
Submitted:
30 June 2023
Posted:
30 June 2023
You are already at the latest version
Abstract
Spermatogonial stem cells (SSCs) are the only primitive spermatogonial cells in males that can naturally transmit genetic information to their offspring and replicate throughout their lives. Phospholipase D family member 6 (PLD6) has recently been found to be a surface marker for SSCs in mice and boars; however, it has not been validated in cattle. The results of RT-PCR and qRT-PCR found that the relative expression of PLD6 gene in the testicular tissues of 2-year-old Simmental calves was significantly higher than that of 6-month-old calves. Immunofluorescent staining further verified the expression of PLD6 protein in bovine spermatogenic cells like germ cell marker VASA. Based on multiple bioinformatic databases, PLD6 is a conservative protein which has high homology with mouse Q5SWZ9 protein. It is closely involved in the normal functioning of the reproductive system. Molecular dynamics simulation analyzed the binding of PLD6 as a phospholipase to cardiolipin (CL), and PLD6-CL complex showed high stability. The protein interaction network analysis showed that there is a significant relationship between PLD6 and piRNA binding protein. PLD6 acts as an endonuclease and participates in piRNA production. In addition, PLD6 in bovine and mouse testes has a similar expression pattern with the spermatogonium-related genes VASA and PIWIL2. In conclusion, these analyses imply that PLD6 has a relatively high expression in bovine testes and could be used as a biomarker for spermatogenic cells including SSCs.
Keywords:
Bovine
; Bioinformatics
; Phospholipase D family member 6
; Testes
; Spermatogonial stem cells
1. Introduction
Phospholipase D6 (PLD6) is the sixth member of the phospholipase superfamily, known as Zucchini (also known as MitoPLD) in Drosophila. The main function of PLD6 appears to be the production of signaling lipid phosphatidic acid (PA) on mitochondrial surface, which is involved in the regulation of mitochondrial division and fusion, and thus functions as a phospholipase [1,2]. The function and dynamics of mitochondria are closely related to multiple biological processes such as cell growth, proliferation and differentiation [3,4]. Additionally, PLD6 is also involved in the production of primary PIWI-interacting RNA (piRNA) which regulates male germ cell development and genome stability. Whereas in the primary piRNA biogenesis, PLD6 functions as a nucleic acid endonuclease [5,6,7]. In male mice deprived of PLD6, the quantity of piRNAs during spermatogenesis is drastically reduced, meiosis is blocked, and ultimately the main phenotype is sterility [8,9]. These evidences suggest that PLD6 plays an essential role in the development of male germ cells.
Spermatogenesis is a complex physiological process that starts with the differentiation of spermatogonia and ends with the formation of mature sperm. Spermatogonia reside in the base of the seminiferous epithelium and are generally divided into three categories, type A, intermediate and type B spermatogonia. For most animals, a small population in type A spermatogonia are regarded as spermatogonial stem cells (SSCs). As the only primitive germ cells in male, SSCs transmit genetic information to their offspring and replicate throughout the lives by self-renewal, proliferation and differentiation. They play an irreplaceable role in the spermatogenesis and germline evolution [10,11]. It is generally believed that SSCs reside in microenvironments/niches of the seminiferous tubules. These niches consist of Sertoli cells, Leydig cells, the basement membrane and peritubular myoid cells, providing SSCs with a microenvironment for growth and growth factors needed to maintain their plasticity [12]. Although they are the foundation of continuous spermatogenesis, SSCs have relatively low numbers in the testis, approximately accounting for only 0.02-0.03% in the cell suspension of whole testis [13]. Thus, identification of these cells by varied methods including biomarker validation is essential to elucidate their functions. At present, surface labeling of SSCs has made significant progress in rodents [14,15]. In domestic animals, some markers such as GFRα-1, PLZF [16], THY1, and UCHL1 [17] have been used to enrich and identify the undifferentiated spermatogonia. Recent studies have shown that PLD6 is expressed in the plasma membrane of mouse SSCs [18] and can be a molecular marker for pre-sexually mature porcine SSCs [19]. However, it has not been confirmed yet in bovine undifferentiating spermatogonia including SSCs.
In this study, reverse transcriptase polymerase chain reaction (RT-PCR), quantitative real-time PCR (qRT-PCR) and immunostaining were used to reveal the expression and localization of PLD6 in bovine testes. The bioinformatics techniques were further employed to investigate the secondary structure and molecular characteristics, as well as possible functions and signaling pathways of PLD6 in bovine spermatogenesis.
2. Results
2.1. Results
2.1.1. Expression of PLD6 in Bovine Testes
The RT-PCR and qRT-PCR analyses revealed that, with GAPDH as the internal reference, PLD6, UCHL1 and the germ cell marker VASA all transcriptionally expressed in both 6-month-old and 2-year-old bovine testes (Figure 1a,b). Immunofluorescent staining showed that PLD6 was observed in all germ cells similar to VASA in the seminiferous epithelium, while UCHL1 was mainly detected in spermatogonia on the basement membrane of the seminiferous tubules (Figure 1c).
2.1.2. Phylogenetic and Structural Features of Bovine PLD6
The phylogenetic tree was constructed by Neighbour-Joining, all the bootstrap values were > 70, indicating that the phylogenetic tree has high credibility. It demonstrated that PLD6 protein of our study grouped together with bovine BIBE10 protein of the same genus, indicating a similar degree of evolution. Additionally, bovine PLD6 exhibited a close relation to mouse Q5SWZ9 protein, with a bootstrap value of 100. The length of the branch in the tree was correlated to the level of genetic variability, indicating that bovine PLD6 is more evolved than that in mouse, but less evolved than that in Homo sapiens (Figure 2a). Evolutionary trace analysis (Table 2) showed that PLD6 contained 34 important trace residues that are subject to evolutionary pressure (Importance score < 25%), in which 17 residues are subject to the most prominent selection pressure (Score < 5%). The SLAC algorithm revealed that the positively selected sites account for 19.33%, while the purification selection accounts for 30.57%.

Table 1.
Primers used for qRT-PCR.
| Gene | Primer Sequence(5′→3′) | Fragment Size (bp) | Gene ID | Accession Number |
|---|---|---|---|---|
| GAPDH | F: TGACCCCTTCATTGACCTTC | 250 | 281181 | NM_001034034 |
| R: TACTCAGCACCAGCATCACC | ||||
| VASA | F: TTGGGACTTGTGTAAGAGCTGT | 98 | 493725 | NM_001278643 |
| R: CTTGACTTGTTTGAGGC | ||||
| UCHL1 | F: CCTTCCCTGGGCAGGTGCGCGC | 280 | 514394 | NM_001046172 |
| R: GGCTGTAGAACGCAAGAA | ||||
| PLD6 | F: GTGGACAGGAAGGTGCTGAT | 108 | 526651 | NM_001271990 |
| R: TACTCCTCGTCCTCCACGAT |
As shown in Figure 2b, homology modeling and conservation analysis exhibited that PLD6 protein has a “bell-shaped” structure, including 37.72% α-helix, 9.25% β-sheet, 43.64% random coil and other secondary structures. The neural convolutional network predicts the solvent accessibility of amino acid residues, showing that 41.36% PLD6 amino acids have low solvent accessibility and are buried (marked “b”), while 58.63% of PLD6 amino acids are exposed (marked “e”). The structural and functional properties of conserved residues were analyzed by convolutional neural network (CNN) of the Consurf server. Among them, residues that are highly conserved and exposed on the protein surface (functional residue, marked “f”), are thought to be involved in the biological functions of PLD6, and residues that are highly conserved and buried inside the protein (structural residue, marked “s”), are important for the maintenance of the structural features of the protein. This finding well constructs the correlation between protein structural features and function. Most of the conservative amino acids are located on the surface, which is speculated to be involved in the functioning of PLD6. Like most PLD family members, PLD6 has several highly conserved regions (Figure 2b), including Val41-Ser46 (VLFFPS), Glu91-Ser99 (ELCLFAFSS), Met151-Ala156 (MHHKFA) and Leu163-Trp170 (LITGSLNW). The prediction of subcellular location by PSORT Prediction server shows that PLD6 is located in the outer mitochondrial membrane, and the Val10-Val32 sequence is a transmembrane segment by TMHMM 2.0 analysis.
2.1.3. Molecular Dynamics Simulation and Binding Mode Analysis
Molecular dynamics simulation and binding mode analysis were all implemented using Gromacs software, which has built-in multiple commands. The analysis shows that the complexes PLD6-CL exhibit a high stability during the simulation with a mean Root-Mean-Square Deviation (RMSD) value of 0.275 ± 0.042 nm (max = 0.363 nm). It was noteworthy that the RMSD values of the system gradually decreased after 35 ns, with a more pronounced conformational shift at about 115 ns, and maintained a significant steady state after 150 ns until 200 ns (Figure 3a). The radius of gyration (Rg) reflects the effect of CL molecules on the tightness of PLD6 folding, with values ranging from 1.548-1.667 nm (Figure 3b). Moreover, it was not difficult to find that the above structures have a high degree of overlap, i.e., when superimposing and comparing the PLD6 structures at 0-200 ns, that was, the binding of CL molecule did not significantly affect the PLD6 conformation, and even increased the folding tightness and orderliness (Figure 3g). Meanwhile, the root means square fluctuation (RMSF) values of PLD6 in the complex were in the range of 0.052-0.921 nm, as shown in Figure 3c, and PLD6 contains a region with prominent structural flexibility in one segment (Leu55-Gly72). Overlapping the conformations at different moments, it was also found that the above region showed obvious structural changes, but the region that played the function of phospholipase has a high stability, which was favorable for the binding of CL molecule and hydrolysis reaction.
In addition, the relationship between protein Gibbs free energy and RMSD and Rg were revealed by using principal component analysis (PCA) method, so as to clarify the correlation between relative free energy and protein conformation. The red region was the energy peak, the transition region fading from red to blue was the energy mound, which can be considered as the energy potential connecting the sub-stable states, and the dark blue part served as the energy lowland, indicating the aggregation region of the minimum conformation of relative Gibbs free energy (Figure 3d). Subsequently, the minimum energy conformation was extracted and the binding mode analysis was performed. From Figure 3e, it could be seen that the O atoms abundant in the end groups of CL molecule could bind to residues Tyr150, His152, His153, Lys154, Thr165, Gly166 and Asn181 of the functional region of PLD6 phospholipase and form 9 hydrogen bonds, and it was worth noting that the formed hydrogen bonds were symmetrically distributed around the CL molecule, and this binding pattern made the force more uniform. During the simulation, it was also found that PLD6 formed about 7-10 hydrogen bonds with CL stably. Meanwhile, the aromatic ring of cationic residue His-153 forms a salt bridge with the P atom in the CL molecule, which was conducive to the stable fitting of the CL molecule with certain structural flexibility in this region and provided a structural basis for the subsequent hydrolysis reaction. In addition, consistent with the findings in RMSF, frequent transitions between secondary structures such as the turn and bend were also evident in the Leu55-Gly72 region (Figure 3f).
