, C.; Silva Da Rocha Costa, R.; Nieri, E. M.; De Carvalho, D.; Brasil, J. E.; Pinto, P.; Rodrigues, A.; Neto, S.; Brondani, G. E. Use of Biodegradable Polyester-Based Microvessels for Micropropagation of Mature Eucalyptus Microcorys. N Z J For Sci 52, 1–13. [CrossRef]
- Skupa Petr and Opatrný, Z. and Opatrný, Z. and P. J. Auxin Biology: Applications and the Mechanisms Behind. In Applied Plant Cell Biology: Cellular Tools and Approaches for Plant Biotechnology; Nick Peter and Opatrny, Z., Ed.; Springer Berlin Heidelberg: Berlin, Heidelberg, 2014; pp. 69–102. [Google Scholar] [CrossRef]
- Teale, W. D.; Paponov, I. A.; Palme, K. Auxin in Action: Signalling, Transport and the Control of Plant Growth and Development. Nat Rev Mol Cell Biol 2006, 7, 847–859. [Google Scholar] [CrossRef]
- Bunn, E. Development of in Vitro Methods for Ex Situ Conservation of Eucalyptus Impensa, an Endangered Mallee from Southwest Western Australia. Plant Cell Tissue Organ Cult 2005, 83, 97–102. [Google Scholar] [CrossRef]
- Kaur, S. In Vitro Regeneration of Shoots From Nodal Explants of Dendrobium Chrysotoxum Lindl. Journal of Horticultural Research 2017, 25. [Google Scholar] [CrossRef]
- WOODWARD, A. W.; BARTEL, B. Auxin: Regulation, Action, and Interaction. Ann Bot 2005, 95, 707–735. [Google Scholar] [CrossRef]
- Chen, J.; Ziv, M. The Effect of Ancymidol on Hyperhydricity, Regeneration, Starch and Antioxidant Enzymatic Activities in Liquid-Cultured Narcissus. Plant Cell Rep 2001, 20, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Brondani, G. E.; de Wit Ondas, H. W.; Baccarin, F. J. B.; Gonçalves, A. N.; de Almeida, M. Micropropagation of Eucalyptus Benthamii to Form a Clonal Micro-Garden. In Vitro Cellular & Developmental Biology - Plant 2012, 48, 478–487. [Google Scholar] [CrossRef]
- Gomes, F.; Canhoto, J. M. Micropropagation of Eucalyptus Nitens Maiden (Shining Gum). In Vitro Cellular & Developmental Biology - Plant 2003, 39, 316–321. [Google Scholar] [CrossRef]
- Ngomuo, M.; Mneney, E.; Ndakidemi, P. A. The <I>in Vitro</I> Propagation Techniques for Producing Banana Using Shoot Tip Cultures. Am J Plant Sci 2014, 05, 1614–1622. [Google Scholar] [CrossRef]
- Al-Aizari, A. A.; Al-Obeed, R. S.; Mohamed, M. A. H. Improving Micropropagation of Some Grape Cultivars via Boron, Calcium and Phosphate. Electronic Journal of Biotechnology 2020, 48, 95–100. [Google Scholar] [CrossRef]
- George Edwin, F. and Hall, M. A. and K. G.-J. De. The Components of Plant Tissue Culture Media I: Macro- and Micro-Nutrients. In Plant Propagation by Tissue Culture: Volume 1. The Background; George Edwin F. and Hall, M. A. and K. G.-J. De, Ed.; Springer Netherlands: Dordrecht, 2008; pp. 65–113. [Google Scholar] [CrossRef]
- Pérez-Tornero, O.; Burgos, L. Different Media Requirements for Micropropagation of Apricot Cultivars. Plant Cell Tissue Organ Cult 2000, 63, 133–141. [Google Scholar] [CrossRef]
- WHITE, P. J.; BROADLEY, M. R. Calcium in Plants. Ann Bot 2003, 92, 487–511. [Google Scholar] [CrossRef]
- Al-Mayahi, A. M. W. Effect of Calcium and Boron on Growth and Development of Callus and Shoot Regeneration of Date Palm Barhee. Canadian Journal of Plant Science 2020, 100, 357–364. [Google Scholar] [CrossRef]
