
Submitted:
30 June 2023
Posted:
30 June 2023
You are already at the latest version
Abstract
Malignant Pleural Mesothelioma (MPM) is an aggressive cancer associated to asbestos exposure. MPM pathogenesis has been related both to oxidative stress, evoked by and in response to asbestos fibers exposure, and the Epithelial Mesenchymal Transition (EMT), an event induced by oxidative stress itself and related to cancer proliferation and metastasis. Asbestos related primary oxidative damage is counteracted in the lung by various redox-sensitive factors, often hyperactivated in some cancers. Among these redox-sensitive factors, Apurinic-apyrimidinic endonuclease 1 (APE-1)/Redox effector factor 1 (Ref-1) has been demonstrated to be overexpressed in MPM and lung cancer, but the molecular mechanism has not yet fully understood. Moreover, asbestos exposure has been associated to induced EMT event, via some EMT transcription factors, such as Twist, Zeb-1 and Snail-1, in a possible crosstalk with oxidative stress and inflammation events. To demonstrate this hypothesis, we inhibited/silenced Ref-1 in MPM cells: as a consequence, both EMT (Twist, Zeb-1 and Snail-1) markers and cellular migration/proliferation were significantly inhibited. Take as a whole, these results show, for the first time, a crosstalk between oxidative stress and EMT in MPM carcinogenesis and invasiveness, so improving the knowledge to better address a preventive and therapeutic approach against this aggressive cancer.
Keywords:
malignant pleural mesothelioma
; oxidative stress
; redox-sensitive factors
; asbestos
; epithelial mesenchymal transition
; proliferation
1. Introduction
Malignant pleural mesothelioma (MPM) is a rare but very aggressive tumor well known for its fatal outcome and in association to asbestos exposure. MPM is a tumor that originates from mesothelial cells lining pleural cavity and is characterized by a long latency period [1]. MPM is histologically distinct in three major subtypes: epithelioid, sarcomatous and biphasic, different from a prognostic point of view [2]. Due to the aggressiveness of this type of tumor, conventional therapies (chemotherapy, radiotherapy and surgical resection) are unsatisfactory [3] and the median survival of these patients is approximately 8-14 months. Moreover, current available predictive and diagnostic markers are few and not effective [1].
The main etiological factor of MPM is the exposure to asbestos: even if the pathogenetic mechanisms involved are not yet fully understood, it has been widely reported that asbestos fibers in mesothelial cells evoke some events that have been associated to MPM development, such as chronic inflammation, accentuated oxidative stress derived from Reactive Oxygen Species (ROS) generation, and the event called Epithelial Mesenchymal Transition (EMT) [4,5]. Particularly, asbestos exposure has been widely demonstrated to primarily induce oxidative stress in the mesothelium, by fibers itself and/or consequently to a pulmonary defensive cellular response: so, MPM onset has been linked to a strong oxidative damage in the microenvironment, event counteracted by antioxidant systems at the pulmonary level, that however could fail and so promote MPM development [6].
As oxidative stress, EMT is an event recently associated to tumorigenesis and metastasis [7]. EMT is a pathophysiological reversible process during which some inducers, such as Transforming Growth Factor β (TGF-β), promote the transition of epithelial cells into mesenchymal cells, via downregulation of some epithelial markers (e.g., E-cadherin) and by increasing the mesenchymal ones (e.g., Fibronectin, Vimentin) [8]. The different expression of these markers is regulated by specific EMT-transcription factors (EMT-TFs), driven by TGF-β signaling, including Twist, Zeb-1 and Snail-1 [8], and thus inducing E-cadherin inhibition. Consequently, cells manifest proliferative and migratory phenotypes, apoptosis resistance and extracellular matrix (ECM) production [9], that together could drive tumoral invasiveness and metastasis.
A consistent body of evidence confirms this correlation between EMT and tumorigenesis in different types of pulmonary tumors, such as Non-Small Cell Lung Cancer (NSCLC) [10,11,12,13,14], and in MPM also in association with asbestos exposure, where asbestos fibers induce EMT in human mesothelial cells via TGF-β signaling [15]. In addition, it was observed in MPM samples that a combination of γ-catenin downregulation and Twist overexpression can be considered a prognostic pattern for patients with MPM [16]. Moreover, also the association between EMT and lung progression was investigated by Liu et al. [17], which demonstrated a synergy between IL-6/JAK/STAT3 and TGF-β/Smad signaling in inducing EMT in lung cancer cell lines.
Thus, oxidative stress and EMT are two events that could interplay in mediating tumorigenesis and/or metastasis. Concerning these events, an EMT-oxidative stress link is particularly evident in different types of tumors: in this regard, it has been shown that ROS production promote EMT through a GSK-3β-mediated mechanism [9] and, moreover, the EMT-oxidative stress link is even more evident considering some proteins, such as heat shock protein 70 (Hsp70), which has a critical role in oxidative stress and EMT in MPM cell lines, via TGF-β pathway [18]. Then, it has been demonstrated that, in human MPM cells, hydrogen peroxide induces overexpression of EMT-related genes [19] and, on the other hand, inhibition of TGF-β signaling and treatment with antioxidants prevent the oxidative stress driven EMT [20].
Our research group has previously demonstrated the overexpression in MPM of some redox-sensitive transcription factors that control the cellular antioxidant defenses [21], such as Nuclear factor erythroid 2-related factor 2 (Nrf2 o NFE2L2)/Kelch-like protein ECH-associated protein 1 (KEAP-1), Apurinic-apyrimidinic endonuclease 1 (APE-1)/Redox effector factor 1 (Ref-1) and Forkhead box protein M1 (FOXM1) [21]. Among these factors, APE-1/Ref-1 (also simply called Ref-1) is a multifunctional enzyme involved in the base excision repair (BER) pathway, crucial for the repair of oxidative and alkylation DNA damage [22], and a reductive activator of transcription factors, such as Activator Protein-1 (AP-1), Hypoxia-Inducible Factor 1 α (HIF-1α), p53 and Nuclear Factor kappa B (NF-kB) [21]. In oxidative stress condition, high levels of ROS elicit Ref-1 overexpression and BER pathway activation, and some tumors present basally Ref-1 overexpressed, thus consequently supporting tumor progression and chemoresistance [23]. As demonstrated, Ref-1 expression affects tumor response to treatment, such as photodynamic therapy [24] or cisplatin-based adjuvant therapy in NSCLC patients [22]. In the latter paper, Ref-1 overexpression is associated with resistance to cisplatin and, on the contrary, the Ref-1 downregulation by siRNA enhances sensitivity of A549 NSCLC cells to chemotherapy [22]. Furthermore, the combination of Ref-1 shRNA knockdown and oxymatrine, decreases A549 cellular proliferation [24], and the treatment of A549 cells with a specific Ref-1 inhibitor, inhibits tumor growth and progression [24]. Finally, different studies demonstrated that Ref-1 silencing improves sensitivity to different DNA damaging agents in various types of tumors [22,25–27].
In this view, until now, no evidence has been correlated to a possible crosstalk between Ref-1 and EMT in MPM. Particularly, Ref-1 could be a crucial point in MPM onset and metastasis, and could be considered a new possible predictive/prognostic marker or a therapeutic target to hit given the failure of the treatments currently available. Take as a whole, this research has been addressed to elucidate this possible crosstalk between oxidative stress and EMT in driving tumorigenesis and metastasis of MPM, by focusing the role of Ref-1 as central point in this crosstalk.
2. Results
2.1. Overexpressed Ref-1 in MPM cells is downregulated after co-incubation with E3330 specific Ref-1 inhibitor.
Primarily, we confirmed previous results obtained by our research group evaluating Ref-1 expression in MSTO-211H mesothelioma cells, compared to human mesothelial cells Met-5A, and NSCLC A549 cells, used as positive control of Ref-1 overexpression in cancer: our results clearly demonstrated significant Ref-1 overexpression in MSTO-211H towards Met-5A cells (Figure 1A-B).
Then, we treated MSTO-211H and A549 cells with different concentrations of E3330 specific Ref-1 inhibitor (as described in Materials & Methods): both MSTO-211H and A549 cell lines showed a significant decreased Ref-1 expression after E3330 treatment, particularly at the higher dose (50 µM) tested (Figure 2A-B).