2.1.4. PPI Network
The final average clustering coefficient (0.808) was obtained from the constructed PPI network, and the PPI enrichment P-value was < 1.0e-16. A total of 60 related proteins were enriched, which were clustered into 5 main sub-networks (Figure 4a). The Top 10 Hub protein including PIWIL4, TDRD9, MAEL, ASZ1, VASA (DDX4), GK2, MGLL, TDRD5, TDRD6 and HENMT1 were screened (Figure 4b). Obviously, PLD6 has a significant interaction with the above-mentioned protein. It is well known that protein with strong associations is often related in functions. Most of these proteins are related to the silencing of PIWI/piRNA pathway genes with promoter hypermethylation, indicating that PLD6 is mainly involved in piRNA metabolism.
Note: The color is assigned according to the degree of connection in the figure. The darker the color, the more critical the site is in the interaction network; the thicker the connection, the stronger the interaction between the two proteins.
2.1.5. GO and KEGG Analysis
As shown in Figure 5, GO analysis was used to annotate PLD6 from three aspects, biological process (BP), molecular function (MF), and cell composition (CC). The BPs of PLD6 are mainly the formation of P granule organization, piRNA metabolism, DNA methylation and male meiosis during the gamete formation. The formation of P granule organization promotes the production of polar granules in primordial germ cells of many organisms, while piRNA regulates the growth and development of germ cells and stem cells [31,32]. Additionally, PLD6 can promote DNA methylation during gametogenesis and facilitate the establishment of DNA methylation patterns in gametes [33]. The MF of PLD6 mainly involves the phospholipase activity and participates in the catalytic hydrolysis of phosphodiester. For CC, PLD6 is mainly enriched in multiple membrane structures such as the outer mitochondrial membrane, outer cell membrane, and endoplasmic reticulum.
The regulation and metabolic pathways involving PLD6 were sorted from KEGG database (Table 3). PLD6 is mainly involved in the metabolism of glycerophospholipid, aminophosphonate, amino acid, and ether lipid as well as GnRH signaling pathway in cattle. The participation rate of PLD6 in the glycerophospholipid metabolism pathway is 61.08%. As a phospholipase, PLD6 can hydrolyze cardiolipin (CL) on the outer mitochondrial membrane to produce phosphatidic acid, and participates in the regulation of mitochondrial division and fusion [34].
2.1.6. Multi-Sequence Alignment and Expression Analysis of PLD6 by GEO Mining
Multi-sequence alignment (Figure 6a) shows that the amino acid sequence of bovine PLD6 has a high similarity with that of mouse (Mean identity: 83.33%, Mean similarity: 94.59%). The sequences of Arg35-Arg218 regions were almost identical except for a few amino acids. The functional site of PLD6 playing catalytic role is also the same. The GSE41637 data set in the GEO database and the ENSBTAG8009 entry in the EMBL-EBI database contain the transcriptomics sequencing data of various tissues of cattle and mouse. They were employed for tissue-specific analysis of PLD6, showing its expression levels both in bovine testes and mouse testes are the highest, approximately 30-196 times compared with those in other tissues (Figure 6b). This suggests that PLD6 is specifically expressed in bovine and mouse testicular tissues. The GSE926 data set records the relative expression of genes in mouse testes at 3, 6, 8, 10, 14, 18, 20, 30, 35, and 56 d (postnatal day). Among these genes, the expression of SSC-specific marker GFRα1 shows a decreasing trend from 3 d to 20 d, then maintains a certain level afterwards. The germ cell marker VASA and PIWI12 show an up-regulated expression from 3 d to 56 d, with a peak at 18-20 d, then decrease to a certain level and remain. We noticed that the transcription of PLD6 also shows a similar trend from 3 d to 56 d, indicating it has the similar expression pattern with VASA and PIWI12 (Figure 6c).
3. Discussion
The wall of the seminiferous tubule is composed of multiple layers of cells, which can be divided into spermatogenic cells and sustentacular/Sertoli cells. Before sexual maturity, there are only spermatogonia in the seminiferous tubules except for immature Sertoli cells; after sexual maturity, there are spermatogonia, primary spermatocytes, secondary spermatocytes, spermatids and sperm from the base of the seminiferous tubules to the lumen [35]. The formation process from spermatogonia to sperm is spermatogenesis, which ultimately produces mature male gametes, and SSCs are the source of spermatogenesis.
Previous studies and the data mining in MGI database (MGI, http://www.informatics.jax.org/) indicate that the absence of PLD6 significantly affects the development and function of the reproductive system (Figure 7). The phenotypic changes of PLD6tm1.1Hsas and PLD6tm1.1Mafr mutant mice are mainly abnormal sperm and seminiferous tubules, developmental disorders and dysfunction of the reproductive system [9]. Among them, azoospermia and absence of epididymis are mainly manifested in 7-week-old mice. Physiological abnormalities of the testis are indicated by severe damage to the RNA interacting with PIWI. Abnormal sperm morphology is manifested by presenting atypical spermatocytes which have condensed or swollen nucleus in the seminiferous tubules. Small testes and arrested male meiosis are also seen in PLD6 mutants. In addition, studies have shown that PLD6 is expressed in the plasma membrane of mouse SSCs [18], and it has also been suggested as a molecular marker for SSCs in pre-sexual boars [19]. In this study, we utilized combined experimental verification and biometric analysis to investigate the potential role of PLD6 in bovine spermatogenesis.
Firstly, we verified the expression of PLD6/PLD6 in bovine testes both transcriptionally and immunohistochemically. Transcriptional analysis verified the expression of PLD6 mRNA in bovine testis tissue, like undifferentiated spermatogonia marker UCHL1 and germ cell marker VASA (Figure 1a,b). Immunostaining further indicates PLD6 expression in bovine spermatogenic cells like VASA (Figure 1c). Thus, it could be used as a potential molecular marker for SSCs in sexually immature bovine testis tissues like in porcine testis [19].
PLD6 is a conserved protein. Phylogenetic analysis revealed that bovine PLD6 had the highest homology with the homologous EIBE10, and the Q5SWZ9 protein from mice came next (Figure 2a). Evolutionary trace analysis assigned importance values to amino acid residues, with lower values indicating lower variability during the evolution of the residue and greater importance for protein structure and function. The PLD6 protein contained 34 important and evolutionarily conserved trace residues, among which 17 residues were under the most prominent selection pressure (Table 2). To further investigate and quantify the magnitude of evolutionary pressure, the SLAC method was used to infer the substitution rate of gene sites, and the Ka/Ks ratio was calculated to determine the ratio of Ka and Ks. The smaller the Ka/Ks ratio, the lower the selection pressure and the more conserved the site. Positive selection sites account for 19.33%, and purifying selection sites account for 30.57%. We further predicted the secondary structure of PLD6 and analyzed the solvent accessibility of amino acid residues using a convolutional neural network. It was found that 41.36% of the PLD6 amino acid residues have low solvent accessibility and are located internally (labeled as “b”), while 58.63% of the residues are located on the surface (labeled as “e”). It should be noted that surface-exposed residues with high conservation (labeled as “f”) are closely related to the protein’s function, while conserved residues with high solvent accessibility (labeled as “s”) are important for maintaining specific structures [36] (Figure 2b). Like most members of the phospholipase D family, PLD6 protein contains several highly conserved regions, and these regions are listed in the results. In addition, we predicted the subcellular localization of PLD6 and found that it is located in the outer mitochondrial membrane, which provides a reference for its biological function as a molecular marker. Up to now, similar structure elucidation studies was reported only for mouse PLD6 (5,7), however no information is available for bovine PLD6.
Based on the molecular characterization and structure-function analysis, we found that PLD6 has a conserved H(x)K(x4)D motif (characterizing an enzymatically active region), which is hypothesized to interact with cardiolipin (CL) to generate the signaling molecule phosphatidic acid (PA) during the development of round to elongated spermatozoa, thereby inducing mitochondrial fusion. Molecular dynamics simulations provide more high-resolution details on time scales, conformational changes and energies, which we used to analyze the binding of PLD6 as a phospholipase to cardiolipin (CL). During the 200 ns simulation, the PLD6-CL complex exhibited high stability and binding to the CL molecule did not significantly affect the protein conformation of PLD6. In addition, PLD6 contains a segment with prominent structural flexibility, Leu55-Gly72, which is a Zinc ion-binding region, and a highly conserved and structurally stable amino acid in the Asp147-Ala174 region (a phospholipase active region). The above features are also beneficial for the phospholipid-catalyzed hydrolysis function. Notably, in the lowest energy conformation, seven residues, including Tyr150, His152, His153, and Lys154, etc. interact with the CL molecule in a hydrogen bond, and all of them are highly conserved (Figure 3). This finding suggests that there is a significant correlation between the structural features we elucidated and their biological functions, and these crucial residues could also be a key breakthrough in the structural study of PLD6 [37].
Studying protein interactions is beneficial in enhancing our comprehension of the connections between protein functions. The PPI network are composed of proteins that interact with each other through interactions which can be used to reveal life processes such as biological signalling, regulation of gene expression, energy and material metabolism, and cell cycle regulation. Proteins do not perform their functions alone, but work together with several proteins to accomplish biological events. The existence of interacting proteins is often functionally synergistic and coherent, and together they influence the biological processes, molecular functions and cellular components. Systematic analysis of protein interactions in biological systems provides a higher dimensional insight into how proteins work. In this study, we searched for a collection of proteins with PLD6 as the core through PPI network, clarified the regulatory pathways involved in PLD6 from a global perspective, and analysed their roles in spermatogenesis. The analysis indicates that PLD6 is closely related to the screened top 10 Hub proteins, suggesting that they are also strongly functionally related. Studies have shown that male sterility is associated with promoter hypermethylation-related silencing of PIWI/piRNA pathway genes, including ASZ1, HENMT1, DDX4, PLD6, MAEL, TDRD1 and TDRD5 [38,39,40,41]. PIWIL14 is essential for ensuring piRNA maturation by participating in primary or secondary biogenesis pathways [42]. It is also reported that PLD6 is synergistically involved in the formation of sperm mitochondrial sheaths with glycerol kinase 2 [4].
Furthermore, GO analysis indicates that the cellular components of PLD6 are mainly enriched in multiple membrane structures such as the outer mitochondrial membrane, the outer cell membrane and the endoplasmic reticulum. Studies reported that ectopic overexpression of PLD6 is localized in the outer mitochondrial membrane [43]. The endogenous PLD6 is localized in the plasma membrane of testicular germ cells [18,19], and is expressed in the Golgi apparatus of testicular germ cells, especially in sperm cells [44]. Subcellular fractionation of testicular cells showed that PLD6 is also expressed in other cellular components, such as cytoplasm, cell membrane, and nuclear components [9]. These evidences are consistent with our analysis and the immunostaining observations.