- Brunoni, F.; Rolli, E.; Dramis, L.; Incerti, M.; Abarca, D.; Pizarro, A.; Diaz-Sala, C.; Ricci, A. Adventitious Rooting Adjuvant Activity of 1,3-Di(Benzo[d]Oxazol-5-Yl)Urea and 1,3-Di(Benzo[d]Oxazol-6-Yl)Urea: New Insights and Perspectives. Plant Cell, Tissue and Organ Culture (PCTOC) 2014, 118, 111–124. [Google Scholar] [CrossRef]
- Lopes da Silva, A. L.; Gollo, A.; Brondani, G.; Horbach, M.; Oliveira, L.; Machado, M.; LIMA, K.; Costa, J. Micropropagation of Eucalyptus Saligna Sm. from Cotyledonary Nodes. Pak J Bot 2015, 47, 311–318. [Google Scholar]
- Sharma, S.; Ramamurthy, V. Micropropagation of 4-Year-Old Elite Eucalyptus Tereticornis Trees. Plant Cell Rep 2000, 19, 511–518. [Google Scholar] [CrossRef]
- Gago, D.; Vilavert, S.; Bernal, M. Á.; Sánchez, C.; Aldrey, A.; Vidal, N. The Effect of Sucrose Supplementation on the Micropropagation of Salix Viminalis L. Shoots in Semisolid Medium and Temporary Immersion Bioreactors. Forests 2021, 12. [Google Scholar] [CrossRef]
- Joshi, I.; Bisht, P.; Sharma, V. K.; Uniyal, D. P. In Vitro Clonal Propagation of Mature Eucalyptus F1 Hybrid (Eucalyptus Tereticornis Sm. x E. Grandis Hill Ex. Maiden). Silvae Genet 2003, 52, 110–113. [Google Scholar]
- Trueman, S. J.; Richardson, D. M. In Vitro Propagation of Corymbia Torelliana C. Citriodora (Myrtaceae) via Cytokinin-Free Node Culture. Aust J Bot 2007, 55, 471–481. [Google Scholar] [CrossRef]
- Sandhu, M.; Wani, S. H.; Jiménez, V. M. In Vitro Propagation of Bamboo Species through Axillary Shoot Proliferation: A Review. Plant Cell, Tissue and Organ Culture (PCTOC) 2018, 132, 27–53. [Google Scholar] [CrossRef]
- Nadgauda, R. S.; Parasharami, V. A.; Mascarenhas, A. F. Precocious Flowering and Seeding Behaviour in Tissue-Cultured Bamboos. Nature 1990, 344, 335–336. [Google Scholar] [CrossRef]
- Shen, G.; Tan, S.; Sun, X.; Chen, Y.; Li, B. Experimental Evidence for the Importance of Light on Understory Grass Communities in a Subtropical Forest. Front Plant Sci 2020, 11. [Google Scholar] [CrossRef]
- Mankessi, F.; Saya, A.; Baptiste, C.; Nourissier, S.; Monteuuis, O. In Vitro Rooting of Genetically Related Eucalyptus Urophylla × Eucalyptus Grandis Clones in Relation to the Time Spent in Culture. Trees 2009, 23, 931–940. [Google Scholar] [CrossRef]
- Fett-Neto, A. G.; Fett, J. P.; Goulart, L. W. V.; Pasquali, G.; Termignoni, R. R.; Ferreira, A. G. Distinct Effects of Auxin and Light on Adventitious Root Development in Eucalyptus Saligna and Eucalyptus Globulus. Tree Physiol 2001, 21, 457–464. [Google Scholar] [CrossRef]
- Fogaça, C. M.; Fett-Neto, A. G. Role of Auxin and Its Modulators in the Adventitious Rooting of Eucalyptus Species Differing in Recalcitrance. Plant Growth Regul 2005, 45, 1–10. [Google Scholar] [CrossRef]
- Watt, M. P.; Thokoane, N. L.; Mycock, D.; Blakeway, F. In Vitro Storage of Eucalyptus Grandis Germplasm under Minimal Growth Conditions. Plant Cell Tissue Organ Cult 2000, 61, 161–164. [Google Scholar] [CrossRef]
- Xiao, Y.; Niu, G.; Kozai, T. Development and Application of Photoautotrophic Micropropagation Plant System. Plant Cell, Tissue and Organ Culture (PCTOC) 2011, 105, 149–158. [Google Scholar] [CrossRef]