2.2. Overexpressed Ref-1 in MPM and A549 cells is downregulated after siRNA transfection.
Considering results obtained with E3330 inhibitor, we performed experiments in the attempt to silence Ref-1. We transfected MSTO-211H and A549 cells with Ref-1 siRNA, as described in Materials & Methods. Then, we evaluated whether Ref-1 downregulation occurred: as shown in Figure 3 (A-B), we obtained a significant decrease of Ref-1 expression in both in MSTO-211H and A549 cells, thus confirming the siRNA effective knockdown.
2.3. Overexpressed EMT-TFs in MPM are downregulated after both co-incubation with E3330 specific Ref-1 inhibitor and siRNA Ref-1.
We investigated the possible crosstalk between oxidative stress and EMT. We addressed our experiments focusing on MPM, by evaluating the main three EMT-TFs Twist, Zeb-1, and Snail-1, already shown to be overexpressed in MPM [16,28,29] and in mesothelial cells after asbestos fibers exposure [15]. So, we evaluated these three EMT-TFs in our MPM cellular model both after Ref-1 inhibitor E3330 incubation (at the higher dose 50 µM) and siRNA knockdown: only Twist resulted significantly downregulated after E3330 treatment (Figure 4A-B), while Zeb-1 and Snail-1 were only partially downregulated, but not significantly.
However, our results showed a significant decrease in all three EMT-TFs Twist, Zeb-1 and Snail-1 in MSTO-211H cells after siRNA Ref-1 transfection (Figure 5A-B).
2.4. Effect of Ref-1 inhibition/knockdown on cellular proliferation.
To test the possible effect of this crucial crosstalk on cancer cell migration and invasiveness, mediated by EMT-induced event, we firstly evaluated the cellular motility after Ref-1 silencing, by a scratch assay performed on both MSTO-211H and A549 cell lines, used as positive cellular control of Ref-1 overexpression in cancer. The results shown in Figure 6 revealed that in MSTO-211H and A549 cells the siRef-1 transfection reduced significantly the migration rate by 31% and 81% respectively after 24 hrs (T1) of silencing (Figure 6A-B).
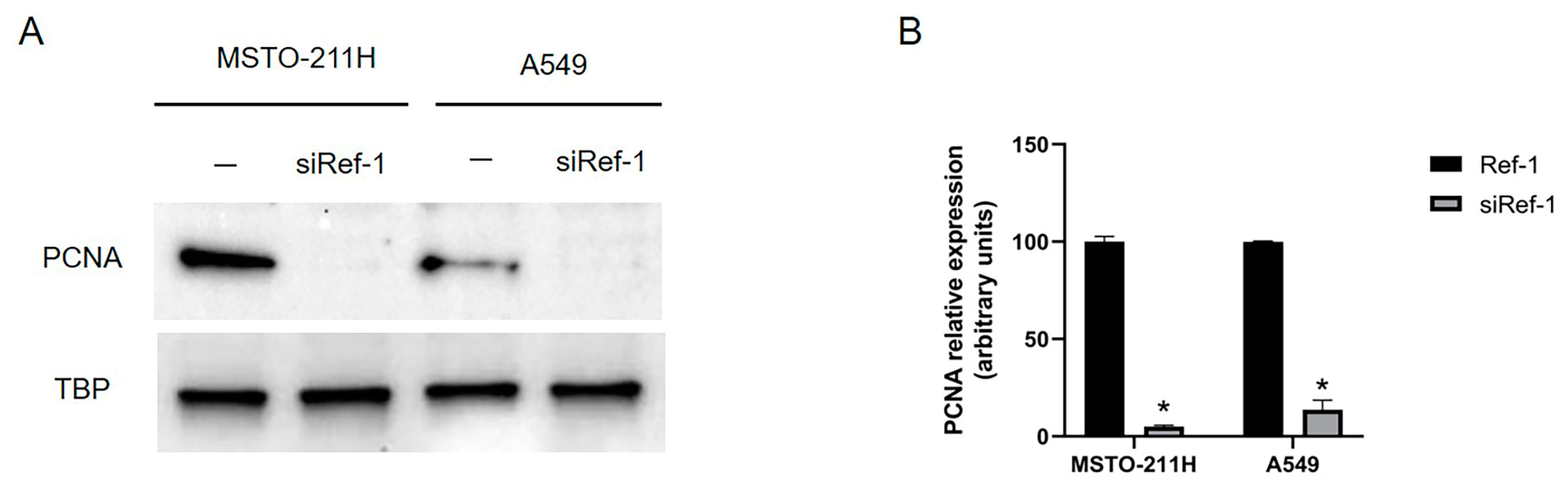
Our evidence was confirmed by investigating, both in MSTO-211H and A549 cells after siRNA Ref-1 transfection, also the cellular proliferation, by evaluating the expression of the Proliferating Cell Nuclear Antigen (PCNA), a protein involved in cell replication processes and a known marker of proliferation. As shown in Figure 7 (A-B), Ref-1 silencing strongly inhibited PCNA expression in both MSTO-211H and A549 cells, thus confirming our results concerning cell migration.
3. Discussion
MPM is a highly aggressive cancer with limited therapeutic possibilities [3]. The development of MPM is closely related to asbestos exposure and the long latency time, as well as its aggressiveness, make increasingly necessary to identify specific predictive markers and new therapeutic targets useful in counteracting this cancer. The pathogenesis of MPM is characterized by numerous and complex molecular events, particularly related to a previous exposure to a strong oxidant as asbestos, and by different deregulated physiological events such as particularly the EMT, a process involved in inflammation, tumorigenesis and invasiveness [30].
Different studies have demonstrated how the primary toxic role of asbestos exposure at level of the mesothelium is crucial, where a consequent inflammatory status, due to cytokines released in response to fibers, was associated to the production of ROS, which in turn promotes an oxidative stress status and stimulates, mainly through the redox-sensitive transcription factor NF-kB, cell survival and the development of a tumor microenvironment [4]. Recently, a lot of paper focused on the crucial role of oxidative stress in cancer: particularly, Cheung and Vousden [31] defined a complex and not well understood molecular system involved in correlating oxidative stress and tumor development and progression, highlighting the role of multiple responses to oxidative stress as a central event in driving tumorigenesis and promote invasiveness.
In this field, our research group demonstrated the overexpression of different redox-sensitive transcription factors (Nrf-2, Ref-1 and FOXM1) in MPM towards not transformed mesothelial cells [21], confirming the hypothesis of the fundamental role of oxidative stress, induced primarily by asbestos exposure, in MPM onset [21]. Among these factors, overexpressed Ref-1 has been recently proposed as a possible marker in different tumors, such as in oral carcinoma [32], or a pharmacological target, such as in pancreatic [33], cutaneous [34], and lung [35] cancers. Particularly in the latter paper, Ref-1 overexpression in NSCLC has been associated to EMT, via a molecular mechanism mediated by TGF-β signaling regulation [35].
Until now, no clearly evidences of a possible crosstalk between oxidative stress and EMT has been demonstrated in MPM, especially considering the central role of asbestos in mediating both events: in mesothelial cells exposed to asbestos fibers, our research group demonstrated asbestos induces EMT in Met-5A cells through the secretion of the TGF-β factor, which in turn mediates the downregulation of epithelial markers (i.e. E-cadherin) via the SMAD-mediated pathway and the upregulation of its downstream transcription factors such as Twist, Zeb-1 and Snail-1, thus promoting EMT [15]. Consequently, the present work has been addressed to evaluate whether this crosstalk between oxidative stress and EMT could be a possible mechanism in MPM able to mediate its carcinogenesis and/or invasiveness, through a molecular study in cellular models of those key factors involved in both cellular processes.
In the light of previous data, we primarily performed experiment to confirm Ref-1 overexpression in MSTO-211H mesothelioma cells towards Met-5A mesothelial not transformed cells, and NSCLC cellular model, used as positive control of Ref-1 overexpression: as expected, we clearly stated a significant Ref-1 overexpression in MSTO-211H and A549 cells, but not in Met-5A not transformed mesothelial cells. Concerning EMT in MPM, some data in literature demonstrated the main TGF-β related transcription factors Twist, Zeb-1, and Snail-1 are overexpressed in MPM and other tumors, by a mechanism mediated by TGF-β, and resulting in downregulation of the key EMT epithelial marker E-cadherin, in fact under-expressed in MPM [17,18,36], as shown previously also by our research group, which demonstrated all Twist, Zeb-1, and Snail-1 factors are significantly overexpressed in mesothelial cells after asbestos exposure, with a consequent E-cadherin downregulation [15].