Sperm needs to rearrange organelles such as mitochondria in the final stage of maturation. Abnormal mitochondria can cause male sterility, leading to oligospermia and asthenospermia [45]. However, the dynamic regulation mechanism of mitochondria during spermatogenesis is still unclear. The enrichment of KEGG pathway in this study shows that PLD6 is mainly involved in the metabolism of glycerophospholipids, and the participation rate is as high as 61.08%. As a phospholipase, PLD6 mainly hydrolyzes the cardiolipin (CL) on the outer mitochondrial membrane to produce phosphatidic acid (PA), which in turn absorbs the phosphatase Lipin 1, converts PA into diglycerides and promotes mitochondrial division [2,8]. Additionally, PLD6 can continuously convert 1,2-diacylsn-glycerol into phosphatidylethanolamine and promote the formation of sperm mitochondrial sheaths to ensure the normal morphology of mitochondria and sperm tails [1,46]. PLD6 can also interact with oil kinase (Gyk)-like protein kinase 1 (Gykl1) and glycerol kinase 2 (Gk2) to induce the aggregation of PLD6 and phosphatidic acid (PA)-dependent mitochondria in cells. Both Gykl1 and Gk2 are specifically located in the mitochondria of sperm. Male mice lacking Gykl1 or Gk2 exhibit sperm tail defects, abnormal mitochondrial morphology, and disordered mitochondrial sheath formation [4]. PLD6 overexpression promotes mitochondrial aggregation, while PLD6 deletion results in disappearance of mitochondrial aggregation and tends to split [1,8]. Therefore, PLD6 plays a lipase function in the testis by participating in the regulation of mitochondrial division and fusion.
The GO analysis also revealed that the biological process of PLD6 is mainly involved in the metabolic process of Piwi-interacting RNA (piRNA). The piRNA is a type of small RNA with a length of about 30 nt isolated from mammalian germ cells, and mainly exists in mammalian germ cells and stem cells. It maintains the integrity of the animal germline genome by silencing transposons [47]. PLD6 has been proven to be a backbone-non-specific, single strand-specific nuclease, and its effect on germ cell development can be traced back to its function in the piRNA pathway. It cleaves either RNA or DNA substrates with similar affinity. The production of 5’ phosphate and 3’ hydroxyl termini suggests that it can directly participate in the processing of primary piRNA transcripts [7]. In mouse germ cells, PLD6 plays a conserved role in the piRNA production pathway and links lipid metabolism signals on the mitochondrial membrane with small RNA biogenesis [8,48,49]. In addition, the male PLD6-/- mice showed dramatically decreased piRNAs, damaged structure of the nucleoid body, blocked spermatogenesis at pachytene stage, and infertility [8,9]. These reports strongly support the PLD6 related phenotype annotation in the present study. The above analysis and evidences imply that PLD6 might regulate bovine spermatogenesis through piRNA pathway.
Next, we used multi-sequence alignment to further explore the amino acid sequence of bovine PLD6, which has a high similarity with that of mouse PLD6, also indicating similar functional site of PLD6 playing catalytic role. Subsequently, we analyzed the tissue-specificity of PLD6 in cattle and mouse by GEO deep mining, which showed a significant highest expression in bovine testes and mouse testes. In testes, the specific expression pattern of SSC molecular markers can reflect the differentiation state of germ cells [50]. Studies demonstrated that the seminiferous epithelium at early prepubertal stages in bovine consists of gonocytes (prospermatogonia), SSCs and immature Sertoli cells [16,46]. Therefore, there are relatively more germline stem cells in early prepubertal testis and they can be marked by VASA, PIWI12, UCHL1, PLZF, GFRα-1[16,17,51,52], etc. In mouse, SSCs first appeared at 16.5d (E16.5d) embryonic stage. Cell proliferation experiments found that the number of SSCs increased significantly from E16.5d to 2d after birth, and their proportion in the total number of germ cells also increased [53]. By GEO database mining, the present study found that the transcription level of PLD6 is low in the embryonic stage, and slightly increases at E15.5-E16.5d, remarkably rises on the 14th day after birth, significantly increased and reached at peak on the 18th-20th day after birth. This expression trend and pattern of PLD6 is very similar to those of VASA and PIWI12. The above analysis indicates that PLD6 has testicular tissue specificity, and its expression pattern is consistent with that of germ cell marker genes. PLD6 reflects the differentiation of SSCs, and it could be a potential marker for bull germ cells including SSCs at certain developmental stage.
4. Materials and Methods
4.1. Animal Tissues
The animal tissues were from 6-month-old and 2-year-old healthy Simmental calves/bulls provided by Jilin Changchun Haoyue Islamic Meat Co., Ltd (Changchun, China). The testicular tissue in each group was from three animals, and six testicular samples were collected from bilateral testicles. Briefly, the testicles were immediately transported to the laboratory from the slaughter house using an insulated container having ice, soaked in 75% alcohol for 10 min, then washed twice with phosphate buffer saline (PBS) containing 1% Penicillin-Streptomycin (Gibco, CA, USA). The epididymis and tunica albuginea were peeled off aseptically, the testes were cut into 1 cm3 pieces and put into freezing vials containing 1 ml freezing medium as reported previously [20]. Subsequently, the samples were stored in liquid nitrogen for later use. The study was permitted by the Jilin University Institutional Animal Care and Use Committee for use of animals/tissues (SY201903002).
4.2. RT-PCR and qRT-PCR
The total RNAs were extracted from above frozen-thawed bovine testicular tissues according to the instructions of the Animal RNA Extraction Kit (Beyotime, Shanghai, China), and the cDNAs were obtained by using RNA Reverse Transcription Kit (TransGen Biotech, Beijing, China). The primers (Table 1) were designed according to the gene sequences in the NCBI database and synthesized by Sangon Biotech (Shanghai, China). Reversed transcription-polymerase chain reaction (RT-PCR) and quantitative real-time PCR (qRT-PCR) reaction systems of 20 μl were prepared and the reactions were carried out according to the manufacturer’s instructions (TransGen Biotech, Beijing, China). The experiments were repeated three times independently, and the relative gene expressions were calculated according to the formula 2−△△CT. Data were expressed as mean ± SEM.
4.3. Immunostaining
Paraffin sections (4 μm-thick) were prepared from the frozen-thawed testicular tissues. Briefly, the tissues were fixed with 4% paraformaldehyde in PBS for 24 h. Gradual dehydration was achieved by immersing in 75% ethanol for 4 h, 85% ethanol for 2 h, 90% ethanol for 2 h, 95% ethanol for 1 h, 100% ethanol two times for 30 min each. After incubation with xylene (2×10 min), the tissues were immersed in a 1:1 mixture of paraffin: xylene for 30 min, in 100% paraffin twice (each for 1 h), at 56-60°C. Subsequently the tissue samples were solidified and sliced. The sections were deparaffinized, rehydrated, and further subjected to ethylene diamine tetraacetic acid (EDTA) antigen retrieval buffer (pH 8.0). Then the sections were blocked with 3% bovine serum albumin (w/v) for 30 min. Subsequently, they were incubated overnight at 4°C with the following primary antibodies, mouse anti-UCHL1 (1:2000; Boster, Wuhan, China), rabbit anti-VASA (1:200; GeneTex, Shanghai, China) and rabbit anti-PLD6 (1:200; Abcam, Cambridge, UK). Next day, the sections were incubated with Alexa Fluor 594-conjugated goat anti-rabbit/mouse IgG (1:500; Proteintech, Rosemont, USA) for 60 min. The sections were again incubated in EDTA antigen retrieval buffer and heated in a microwave oven to remove the primary and secondary antibodies that had bind to the tissues. To visualize the nuclei, the sections were counterstained with 4’, 6-diamidino-2-phenylindole (DAPI, Beyotim Biotechnology, Shanghai, China) and incubated at room temperature for 15 min. Finally, the sections were covered with anti-quenching tablets and observed under a Nikon 80i fluorescence microscope. The sections stained with isotype IgG served as the negative control.
4.4. Bioinformatics
4.4.1. Phylogenetic Analysis
Evolutionary Trace Server (ETS, http://mordred.bioc.cam.ac.uk/~jiye/evoltrace/ evoltrace.html) was used for PLD6 phylogenetics. Single Likelihood Ancestor Counting (SLAC) algorithm module of Datamonkey Adaptive Evolution Server (http://www.datamonkey.org/) was employed to analyze the evolutionary pressure of its coding genes. The non-synonymous rate (Ka), synonymous rate (Ks), and Ka/Ks ratio were calculated as previously described [21]. The Ka/Ks > 1 indicates the target gene is subject to positive selection, Ka/Ks = 1 represents neutral evolution, while Ka/Ks < 1 suggests the gene is subject to purify selection. Thus Ka/Ks ratio indicated the choice of gene locus. Additionally, the software ClustalX 1.83 [22] and MEGA7.0 [23] as well as iTOL (https://itol.embl.de/) were used to construct and visualize the phylogenetic tree.
4.4.2. Protein Structure Analysis
The Predict Protein (http://www.predictprotein.org/) was used to analyze the secondary structure of PLD protein. The SWISS-MODEL (http://swissmodel.expasy.org/) and Consurf Web Server (http://consurf.tau.ac.il/) were employed to perform the homology modeling and conservative analysis [24]. The conservation and solvent accessibility of the amino acids were mapped to the protein structure. The relationship between the solvent accessibility of PLD6 amino acid residues and its structure as well as function was predicted based on the neural convolutional networks. The key amino acids were further annotated with visualization software PyMOL. The PSORT Prediction (http://psort1.hgc.jp/form.html) was used to predict the subcellular localization of PLD6, and to analyze its related biological behaviors as molecular markers [25]. The transmembrane region prediction was performed with TMHMM 2.0 (https://services.healthtech.dtu.dk/services/TMHMM-2.0/).
4.4.3. Molecular Docking
Utilizing PLD6 as the receptor protein, the three-dimensional (3D) structure of the bovine-derived PLD6 was obtained by performing homology modeling with SWISS-MODEL Server (https://swissmodel.expasy.org/). The structure file of cardiolipin (CL) was downloaded from the PubChem (https://pubchem.ncbi.nlm.nih.gov/) database, energy minimization based on the MMFF94 force field was performed and structure files were generated that could be used for docking. Subsequently, the AutoDock Tools 1.5.6 program was used to remove redundant solvent molecules, balance the charge and add hydrogen atoms for the receptor protein PLD6. At the same time, the center coordinates of the docking and the Grid box were set. Semi-flexible molecular docking of CL molecule to PLD6 was performed with the help of AutoDock Vina program [26]. The final results that consider scoring and conformational fit were selected and visualized using the PyMOL v2.4.0 program [27].