- Miranda, N. A.; Xavier, A.; Otoni, W. C.; Gallo, R.; Gatti, K. C.; de Moura, L. C.; Souza, D. M. S. C.; Maggioni, J. H.; Santos, S. S. de O. Quality and Intensity of Light in the In Vitro Development of Microstumps of Eucalyptus Urophylla in a Photoautotrophic System. Forest Science 2020, 66, 754–760. [Google Scholar] [CrossRef]
- Frade, S. R. do P.; Souza, D. M. S. C.; Fernandes, S. B.; Avelar, M. L. M.; Molinari, L. V.; Gonçalves, D. S.; Magalhães, T. A.; Brondani, G. E. Spectral Quality Influence on in Vitro Morphophysiological Responses of Eucalyptus Dunnii Maiden and Eucalyptus Grandis w.Hill Ex Maiden x E. Urophylla s.t.Blake. N Z J For Sci 2023, 53. [Google Scholar] [CrossRef]
- Souza, D. M. S. C.; Avelar, M. L. M.; Fernandes, S. B.; Silva, E. O.; Duarte, V. P.; Molinari, L. V.; Brondani, G. E. Spectral Quality and Temporary Immersion Bioreactor for in Vitro Multiplication of Eucalytpus Grandis × Eucalyptus Urophylla. 3 Biotech 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Matheus, D.; Souza, S. C.; Martins, A. R.; Fernandes, S. B.; Lopes Martins Avelar, M.; Vaz Molinari, L.; Santos Gonçalves, D.; Brondani, G. E. In Vitro Multiplication of Eucalyptus Pilularis and Eucalyptus Grandis x E. Urophylla (Urograndis Eucalypt): Effect of Light Quality in Temporary Immersion Bioreactor; 2022; Vol. 20.
- Roque-Borda, C. A.; Kulus, D.; de Souza, A.; Kaviani, B.; Vicente, E. F. Cryopreservation of Agronomic Plant Germplasm Using Vitrification-Based Methods: An Overview of Selected Case Studies. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Padayachee, K.; Watt, M. P.; Edwards, N.; Mycock, D. J. Cryopreservation as a Tool for the Conservation of Eucalyptus Genetic Variability: Concepts and Challenges. Southern Forests: a Journal of Forest Science 2009, 71, 165–170. [Google Scholar] [CrossRef]
- Blakesley, D.; Kiernan, R. Cryopreservation of Axillary Buds of a Eucalyptus Grandis x Eucalyptus Camaldulensis Hybrid. Cryo Letters 2001, 22, 13–18. [Google Scholar] [PubMed]
- Hung, C. D.; Trueman, S. J. Preservation of Encapsulated Shoot Tips and Nodes of the Tropical Hardwoods Corymbia Torelliana × C. Citriodora and Khaya Senegalensis. Plant Cell, Tissue and Organ Culture (PCTOC) 2012, 109, 341–352. [Google Scholar] [CrossRef]
- Costa Souza, D. M. S.; Fernandes, S. B.; Avelar, M. L. M.; do Prado Frade, S. R.; Molinari, L. V.; Gonçalves, D. S.; Pereira Pinto, J. E. B.; Brondani, G. E. Light Quality in Micropropagation of Eucalyptus Grandis × Eucalyptus Urophylla. Scientia Forestalis/Forest Sciences 2020, 48. [Google Scholar] [CrossRef]
Figure 1.
Steps involved in Eucalypt micropropagion starting with collection and preparation of explants that could be axial leaf, shoot tips, buds or seeds, followed by inoculating and culturing in specific media with optimized concentration of the hormones and other nutritional supplements based on the requirement of the plant as well envionmental conditions. After shoot proliferation the raised plants are subjected to root proliferation and root hardening in the field conditions or else the germplasm of the raised plants could be preserved for future use in similar related protocols.
Figure 1.