The next step in trying to demonstrate this crosstalk between oxidative stress and EMT was, first of all, to inhibit Ref-1 by, on the one hand, co-incubating MPM and A549 cells with different concentrations of the well documented E3330 specific inhibitor, and, on the other hand, via silencing directly Ref-1 (siRNA). Our results shown a significant reduction of Ref-1 expression, particularly at the higher dose of E3330 inhibitor tested, both in MSTO-211H and A549 cells, and, at the same time, Ref-1 silencing was effective, demonstrating a significant downregulation of Ref-1 after a siRNA approach.
In support of the observed results related to Ref-1 role in tumorigenesis and invasiveness, we wondered if, after Ref-1 inhibition, the consequent Ref-1 downregulation could in fact also block the EMT. Then, by investigating this point in MSTO-211H cells, we evaluated whether Twist, Zeb-1 and Snail-1 factors could be affected by Ref-1 inhibition/silencing: the results clearly showed a strong and significant reduction of Twist, Zeb-1 and Snail-1 expression when Ref-1 turns out to be silenced, while only Twist was significantly downregulated after E3330 incubation, but not significantly Zeb-1 and Snail-1, nevertheless confirming this effective crosstalk between oxidative stress and EMT in MPM carcinogenesis and invasiveness.
Recently, the use of E3330 inhibitor in pancreatic cancer has been shown to promote a tumor growth reduction [33] and, also in A549 cells and bladder cancer, E3330 treatment has been demonstrated to be effective in attenuating cellular proliferation [24,37], so as Ref-1 shRNA in A549 cells [24]. In our cellular models, we also observed a significant inhibition of cellular proliferation particularly after Ref-1 silencing, by evaluating PCNA expression as index of tumor growth, and the cellular motility by the scratch assay: our results showed a significant reduction of both PCNA expression and cellular migration in MSTO-211H and A549 cells after Ref-1 silencing, thus focusing the crucial role of Ref-1 in controlling cancer proliferation and migration, the latter one particularly by blocking EMT via Twist inhibition. As further confirmation of our results, in literature a reduction of the expression of EMT mesenchymal markers N-cadherin, Vimentin and Snail-1 has been highlighted in cutaneous squamous carcinoma cells following silencing of Ref-1 [34]. Moreover, Yang et al. [35] showed Ref-1 inhibition suppresses EMT in NSCLC, through TGF-β signaling, and restores chemosensitivity. This evidence allows to confirm the central role of Ref-1 in driving EMT and its crosslink with oxidative stress, thus, consequently, in mediating tumorigenesis and invasiveness, particularly via Twist factor. Thus, the block of Ref-1 could not only restore the sensitivity to oxidative stress induced by chemotherapeutics, but also could avoid EMT driven invasiveness and, at the same time, contrast tumor proliferation.
Take as a whole, although there are still many points to be clarified on the molecular mechanism involved in the development and invasiveness of asbestos-induced MPM, we can state that Ref-1 can be considered a promising predictive/prognostic marker and a good therapeutic target in MPM, in the attempt to better counteract this so aggressive cancer by improving the prognosis and the pharmacological approach, particularly important when foreseeing the growing increase of MPM in the next years.
4. Materials and Methods
4.1. Cell cultures
Experiments were performed on cell lines MSTO-211H (human biphasic mesothelioma cells), A549 (human lung adenocarcinoma) and Met-5A (human mesothelial cells): all cells were purchased from American Type Culture Collection (ATCC; Manassas, VA, USA) and grown in RPMI 1640 medium or Ham’s F12 medium supplemented with 10% fetal bovine serum, 1% penicillin and streptomycin. The cells were maintained at 37 °C in a humidified atmosphere of 5% CO2 and 95% air.
4.2. E3330 Ref-1 inhibitor
MSTO-211H cells were incubated with E3330 Ref-1 inhibitor, provided by Sigma-Aldrich (St Louis, MO, USA). E3330 Ref-1 inhibitor powder was previously resuspended with dimethyl sulfoxide (DMSO), then cells were seeded into 6-well plates at a density of 100.000 cells/well and treated with different concentrations of E3330 Ref-1 inhibitor (20, 30 or 50 μmol/L). After 48 h incubation at 37°C, cells were processed as described below.
4.3. siRNA Ref-1
siRNA targeting Ref-1 (siRef-1) was purchased by Santa Cruz Biotecnology (sc-29470; Santa Cruz Biotechnology, CA, USA). Cells were seeded into 6-well plates at a density of 100.000 cells/well. After 24h, at 50% confluency, transfection was performed with jetPRIME® transfection reagent (Polyplus transfection, France): in each well, the medium was replaced with fresh medium containing a transfection mix of jetPRIME® buffer, siRef-1 (60nM) and jetPRIME® reagent. MSTO-211H and A549 cells were treated at 37 °C for 48 h, then cells were processed as described below.
4.4. Nuclear protein extraction
Nuclear protein extraction was performed using Active Motif nuclear extraction kit (Active Motif, La Hulpe, Belgium): cells were washed with a solution of PBS and phosphatase inhibitors on ice and, subsequently, detached from the plates and transferred into iced microtubes, in PBS with phosphatase inhibitors, and then were centrifuged at 13.000 g at 4 °C, for 5 minutes. The supernatant was then removed and the pellet was resuspended in Hypotonic Buffer, incubated in ice for 15 minutes. Next, 10% NP-40 detergent was added, and the tubes were centrifuged at 13.000 g at 4 °C for 30 seconds. The supernatant, with cytoplasmic content, obtained was collected in other tubes and stored at -80 °C. The nuclear pellet was resuspended in a Lysis Buffer (10mM DTT, Lysis buffer and protease inhibitors), then the suspension was incubated on ice for 30 minutes under shaking. Subsequently, further centrifugation was carried out for 10 minutes at 13.000 g at 4 °C, then the supernatant was collected and stored at -80 °C. Finally, protein quantification was performed spectrophotometrically at 595 nm by a Synergy HT microplate reader (Bio-Tek Instruments, Winooski, VT) with Bradford assay.
4.5. Western blot Analysis
For Western Blotting evaluation, nuclear extracts previously obtained were added to Laemmli Buffer 5X (containing 1.5 M Tris pH 6.8, glycerol, β-mercaptoethanol, SDS and bromophenol blue) and subjected to a gradient 4-20% SDS-PAGE using polyacrylamide gels (Mini-ProteanTGX stain-Free, BioRad). The transfer was carried out, through Trans-Blot Turbo Transfer System, on polyvinylidene fluoride (PVDF) membranes (TransBlot Turbo, BioRad) activated in methanol. Then, the membranes were incubated with Block solution (0.1% PBS-Tween and 5% nonfat dry milk) and were decorated for 1h with the diluted primary antibody of interest in 0.1% PBS-Tween and 5% nonfat dry milk. After overnight antibody incubation and serial washes with 0.1% PBS-Tween, the membranes were incubated for 1 hour with peroxidase-conjugated sheep anti-mouse or sheep anti-rabbit IgG antibody (Amersham International, Little Chalfont, UK) diluted 1:3.000 in 0.1% PBS-Tween with 5% nonfat dry milk. After further washing with 0.1% PBS-Tween, proteins were detected by Enhanced Chemiluminescence (ECL), (Perkin Elmer, Waltham, MA, USA). The chemiluminescence signal was then read through the ChemiDoc Touch Imaging System (Biorad). Bio-Rad Image Lab Software 5.1 was used for image processing and densitometric analysis.
The following antibodies were probed: anti-Ref-1, anti-TATA-binding protein (TBP), anti-Twist, anti-Zeb-1, anti-Snail-1 and anti-Proliferating Cell Nuclear Antigen (PCNA). All antibodies were provided by Santa Cruz Biotechnology, Inc. (CA, USA).