4.4.4. Molecular Dynamics Simulation
The all-atom molecular dynamics simulation program GROMACS 2022.03 [28] was used to perform all-atom molecular dynamics simulations of the docked complex of PLD6 and CL as the initial conformation, with a view to analyzing the mechanism of action present and verifying the reliability of the binding mode. Simulations of the complexes were carried out based on the Amber 99SB-ILDN force field, using the Antechamber [29] and Acpype [30] programs to generate topology files of the ligand molecules. A TIP3P water model and a dodecahedral solvation box were chosen and the system boundary was set at a closest distance to the protein of 1.5 nm. One Na+ was randomly added to neutralise the system charge, and the energy of the system was minimized using a maximum of 50,000 steps of the fastest descent algorithm. The system was then ramped up to 310 K in a 500 ps simulation under a regular ensemble (NVT) and continued for 500 ps under an isothermal isobaric ensemble (NPT). After the system has reached equilibrium, the bond lengths and bonding interactions were constrained by the LINCS algorithm, with step intervals set to 2 fs and 200 ns of unconstrained dynamic simulations. The trajectory of the calibration periodicity was analyzed by the GROMACS tools, in which the root mean square deviation (RMSD) indicates the degree of variation in molecular structure to measure the stability of the complex system; the root mean square fluctuation (RMSF) shows the structural flexibility of the protein and the fluctuation of residues; the radius of gyration (Rg) reflects the tightness of the protein folding during the simulation; The clustering results of the lowest Gibbs free energy were presented by free energy morphology and the lowest energy conformation was captured to analyze the binding mode of CL molecule to PLD6 protein.
4.4.5. Protein-Protein Interaction (PPI) Network
The PPI network involving PLD6 was analyzed and visualized by the STRING (https://string-db.org) and Metascape databases. The STRING default medium confidence score ≥0.4, high confidence score ≥0.7, and highest confidence score ≥0.9 were used to create PPI network. Notably, the mean confidence score of the proteins interacting with PLD6 was 0.736, which reached the high confidence level of the STRING database, indicating that the PLD6-based PPI network is able to enrich the truly relevant proteins. The Max number of interactors (both the 1st and 2nd shells) was set ≤ 50, and the Markov Clustering algorithm (MCL) was used to cluster the proteins with close functions, so as to obtain the key proteins and sub-networks in the network. Subsequently, the Cytoscape plug-in Cytohubba was used to screen the proteins with the degree of connectivity at the top 10 in the protein network as Hub proteins (Highly connected proteins).
4.4.6. GO and KEGG Analysis
The DAVID (https://david.ncifcrf.gov/), Uniprot (https://www.uniprot.org/), Metascape (https://metascape.org/), and GSEA (Gene Set Enrichment Analysis) databases were used to search and enrich the function as well as biological significance of PLD6 at the Gene Ontology (GO) level. The pathways involving PLD6 were associated and analyzed through the Kyoto Encyclopedia of Genes and Genomes (KEGG, https://www.kegg.jp/) database and TBtools program.
4.4.7. Multiple Sequence Alignment and GEO Database Mining
The sequence files were obtained from the NCBI and UniProt databases, and aligned using the Clustal Omega program. The alignment results were imported into the ESPript3 Server (https://espript.ibcp.fr/ESPript/cgi-bin/ESPript.cgi) and annotated. The transcriptomics sequencing (RNA-Seq) data of testicular tissues in GEO database (https://www.ncbi.nlm.nih.gov/geo/) were searched. The GEO2R and R language were used to process the sequencing data. Taking |logFC| > 1.5 and P < 0.05 as the significant standard, the differentially expressed genes (DEGs) in each data set were screened. Finally, the expression patterns of PLD6 in testicular tissues of cattle and mouse in the RNA-Seq data were analyzed by using the Expression Atlas online platform (https://www.ebi.ac.uk/gxa/home) in EMBL-EBI database (https://elixir-europe.org/).
4.5. Statistical Analysis
Statistical significance was compared among the groups. SPSS 22.0 and GraphPad Prism 8.3.0 software were used for data statistics and graphing. T test was used for significance analysis of univariate experiment. *P < 0.05, **P < 0.01 and ***P < 0.001 indicate there is a statistical difference, or the difference is extremely significant.
5. Conclusions
In conclusion, we verified the expression of PLD6/PLD6 in bovine testis, analyzed its molecular features and predicted its possible functions in spermatogenesis by multiple bioinformatics. On the one hand, PLD6 might combine with cardiolipin as a phospholipase to induce mitochondrial fusion; on the other hand, PLD6 might participate in the processing of primary piRNA transcripts as an endonuclease and have a significant relationship with piRNA binding protein. We have found that, overall,PLD6 has a relatively high expression in bovine testes and could be used as a biomarker for spermatogenic cells including SSCs. Nevertheless, more and direct methods such as RNA interference and gene editing might be used to further elucidate the function of bovine PLD6 in future.
Author Contributions
Conceptualization, R.Y. and B.-Y.Z.; methodology, R.Y. and B.-Y.Z; software, R.Y.; validation, W.-Q.Z., C.-L.Z. and L.-X.C.; formal analysis, Y.-S.Z.; investigation, Y.-Q.W. and Y.Z.; resources, C.-L.Z. and L.-X.C.; data curation, B.T.; writing—original draft preparation, R.Y. and B.-Y.Z; writing—review and editing, R.Y., A.R. and X.-M.Z.; visualization, X.-M.Z.; supervision, B.T.; project administration, X.-M.Z.; funding acquisition, X.-M.Z.; All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Natural Science Foundation of China, grant number 32172803 and 31872434.
Institutional Review Board Statement
The animal study protocol was approved by the Jilin University Institutional Animal Care and Use Committee for use of animals/tissues (SY201903002).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cazzolli, R.; Shemon, A.N.; Fang, M.Q.; Hughes, W.E. Phospholipid signalling through phospholipase D and phosphatidic acid. Iubmb Life 2006, 58, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Li, F.; Zhang, L.; Lei, P.; Zheng, Y.; Zeng, W. Stage-specific embryonic antigen 4 is a membrane marker for enrichment of porcine spermatogonial stem cells. Andrology 2020, 8, 1923–1934. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Chi, X.; Wang, Y.; Heng, B.C.; Wei, Y.; Zhang, X.; Zhao, H.; Yin, Y.; Deng, X. Mitochondria transfer enhances proliferation, migration, and osteogenic differentiation of bone marrow mesenchymal stem cell and promotes bone defect healing. Stem Cell Res. Ther. 2020, 11, 245. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liang, P.; Huang, Y.; Li, M.; Zhang, X.; Ding, C.; Feng, J.; Zhang, Z.; Zhang, X.; Gao, Y. Glycerol kinase-like proteins cooperate with Pld6 in regulating sperm mitochondrial sheath formation and male fertility. Cell Discov. 2017, 3, 17030. [Google Scholar] [CrossRef]
- Ipsaro, J.J.; Haase, A.D.; Knott, S.R.; Joshua-Tor, L.; Hannon, G.J. The structural biochemistry of Zucchini implicates it as a nuclease in piRNA biogenesis. Nature 2012, 491, 279–283. [Google Scholar] [CrossRef]
- Kabayama, Y.; Toh, H.; Katanaya, A.; Sakurai, T.; Chuma, S.; Kuramochi-Miyagawa, S.; Saga, Y.; Nakano, T.; Sasaki, H. Roles of MIWI, MILI and PLD6 in small RNA regulation in mouse growing oocytes. Nucleic Acids Res. 2017, 45, 5387–5398. [Google Scholar] [CrossRef]
- Nishimasu, H.; Ishizu, H.; Saito, K.; Fukuhara, S.; Kamatani, M.K.; Bonnefond, L.; Matsumoto, N.; Nishizawa, T.; Nakanaga, K.; Aoki, J. Structure and function of Zucchini endoribonuclease in piRNA biogenesis. Nature 2012, 491, 284–287. [Google Scholar] [CrossRef]
- Huang, H.; Gao, Q.; Peng, X.; Choi, S.Y.; Sarma, K.; Ren, H.; Morris, A.J.; Frohman, M.A. piRNA-associated germline nuage formation and spermatogenesis require MitoPLD profusogenic mitochondrial-surface lipid signaling. Dev. Cell 2011, 20, 376–387. [Google Scholar] [CrossRef]
- Watanabe, T.; Chuma, S.; Yamamoto, Y.; Kuramochi-Miyagawa, S.; Totoki, Y.; Toyoda, A.; Hoki, Y.; Fujiyama, A.; Shibata, T.; Sado, T. MITOPLD is a mitochondrial protein essential for nuage formation and piRNA biogenesis in the mouse germline. Dev. Cell 2011, 20, 364–375. [Google Scholar] [CrossRef]
- Subash, S.K.; Kumar, P.G. Self-renewal and differentiation of spermatogonial stem cells. Front. Biosci.-Landmrk. 2021, 26, 163–205. [Google Scholar] [CrossRef]
- Zhang, P.; Qin, Y.; Zheng, Y.; Zeng, W. Phospholipase D Family Member 6 Is a Surface Marker for Enrichment of Undifferentiated Spermatogonia in Prepubertal Boars. Stem Cells Dev. 2018, 27, 55–64. [Google Scholar] [CrossRef]
- Law, N.C.; Oatley, M.J.; Oatley, J.M. Developmental kinetics and transcriptome dynamics of stem cell specification in the spermatogenic lineage. Nat. Commun. 2019, 10, 2787. [Google Scholar] [CrossRef] [PubMed]
- de Rooij, D.G.; Russell, L.D. All you wanted to know about spermatogonia but were afraid to ask. J. Androl. 2000, 21, 776–798. [Google Scholar] [CrossRef]