Steps involved in Eucalypt micropropagion starting with collection and preparation of explants that could be axial leaf, shoot tips, buds or seeds, followed by inoculating and culturing in specific media with optimized concentration of the hormones and other nutritional supplements based on the requirement of the plant as well envionmental conditions. After shoot proliferation the raised plants are subjected to root proliferation and root hardening in the field conditions or else the germplasm of the raised plants could be preserved for future use in similar related protocols.
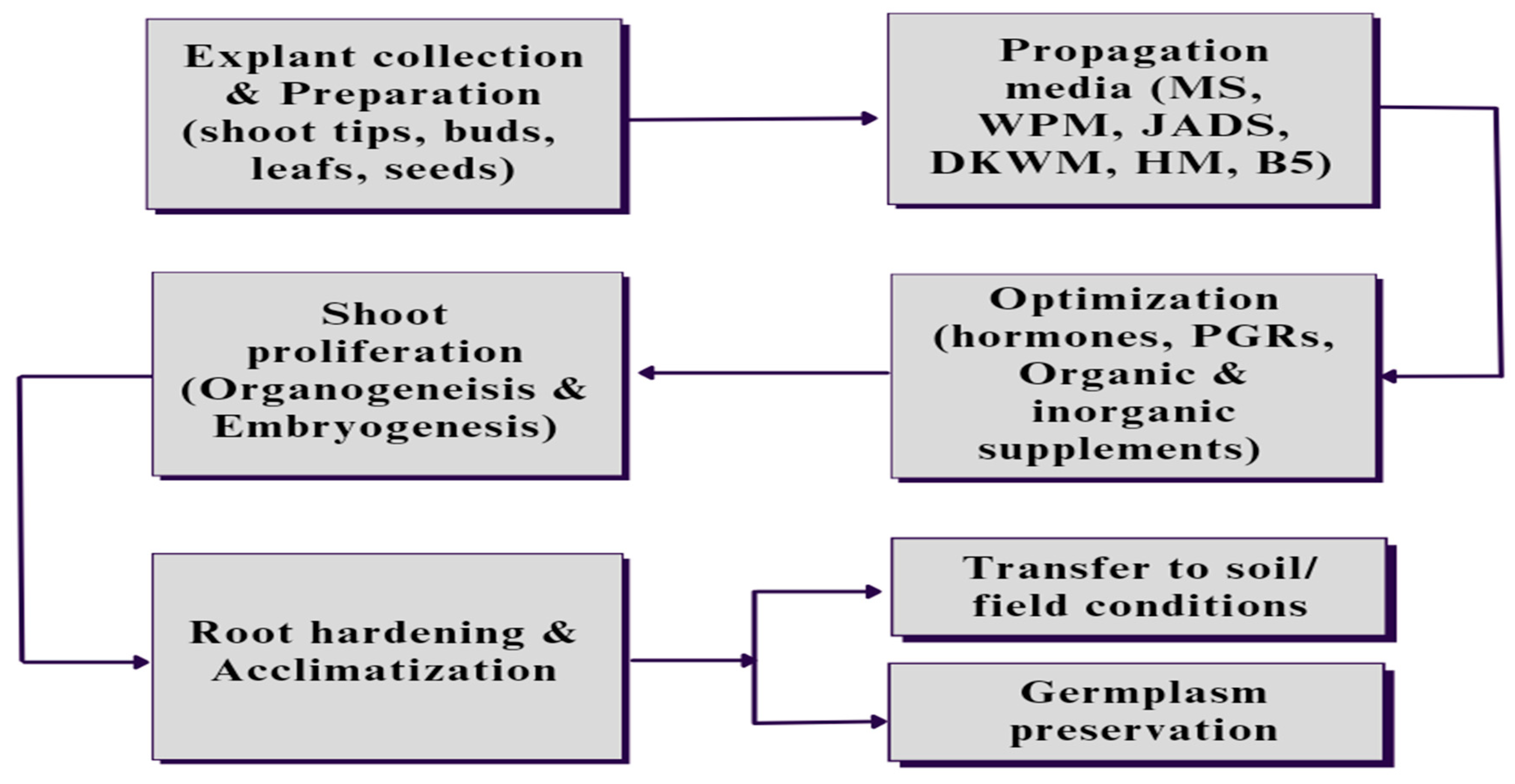
Figure 2.
Factors affecting the clonal propagation of Eucalypt.