4.6. Migration assay
MSTO-211H and A549 cells grew for 48 hrs in medium transfected or not with siRef-1, until confluence. A scratch wound was generated with a pipette tip. After rinsing with medium to remove detached cells, low serum medium (1% FBS) was added. Photographs were taken of each well immediately (T0) and after 24 hrs (T1), using a Leica DMRXA camera (Leica Microsystems, Milan, Italy). Images were analyzed using ImageJ Software (http://rsb.info.nih.gov/ij/). The distance that cells migrated through the area created by scratching was determined by measuring the wound width at T1 and subtracting it from the wound width at the start. The relative migration rate was calculated by setting the percentage of migration of the control cells at time T1 and comparing the percentage of migration of the cells after Ref-1 silencing to this value. The results were representative of three independent experiments.
4.7. Statistical Analysis
Experiments were repeated at three times. Statistical analysis of the results was performed by a one-way analysis of variance (ANOVA) and Tukey test, using GraphPad Prism software (v6.01, San Diego, CA, USA).
Author Contributions
Conceptualization, E.A.; methodology, V.R., G.Z. and M.L.P.; software, V.R.; validation, V.R. and M.L.P.; formal analysis, V.R. and M.L.P.; investigation, E.A. and V.R.; resources, C.R.; data curation, V.R. and G.Z.; writing—original draft preparation, E.A. and V.R.; writing—review and editing, E.A. and V.R.; visualization, V.R. and G.Z.; supervision, E.A.; project administration, E.A. and C.R; funding acquisition, E.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Italian Ministry of University and Research (EX60% Funding 2019 to E.A.), grant “ALDE_RILO_19_01”.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated or analyzed during this study are included in this published article.
Acknowledgments
We acknowledge Costanzo Costamagna, Dept. of Oncology, University of Torino, for the technical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nicolini, F.; Bocchini, M.; Bronte, G.; Delmonte, A.; Guidoboni, M.; Crinò, L.; Mazza, M. Malignant Pleural Mesothelioma: State-of-the-Art on Current Therapies and Promises for the Future. Frontiers in Oncology 2020, 9. [Google Scholar] [CrossRef]
- Brcic, L.; Kern, I. Clinical Significance of Histologic Subtyping of Malignant Pleural Mesothelioma. Transl Lung Cancer Res 2020, 9, 924–933. [Google Scholar] [CrossRef]
- Bibby, A.C.; Tsim, S.; Kanellakis, N.; Ball, H.; Talbot, D.C.; Blyth, K.G.; Maskell, N.A.; Psallidas, I. Malignant Pleural Mesothelioma: An Update on Investigation, Diagnosis and Treatment. Eur Respir Rev 2016, 25, 472–486. [Google Scholar] [CrossRef]
- Kamp, D.W. Asbestos-Induced Lung Diseases: An Update. Translational Research 2009, 153, 143–152. [Google Scholar] [CrossRef]
- Ramundo, V.; Zanirato, G.; Aldieri, E. The Epithelial-to-Mesenchymal Transition (EMT) in the Development and Metastasis of Malignant Pleural Mesothelioma. IJMS 2021, 22, 12216. [Google Scholar] [CrossRef]
- Chew, S.H.; Toyokuni, S. Malignant Mesothelioma as an Oxidative Stress-Induced Cancer: An Update. Free Radical Biology and Medicine 2015, 86, 166–178. [Google Scholar] [CrossRef]
- Chaffer, C.L.; San Juan, B.P.; Lim, E.; Weinberg, R.A. EMT, Cell Plasticity and Metastasis. Cancer and Metastasis Reviews 2016, 35, 645–654. [Google Scholar] [CrossRef]
- Dongre, A.; Weinberg, R.A. New Insights into the Mechanisms of Epithelial–Mesenchymal Transition and Implications for Cancer. Nature Reviews Molecular Cell Biology 2019, 20, 69–84. [Google Scholar] [CrossRef]
- Cannito, S.; Novo, E.; di Bonzo, L.V.; Busletta, C.; Colombatto, S.; Parola, M. Epithelial–Mesenchymal Transition: From Molecular Mechanisms, Redox Regulation to Implications in Human Health and Disease. Antioxidants & Redox Signaling 2010, 12, 1383–1430. [Google Scholar] [CrossRef]
- Xiao, D.; He, J. Epithelial mesenchymal transition and lung cancer. J Thorac Dis 2010, 2, 154–159. [Google Scholar] [CrossRef]
- Legras, A.; Pécuchet, N.; Imbeaud, S.; Pallier, K.; Didelot, A.; Roussel, H.; Gibault, L.; Fabre, E.; Le Pimpec-Barthes, F.; Laurent-Puig, P.; et al. Epithelial-to-Mesenchymal Transition and MicroRNAs in Lung Cancer. Cancers 2017, 9, 101. [Google Scholar] [CrossRef]
- Mahmood, M.Q.; Ward, C.; Muller, H.K.; Sohal, S.S.; Walters, E.H. Epithelial Mesenchymal Transition (EMT) and Non-Small Cell Lung Cancer (NSCLC): A Mutual Association with Airway Disease. Medical Oncology 2017, 34, 45. [Google Scholar] [CrossRef]
- Schramm, A.; Opitz, I.; Thies, S.; Seifert, B.; Moch, H.; Weder, W.; Soltermann, A. Prognostic Significance of Epithelial–Mesenchymal Transition in Malignant Pleural Mesothelioma. European Journal of Cardio-Thoracic Surgery 2010, 37, 566–572. [Google Scholar] [CrossRef]
- Wu, L.; Amjad, S.; Yun, H.; Mani, S.; de Perrot, M. A Panel of Emerging EMT Genes Identified in Malignant Mesothelioma. Scientific Reports 2022, 12, 1007. [Google Scholar] [CrossRef]
- Turini, S.; Bergandi, L.; Gazzano, E.; Prato, M.; Aldieri, E. Epithelial to Mesenchymal Transition in Human Mesothelial Cells Exposed to Asbestos Fibers: Role of TGF-β as Mediator of Malignant Mesothelioma Development or Metastasis via EMT Event. IJMS 2019, 20, 150. [Google Scholar] [CrossRef]
- Iwanami, T.; Uramoto, H.; Nakagawa, M.; Shimokawa, H.; Yamada, S.; Kohno, K.; Tanaka, F. Clinical Significance of Epithelial-Mesenchymal Transition-Associated Markers in Malignant Pleural Mesothelioma. Oncology 2014, 86, 109–116. [Google Scholar] [CrossRef]
- Liu, R.-Y.; Zeng, Y.; Lei, Z.; Wang, L.; Yang, H.; Liu, Z.; Zhao, J.; Zhang, H.-T. JAK/STAT3 Signaling Is Required for TGF-β-Induced Epithelial-Mesenchymal Transition in Lung Cancer Cells. Int J Oncol 2014, 44, 1643–1651. [Google Scholar] [CrossRef]
- Liu, J.; Bao, J.; Hao, J.; Peng, Y.; Hong, F. HSP70 Inhibits High Glucose-Induced Smad3 Activation and Attenuates Epithelial-to-Mesenchymal Transition of Peritoneal Mesothelial Cells. Mol Med Rep 2014, 10, 1089–1095. [Google Scholar] [CrossRef]