- Kanatsu-Shinohara, M.; Chen, G.; Morimoto, H.; Shinohara, T. CD2 is a surface marker for mouse and rat spermatogonial stem cells. J. Reprod. Dev. 2020, 66, 341–349. [Google Scholar] [CrossRef]
- Mutoji, K.; Singh, A.; Nguyen, T.; Gildersleeve, H.; Kaucher, A.V.; Oatley, M.J.; Oatley, J.M.; Velte, E.K.; Geyer, C.B.; Cheng, K. TSPAN8 Expression Distinguishes Spermatogonial Stem Cells in the Prepubertal Mouse Testis. Biol. Reprod. 2016, 95, 117. [Google Scholar] [CrossRef]
- Cai, H.; Jiang, Y.; Zhang, S.; Cai, N.N.; Zhu, W.Q.; Yang, R.; Tang, B.; Li, Z.Y.; Zhang, X.M. Culture bovine prospermatogonia with 2i medium. Andrologia 2021, 53, e14056. [Google Scholar] [CrossRef]
- Zhao, H.; Nie, J.; Zhu, X.; Lu, Y.; Liang, X.; Xu, H.; Yang, X.; Zhang, Y.; Lu, K.; Lu, S. In vitro differentiation of spermatogonial stem cells using testicular cells from Guangxi Bama mini-pig. J. Vet. Sci. 2018, 19, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Guo, Y.; Zheng, B.; Shao, B.; Jiang, M.; Wang, G.; Zhou, T.; Wang, L.; Zhou, Z.; Guo, X. Establishment of a proteome profile and identification of molecular markers for mouse spermatogonial stem cells. J. Cell. Mol. Med. 2015, 19, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Qin, Y.; Zheng, Y.; Zeng, W. Phospholipase D Family Member 6 Is a Surface Marker for Enrichment of Undifferentiated Spermatogonia in Prepubertal Boars. Stem Cells Dev. 2018, 27, 55–64. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhu, W.Q.; Zhu, X.C.; Cai, N.N.; Yang, R.; Cai, H.; Zhang, X.M. Cryopreservation of calf testicular tissues with knockout serum replacement. Cryobiology 2020, 92, 255–257. [Google Scholar] [CrossRef]
- Geng, N.; Sun, G.; Liu, W.J.; Gao, B.C.; Sun, C.; Xu, C.; Hua, E.; Xu, L. Distribution, Phylogeny and Evolution of Clinical and Environmental Vibrio vulnificus Antibiotic-Resistant Genes. Evol. Bioinform. Online 2022, 18, 1041271744. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple sequence alignment using ClustalW and ClustalX. Curr. Protoc. Bioinform. 2002, Chapter 2, 2–3. [Google Scholar] [CrossRef]
- Hall, B.G. Building phylogenetic trees from molecular data with MEGA. Mol. Biol. Evol. 2013, 30, 1229–1235. [Google Scholar] [CrossRef] [PubMed]
- van der Weg, K.J.; Gohlke, H. TopEnzyme: A framework and database for structural coverage of the functional enzyme space. Bioinformatics 2023, 39, btad116. [Google Scholar] [CrossRef]
- Chen, Y.; Yu, P.; Luo, J.; Jiang, Y. Secreted protein prediction system combining CJ-SPHMM, TMHMM, and PSORT. Mamm. Genome 2003, 14, 859–865. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Rigsby, R.E.; Parker, A.B. Using the PyMOL application to reinforce visual understanding of protein structure. Biochem. Mol. Biol. Educ. 2016, 44, 433–437. [Google Scholar] [CrossRef] [PubMed]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E.; Oak Ridge National Lab. ORNL, O.R.T.U.; Teoretisk, F.; Science For Life Laboratory, S. et al. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef]
- Wang, J.; Wolf, R.M.; Caldwell, J.W.; Kollman, P.A.; Case, D.A. Development and testing of a general amber force field. J. Comput. Chem. 2004, 25, 1157–1174. [Google Scholar] [CrossRef]
- Sousa, D.S.A.; Vranken, W.F. ACPYPE—AnteChamber PYthon Parser interfacE. BMC Res. Notes 2012, 5, 367. [Google Scholar] [CrossRef]
- Brangwynne, C.P.; Eckmann, C.R.; Courson, D.S.; Rybarska, A.; Hoege, C.; Gharakhani, J.; Julicher, F.; Hyman, A.A. Germline P granules are liquid droplets that localize by controlled dissolution/condensation. Science 2009, 324, 1729–1732. [Google Scholar] [CrossRef] [PubMed]
- Lehtiniemi, T.; Kotaja, N. Germ granule-mediated RNA regulation in male germ cells. Reproduction 2018, 155, R77–R91. [Google Scholar] [CrossRef]
- Zhang, G.W.; Wang, L.; Chen, H.; Guan, J.; Wu, Y.; Zhao, J.; Luo, Z.; Huang, W.; Zuo, F. Promoter hypermethylation of PIWI/piRNA pathway genes associated with diminished pachytene piRNA production in bovine hybrid male sterility. Epigenetics-US 2020, 15, 914–931. [Google Scholar] [CrossRef]
- Ipsaro, J.J.; Haase, A.D.; Knott, S.R.; Joshua-Tor, L.; Hannon, G.J. The structural biochemistry of Zucchini implicates it as a nuclease in piRNA biogenesis. Nature 2012, 491, 279–283. [Google Scholar] [CrossRef]
- Nakata, H. Morphology of mouse seminiferous tubules. Anat. Sci. Int. 2019, 94, 1–10. [Google Scholar] [CrossRef]
- Kulandaisamy, A.; Lathi, V.; ViswaPoorani, K.; Yugandhar, K.; Gromiha, M.M. Important amino acid residues involved in folding and binding of protein-protein complexes. Int. J. Biol. Macromol. 2017, 94, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Nishimasu, H.; Ishizu, H.; Saito, K.; Fukuhara, S.; Kamatani, M.K.; Bonnefond, L.; Matsumoto, N.; Nishizawa, T.; Nakanaga, K.; Aoki, J. Structure and function of Zucchini endoribonuclease in piRNA biogenesis. Nature 2012, 491, 284–287. [Google Scholar] [CrossRef]
- Ikeda, S.; Tanaka, K.; Ohtani, R.; Kanda, A.; Sotomaru, Y.; Kono, T.; Obata, Y. Disruption of piRNA machinery by deletion of ASZ1/GASZ results in the expression of aberrant chimeric transcripts in gonocytes. J. Reprod. Dev. 2022, 68, 125–136. [Google Scholar] [CrossRef]
- Hutt, K.J.; Lim, S.L.; Zhang, Q.H.; Gonzalez, M.; O’Connor, A.E.; Merriner, D.J.; Liew, S.H.; Al-Zubaidi, U.; Yuen, W.S.; Adhikari, D. HENMT1 is involved in the maintenance of normal female fertility in the mouse. Mol. Hum. Reprod. 2021, 27, gaab061. [Google Scholar] [CrossRef] [PubMed]
- Hempfling, A.L.; Lim, S.L.; Adelson, D.L.; Evans, J.; O’Connor, A.E.; Qu, Z.P.; Kliesch, S.; Weidner, W.; O’Bryan, M.K.; Bergmann, M. Expression patterns of HENMT1 and PIWIL1 in human testis: Implications for transposon expression. Reproduction 2017, 154, 363–374. [Google Scholar] [CrossRef]
- Ding, D.; Liu, J.; Midic, U.; Wu, Y.; Dong, K.; Melnick, A.; Latham, K.E.; Chen, C. TDRD5 binds piRNA precursors and selectively enhances pachytene piRNA processing in mice. Nat. Commun. 2018, 9, 127. [Google Scholar] [CrossRef]
- Iwasaki, Y.W.; Siomi, M.C.; Siomi, H. PIWI-Interacting RNA: Its Biogenesis and Functions. Annu. Rev. Biochem. 2015, 84, 405–433. [Google Scholar] [CrossRef]
- Philip, F.; Ha, E.E.; Seeliger, M.A.; Frohman, M.A. Measuring Phospholipase D Enzymatic Activity Through Biochemical and Imaging Methods. Methods Enzymol. 2017, 583, 309–325. [Google Scholar] [CrossRef]
- Riew, T.R.; Kim, S.; Jin, X.; Kim, H.L.; Hwang, W.C.; Kang, M.; Yang, E.S.; Lee, M.Y.; Min, D.S. Cellular and subcellular localization of endogenous phospholipase D6 in seminiferous tubules of mouse testes. Cell Tissue Res. 2021, 385, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Ramalho-Santos, J.; Varum, S.; Amaral, S.; Mota, P.C.; Sousa, A.P.; Amaral, A. Mitochondrial functionality in reproduction: From gonads and gametes to embryos and embryonic stem cells. Hum. Reprod. Update 2009, 15, 553–572. [Google Scholar] [CrossRef] [PubMed]
- Ha, E.E.; Frohman, M.A. Regulation of mitochondrial morphology by lipids. Biofactors 2014, 40, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Ozata, D.M.; Gainetdinov, I.; Zoch, A.; O’Carroll, D.; Zamore, P.D. PIWI-interacting RNAs: Small RNAs with big functions. Nat. Rev. Genet. 2019, 20, 89–108. [Google Scholar] [CrossRef]
- Aravin, A.A.; Chan, D.C. piRNAs meet mitochondria. Dev. Cell 2011, 20, 287–288. [Google Scholar] [CrossRef]
- Gao, Q.; Frohman, M.A. Roles for the lipid-signaling enzyme MitoPLD in mitochondrial dynamics, piRNA biogenesis, and spermatogenesis. BMB Rep. 2012, 45, 7–13. [Google Scholar] [CrossRef]
- Helsel, A.R.; Yang, Q.; Oatley, M.J.; Lord, T.; Sablitzky, F.; Oatley, J.M. ID4 levels dictate the stem cell state in mouse spermatogonia. Development 2017, 144, 624–634. [Google Scholar] [CrossRef]
- Cai, H.; Tang, B.; Wu, J.Y.; Zhao, X.X.; Wang, Z.Z.; An, X.L.; Lai, L.X.; Li, Z.Y.; Zhang, X.M. Enrichment and in vitro features of the putative gonocytes from cryopreserved testicular tissue of neonatal bulls. Andrology-US 2016, 4, 1150–1158. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Choi, Y.R.; Kim, B.J.; Jung, S.E.; Kim, S.M.; Jin, J.H.; Yun, M.H.; Kim, S.U.; Kim, Y.H.; Hwang, S. GDNF family receptor alpha 1 is a reliable marker of undifferentiated germ cells in bulls. Theriogenology 2019, 132, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Law, N.C.; Oatley, M.J.; Oatley, J.M. Developmental kinetics and transcriptome dynamics of stem cell specification in the spermatogenic lineage. Nat. Commun. 2019, 10, 2787. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
PLD6 expression in bovine testis: (a) Electrophoresis of RT-PCR products (PLD6, VASA, UCHL1); (b) mRNA relative levels of PLD6, VASA, UCHL1 by qRT-PCR analysis (*P < 0.05, ***P < 0.001); (c) Immunofluorescent staining of PLD6, VASA and UCHL1 (red: PLD6, VASA and UCHL1 positive staining; blue: DAPI counterstained nuclei; purple: merged images). Scale bars = 50 μm.
Figure 1.
PLD6 expression in bovine testis: (a) Electrophoresis of RT-PCR products (PLD6, VASA, UCHL1); (b) mRNA relative levels of PLD6, VASA, UCHL1 by qRT-PCR analysis (*P < 0.05, ***P < 0.001); (c) Immunofluorescent staining of PLD6, VASA and UCHL1 (red: PLD6, VASA and UCHL1 positive staining; blue: DAPI counterstained nuclei; purple: merged images). Scale bars = 50 μm.