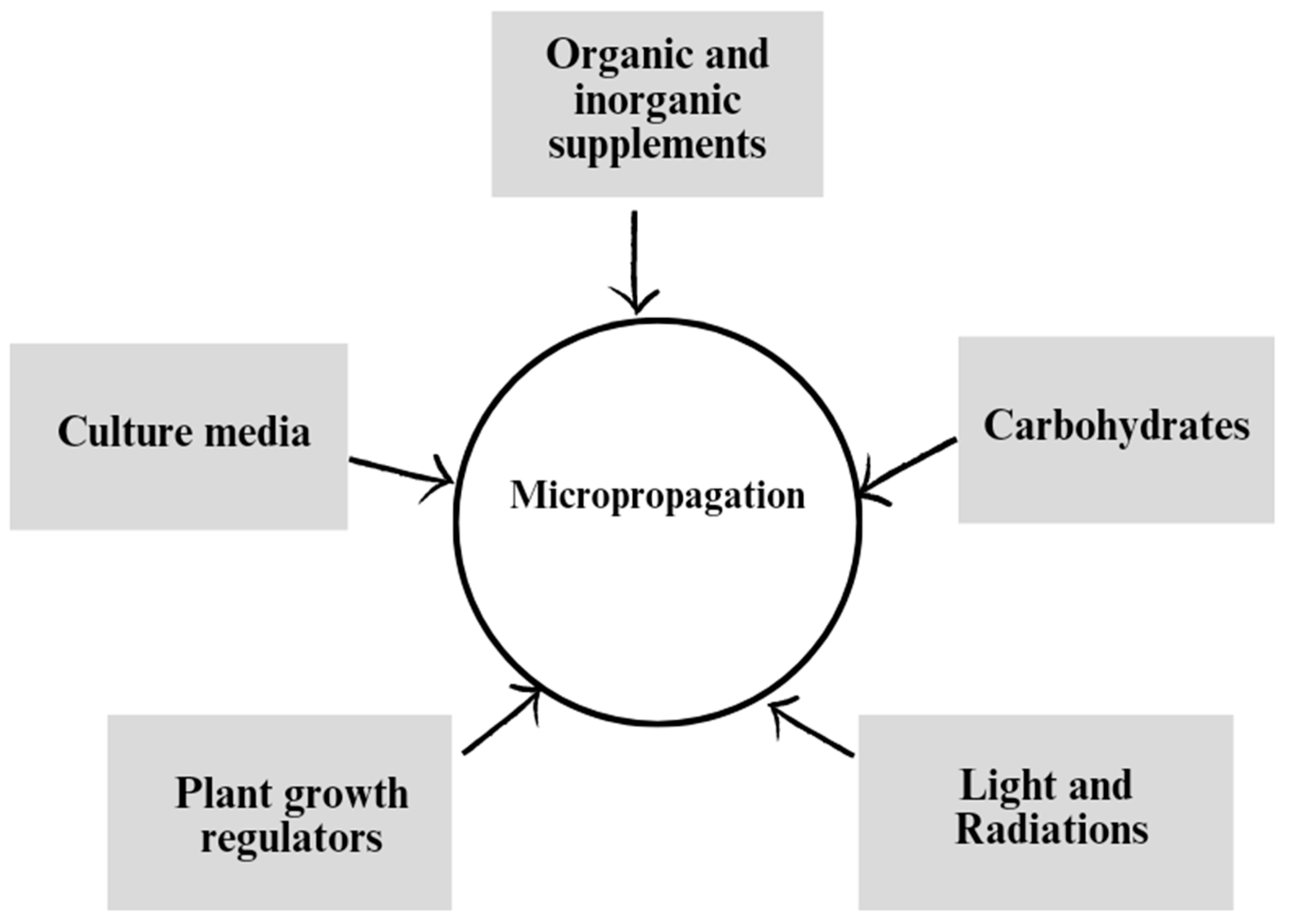

Table 1.