- Hydrogen Peroxide Promotes Epithelial to Mesenchymal Transition and Stemness in Human Malignant Mesothelioma Cells. Asian Pacific Journal of Cancer Prevention 2013, 14, 3625–3630. [CrossRef]
- Gorowiec, M.R.; Borthwick, L.A.; Parker, S.M.; Kirby, J.A.; Saretzki, G.C.; Fisher, A.J. Free Radical Generation Induces Epithelial-to-Mesenchymal Transition in Lung Epithelium via a TGF-Β1-Dependent Mechanism. Free Radical Biology and Medicine 2012, 52, 1024–1032. [Google Scholar] [CrossRef]
- Schiavello, M.; Gazzano, E.; Bergandi, L.; Silvagno, F.; Libener, R.; Riganti, C.; Aldieri, E. Identification of Redox-Sensitive Transcription Factors as Markers of Malignant Pleural Mesothelioma. Cancers 2021, 13, 1138. [Google Scholar] [CrossRef]
- Wang, D.; Xiang, D.-B.; Yang, X.; Chen, L.-S.; Li, M.-X.; Zhong, Z.-Y.; Zhang, Y.-S. APE1 Overexpression Is Associated with Cisplatin Resistance in Non-Small Cell Lung Cancer and Targeted Inhibition of APE1 Enhances the Activity of Cisplatin in A549 Cells. Lung Cancer 2009, 66, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Caston, R.A.; Gampala, S.; Armstrong, L.; Messmann, R.A.; Fishel, M.L.; Kelley, M.R. The Multifunctional APE1 DNA Repair–Redox Signaling Protein as a Drug Target in Human Disease. Drug Discovery Today 2021, 26, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Shah, F.; Logsdon, D.; Messmann, R.A.; Fehrenbacher, J.C.; Fishel, M.L.; Kelley, M.R. Exploiting the Ref-1-APE1 Node in Cancer Signaling and Other Diseases: From Bench to Clinic. npj Precision Oncology 2017, 1, 19. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Luo, M.; Kelley, M.R. Human Apurinic Endonuclease 1 (APE1) Expression and Prognostic Significance in Osteosarcoma: Enhanced Sensitivity of Osteosarcoma to DNA Damaging Agents Using Silencing RNA APE1 Expression Inhibition. Molecular Cancer Therapeutics 2004, 3, 679–686. [Google Scholar] [CrossRef]
- Xiang, D.-B.; Chen, Z.-T.; Wang, D.; Li, M.-X.; Xie, J.-Y.; Zhang, Y.-S.; Qing, Y.; Li, Z.-P.; Xie, J. Chimeric Adenoviral Vector Ad5/F35-Mediated APE1 SiRNA Enhances Sensitivity of Human Colorectal Cancer Cells to Radiotherapy in Vitro and in Vivo. Cancer Gene Therapy 2008, 15, 625–635. [Google Scholar] [CrossRef]
- Yang, Z.-Z.; Chen, X.-H.; Wang, D. Experimental Study Enhancing the Chemosensitivity of Multiple Myeloma to Melphalan by Using a Tissue-Specific APE1-Silencing RNA Expression Vector. Clinical Lymphoma and Myeloma 2007, 7, 296–304. [Google Scholar] [CrossRef]
- Fassina, A.; Cappellesso, R.; Guzzardo, V.; Dalla Via, L.; Piccolo, S.; Ventura, L.; Fassan, M. Epithelial–Mesenchymal Transition in Malignant Mesothelioma. Modern Pathology 2012, 25, 86–99. [Google Scholar] [CrossRef]
- Merikallio, H.; Pääkkö, P.; Salmenkivi, K.; Kinnula, V.; Harju, T.; Soini, Y. Expression of Snail, Twist, and Zeb1 in Malignant Mesothelioma. APMIS 2013, 121, 1–10. [Google Scholar] [CrossRef]
- Mehta, A.; Dobersch, S.; Romero-Olmedo, A.J.; Barreto, G. Epigenetics in Lung Cancer Diagnosis and Therapy. Cancer and Metastasis Reviews 2015, 34, 229–241. [Google Scholar] [CrossRef]
- Cheung, E.C.; Vousden, K.H. The Role of ROS in Tumour Development and Progression. Nature Reviews Cancer 2022, 22, 280–297. [Google Scholar] [CrossRef]
- Xie, J.; Li, Y.; Kong, J.; Li, C. Apurinic/Apyrimidinic Endonuclease 1/Redox Factor-1 Could Serve as a Potential Serological Biomarker for the Diagnosis and Prognosis of Oral Squamous Cell Carcinoma. Journal of Oral and Maxillofacial Surgery 2019, 77, 859–866. [Google Scholar] [CrossRef]
- Gampala, S.; Shah, F.; Lu, X.; Moon, H.; Babb, O.; Umesh Ganesh, N.; Sandusky, G.; Hulsey, E.; Armstrong, L.; Mosely, A.L.; et al. Ref-1 Redox Activity Alters Cancer Cell Metabolism in Pancreatic Cancer: Exploiting This Novel Finding as a Potential Target. J Exp Clin Cancer Res 2021, 40, 251. [Google Scholar] [CrossRef]
- Deng, X.; Zhen, P.; Niu, X.; Dai, Y.; Wang, Y.; Zhou, M. APE1 Promotes Proliferation and Migration of Cutaneous Squamous Cell Carcinoma. Journal of Dermatological Science 2020, 100, 67–74. [Google Scholar] [CrossRef]
- Yang, X.; Peng, Y.; Jiang, X.; Lu, X.; Duan, W.; Zhang, S.; Dai, N.; Shan, J.; Feng, Y.; Li, X.; et al. The Regulatory Role of APE1 in Epithelial-to-Mesenchymal Transition and in Determining EGFR-TKI Responsiveness in Non-Small-Cell Lung Cancer. Cancer Medicine 2018, 7, 4406–4419. [Google Scholar] [CrossRef]
- Menju, T.; Date, H. Lung Cancer and Epithelial-Mesenchymal Transition. General Thoracic and Cardiovascular Surgery 2021, 69, 781–789. [Google Scholar] [CrossRef]
- Fishel, M.L.; Xia, H.; McGeown, J.; McIlwain, D.W.; Elbanna, M.; Craft, A.A.; Kaimakliotis, H.Z.; Sandusky, G.E.; Zhang, C.; Pili, R.; et al. Antitumor Activity and Mechanistic Characterization of APE1/Ref-1 Inhibitors in Bladder Cancer. Molecular Cancer Therapeutics 2019, 18, 1947–1960. [Google Scholar] [CrossRef]
Figure 1.
Ref-1 expression in MeT-5A, MSTO-211H and A549 cells. Ref-1 expression (37 kDa) was evaluated (A) by Western blotting on nuclear extracts of MeT-5A, MSTO-211H and A549 cells. TBP (36 kDa) was used as loading control. The image is representative of three independent experiments that produced similar results. (B) Densitometry data are presented as the percent decrease or increase in the protein levels. Data are presented as means ± SD (n=3). One-way ANOVA test: **p < 0.0001, *p < 0.001.
Figure 1.
Ref-1 expression in MeT-5A, MSTO-211H and A549 cells. Ref-1 expression (37 kDa) was evaluated (A) by Western blotting on nuclear extracts of MeT-5A, MSTO-211H and A549 cells. TBP (36 kDa) was used as loading control. The image is representative of three independent experiments that produced similar results. (B) Densitometry data are presented as the percent decrease or increase in the protein levels. Data are presented as means ± SD (n=3). One-way ANOVA test: **p < 0.0001, *p < 0.001.
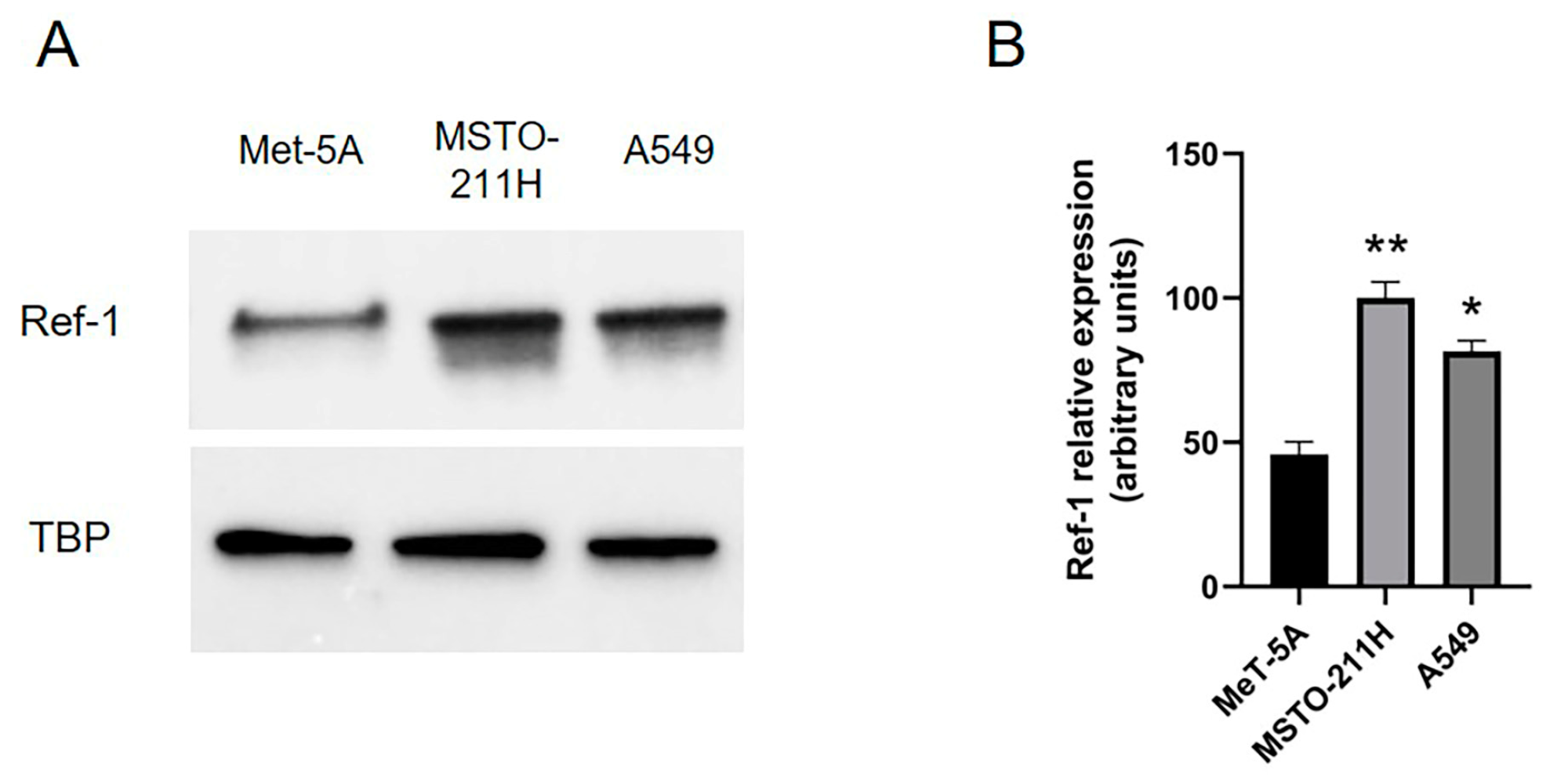
Figure 2.