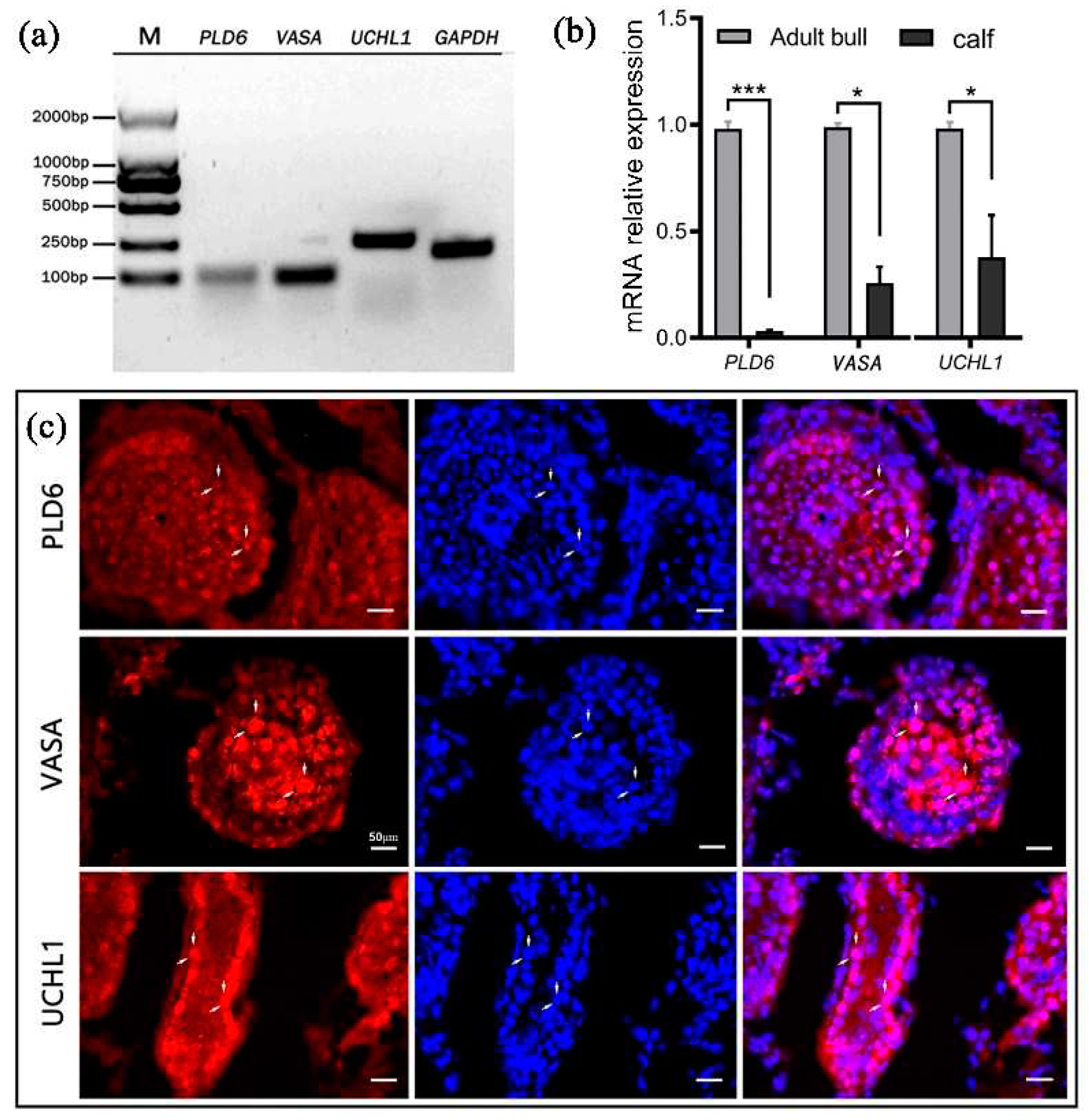
Figure 2.
Phylogenetic and structure analysis of PLD6: (a) Evolutionary Trace Server was used for PLD6 phylogenetics. Single Likelihood Ancestor Counting algorithm module of Datamonkey Adaptive Evolution Server was employed to analyze the evolutionary pressure. ClustalX 1.83, MEGA7.0 and iTOL were used to construct and visualize the phylogenetic tree. (Bar: Nucleotide divergence, numbers at the nodes indicate the bootstrap values); (b) Protein structure analysis of PLD6 including homology modeling and conservative analysis by SWISS-MODEL & convolutional neural network of Consurf Web Server (left), and structure prediction by the Predict Protein (right, showing two opposite sides); e: an exposed amino acid residue; b: a buried residue with low solvent accessibility; f: a highly conserved and exposed residue; s: a highly conserved and buried residue.
Figure 2.
Phylogenetic and structure analysis of PLD6: (a) Evolutionary Trace Server was used for PLD6 phylogenetics. Single Likelihood Ancestor Counting algorithm module of Datamonkey Adaptive Evolution Server was employed to analyze the evolutionary pressure. ClustalX 1.83, MEGA7.0 and iTOL were used to construct and visualize the phylogenetic tree. (Bar: Nucleotide divergence, numbers at the nodes indicate the bootstrap values); (b) Protein structure analysis of PLD6 including homology modeling and conservative analysis by SWISS-MODEL & convolutional neural network of Consurf Web Server (left), and structure prediction by the Predict Protein (right, showing two opposite sides); e: an exposed amino acid residue; b: a buried residue with low solvent accessibility; f: a highly conserved and exposed residue; s: a highly conserved and buried residue.
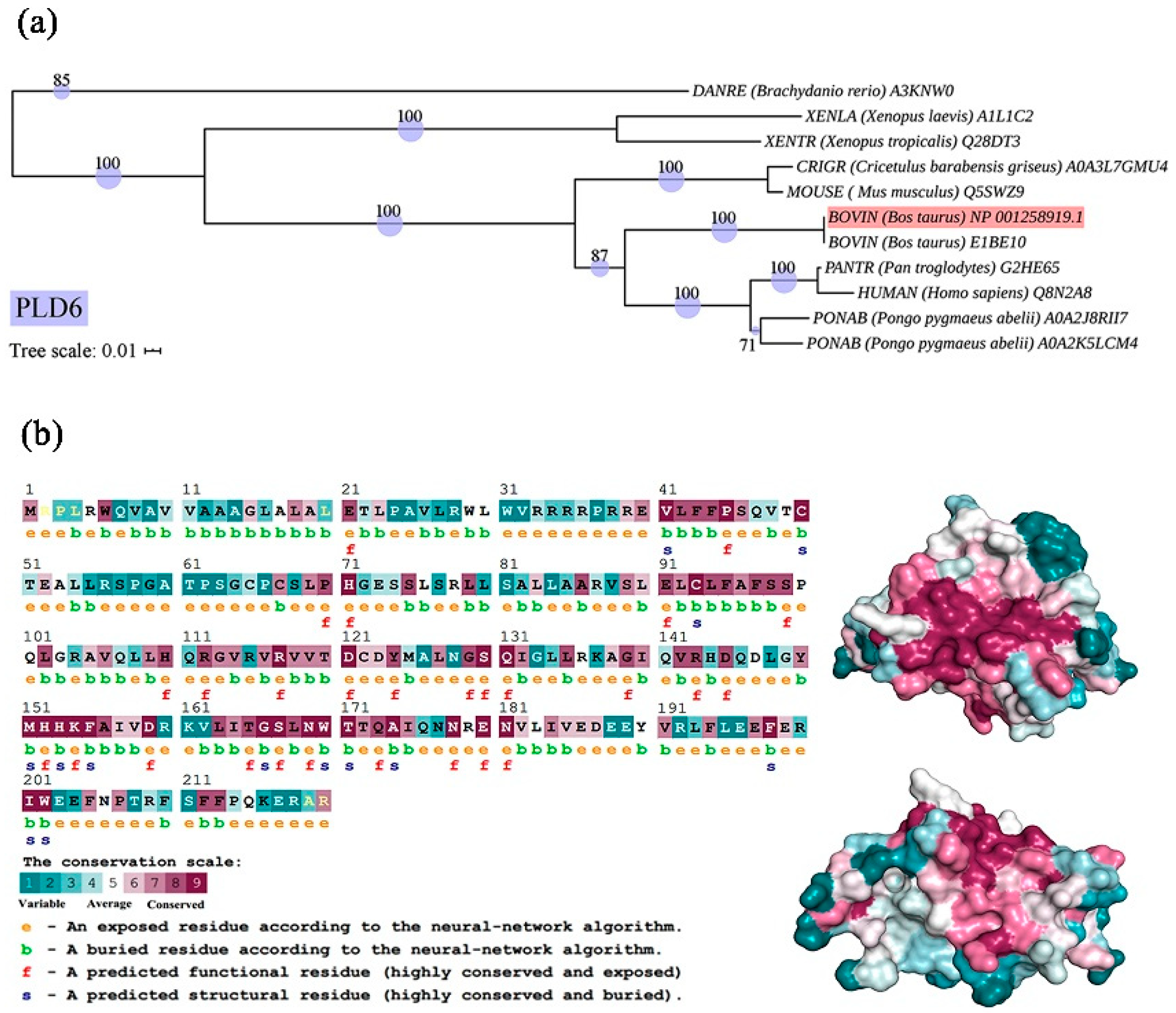
Figure 3.
Molecular dynamics simulation of the PLD6-CL complex: (a) RMSD plot of PLD6-CL complex; (b) Rg plot of PLD6-CL complex; (c) RMSF and structural overlap plot of PLD6; (d) PLD6-CL complex free energy landscape (FEL); (e) Binding conformation of CL to residues inside the phosphodiesterase domain of PLD6; (f) Allosteric diagram of the secondary structure of PLD6; (g) Conformational differences of PLD6 at 0-200 ns.
Figure 3.
Molecular dynamics simulation of the PLD6-CL complex: (a) RMSD plot of PLD6-CL complex; (b) Rg plot of PLD6-CL complex; (c) RMSF and structural overlap plot of PLD6; (d) PLD6-CL complex free energy landscape (FEL); (e) Binding conformation of CL to residues inside the phosphodiesterase domain of PLD6; (f) Allosteric diagram of the secondary structure of PLD6; (g) Conformational differences of PLD6 at 0-200 ns.