The above table highlights various sources of explants, culture media, additives etc. that have been previously used in Eucalypt micropropagation. Abbreviations used in the table are, 2,4-D- 2,4-Dichlorophenoxyacetic acid; ABA- Abscisic acid; BA- 6-benzyladenine; BAP- 6-benzylamino purine; CaCl2- Calcium chloride ; CO2 – Carbon dioxide; Cu- Copper; DTE/ DTT- Dithioerythritol; DKW- Driver and Kuniyaki Walnut; EDM- E. Dunnii basal medium; Fe- Iron; GA3- Gibberellic acid; HCl- Hydrochloric acid; IAA- Indole-3-acetic acid; IBA-Indole-3-butyric acid; JADS- Juan, Antonio, Diva and Silvian; K- Potassium; LED- Light Emitting Diode; Mn- Manganese; NAA- α-naphthaleneacetic acid; PGRs- Plant Growth Regulators; PSV2- Plant Vitrification Solution 2; PVP- Polyvinylpyrrolidone; PVPP- Polyvinylpolypyrrolidone; QL- Quoirin & Lepoivre medium; S- sulfur; SH- Schenk and Hildebrandt; TDZ- Thidizuron; WPM- Woody Plant Medium; Zn- Zinc.
Table 1.
The above table highlights various sources of explants, culture media, additives etc. that have been previously used in Eucalypt micropropagation. Abbreviations used in the table are, 2,4-D- 2,4-Dichlorophenoxyacetic acid; ABA- Abscisic acid; BA- 6-benzyladenine; BAP- 6-benzylamino purine; CaCl2- Calcium chloride ; CO2 – Carbon dioxide; Cu- Copper; DTE/ DTT- Dithioerythritol; DKW- Driver and Kuniyaki Walnut; EDM- E. Dunnii basal medium; Fe- Iron; GA3- Gibberellic acid; HCl- Hydrochloric acid; IAA- Indole-3-acetic acid; IBA-Indole-3-butyric acid; JADS- Juan, Antonio, Diva and Silvian; K- Potassium; LED- Light Emitting Diode; Mn- Manganese; NAA- α-naphthaleneacetic acid; PGRs- Plant Growth Regulators; PSV2- Plant Vitrification Solution 2; PVP- Polyvinylpyrrolidone; PVPP- Polyvinylpolypyrrolidone; QL- Quoirin & Lepoivre medium; S- sulfur; SH- Schenk and Hildebrandt; TDZ- Thidizuron; WPM- Woody Plant Medium; Zn- Zinc.
| Explant-source/type | Culture media/PGRs/additives | Experimental outcomes/remarks/productivity/root hardening etc. | References |
|---|---|---|---|
| Nodal segments | MS media, WPM, 0.5 mgL-1 IBA, 1.0mgL-1 BAP, B5, NAA, Kinetin | Full strength MS media supplemented with 1.0mgL-1 BAP showed best shoot elongation; ½ MS media supplemented with 0.5 mgL-1 IBA showed best rooting; the resulting hybrid produced yields 3-5 times more wood than parental species | [5] |
| Nodal segments (30–32-year trees) | MS media, WPM, B5 medium, SH medium, IBA, BAP, NAA, | Best in-vitro studies were obtained from the explants collected during the period of January to February and August to September; ½ MS media + BAP resulted in best rooting (92%); Up to 98% plants survived acclimatization | [6] |
| Stem cuttings and axillary buds | MS Media, 2.22 µm BA, 1.16µm Kinetin, 0.029µm gibberellic acid, 400mgL-1 PVP, 30gL-1 sucrose | Micropropagation and micro cutting showed higher adventitious rooting (24.8-100% and 43-95% respectively) than stem cuttings (9.3-75.5%) | [12] |
| Maintained Elite clones (KE8, CE2, T1, and Y8) | Basal MS media, 2.5 µM BA, 0.5 µM NAA, D-mannitol (0, 250, 500, 750, and 1000 mM) | Culture Growth Index of all clones was reduced significantly because of drought stress | [17] |
| - | Full and ½ strength MS media | Concentrations of IAA, IBA as well the stem anatomy has no effect on the rooting potential of shoots | [22] |
| Nodal cuttings | IAA, IBA (0, 1, 3, 8gKg-1) | The position from which explant is harvested can affect the rooting potential and seed vigor. Explants obtained from 7/8 and 9/10 apical position showed enhanced rooting and shooting | [25] |