Effect of Ref-1 inhibitor (E3330) on MSTO-211H and A549 cells. MSTO-211H and A549 cells were incubated for 48 hours with (+) or without (-, Ctrl) increasing concentrations (20, 30 or 50 μM) of the specific Ref-1 inhibitor E3330. (A) Ref-1 expression (37 kDa) was evaluated by Western blotting on nuclear extracts of MSTO-211H and A549 cells. TBP (36 kDa) was used as a loading control. The image is representative of three independent experiments that produced similar results. (B) Densitometry data are presented as the percent decrease or increase in the protein levels versus the respective control. Data are presented as means ± SD (n=3). One-way ANOVA test: MSTO-211H treated cells vs untreated cells **p<0.005, *p<0.05; A549 treated cells vs untreated cells #p<0.001.
Figure 2.
Effect of Ref-1 inhibitor (E3330) on MSTO-211H and A549 cells. MSTO-211H and A549 cells were incubated for 48 hours with (+) or without (-, Ctrl) increasing concentrations (20, 30 or 50 μM) of the specific Ref-1 inhibitor E3330. (A) Ref-1 expression (37 kDa) was evaluated by Western blotting on nuclear extracts of MSTO-211H and A549 cells. TBP (36 kDa) was used as a loading control. The image is representative of three independent experiments that produced similar results. (B) Densitometry data are presented as the percent decrease or increase in the protein levels versus the respective control. Data are presented as means ± SD (n=3). One-way ANOVA test: MSTO-211H treated cells vs untreated cells **p<0.005, *p<0.05; A549 treated cells vs untreated cells #p<0.001.
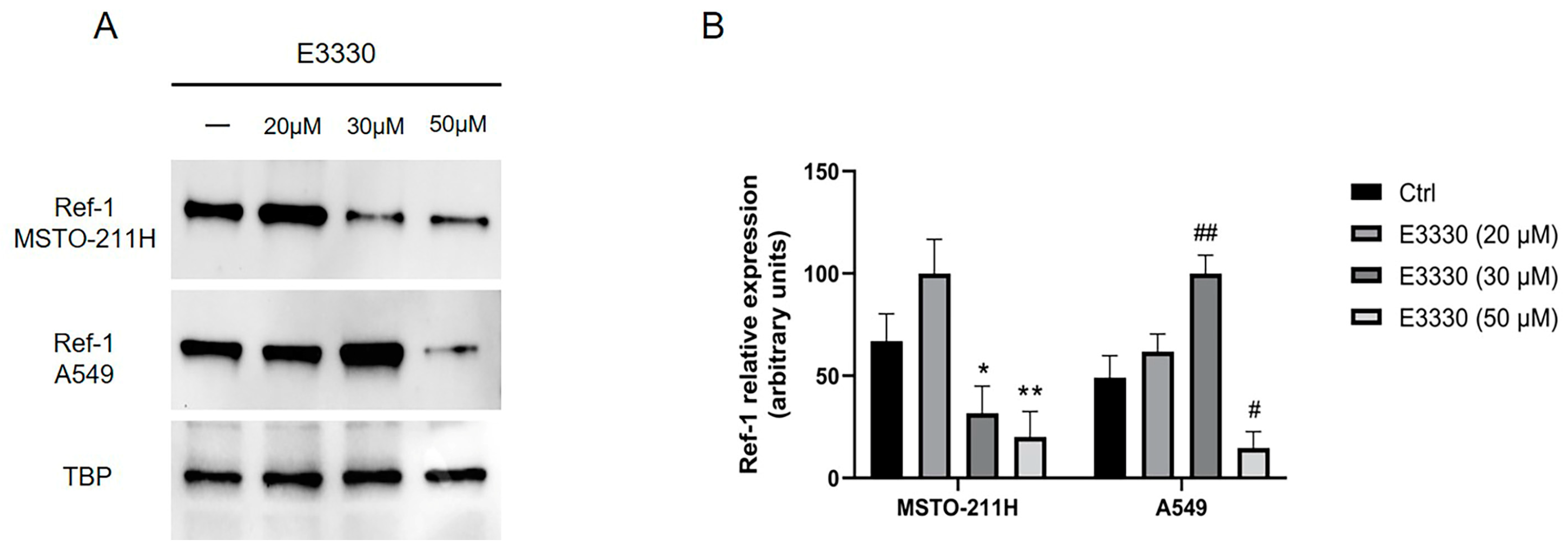
Figure 3.
Ref-1 siRNA knockdown in MSTO-211H and A549 cells. MSTO-211H and A549 cells were transfected for 48 h without (-) or with 60 nM Ref-1 targeted siRNA. (A) Ref-1 expression (37 kDa) was evaluated by Western blotting on nuclear extracts of MSTO-211H and A549 cells. TBP (36 kDa) was used as a loading control. The image is representative of three independent experiments that produced similar results. (B) Densitometry data are presented as the percent decrease or increase in the protein levels versus the respective control. Data are presented as means ± SD (n=3). One-way ANOVA test: siRef-1 transfected cells vs ctrl cells *p<0.0001.
Figure 3.
Ref-1 siRNA knockdown in MSTO-211H and A549 cells. MSTO-211H and A549 cells were transfected for 48 h without (-) or with 60 nM Ref-1 targeted siRNA. (A) Ref-1 expression (37 kDa) was evaluated by Western blotting on nuclear extracts of MSTO-211H and A549 cells. TBP (36 kDa) was used as a loading control. The image is representative of three independent experiments that produced similar results. (B) Densitometry data are presented as the percent decrease or increase in the protein levels versus the respective control. Data are presented as means ± SD (n=3). One-way ANOVA test: siRef-1 transfected cells vs ctrl cells *p<0.0001.