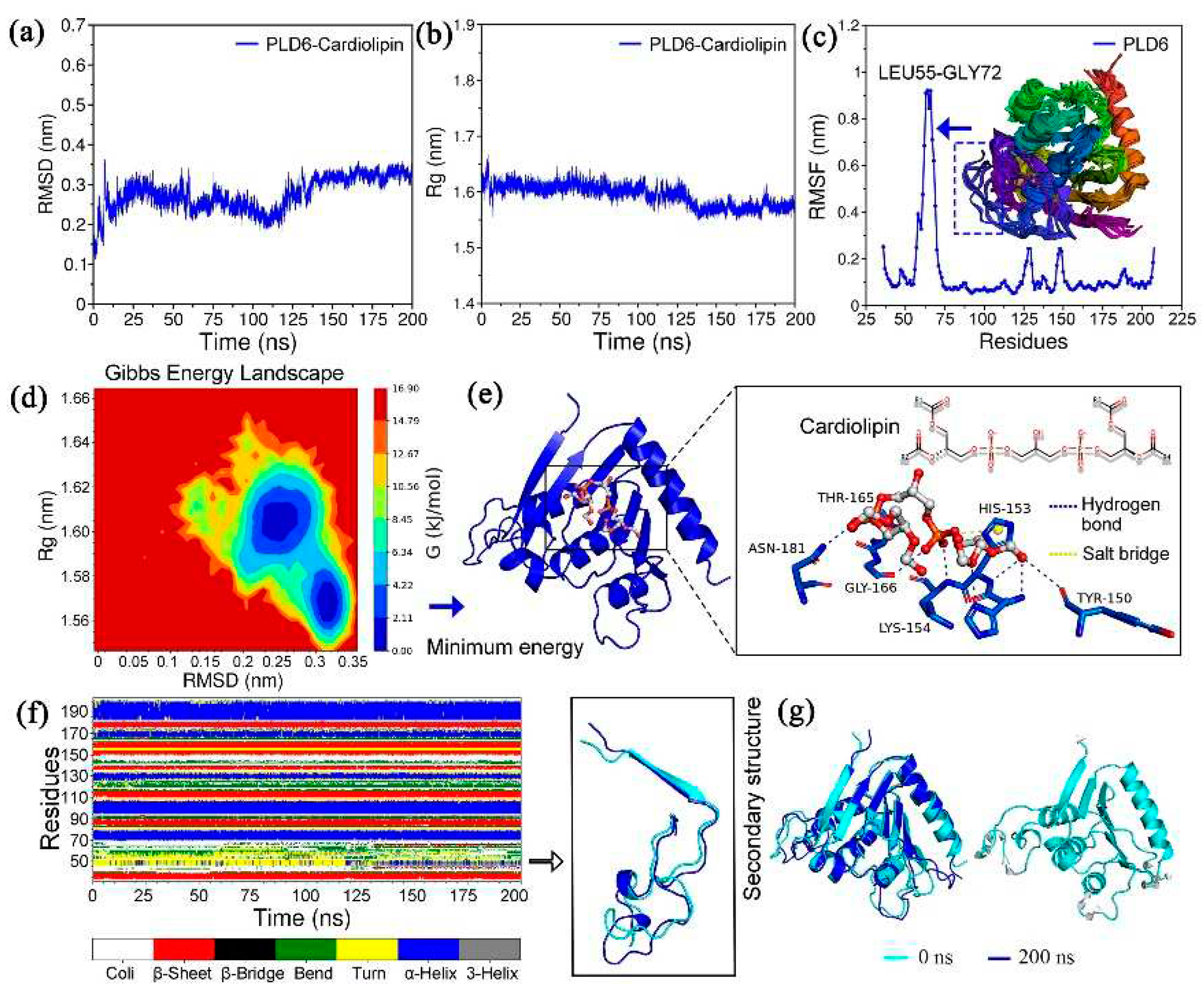
Figure 4.
Protein-protein interaction (PPI) network: (a) Analysis of PLD6 protein interaction network. Protein interaction network analysis is enriched to 60 protein, which are divided into 5 main sub-networks by algorithm clustering; (b) The top 10 Hub protein involving PLD6 were screened out, namely PIWIL4, TDRD9, MEAL, ASZ1, VASA (DDX4), GK2, MGLL, TDRD5, TDRD6 and HENMT1.
Figure 4.
Protein-protein interaction (PPI) network: (a) Analysis of PLD6 protein interaction network. Protein interaction network analysis is enriched to 60 protein, which are divided into 5 main sub-networks by algorithm clustering; (b) The top 10 Hub protein involving PLD6 were screened out, namely PIWIL4, TDRD9, MEAL, ASZ1, VASA (DDX4), GK2, MGLL, TDRD5, TDRD6 and HENMT1.
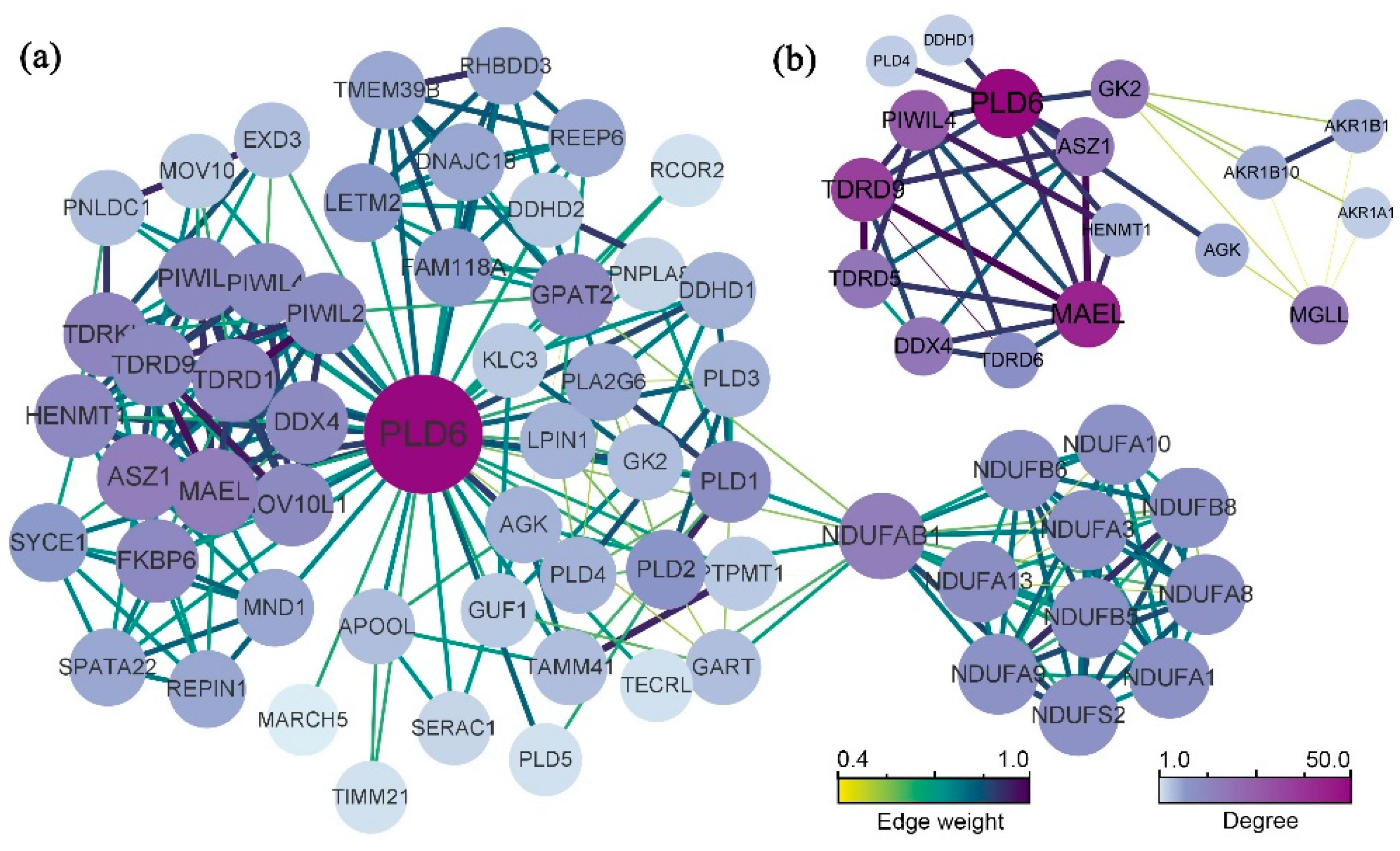
Figure 5.
GO enrichment analysis of PLD6. The biological function of PLD6 is annotated from the three aspects of biological process (BP), molecular function (MF) and cell composition (CC). The greater the strength value, the higher the participation of PLD6 protein in this part.
Figure 5.
GO enrichment analysis of PLD6. The biological function of PLD6 is annotated from the three aspects of biological process (BP), molecular function (MF) and cell composition (CC). The greater the strength value, the higher the participation of PLD6 protein in this part.
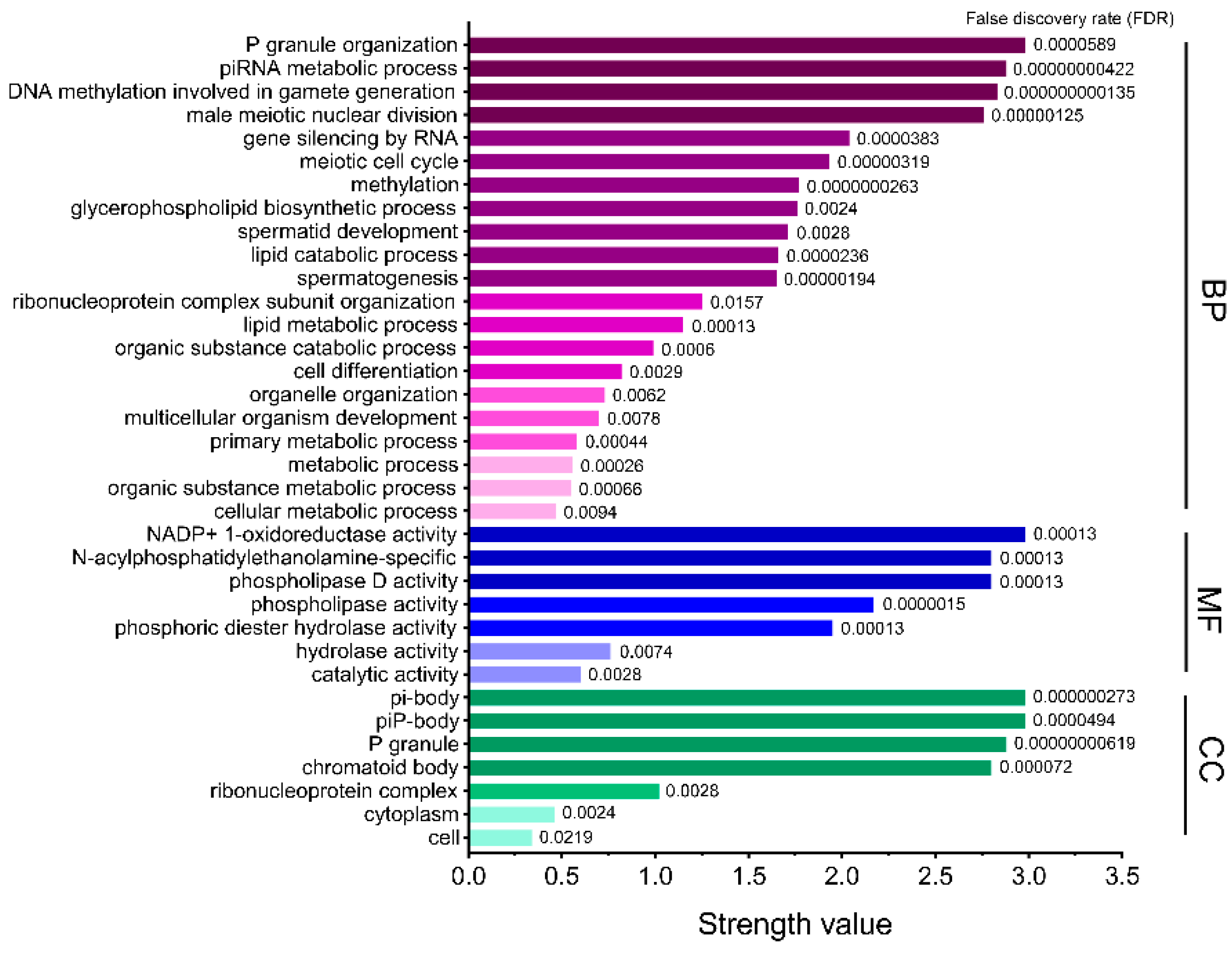
Figure 6.