| Cuttings from 6-month-old parental plants | MS media, IAA, meta-topolin, kinetin, BAP, vit. B5, Biotin, Sucrose | 0.5mgL-1 meta-topolin and 1mgL-1 IAA enhanced shoot elongation as well bud proliferation while 0.5mgL-1 IAA resulted in the most consistent rooting percentages. Moreover, equal expression of AUX1 and PIN1 transporter genes increased responsiveness towards PGRs | [29] |
| Nodal segments | MS media, 58mM sucrose, 0.5 μM NAA, 2.5 μM BA, | Media supplemented with 1.0 μM 2,4-D, 5.0 μM BA and 500 mgL-1 cefotaxime showed maximum (44.6%) shoot bud organogenesis | [33] |
| Nodal segments | MS media, 2mgL-1 BAP, 0.1mgL-1 NAA | Media supplemented with 0.5mgL-1 showed best shoot elongation, 1/2MS + 1mgL-1 IAA Showed best root induction and elongation, and direct regeneration was observed in MS + 20:1 BAP: NAA | [34] |
| Young shoot segments | WPM, MS media, 2iP, NAA, BAP, sucrose | Media suppelmented with BA resulted in 99% shoot proliferation, media supplemented with 2iP resulted in 93% shoot regeneration and IBA promoted rooting in 60% of the clones. | [35] |
| Nodal segments | EDM basal media (a novel basal media for E. dunnii) supplemented with 20gL-1 sucrose and without PGRs | Higher Fe, Cu, Zn and Mn concentrations in EDMm media increased rooting. Moreover, high S and K concentrations in EDMm increased growth rate and multiplication. Also no Fe chlorosis / oxidation was observed in shoots cultured on EDMm | [37] |
| Zygotic embryo | One of the following media and growth regulators: ½ or full-strength MS media/ WPM/ B5/ DKW/ JADS media/ 3mgL-1 NAA/ 10mL-1 silver nitrate/ 0.5mgL-1 DTT/ 100mgL-1 ascorbic acid/ 0.5mgL-1 DTE/ 1%m/v PVP/ 1% m/v PVPP/ 0.01% w/v activated charcoal | The best media for somatic embryogenesis was B5 and MS. Moreover, Silver nitrate, activated charcoal and DTE reduced browning of explants | [41] |
| Somatic embryos | MS media supplemented with 3mgL-1 NAA | MS media scarce of PGRs is highly efficient to promote cotyledonary embryo proliferation and germination | [42] |
| Zygotic embryo cotyledons | Hormone free MS media | Reserve accumulation of mature zygotic embryos was analyzed. Cotyledonary somatic embryos possess low density of starch and no lipids/ proteins. | [43] |
| Axillary shoots | ½ MS, 4.4μM, 1μM NAA, 1gL-1 sucrose | WPM and QL media supplemented with Gibberlic acids showed enhanced shoot proliferation, ½ WPM supplemented with 20 μM IBA showed enhanced rooting, 67% Plantlet hardening was achieved | [47] |
| Shoot segments | MS media, 0.02mgL-1 IBA | Vitron vessel placed in Low Photon Flux density at 3000ppm CO2 for 24 hours/ day yielded best growth and quality of plantlets | [52] |
| Seedlings raised from seeds | - | Low temperatures 18°C/13°C to 23°C/18°C (day/night) reduced the number of harvested cuttings however did not affect the percentage of roots proliferated from cuttings. While, increasing temperature 33°C/28°C resulted in an increased number of cutting per stock plant. | [53] |
| Hypocotyl segments and cotyledonary leaves | MS media supplemented with different concentrations of NAA and TDZ, 0.8gL-1 PVP, 0.1gL-1 biotin, 0.1gL-1 calcium pantothenate, 30gL-1 sucrose | 0.44 µM BAP increased regeneration of adventitious buds | [59] |
| Zygotic embryos and cotyledons | MS media supplemented with 3gL-1 sucrose and different concentrations of NAA, 2,4-D, BA, ABA | 1mgL-1 NAA resulted in maximum callus induction, the frequency of callus proliferation depends on the age of explant with 10-year-old explants showing maximum proliferation, highest frequency of somatic embryogenesis was observed in callus from mature zygotic embryos, low ABA concentrations increased number of somatic embryos | [60] |
| Nodal segments | 1/2 MS supplemented with different concentrations of BAP, NAA and GA3 | 0.050 mgL-1 BAP achieved optimal bud proliferation + 0.50 mgL-1 NAA while ½ MS media supplemented with 0.2-1 and 0.10 mgL-1 GA3 + 0.10 mgL-1 BAP showed highest shoot elongation | [61] |