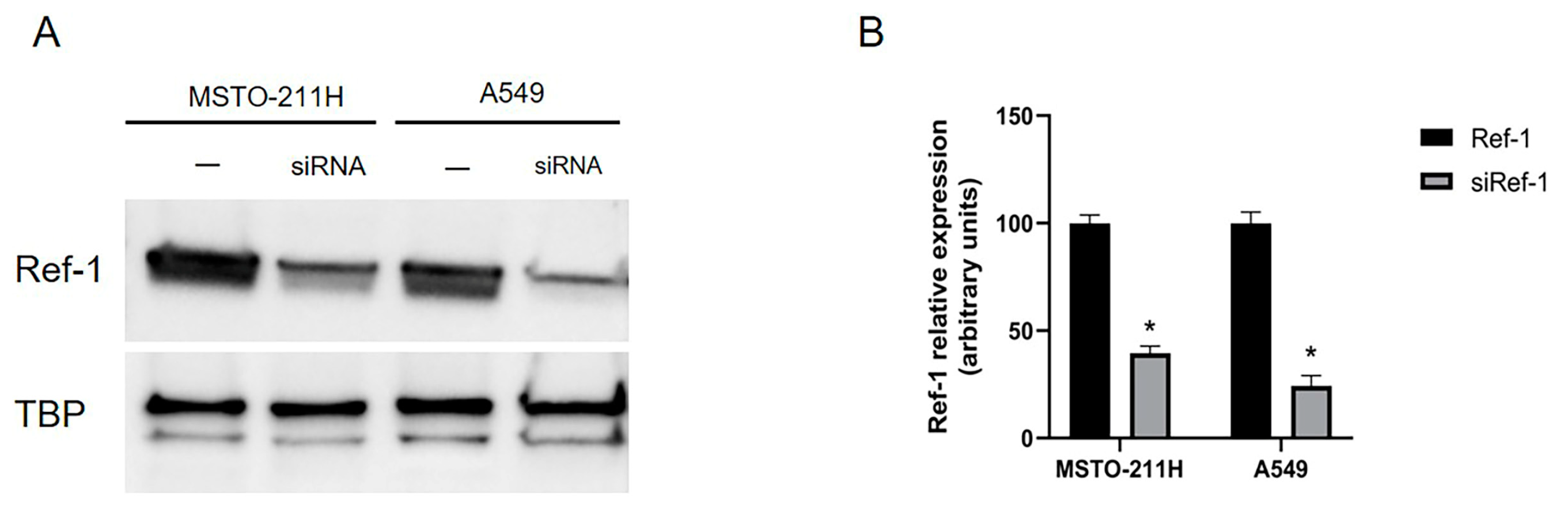
Figure 4.
Effect of Ref-1 inhibition in MSTO-211H cells on EMT-TFs. MSTO-211H cells were incubated for 48 hours with (+) or without (-) 50 μM of E3330 Ref-1 inhibitor. (A) Twist (28 kDa), Zeb-1 (124 kDa) and Snail-1 (29 kDa) EMT-TFs expression was evaluated by Western blotting on nuclear extracts of MSTO-211H cells. TBP (36 kDa) was used as a loading control. The image is representative of three independent experiments that produced similar results. (B) Densitometry data are presented as the percent decrease or increase in the protein levels versus the respective control. Data are presented as means ± SD (n=3). One-way ANOVA test: E3330 treated cells vs ctrl cells *p<0.0001.
Figure 4.
Effect of Ref-1 inhibition in MSTO-211H cells on EMT-TFs. MSTO-211H cells were incubated for 48 hours with (+) or without (-) 50 μM of E3330 Ref-1 inhibitor. (A) Twist (28 kDa), Zeb-1 (124 kDa) and Snail-1 (29 kDa) EMT-TFs expression was evaluated by Western blotting on nuclear extracts of MSTO-211H cells. TBP (36 kDa) was used as a loading control. The image is representative of three independent experiments that produced similar results. (B) Densitometry data are presented as the percent decrease or increase in the protein levels versus the respective control. Data are presented as means ± SD (n=3). One-way ANOVA test: E3330 treated cells vs ctrl cells *p<0.0001.
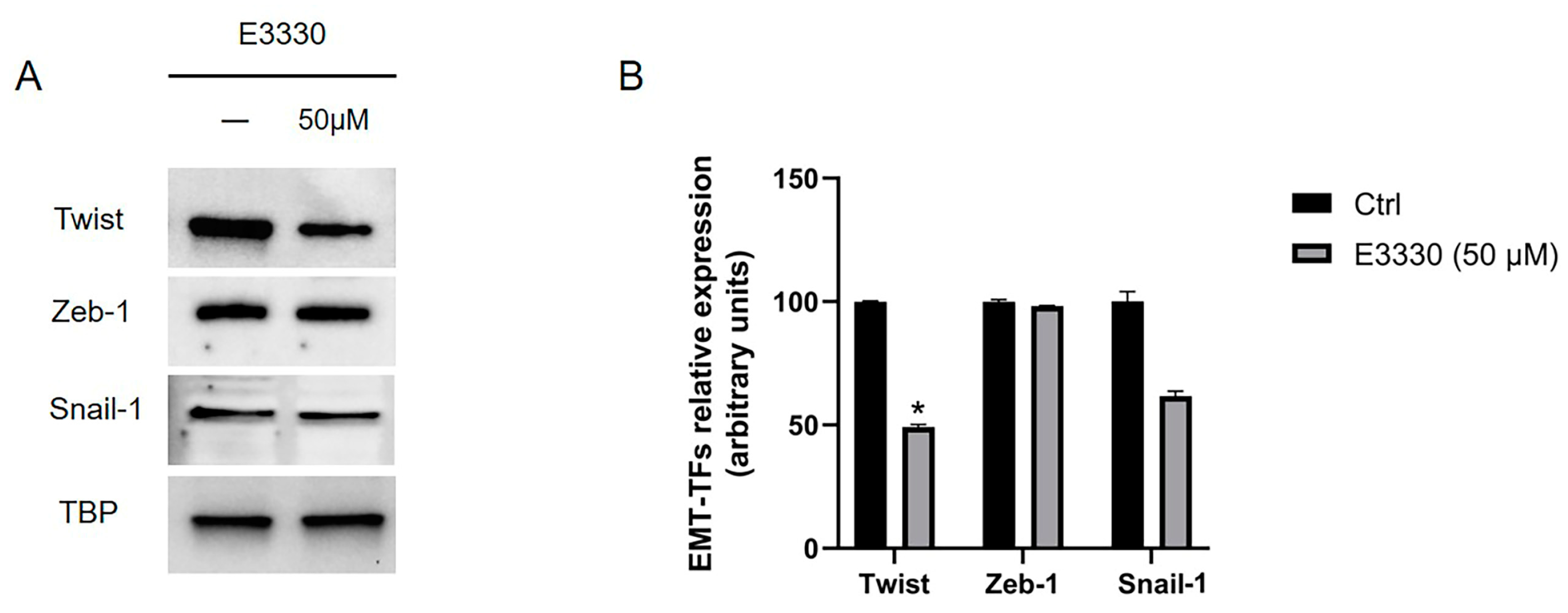
Figure 5.
Effect of Ref-1 siRNA knockdown in MSTO-211H cells on EMT-TFs. MSTO-211H cells were transfected for 48 h without (-) or with 60 nM Ref-1 targeted siRNA. (A) Twist (28 kDa), Zeb-1 (124 kDa) and Snail-1 (29 kDa) EMT-TFs expression was evaluated by Western blotting on nuclear extracts of MSTO-211H cells. TBP (36 kDa) was used as a loading control. The image is representative of three independent experiments that produced similar results. (B) Densitometry data are presented as the percent decrease or increase in the protein levels versus the respective control. Data are presented as means ± SD (n=3). One-way ANOVA test: siRef-1 transfected cells vs ctrl cells **p<0.0001 *p<0.001.
Figure 5.
Effect of Ref-1 siRNA knockdown in MSTO-211H cells on EMT-TFs. MSTO-211H cells were transfected for 48 h without (-) or with 60 nM Ref-1 targeted siRNA. (A) Twist (28 kDa), Zeb-1 (124 kDa) and Snail-1 (29 kDa) EMT-TFs expression was evaluated by Western blotting on nuclear extracts of MSTO-211H cells. TBP (36 kDa) was used as a loading control. The image is representative of three independent experiments that produced similar results. (B) Densitometry data are presented as the percent decrease or increase in the protein levels versus the respective control. Data are presented as means ± SD (n=3). One-way ANOVA test: siRef-1 transfected cells vs ctrl cells **p<0.0001 *p<0.001.