Dynamic analysis of PLD6/PLD6 expression level: (a) Bovine and murine amino acid multiple sequence alignment of PLD6 protein; (b) RNA-Seq-Expression of PLD6 in bovine and mouse organs including brain, colon, heart, kidney, liver, lung, muscle, spleen and testis, in which testis tissue is the most prominent; (c) Expression of PLD6, VASA, PIWIL2 and GFRα1 in mouse testes at 3, 6, 8, 10, 14, 18, 20, 30, 35 and 56 d. The upwards of the 0-tick mark means that the gene transcription level is up-regulated, and the downwards of the 0 scale means that the gene transcription level is down-regulated. The heat map indicates the transcription level of PLD6 at different time points.
Figure 6.
Dynamic analysis of PLD6/PLD6 expression level: (a) Bovine and murine amino acid multiple sequence alignment of PLD6 protein; (b) RNA-Seq-Expression of PLD6 in bovine and mouse organs including brain, colon, heart, kidney, liver, lung, muscle, spleen and testis, in which testis tissue is the most prominent; (c) Expression of PLD6, VASA, PIWIL2 and GFRα1 in mouse testes at 3, 6, 8, 10, 14, 18, 20, 30, 35 and 56 d. The upwards of the 0-tick mark means that the gene transcription level is up-regulated, and the downwards of the 0 scale means that the gene transcription level is down-regulated. The heat map indicates the transcription level of PLD6 at different time points.
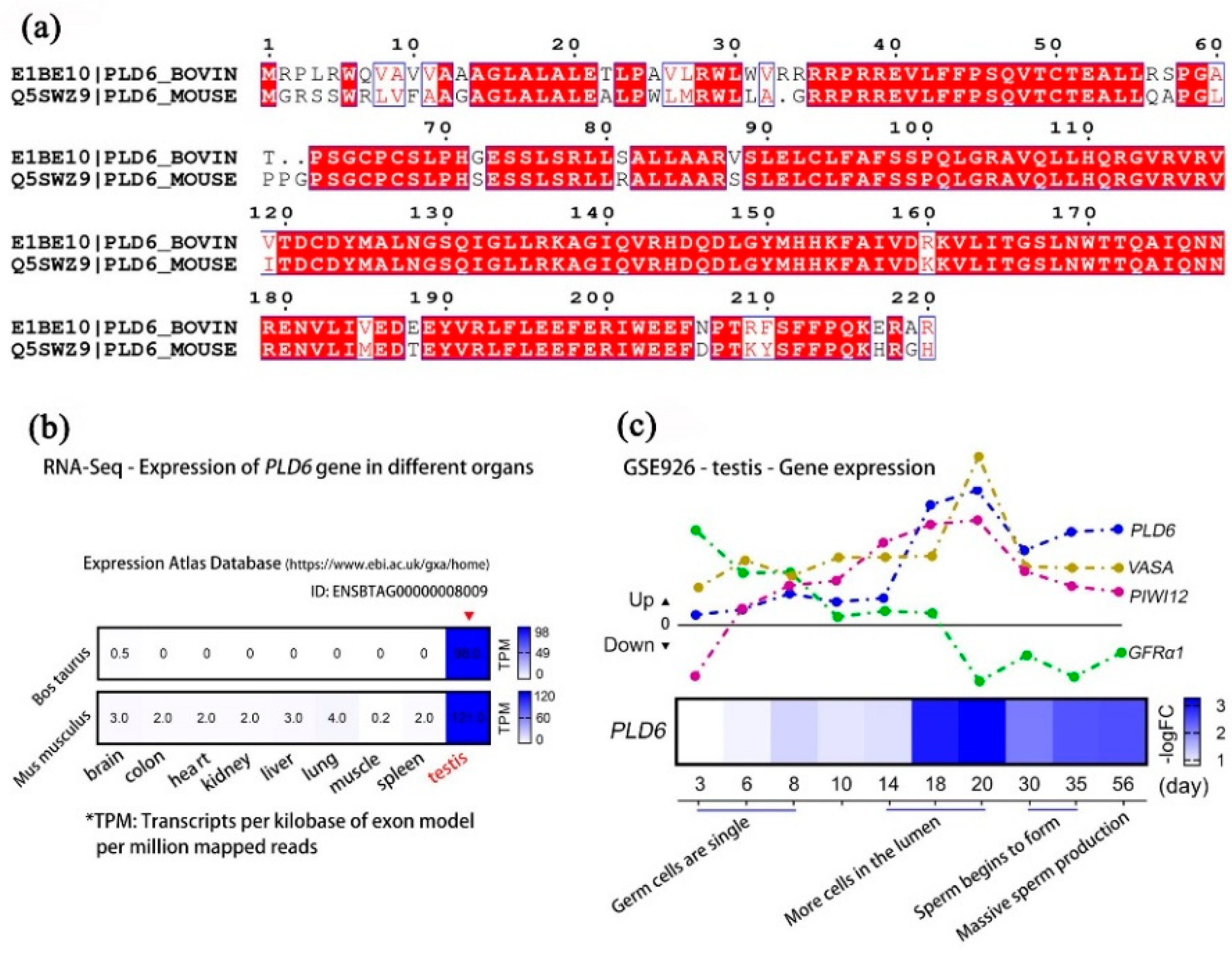
Figure 7.
Phenotypic changes in PLD6tm1.1Hsas and PLD6tm1.1Mafr mice based on the references and MGI database.
Figure 7.
Phenotypic changes in PLD6tm1.1Hsas and PLD6tm1.1Mafr mice based on the references and MGI database.
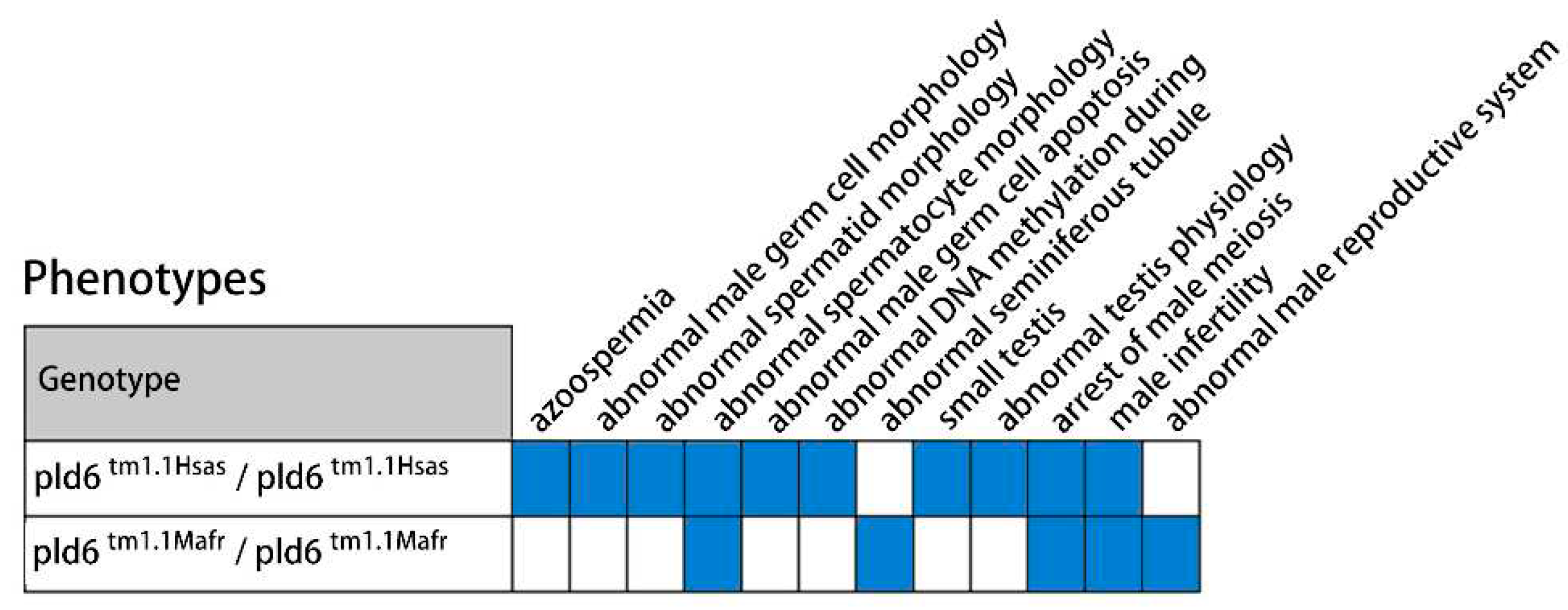
Table 2.
The residues in PLD6 classified by importance and evolutionary pressure.
| Importance Score | Number | Trace Residues |
|---|---|---|
| < 25% | 34 | 44, 45, 69, 71, 72, 83, 86, 93, 95, 97, 106, 109, 112, 113, 114, 116, 121, 151, 152, 154, 155, 159, 166, 167, 169, 170, 171, 174, 178, 180, 181, 194, 198, 202 |
| < 5% | 17 | 86, 93, 95, 112, 121, 152, 154, 159, 166, 167, 169, 170, 174, 178, 180, 198, 202 |
Table 3.
KEGG bioaccumulation analysis of PLD6.
| Potential Biomarker | Proteins Involved(%) | KEGG Pathway ID | Description |
|---|---|---|---|
| PLD6 | 61.08 | map 00564 | Glycerophospholipid metabolism |
| 14.77 | map 00440 | Aminophosphonate metabolism | |
| 9.55 | map 00260 | Glycine, serine and threonine metabolism | |
| 7.03 | map 00565 | Ether lipid metabolism | |
| 7.03 | map 04912 | GnRH signaling pathway | |
| 0.18 | map 00623 | 2,4-Dichlorobenzoate degradation | |
| 0.18 | map 00960 | Alkaloid biosynthesis II | |
| 0.18 | map 00650 | Butanoate metabolism |
Table 4.
Species information in phylogenetic analysis.
| Mnemonic name | Species | Common name | Accession number |
|---|---|---|---|
| DANRE | Danio rerio | Zebrafish | A3KNW0 |
| XENLA | Xenopus laevis | African clawed frog | A1L1C2 |
| XENTR | Xenopus tropicalis | Western clawed frog | Q28DT3 |
| CRIGR | Cricetulus griseus | Chinese hamster | A0A3L7GMU4 |
| MOUSE | Mus musculus | Mouse | Q5SWZ9 |
| BOVINE | Bos taurus | Bovine | NP_001258919.1 |
| BOVINE | Bos taurus | Bovine | E1BE10 |
| PANTR | Pan troglodytes | Chimpanzee | G2HE65 |
| HUMAN | Homo sapiens | Human | Q8N2A8 |
| PONAB | Pongo abelii | Sumatran orangutan | A0A2J8RII7 |
| PONAB | Cercocebus atys | Sooty mangabey | A0A2K5LCM4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