| Nodal segments | MS media without PGRs | Media free from GA3 + BAP resulted in best shoot elongation, WPM + 0.05mgL-1 NAA + 0.5mgL-1 BAP resulted in maximum axillary bud proliferation | [71] |
| Nodal segments | ½ MS media, De-Fossard Medium, 0.9 µgL-1 BA, 0.5 µM NAA | Best multiplication rate (2.25) was achieved, 93% of the plants survived acclimatization | [72] |
| Nodal segments | MS media supplemented with 0.05 μM NAA, 0.4 μM BA, 1mgL-1 nicotinic acid, 1mgL-1 pyridoxine-HCl, 1mgL-1 thiamine, 2mgL-1 glycine, 50mgL-1 myo-inositol, 30gL-1 sucrose | Endogenous rhythms cause time related fluctuations resulting in rooting variations among closely related genotypes | [88] |
| Epicotyl segments | ½ MS supplemented with 1/6x CaCl2, 2% (w/v) sucrose | Auxin reduced mean rooting time, light conditions did not affect the rooting efficiency, with increased age decreased rooting capability was observed | [89] |
| Axillary buds | ½ MS supplemented with 1gL-1 ABA | Encapsulation by calcium alginate and storing under low light intensities resulted in the preservation of cultures for up to 3 months without affecting its viability | [91] |
| Apical shoots | MS media supplemented with 0.04mgL-1 BA, 1% Sucrose, with/ without charcoal | 38%-85% survival was observed with plants exposed to PSV2 for 30 min in liquid nitrogen | [99] |
| Nodal segments | MS media supplemented with 30gL-1 sucrose | Best in-vitro establishment, multiplication, shooting, and rooting was achieved by Red-Blue LEDs and Fluorescent lamps | [101] |
Table 2.
The above table highlights the media composition, Additives, and sterilant used in various successful attempts of Eucalypt hybrid production.
Table 2.
The above table highlights the media composition, Additives, and sterilant used in various successful attempts of Eucalypt hybrid production.
| Species | Explant | Sterilant | Media | PGR (if any) | Area studied | Scope of work | References |
|---|---|---|---|---|---|---|---|
| E. camaldulensis × E. tereticornis and E. torelliana × E. citriodora | Nodal segments from mature trees (30-32 yrs) | 0.15% HgCl2 |
MS medium, WPM, SH medium, B5 medium | BAP, NAA | Hybridisation of Eucalypt species | Two hybrids developed that showed suprior performance than parental genotypes | [17] |
| E. grandis × E. nitens | Axial buds | 10g l-1 CaOCl | MS media | BAP, IAA, metatopolin, kinetin | Individual evalualtion of each stage of micropropagation | Auxins are principal components of media and expression of different auxin transporters might be used as markers to identify Eucalypt spp amenable for micropropagation | [29] |
| E. erythronema × E. stricklandii | Seedlings germinated in vitro | 3% NaOCl | MS media supplemented with sucrose 30gL-1 | IBA, NAA, Gibberlic acids | First micropropagation report of ornamemtal Eucalypt spp. | Successful micorpropagation from juvenile seedlings acheived | [47] |
| E.benthamii × E. dunni | Nodal segments from 1 year old plants | NaOCl | ½ strength MS media | PVP40, NAA, BAP | Optimization of chlorine concentration for explant sterlisation and optimum ratio of PGRs for shoot elongation | 0.5% NaOCl is suggested for nodal segments ; 0.50 mg L-1 BAP + 0.05 mg L-1 NAA provides highest number of bud proliferation | [61] |
| E. erythronema × E. stricklandii | Nodal segments | 1% NaOCl | MS media supplemented with sucrose 30gL-1 | 0.05μM NAA and 2.22μM BAP | Effect of different light intensities on micropropagation efficiency | Red/ blue LEDs and Florescent light results in higher vigor, High photosynthesis, increased shoot and root proliferation | [101] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