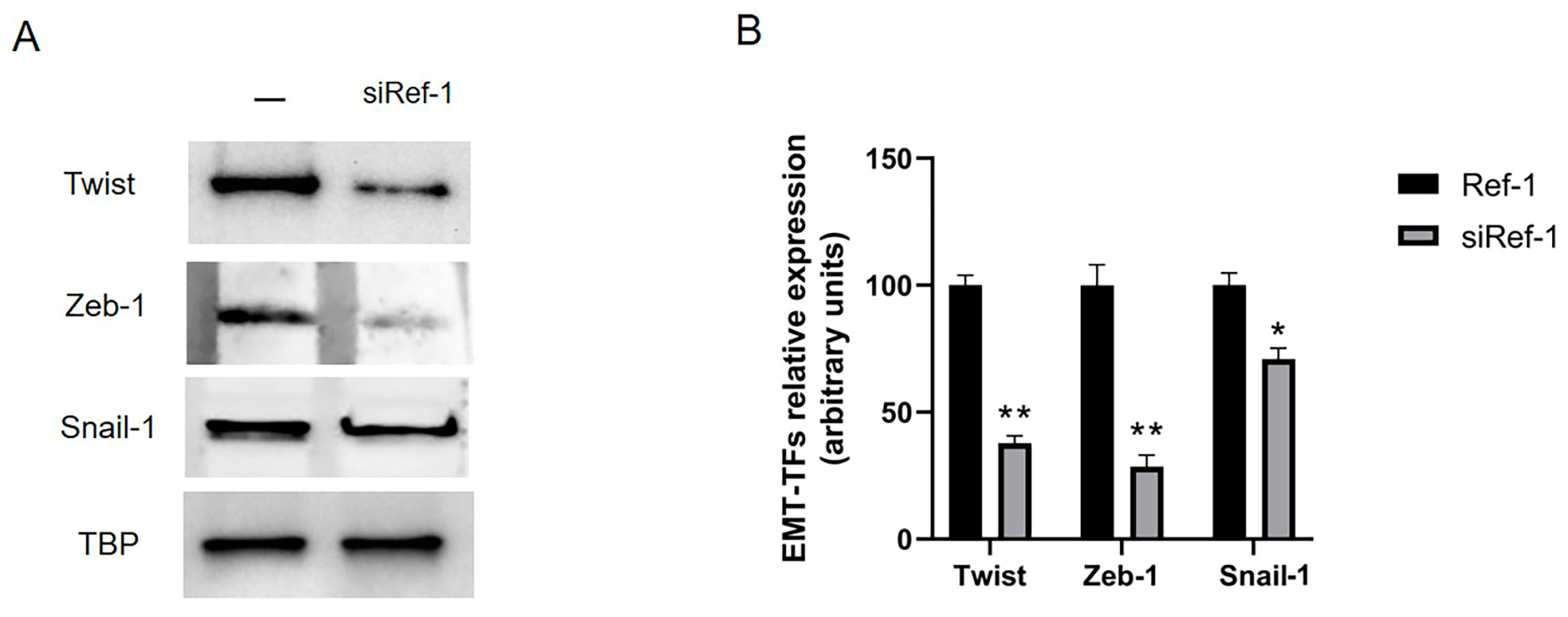
Figure 6.
The effect of Ref-1 siRNA on cell migration. (A) Scratch assay assessed on MSTO-211H and A549 cells after 48 h without (-) or with 60 nM Ref-1 targeted siRNA. The cells were microscopically analyzed at the time of the scratch (T0) and after 24 hrs (T1). (B) The relative migration rate was calculated by setting the percentage of migration of the cells after siRef-1 to this value. The experiments were performed in triplicate and the mean values ± SD were plotted in the relative graph. One-way ANOVA test: siRef-1 transfected cells vs ctrl cells *p<0.0001.
Figure 6.
The effect of Ref-1 siRNA on cell migration. (A) Scratch assay assessed on MSTO-211H and A549 cells after 48 h without (-) or with 60 nM Ref-1 targeted siRNA. The cells were microscopically analyzed at the time of the scratch (T0) and after 24 hrs (T1). (B) The relative migration rate was calculated by setting the percentage of migration of the cells after siRef-1 to this value. The experiments were performed in triplicate and the mean values ± SD were plotted in the relative graph. One-way ANOVA test: siRef-1 transfected cells vs ctrl cells *p<0.0001.
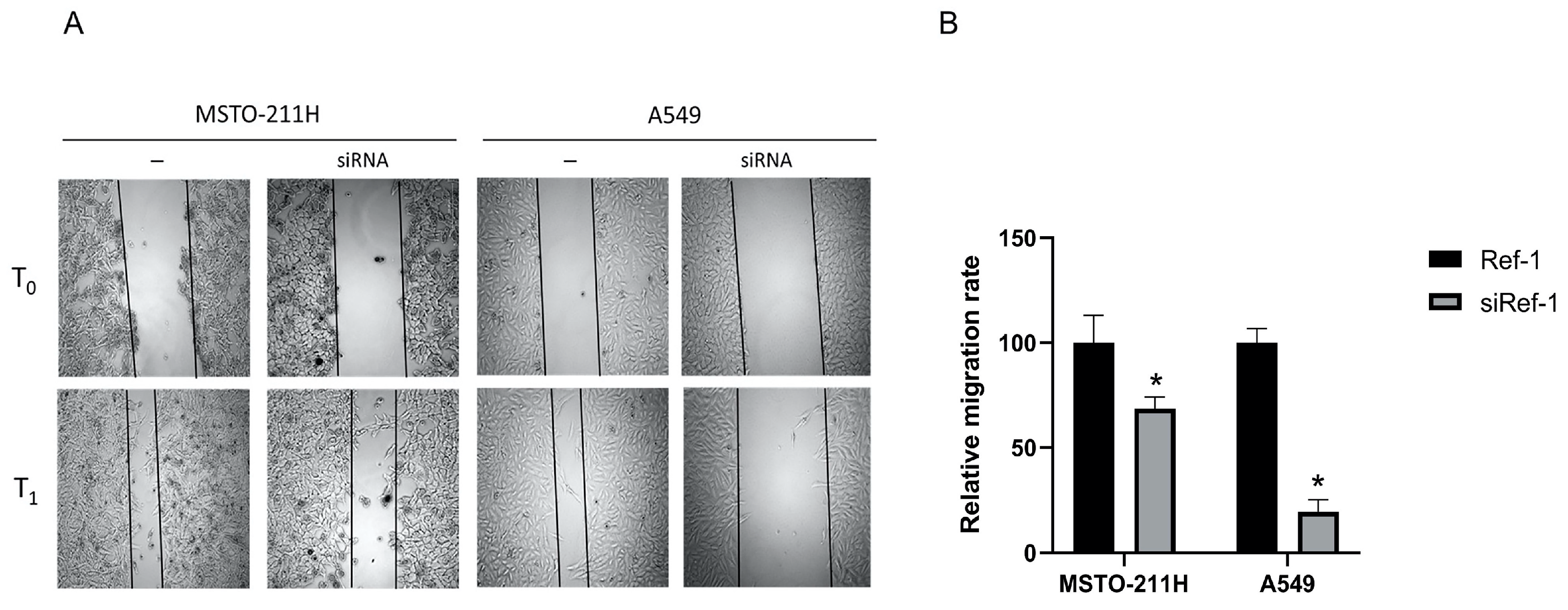
Figure 7.
Effect of Ref-1 siRNA knockdown in MSTO-211H and A549 cells on cell proliferation. MSTO-211H and A549 cells were transfected for 48 h without (-) or with 60 nM Ref-1 targeted siRNA and it was evaluated the expression of PCNA proliferation marker. (A) PCNA expression (36 kDa) was evaluated by Western blotting on nuclear extracts of MSTO-211H and A549 cells. TBP (36 kDa) was used as a loading control. The image is representative of three independent experiments that produced similar results. (B) Densitometry data are presented as the percent decrease or increase in the protein levels versus the respective control. Data are presented as means ± SD (n=3). One-way ANOVA test: siRef-1 transfected cells vs ctrl cells *p<0.0001.
Figure 7.
Effect of Ref-1 siRNA knockdown in MSTO-211H and A549 cells on cell proliferation. MSTO-211H and A549 cells were transfected for 48 h without (-) or with 60 nM Ref-1 targeted siRNA and it was evaluated the expression of PCNA proliferation marker. (A) PCNA expression (36 kDa) was evaluated by Western blotting on nuclear extracts of MSTO-211H and A549 cells. TBP (36 kDa) was used as a loading control. The image is representative of three independent experiments that produced similar results. (B) Densitometry data are presented as the percent decrease or increase in the protein levels versus the respective control. Data are presented as means ± SD (n=3). One-way ANOVA test: siRef-1 transfected cells vs ctrl cells *p<0.0001.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



